Submitted:
30 July 2025
Posted:
31 July 2025
You are already at the latest version
Abstract
Age-related macular degeneration (AMD) is a leading cause of irreversible central vision loss characterized by complex multifactorial pathogenesis involving genetic, inflammatory, oxidative, and environmental factors. This review provides a comprehensive translational overview of biomarkers associated with both dry and wet forms of AMD, integrating evidence across human studies and preclinical mouse models, including chemical, genetic, and laser-induced paradigms. We summarize key tissue, fluid, and systemic biomarkers linked to oxidative stress, inflammation, complement activation, extracellular matrix remodeling, angiogenesis, and gut microbiota alterations. Emphasis is placed on parallels and discrepancies between human AMD and animal models, highlighting opportunities and challenges in biomarker validation and application. The review underscores the potential of integrated biomarker profiles from ocular tissues, blood, tear fluid, aqueous and vitreous humor, and gut microbiome samples for advancing early diagnosis, therapeutic monitoring, and personalized treatment strategies. Future directions emphasize bridging experimental and clinical data to enhance biomarker-driven precision medicine in AMD.
Keywords:
biomarkers
; retina
; age-related degeneration
; humans and mice
Introduction
Dry age-related macular degeneration (dry AMD), the most prevalent form of AMD, represents a progressive neurodegenerative condition of the macula, affecting approximately 85–90% of AMD patients [1]. Non-exudative AMD is characterized by the gradual accumulation of drusen and progressive photoreceptor degeneration. In its advanced stage, known as geographic atrophy (GA), there is marked atrophy of the retinal pigment epithelium (RPE) and underlying choroid. GA represents a leading cause of irreversible central vision loss in the elderly population [2,3,4]. Patients with AMD may be thus classified as having early stage disease (early/intermediate dry AMD), in which visual function is generally not affected, or late AMD (generally characterized as either “wet” neovascular AMD, “dry” atrophic AMD or both), in which central vision is severely compromised [5]. Currently, wet AMD can be managed with anti-vascular endothelial growth factor (anti-VEGF, Vascular Endothelial Growth Factor) therapies, which help control disease progression and preserve vision. Although recent FDA-approved therapies for GA, the advanced non-exudative stage of AMD, can slow disease progression, no treatments are currently available that restore vision already lost due to advanced atrophy, highlighting a critical unmet clinical need [5]. The disease arises from a complex interplay of genetic susceptibility, chronic inflammation, oxidative stress, and mitochondrial dysfunction, with strong associations identified in genome-wide studies involving variants in CFH, ARMS2/HTRA1, and C3 [6,7]. Despite the significant burden of non-exudative AMD, no approved treatments currently exist to halt or reverse disease progression in its early and intermediate stages. Although complement inhibitors have recently been approved to slow the progression of GA, the advanced form of non-exudative AMD, a critical need remains for effective therapies that restore lost vision, as well as for early diagnostic and prognostic biomarkers to improve patient outcomes. Biomarker discovery efforts in recent years have focused on a wide array of biological sources, including blood, plasma, aqueous humor, tear fluid, and gut microbiota, to identify molecular signatures associated with disease onset, progression, and therapeutic response [8,9,10]. Understanding and validating these biomarkers in both human patients and animal models will be key to advancing precision medicine and developing targeted interventions for dry AMD. The integration of genetic mouse models with the analysis of systemic biofluids and gut-derived biomarkers provides a powerful framework for studying AMD pathogenesis. These biomarkers not only validate disease mechanisms at the ocular–systemic interface but also offer translational potential for the development of early diagnostics and personalized treatment strategies.
Chemical AMD Models for Biomarker Discovery
Chemically induced models of AMD provide powerful platforms to study the molecular and cellular mechanisms underlying disease progression. These models allow controlled induction of oxidative stress, photoreceptor apoptosis, or RPE degeneration, mimicking key features of both dry and neovascular (wet) AMD. Importantly, they have been instrumental in identifying tissue and circulating biomarkers involved in inflammation, oxidative damage, angiogenesis, and cell death. Several chemical agents are used to reproduce different aspects of AMD pathology. Sodium iodate (NaIO₃) is the most widely employed compound to selectively damage the RPE, leading to secondary photoreceptor degeneration and chronic inflammation, thereby modeling features of dry AMD [11]. Moreover, Cho et al. shown that NaIO₃ induces alterations in the expression of microRNAs (miRNAs), small non-coding RNAs that regulate gene expression and modulate pathological processes. Specifically, NaIO₃-induced RPE dysfunction has been associated with the deregulation of miR-146a-5p, which contributes to RPE degeneration and retinal inflammation [12]. miR-21-5p has also been identified as a promoter of necroptosis in RPE cells in NaIO₃-induced AMD models, through negative regulation of the Peli1 gene, thereby exacerbating cellular damage [13]. N-methyl-N-nitrosourea (MNU), a DNA-alkylating agent, induces rapid and selective apoptosis of photoreceptors, replicating retinal thinning and neuroinflammatory responses [14]. All-trans-retinal (atRAL) and bis-retinoids like A2E accumulate in the RPE and contribute to oxidative stress, lipofuscin formation, and complement activation, mimicking lipofuscin-driven toxicity observed in aging and dry AMD [15,16]. To model oxidative stress–related mechanisms more directly, hydrogen peroxide (H₂O₂) is widely used in vitro to induce acute oxidative damage in RPE cells or retinal explants [17]. H₂O₂ exposure leads to mitochondrial dysfunction, tight junction disruption, and apoptosis, making it a valuable system for studying early degenerative changes and testing antioxidant therapies. For modeling neovascular AMD (nAMD), intraocular injections of VEGF, Fibroblast Growth Factor 2 (FGF2), or inflammatory cytokines such as Tumor Necrosis Factor (TNF)-α and interleukin (IL)-1β are used to promote neovascularization and breakdown of the blood–retinal barrier [17,18,19]. Another widely used approach to mimic retinal hypoxia and angiogenic signaling involves the use of cobalt chloride (CoCl₂), a hypoxia-mimetic agent that stabilizes HIF-1α and induces the expression of VEGF and other pro-angiogenic factors [20]. CoCl₂ administration leads to neovascularization, gliosis, and photoreceptor loss, providing a useful model for studying hypoxia-driven mechanisms in nAMD [20,21]. This section outlines the most relevant chemically induced AMD models and summarizes the associated biomarkers across various biological compartments.
Retina and RPE-Choroid Tissue Biomarkers
In chemically induced models of AMD, including those using NaIO₃, MNU, atRAL, and bis-retinoids such as A2E, extensive analyses of retina and RPE–choroid complexes have revealed multiple biomarkers indicative of oxidative stress, inflammation, cell death, and extracellular matrix remodeling. In NaIO₃-treated rodents, oxidative stress markers such as 4-hydroxynonenal (4-HNE), 8-hydroxy-2'-deoxyguanosine (8-OHdG), and nitrotyrosine are consistently upregulated in RPE and photoreceptor layers, indicating ROS-mediated cellular injury [22] (Figure 1). Apoptotic markers, including cleaved caspase-3, Bcl-2-associated X protein (BAX), and cytochrome c, are also elevated, particularly in the outer nuclear layer and RPE, reflecting photoreceptor degeneration [14,23]. Inflammatory mediators such as TNF-α, IL-1β, IL-6, Monocyte Chemoattractant Protein (MCP)-1, and Glial Fibrillary Acidic Protein (GFAP) (indicative of Müller cell gliosis) are increased in both NaIO₃ and MNU models [24] (Figure 1). In models using A2E or atRAL, accumulation of these bis-retinoids in RPE cells leads to lysosomal dysfunction and upregulation of complement components like C3 and CFH, as well as lipid peroxidation products, closely mimicking the chronic features of dry AMD [25]. Additionally, altered expression of structural and extracellular matrix proteins such as MMP-2, MMP-9, and TIMP-3 has been observed, reflecting Bruch’s membrane (BrM) remodeling and RPE–choroid barrier disruption [22,24] (Figure 1). Additional models employing CoCl₂, a chemical hypoxia mimetic, simulate ischemic damage and HIF-1α pathway activation, leading to VEGF upregulation, glial activation, and neuronal loss in the outer retina [26] (Figure 1). Notably, CoCl₂ exposure alters the expression of several microRNAs (miRNAs) involved in the regulation of oxidative stress, angiogenesis, and cell survival. Specifically, miR-210-5p, miR-21a-5p, miR-29a-3p, and miR-16-5p are significantly upregulated, while miR-183-5p is downregulated, highlighting the contribution of non-coding RNA networks to hypoxia-induced retinal damage [27] (Figure 1). Similarly, exogenous administration of 4-HNE or H₂O₂ directly induces oxidative stress in retinal explants or in vivo, resulting in mitochondrial dysfunction, RPE barrier disruption, and increased expression of pro-apoptotic proteins and pro-inflammatory [28]. Additionally, these stressors modulate autophagic flux and have been used to study the interplay between oxidative damage and defective clearance mechanisms in AMD. Altered expression of structural and ECM proteins such as MMP-2, MMP-9, and Tissue Inhibitor of Metalloproteinase (TIMP)-3 has been consistently observed across models, reflecting BrM remodeling and RPE–choroid barrier disruption [28,29]. In the VEGF-induced model, localized overexpression of VEGF leads to RPE and photoreceptor layer disorganization, increased levels of the peroxynitrite nitrotyrosine, infiltration of CD68⁺ macrophages, upregulation of HIF-1α, ANGPT2, and MMP-9 in the choroid and retina [30,31,32] (Figure 1).
Blood and Plasma Biomarkers
In the NaIO₃ model, which induces selective RPE damage and secondary photoreceptor loss, systemic analyses of plasma have identified a distinct inflammatory signature. Proteomic and cytokine profiling revealed increased levels of complement proteins (C3, C4b, factor B) and serum amyloid A (SAA), indicative of innate immune activation [33]. Concurrently, elevated plasma concentrations of IL-6 and MCP-1/CCL2 reflect a chronic inflammatory state associated with retinal degeneration [33] (Figure 1). Similarly, in CoCl₂-treated animals plasma levels of VEGF, IL-6, and erythropoietin (EPO) are significantly upregulated, suggesting systemic responses to oxidative stress and Hypoxia-Inducible Factor (HIF)-1α pathway activation [34] (Figure 1).
Stool and Gut Microbiome Biomarkers
Emerging evidence from chemically induced models of AMD, particularly those using NaIO₃ or oxidative stress-inducing agents, highlights the gut–retina axis as a contributor to retinal degeneration. In NaIO₃-treated mice, 16S rRNA sequencing of fecal samples revealed pronounced gut dysbiosis, characterized by a decrease in Firmicutes and an increase in Bacteroidetes and Proteobacteria, microbial shifts commonly linked to pro-inflammatory states [35]. This imbalance was associated with reduced levels of short-chain fatty acids (SCFAs), especially butyrate, and elevated concentrations of lipopolysaccharide (LPS) in both stool and plasma, suggesting increased intestinal permeability and systemic inflammation. These changes correlated with retinal thinning and enhanced expression of inflammatory cytokines such as IL-6 and TNF-α in the retina [35] (Figure 1). Therapeutic interventions with probiotics or Short-Chain Fatty Acids (SCFAs) have shown protective effects on the retina, supporting a functional connection between gut microbial metabolites and retinal health [36]. Notably, in the NaIO₃-induced mouse model, combined treatment with Lactobacillus fermentum NS9 and Aronia anthocyanidin extract (AAE) produced beneficial effects both at the retinal level and in modulating gut microbiota composition. Fecal metagenomic analysis revealed a significant enrichment of the genus Parasutterella, particularly Parasutterella excrementihominis, in the AAE+LF group compared to both the disease model and the AAE-only group (Figure 1). Parasutterella has been previously associated with immunomodulatory activity and the metabolism of bile acids and amino acids, and its increased abundance may contribute to reduced oxidative stress and systemic inflammation [35].
Genetic Mouse Models for AMD
Understanding the complex pathophysiology of AMD has greatly advanced through the development of genetically engineered mouse models that mimic key features of both dry and neovascular AMD. These models enable detailed investigation of the molecular and cellular processes underlying disease onset and progression, particularly within the retina and RPE-choroid complex. Inflammation-driven models such as CC chemokine ligand 2 (Ccl2)⁻/⁻, C-X3-C Motif Chemokine Receptor 1 (Cx3cr1)⁻/⁻, and Ccl2⁻/⁻Cx3cr1⁻/⁻ replicate aspects of dry AMD by exhibiting subretinal microglial accumulation, upregulation of pro-inflammatory cytokines, and complement activation [37,38,39]. Oxidative stress models, including Sod1⁻/⁻ mice, demonstrate how impaired antioxidant defense leads to RPE degeneration and photoreceptor loss through increased production of oxidative biomarkers such as 4-HNE and 8-OHdG [40,41,42]. Meanwhile, models involving HtrA1 overexpression and Timp3 mutations (e.g., S179C) highlight the role of extracellular matrix remodeling in AMD pathogenesis [43,44,45]. On the neovascular front, Very low density lipoprotein receptor (Vldlr)⁻/⁻ mice spontaneously develop subretinal neovascularization resembling type 2 macular neovascularization [46]. Importantly, emerging evidence from C3-deficient and Complement factor H (CFH)-variant mouse models suggests a functional link between AMD-associated genetic variants and alterations in the gut microbiota, reinforcing the role of systemic factors in modulating retinal disease [47,48]. Collectively, these models provide a valuable experimental framework for dissecting the multifactorial nature of AMD and for identifying novel therapeutic targets across inflammatory, oxidative, angiogenic, and systemic pathways.
Retina and RPE-Choroid Tissue Biomarkers
Genetically engineered mouse models of AMD have provided valuable insights into molecular and cellular alterations occurring within the retina and RPE–choroid complex during disease progression. In Ccl2⁻/⁻, Cx3cr1⁻/⁻, and Ccl2⁻/⁻ Cx3cr1⁻/⁻ mice, which exhibit features reminiscent of dry AMD, histological and transcriptomic analyses revealed elevated expression of inflammatory markers such as TNF-α, IL-1β, and MCP-1 in the RPE and subretinal space, along with microglial accumulation and complement activation [38,39,49] (Figure 2). Increased GFAP expression indicates Müller cell gliosis, while upregulation of Iba1 and CD68 highlights enhanced microglial activity and infiltration. In Sod1⁻/⁻ mice, characterized by impaired oxidative defense, the retina shows elevated levels of oxidative stress biomarkers including 4-HNE, 8-OHdG, and nitrotyrosine, particularly in the photoreceptor layer and RPE, correlating with progressive degeneration [41,42] (Figure 2). Liu et al. [50] identified interleukin-1 receptor–associated kinase M (IRAK-M) as a critical immunoregulatory protein in the RPE that progressively declines with age. Rare genetic variants in IRAK3, the gene encoding IRAK-M, were found to be associated with an increased risk of developing AMD (Figure 2). Analyses of human tissues and genetic mouse models revealed that IRAK-M expression in the RPE diminishes with aging and oxidative stress, with a more pronounced reduction in AMD-affected tissues. Mice lacking Irak3 exhibited early-onset outer retinal degeneration, which was further aggravated by oxidative insults. The loss of IRAK-M disrupted RPE homeostasis, leading to mitochondrial dysfunction, cellular senescence, and abnormal cytokine production [50]. Conversely, IRAK-M overexpression conferred protection to RPE cells against oxidative and immune stress [50]. Notably, subretinal delivery of adeno-associated virus (AAV) expressing human IRAK3 mitigated light-induced retinal damage in wild-type mice and reduced age-related degeneration in Irak3-deficient mice. These findings suggest that restoring IRAK-M levels in the RPE may help counteract chronic inflammation and cellular dysfunction in AMD, offering a promising therapeutic strategy for retinal degeneration. Models overexpressing HtrA1 or carrying disease-associated mutations in Timp3 (e.g., S179C) display thickening of BrM, accumulation of basal deposits, and upregulation of extracellular matrix remodeling enzymes such as MMP-2 and MMP-9 in BrM [44,51] (Figure 2). In Vldlr⁻/⁻ mice, which spontaneously develop subretinal neovascularization resembling wet AMD, there is increased retinal expression of VEGF, HIF-1α, and Angiopoietin (ANGPT) 2, alongside infiltration of CD68⁺ macrophages and photoreceptor disorganization [46,52] (Figure 2).
Despite the extensive use of genetically engineered mouse models to study the molecular and cellular mechanisms of AMD, there is a notable lack of published data on body fluid biomarkers, particularly in the context of these models. Specifically, blood and plasma biomarkers, such as complement components, inflammatory cytokines, or oxidative stress markers, are rarely measured or reported in a systematic manner. Similarly, tear fluid biomarkers remain virtually unexplored in murine models due to technical limitations in sample collection and the small volume of tears in rodents. Additionally, aqueous and vitreous humor biomarkers, which are clinically relevant in human AMD for tracking intraocular inflammation, angiogenesis, and complement activity, are seldom analyzed in mouse models, largely because of the challenges associated with fluid extraction from small eyes. This gap in biomarker profiling in murine systems limits the ability to directly translate findings from genetic models to clinical diagnostics and hinders the development of non-invasive monitoring strategies in preclinical AMD research. Addressing this deficiency through improved micro-sampling techniques and proteomic or transcriptomic analyses could substantially enhance the translational value of these models.
Stool and Gut Microbiome Biomarkers
Zysset-Burri et al. [53] demonstrated that complement C3-deficient mice exhibited a higher Firmicutes-to-Bacteroides ratio compared to wild-type controls, a microbial profile resembling that commonly observed in patients with nAMD (Figure 2). In human studies, the CFH3 single nucleotide polymorphism (SNP) was significantly associated with nAMD and positively correlated with increased abundance of Negativicutes. Additionally, individuals carrying the CFH3 variant showed elevated levels of Clostridiales, whereas those with CFH1 and CFH2 variants exhibited a negative correlation with this microbial group. These findings support a functional interaction between host genetics and the gut microbiome in AMD, suggesting that genetically driven alterations in microbial composition may influence complement system activation and contribute to disease progression [47,54] A deeper understanding of this gene–microbiota interplay will be essential for developing personalized therapeutic strategies targeting AMD pathogenesis.
Laser-Induced Neovascularization Model
The laser-induced neovascularization model is the most widely used experimental approach for studying wet AMD [55,56]. In this model, laser burns are applied to the mouse retina using a focused green Argon laser or a similar photocoagulation system to rupture BrM, which triggers the growth of abnormal new blood vessels from the choroid into the subretinal space [57]. As a result, subretinal neovascularization develops, often accompanied by vascular leakage and local inflammation [58]. This process closely mimics key pathological hallmarks of human wet AMD, such as MNV, vascular leakage, and subretinal fibrosis, making the model highly relevant for investigating disease mechanisms and testing new therapies [59,60].
The main advantages of the laser-induced neovascularization model are its reproducibility, rapid induction of lesions, and compatibility with a wide range of genetic backgrounds and imaging techniques [61]. However, it also has limitations: it represents an acute injury rather than the slow, age-related progression seen in human AMD, and mice lack a macula, the central retinal region most affected in patients [58,62]. Despite these differences, the model reliably recapitulates the angiogenic, inflammatory, and fibrotic responses central to human wet AMD, and has been instrumental in the preclinical development of anti-VEGF therapies and the identification of disease biomarkers, supporting its continued relevance for translational AMD research [63].
Because the laser-induced neovascularization closely mirrors the molecular and cellular changes seen in human AMD, it serves as a powerful platform for biomarker discovery. Using this model, researchers have identified a range of tissue and fluid biomarkers that reflect disease activity and offer insights into underlying mechanisms. The sections below highlight key biomarkers identified in the retina, blood, tear fluid, ocular humors, and even the gut microbiome, emphasizing their relevance for translational AMD research.
Retina and RPE-Choroid Tissue Biomarkers
Laser-induced neovascularization in mice leads to profound molecular changes in the retina and RPE-choroid complex. Transcriptomic and proteomic analyses reveal upregulation of angiogenic genes such as VEGFA, a primary driver of neovascularization and a direct target of anti-VEGF therapies in humans [64]. Inflammatory genes, including Ccl2 (MCP-1), Ccl8, and Cxcl9, are also highly expressed, reflecting immune cell recruitment and a pro-inflammatory microenvironment [65,66] (Figure 3). These responses are not unique to mice; comparative studies show that many of these genes are conserved and similarly upregulated in human AMD lesions, underscoring their translational relevance [59]. Additionally, extracellular matrix remodeling genes such as Comp, Lrcc15, Fndc1, and Thbs2 are elevated, contributing to fibrosis, a process that can limit the effectiveness of anti-angiogenic treatments and lead to vision loss in patients [66]. MicroRNAs (miRNAs) including miR-505, miR-155, miR-342-5p, and miR-126-3p are also differentially expressed, acting as post-transcriptional regulators of these pathogenic pathways [30] (Figure 3). The identification of these conserved molecular signatures supports the model’s use for identifying new therapeutic targets and for understanding the mechanisms underlying neovascularization and fibrosis in AMD.
Blood and Plasma Biomarkers
In the laser-induced neovascularization mouse model, the search for systemic, minimally invasive biomarkers has focused on circulating miRNAs due to their stability and regulatory roles in angiogenesis and inflammation. Recent studies have identified robust dysregulation of mmu-miR-486a-5p and mmu-miR-92a-3p in both the blood and RPE/choroid tissues following laser-induced neovascularization, as confirmed by RNA sequencing and RT-qPCR [67] (Figure 3). These miRNAs are functionally relevant, with evidence that mmu-miR-486a-5p and mmu-miR-92a-3p modulate microglial cell viability and mobility, processes implicated in neovascularization pathogenesis [67]. Importantly, similar miRNA alterations have been reported in plasma samples from patients with neovascular AMD, highlighting their translational potential as biomarkers for disease activity, early detection, and therapeutic monitoring [67]. The accessibility of blood sampling further supports the clinical utility of these biomarkers, allowing for longitudinal assessment without invasive ocular procedures. Additionally, panels of miRNAs may improve diagnostic accuracy beyond individual markers. The conservation of miRNA dysregulation between the laser-induced neovascularization and human AMD underscores the value of this model for biomarker discovery and validation in translational research [67].
Tear Fluid Biomarkers
Tear fluid has recently emerged as a valuable, non-invasive source of biomarkers for neovascular AMD. Studies using the mouse model have demonstrated that VEGF concentrations are significantly increased in tears following neovascularization induction, mirroring findings in human patients with active neovascular AMD [68] (Figure 3). The primary source of this tear VEGF is the choroid-RPE complex, and its presence in tears reflects ongoing angiogenic and inflammatory activity within the eye. The ease and repeatability of tear sampling make it particularly attractive for longitudinal disease monitoring and assessing therapeutic response. Notably, clinical studies have also reported that tear VEGF levels can differ by sex, suggesting potential for personalized biomarker strategies. The parallel elevation of tear VEGF in both the murine model and human AMD underscores the translational promise of tear-based assays, which could facilitate routine, non-invasive monitoring of disease activity and treatment efficacy in clinical practice [68].
Aqueous and Vitreous Humor Biomarkers
The analysis of aqueous and vitreous humor has provided important insights into intraocular biomarkers for neovascular AMD. In the laser-induced neovascularization model, levels of angiogenic and fibrotic proteins such as VEGF and thrombospondin 2 (THBS2) are significantly elevated following neovascularization induction [69] (Figure 3). These proteins are well-established mediators of pathological neovascularization and tissue remodeling in AMD. Measurement of VEGF and related factors in ocular fluids is already integrated into clinical practice, particularly for patients undergoing intravitreal injections or ocular surgery. The ability of the murine model to recapitulate these biomarker changes further validates its utility for preclinical evaluation of anti-angiogenic and anti-fibrotic therapies. Moreover, the parallel findings in both animal models and human patients support the ongoing development of aqueous and vitreous fluid-based biomarkers for monitoring disease activity and therapeutic response in neovascular AMD [59,70].
Stool and Gut Microbiome Biomarkers
Recent studies using the mouse model have revealed that the gut microbiome may play a significant role in retinal disease processes. During neovascularization development, researchers have observed notable shifts in the composition of gut bacteria, particularly increases in genera such as Lachnospiraceae UCG-001 and Candidatus Saccharimonas, which correlate with distinct changes in fecal metabolite profiles [71] (Figure 3). These microbial alterations are thought to influence immune cell activation and systemic inflammation, potentially affecting the retinal environment and modulating the severity of neovascularization. Members of the Lachnospiraceae family are recognized for their ability to produce SCFAs, which can regulate host immunity and maintain epithelial barrier integrity [72] (Figure 3). Additionally, metabolic products from these bacteria may impact angiogenic signaling pathways relevant to AMD pathogenesis. The gut-retina axis, therefore, represents a promising area for biomarker discovery and therapeutic intervention. Although the direct clinical application of stool-based biomarkers in AMD patients is still being explored, these findings highlight the systemic nature of ocular diseases and suggest that gut microbiome profiling could eventually aid in risk assessment, disease monitoring, or even in tailoring microbiome-targeted therapies for AMD [73].
Together, these findings demonstrate that the laser-induced neovascularization model not only recapitulates key features of human neovascular AMD but also provides a robust platform for the discovery and validation of clinically relevant biomarkers across multiple tissues and biological fluids, advancing both mechanistic understanding and translational research in AMD [67,68].
Biomarkers in “Dry” AMD in Humans
Dry AMD, also known as non-neovascular or atrophic AMD, is the most prevalent form of the disease, accounting for up to 90% of cases. It primarily affects individuals over the age of 60 and is characterized by a gradual loss of central vision due to progressive degeneration of the macula, the central region of the retina responsible for detailed visual tasks [5]. A hallmark of early dry AMD is the accumulation of extracellular deposits known as drusen, which form between the RPE and BrM. These deposits, composed of lipids, proteins, and complement factors, are thought to arise from impaired fluid and metabolite transport across BrM, leading to metabolic stress in the RPE. The pathogenesis of dry AMD begins with a thickening of BrM, primarily due to the buildup of lipid- and protein-rich material. This accumulation disrupts the normal exchange of nutrients and waste products between the choroid and the retina, compromising RPE homeostasis. As a result, oxidative stress increases within RPE cells, leading to the accumulation of lipofuscin, an autofluorescent byproduct of incomplete lysosomal degradation, that further impairs lysosomal function and cholesterol metabolism. In response to cellular stress, RPE cells may release membranous vesicles such as exosomes, which are believed to contribute to sub-RPE deposit formation. Moreover, the presence of serum-derived proteins in drusen suggests that systemic factors can infiltrate the sub-RPE space, likely due to BrM dysfunction, further promoting deposit accumulation [5]. Over time, these pathogenic processes can lead to GA, the advanced form of dry AMD, which involves the irreversible loss of RPE cells, photoreceptors, and the underlying choriocapillaris. This results in permanent central vision loss, for which no approved curative treatments currently exist [74]. Given the absence of approved treatments for dry AMD, particularly in advanced stages, there is a growing emphasis on the identification of early and predictive biomarkers that can aid in diagnosis, monitor disease progression, and guide therapeutic development.
Retina and RPE-Choroid Tissue Biomarkers
Extensive molecular profiling of the retina and RPE-choroid complex in human dry AMD has identified numerous tissue-level biomarkers reflecting key pathogenic processes, such as oxidative stress, inflammation, lipid metabolism and extracellular matrix remodeling. Clinical studies involving AMD patients and post-mortem donor eyes have provided strong evidence for the accumulation of oxidative stress-related DNA, protein, and lipid biomarkers in AMD pathology. Among these, oxidative DNA damage, particularly the formation of 8-OHdG, has been prominently observed in AMD donor eyes, with higher levels detected in cases of dry AMD with RPE atrophy [75] (Figure 4). Given the abundance of docosahexaenoic acid (DHA) in the lipid-rich outer segments of photoreceptors [76], oxidative degradation of DHA has been implicated in AMD, as demonstrated by increased levels of carboxyethyl pyrrole (CEP) protein adducts, a marker of DHA peroxidation, within BrM of AMD donor eyes [77,78] (Figure 4). Notably, CEP levels were shown to be approximately 60% higher in dry AMD eyes compared to controls, as measured by ELISA [79,80]. However, the levels of 4-hydroxy-2-nonenal (4-HNE) modified proteins did not differ significantly across retinal regions or among the histological stages of AMD [81]. A separate study integrating proteomic and genomic analyses found that increased plasma CEP levels in AMD were associated with elevated expression of associated with age-related maculopathy susceptibility 2 (ARMS2), CFH, and complement C3, further linking oxidative stress to genetic susceptibility in dry AMD [82] (Figure 4). In addition to ocular findings, systemic markers of oxidative stress such as malondialdehyde (MDA), 8-OHdG, and protein carbonyls were significantly elevated in the serum of AMD patients, suggesting that oxidative damage in AMD is not confined to the retina but reflects a broader systemic oxidative imbalance [83] (Figure 4). Additionally, impaired extracellular matrix (ECM) turnover and thickening of BrM are early hallmarks of AMD, closely associated with altered activity of MMPs. Analyses of human AMD eyes have demonstrated elevated expression and accumulation of MMP-2 and MMP-9 in BrM and RPE choroid tissue, accompanied by deposit buildup beneath the RPE, evidence of disrupted ECM remodeling [6,84] (Figure 4). MMP-2, primarily secreted by RPE cells, becomes dysregulated in AMD, leading to excessive collagen IV accumulation and basal deposits [84]. Although total levels of activated MMP-2 and MMP-9 may actually decrease in AMD donor eyes, an increase in high-molecular-weight proenzyme complexes likely hinders their activation, contributing to ECM thickening [85]. Collectively, these findings highlight the critical disruption of MMP-mediated ECM turnover as a key tissue biomarker and potential therapeutic target.
Blood and Plasma Biomarkers
In AMD, increasing evidence suggests that systemic biomarkers detectable in blood and plasma reflect underlying pathogenic processes such as complement activation, oxidative stress, and chronic inflammation. Among the most consistently reported are components of the complement system, including elevated levels of C3a, C5a, and factor B, as well as reduced levels or functional impairment of CFH, particularly in individuals carrying the AMD-associated CFH Y402H polymorphism [86,87]. Inflammatory markers such as C-reactive protein (CRP), IL-6, and TNF-α are also elevated in the plasma of patients with dry AMD and have been linked to disease progression and genetic [88] (Figure 4). Oxidative stress-related plasma biomarkers, including malondialdehyde (MDA), 8-OHdG, and CEP-adducts, are found at significantly higher levels in dry AMD patients compared to age-matched controls, indicating systemic lipid peroxidation and DNA damage [77,89] (Figure 4). Multivariate analysis revealed that dry AMD was an independent factor associated with increased expression of miR-23a-3p, miR-126-3p, miR-126-5p, miR-146a, miR-191-5p and decreased expression of miR-16-5p, miR-17-3p, miR-17-5p. Six miRNAs were differentially expressed in peripheral blood nuclear cells of AMD subtypes: four were increased in dry AMD patients (miR-126-3p, miR-126-5p, miR-150-5p, miR-155-5p) whereas two were increased in wet AMD patients (miR-30b, miR-191-5p) [90]. Visual acuity and expression of miR-126-3p, miR-126-5p, miR-155-5p were positively correlated, whereas visual acuity and miR-191-5p expression were negatively correlated [90,91,92] (Figure 4). In addition, miR-27a-3p was suggested as a potential diagnostic biomarker for both wet and dry AMD in whole blood [90].
These circulating biomarkers not only provide insights into systemic disease mechanisms but may also serve as accessible, non-invasive tools for risk stratification, early detection, and monitoring of therapeutic response in dry AMD.
Tear Fluid Biomarkers
Tear fluid biomarkers have emerged as a promising, non-invasive source for detecting molecular changes associated with AMD. A study from Winiarczyk et al. [93] analyzed tear films from a total of 22 patients (8 with wet AMD, 6 with dry AMD, and 8 control individuals. 2D electrophoresis was used to separate tear film proteins prior to their identification with matrix-assisted laser desorption/ionization time of flight spectrometer (MALDI-TOF/TOF). They observed that dry AMD patients showed 97 specific proteins related to 44 pathway patterns and that these proteins were involved in more pathways than those identified in the wet AMD [93]. The two most widely expressed pathways were the Wnt signaling pathway and the Huntington disease pathway. It is noteworthy that there was a major representation of proteins involved in oxidative stress, inflammation, and proteolysis, e.g., the autophagy-related PI3K pathway. Autophagy failure has been reported to be associated with AMD development [94]. Notably, eight proteins, including shootin-1, histatin-3, and prolactin-inducible protein 1, were uniquely upregulated in AMD tears, suggesting their potential as disease-specific biomarkers [93] (Figure 4). These proteins are involved in pathways related to inflammation, apoptosis, and angiogenesis, reflecting underlying AMD pathophysiology. The study also highlighted the presence of signaling molecules such as STAT3 and FGFR1 linked to neovascular processes in wet AMD [93,95] (Figure 4). Mass spectrometry-based proteomics has further advanced tear biomarker discovery, enabling sensitive and quantitative profiling of tear proteins in ocular diseases including AMD [96]. These findings underscore the utility of tear fluid proteomics for early diagnosis, monitoring disease progression, and evaluating therapeutic response in AMD.
Aqueous and Vitreous Humor Biomarkers
Aqueous and vitreous humor biomarkers have gained increasing attention in dry AMD research due to their proximity to retinal tissues and potential to reflect local pathological changes. Proteomic analyses of aqueous humor from AMD patients have identified significant alterations in proteins involved in inflammation, extracellular matrix remodeling, and immune regulation. For example, Huang et al. [97] used affinity-based proteomics to discover 82 proteins significantly altered in geographic atrophy (GA), a late dry AMD form, highlighting Secreted Modular Calcium-binding protein 2 (SMOC2) and IL-6 as promising biomarkers linked to RPE dysfunction and inflammation (Figure 4). Other studies confirmed elevated levels of complement and coagulation cascade proteins, such as clusterin and serpin A4, in aqueous humor, in dry AMD patients suggesting dysregulated protein metabolism and immune activation [98,99] (Figure 4). Additionally, increased VEGF levels have been detected in aqueous humor of dry AMD patients with subretinal drusen deposits, implicating angiogenic signaling even in early disease [100]. Although vitreous humor is less frequently studied in dry AMD, proteomic profiling indicates changes in complement factors and neurodegeneration-related proteins, supporting its role as a biomarker source [101]. Collectively, aqueous and vitreous humor biomarker studies provide valuable insights into local molecular alterations in dry AMD, offering potential for improved diagnosis and monitoring of disease progression.
Stool and Gut Microbiome Biomarkers
Emerging evidence from recent studies supports a significant association between gut microbiota composition and dry AMD. Using metagenomic and Mendelian randomization approaches, specific gut microbial taxa have been causally linked to dry AMD risk. Notably, families and genera such as Peptococcaceae, Bilophila, Faecalibacterium, and Roseburia have been associated with an increased risk of developing dry AMD, whereas Candidatus Soleaferrea, Desulfovibrio, and the Eubacterium ventriosum group appear to exert protective effects [102] (Figure 4). These microbial shifts are thought to contribute to systemic inflammation and retinal degeneration via the gut-retina axis. Mechanistically, dysbiosis may promote increased gut permeability and elevated lipopolysaccharide (LPS) translocation, triggering chronic low-grade inflammation that affects retinal health. Metabolomic analyses of stool samples from dry AMD patients reveal altered profiles of microbial metabolites, including bile acids, tryptophan catabolites, and branched-chain amino acids, which are involved in immune regulation and oxidative stress pathways relevant to AMD pathogenesis [53,103]. Moreover, metabolomic analyses of fecal samples from patients with dry AMD reveal altered profiles of microbial metabolites such as bile acids, tryptophan catabolites, and branched-chain amino acids, all of which are involved in immune regulation and oxidative stress pathways relevant to the pathogenesis of the disease [102].
Biomarkers in “Wet” AMD in Humans
Neovascular, or wet AMD is a leading cause of irreversible central vision loss in the elderly population worldwide. The disease is characterized by the growth of abnormal choroidal blood vessels through BrM into the subretinal space, resulting in exudation, hemorrhage, and ultimately fibrotic scarring that damages the macula [104]. While anti-VEGF therapies have transformed the management of wet AMD, inter-individual variability in disease progression and treatment response remains a significant challenge. This has driven the search for robust biomarkers that can support early diagnosis, predict prognosis, guide therapeutic decisions, and monitor disease activity. In humans, biomarker discovery has leveraged advances in ocular imaging, molecular profiling of ocular and systemic fluids, and genetic analysis, providing a multidimensional understanding of disease pathogenesis and progression.
Retinal and RPE-Choroid Tissue Biomarkers
Direct sampling of retinal and RPE-choroid tissues in human patients is limited to rare surgical specimens or postmortem studies, but advances in non-invasive imaging have enabled the identification of surrogate tissue biomarkers. Spectral-domain optical coherence tomography (SD-OCT) has become central to the clinical management of wet AMD, offering high-resolution visualization of retinal and subretinal structures. Key SD-OCT biomarkers include the presence and volume of intraretinal fluid (IRF), subretinal fluid (SRF), and pigment epithelial detachments (PEDs), all of which are strongly associated with active neovascularization and worse visual prognosis [105]. Quantitative changes in central retinal thickness and the dynamics of fluid compartments are routinely used to guide anti-VEGF therapy and assess treatment response [106]. Hyperreflective foci, seen as discrete hyperreflective dots within the neurosensory retina, have been linked to activated microglia or lipoprotein aggregates and are associated with chronic inflammation, photoreceptor degeneration, and progression to fibrosis [107]. Outer retinal tubulations and subretinal hyperreflective material are additional OCT features that reflect photoreceptor degeneration and chronicity of disease [108]. Beyond OCT, fundus autofluorescence and fluorescein angiography provide further insight into RPE health and the extent of neovascularization, although their use as quantitative biomarkers is more limited [109]. While direct molecular profiling of retinal tissue is rare in humans, transcriptomic and proteomic studies of surgically excised neovascularization membranes have revealed upregulation of angiogenic (VEGFA), inflammatory (CCL2, CXCL9), and extracellular matrix remodeling genes (THBS2, MMPs), mirroring findings in animal models [110] (Figure 5).
Blood, Plasma, and Serum Biomarkers
Systemic biomarkers, particularly circulating miRNAs, have emerged as promising minimally invasive tools for wet AMD. Multiple studies have identified distinct miRNA signatures in whole blood, plasma, and serum of patients with neovascular AMD. For example, elevated levels of miR-27a-3p, miR-29b-3p, and miR-195-5p have been reported in whole blood, while miR-23a-3p, miR-30b, miR-191-5p, and miR-223-3p are upregulated in peripheral blood nuclear cells [67] (Figure 5). Conversely, miR-16-5p, miR-17-3p, miR-150-5p, and miR-155-5p are downregulated in wet AMD compared to healthy controls [67]. In plasma, increased miR-16-5p, miR-30b, and miR-191-5p, along with decreased miR-23a-3p, have been observed, with several miRNAs (such as miR-26b-5p and miR-27b-3p) appearing specific to the neovascular form of the disease [111]. Serum analyses have revealed reduced levels of miR-34a-5p, miR-126-3p, miR-145-5p, and miR-205-5p, some of which target VEGFA, the principal driver of neovascularization [92] (Figure 5). These miRNAs are functionally implicated in angiogenesis, inflammation, and extracellular matrix remodeling, and their conservation between human patients and animal models underscores their translational potential.
Beyond miRNAs, systemic markers of inflammation and oxidative stress have been investigated in wet AMD. Elevated plasma levels of CRP, complement activation fragments (e.g., C3a, C5a), and pro-inflammatory cytokines (such as IL-6 and TNF-α) have been associated with increased risk and activity of neovascular AMD, reflecting the role of systemic immune dysregulation in disease pathogenesis [112] (Figure 5). Markers of oxidative stress, including malondialdehyde and advanced glycation end products, are also elevated in AMD patients, supporting the contribution of chronic oxidative injury to retinal degeneration [113].
Tear Fluid Biomarkers
Tear fluid has recently gained attention as a non-invasive source of biomarkers for wet AMD. Elevated concentrations of VEGF have been detected in the tears of patients with active neovascular AMD, paralleling findings from preclinical models [68] Tear VEGF levels correlate with intraocular disease activity and may exhibit sex-specific differences, suggesting potential for personalized disease monitoring [68] (Figure 5). The accessibility and repeatability of tear sampling make it particularly attractive for longitudinal studies and for assessing therapeutic response in clinical settings. Other tear-based markers, such as inflammatory cytokines and matrix metalloproteinases, are under investigation, but their diagnostic and prognostic value in AMD remains to be fully established [68].
Aqueous and Vitreous Humor Biomarkers
Analysis of intraocular fluids, particularly aqueous and vitreous humor, has provided direct evidence of the molecular milieu associated with wet AMD. VEGF levels are consistently elevated in both aqueous and vitreous samples from patients with neovascular AMD, reflecting ongoing angiogenic activity and forming the basis for anti-VEGF therapeutic strategies [114]. In addition to VEGF, increased concentrations of fibrotic mediators such as thrombospondin 2 (THBS2), as well as various interleukins and matrix metalloproteinases (MMP-9), have been reported [115,116] (Figure 5). These factors are involved in tissue remodeling, inflammation, and fibrosis, processes central to disease progression and vision loss. Proteomic analyses of vitreous samples have further identified dysregulation of proteins involved in complement activation, lipid metabolism, and extracellular matrix turnover, highlighting the complex molecular environment of the neovascular retina [117].
Genetic Biomarkers
Genetic association studies have identified several robust risk loci for AMD, most notably single nucleotide polymorphisms in the CFH gene and the ARMS2/HTRA1 locus (Figure 5). These variants account for a substantial proportion of the heritable risk for AMD and are increasingly used to stratify patients in both clinical and research settings [6]. Additional genetic markers under investigation include variants in genes involved in lipid metabolism (e.g., APOE, LIPC), extracellular matrix regulation (e.g., TIMP3), and angiogenic signaling pathways (e.g., VEGFA), further expanding the landscape of genetic risk assessment in AMD [118,119,120] (Figure 5). Polygenic risk scores integrating multiple loci are being developed to improve predictive accuracy for disease onset and progression [121].
Gut Microbiome Biomarkers
Emerging evidence suggests that the gut microbiome may influence AMD pathogenesis in humans, as in animal models. Distinct microbial signatures and metabolite profiles have been associated with AMD risk, although the clinical application of stool-based biomarkers remains in its infancy [47]. Studies have reported alterations in the relative abundance of specific bacterial genera, such as increased Lachnospiraceae and decreased Bacteroidetes, in AMD patients compared to controls (Figure 5). These microbial shifts may modulate systemic inflammation and immune responses, ultimately impacting retinal health through the gut-retina axis [122].
Collectively, these findings demonstrate that wet AMD in humans is associated with a diverse array of biomarkers detectable through advanced imaging, systemic and ocular fluid analysis, and genetic profiling. Many of these biomarkers are conserved across preclinical models and human disease, supporting their utility in translational research and personalized medicine for AMD [68].
Comparative Analysis of Biomarker Profiles in Chemically Induced Models and Human Dry AMD
Comparative analyses between chemically induced mouse models and human dry AMD revealed substantial overlap in biomarker profiles across species and systems. For example, oxidative stress markers such as 8-OHdG and lipid peroxidation products like 4-HNE and CEP are consistently elevated in both human AMD donor tissues and chemically treated animal models [75]. Similarly, upregulation of pro-apoptotic markers (cleaved caspase-3, Bcl-2-associated X protein) and inflammatory cytokines (IL-6, TNF-α, MCP-1) is observed in the retina and plasma of AMD patients as well as in NaIO₃ and MNU models [24]. Matrix metalloproteinases (MMP-2, MMP-9) involved in BrM remodeling are also dysregulated across both settings [84]. Furthermore, complement activation (e.g., C3, Factor B), systemic oxidative stress markers (MDA, 8-OHdG), and specific microRNAs such as miR-21 and miR-146a are common to both human AMD and chemically induced models [33,90,123]. While direct proteomic profiling of tear fluid and intraocular humors has been extensively performed in AMD patients, such analyses remain limited or absent in mouse models. Nonetheless, gut microbiota alterations and reduced SCFA levels observed in NaIO₃-treated mice closely mirror findings in human dry AMD, supporting the relevance of the gut-retina axis in disease progression [35,102].
Comparison of Biomarker Findings in the Genetic Mouse Model and Human Dry AMD
Comparative studies between genetic mouse models and human dry AMD have revealed substantial overlap in molecular biomarkers associated with disease pathogenesis, particularly in the areas of oxidative stress, complement activation, and inflammation. In both human donor eyes and mouse models such as Sod⁻/⁻ and Sod2 knockdown, oxidative stress biomarkers, including 4-HNE, 8-OHdG, and CEP adducts, are elevated in the retina and RPE-choroid complex, reflecting conserved mechanisms of photoreceptor and RPE damage [41,42,124]. Pro-inflammatory cytokines such as IL-6, IL-1β, and TNF-α are consistently upregulated in both human dry AMD tissues and inflammatory mouse models like Irak3⁻/⁻, supporting a shared inflammatory signature [38,39,50]. However, discrepancies remain; for instance, while human studies report elevated plasma CRP and systemic oxidative markers, these are not consistently measured in mouse models due to sampling limitations. Additionally, features like geographic atrophy, a hallmark of late-stage dry AMD in humans, are incompletely replicated in mice. Nevertheless, the convergence of key biomarkers across species underscores the translational value of murine models for elucidating mechanisms and testing potential therapies in dry AMD. Emerging evidence from both human studies and genetic mouse models supports a critical role for the gut microbiota in modulating AMD, particularly through biomarkers linked to inflammation, complement activation, and metabolic imbalance. In human AMD patients, especially those with nAMD, gut microbiome profiling has revealed an increased Firmicutes/Bacteroidetes (F/B) ratio, reduced microbial diversity, and enrichment of pro-inflammatory taxa such as Proteobacteria, Actinobacteria, and Clostridiales [53,54,125]. These microbial shifts are often accompanied by decreased levels of SCFA-producing bacteria, such as Akkermansia muciniphila and Ruminococcaceae, which are known to maintain gut barrier integrity and regulate immune responses. Remarkably, similar dysbiotic signatures have been observed in genetically modified mouse models, such as Cfh⁻/⁻, Cfh-H402H, and C3⁻/⁻ mice, particularly when combined with high-fat or high-glycemic diets [47,53,54,72]. These models demonstrate elevated F/B ratios, loss of A. muciniphila, and increased abundance of Negativicutes and Clostridiales, echoing the gut microbial imbalances seen in AMD patients [53,103,126]. Furthermore, certain CFH genetic variants in humans have been correlated with specific microbiome alterations, suggesting a gene–microbiota interaction that may influence systemic inflammation and complement dysregulation [53,126]. Collectively, these findings underscore the potential of gut microbiota-derived biomarkers, detectable in stool or plasma, as non-invasive indicators of AMD risk and progression, while also highlighting the value of murine models for mechanistic studies of the gut-retina axis.
Comparison of Biomarker Findings in the Mouse Model and Human Wet AMD
The laser-induced neovascularization in mice and human wet AMD share fundamental pathological features, including abnormal angiogenesis, inflammation, and extracellular matrix remodeling, which are reflected in their biomarker profiles. Both systems exhibit marked upregulation of VEGF, a critical driver of neovascularization targeted by current therapies [127]. Inflammatory chemokines such as CCL2 and CXCL9 are elevated in both the mouse model and human disease, facilitating immune cell recruitment and sustaining a pro-inflammatory microenvironment [128,129]. Similarly, proteins involved in extracellular matrix remodeling, including HBS2 and matrix metalloproteinases, are increased, contributing to fibrosis and tissue remodeling in both contexts [115,130]. At the systemic level, miRNAs dysregulated in the blood and ocular tissues of mice, such as miR-155 and miR-126-3p, have also been identified in the plasma or serum of patients with wet AMD, supporting their conserved role as regulators of angiogenesis and inflammation [67]. Elevated VEGF levels in ocular fluids, including aqueous humor, vitreous, and tear fluid, are consistently observed in both models, underscoring their translational relevance as minimally invasive biomarkers [68]. Additionally, oxidative stress markers and complement activation components are implicated in disease pathogenesis in both species [131].
Despite these parallels, important differences limit the direct translation of findings from the mouse model to human disease. Anatomically, mice lack a macula, the specialized central retinal region predominantly affected in human AMD, restricting the model’s ability to fully replicate the spatial and functional consequences of macular pathology [132]. The laser-induced neovascularization represents an acute injury with rapid lesion formation and resolution, whereas human wet AMD is a chronic, progressive disorder characterized by sustained low-grade inflammation and gradual tissue remodeling over years [55]. Species-specific differences in immune system regulation, lipid metabolism, and extracellular deposit formation further complicate direct comparisons. Notably, mice do not develop drusen, the extracellular deposits that are hallmark features of early and intermediate human AMD but instead accumulate basal laminar deposits with only partial compositional overlap [40]. The complexity of gene expression changes in human AMD, influenced by genetic risk factors such as variants in CFH and ARMS2/HTRA1 and environmental exposures, is not fully recapitulated in the mouse model [133]. Furthermore, systemic biomarker profiles in humans are affected by aging, comorbidities, and genetic diversity, factors that are controlled or absent in laboratory mice, which may result in species-specific differences in circulating miRNAs and protein biomarkers [134].
Conclusions
The identification and validation of reliable biomarkers are crucial for improving the diagnosis, monitoring, and treatment of AMD, particularly in its dry form, which still lacks curative therapies. Different biomarkers, linked to oxidative stress, inflammation, complement activation, extracellular matrix remodeling, angiogenesis, and gut microbiota alterations, are conserved across human AMD and preclinical models, including genetically engineered mice, chemically induced models, and laser-induced neovascularization. Each model captures specific aspects of disease pathophysiology; chemically induced models offer a reproducible platform for dissecting molecular mechanisms. A notable overlap in biomarker signatures across retina/RPE–choroid tissue, blood, aqueous and vitreous humor, tear fluid, and feces emphasizes their translational relevance. Nonetheless, limitations persist, especially concerning the analysis of body fluids in animal models and the incomplete representation of human macular anatomy and disease chronicity. While the mouse model shares key angiogenic, inflammatory, and remodeling features with neovascular AMD, anatomical and temporal discrepancies, along with interspecies differences, require careful interpretation [67]. Thus, although preclinical models remain indispensable for mechanistic and biomarker discovery, their findings must be validated in human cohorts to ensure clinical applicability. Ultimately, integrating experimental and patient-derived data will advance the development of non-invasive, accessible, and personalized biomarker-driven strategies for AMD management.
References
- Wong, W. L. et al. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: a systematic review and meta-analysis. The Lancet Global Health 2, e106–e116 (2014). [CrossRef]
- Curcio, C. A., Kar, D., Owsley, C., Sloan, K. R. & Ach, T. Age-Related Macular Degeneration, a Mathematically Tractable Disease. Invest Ophthalmol Vis Sci 65, 4 (2024). [CrossRef]
- Choudhary, M. & Malek, G. A Review of Pathogenic Drivers of Age-Related Macular Degeneration, Beyond Complement, and Potential Endpoints to Test Therapeutic Interventions in Preclinical Studies. Adv Exp Med Biol 1185, 9–13 (2019). [CrossRef]
- Wong, J. H. C. et al. Exploring the pathogenesis of age-related macular degeneration: A review of the interplay between retinal pigment epithelium dysfunction and the innate immune system. Front Neurosci 16, 1009599 (2022). [CrossRef]
- Bowes Rickman, C., Farsiu, S., Toth, C. A. & Klingeborn, M. Dry Age-Related Macular Degeneration: Mechanisms, Therapeutic Targets, and Imaging. Invest Ophthalmol Vis Sci 54, ORSF68–ORSF80 (2013). [CrossRef]
- Fritsche, L. G. et al. A large genome-wide association study of age-related macular degeneration highlights contributions of rare and common variants. Nat Genet 48, 134–143 (2016). [CrossRef]
- Hageman, G. S. et al. Clinical validation of a genetic model to estimate the risk of developing choroidal neovascular age-related macular degeneration. Hum Genomics 5, 420–440 (2011). [CrossRef]
- Lad, E. M., Finger, R. P. & Guymer, R. Biomarkers for the Progression of Intermediate Age-Related Macular Degeneration. Ophthalmol Ther 12, 2917–2941 (2023). [CrossRef]
- Li, S. et al. Serum metabolite biomarkers for the early diagnosis and monitoring of age-related macular degeneration. Journal of Advanced Research (2024). [CrossRef]
- Borrelli, E. et al. Deep Learning in Neovascular Age-Related Macular Degeneration. Medicina (Kaunas) 60, 990 (2024). [CrossRef]
- Anderson, B. D., Lee, T. T., Bell, B. A., Wang, T. & Dunaief, J. L. Optimizing the sodium iodate model: effects of dose, gender, and age. Exp Eye Res 239, 109772 (2024). [CrossRef]
- Exploring the Role of Exosomal miRNA-146a-5p in Sodium Iodate-Induced Retinal Pigment Epithelial Dysfunction | IOVS | ARVO Journals (2024).
- Upadhyay, M. & Bonilha, V. L. Regulated cell death pathways in the sodium iodate model: Insights and implications for AMD. Experimental Eye Research 238, 109728 (2024). [CrossRef]
- Chen, Y., Liu, S., Hu, D., Xing, Y. & Shen, Y. N -methyl- N -nitrosourea-induced retinal degeneration in mice. Experimental Eye Research 121, 102–113 (2014).
- Zhao, J. et al. Aberrant Buildup of All-Trans-Retinal Dimer, a Nonpyridinium Bisretinoid Lipofuscin Fluorophore, Contributes to the Degeneration of the Retinal Pigment Epithelium. Investigative Ophthalmology & Visual Science 58, 1063–1075 (2017). [CrossRef]
- Zhang, J. et al. Protective effect of autophagy on human retinal pigment epithelial cells against lipofuscin fluorophore A2E: implications for age-related macular degeneration. Cell Death Dis 6, e1972–e1972 (2015). [CrossRef]
- Chen, X. et al. Hydrogen peroxide-induced oxidative damage and protective role of peroxiredoxin 6 protein via EGFR/ERK signaling pathway in RPE cells. Front Aging Neurosci 15, 1169211 (2023). [CrossRef]
- Kaczara, P., Sarna, T. & Burke, J. M. Dynamics of H2O2 Availability to ARPE-19 Cultures in Models of Oxidative Stress. Free Radic Biol Med 48, 1064–1070 (2010). [CrossRef]
- You, L., Zhao, W., Li, X., Yang, C. & Guo, P. Tyrosol protects RPE cells from H2O2-induced oxidative damage in vitro and in vivo through activation of the Nrf2/HO-1 pathway. European Journal of Pharmacology 991, 177316 (2025). [CrossRef]
- Hara, A. et al. A new model of retinal photoreceptor cell degeneration induced by a chemical hypoxia-mimicking agent, cobalt chloride. Brain Research 1109, 192–200 (2006). [CrossRef]
- Muñoz-Sánchez, J. & Chánez-Cárdenas, M. E. The use of cobalt chloride as a chemical hypoxia model. Journal of Applied Toxicology 39, 556–570 (2019). [CrossRef]
- Montezuma, S. R., Sobrin, L. & Seddon, J. M. Review of Genetics in Age Related Macular Degeneration. Seminars in Ophthalmology 22, 229–240 (2007). [CrossRef]
- Hanus, J., Anderson, C. & Wang, S. RPE Necroptosis in Response to Oxidative Stress and in AMD. Ageing Res Rev 24, 286–298 (2015). [CrossRef]
- Enzbrenner, A. et al. Sodium Iodate-Induced Degeneration Results in Local Complement Changes and Inflammatory Processes in Murine Retina. Int J Mol Sci 22, 9218 (2021). [CrossRef]
- Liu, R. T. et al. Inflammatory Mediators Induced by Amyloid-Beta in the Retina and RPE In Vivo: Implications for Inflammasome Activation in Age-Related Macular Degeneration. Invest Ophthalmol Vis Sci 54, 2225–2237 (2013). [CrossRef]
- Cervellati, F. et al. Hypoxia induces cell damage via oxidative stress in retinal epithelial cells. Free Radic Res 48, 303–312 (2014). [CrossRef]
- Lazzara, F. et al. Stabilization of HIF-1α in Human Retinal Endothelial Cells Modulates Expression of miRNAs and Proangiogenic Growth Factors. Front Pharmacol 11, 1063 (2020). [CrossRef]
- Tong, Y. et al. Comparative mechanistic study of RPE cell death induced by different oxidative stresses. Redox Biol 65, 102840 (2023). [CrossRef]
- Yang, H.-J. et al. 4-HNE induces proinflammatory cytokines of human retinal pigment epithelial cells by promoting extracellular efflux of HSP70. Exp Eye Res 188, 107792 (2019). [CrossRef]
- Penn, J. S. et al. Vascular Endothelial Growth Factor in Eye Disease. Prog Retin Eye Res 27, 331–371 (2008). [CrossRef]
- Spilsbury, K., Garrett, K. L., Shen, W. Y., Constable, I. J. & Rakoczy, P. E. Overexpression of vascular endothelial growth factor (VEGF) in the retinal pigment epithelium leads to the development of choroidal neovascularization. Am J Pathol 157, 135–144 (2000). [CrossRef]
- Luo, L. et al. Photoreceptor avascular privilege is shielded by soluble VEGF receptor-1. eLife 2, e00324 (2013). [CrossRef]
- Hanus, J., Anderson, C., Sarraf, D., Ma, J. & Wang, S. Retinal pigment epithelial cell necroptosis in response to sodium iodate. Cell Death Discov 2, 16054 (2016). [CrossRef]
- Gu, Y., Liu, W., Liu, G., Li, X. & Lu, P. Assessing the protective effects of cryptotanshinone on CoCl2-induced hypoxia in RPE cells. Mol Med Rep 24, 739 (2021). [CrossRef]
- Xing, Y. et al. Combination of Lactobacillus fermentum NS9 and aronia anthocyanidin extract alleviates sodium iodate-induced retina degeneration. Sci Rep 13, 8380 (2023). [CrossRef]
- Nguyen, Y. et al. Unveiling the gut-eye axis: how microbial metabolites influence ocular health and disease. Front Med (Lausanne) 11, 1377186 (2024). [CrossRef]
- Raoul, W. et al. CCL2/CCR2 and CX3CL1/CX3CR1 chemokine axes and their possible involvement in age-related macular degeneration. J Neuroinflammation 7, 87 (2010). [CrossRef]
- Vessey, K. A. et al. Ccl2/Cx3cr1 knockout mice have inner retinal dysfunction but are not an accelerated model of AMD. Invest Ophthalmol Vis Sci 53, 7833–7846 (2012). [CrossRef]
- Ross, R. J. et al. Immunological protein expression profile in Ccl2/Cx3cr1 deficient mice with lesions similar to age-related macular degeneration. Exp Eye Res 86, 675–683 (2008). [CrossRef]
- Imamura, Y. et al. Drusen, choroidal neovascularization, and retinal pigment epithelium dysfunction in SOD1-deficient mice: a model of age-related macular degeneration. Proc Natl Acad Sci U S A 103, 11282–11287 (2006). [CrossRef]
- Hashizume, K. et al. Retinal Dysfunction and Progressive Retinal Cell Death in SOD1-Deficient Mice. Am J Pathol 172, 1325–1331 (2008). [CrossRef]
- Zhu, Y. et al. Mice With a Combined Deficiency of Superoxide Dismutase 1 (Sod1), DJ-1 (Park7), and Parkin (Prkn) Develop Spontaneous Retinal Degeneration With Aging. Invest Ophthalmol Vis Sci 60, 3740–3751 (2019). [CrossRef]
- Lin, M. K. et al. HTRA1, an age-related macular degeneration protease, processes extracellular matrix proteins EFEMP1 and TSP1. Aging Cell 17, e12710 (2018). [CrossRef]
- Vierkotten, S., Muether, P. S. & Fauser, S. Overexpression of HTRA1 leads to ultrastructural changes in the elastic layer of Bruch’s membrane via cleavage of extracellular matrix components. PLoS One 6, e22959 (2011). [CrossRef]
- Chen, C.-Y. et al. N-Terminomics identifies HtrA1 cleavage of thrombospondin-1 with generation of a proangiogenic fragment in the polarized retinal pigment epithelial cell model of age-related macular degeneration. Matrix Biol 70, 84–101 (2018). [CrossRef]
- Hu, W. et al. Expression of VLDLR in the retina and evolution of subretinal neovascularization in the knockout mouse model’s retinal angiomatous proliferation. Invest Ophthalmol Vis Sci 49, 407–415 (2008). [CrossRef]
- Zinkernagel, M. S. et al. Association of the Intestinal Microbiome with the Development of Neovascular Age-Related Macular Degeneration. Sci Rep 7, 40826 (2017). [CrossRef]
- Bringer, M.-A., Gabrielle, P.-H., Bron, A. M., Creuzot-Garcher, C. & Acar, N. The gut microbiota in retinal diseases. Exp Eye Res 214, 108867 (2022). [CrossRef]
- Badia, A. et al. Transcriptomics analysis of Ccl2/Cx3cr1/Crb1rd8 deficient mice provides new insights into the pathophysiology of progressive retinal degeneration. Exp Eye Res 203, 108424 (2021). [CrossRef]
- Liu, J. et al. Replenishing IRAK-M expression in retinal pigment epithelium attenuates outer retinal degeneration. Sci Transl Med 16, eadi4125 (2024). [CrossRef]
- Jones, A. et al. Increased expression of multifunctional serine protease, HTRA1, in retinal pigment epithelium induces polypoidal choroidal vasculopathy in mice. Proc Natl Acad Sci U S A 108, 14578–14583 (2011). [CrossRef]
- Cai, X., Seal, S. & McGinnis, J. F. Sustained inhibition of neovascularization in vldlr-/- mice following intravitreal injection of cerium oxide nanoparticles and the role of the ASK1-P38/JNK-NF-κB pathway. Biomaterials 35, 249–258 (2014). [CrossRef]
- Zysset-Burri, D. C. et al. Associations of the intestinal microbiome with the complement system in neovascular age-related macular degeneration. NPJ Genom Med 5, 34 (2020). [CrossRef]
- Rowan, S. & Taylor, A. Gut microbiota modify risk for dietary glycemia-induced age-related macular degeneration. Gut Microbes 9, 452–457 (2018). [CrossRef]
- Shah, R. S., Soetikno, B. T., Lajko, M. & Fawzi, A. A. A Mouse Model for Laser-induced Choroidal Neovascularization. J Vis Exp 53502 (2015) doi:10.3791/53502. [CrossRef]
- Jimenez, A. I. et al. Optimization and characterization of an improved laser-induced choroidal neovascularization animal model for the study of retinal diseases. Investigative Ophthalmology & Visual Science 64, 2112 (2023).
- Gong, Y. et al. Optimization of an Image-Guided Laser-Induced Choroidal Neovascularization Model in Mice. PLOS ONE 10, e0132643 (2015). [CrossRef]
- Salas, A. et al. Neovascular Progression and Retinal Dysfunction in the Laser-Induced Choroidal Neovascularization Mouse Model. Biomedicines 11, 2445 (2023). [CrossRef]
- Liu, Y.-S. et al. Comparative Analysis of Molecular Landscape in Mouse Models and Patients Reveals Conserved Inflammation Pathways in Age-Related Macular Degeneration. Investigative Ophthalmology & Visual Science 65, 13 (2024). [CrossRef]
- Wolf, J. et al. Comparative transcriptome analysis of human and murine choroidal neovascularization identifies fibroblast growth factor inducible-14 as phylogenetically conserved mediator of neovascular age-related macular degeneration. Biochim Biophys Acta Mol Basis Dis 1868, 166340 (2022). [CrossRef]
- Toma, H. S., Barnett, J. M., Penn, J. S. & Kim, S. J. Improved assessment of laser-induced choroidal neovascularization. Microvasc Res 80, 295–302 (2010). [CrossRef]
- Iwanishi, H. et al. Delayed regression of laser-induced choroidal neovascularization in TNFα-null mice. Journal of Cellular and Molecular Medicine 26, 5315–5325 (2022). [CrossRef]
- Lambert, V. et al. Laser-induced choroidal neovascularization model to study age-related macular degeneration in mice. Nat Protoc 8, 2197–2211 (2013). [CrossRef]
- Hu, Y. et al. Proteotranscriptomic analyses reveal distinct interferon-beta signaling pathways and therapeutic targets in choroidal neovascularization. Front. Immunol. 14, (2023). [CrossRef]
- Apte, R. S., Richter, J., Herndon, J. & Ferguson, T. A. Macrophages Inhibit Neovascularization in a Murine Model of Age-Related Macular Degeneration. PLoS Med 3, e310 (2006). [CrossRef]
- Brandli, A., Khong, F. L., Kong, R. C. K., Kelly, D. J. & Fletcher, E. L. Transcriptomic analysis of choroidal neovascularization reveals dysregulation of immune and fibrosis pathways that are attenuated by a novel anti-fibrotic treatment. Sci Rep 12, 859 (2022). [CrossRef]
- Kiel, C. et al. A Circulating MicroRNA Profile in a Laser-Induced Mouse Model of Choroidal Neovascularization. Int J Mol Sci 21, 2689 (2020). [CrossRef]
- Moshtaghion, S. M. M. et al. VEGF in Tears as a Biomarker for Exudative Age-Related Macular Degeneration: Molecular Dynamics in a Mouse Model and Human Samples. Int J Mol Sci 26, 3855 (2025). [CrossRef]
- dos Santos, F. M. et al. Vitreous humor proteome: unraveling the molecular mechanisms underlying proliferative and neovascular vitreoretinal diseases. Cell Mol Life Sci 80, 22 (2022). [CrossRef]
- Oca, A. I. et al. Predictive Biomarkers of Age-Related Macular Degeneration Response to Anti-VEGF Treatment. Journal of Personalized Medicine 11, 1329 (2021). [CrossRef]
- Ratnapriya, R. & Chew, E. Y. Age-related macular degeneration-clinical review and genetics update. Clin Genet 84, 160–166 (2013). [CrossRef]
- Andriessen, E. M. et al. Gut microbiota influences pathological angiogenesis in obesity-driven choroidal neovascularization. EMBO Mol Med 8, 1366–1379 (2016). [CrossRef]
- Zhang, H. & Mo, Y. The gut-retina axis: a new perspective in the prevention and treatment of diabetic retinopathy. Front. Endocrinol. 14, (2023). [CrossRef]
- Bakri, S. J. et al. Geographic atrophy: Mechanism of disease, pathophysiology, and role of the complement system. J Manag Care Spec Pharm 29, S2–S11 (2023). [CrossRef]
- Lau, L.-I., Liu, C. J. & Wei, Y.-H. Increase of 8-hydroxy-2’-deoxyguanosine in aqueous humor of patients with exudative age-related macular degeneration. Invest Ophthalmol Vis Sci 51, 5486–5490 (2010). [CrossRef]
- Shen, J. K. et al. Oxidative damage in age-related macular degeneration. Histol Histopathol 22, 1301–1308 (2007). [CrossRef]
- Crabb, J. W. et al. Drusen proteome analysis: an approach to the etiology of age-related macular degeneration. Proc Natl Acad Sci U S A 99, 14682–14687 (2002). [CrossRef]
- Lu, L. et al. Synthesis and structural characterization of carboxyethylpyrrole-modified proteins: mediators of age-related macular degeneration. Bioorg Med Chem 17, 7548–7561 (2009). [CrossRef]
- Gu, X. et al. Carboxyethylpyrrole protein adducts and autoantibodies, biomarkers for age-related macular degeneration. J Biol Chem 278, 42027–42035 (2003). [CrossRef]
- Renganathan, K. et al. CEP Biomarkers as Potential Tools for Monitoring Therapeutics. PLoS One 8, e76325 (2013). [CrossRef]
- Ethen, C. M., Reilly, C., Feng, X., Olsen, T. W. & Ferrington, D. A. Age-related macular degeneration and retinal protein modification by 4-hydroxy-2-nonenal. Invest Ophthalmol Vis Sci 48, 3469–3479 (2007). [CrossRef]
- Gu, J. et al. Proteomic and genomic biomarkers for age-related macular degeneration. Adv Exp Med Biol 664, 411–417 (2010). [CrossRef]
- Totan, Y. et al. Oxidative macromolecular damage in age-related macular degeneration. Curr Eye Res 34, 1089–1093 (2009). [CrossRef]
- Nita, M., Strzałka-Mrozik, B., Grzybowski, A., Mazurek, U. & Romaniuk, W. Age-related macular degeneration and changes in the extracellular matrix. Med Sci Monit 20, 1003–1016 (2014). [CrossRef]
- Hussain, A. A., Lee, Y., Zhang, J.-J. & Marshall, J. Disturbed Matrix Metalloproteinase Activity of Bruch’s Membrane in Age-Related Macular Degeneration. Investigative ophthalmology & visual science 52, 4459- (2011). [CrossRef]
- Hageman, G. S. et al. A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc Natl Acad Sci U S A 102, 7227–7232 (2005). [CrossRef]
- Reynolds, R. et al. Plasma complement components and activation fragments: associations with age-related macular degeneration genotypes and phenotypes. Invest Ophthalmol Vis Sci 50, 5818–5827 (2009). [CrossRef]
- Ildefonso, C. J., Biswal, M. R., Ahmed, C. M. & Lewin, A. S. The NLRP3 Inflammasome and its Role in Age-Related Macular Degeneration. Adv Exp Med Biol 854, 59–65 (2016). [CrossRef]
- Szaflik, J. P. et al. DNA damage and repair in age-related macular degeneration. Mutat Res 669, 169–176 (2009). [CrossRef]
- Martinez, B. & Peplow, P. V. MicroRNAs as diagnostic and prognostic biomarkers of age-related macular degeneration: advances and limitations. Neural Regen Res 16, 440–447 (2020). [CrossRef]
- Berber, P., Grassmann, F., Kiel, C. & Weber, B. H. F. An Eye on Age-Related Macular Degeneration: The Role of MicroRNAs in Disease Pathology. Mol Diagn Ther 21, 31–43 (2017). [CrossRef]
- Cruz-Aguilar, M., Groman-Lupa, S. & Jiménez-Martínez, M. C. MicroRNAs as potential biomarkers and therapeutic targets in age-related macular degeneration. Front Ophthalmol (Lausanne) 3, 1023782 (2023). [CrossRef]
- Winiarczyk, M. et al. Tear film proteome in age-related macular degeneration. Graefes Arch Clin Exp Ophthalmol 256, 1127–1139 (2018). [CrossRef]
- Kaarniranta, K., Tokarz, P., Koskela, A., Paterno, J. & Blasiak, J. Autophagy regulates death of retinal pigment epithelium cells in age-related macular degeneration. Cell Biol Toxicol 33, 113–128 (2017). [CrossRef]
- Zhou, L. & Beuerman, R. W. The power of tears: how tear proteomics research could revolutionize the clinic. Expert Rev Proteomics 14, 189–191 (2017). [CrossRef]
- Zhou, L. et al. Identification of tear fluid biomarkers in dry eye syndrome using iTRAQ quantitative proteomics. J Proteome Res 8, 4889–4905 (2009). [CrossRef]
- Huang, K. et al. Proteomics approach identifies aqueous humor biomarkers in retinal diseases. Commun Med (Lond) 5, 134 (2025). [CrossRef]
- Rinsky, B. et al. Analysis of the Aqueous Humor Proteome in Patients With Age-Related Macular Degeneration. Invest Ophthalmol Vis Sci 62, 18 (2021). [CrossRef]
- Baek, J.-H. et al. Quantitative proteomic analysis of aqueous humor from patients with drusen and reticular pseudodrusen in age-related macular degeneration. BMC Ophthalmol 18, 289 (2018). [CrossRef]
- Yoon, E. G., Nam, K. T., Choi, M., Choi, K.-E. & Yun, C. Aqueous Humor Levels of Vascular Endothelial Growth Factor in Patients With Dry Age-Related Macular Degeneration and Subretinal Drusenoid Deposits. Invest Ophthalmol Vis Sci 66, 10 (2025). [CrossRef]
- García-Quintanilla, L. et al. Recent Advances in Proteomics-Based Approaches to Studying Age-Related Macular Degeneration: A Systematic Review. Int J Mol Sci 23, 14759 (2022). [CrossRef]
- Huang, H.-Y., Wang, J., Qin, B. & Tan, Y. Investigating the causal link between gut microbiota and dry age-related macular degeneration: a bidirectional Mendelian randomization study. Int J Ophthalmol 17, 1723–1730 (2024). [CrossRef]
- Xiao, J., Zhang, J. Y., Luo, W., He, P. C. & Skondra, D. The Emerging Role of Gut Microbiota in Age-Related Macular Degeneration. Am J Pathol 193, 1627–1637 (2023). [CrossRef]
- Ambati, J. & Fowler, B. J. Mechanisms of age-related macular degeneration. Neuron 75, 26–39 (2012). [CrossRef]
- Wong, D. T. et al. Imaging Biomarkers and Their Impact on Therapeutic Decision-Making in the Management of Neovascular Age-Related Macular Degeneration. Ophthalmologica 244, 265–280 (2021). [CrossRef]
- Kaiser, P. K. et al. RETINAL FLUID AND THICKNESS AS MEASURES OF DISEASE ACTIVITY IN NEOVASCULAR AGE-RELATED MACULAR DEGENERATION. Retina 41, 1579–1586 (2021). [CrossRef]
- Rispoli, M. et al. Practical guidance for imaging biomarkers in exudative age-related macular degeneration. Surv Ophthalmol 68, 615–627 (2023). [CrossRef]
- Litts, K. M. et al. Quantitative Analysis of Outer Retinal Tubulation in Age-Related Macular Degeneration From Spectral-Domain Optical Coherence Tomography and Histology. Investigative Ophthalmology & Visual Science 57, 2647–2656 (2016). [CrossRef]
- Hammer, M. et al. Fundus autofluorescence intensity, lifetime, and spectral imaging in age-related macular degeneration. Experimental Eye Research 258, 110500 (2025). [CrossRef]
- Schlecht, A. et al. Transcriptomic Characterization of Human Choroidal Neovascular Membranes Identifies Calprotectin as a Novel Biomarker for Patients with Age-Related Macular Degeneration. The American Journal of Pathology 190, 1632–1642 (2020). [CrossRef]
- ElShelmani, H., Brennan, I., Kelly, D. J. & Keegan, D. Differential Circulating MicroRNA Expression in Age-Related Macular Degeneration. Int J Mol Sci 22, 12321 (2021). [CrossRef]
- Kauppinen, A., Paterno, J. J., Blasiak, J., Salminen, A. & Kaarniranta, K. Inflammation and its role in age-related macular degeneration. Cell Mol Life Sci 73, 1765–1786 (2016). [CrossRef]
- Beatty, S., Koh, H., Phil, M., Henson, D. & Boulton, M. The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv Ophthalmol 45, 115–134 (2000). [CrossRef]
- Proteomics approach identifies aqueous humor biomarkers in retinal diseases | Communications Medicine. https://www.nature.com/articles/s43856-025-00862-2.
- Caban, M., Owczarek, K. & Lewandowska, U. The Role of Metalloproteinases and Their Tissue Inhibitors on Ocular Diseases: Focusing on Potential Mechanisms. Int J Mol Sci 23, 4256 (2022). [CrossRef]
- Wilson, S. et al. Correlation of Aqueous, Vitreous, and Serum Protein Levels in Patients With Retinal Diseases. Transl Vis Sci Technol 12, 9 (2023). [CrossRef]
- Nobl, M. et al. Proteomics of vitreous in neovascular age-related macular degeneration. Experimental Eye Research 146, 107–117 (2016). [CrossRef]
- Fritsche, L. G. et al. Seven new loci associated with age-related macular degeneration. Nat Genet 45, 433–439 (2013). [CrossRef]
- Kaur, I., Rathi, S. & Chakrabarti, S. Variations in TIMP3 are associated with age-related macular degeneration. Proceedings of the National Academy of Sciences 107, E112–E113 (2010). [CrossRef]
- Churchill, A. J. et al. VEGF polymorphisms are associated with neovascular age-related macular degeneration. Hum Mol Genet 15, 2955–2961 (2006). [CrossRef]
- Yu, C. et al. Predictive Performance of an Updated Polygenic Risk Score for Age-Related Macular Degeneration. Ophthalmology 131, 880–891 (2024). [CrossRef]
- Schiavone, N. et al. Exploring the Gut Microbiota–Retina Axis: Implications for Health and Disease. Microorganisms 13, 1101 (2025). [CrossRef]
- Shu, Y. et al. MiR-21-5p promotes RPE cell necroptosis by targeting Peli1 in a rat model of AMD. In Vitro Cell Dev Biol Anim (2025) doi:10.1007/s11626-025-01064-9. [CrossRef]
- Merkle, C. W., Augustin, M., Harper, D. J., Glösmann, M. & Baumann, B. Degeneration of Melanin-Containing Structures Observed Longitudinally in the Eyes of SOD1-/- Mice Using Intensity, Polarization, and Spectroscopic OCT. Transl Vis Sci Technol 11, 28 (2022). [CrossRef]
- Zhang, Y. et al. Alterations of the intestinal microbiota in age-related macular degeneration. Front Microbiol 14, 1069325 (2023). [CrossRef]
- Luo, W. & Skondra, D. Implication of gut microbiome in age-related macular degeneration. Neural Regen Res 18, 2699–2700 (2023). [CrossRef]
- Fabian-Jessing, B. K. et al. Animal Models of Choroidal Neovascularization: A Systematic Review. Invest Ophthalmol Vis Sci 63, 11 (2022). [CrossRef]
- Choi, Y. J., Lim, D., Byeon, S. H., Shin, E.-C. & Chung, H. Chemokine Receptor Profiles of T Cells in Patients with Age-Related Macular Degeneration. Yonsei Med J 63, 357–364 (2022). [CrossRef]
- Sennlaub, F. et al. CCR2(+) monocytes infiltrate atrophic lesions in age-related macular disease and mediate photoreceptor degeneration in experimental subretinal inflammation in Cx3cr1 deficient mice. EMBO Mol Med 5, 1775–1793 (2013). [CrossRef]
- Masli, S., Sheibani, N., Cursiefen, C. & Zieske, J. Matricellular protein thrombospondin : influence on ocular angiogenesis, wound healing and immuneregulation. Curr Eye Res 39, 759–774 (2014). [CrossRef]
- Abokyi, S., To, C.-H., Lam, T. T. & Tse, D. Y. Central Role of Oxidative Stress in Age-Related Macular Degeneration: Evidence from a Review of the Molecular Mechanisms and Animal Models. Oxidative Medicine and Cellular Longevity 2020, 7901270 (2020). [CrossRef]
- Huberman, A. D. & Niell, C. M. What can mice tell us about how vision works? Trends Neurosci 34, 464–473 (2011). [CrossRef]
- Voigt, A. P. et al. Single-cell transcriptomics of the human retinal pigment epithelium and choroid in health and macular degeneration. Proceedings of the National Academy of Sciences 116, 24100–24107 (2019). [CrossRef]
- Chakravarthy, U. et al. Clinical risk factors for age-related macular degeneration: a systematic review and meta-analysis. BMC Ophthalmol 10, 31 (2010). [CrossRef]
Figure 1.
Schematic representation of the main findings regarding biomarkers in chemically -induced AMD mice models using ocular tissues/fluids as a source. Created using BioRender.com.
Figure 1.
Schematic representation of the main findings regarding biomarkers in chemically -induced AMD mice models using ocular tissues/fluids as a source. Created using BioRender.com.
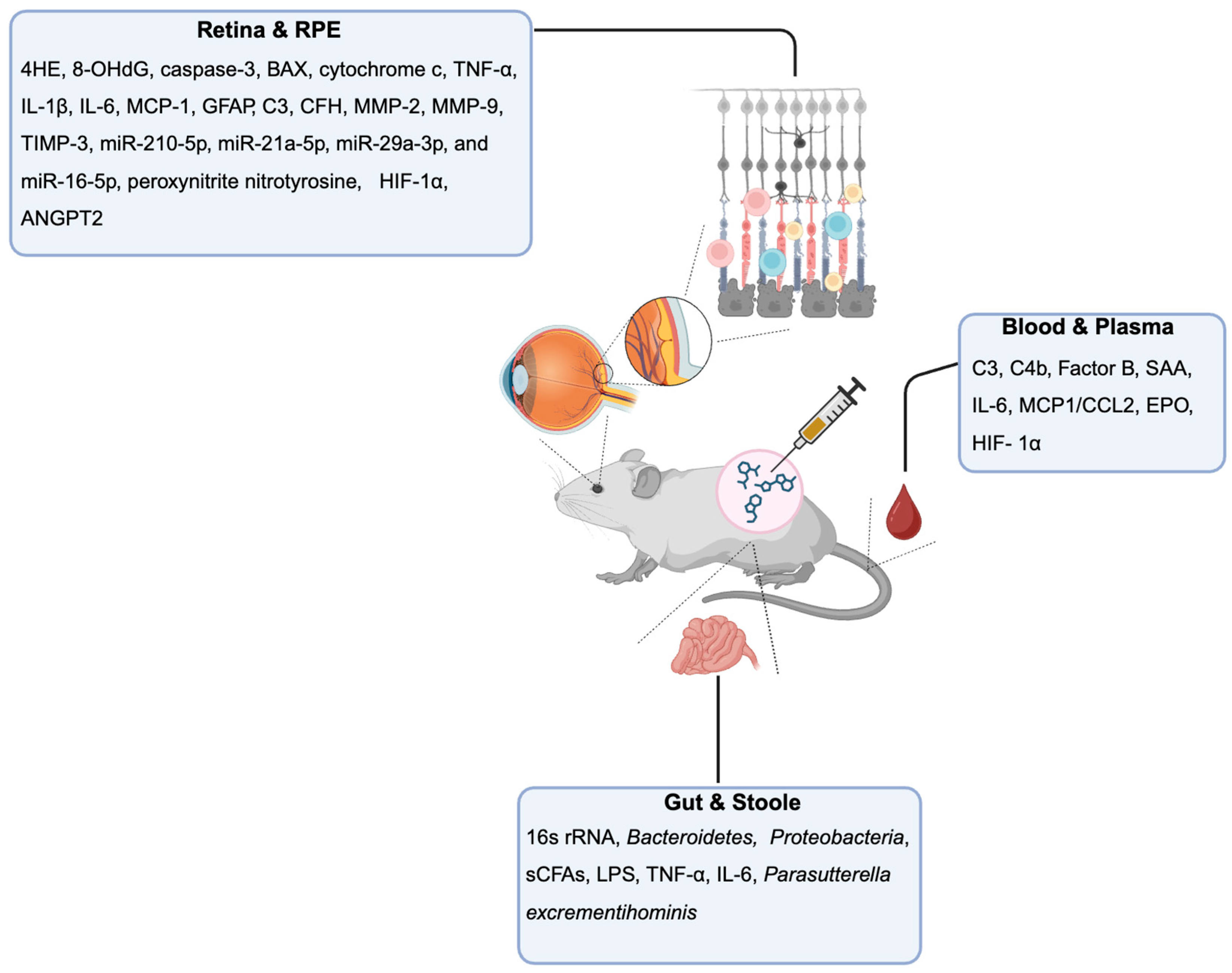
Figure 2.
Schematic representation of the main findings regarding biomarkers in genetic AMD mice models using ocular tissues/fluids as a source. Created using BioRender.com.
Figure 2.
Schematic representation of the main findings regarding biomarkers in genetic AMD mice models using ocular tissues/fluids as a source. Created using BioRender.com.
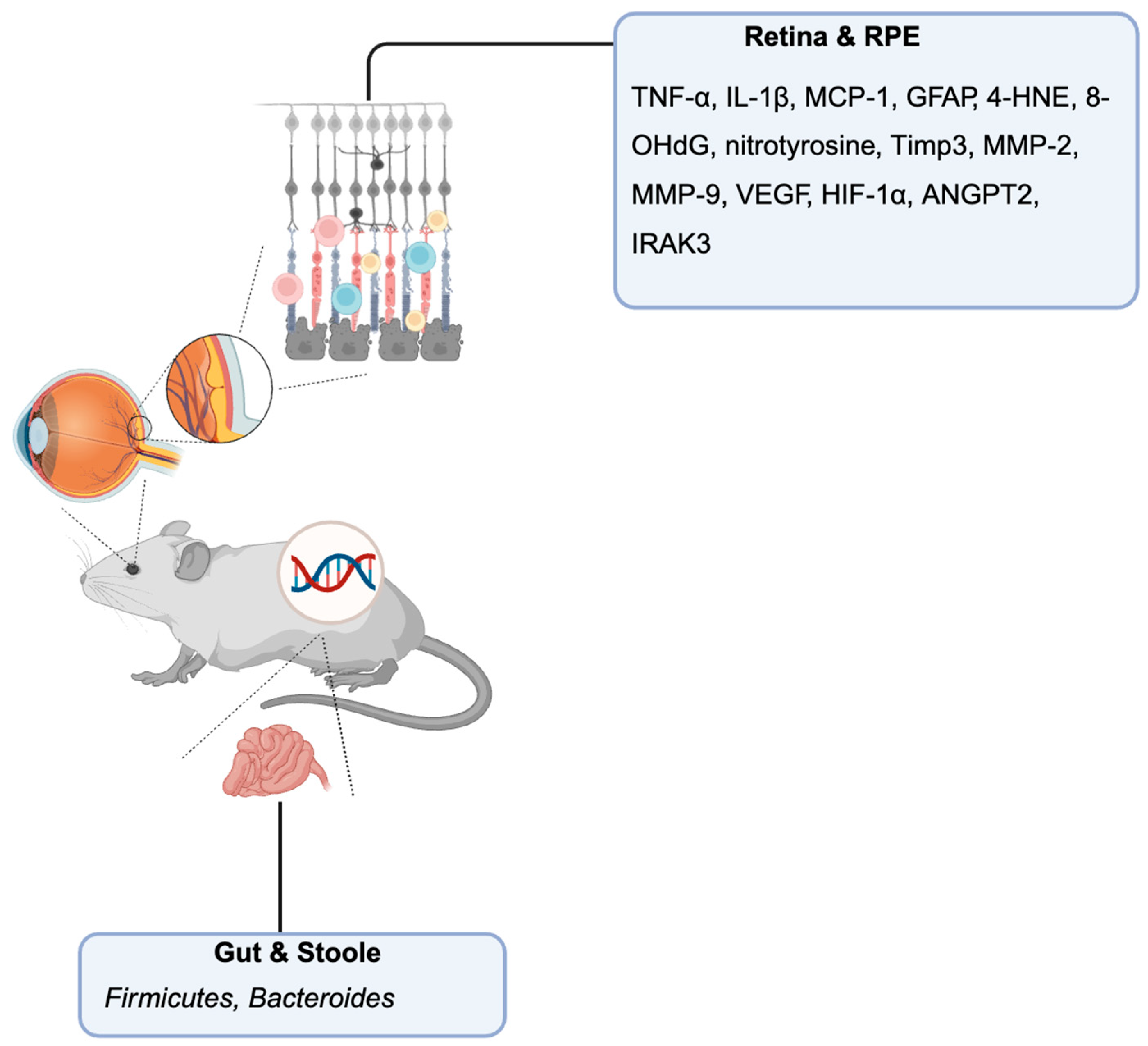
Figure 3.
Schematic representation of the main findings regarding biomarkers in laser-induced neovascularization model using ocular tissues/fluids as a source. Created using BioRender.com.
Figure 3.
Schematic representation of the main findings regarding biomarkers in laser-induced neovascularization model using ocular tissues/fluids as a source. Created using BioRender.com.
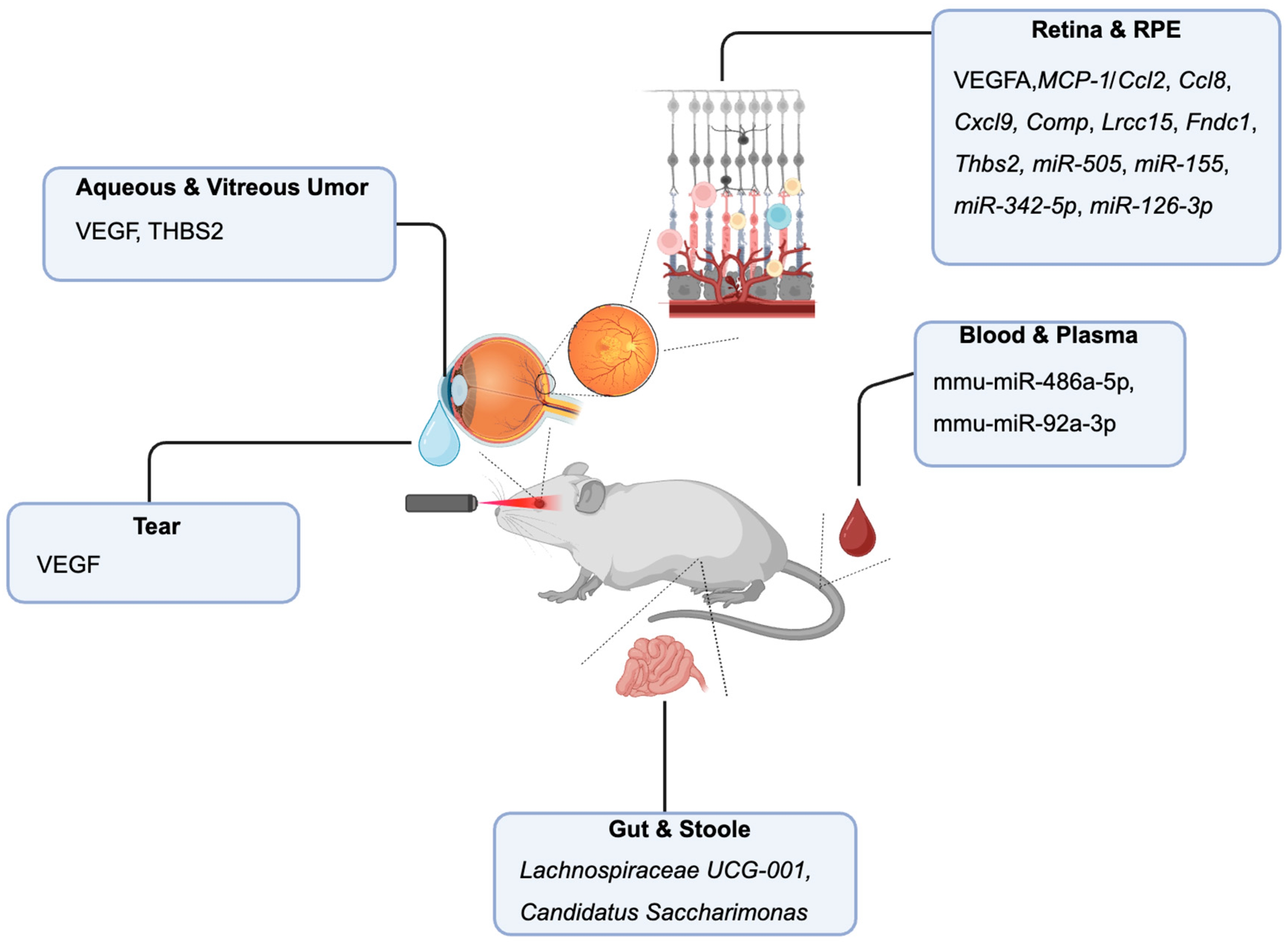
Figure 4.
Schematic representation of the main findings regarding biomarkers in human dry AMD using ocular tissues/fluids as a source. Created using BioRender.com.
Figure 4.
Schematic representation of the main findings regarding biomarkers in human dry AMD using ocular tissues/fluids as a source. Created using BioRender.com.
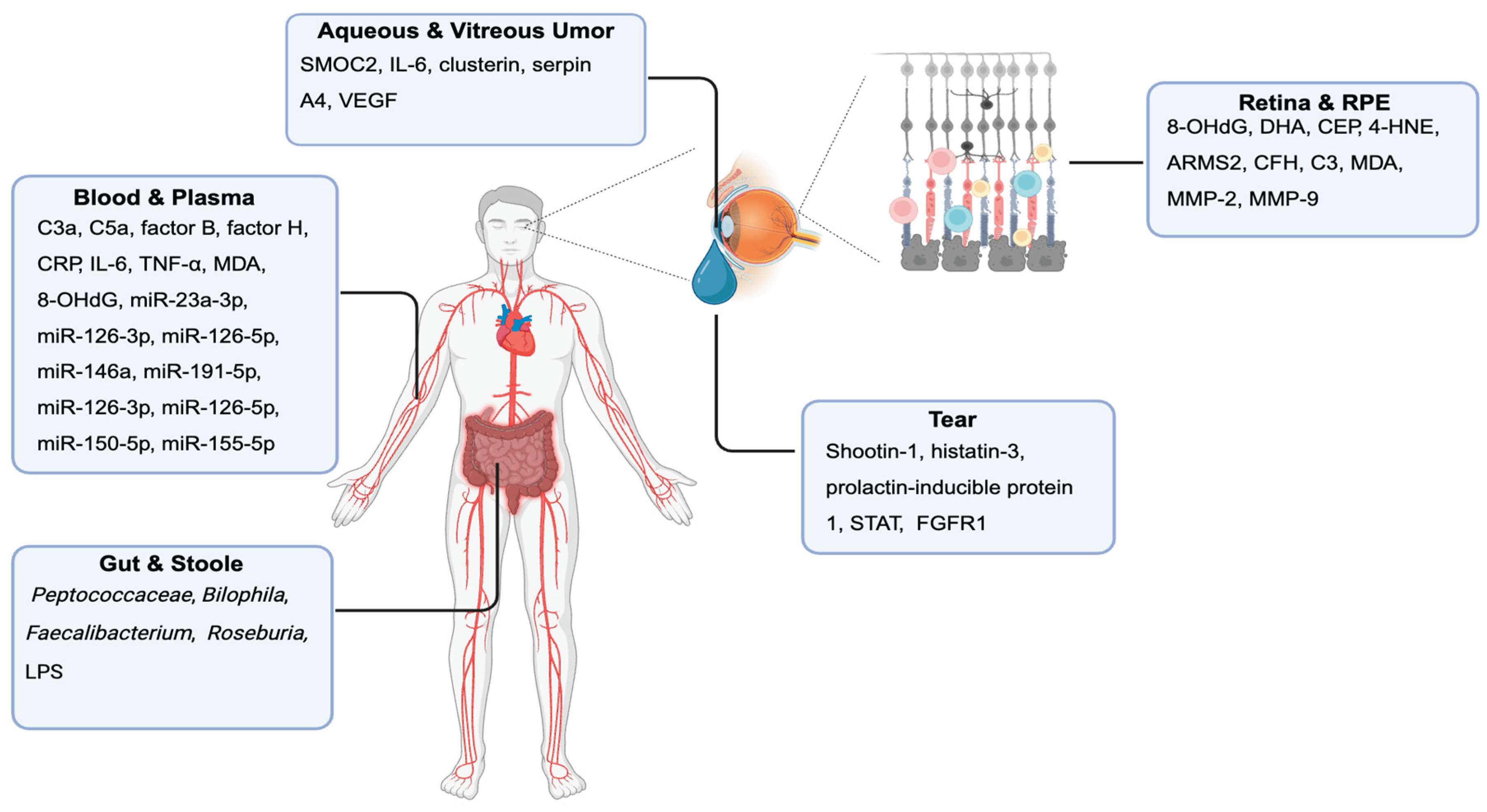
Figure 5.
Schematic representation of the main findings regarding biomarkers in human wet AMD using ocular tissues/fluids as a source. Created using BioRender.com.
Figure 5.
Schematic representation of the main findings regarding biomarkers in human wet AMD using ocular tissues/fluids as a source. Created using BioRender.com.
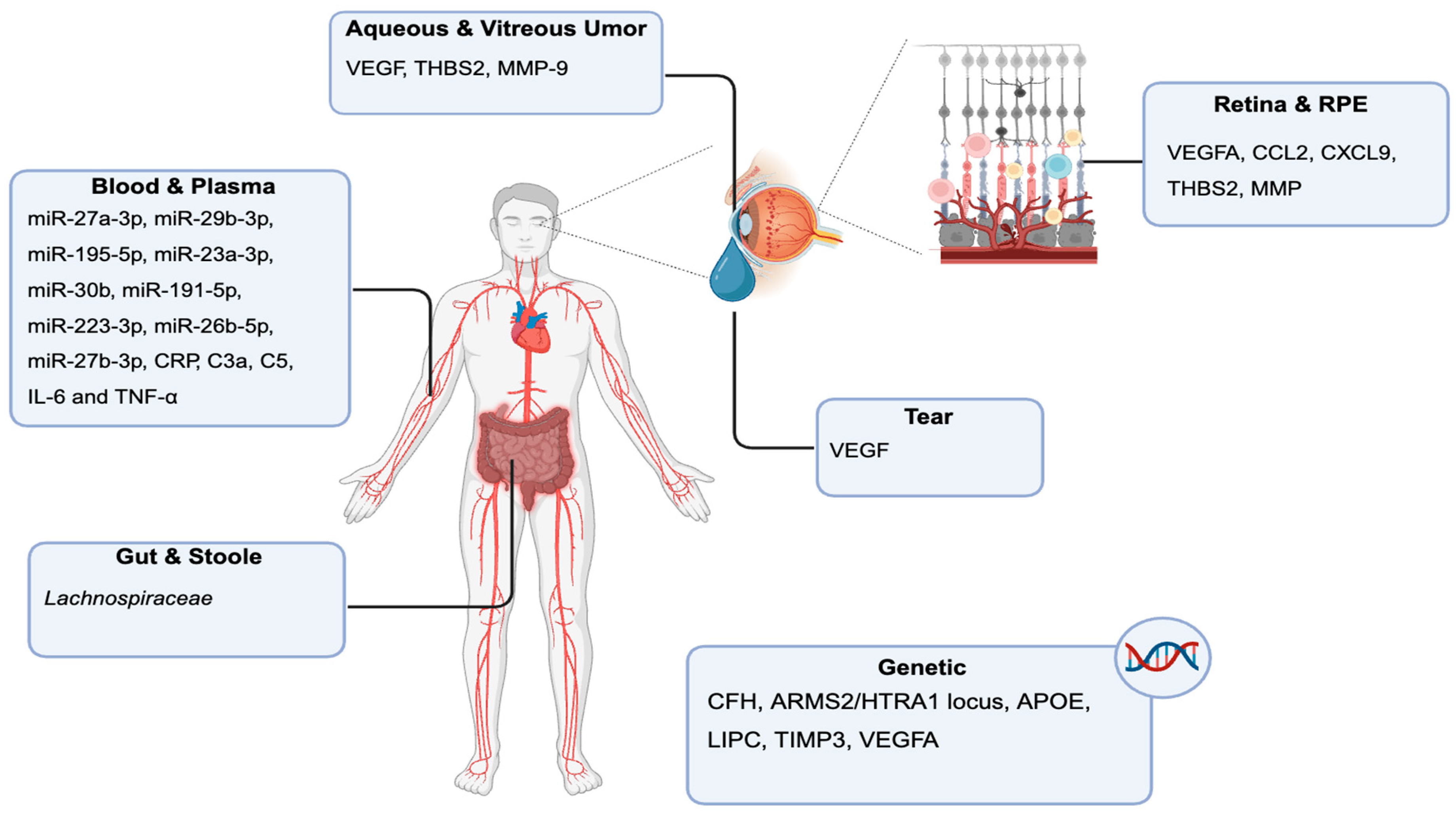
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



