Submitted:
30 July 2025
Posted:
31 July 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Sex-based immunological dimorphisms significantly influence the outcome of vac-cination, yet the molecular mediators underpinning these differences remain largely elusive. MicroRNAs (miRNAs), key post-transcriptional regulators of gene expression, have emerged as critical modulators of innate and adaptive immune responses. In this study, we investigated the expression profile of selected circulating miRNAs as poten-tial biomarkers of sex-specific humoral responses to the mRNA COVID-19 vaccine in a cohort of health care workers. Plasma samples were collected longitudinally at a de-fined time point (average 71 days) post-vaccination and analyzed using RT-qPCR to quantify a panel of immune-relevant miRNAs. Anti-spike (anti-S) IgG titers were measured by chemiluminescent immunoassays. Our results revealed sex-dependent differences in miRNA expression dynamics, with miR-221-3p, miR-148a-3p signifi-cantly overexpressed in vaccinated female HCWs and miR-155-5p overexpressed in vaccinated males. MiR-148a-3p showed a significant association with anti-S/RBD IgG levels in a sex-specific manner.
Bioinformatic analysis for miRNAs targets indicated distinct regulatory networks and pathways involved in innate and adaptive immune responses, potentially underlying the differential immune activation observed between males and females. These find-ings support the utility of circulating miRNAs as minimally invasive biomarkers for monitoring and predicting sex-specific vaccine-induced immune responses and pro-vide mechanistic insights that may inform tailored vaccination strategies.
Keywords:
health care workers
; vaccine
; COVID-19
; sex
; anti-S/RBD
; microRNA
1. Introduction
The unprecedented global impact of the COVID-19 pandemic has accelerated the development and deployment of vaccines, particularly mRNA-based vaccines, which have proven highly effective in preventing severe disease caused by SARS-CoV-2 [1]. However, emerging evidence suggests that the immune response to these vaccines is not uniform across all individuals, with notable differences observed between males and females [2]. In particular, females generally exhibit higher antibody levels and stronger humoral responses than males [3,4,5].
In recent years, significant progress has been made in detecting biomarkers for vaccine immunogenicity and efficacy [6]. In this context, microRNAs (miRNAs), a class of small, non-coding RNAs involved in the regulation of gene expression at the post-transcriptional level [7], have been shown to be attractive candidates [8,9]. MiRNAs are known to modulate key aspects of the innate and adaptive immune responses, including the activation and differentiation of immune cells, cytokine production, and the regulation of antibody production; they can be secreted and circulate in body fluids, associated with microvesicles, apoptotic bodies, exosomes, making them stable and easily measurable in blood [10,11]. Given these features, miRNAs are increasingly recognized as potential biomarkers for various diseases and immune responses, including vaccine-induced immunity. Additionally, in humans, around 118 microRNAs are located on the X chromosome (roughly 10% of the total known microRNAs), according to the miRBase database and only 4 encoded on the Y chromosome; the X chromosome’s microRNAs are thought to play important roles in various cellular processes, including immunity and cancer [12]. Given the above-mentioned crucial role of microRNAs in regulating important cellular processes and pathways, such as apoptosis, differentiation, oxidative stress, etc., and in immune responses during infection and vaccination, females have a greater regulatory capacity than males.
Despite the growing interest in the role of miRNAs as predictive markers of vaccine response, there is limited understanding of how specific miRNAs might contribute to sex-specific differences in the antibody response to the COVID-19 mRNA vaccine.
This study aims to profile selected miRNAs, known to take part in immune responses to vaccinations based on previous reports [13,14,15,16,17,18], to determine their potential as markers of sex-specific anti-S/RBD response to the COVID-19 mRNA vaccine. By analyzing miRNA expression in vaccinated individuals, we seek to identify those that associate with differences in anti-S antibody titers between COVID-19 vaccinated male and female individuals
2. Results
2.1. Description of the Study Population
The study population was composed of 128 healthcare workers (HCWs), 47 men (36.7%) and 81 women (63.3%), working at the Bambino Gesù Children’s Hospital in Rome, Italy. All participants had no history of SARS-CoV-2 infection and had received two doses of the mRNA vaccine. Overall, the median age was 45 years, slightly higher among female workers (47 years) than male workers (42 years) (Table 1). The majority of HCWs belonged to the oldest age group (46-72 years) for females (70.5%), and the youngest age group (23–45 years) for males (43.3%), according to the age-based classification of the research population.
2.2. Sex Differences in Humoral Response to COVID-19 Vaccination
The level of anti-S/RBD antibodies was measured 71 days post-second dose of the COVID-19 vaccine.
As shown in Table 2, vaccinated female HCWs showed significantly higher anti-S/RBD levels (geometric mean 1069 AU/ml, 95% CI: 925.8- 1234 AU/ml) than male HCWs (geometric mean 713.9 AU/ml, 95% CI: 509.1-1001 AU/ml). Considering age at vaccination, elder male and female HCWs tended to have lower anti-S/RBD antibody (Ab) concentrations (979 AU/ml in females and 526 AU/ml in males) than those of younger subjects (1031 AU/ml in females and 863 AU/ml in males), although the differences observed were not statistically significant (p>0,05).
2.3. miRNA Expression Profile in Plasma from COVID-19 Vaccinated Male and Female HCWs and Their Association with Anti-S/RBD Levels
Six miRNAs (miR-148a-3p, miR-221-3p, miR-150-5p, miR-155-5p, miR-223-3p, and miR-98-5p) were analyzed as potential sex-specific markers of the COVID-19 vaccine response. These miRNAs were selected based on their reported activity in immune responses to vaccinations (all the selected miRNAs), being encoded on the X chromosome (miR-223-3p and miR-98-5p), or having their expression regulated by sex hormones (all the selected miRNAs).
Significantly higher expression levels of most of the circulating miRNAs analyzed (miR-148a-3p, miR-150-5p, miR-155-5p, miR-98-5p, and miR-223-3p) were detected in the vaccinated total workers cohort compared to the unvaccinated controls, as shown in Figure 1A. The only exception was miR-221-3p, whose expression level seemed to be the same in vaccinated and non-vaccinated HCWs.
However, sex-specific analysis (Figure 1B) revealed that some of the microRNAs had unequal plasma concentrations in male and female vaccinated HCWs. Specifically, miR-148a-3p and miR-221-3p levels increased in female HCWs but remained the same as unvaccinated and sex-matched control subjects in male HCWs after COVID-19 vaccination. Additionally, miR155-5p was significantly overexpressed in vaccinated males, not in females.
To investigate the possible association between miRNAs that showed sex-different expression in vaccinated HCWs (miR-148a-3p, miR-221-3p and miR-155-5p) and anti-S/RBD response following COVID-19 vaccination, we initially grouped subjects with microRNAs fold-changes ≥1,5, that means an increase in miRNA levels, and < 1.5, that means no variation in miRNA expression, comparing vaccinated and unvaccinated HCWs. Anti-S/RBD levels were subsequently compared between the two groups. In the overall vaccinated workers, the group having a fold variation for miR-148a-3p ≥1,5, showed significantly higher levels of anti-S/RBD antibodies compared to those in the group with < 1.5-fold changes (Figure 2A).
Sex-disaggregated analysis revealed that only female HCWs in the group with miR-148a-3p fold changes ≥1.5 showed significantly higher levels of the anti-S/RBD compared to those in the group with <1.5-fold changes (Figure 2B). A similar trend was observed in male workers, but without statistical significance (Figure 2C).
In contrast, neither the entire cohort nor the sex-disaggregated population of health care professionals showed a statistically significant difference in anti-S/RBD distributions when the same categorization based on fold variation (≥1.5 and <1.5) was applied for miR-221-3p and miR-155-5p (Figures 2 D-I), although they showed a trend similar to that observed with miR-148a-3p.
These results raise the possibility that miR-148a-3p may have a sex-specific role in the anti-S response to COVID-19 immunization.
In Figure S1, the results of a simple regression model are shown, further indicating a strong statistical association between anti-S/RBD antibodies and a ≥1.5-fold change in circulating miR-148a-3p. This association was observed following COVID-19 vaccination, but only in female healthcare workers.
2.4. Targets of miR-148a-3p
To ascertain the biological implications of the variation in miR-148a-3p expression associated with sex, we first constructed a network of the miRNA-target interactions that have been experimentally verified, followed by a functional enrichment analysis of the KEGG pathways that comprise the targets (Table S1 and Table 3).
Target genes of miR-148a-3p were discovered to control a total of 75 significantly enriched pathways (FDR < 0.05), as shown in Supplementary Table 1. It’s interesting to note that several of these pathways were engaged in modulating the immune response and/or were often involved in response to several viral infections (Table 3). These include: Epstein-Barr virus infection, FoxO signaling pathway, Human T-cell leukemia virus infection, Human papillomavirus infection, TGF-beta signaling pathway, PI3K-Akt signaling pathway”, Herpes simplex virus 1 infection, MAPK signaling pathway, Measles, Human cytomegalovirus infection, Apoptosis, Hepatitis B, Adipocytokine signaling pathway, p53 signaling pathway, Cellular senescence, Cell cycle, mTOR signaling pathway, Chemochine signaling pathway, Th17 cell differentiation, Regulation of actin cytoskeleton, Antigen processing and presentation.
2.5. Targets of miR-221-3p and miR-155-5p
We further analyzed the biological significance of miR-221-3p and miR-155-5p, whose expression levels were significantly different between male and female vaccinated HCWs.
miR-221-3p has been reported to promote B cell proliferation and class switch recombination in vitro, by targeting Foxp1 and Arid1a as regulators of Ig class switch recombination (CSR), as well as IgE production in allergic hypersensitivity, which suggests a role for this miRNA as a regulator of cell production of antibodies associated with allergy [21].
Table S2 and Table 4 show the KEGG pathways and miR-221-3p targets involved, indicating that miR-221-3p has a role in the regulation of the immune responses (FoxO signaling, Toll–like receptor signaling, leukocyte migration, Th17 differentiation, chemokine signaling), sex hormones pathways, cell senescence, and apoptosis, among others.
MiR-155-5p is strongly expressed by hematopoietic cells and acts as a master regulator of immune responses, finely tuning both the innate and adaptive immune systems. [22]. Its targets do indeed play a role in controlling inflammatory pathways, responses to viral infections, and Toll-like receptor signaling (TNF signalling pathway, response to HBV, HCV, EBV viruses, and Toll-like receptors pathways) (Table 5 and Table S3).
Results presented in Figure 3 highlight experimentally validated targets shared between miR-148a-3p and miR-221-3p, which we found overexpressed in plasma of female HCWs. CDK1B and BCL2L11 emerge as the exclusive, strongly validated common targets between hsa-miR-148a-3p and miR-221-3p, that are components of the cell cycle and apoptosis pathways.
3. Discussion
The present study pointed out a sex-specific response to the two-dose requirement of COVID-19 vaccination in a cohort of healthcare workers. Female individuals showed higher levels of anti-S/RBD antibodies about 71 days apart of vaccination, compared to males. This finding aligns with our previous research, where we reported a stronger antibody response in females than in males following COVID-19 mRNA vaccine [5].
Age-related differences in antibody response to vaccination were not observed in either the whole population or sex-disaggregated analysis. This is likely because most of the population was under 45 years old.
The molecular basis of sex disparity in immune responses includes genetic, hormonal, and epigenetic mechanisms. Among the latter, DNA methylation patterns and microRNAs regulation of gene expression are reported as potential mechanisms of sex differences in pathophysiology and immune responses. In the present study, we investigated a profile of six microRNAs selected for their role in immune cells regulation, and as being encoded on the X chromosome and/or being regulated by sex hormones.
MicroRNAs have been previously investigated as potential markers of vaccine response. The temporal and vaccine-specific expression of microRNAs has been connected to the effectiveness of vaccines or the adverse events linked to vaccination. Atherton et al. considered microRNAs as promising biomarkers that could provide crucial information for vaccine development after identifying microRNA patterns specific to several vaccine types [7]. MicroRNAs found in extracellular vesicles are essential tools for enhancing vaccine efficacy and serving as indicators of immunological response and adverse outcomes after vaccination, according to Oshiumi H. [23]. Furthermore, small regulatory microRNAs crucially regulate cytokines and other significant immune mediators’ production and functions [23,25]. Moreover, correlation between B-cell specific microRNAs and neutralizing antibody response intensity after measles vaccination has been reported, suggesting that B cell-specific miRNAs may serve as useful predictive biomarkers of vaccine humoral immune response [15,17].
Although it is known that microRNAs are expressed differently in males and females, the influence and modification of the immunological response to vaccinations by microRNAs in a sex-specific manner have not been well-documented. This sex-biased microRNA expression has been observed in both invertebrates and higher species and is attributed to hormonal and genetic differences between the sexes [26,27]. Specifically, sex steroid hormones, such as estrogens, play a role in regulating miRNA expression [28]. Additionally, there is a difference in the density of miRNAs encoded in sex chromosomes, with the X chromosome containing a significantly higher density of miRNAs than the Y chromosome [12]. MicroRNAs are crucial regulators of gene expression and cell functions, suggesting that females may have more precise regulation of genes involved in immune responses.
In the present study, we were able to identify a sex-specific miRNA signature of response to COVID-19 vaccination with two doses of mRNA vaccine.
We further detected that levels of circulating miR-148a-3p was associated with higher levels of anti-S response in females more significantly than in males.
miR-148a-3p, encoded on chromosome 7, has been reported to regulate antigen presentation of TLR-triggered dendritic cells and to negatively regulate innate immune response. Considering that dendritic cells are crucial for activation of B-cells through naive T-cell priming, it is conceivable that miR-148a could affect the antibody levels after vaccination [29]. Furthermore, Pracht et al. found that miR-148a-3p promoted differentiation of B cells to antibody secreting plasma cells (PC) and modulated PC survival [30]. miR-148a has been reported to be inducible by androgen in prostate cancer cell lines and to be inhibited by estradiol in breast cancer cells, highlighting its sex-specific role in carcinogenesis [31,32]
Here we found that miR-148a-3p, overexpressed in vaccinated female HCW and positively associated to higher antibody levels in response to vaccination, was involved in the regulation of several pathways, identified by the Mienturnet enrichment analysis, including genes of the innate immune responses to several viral infections, of the cell cycle and apoptosis, of the chemokines and cytokines signaling pathways, as well as antigen presentation and processing, suggesting its multiple regulatory functions.
It should be considered that the same miRNA can have distinct effects depending on the cell type. For miRNA-mediated regulation to be effective, its target genes must be expressed at appropriate levels in the given cell type. If a target gene is not highly expressed in a particular cell type, the miRNA may not have a noticeable effect on it. Thus, the impact of miRNA regulation could be amplified in certain cell types where its targets are more abundant. Therefore, understanding the precise mechanisms by which miR-148a-3p exerts its effect on the humoral response to the COVID-19 vaccine requires a more detailed and in-depth evaluation.
Interestingly, miR-221-3p, miR-155-5p, and miR-150-5p expression levels were also significantly different in male and female HCWs vaccinated with COVID-19 mRNA vaccines. Specifically, miR-221-3p was overexpressed in vaccinated females compared to the unvaccinated and to vaccinated males. miR-155-5p was overexpressed in vaccinated males compared to females. Although we could not detect a statistically significant association between the variation of miR-221-3p and miR150-5p upon vaccination and antibody levels, it is intriguing that all these microRNAs have a role in immune function. MiR-221-3p has been shown to modulate inflammatory responses in M1 and M2 macrophages activated by TLR4 [33], and its dysregulated expression has been reported to associate to severe COVID-19 [34]. Specifically, miR-221-3p targets molecules of the inflammatory pathways, such as TLRs, NF-kB, cytokines, and chemokines. Thus, the higher miR-221-3p level in vaccinated females may conceivably affect activation of the innate immune response to vaccine antigens.
miR-148a-3-p and miR-221-5-p, overexpressed in female plasma, target two common genes, BCL2L11 and CDKN1B, that are involved in apoptosis and cell cycle regulation, and are reported regulate the immune response and antibody production. We can hypothesize that, because of the miRNAs upregulation in females, the targets will be downregulated with subsequent activation of B cell proliferation and antibodies production more actively in female than in male vaccinees.
MiR-155-5p is one of the most studied microRNAs. It is highly expressed in hematopoietic cells, where it plays a key role in inflammation and immunity [22,35]. Its targets are involved in the regulation of inflammatory pathways, the response to viral infections, and Toll-like receptor (TLR) signaling. It acts as a key regulator of both innate and adaptive immunity, influencing various immune cell types, including T cells, B cells, macrophages, and dendritic cells. miR-155 helps amplify inflammatory signals during immune responses, such as in infections or autoimmune diseases. For example, it targets molecules like SOCS1 (Suppressor of Cytokine Signaling 1) and PTEN, which are negative regulators of inflammatory signaling pathways. By downregulating these suppressors, MiR-155-5p promotes the activation of inflammatory cytokines like TNF-α, IL-6, and IL-1β.
So far, a limited number of studies have documented variations in microRNAs after vaccination [13,14,15,16,17,18]. In line with our results, Miyashita et al. [13] were able to identify that miR-148a was involved in antibody production in response to the COVID-19 mRNA vaccine, and miR-92a-2-5p was related to cytokines and adverse reactions to vaccination. An association between miRNA expression and local adverse reactions to influenza vaccination has also been reported [24], with levels of serum vesicle miR-451a negatively correlated to inflammatory cytokines after seasonal influenza vaccination. The aforementioned studies analyzed microRNAs before vaccination, making them predictive of the response to vaccination. In the present study, we analyzed miRNA levels in the plasma 71 days post-second dose of the COVID-19 vaccine, providing an observational report on microRNA expression associated with the vaccine response. This can be valuable as it provides evidence of miRNA associated with antibody production in response to vaccination, but it is also limited by the lack of comparative evaluation before and after vaccination. Another limitation of this study is the small sample size, particularly as we disaggregated the analysis by sex. Nevertheless, we consider the analysis of data by sex to be a strength and original aspect of the study compared to previously published reports.
4. Materials and Methods
4.1. Population and Study Design
In this prospective, observational study, we recruited 128 healthcare workers, working at the Bambino Gesù Children Hospital in Rome, who received the 2-dose schedule of BNT162b COVID-19 vaccine in January-March 2021, and have never had a SARS-CoV-2 infection.
Plasma samples were collected from the recruited health care workers (HCWs) between 50 and 100 days (median 71 days) after second dose of the COVID-19 vaccine, concomitantly with periodic health surveillance, and used to evaluate plasma levels of antibodies against the SARS-CoV-2 spike/RBD antigen (anti-S/RBD), by a commercial serologic assay (AdviseDx SARS-CoV-2 immunoglobulin (Ig)G II assay, ARCHITECT®, i2000sr Abbott Diagnostics), according to the manufacturers’ instructions.
Demographic characteristics of the enrolled subjects are reported in Table 1
This study was conducted after obtaining ethical approval from ISS (AOO-ISS 09/05/2021–0017778); all participants provided written informed consent.
Before enrollment, all study participants provided written informed consent.
4.2. RNA Extraction, cDNA Synthesis and Quantitative Analysis by qRT-PCR of the Selected microRNAs
The level of selected circulating miRNAs was evaluated in plasma samples from a subgroup of 86 HCWs (43 males, 43 females).
Total RNA from plasma samples was extracted using a “Plasma/Serum Isolation kit” (Norgen), according to the manufacturer’s instructions. miRNAs were reverse transcribed by “TaqMan™ Advanced miRNA cDNA Synthesis Kit” (Thermo Fisher TM Waltham, MA, USA). Quantitative expression of the selected microRNAs was carried out by Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) using specific inventoried Advanced TaqMan MicroRNA Assays (Thermo Fisher TM Waltham, MA, USA). The reactions were run in a Quant Studio 12K Flex (Thermo Fisher TM Waltham, MA, USA) qPCR Real-Time PCR machine. Fold expression changes of miRNAs, relative to sex-matched controls who didn’t receive the COVID-19 vaccine, were determined by the 2−ΔΔCt method, after normalization to the spike-in cel-miR-39. A 1.5-fold change increase of miRNA level or a 0.5-fold change decrease were considered significant variations in the relative quantitation of microRNAs between vaccinated and unvaccinated subjects.
4.3. Statistical Analyses
Anti-S/RBD concentrations (AU/ml) were reported as geometric means with their confidence intervals (95 % CI).
When the variables of interest were not normally distributed, analyses were performed after a natural logarithmic transformation, and when comparing groups, we applied t-tests; whilst we performed Mann-Whitney non-parametric tests for original data. A p-value of <0.05 was considered statistically significant.
In particular, for each miRNA, we created a categorical variable choosing the value of 1.5-fold change (FC) between vaccinated and unvaccinated controls, which is considered the biological significance threshold, as a cut-off.
To study the effect of miRNAs on anti-S/RBD antibodies, we applied a simple regression model considering the natural logarithm (ln) of anti-S/RBD antibody level as the response variable and miRNAs fold change as a categorical covariate; we showed LS-means (Least-Square means) of the response variable by miR cutoff, overall and by sex. In particular, we applied the Laplace approximation to fit the regression models.
Statistical analysis was performed with SAS version 9.4 and GraphPad Prism (version 5.0).
4.4. MicroRNA-Target Interaction Network
The miRNA-target interactions were analyzed by using MIENTURNET (MicroRNA ENrichment TURned NETwork; http://userver.bio.uniroma1.it/apps/mienturnet/) [19], a web application that enables users to apply filters based on evidence categories from the miRTarBase database, distinguishing between ‘Strong’ experimental methods (e.g., Luciferase assay, Western), ‘Weak’ evidence (e.g., CLIP), or a combination of both (‘Strong and Weak’). In this work, we have used ‘Strong’ experimental method and set the threshold for the False Discovery Rate (FDR) as 0.05.
4.5. Functional Enrichment Analysis
The functional enrichment analysis was performed by querying the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway through the MIENTURNET web tool; p-values were adjusted with the Benjamini–Hochberg method, and a threshold (FDR) equal to 0.05 was set to identify functional annotations significantly enriched among genes of the input list [20].
5. Conclusions
In conclusion, our study offers an original contribution to understanding microRNAs associated with the immune response to COVID-19 vaccination, highlighting the potential role of specific microRNAs in antibody production. Understanding these associations could not only enhance our knowledge of the immune response to COVID-19 vaccines but also contribute to pave the way for more personalized, sex-specific approaches to vaccination, ultimately enhancing the effectiveness of vaccines in diverse populations and possibly reducing the adverse events to vaccinations. Although limited by a small sample size and the absence of pre-vaccination data, the post-vaccination analysis and sex-stratified approach, which complement previous predictive studies, represent innovative and valuable aspects of the work. Future longitudinal studies with larger cohorts will be essential to confirm and extend these findings.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1. Association between miR-148a-3p fold change and anti-S/RBD level. Simple regression models applied using ln (Anti-S/RBD antibody) levels as response variable and miR-148a-3p (>= 1.5 vs. <1.5 fold change versus unvaccinated control) as covariate; LS-means (Least-Squares means) with its 95% CI are shown in each panel: A )overall study population; test for difference = + 0.370, p-val =0.014; B) female HCWs; test for difference = + 0.547 p-val =0.044; C) male HCWs; test for difference = + 0.337 p-val =0.102. P-values were calculated by Mann–Whitney U test. * p < 0.05; Table S1. Pathway enrichment results for miR-148a-3p target genes; Table S2. Pathway enrichment results for miR-221-3p target genes; Table S3. Pathway enrichment results for miR-155-5p target genes.
Author Contributions
Conceptualization, A.R.; methodology, M.D.; formal analysis, S.A. and M.D.; investigation, S.A. and E.I.; resources P.T., N.V., R.C. and S.Z.; data curation P.T. and N.V.; writing—original draft preparation NVand S.A.; writing—review and editing, A.R. and P.T.; visualization, S.A.; supervision, A.R.; project administration, A.R.; funding acquisition, A.R., P.T. and S.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by BRIC-INAIL- ID 35/2022 and BRIC-INAIL ID 27/2019 fundings to AR and PT; by the European Union funding within the Ministero dell’Università e della Ricerca Piano Nazionale di Ripresa e Resilienza Extended Partnership initiative on Emerging Infectious Diseases (project No. PE00000007, INF-ACT).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by Ethics Committee of ISS (AOO-ISS 09/05/2021–0017778) on 2021-05-09.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available upon request from the corresponding authors.
Acknowledgments
The authors would like to thank Dr. Reparata Rosa Di Prinzio, Dr Maria Rosaria Vinci, Dr. Christian Albano, IRCCS Bambino Gesù Children’s Hospital, Rome, for antibody determination, collection of plasma samples, anagraphic and anamnestic data from HCWs. The authors are grateful to Fabiola and Daniela Diamanti for their qualified technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gote, V.; Bolla, P.K.; Kommineni, N.; Butreddy, A.; Nukala, P.K.; Palakurthi, S.S.; Khan, W. A Comprehensive Review of mRNA Vaccines. Int. J. Mol. Sci. 2023, 24, 2700. [Google Scholar] [CrossRef]
- Bachmann, M.; Gültekin, N; Stanga, Z, Fehr, J. S.; Ülgür, I.I.; Schlagenhauf, P. Disparities in response to mRNA SARS-CoV-2 vaccines according to sex and age: A systematic review. New Microbes New Infect. 2024, 63, 101551. [Google Scholar] [CrossRef]
- Nam, M.; Yun, S.G.; Kim, S.; Kim, C.G.; Cha, J.H.; Lee, C.; Kang, S.; Park, S.G.; Kim, S.B.; Lee, K.; et al. Humoral and Cellular Immune Responses to Vector, Mix-and-Match, or mRNA Vaccines against SARS-CoV-2 and the Relationship between the Two Immune Responses. Microbiol Spectr. 2022, 10, e02495–21. [Google Scholar] [CrossRef]
- Gu, Y.; Shunmuganathan, B.; Qian, X.; Gupta, R.; Tan, R.S.W.; Kozma, M.; Purushotorman, K.; Murali, T.M.; Tan, N.Y.J.; Preiser, P.R.; et al. Employment of a high throughput functional assay to define the critical factors that influence vaccine induced cross-variant neutralizing antibodies for SARS-CoV-2. Sci Rep. 2023, 13, 21810. [Google Scholar] [CrossRef] [PubMed]
- Anticoli, S.; Dorrucci, M.; Iessi, E.; Chiarotti, F.; Di Prinzio, R. R.; Vinci, M. R.; Zaffina, S.; Puro, V.; Colavita, F.; Mizzoni, K.; et al. Association between sex hormones and anti-S/RBD antibody responses to COVID-19 vaccines in healthcare workers. Hum Vaccin Immunother. 2023, 19, 2273697. [Google Scholar] [CrossRef]
- Van Tilbeurgh, M.; Lemdani, K.; Beignon, A.S.; Chapon, C.; Tchitchek, N.; Cheraitia, L.; Marcos Lopez, E.; Pascal, Q.; Le Grand, R.; Maisonnasse, P.; et al. Predictive Markers of Immunogenicity and Efficacy for Human Vaccines. Vaccines 2021, 9, 579. [Google Scholar] [CrossRef]
- Atherton, L.J.; Jorquera, P.A.; Bakre, A.A.; Tripp, R.A. Determining Immune and miRNA Biomarkers Related to Respiratory Syncytial Virus (RSV) Vaccine Types. Front Immunol. 2019, 10, 2323. [Google Scholar] [CrossRef]
- Lin, Y.-P.; Hsieh, Y.-S.; Cheng, M.-H.; Shen, C.-F.; Shen, C.-J.; Cheng, C.-M. Using MicroRNA Arrays as a Tool to Evaluate COVID-19 Vaccine Efficacy. Vaccines 2022, 10, 1681. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front Endocrinol (Lausanne) 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xu, X.; Xiao, L.; Wang, L.; Qiang, S. The Role of microRNA in the Inflammatory Response of Wound Healing. Front Immunol. 2022, 13, 852419. [Google Scholar] [CrossRef] [PubMed]
- Gaál, Z. Role of microRNAs in Immune Regulation with Translational and Clinical Applications. Int J Mol Sci. 2024, 25, 1942. [Google Scholar] [CrossRef]
- Di Palo, A.; Siniscalchi, C.; Salerno, M.; Russo, A.; Gravholt, C.H.; Potenza, N. What microRNAs could tell us about the human X chromosome. Cell Mol Life Sci 2020, 77, 4069–4080. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, Y.; Yoshida, T.; Takagi, Y.; Tsukamoto, H.; Takashima, K.; Kouwaki, T.; Makino, K.; Fukushima, S.; Nakamura, K.; Oshiumi, H. Circulating extracellular vesicle microRNAs associated with adverse reactions, proinflammatory cytokine, and antibody production after COVID-19 vaccination. NPJ Vaccines 2022, 7, 16. [Google Scholar] [CrossRef]
- de Candia, P.; Torri, A.; Gorletta, T.; Fedeli, M.; Bulgheroni, E.; Cheroni, C.; Marabita, F.; Crosti, M.; Moro, M.; Pariani, E.; et al. Intracellular modulation, extracellular disposal and serum increase of MiR-150 mark lymphocyte activation. PLoS One 2013, 8, e75348. [Google Scholar] [CrossRef] [PubMed]
- Haralambieva, I.H.; Ratishvili, T.; Goergen, K.M.; Grill, D.E.; Simon, W.L.; Chen, J.; Ovsyannikova, I.G.; Poland, G.A.; Kennedy, R.B. Effect of lymphocyte miRNA expression on influenza vaccine-induced immunity. Vaccine 2025, 55, 127023. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Chen, S.; Liu, L.; Zhao, Y.; Lin, W.; Ni, J. Increased serum microRNA-155 level associated with nonresponsiveness to hepatitis B vaccine. Clin Vaccine Immunol. 2013, 20, 1089–1091. [Google Scholar] [CrossRef]
- Haralambieva, I.H.; Kennedy, R.B.; Simon, W.L.; Goergen, K.M.; Grill, D.E.; Ovsyannikova, I.G.; Poland, G.A. Differential miRNA expression in B cells is associated with inter-individual differences in humoral immune response to measles vaccination. PLoS One 2018, 13, e0191812. [Google Scholar] [CrossRef]
- Vianello, E.; Persson, J.; Andersson, B.; van Veen, S.; Dias, T.L.; Santoro, F.; Östensson, M.; Obudulu, O.; Agbajogu, C.; Torkzadeh, S. Global blood miRNA profiling unravels early signatures of immunogenicity of Ebola vaccine rVSVΔG-ZEBOV-GP. iScience 2023, 26, 108574. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Licursi, V.; Conte, F.; Fiscon, G.; Paci, P. MIENTURNET: an interactive web tool for microRNA-target enrichment and network-based analysis. BMC Bioinformatics 2019, 20, 545. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Kim, M.S. Exposure to a mixture of heavy metals induces cognitive impairment: Genes and microRNAs involved. Toxicology 2022, 471, 153164. [Google Scholar] [CrossRef]
- Wigton, E.J.; Mikami, Y.; McMonigle, R.J.; Castellanos, C.A.; Wade-Vallance, A.K.; Zhou, S.K.; Kageyama, R.; Litterman, A.; Roy, S.; Kitamura, D.; et al. MicroRNA-directed pathway discovery elucidates an miR-221/222-mediated regulatory circuit in class switch recombination. J Exp Med. 2021, 218, e20201422. [Google Scholar] [CrossRef]
- Jafarzadeh, A.; Naseri, A.; Shojaie, L.; Nemati, M.; Jafarzadeh, S.; Bannazadeh Baghi, H.; Hamblin, M.R.; Akhlagh, S.A.; Mirzaei, H. MicroRNA-155 and antiviral immune responses. Int Immunopharmacol. 2021, 101(Pt A), 108188. [Google Scholar] [CrossRef]
- Oshiumi, H. Circulating Extracellular Vesicles Carry Immune Regulatory miRNAs and Regulate Vaccine Efficacy and Local Inflammatory Response After Vaccination. Front Immunol. 2021, 12, 685344. [Google Scholar] [CrossRef]
- Miyashita, Y.; Ishikawa, K.; Fukushima, Y.; Kouwaki, T.; Nakamura, K.; Oshiumi, H. Immune-regulatory microRNA expression levels within circulating extracellular vesicles correspond with the appearance of local symptoms after seasonal flu vaccination. PLoS ONE 2019, 14, e0219510. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, C.M.; Podyminogin, R.L.; Navarro, G.; Zhao, G.W.; Askovich, P.S.; Weiss, M.J.; Aderem, A. miR-451 Regulates Dendritic Cell Cytokine Responses to Influenza Infection. J Immunol. 2012, 189, 5965–5975. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Eghbali, M. Influence of sex differences on microRNA gene regulation in disease. Biol Sex Differ. [CrossRef]
- Zhong, B.; Cui, C.; Cui, Q. Identification and Analysis of Sex-Biased MicroRNAs in Human Diseases. Genes 2023, 14, 1688. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M. miRNAs regulated by estrogens, tamoxifen, and endocrine disruptors and their downstream gene targets. Mol Cell Endocrinol. 2015, 418 Pt 3(0 3), 273–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhan, Z.; Xu, L.; Ma, F.; Li, D.; Guo, Z.; Li, N.; Cao, X. MicroRNA-148/152 Impair Innate Response and Antigen Presentation of TLR-Triggered Dendritic Cells by Targeting Camkiiα. J Immunol. 2010, 185, 7244–7251. [Google Scholar] [CrossRef]
- Pracht, K.; Meinzinger, J.; Schulz, S.R.; Daum, P.; Côrte-Real, J.; Hauke, M.; Roth, E.; Kindermann, D.; Mielenz, D.; Schuh, W.; et al. miR-148a controls metabolic programming and survival of mature CD19-negative plasma cells in mice. Eur J Immunol. 2021, 51, 1089–1109. [Google Scholar] [CrossRef]
- Murata, T.; Takayama, K.; Katayama, S.; Urano, T.; Horie-Inoue, K.; Ikeda, K.; Takahashi, S.; Kawazu, C.; Hasegawa, A.; Ouchi, Y.; et al. miR-148a is an androgen-responsive microRNA that promotes LNCaP prostate cell growth by repressing its target CAND1 expression. Prostate Cancer Prostatic Dis. [CrossRef]
- Tao, S.; He, H.; Chen, Q.; Yue, W. GPER mediated estradiol reduces miR-148a to promote HLA-G expression in breast cancer. Biochem Biophys Res Commun. 2014, 451, 74–78. [Google Scholar] [CrossRef]
- Quero, L.; Tiaden, A.N.; Hanser, E.; Roux, J.; Laski, A.; Hall, J.; Kyburz, D. miR-221-3p Drives the Shift of M2-Macrophages to a Pro-Inflammatory Function by Suppressing JAK3/STAT3 Activation. Front Immunol. 2020, 10, 3087. [Google Scholar] [CrossRef] [PubMed]
- Gaytán-Pacheco, N.; Ibáñez-Salazar, A.; Herrera-Van Oostdam, A.S.; Oropeza-Valdez, J.J.; Magaña-Aquino, M.; Adrián López, J.; Monárrez-Espino, J.; López-Hernández, Y. miR-146a, miR-221, and miR-155 are Involved in Inflammatory Immune Response in Severe COVID-19 Patients. Diagnostics (Basel). 2022, 13, 133. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hu, J.; Huang, S.; Liu, X.; Zhang, Y.; Wei, S.; Hu, X. miR-155: An Important Role in Inflammation Response. J Immunol Res. 2022, 2022, 7437281. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Relative quantitation of the selected microRNAs measured by qRT-PCR. Plasma level of the selected microRNAs in COVID-19 vaccinated HCWs: A) in overall under study population; the values of fold change for each sample are shown relative to the mean CT values of the unvaccinated control population, shown by dotted horizontal bar; B) in male and female HCWs; the values of fold change for each sample are shown relative to the mean CT values of the unvaccinated and sex-matched control. The mean ± SD values are shown. Statistically significant differences between sexes were evaluated by the Mann-Whitney U test. *** p < 0.001. Obtained with GraphPad Prism (version 5.0).
Figure 1.
Relative quantitation of the selected microRNAs measured by qRT-PCR. Plasma level of the selected microRNAs in COVID-19 vaccinated HCWs: A) in overall under study population; the values of fold change for each sample are shown relative to the mean CT values of the unvaccinated control population, shown by dotted horizontal bar; B) in male and female HCWs; the values of fold change for each sample are shown relative to the mean CT values of the unvaccinated and sex-matched control. The mean ± SD values are shown. Statistically significant differences between sexes were evaluated by the Mann-Whitney U test. *** p < 0.001. Obtained with GraphPad Prism (version 5.0).
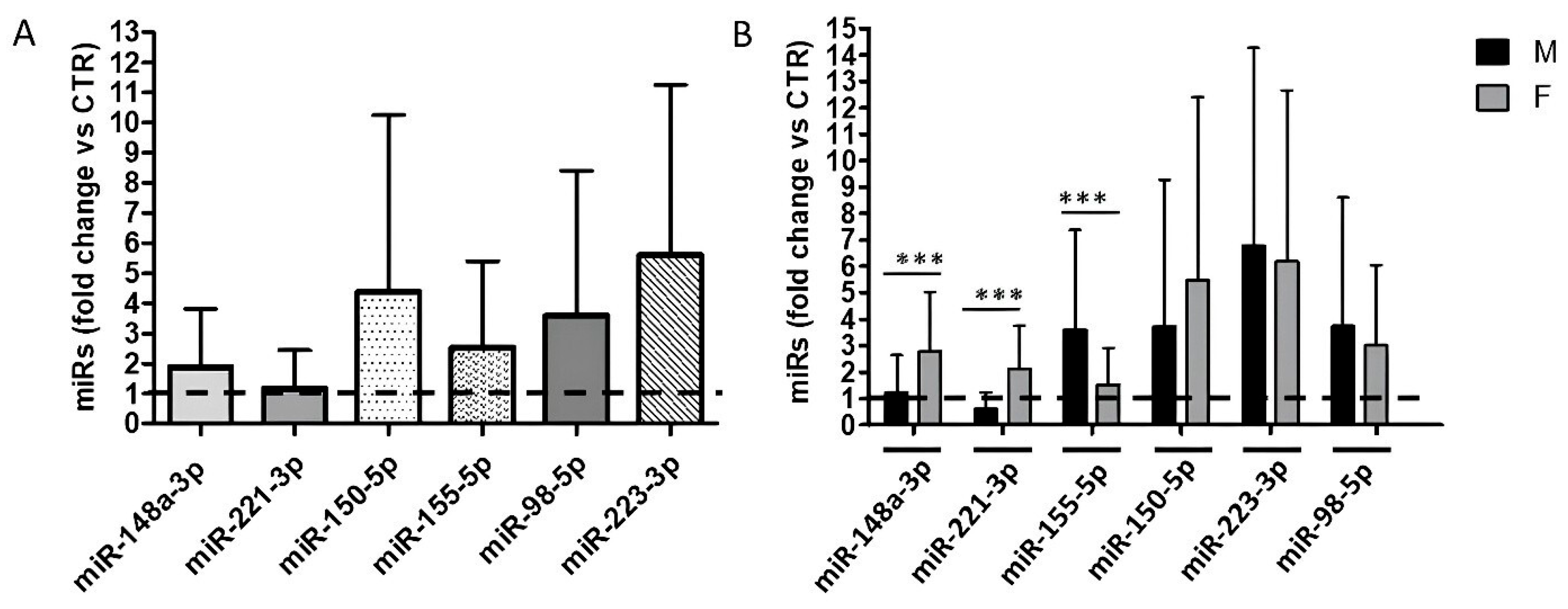
Figure 2.
Association between circulating microRNA fold changes and anti-S/RBD antibody levels after vaccination. Box-plots of Ln (Anti-S/RBD antibody levels) distributions according to the two groups with (A-C) ≥1.5 and < 1.5 miR-148a-3p fold changes between vaccinated and unvaccinated total HCWs (A) and unvaccinated total HCWs (D), female (E) and male HCWs (F); (G-I) ≥1.5 and < 1.5 miR-155-5p fold changes between vaccinated and unvaccinated total HCWs (G), female (H) and male HCWs (I). The boxes represent the interquartile ranges (IQR): the medians are the line inside, the I quartile is the bottom edge, and III quartile is the top edge; the lines that extend from each box indicate the range of values that are outside of the IQR, and that do not exceed 1.5-fold the IQR. Differences between sexes were tested by t-test. * p < 0.05.
Figure 2.
Association between circulating microRNA fold changes and anti-S/RBD antibody levels after vaccination. Box-plots of Ln (Anti-S/RBD antibody levels) distributions according to the two groups with (A-C) ≥1.5 and < 1.5 miR-148a-3p fold changes between vaccinated and unvaccinated total HCWs (A) and unvaccinated total HCWs (D), female (E) and male HCWs (F); (G-I) ≥1.5 and < 1.5 miR-155-5p fold changes between vaccinated and unvaccinated total HCWs (G), female (H) and male HCWs (I). The boxes represent the interquartile ranges (IQR): the medians are the line inside, the I quartile is the bottom edge, and III quartile is the top edge; the lines that extend from each box indicate the range of values that are outside of the IQR, and that do not exceed 1.5-fold the IQR. Differences between sexes were tested by t-test. * p < 0.05.
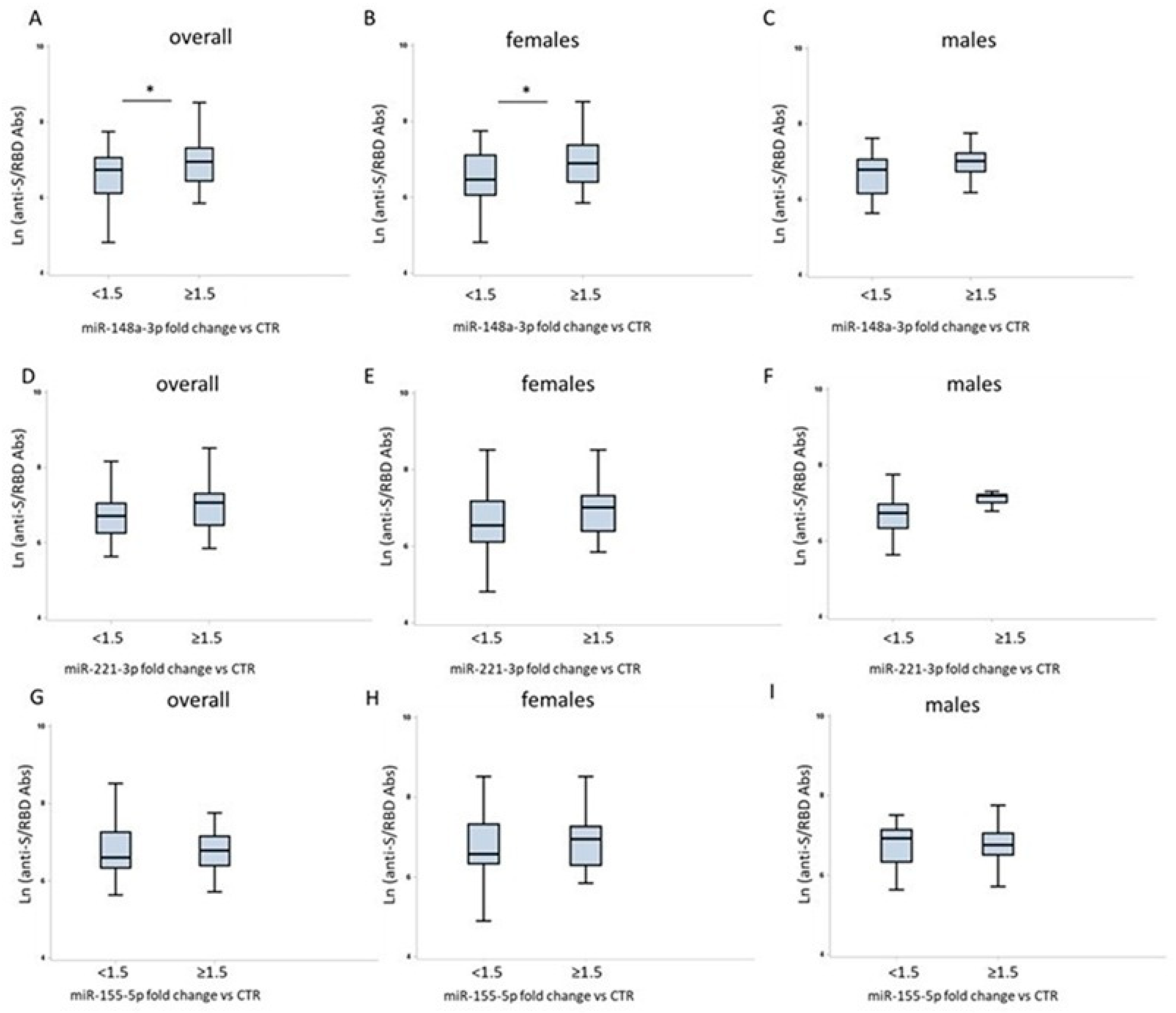
Figure 3.
Network analysis and target mRNAs shared between miR-148a-3p and miR-221-3p. The miRNA-target interactions were analyzed by using MIENTURNET. Only targets with “strong interaction” have been considered.
Figure 3.
Network analysis and target mRNAs shared between miR-148a-3p and miR-221-3p. The miRNA-target interactions were analyzed by using MIENTURNET. Only targets with “strong interaction” have been considered.
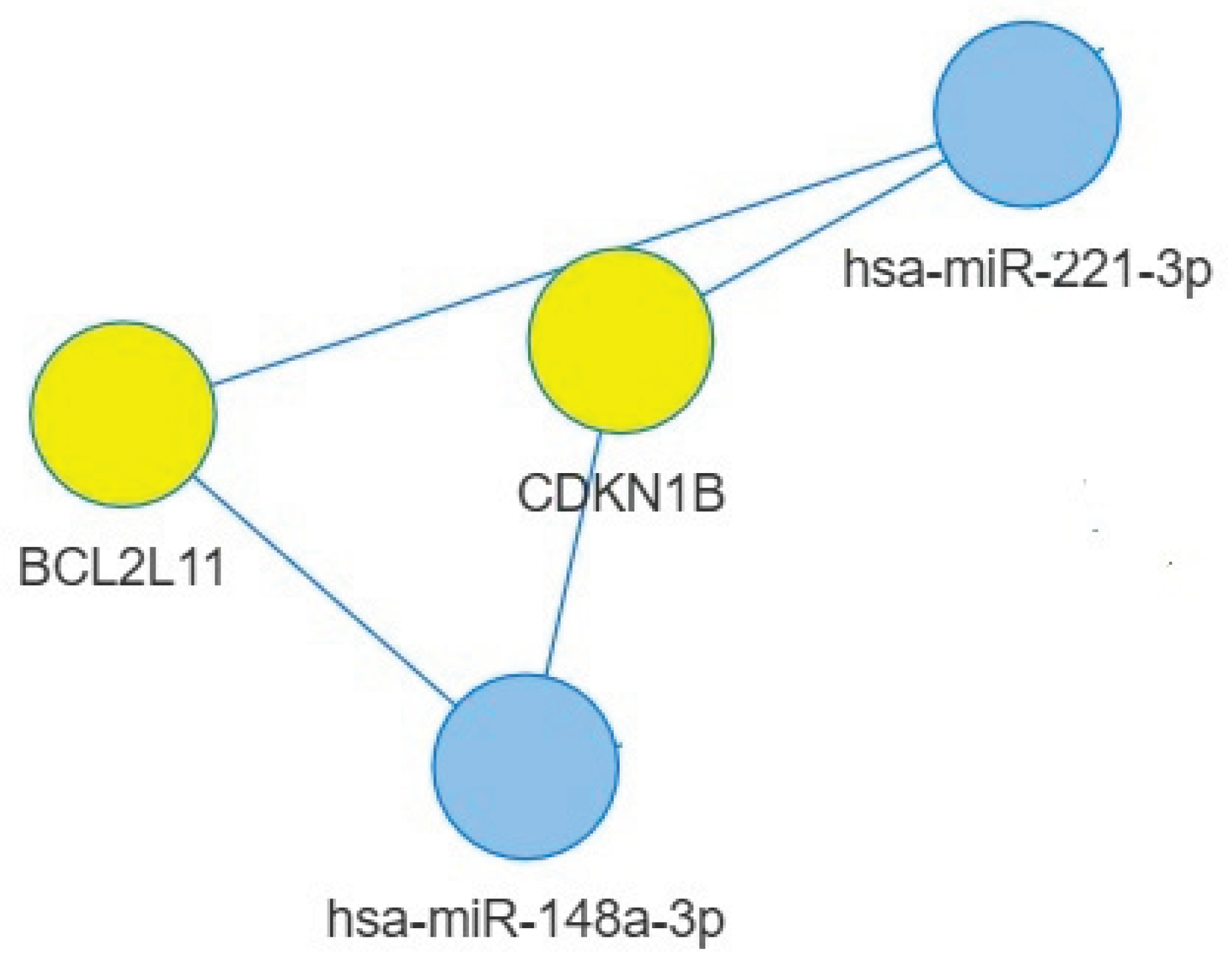

Table 1.
Demographic characteristics of the study population.
| Study population | Males | Females | |
| n = 128 | 47 (36.7 %) | 81 (63.3 %) | |
| Age (years), median (IQR); (range) |
45 (36–54); (23–72) |
42 (36–51); (26–72) |
47 (35.5–54.5); (23–64) |
| age groups | |||
| 23–45 years (n) | 67 (52.3 %) | 29 (43.3 %) | 38 (56.7 %) |
| 46–72 years (n) | 61 (47.7 %) | 18 (29.5 %) | 43 (70.5 %) |
| Interval (days) median (IQR); (range) |
71 (70-79); (55-100) |
71 (70-81); (55-100) |
71 (69-78); (56-100) |
IQR: interquartile range; Interval: time interval between the second vaccine dose and anti-S/RBD testing.
Table 2.
Anti-S/RBD concentrations by sex and age. Geometric means (GMT) of anti-S/RBD levels (AU/mL) after the second dose of COVID-19 vaccine (median time interval: 71 days); p-values refer to the comparison among groups.
Table 2.
Anti-S/RBD concentrations by sex and age. Geometric means (GMT) of anti-S/RBD levels (AU/mL) after the second dose of COVID-19 vaccine (median time interval: 71 days); p-values refer to the comparison among groups.
|
Anti-S/RBD titer (AU/L) GMT (95% CI) |
p-value | |
| All subjects | 921.5 | |
| (790.7-1074) | ||
| F | 1069 (925.8-1234) |
0.0123 |
| M | 713,9 | |
| (509.1-1001) | ||
| Age | ||
| ≤ 45 years | 1031 | 0.42 |
| (925.8-1234) | ||
| > 45 years | 713.9 | |
| (509.1-1001) | ||
| Sex and age | ||
| F≤ 45 years | 1031 | 0.37 |
| 870.7-1220 | ||
| F> 45 years | 978.9 | |
| 830.9-1153 | ||
| M≤ 45 years | 862.7 | 0.48 |
| (696.5-1068) | ||
| M> 45 years | 526.2 | |
| (225.6-1227) | ||
Table 3.
KEGG pathways enrichment analysis for miR-148a-3p. Statistically significant pathways (FDR ≤ 0.05) involved immune response modulation and/or frequently altered in response to viral infections.
Table 3.
KEGG pathways enrichment analysis for miR-148a-3p. Statistically significant pathways (FDR ≤ 0.05) involved immune response modulation and/or frequently altered in response to viral infections.
| Description | KEGG ID | p-value | FDR * | Genes |
| Epstein-Barr virus infection | hsa05169 | 8,74364E-08 | 2,49E-06 | HLA-G/BCL2/CDKN1B/RUNX3/ BCL2L11/PDIA3/STAT3/IKBKB/BAX |
| FoxO signaling pathway | hsa04068 | 8,86186E-07 | 1,37E-05 | IRS1/CDKN1B/BCL2L11/S1PR1/STAT3/ TGFB2/IKBKB |
| Human T-cell leukemia virus 1 infection | hsa05166 | 2,79378E-05 | 0,000259 | HLA-G/MMP7/SMAD2/TGFB2/IKBKB/ BAX/NRP1 |
| Human papillomavirus infection | hsa05165 | 4,59073E-05 | 0,000387 | HLA-G/WNT10B/CDKN1B/ITGB8/ITGA5/ WNT1/IKBKB/BAX |
| TGF-beta signaling pathway | hsa04350 | 7,06962E-05 | 0,000504 | TGIF2/ACVR1/ROCK1/SMAD2/TGFB2 |
| PI3K-Akt signaling pathway | hsa04151 | 8,29241E-05 | 0,000517 | IRS1/BCL2/CDKN1B/ITGB8/ITGA5/MET/ BCL2L11/IKBKB |
| Herpes simplex virus 1 infection | hsa05168 | 8,36848E-05 | 0,000517 | HLA-G/BCL2/ITGA5/PDIA3/IKBKB/BAX |
| MAPK signaling pathway | hsa04010 | 0,000177249 | 0,000912 | RPS6KA5/CDC25B/MAP3K4/MET/MAP3K9/ TGFB2/IKBKB |
| Measles | hsa05162 | 0,000232811 | 0,001135 | BCL2/CDKN1B/STAT3/IKBKB/BAX |
| Human cytomegalovirus infection | hsa05163 | 0,000274049 | 0,001156 | HLA-G/ROCK1/PDIA3/STAT3/IKBKB/BAX |
| Apoptosis - multiple species | hsa04215 | 0,000288247 | 0,001161 | BCL2/BCL2L11/BAX |
| Hepatitis B | hsa05161 | 0,000486342 | 0,001733 | BCL2/STAT3/TGFB2/IKBKB/BAX |
| Apoptosis | hsa04210 | 0,00219604 | 0,005812 | BCL2/BCL2L11/IKBKB/BAX |
| Adipocytokine signaling pathway | hsa04920 | 0,002859899 | 0,006971 | IRS1/STAT3/IKBKB |
| p53 signaling pathway | hsa04115 | 0,003478888 | 0,007952 | BCL2/SERPINE1/BAX |
| Cellular senescence | hsa04218 | 0,003692136 | 0,007952 | HLA-G/SERPINE1/SMAD2/TGFB2 |
| Cell cycle | hsa04110 | 0,003777087 | 0,007952 | CDC25B/CDKN1B/SMAD2/TGFB2 |
| mTOR signaling pathway | hsa04150 | 0,003777087 | 0,007952 | IRS1/WNT10B/WNT1/IKBKB |
| Chemokine signaling pathway | hsa04062 | 0,00765295 | 0,014769 | VAV2/ROCK1/STAT3/IKBKB |
| Th17 cell differentiation | hsa04659 | 0,009837214 | 0,017193 | SMAD2/STAT3/IKBKB |
| Hormone signaling | hsa04081 | 0,011813777 | 0,020265 | CCKBR/IRS1/ACVR1/STAT3 |
| Regulation of actin cytoskeleton | hsa04810 | 0,014353241 | 0,024174 | ITGB8/VAV2/ITGA5/ROCK1 |
| Antigen processing and presentation | hsa04612 | 0,043197093 | 0,053352 | HLA-G/PDIA3 |
*FDR: False Discovery Rate.
Table 4.
KEGG pathways enrichment analysis for miR-221-3p targets. Statistically significant pathways (FDR ≤ 0.05) involved in immune response modulation and/or hormone signaling pathways.
Table 4.
KEGG pathways enrichment analysis for miR-221-3p targets. Statistically significant pathways (FDR ≤ 0.05) involved in immune response modulation and/or hormone signaling pathways.
| Description | KEGG ID | p-value | FDR* | Genes | |||
| FoxO signaling pathway | hsa04068 | 1,02E-07 | 4,28E-06 | CDKN1B/BCL2L11/FOXO3/TNFSF10/BNIP3/ PTEN/SIRT1/MDM2/PIK3R1 |
|||
| Cellular senescence | hsa04218 | 4,82E-06 | 4,84E-05 | FOXO3/PTEN/TP53/ETS1/RB1/SIRT1/MDM2/PIK3R1 | |||
| Apoptosis | hsa04210 | 1,87E-05 | 0,000141 | BCL2L11/BBC3/TNFSF10/FOS/TP53/APAF1/PIK3R1 | |||
| Mitophagy - animal | hsa04137 | 4,31E-05 | 0,000217 | FOXO3/BNIP3L/TBK1/BNIP3/TP53/BECN1 | |||
| Autophagy - animal | hsa04140 | 7,54E-05 | 0,000325 | DDIT4/TBK1/BNIP3/PTEN/RAB1A/PIK3R1/BECN1 | |||
| p53 signaling pathway | hsa04115 | 9,46E-05 | 0,000329 | BBC3/PTEN/TP53/APAF1/MDM2 | |||
| PI3K-Akt signaling pathway | hsa04151 | 0,00034 | 0,000906 | CDKN1B/BCL2L11/FOXO3/KIT/DDIT4/PTEN/ TP53/MDM2/PIK3R1 |
|||
| Cell cycle | hsa04110 | 0,000409 | 0,00103 | CDKN1B/CDKN1C/TP53/WEE1/RB1/MDM2 | |||
| TNF signaling pathway | hsa04668 | 0,000815 | 0,001891 | ICAM1/FOS/SELE/PIK3R1/SOCS3 | |||
| mTOR signaling pathway | hsa04150 | 0,002861 | 0,004887 | DDIT4/PTEN/DVL2/PIK3R1/GRB10 | |||
| Toll-like receptor signaling pathway | hsa04620 | 0,004558 | 0,007239 | TBK1/FOS/TICAM1/PIK3R1 | |||
| Leukocyte transendothelial migration | hsa04670 | 0,005682 | 0,008869 | ICAM1/PIK3R1/MMP2/CXCL12 | |||
| Thyroid hormone signaling pathway | hsa04919 | 0,006783 | 0,010066 | ESR1/TP53/MDM2/PIK3R1 | |||
| Estrogen signaling pathway | hsa04915 | 0,010646 | 0,014602 | FOS/ESR1/PIK3R1/MMP2 | |||
| JAK-STAT signaling pathway | hsa04630 | 0,020074 | 0,025239 | PIK3R1/STAT5A/SOCS3/SOCS1 | |||
| NF-kappa B signaling pathway | hsa04064 | 0,027183 | 0,031548 | ICAM1/TICAM1/CXCL12 | |||
| Th17 cell differentiation | hsa04659 | 0,029922 | 0,033859 | FOS/RUNX1/STAT5A | |||
| Chemokine signaling pathway | hsa04062 | 0,031384 | 0,034647 | FOXO3/PAK1/PIK3R1/CXCL12 | |||
|
MAPK signaling pathway |
hsa04010 | 0,037664 | 0,040358 | KIT/FOS/TP53/PAK1/STMN1 | |||
|
AMPK signaling pathway |
hsa04152 | 0,039796 | 0,040938 | FOXO3/SIRT1/PIK3R1 | |||
| T cell receptor signaling pathway | hsa04660 | 0,039796 | 0,040938 | FOS/PAK1/PIK3R1 | |||
*FDR: False Discovery Rate.
Table 5.
KEGG pathways enrichment for miR-155-5p. Statistically significant pathways (FDR ≤ 0.05) involved in immune response modulation and/or hormone signaling pathways and/or frequently altered in response to viral infections.
Table 5.
KEGG pathways enrichment for miR-155-5p. Statistically significant pathways (FDR ≤ 0.05) involved in immune response modulation and/or hormone signaling pathways and/or frequently altered in response to viral infections.
| Description | KEGG ID | p-value | FDR * | Genes |
| Hepatitis B | hsa05161 | 6,35E-13 | 2,25E-11 | TAB2/IKBKE/KRAS/JUN/FADD/MYD88/YWHAZ/ SMAD4/APAF1/SMAD3/CXCL8/NFKB1/E2F2/PIK3R1/FOS/MAPK14/MYC/MAPK13/STAT1/ CASP3 |
| Epstein-Barr virus infection | hsa05169 | 4,26E-11 | 1,13E-09 | TAB2/IKBKE/ICAM1/JUN/FADD/MYD88/RAC1/ SAP30L/MAP3K14/APAF1/CCND1/NFKB1/E2F2/PIK3R1/MAPK14/MYC/MAPK13/STAT1/ CCND2/CASP3 |
| Cellular senescence | hsa04218 | 2,48E-09 | 2,93E-08 | RHEB/FOXO3/KRAS/ETS1/SMAD2/SMAD3/CCND1/CXCL8/NFKB1/E2F2/PIK3R1/MAPK14/MYC/MAPK13/CCND2/PTEN |
| TNF signaling pathway | hsa04668 | 4,03E-09 | 3,89E-08 | TAB2/CEBPB/EDN1/ICAM1/SELE/JUN/FADD/MAP3K14/ NFKB1/PIK3R1/FOS/MAPK14/MAPK13/CASP3 |
| Toll-like receptor signaling pathway | hsa04620 | 1,26E-08 | 9,28E-08 | TAB2/IKBKE/JUN/FADD/MYD88/RAC1/CXCL8/NFKB1/PIK3R1/FOS/MAPK14/MAPK13/STAT1 |
|
IL-17 signaling pathway |
hsa04657 | 2,44E-08 | 1,62E-07 | TAB2/CEBPB/IKBKE/JUN/FADD/IL17RB/CXCL8/NFKB1/ FOS/MAPK14/MAPK13/CASP3 |
| Hepatitis C | hsa05160 | 1,68E-07 | 9,4E-07 | CLDN1/IKBKE/KRAS/FADD/YWHAZ/APAF1/CCND1/ NFKB1/E2F2/PIK3R1/MYC/STAT1/CASP3/ NR1H3 |
| Measles | hsa05162 | 2,35E-07 | 1,25E-06 | TAB2/IKBKE/JUN/FADD/MYD88/APAF1/CCND1/NFKB1/PIK3R1/FOS/STAT1/CCND2/CASP3 |
|
Th17 cell differentiation |
hsa04659 | 9,74E-07 | 4,14E-06 | IFNGR1/SMAD2/JUN/SMAD4/SMAD3/NFKB1/ FOS/MAPK14/HIF1A/MAPK13/STAT1 |
|
FoxO signaling pathway |
hsa04068 | 1,03E-06 | 4,23E-06 | FOXO3/KRAS/BCL6/SMAD4/GABARAPL1/SMAD3/ CCND1/PIK3R1/MAPK14/MAPK13/CCND2/ PTEN |
|
T cell receptor signaling pathway |
hsa04660 | 3E-06 | 1,1E-05 | RHOA/KRAS/JUN/CARD11/MAP3K14/PAK2/NFKB1/PIK3R1/FOS/MAPK14/MAPK13 |
|
MAPK signaling pathway |
hsa04010 | 4,1E-06 | 1,28E-05 | TAB2/FGF7/KRAS/CSF1R/JUN/MAP3K10/MYD88/RAC1/RAPGEF2/MAP3K14/PAK2/NFKB1/FOS/MAPK14/MYC/MAPK13/CASP3 |
| NOD-like receptor signaling pathway | hsa04621 | 7,68E-06 | 2,27E-05 | TAB2/PKN2/RHOA/IKBKE/JUN/FADD/MYD88/GABARAPL1/CXCL8/NFKB1/MAPK14/MAPK13/STAT1 |
| Cell cycle | hsa04110 | 3,56E-05 | 8,22E-05 | TRIP13/SMAD2/ANAPC16/YWHAZ/SMAD4/ SMAD3/WEE1/CCND1/E2F2/MYC/CCND2 |
| PI3K-Akt signaling pathway | hsa04151 | 4,75E-05 | 0,000108 | RHEB/PKN2/FGF7/FOXO3/KRAS/MYB/CSF1R/ NOS3/YWHAZ/RAC1/CCND1/NFKB1/PIK3R1/ MYC/RPTOR/CCND2/PTEN |
| Influenza A | hsa05164 | 8,15E-05 | 0,00017 | IKBKE/IFNGR1/ICAM1/FADD/MYD88/APAF1/ CXCL8/NFKB1/PIK3R1/STAT1/CASP3 |
|
B cell receptor signaling pathway |
hsa04662 | 8,48E-05 | 0,000172 | INPP5D/KRAS/JUN/CARD11/RAC1/NFKB1/ PIK3R1/FOS |
| Apoptosis | hsa04210 | 0,000274 | 0,000448 | KRAS/JUN/FADD/MAP3K14/APAF1/NFKB1/ PIK3R1/FOS/CASP3 |
| TGF-beta signaling pathway | hsa04350 | 0,000281 | 0,000453 | SMAD5/SMAD1/RHOA/SMAD2/SKI/SMAD4/ SMAD3/MYC |
| Coronavirus disease - COVID-19 | hsa05171 | 0,000346 | 0,000536 | TAB2/AGTR1/IKBKE/JUN/MYD88/CXCL8/NFKB1/ PIK3R1/FOS/MAPK14/MAPK13/STAT1 |
| Th1 and Th2 cell differentiation | hsa04658 | 0,000617 | 0,000887 | IFNGR1/JUN/NFKB1/FOS/MAPK14/MAPK13/ STAT1 |
| RIG-I-like receptor signaling pathway | hsa04622 | 0,000892 | 0,001193 | IKBKE/FADD/CXCL8/NFKB1/MAPK14/MAPK13 |
|
JAK-STAT signaling pathway |
hsa04630 | 0,001268 | 0,001607 | SOCS1/IFNGR1/IL13RA1/CCND1/PIK3R1/MYC/ SOCS6/STAT1/CCND2 |
|
NF-kappa B signaling pathway |
hsa04064 | 0,00127 | 0,001607 | TAB2/ICAM1/MYD88/CARD11/MAP3K14/ CXCL8/NFKB1 |
| Mitophagy - animal | hsa04137 | 0,00127 | 0,001607 | FOXO3/KRAS/JUN/MITF/GABARAPL1/TOMM20/HIF1A |
| Human papillomavirus infection | hsa05165 | 0,002137 | 0,002497 | APC/RHEB/IKBKE/KRAS/CSNK1A1/FADD/ CCND1/NFKB1/PIK3R1/STAT1/CCND2/CASP3/PTEN |
|
Leukocyte transendothelial migration |
hsa04670 | 0,002257 | 0,002608 | CLDN1/RHOA/ICAM1/RAC1/PIK3R1/MAPK14/MAPK13 |
| Autophagy - animal | hsa04140 | 0,005021 | 0,005561 | RHEB/KRAS/GABARAPL1/VPS18/PIK3R1/HIF1A/RPTOR/PTEN |
|
p53 signaling pathway |
hsa04115 | 0,006329 | 0,006662 | APAF1/CCND1/CCND2/CASP3/PTEN |
|
Chemokine signaling pathway |
hsa04062 | 0,010868 | 0,010727 | RHOA/FOXO3/KRAS/RAC1/CXCL8/NFKB1/PIK3R1 /STAT1 |
|
Apoptosis – multiple species |
hsa04215 | 0,013437 | 0,012642 | FADD/APAF1/CASP3 |
|
Natural killer cell mediated cytotoxicity |
hsa04650 | 0,018674 | 0,016969 | IFNGR1/KRAS/ICAM1/RAC1/PIK3R1/CASP3 |
| Hormone signaling | hsa04081 | 0,056541 | 0,047333 | SMAD5/SMAD1/RHOA/AGTR1/SMAD4/PIK3R1/THRB |
*FDR: False Discovery Rate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



