Submitted:
31 July 2025
Posted:
31 July 2025
You are already at the latest version
Abstract
Respiratory tract infections (RTIs) remain a major cause of global morbidity and mortality, yet accurate pathogen identification and risk stratification continue to pose clinical challenges. Lipidomics—the comprehensive analysis of lipid species within biological systems—has emerged as a promising tool to unravel host–pathogen interactions and reveal novel diagnostic and prognostic biomarkers. This systematic review synthesizes evidence from nine original studies applying mass spectrometry–based lipidomic profiling in human RTIs, including community-acquired pneumonia (CAP), ventilator-associated pneumonia (VAP), and coronavirus disease 2019 (COVID-19). Across diverse study designs, sample types, and analytical platforms, consistent alterations in lipid metabolism were observed. Perturbations in phospholipid classes, particularly phosphatidylcholines (PCs) and lysophosphatidylcholines (LPCs), were frequently associated with disease severity and immune activation. The ratios of PC to LPC and phosphatidylethanolamine (PE) to lysophosphatidylethanolamine (LPE) emerged as markers of inflammatory remodeling. Sphingolipids—including sphingomyelins (SMs) and sphingosine-1-phosphate (S1P)—were identified as key modulators of monocyte and neutrophil activation. Fatty acid–derived lipid mediators such as oxylipins (e.g., 12,13-epoxyoctadecenoic acid and 15-hydroxyeicosatetraenoic acid) and acylcarnitines reflected pathogen-specific immune responses and mitochondrial dysfunction. Several lipid-based classifiers demonstrated superior diagnostic and prognostic performance compared to conventional clinical scores, including the CURB-65 and pneumonia severity index. However, significant heterogeneity in experimental design, lipid identification workflows, and reporting standards limits inter-study comparability. While preliminary findings support the integration of lipidomics into infectious disease research, larger multi-omic and longitudinal studies are required. This review provides the first comprehensive synthesis of lipidomic alterations in human RTIs and highlights their emerging translational relevance.
Keywords:
lipidomics
; respiratory tract infections
; phosphatidylcholines
; sphingolipids
; biomarkers
; pneumonia
1. Introduction
Respiratory tract infections (RTIs) continue to pose a significant public health burden worldwide, encompassing a broad spectrum of conditions ranging from viral illnesses such as influenza, respiratory syncytial virus (RSV), and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), to bacterial pneumonias caused by pathogens like Streptococcus pneumoniae, Haemophilus influenzae, and Klebsiella pneumoniae [1-4]. These infections can be classified based on their anatomical location—upper or lower respiratory tract—as well as by the setting in which they are acquired, distinguishing community-acquired pneumonia (CAP) from nosocomial infections such as hospital-acquired pneumonia (HAP) and ventilator-associated pneumonia (VAP) [5]. Despite advances in diagnostic microbiology and host biomarker discovery, challenges remain in early detection, pathogen-specific diagnosis, and prognostication of disease severity in both adult and pediatric populations.
In recent years, the emergence of lipidomics as a subfield of metabolomics has offered new opportunities for understanding the pathophysiological basis of infectious diseases [6]. Lipidomics entails the comprehensive characterization of lipid species within biological systems using mass spectrometry–based approaches such as liquid chromatography–mass spectrometry (LC-MS), gas chromatography–mass spectrometry (GC-MS), and shotgun lipidomics [7]. These technologies allow the quantification of hundreds to thousands of lipid molecules, including glycerophospholipids, sphingolipids, fatty acids, and sterols, across various biological matrices [8-10].
Lipidomics has become an increasingly valuable tool for studying host–pathogen interactions, given the essential roles of lipids in membrane structure, energy balance, and immune signaling [11]. Bioactive lipid mediators such as lysophosphatidylcholines (LPCs), sphingolipids, oxylipins, and bile acids are often dysregulated in RTIs, potentially affecting immune responses and disease progression. Integrated omics studies have shown that plasma lipidomic profiles can differentiate viral from bacterial CAP, offering promising diagnostic insights [12]. Specific phosphatidylcholines (PCs) (e.g., PC 16:0_18:1, 36:4, 38:6), identified via ultra-high performance liquid chromatography–tandem mass spectrometry (UHPLC-MS/MS), have been associated with inflammation, oxygenation, and 30-day outcomes, outperforming clinical scores like pneumonia severity index (PSI) and CURB-65 in predicting severe CAP [13].
Evidence from RSV and SARS-CoV-2 studies further supports lipidomics’ diagnostic and prognostic potential. RSV-induced pneumonia alters lipid metabolism in lungs and plasma, especially affecting phospholipids and acylcarnitines [14]. In COVID-19 acute respiratory distress syndrome (ARDS), lipidomic analysis of tracheal aspirates revealed distinct lipid changes associated with VAP, with sphingomyelin (34:1) and PC (O-34:1) outperforming C-reactive protein (CRP) and procalcitonin (PCT) as biomarkers. Similarly, pediatric CAP studies have identified reduced LPCs and increased sphinganine, reflecting inflammation-related metabolic changes [15].
Despite these promising findings, lipidomic applications in RTIs remain underexplored relative to other ‘omics’ disciplines. Heterogeneity in lipidomic platforms, study designs, and clinical endpoints has limited cross-study comparability and translational relevance. Nonetheless, the growing body of literature suggests that lipid profiling may yield novel biomarkers for early diagnosis, disease stratification, and mechanistic understanding of RTIs across diverse patient populations.
The aim of this systematic review is to synthesize existing evidence on the use of lipidomics in respiratory tract infections. Specifically, the review explores lipidomic alterations associated with various pathogens, the biological functions of dysregulated lipids in infection pathophysiology, and the diagnostic and prognostic potential of lipid-based biomarkers in both viral and bacterial RTIs. Furthermore, this work identifies methodological gaps and provide recommendations for future studies aimed at integrating lipidomic tools into clinical infectious disease research.
2. Methods
This systematic review was designed in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) 2020 statement [16]. It has also been registered in the International Prospective Register of Systematic Reviews (PROSPERO) with ID number CRD420251103189.
2.1. Search Strategy
A systematic and comprehensive literature search was performed across four major electronic bibliographic databases—PubMed, Scopus, Web of Science, and EMBASE—from their respective inceptions through June 2025. The search strategy was constructed using a combination of controlled vocabulary terms (e.g., Medical Subject Headings [MeSH]) and relevant free-text terms to maximize sensitivity and capture a broad spectrum of relevant studies.
The core search concepts included lipidomics and respiratory infections. Key search terms encompassed: “lipidomics”, “respiratory tract infection”, “community-acquired pneumonia”, “hospital-acquired pneumonia”, “ventilator-associated pneumonia”, “viral pneumonia”, “bacterial pneumonia”, “COVID-19”, “SARS-CoV-2”, “RSV”, “influenza”, “mass spectrometry”, “ultra-high-performance liquid chromatography”, “liquid chromatography–mass spectrometry”, “gas chromatography–mass spectrometry” “sphingolipids”, “phospholipids”, “oxylipins”, “lysophospholipids”, “bronchoalveolar lavage”, “tracheal aspirate”, “serum”, and “plasma”.
Boolean operators (AND, OR), truncation symbols, and field-specific tags (e.g., [MeSH Terms], [Title/Abstract]) were utilized to enhance the precision and breadth of the search. The full search syntax was adapted to the indexing system of each database. Additionally, the reference lists of all included full-text articles were manually screened to identify potentially eligible studies not captured by electronic search.
2.2. Inclusion and Exclusion Criteria
Studies were eligible for inclusion if they employed any form of lipidomic analysis, whether targeted or untargeted, and presented original research data relevant to RTIs. Eligible study designs included both observational studies, such as cohort, case–control, and cross-sectional studies, and interventional studies, including clinical trials or controlled experimental models. The review considered studies involving human participants (patients or healthy controls), provided the respiratory infection was clearly defined and lipidomic data were reported. Acceptable sample types included serum, plasma, sputum, bronchoalveolar lavage fluid, tracheal aspirates, or other biologically relevant specimens collected in the context of RTIs. To be included, studies had to utilize recognized lipidomic technologies such as LC-MS, GC-MS, nuclear magnetic resonance spectroscopy (NMR), or shotgun lipidomics, and report specific findings on lipid species, lipid class alterations, or lipid pathway perturbations linked to the infection.
Studies were excluded if they did not involve RTIs or if lipidomic analysis was not performed. Publications that focused solely on other omics approaches (e.g., proteomics or transcriptomics) without presenting lipid-specific results were not considered eligible. Secondary literature including reviews, editorials, commentaries, expert opinions, and methodological papers without primary lipidomic data were also excluded. Additionally, case reports, small case series with fewer than five patients, animal studies and conference abstracts that lacked sufficient methodological or results information were not included. Only studies published in English and accessible in full-text format were considered for final inclusion.
2.3. PRISMA Process
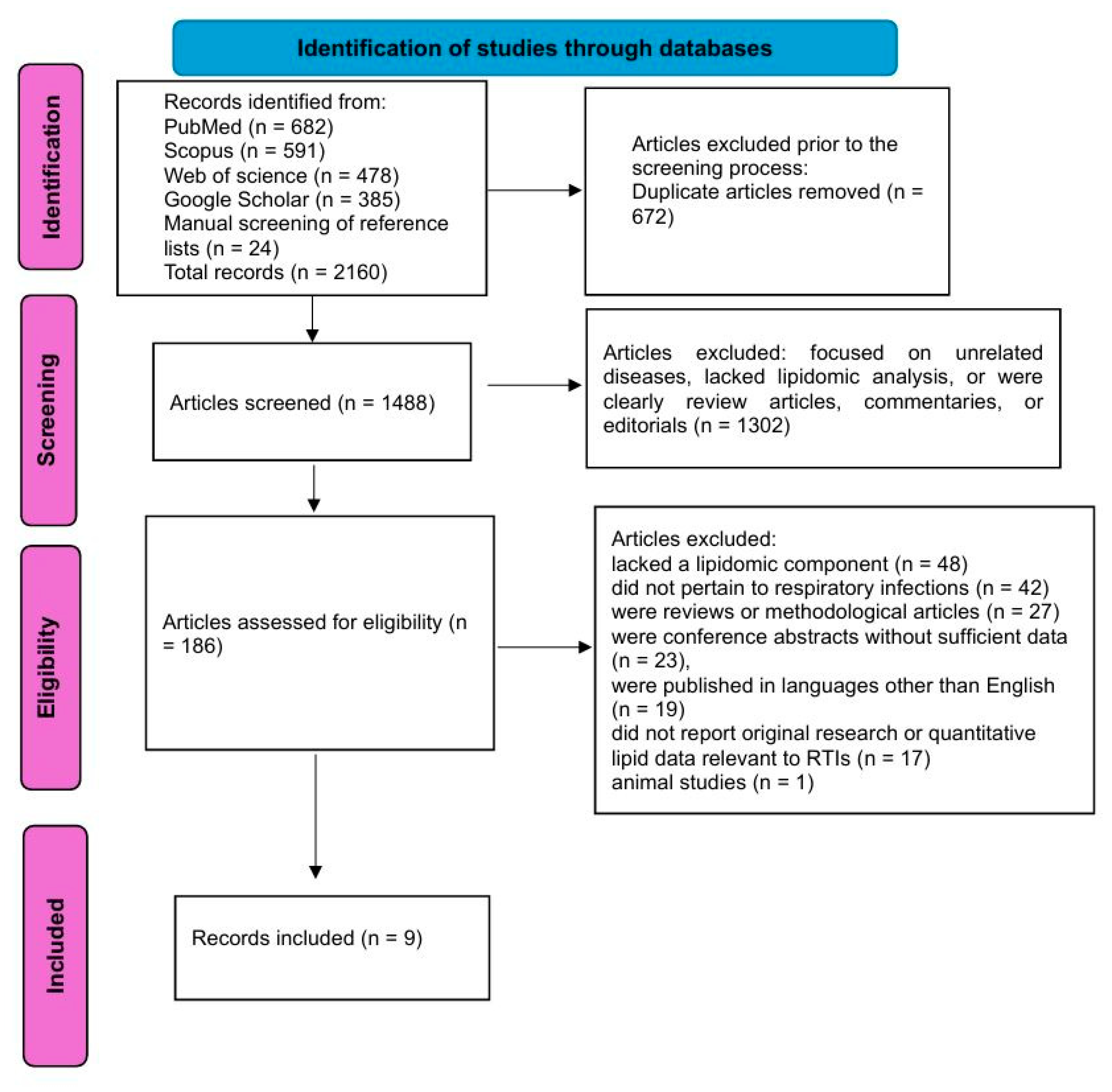
The literature search yielded a total of 2136 records across four major databases: PubMed (682), Scopus (591), Web of Science (478), and Google Scholar (385). An additional 24 articles were identified through manual screening of reference lists from relevant reviews and included studies, bringing the total to 2160 records.
After the removal of 672 duplicate entries, 1488 unique records remained for title and abstract screening. This initial screening was conducted independently by two reviewers, resulting in the exclusion of 1302 records that did not meet the inclusion criteria. These excluded records primarily focused on unrelated diseases, lacked lipidomic analysis, or were clearly review articles, commentaries, or editorials.
A total of 186 full-text articles were retrieved and assessed for eligibility. During full-text screening, 176 articles were excluded for the following reasons: 48 studies lacked a lipidomic component, 42 did not pertain to respiratory infections, 27 were reviews or methodological articles, 23 were conference abstracts without sufficient data, 19 were published in languages other than English, 17 did not report original research or quantitative lipid data relevant to RTIs and one was an animal study.
Following this selection process, 9 studies met all predefined inclusion criteria and were included in the final qualitative synthesis.
Figure 1.
The flowchart of the study selection process.
2.4. Quality Assessment
The methodological quality of the included studies was assessed using the Newcastle–Ottawa Scale (NOS), a validated tool specifically designed for evaluating the risk of bias in non-randomized observational studies, including cohort and case–control designs [17].
2.5. Data Extraction
A standardized data extraction form was used to record study characteristics, including author, year of publication, country, study design, infection type, sample source, analytical platform (e.g., LC-MS, GC-MS), number and classes of lipid species identified, and major lipidomic findings. Outcomes of interest included the identification of specific lipids associated with infection, the use of lipidomics for differential diagnosis or prognosis, and mechanistic insights into host–pathogen interactions mediated by lipids.
2.6. Data Synthesis
Given the heterogeneity in study populations, lipidomic techniques, and outcome measures, a meta-analysis was not performed. Instead, findings were synthesized narratively, with particular attention to patterns of lipid dysregulation across different pathogens, biological plausibility of identified lipids, and consistency of results across studies.
3. Results
3.1. Study Characteristics and Analytical Platforms
The included studies [12, 13, 15, 18-23] span diverse geographic regions and clinical settings, with cohorts recruited from Spain, China, Germany, France, Greece, and the Netherlands. Study designs ranged from prospective observational cohorts and retrospective post-hoc analyses to cross-sectional and pilot investigations. Patient populations encompassed a wide spectrum: adults and children, hospitalized and ICU patients, and those with varying degrees of disease severity, including CAP, COVID-19, VAP, and sepsis. Sample types varied across studies and included serum, plasma, tracheal aspirates, and isolated immune cells (monocytes and neutrophils), reflecting both systemic and compartment-specific lipid responses. Some studies incorporated validation cohorts (e.g., CAPSOD and EARLI), while others integrated multi-timepoint sampling (e.g., Days 1, 3, 6, 12), enabling the capture of dynamic lipidomic trajectories over the course of illness. Most studies employed untargeted lipidomic approaches, utilizing LC-MS/MS or UHPLC-HRMS platforms, ensuring broad coverage of lipid classes. Some studies further integrated proteomics, transcriptomics, or metabolomics, offering mechanistic insights. Importantly, sample types varied—ranging from serum and plasma to tracheal aspirates and isolated immune cells—reflecting diverse anatomical and temporal disease compartments.
Summary of the studies’ characteristics is displayed in Table 1.
3.2. Identified Lipid Biomarkers in RTIs
PCs and LPCs were the most frequently investigated lipid classes, with multiple studies reporting decreased LPC and variable PC levels in patients with CAP and COVID-19. Notably, specific phosphatidylcholine species such as PC(18:2_20:4), PC(36:4), and PC(38:6) were found to be significantly reduced in severe CAP and were independently associated with disease severity, oxygen requirements, and procalcitonin levels. In particular, PC(18:2_20:4) demonstrated high diagnostic accuracy with an area under the curve (AUC) of 0.954 [13].
Similarly, decreased LPC(22:6-sn2) and increased PC(36:1) were among the most discriminatory lipid alterations in hospitalized COVID-19 patients, with the LPC/PC ratio achieving an AUC > 0.95 for disease differentiation [18]. Other studies reported that the PC/LPC and phosphatidylethanolamine/ lysophosphatidylethanolamine (PE/LPE) ratios were positively correlated with systemic inflammation markers and predictive of mortality [22].
Sphingolipids, particularly sphingomyelins and ceramides, were also frequently altered. Sphingomyelin (SM) (34:1) and PC(O-34:1) emerged as strong predictive biomarkers for VAP in COVID-19-associated ARDS, with area under the receiver operating characteristic curve (AUROC) values of 0.85 and 0.83, respectively [21]. In a complementary study, Schuurman et al. (2024) identified significant upregulation of the sphingosine-1-phosphate (S1P) signaling pathway and its regulatory enzymes—sphingosine kinase 1 (SPHK1), UDP-glucose ceramide glucosyltransferase (UGCG), and sphingomyelin phosphodiesterase 1 (SMPD1)—in monocytes and neutrophils of patients with CAP, thereby linking sphingolipid metabolic alterations to pro-inflammatory immune responses.
In terms of fatty acid-derived lipids, oxylipins such as 9- and 13-hydroxyoctadecadienoic acid (9/13-HODE), 15-hydroxyeicosatetraenoic acid (15-HETE), 12,13-epoxyoctadecenoic acid (12,13-EpOME) and 9,10-dihydroxyoctadecenoic acid (9,10-DiHOME) were differentially expressed across viral and bacterial etiologies. Viral CAP was characterized by elevated oxylipins and FA 18:2–containing triglycerides (TGs) and diacylglycerols (DGs), while bacterial CAP showed enrichment in PCs and PC ethers [12].
Several studies identified cholesterol esters as inversely correlated with severity scores and systemic inflammation. In the study by Chouchane et al. [19], a 10% increase in cholesteryl ester (CE) levels by Day 4 in the ICU was associated with a reduced 30-day mortality [odds ration (OR) = 0.84], supporting their prognostic significance.
Notably, lipidomic analyses of tracheal aspirates revealed local lipid remodeling at the infection site. In patients with high VAP suspicion, multiple PC and SM species were upregulated, and these lipid signatures correlated with pathogen type and inflammation [15].
3.3. Role of Lipidomics in Pathophysiological Insights
One of the most consistent findings across studies is the dysregulation of phospholipids, particularly PCs and LPCs, which are integral to membrane integrity, surfactant composition, and inflammatory signaling. Reduced LPC levels, frequently reported in severe CAP and COVID-19, may reflect increased consumption during acute-phase inflammation and impaired resolution of infection [13, 18, 19, 20]. Furthermore, the altered PC/LPC and PE/LPE ratios, which correlated with pro-inflammatory cytokines, CRP levels, and disease severity, suggest that lipid remodeling enzymes such as lysophosphatidylcholine acyltransferase 1/2 (LPCAT1/2) are tightly regulated in infection-induced stress responses [22].
Lipidomics has also shed light on bioactive lipid mediators such as oxylipins, which include hydroxyoctadecadienoic acids (HODEs) and epoxyoctadecenoic acids (EpOMEs). These metabolites, derived from linoleic acid and other polyunsaturated fatty acids via enzymatic oxidation, are known to modulate vascular permeability, leukocyte recruitment, and immune cell activation. Their reduction or elevation in viral versus bacterial infections points to divergent lipid signaling pathways depending on the pathogenic trigger [12, 18].
Another key contribution of lipidomics is the elucidation of sphingolipid metabolism, particularly the role of SMs and S1P in immune regulation. Sphingolipids are not only structural membrane components but also active regulators of neutrophil migration, cytokine release, and lymphocyte trafficking. In CAP patients, upregulation of SPHK1, the enzyme responsible for S1P synthesis, was linked to enhanced monocyte and neutrophil activation, as confirmed by transcriptomic integration and functional inhibition assays [23].
Furthermore, the observation of altered levels of acylcarnitines and triglycerides reflects systemic metabolic reprogramming during infection. Elevated acylcarnitines indicate disrupted mitochondrial β-oxidation and energy stress, while increased circulating triglycerides—particularly in viral CAP and COVID-19—are consistent with hepatic lipogenesis driven by inflammatory cytokines such as interleukin -6 (IL-6) and tumor necrosis factor alpha (TNF-α) [12, 18]. These findings suggest that host lipid metabolism is not merely a bystander but a central player in the host response to infection.
Lipidomic signatures derived from localized samples such as tracheal aspirates further illuminate tissue-specific processes. In VAP, differential expression of surfactant-associated lipids and ether-linked phospholipids pointed to alveolar epithelial injury rather than direct bacterial lipid contribution. These changes were correlated with pathogen-specific profiles, offering insights into pulmonary lipid metabolism at the site of infection [15, 21].
3.4. Lipidomic Alterations by Pathogen Type
In the prospective cohort study by Rischke et al. [12], lipidomic profiling of patients with viral versus bacterial CAP revealed substantial differences. Bacterial CAP was characterized by elevated levels of PCs and ether-linked PCs, consistent with enhanced membrane biosynthesis and neutrophilic activation. In contrast, viral CAP showed increased concentrations of TGs and DGs, particularly those containing linoleic acid (18:2), as well as higher levels of pro-inflammatory oxylipins, including 12,13-EpOME and 9,10-DiHOME. These findings suggest that viral infections are associated with increased lipid catabolism and eicosanoid pathway activation, whereas bacterial infections induce structural lipid accumulation likely due to phagocytic and oxidative burst activity.
Similarly, in a pediatric ICU cohort, Virgiliou et al. [15] demonstrated that lipidomic profiles in VAP varied by pathogen. Blood lipid signatures—including specific PCs, SMs, and TGs—differentiated infections due to Staphylococcus aureus and Klebsiella pneumoniae, supporting the concept of pathogen-specific metabolic remodeling in critically ill children. Notably, PCs (e.g., PC 32:2) and TGs (e.g., TG 48:3) showed good discriminatory performance (AUC > 0.75), indicating the potential of lipidomics for microbial stratification in clinical practice.
Further evidence for pathogen-specific lipid responses was observed in the study by Kassa-Sombo et al. [21], which examined tracheal aspirate lipidomes in COVID-19 ARDS patients with or without VAP. While not explicitly focused on microbial taxonomy, the study identified eight significantly upregulated lipids [including SM(34:1) and PC(O-34:1)] in VAP patients. These lipids were associated with epithelial cell membrane breakdown rather than direct bacterial lipid production, but their expression patterns reflected the presence and severity of superimposed bacterial infection.
3.5. Temporal Dynamics and Prognostic Implications
In a multicenter study of critically ill CAP patients, Chouchane et al. [19] documented time-dependent recovery of specific lipid classes, most notably CE and LPC. More than half (58%) of all quantified lipid species were decreased upon ICU admission. However, restoration of CE levels by Day 4 was independently associated with improved outcomes, with a 10% increase corresponding to a 16% reduction in 30-day mortality risk (OR = 0.84). These findings indicate that early lipid normalization may serve as a surrogate for systemic recovery and immune homeostasis.
Similarly, Schuurman et al. [23] assessed monocyte and neutrophil lipidomes at baseline and one month following CAP. While lipid perturbations in monocytes largely resolved during convalescence, neutrophils exhibited sustained alterations, including persistent elevations in polyunsaturated TGs and DGs. The enduring disruption of neutrophil lipid metabolism, coupled with upregulation of sphingolipid synthesis enzymes (e.g., SPHK1), suggests prolonged immunometabolic dysregulation even after apparent clinical recovery.
In pediatric populations, Virgiliou et al. [15] conducted serial lipidomic analyses in patients with suspected VAP across four timepoints (Days 1, 3, 6, and 12). A gradual increase in PCs and TGs was observed in the high-suspicion group, parallel with microbiological confirmation and clinical deterioration. Specific lipids such as TG(48:3) and SM(40:1) showed increasing discrimination power over time (AUC > 0.75), indicating that lipid signatures may not only track disease progression but also anticipate microbiological confirmation and clinical outcomes before conventional diagnostics.
Prognostically, several studies reported lipid-based models that outperformed traditional clinical scores. In the study by Chen et al. [13], phosphatidylcholine species PC(18:2_20:4) and PC(38:6) achieved AUCs of 0.954 and 0.959, respectively, for predicting CAP severity—superior to CURB-65 and the PSI. Likewise, Ma et al. [22] identified that elevated PC/LPC and PE/LPE ratios were positively associated with disease severity, inflammatory markers, and prolonged hospital stay, and independently predicted 30-day mortality (AUC = 0.838 for PC/LPC ratio).
3.6. Integrative Models and Clinical Translation
Several included studies employed multivariate statistical and machine learning techniques to build predictive models based on lipidomic features. In a retrospective cohort of hospitalized COVID-19 patients, Castané et al. [18] used Monte Carlo simulations, principal component analysis (PCA), and partial least squares discriminant analysis (PLS-DA) to identify a panel of discriminatory lipids, including LPC(22:6-sn2) and PC(36:1). The derived ratio LPC(22:6-sn2)/PC(36:1) yielded an AUC exceeding 0.95 for distinguishing COVID-19 from other infectious or inflammatory conditions, demonstrating the diagnostic superiority of lipidomic models over conventional clinical parameters.
Similarly, Chen et al. [13] developed a ROC-based classifier using individual phosphatidylcholines (e.g., PC(38:6) and PC(18:2_20:4)) that outperformed traditional clinical severity scores such as CURB-65 and PSI, with AUCs > 0.95. These models were not only statistically robust but also biologically interpretable, as the lipid species selected were implicated in inflammation, cellular stress, and membrane dynamics.
The integration of lipidomics with other omics platforms further enhanced discriminatory power and mechanistic understanding. Rischke et al. [12] combined lipidomics with proteomics and metabolomics to differentiate viral from bacterial CAP, demonstrating that pathogen-specific clusters could be generated based on co-expression networks of linoleic acid–derived oxylipins, triglycerides, and host immune markers such as TNF-related apoptosis-inducing ligand (TRAIL) and Lymphocyte-activation gene 3 (LAG-3). Such multi-omic integration supports not only diagnostic classification but also insights into immune-metabolic crosstalk.
4. Discussion
This systematic review synthesized evidence from nine original studies that employed lipidomic analyses in the context of RTIs, including CAP, VAP, and COVID-19. Across diverse populations, analytical platforms, and sample types, a consistent pattern of lipid alterations emerged. Dysregulation of PCs and LPCs was the most commonly reported finding, with decreased LPC levels frequently associated with severe disease. Changes in lipid profiles also demonstrated pathogen-specific patterns, distinguishing viral from bacterial etiologies. Furthermore, temporal dynamics of lipid species were linked to clinical outcomes, with early normalization of certain lipid classes (e.g., cholesterol esters) associated with reduced mortality. Importantly, lipidomic biomarkers often outperformed conventional clinical scores in predictive models, underscoring their potential clinical utility.
Lipidomic analyses have provided critical mechanistic insights into how RTIs disrupt host lipid metabolism. One of the most consistent findings across studies is the downregulation of LPCs and the alteration of PC/LPC and PE/LPE ratios, particularly in severe CAP and COVID-19 cases. These changes suggest involvement of phospholipid remodeling pathways regulated by LPCAT1/2, enzymes known to be modulated in response to inflammatory stress, potentially reflecting the host’s attempt to maintain membrane integrity and immune regulation [13, 18, 19, 22].
Oxylipins, which are oxygenated derivatives of polyunsaturated fatty acids, were also differentially expressed across pathogen types. Elevated concentrations of 12,13-EpOME and 9,10-DiHOME in viral CAP highlight increased cytochrome P450 and epoxide hydrolase activity, which may mediate pro-inflammatory or vasoactive responses. Conversely, reduced levels of 9/13-HODE and 15-HETE observed in bacterial pneumonia may reflect suppressed resolution-phase signaling and altered lipid mediator balance [12, 18].
Additionally, sphingolipid metabolism has emerged as a key regulatory axis in the host response to RTIs. The upregulation of S1P signaling and its biosynthetic enzyme SPHK1, along with changes in specific SM species (e.g., SM 34:1), suggest a mechanistic link to monocyte and neutrophil activation, as well as cytokine production [21, 23]. These alterations likely contribute to broader immunometabolic rewiring and may perpetuate inflammation or immune exhaustion during infection.
Finally, multiple studies reported elevated acylcarnitines and TGs in patients with viral pneumonia, indicating disrupted mitochondrial β-oxidation and enhanced hepatic lipogenesis, respectively—hallmarks of systemic metabolic stress and energy imbalance during acute infection [12, 18].
Despite the promising results, several technical limitations warrant consideration. First, there is significant heterogeneity in analytical platforms (e.g., LC-MS/MS, UHPLC-HRMS, NMR), lipid extraction protocols, and bioinformatic pipelines, which limits direct comparison across studies. Second, many studies utilized untargeted lipidomics, which, while comprehensive, may suffer from challenges in lipid identification, quantification, and inter-laboratory reproducibility. Additionally, normalization techniques, internal standards, and statistical thresholds varied widely across studies, potentially influencing results.
Sample types and timing also varied considerably—ranging from serum to tracheal aspirates and from single to multi-timepoint sampling—which complicates harmonization. Finally, few studies validated their findings in independent cohorts, and even fewer integrated lipidomic data with clinical decision-making tools or prospective outcome measures.
This systematic review has several limitations that should be acknowledged. First, the number of eligible studies was relatively small (n = 9), reflecting the early and still emerging nature of lipidomic applications in RTIs. As a result, some conclusions are based on limited data, and findings may not be generalizable across all RTI subtypes, geographic regions, or patient populations. Second, the heterogeneity in study designs, sample types, disease severity, and lipidomic platforms precluded quantitative synthesis through meta-analysis. Variations in analytical methods—including differences in mass spectrometry instrumentation, lipid extraction protocols, normalization strategies, and data processing workflows—introduce methodological variability that complicates direct comparisons across studies.
Third, while most studies achieved high scores on the Newcastle–Ottawa Scale, many lacked external validation cohorts, used small sample sizes, or did not control for potential confounders such as comorbidities, medication use, or nutritional status, all of which can influence the lipidome. Additionally, few studies conducted longitudinal sampling or integrated lipidomic data with real-time clinical decision-making, limiting insights into the temporal utility of lipid biomarkers in routine care. Fourth, publication bias may have influenced the findings, as studies with null or non-significant results may be underrepresented in the published literature. Moreover, only English-language articles were included, potentially excluding relevant non-English studies. Finally, while this review included both adult and pediatric populations, age-specific differences in lipid metabolism were not separately analyzed due to the limited number of pediatric studies, which may have masked age-related variability in lipidomic signatures.
Despite these limitations, this review provides the first comprehensive synthesis of the current evidence on the role of lipidomics in RTIs. Future studies addressing the above limitations are needed to validate and expand upon these preliminary findings.
Several gaps remain in the current literature. Most notably, large-scale, multi-center studies are lacking, and many of the available data are derived from pilot or single-center cohorts with modest sample sizes. Future research should prioritize validation of lipid-based biomarkers in diverse patient populations and clinical settings. There is also a need for standardized protocols and reporting frameworks for lipidomic studies, akin to the Minimum Information About a Metabolomics Experiment (MIAME) guidelines used in transcriptomics.
Moreover, integrative multi-omics approaches—including transcriptomics, proteomics, and metabolomics—should be employed more consistently to contextualize lipidomic changes within broader biological networks. Functional studies exploring how specific lipid mediators (e.g., oxylipins, sphingolipids) influence immune cell behavior during infection would also enhance mechanistic understanding.
Finally, efforts should be directed toward translating lipidomic findings into clinically applicable tools, such as rapid point-of-care tests or algorithm-based risk stratification models. Collaboration between clinicians, analytical chemists, and computational biologists will be critical to realizing the translational potential of lipidomics in RTIs.
5. Conclusions
This systematic review highlights the emerging role of lipidomics in unraveling the complex host response to respiratory tract infections. Across diverse clinical settings and infection types, lipidomic profiling has revealed consistent perturbations in key lipid classes—particularly phospholipids, sphingolipids, oxylipins, and acylcarnitines—reflecting alterations in membrane remodeling, immune activation, and systemic metabolic stress. The diagnostic and prognostic potential of lipid-based biomarkers, including specific lipid ratios and composite classifiers, appears promising and, in some studies, exceeds the predictive performance of conventional clinical scoring systems. Moreover, the integration of lipidomics with proteomic and transcriptomic data offers mechanistic insights into pathogen-specific immune pathways and points toward precision medicine approaches in infectious disease management. Nevertheless, substantial methodological heterogeneity, limited validation cohorts, and the lack of standardization across platforms currently constrain clinical translation. Future research should focus on multi-center, longitudinal studies with standardized protocols, age- and pathogen-stratified analyses, and integration with other omics technologies. Overall, lipidomics holds substantial potential not only as a tool for biomarker discovery but also for advancing our understanding of host–pathogen interactions in RTIs.
References
- Georgakopoulou, VE. Insights from respiratory virus co-infections. World J Virol. 2024;13(4):98600. [CrossRef]
- Georgakopoulou VE, Lempesis IG, Tarantinos K, Sklapani P, Trakas N, Spandidos DA. Atypical pneumonia (Review). Exp Ther Med. 2024;28(5):424. [CrossRef]
- Hanage WP, Schaffner W. Burden of Acute Respiratory Infections Caused by Influenza Virus, Respiratory Syncytial Virus, and SARS-CoV-2 with Consideration of Older Adults: A Narrative Review. Infect Dis Ther. 2025;14(Suppl 1):5-37. [CrossRef]
- Liapikou A, Torres A. The clinical management of lower respiratory tract infections. Expert Rev Respir Med. 2016;10(4):441-452. [CrossRef]
- Lim, WS. Pneumonia—Overview. Encyclopedia of Respiratory Medicine. 2022:185–97. [CrossRef]
- Wenk, MR. Lipidomics of host-pathogen interactions. FEBS Lett. 2006 ;580(23):5541-51. [CrossRef]
- Han X, Gross RW. Shotgun lipidomics: multidimensional MS analysis of cellular lipidomes. Expert Rev Proteomics. 2005;2(2):253-64. [CrossRef]
- Anh NK, Thu NQ, Tien NTN, Long NP, Nguyen HT. Advancements in Mass Spectrometry-Based Targeted Metabolomics and Lipidomics: Implications for Clinical Research. Molecules. 2024 Dec 16;29(24):5934. [CrossRef]
- Cajka T, Fiehn O. Comprehensive analysis of lipids in biological systems by liquid chromatography-mass spectrometry. Trends Analyt Chem. 2014 Oct 1;61:192-206. [CrossRef]
- Wu Z, Shon JC, Liu KH. Mass Spectrometry-based Lipidomics and Its Application to Biomedical Research. J Lifestyle Med. 2014 Mar;4(1):17-33. [CrossRef]
- Schenck EJ, Plataki M, Wheelock CE. A Lipid Map for Community-acquired Pneumonia with Sepsis: Observation Is the First Step in Scientific Progress. Am J Respir Crit Care Med. 2024;209(8):903-904. [CrossRef]
- Rischke S, Gurke R, Zielbauer AS, Ziegler N, Hahnefeld L, Köhm M, Kannt A, Vehreschild MJ, Geisslinger G, Rohde G, Bellinghausen C, Behrens F, Study Group C. Proteomic, metabolomic and lipidomic profiles in community acquired pneumonia for differentiating viral and bacterial infections. Sci Rep. 2025;15(1):1922. [CrossRef]
- Chen L, Zheng Y, Zhao L, Zhang Y, Yin L, He Y, Ma X, Xu Y, Gao Z. Lipid profiles and differential lipids in serum related to severity of community-acquired pneumonia: A pilot study. PLoS One. 2021;16(3):e0245770. [CrossRef]
- Shan J, Qian W, Shen C, Lin L, Xie T, Peng L, Xu J, Yang R, Ji J, Zhao X. High-resolution lipidomics reveals dysregulation of lipid metabolism in respiratory syncytial virus pneumonia mice. RSC Adv. 2018 Aug 17;8(51):29368-29377. [CrossRef]
- Virgiliou C, Begou O, Ftergioti A, Simitsopoulou M, Sdougka M, Roilides E, Theodoridis G, Gika H, Iosifidis E. Untargeted Blood Lipidomics Analysis in Critically Ill Pediatric Patients with Ventilator-Associated Pneumonia: A Pilot Study. Metabolites. 2024;14(9):466. [CrossRef]
- Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 2021;372:n71. [CrossRef]
- Wells GA, Shea B, O’Connell D, Peterson J, Welch V, Losos M, Tugwell P. The Newcastle-Ottawa Scale (NOS) for assessing the quality of nonrandomised studies in meta-analyses.
- Castañé H, Iftimie S, Baiges-Gaya G, Rodríguez-Tomàs E, Jiménez-Franco A, López-Azcona AF, Garrido P, Castro A, Camps J, Joven J. Machine learning and semi-targeted lipidomics identify distinct serum lipid signatures in hospitalized COVID-19-positive and COVID-19-negative patients. Metabolism. 2022;131:155197. [CrossRef]
- Chouchane O, Schuurman AR, Reijnders TDY, Peters-Sengers H, Butler JM, Uhel F, Schultz MJ, Bonten MJ, Cremer OL, Calfee CS, Matthay MA, Langley RJ, Alipanah-Lechner N, Kingsmore SF, Rogers A, van Weeghel M, Vaz FM, van der Poll T. The Plasma Lipidomic Landscape in Patients with Sepsis due to Community-acquired Pneumonia. Am J Respir Crit Care Med. 2024 Apr 15;209(8):973-986. [CrossRef]
- Saballs M, Parra S, Martínez N, Amigo N, Cabau L, Iftimie S, Pavon R, Gabaldó X, Correig X, Paredes S, Vallvé JM, Castro A. Lipidomic and metabolomic changes in community-acquired and COVID-19 pneumonia. J Lipid Res. 2024;65(9):100622. [CrossRef]
- Kassa-Sombo A, Verney C, Pasquet A, Vaidie J, Brea D, Vasseur V, Cezard A, Lefevre A, David C, Piver E, Nadal-Desbarats L, Emond P, Blasco H, Si-Tahar M, Guillon A. Lipidomic signatures of ventilator-associated pneumonia in COVID-19 ARDS patients: a new frontier for diagnostic biomarkers. Ann Intensive Care. 2025;15(1):78. [CrossRef]
- Ma X, Chen L, He Y, Zhao L, Yu W, Xie Y, Yu Y, Xu Y, Zheng Y, Li R, Gao Z. Targeted lipidomics reveals phospholipids and lysophospholipids as biomarkers for evaluating community-acquired pneumonia. Ann Transl Med. 2022;10(7):395. [CrossRef]
- Schuurman AR, Chouchane O, Butler JM, Peters-Sengers H, Joosten S, Brands X, Haak BW, Otto NA, Uhel F, Klarenbeek A, van Linge CC, van Kampen A, Pras-Raves M, van Weeghel M, van Eijk M, Ferraz MJ, Faber DR, de Vos A, Scicluna BP, Vaz FM, Wiersinga WJ, van der Poll T. The shifting lipidomic landscape of blood monocytes and neutrophils during pneumonia. JCI Insight. 2024;9(4):e164400. [CrossRef]

Table 1.
Summary of the studies’ characteristics.
| First Author | Year | Country | Study Design | Population | Sample Type | Analytical Platform | Lipid Classes Analyzed | Main findings | NOS |
|---|---|---|---|---|---|---|---|---|---|
| Castané [18] | 2022 | Spain | Retrospective post-hoc cohort study | 126 hospitalized COVID-19 patients, 45 hospitalized COVID-19-negative patients (infectious/inflammatory), 50 healthy controls | Serum | Semi-targeted lipidomics by UHPLC-QTOF-MS, machine learning (Monte Carlo, PCA, PLS-DA) | Acylcarnitines, lysophospholipids (LPC, LPE), phosphatidylcholines (PC), oxylipins (e.g., 9/13-HODE, 15-HETE), bile acids, long-chain TGs | COVID-19 and non-COVID inflammatory patients shared a common lipid signature characterized by elevated acylcarnitines and LPE, and decreased 9/13-HODE and 15-HETE. However, specific discrimination between COVID-19 and other conditions was achieved via decreased LPC(22:6-sn2), increased PC(36:1), and changes in secondary bile acids. Arachidonic acid levels were markedly decreased in both COVID-19 and other infectious groups. Machine learning identified oxylipins, carnitines, and phospholipids as key discriminatory features. Alterations in β-oxidation and fatty acid metabolism were prominent. The ratio of LPC22:6-sn2/PC36:1 achieved an AUC > 0.95. No association was found with ICU admission or mortality. |
8/9 |
| Chen [13] | 2021 | China | Cross-sectional pilot study | 28 CAP patients (13 severe, 15 non-severe), 20 matched non-CAP controls | Serum | Untargeted UHPLC-MS/MS lipidomics, OPLS-DA, ROC, multivariate regression | Glycerophospholipids (PC, PE), sphingolipids (SM, HexCer), lysophospholipids (LPC, LPE), diacylglycerols, cholesterol esters, free fatty acids | Lipid profiles differed significantly across NC, NSCAP, and SCAP groups. CAP patients exhibited reduced LPC and PE levels, increased Hex2Cer and cholesterol esters. Four lipids (PC[16:0_18:1], PC[18:2_20:4], PC[36:4], PC[38:6]) outperformed CURB-65 and PSI in ROC analysis for disease severity. PC(18:2_20:4) had AUC 0.954, PC(38:6) had AUC 0.959. PC(18:2_20:4) and PC(36:4) correlated negatively with FiO2 and PCT; PC(16:0_18:1) positively with PCT. Lower PC levels were linked to longer hospital stay and higher 30-day mortality. Combined phospholipid biomarkers showed potential for disease monitoring, diagnosis, and prognosis in CAP. |
7/9 |
| Chouchane [19] | 2024 | Netherlands | Prospective cohort study | 169 ICU patients with CAP sepsis, 51 noninfected ICU controls, 48 outpatient controls; plus two validation cohorts (CAPSOD, EARLI) | Plasma | Untargeted lipidomics via HPLC-MS (1,833 lipid species across 33 classes), data validated in external cohorts | Cholesterol esters, triacylglycerols, phospholipids (PC, PE, LPC), sphingomyelins, ceramides, sulfatides, plasmalogens, lysolipids | Patients with sepsis due to CAP exhibited a profound shift in the plasma lipidome compared to both healthy and noninfected ICU controls. 58% of lipid species were decreased, while 6% increased. Cholesterol esters and lysophospholipids showed strong inverse associations with SOFA score and systemic inflammation. Recovery of specific lipids such as cholesterol esters by Day 4 in ICU was linked to lower 30-day mortality (e.g., OR=0.84 per 10% increase). The lipidomic profile showed partial recovery over time, and a specific TG-rich pattern distinguished CAP from other ICU patients. LPC and Chol-E emerged as key prognostic lipids. Lipid class patterns were validated across CAPSOD and EARLI cohorts. Results support lipidomics as a biomarker and prognostic tool in CAP-associated sepsis. | 9/9 |
| Rischke [12] | 2025 | Germany | Prospective cohort study | 69 patients with community-acquired pneumonia (CAP): 43 viral (incl. COVID-19), 26 bacterial | Plasma (baseline, day 3, day 7) | LC-MS/MS (MxP Quant 500), Olink proteomics, lipid network enrichment (LINEX2), PLS-DA, PCA | Phosphatidylcholines, ether-PCs, lysophosphatidylcholines (LPC), triglycerides, diglycerides, bile acids (GCA, TCA, TCDCA), oxylipins (EpOME, DiHOME) | Distinct plasma lipidomic profiles differentiated viral from bacterial CAP. Bacterial CAP was characterized by elevated PCs and PC-ethers, while viral CAP showed increased TGs, DGs (especially FA 18:2-containing), and linoleic acid–derived oxylipins (12,13-EpOME, 9,10-DiHOME). Proteomic markers like TRAIL, LAG-3, and LAMP3 were elevated in viral CAP, while CLEC4D and EN-RAGE were elevated in bacterial CAP. Integrated clustering of lipidomic, metabolomic, and proteomic analytes supported co-expression of pathogen-specific patterns. PLS-DA and hierarchical clustering identified robust discriminatory features. Findings indicate the potential of lipidomics and multi-omics in pathogen-specific diagnosis and individualized treatment strategies in CAP. |
8/9 |
| Saballs [20] | 2024 | Spain | Prospective observational study | 71 CAP patients, 75 COVID-19 pneumonia patients, 75 healthy controls (age- and sex-matched) | Serum | 1H NMR spectroscopy (Liposcale® assay), BUME extraction, LipSpin, multivariate analysis (PLS-DA, random forest, ROC) | Phosphatidylcholine, lysophosphatidylcholine, PUFA, DHA, esterified/free cholesterol, triglycerides, HDL/LDL/VLDL subclasses, glycoproteins | Both CAP and COVID-19 pneumonia patients exhibited hypolipidemia with reduced levels of HDL-c, phosphatidylcholine, lysophosphatidylcholine, PUFA, and DHA. Severity was associated with increased VLDL-c, IDL-c, LDL-tg/LDL-c, triglycerides, and glycoproteins (GlycA, GlycB, GlycF), along with decreased HDL particles and esterified cholesterol. COVID-19 patients showed more pronounced alterations. A lipidomic-metabolomic model based on PC, glycerophospholipids, creatine, glutamate, isoleucine, alanine, and glycoproteins achieved AUC = 0.935 for etiology classification and 0.931 for severity. Metabolites linked to inflammation and energy metabolism (lactate, glucose, creatine, BCAAs, glutamate) were elevated. Decreased glutamine and DHA in severe COVID-19 suggest impaired immune energy supply and pro-inflammatory resolution failure. These alterations reflect lipid-mediated immune modulation, metabolic reprogramming, and potential for diagnostic/prognostic biomarker development. |
7/9 |
| Kassa-Sombo [21] | 2025 | France | Prospective observational cohort study | 39 patients with COVID-19 ARDS (26 with VAP, 13 without VAP), matched controls | Tracheal aspirate | Untargeted lipidomics via UHPLC-HRMS (Q-Exactive MS), data processed with Workflow4Metabolomics/XCMS | Phosphatidylcholines, phosphatidylethanolamines, sphingomyelins, ether-linked PCs and PEs, lysophospholipids | Significant alterations in the tracheal aspirate lipidome were observed in VAP versus non-VAP COVID-19 ARDS patients (p = 0.003). Among 272 identified lipids, PCs were most frequently dysregulated, with 17 upregulated and 6 downregulated. SM(34:1) and PC(O-34:1) were the most predictive biomarkers for VAP, showing AUROC of 0.85 and 0.83 respectively. Eight key lipids were identified via multivariate analyses (PCA, PLS-DA, OPLS-DA), all upregulated in VAP. Lipidomic changes reflected breakdown of surfactant and pulmonary cells rather than bacterial origin. Combined lipid biomarkers modestly improved diagnostic performance (AUROC up to 0.86). These results suggest that lipidomics of tracheal aspirates offers potential for accurate VAP diagnostics and highlights lipid remodeling at the site of infection. |
8/9 |
| Ma [22] | 2022 | China | Prospective multi-center cohort study | 58 patients with CAP (30 NSCAP, 28 SCAP), 11 healthy controls | Serum | Targeted LC-MS/MS, qRT-PCR validation, GEO database transcriptome analysis | Phosphatidylcholine (PC), lysophosphatidylcholine (LPC), phosphatidylethanolamine (PE), lysophosphatidylethanolamine (LPE) | LPC levels were decreased, while PE, PC, PC/LPC, and PE/LPE ratios were increased in CAP. PE combined with CURB-65 predicted severity (AUC = 0.848), while PC/LPC ratio improved 30-day mortality prediction (AUC = 0.838). PC(36:4) and LPC(18:2/0:0) emerged as species-level biomarkers. Expression of LPCAT2 was upregulated and LPCAT1 downregulated in SCAP; LPCAT2 positively correlated with inflammatory genes, LPCAT1 negatively. Lipid ratios (PC/LPC, PE/LPE) and PE were positively correlated with CRP, neutrophil percentage, and PSI, and negatively with albumin and lymphocytes. Alterations in Lands cycle enzymes support metabolic dysregulation during infection. | 8/9 |
| Schuurman [23] | 2024 | Netherlands | Case-control cohort study | 48 CAP patients (baseline and 1-month follow-up), 25 matched noninfectious controls | Isolated blood monocytes and neutrophils | Untargeted HPLC-MS lipidomics with transcriptomics integration | Sphingolipids (SM, ceramides, S1P), phospholipids (PC, PE), lysophospholipids (LPC, LPE), fatty acids, diacylglycerols (DG), triglycerides (TG), BMPs | Pneumonia significantly altered the lipidomic landscape of monocytes and neutrophils, with distinct profiles. Monocyte lipid changes were mostly decreases in PC, PE, and SM species and resolved after 1 month. In contrast, neutrophils showed persistent changes with increased PC, PE, DG, BMP, and polyunsaturated TGs. Sphingolipid metabolism, particularly S1P signaling, was upregulated. Transcriptomic analysis confirmed upregulation of key enzymes (SPHK1, UGCG, SMPD1). Functional validation showed that inhibiting SPT and SPHK1 blunted cytokine production in both cell types. The study demonstrated a mechanistic link between altered lipid profiles and immune function during CAP. | 9/9 |
| Virgiliou [15] | 2024 | Greece | Prospective pilot cohort study | 20 critically ill pediatric patients (12 high VAP suspicion, 8 low suspicion) | Plasma (blood samples at 4 timepoints: Days 1, 3, 6, 12) | Untargeted LC-HRMS (UPLC-TIMS-TOF/MS), multivariate + univariate analysis, MS-DIAL, Lipostar2 | Phosphatidylcholines (PC), lysophosphatidylcholines (LPC), sphingomyelins (SM), triglycerides (TG), diglycerides (DG), cholesterol esters (CE), carnitines | Untargeted lipidomics revealed 144 blood lipid species in critically ill pediatric patients with VAP suspicion. PCs, SMs, TGs, and DGs were significantly altered between high and low mCPIS groups. High suspicion group showed increased levels of PCs and TGs over time. Discriminatory lipids (e.g., PC 32:2, TG 48:3, SM 40:1) had AUC > 0.75. Specific lipid profiles were associated with culture-confirmed pathogens, including S. aureus and K. pneumoniae. Multivariate models (OPLS-DA) distinguished both VAP severity and pathogen type. Phospholipid shifts correlated with inflammatory markers and may indicate pathogen-specific metabolic remodeling. The study supports lipidomics as a promising diagnostic tool for early VAP identification and microbial stratification in pediatric ICU settings. | 7/9 |
AUC – Area Under the Curve; ARDS – Acute Respiratory Distress Syndrome; BCAAs – Branched-Chain Amino Acids; BMP – Bis(monoacylglycerol)phosphate CAP – Community-Acquired Pneumonia; CE – Cholesteryl Esters; COVID-19 – Coronavirus Disease 2019; CRP – C-Reactive Protein CURB-65 – Confusion, Urea, Respiratory Rate, Blood pressure, Age ≥65; DHA – Docosahexaenoic Acid ; DG – Diacylglycerol; DiHOME – Dihydroxyoctadecenoic Acid; EpOME – Epoxyoctadecenoic Acid; FA – Fatty Acid; FiO₂ – Fraction of Inspired Oxygen; GCA – Glycocholic Acid; GEO – Gene Expression Omnibus ; HDL – High-Density Lipoprotein; HexCer – Hexosylceramide; HRMS – High-Resolution Mass Spectrometry ; ICU – Intensive Care Unit ; IDL – Intermediate-Density Lipoprotein ; LAG-3 – Lymphocyte Activation Gene 3; LAMP3 – Lysosome-Associated Membrane Glycoprotein 3 LC-MS/MS – Liquid Chromatography–Tandem Mass Spectrometry ; LDL – Low-Density Lipoprotein; LINEX2 – Lipid Network Explorer 2; LPC – Lysophosphatidylcholine; LPCAT1/2 – Lysophosphatidylcholine Acyltransferase 1/2; LPE –Lysophosphatidylethanolamine; MaCPIS – Modified Clinical Pulmonary Infection Score; MS-DIAL – Mass Spectrometry–Data Independent AnaLysis; NMR – Nuclear Magnetic Resonance; NOS – Newcastle–Ottawa Scale; OPLS-DA – Orthogonal Partial Least Squares Discriminant Analysis; PCA – Principal Component Analysis; PC – Phosphatidylcholine; PCR – Polymerase Chain Reaction; PE – Phosphatidylethanolamine; PLS-DA – Partial Least Squares Discriminant Analysis; PCT – Procalcitonin; PSI – Pneumonia Severity Index; PUFA – Polyunsaturated Fatty Acid; qRT-PCR – Quantitative Reverse Transcription Polymerase Chain Reaction; ROC – Receiver Operating Characteristic; SCAP – Severe Community-Acquired Pneumonia; SM – Sphingomyelin; SOFA – Sequential Organ Failure Assessment; SPHK1 – Sphingosine Kinase 1; S1P – Sphingosine-1-Phosphate; TG – Triglyceride; TIMS-TOF/MS – Trapped Ion Mobility Spectrometry – Time-of-Flight Mass Spectrometry; TRAIL – TNF-Related Apoptosis-Inducing Ligand; UHPLC – Ultra-High Performance Liquid Chromatography; UHPLC-HRMS – Ultra-High Performance Liquid Chromatography – High Resolution Mass Spectrometry; UPLC – Ultra Performance Liquid Chromatography; VAP – Ventilator-Associated Pneumonia; VLDL – Very Low-Density Lipoprotein; XCMS – Cross-platform Chromatographic Alignment and Peak Picking Software.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



