
Submitted:
29 July 2025
Posted:
30 July 2025
You are already at the latest version
Abstract
Exercise can have a positive therapeutic effect in people with chronic obstructive pulmonary disease (COPD), where skeletal muscle dysfunction is strongly associated with poor prognosis. However, the precise effect of exercise on skeletal muscle mitochondrial biology in COPD is not yet understood. COPD is an inflammatory lung condition that is characterised by obstructed airflow from the lungs. Around 390 million people are affected across the globe, with exposure to environmental tobacco smoke being the greatest risk factor. While it is described as a chronic disease, complications can lead to acute respiratory failure. Different types of physical exercise may have a potential therapeutic benefit, but COPD often limits such activities in affected people. Here we explored the effect that a session of eccentric or concentric cycling can induce in the proteome of mitochondria isolated from skeletal muscle of COPD individuals. We report changes in the abundance of proteins that have been associated with mitochondrial ultrastructure and morphology, with some sex-specific changes, while others are common to both male and female participants. We found that the changes in protein expression are distinct between the different types of cycling exercise and that there are more proteins with significantly altered abundance in response to both types of exercise, in males. Rapid, exercise specific changes in the skeletal muscle mitochondrial proteome clearly demonstrate that treatment strategies for this common and debilitating condition should be managed differently in males and females. Immediate proteomic changes have not been measured previously and could be interpreted as a more ‘reflexive’ effect of exercise, least moderated by background heterogeneity. Our data show that all exercise types probably do not elicit the same quality of therapeutic benefit and therefore should be evaluated carefully before recommendation to people with COPD.
Keywords:
COPD
; mitochondria
; proteomics
; eccentric
; concentric
; skeletal muscle
; sex differences
1. Introduction
Chronic obstructive pulmonary disease (COPD) is a global disease that is estimated to impact 10.3% of people aged 30 to 79 years old, which amounts to over 390 million people[1]. The 2022 report published by the Global Initiative for Chronic Obstructive Lung Disease (GOLD) defines COPD as a “common, preventable, and treatable disease that is characterized by persistent respiratory symptoms and airflow limitation[2]. The most significant risk factor for COPD is exposure to environmental tobacco smoke[3]. The major pathophysiology of COPD is the narrowing of the bronchioles in the lungs, due to inflammation and fibrosis, resulting in impaired airflow and gas exchange[4]. The impaired airflow leads to the characteristic symptom of dyspnoea, while acute respiratory failure is a common event associated with severe exacerbation of the disease[5].
Hypoxia of lung alveoli contributes to the hypoxemia observed in COPD patients[6], which is linked to muscle dysfunction and wasting observed in COPD[7]. Reduced blood oxygen is likely to impact the function of the mitochondria, as they depend upon oxygen delivery to cells, where ATP is produced by oxidative phosphorylation. COPD patients with a low BMI (which could be attributed to sarcopenia) have reduced respiratory chain function in the vastus lateralis when compared with COPD patients with a normal BMI and healthy control trial participants[8]. It has been speculated that skeletal muscle dysfunction could be associated with abnormal mitochondrial adaptation to episodes of cell hypoxia. Mitochondrial oxygen consumption and ATP production in state 3 respiration were both decreased in the vastus lateralis and the external intercostalis of mild to moderate COPD patients undergoing surgery for lung cancer. Also, cytochrome c oxidase activity showed a significant negative relationship with arterial oxygen tension[9].
Mitochondrial function has been clearly connected with COPD pathology since increased levels of uncoupling protein 3 (UCP3) transcription have been measured[8], increased reactive oxygen species (ROS) in both skeletal muscle and bronchial tissue[9,10], decreased mitochondrial membrane potential in bronchial tissue of GOLD grade 2 COPD patient bronchial tissue, and decreased expression of respiratory complex proteins in the bronchial tissue mitochondria of both GOLD grade 2 COPD patients and GOLD grade 3/4 skeletal muscle mitochondria[10]. The role of the inner mitochondrial fusion protein OPA1 has also been studied with respect to the pathophysiology of COPD; short OPA1 isoforms maintain mitochondrial bioenergetic function and cristae structure and appear to be significantly elevated in COPD patient lung tissue[11,12].
While COPD limits the capacity for exercise of those living with the disease[13], exercise therapy has long been considered for the physical and psychosocial wellbeing of patients with this condition[14]. Contemporary studies with exercise regimes that have a low cardiorespiratory demand also demonstrate the benefits of exercise as a therapeutic intervention for COPD[15]. The effectiveness of exercise therapy for other chronic respiratory diseases, including cystic fibrosis and asthma in children, has been reviewed[16]. Until now, changes to the mitochondrial proteome, which could be associated with improved skeletal muscle function resulting from exercise, remains largely undefined in individuals with respiratory diseases such as COPD.
When a muscle is used in exercise then its contraction can be isometric, where its length does not change during contraction, or isotonic, where the muscle can become either shorter or longer while tension is kept constant[17]. Isotonic contraction is defined as concentric, where the muscle shortens while producing force, or eccentric, when it lengthens. As eccentric cycling has been shown to have a lower cardiopulmonary demand than concentric exercise when mechanical loads are matched[18], it presents an easier way to exercise for people who have difficulty with breathing and muscle strength (e.g. in COPD)[19].
Eccentric exercise promotes the highest forces with the lowest energy requirement[20], when compared with concentric exercise. There have been efforts to understand the impact of eccentric exercise on mitochondria in humans and other animals. The results have been mixed, where mitochondrial calcium content has been shown to be elevated, which leads to increased mitochondrial permeability transition pore (MPTP) sensitivity[21]. However, other aspects of mitochondrial physiology remain unaltered, including respiratory chain activity of skeletal muscle from participants with COPD[22], despite suggestions that the high levels of muscle damage associated with eccentric exercise might lead to significant changes[23]. This is in comparison to the impact of concentric exercise on mitochondrial physiology, where a study of rats found that 20 days of concentric exercise training increased maximal respiration in gastrocnemius and soleus muscles but eccentric training did not, and eccentric training caused an increase in H2O2 output in vastus intermedius muscles[24].
Proteomic profiling has been used in studies of COPD, both to develop a deeper understanding of the pathophysiological changes in disease relevant tissues[25,26,27], and as a potential diagnostic tool from minimally invasive sample collection procedures[28]. The mitochondrial proteome has also been widely studied across a range of model organisms[29,30,31,32], as well as different states of human health and disease[33,34,35]. Contemporary methods of label-free mass spectrometry are increasingly used for quantitative proteomic studies in place of more expensive and often time-consuming sample labelling methods[36]. Orbitrap technology has been widely adopted in proteomic studies since the early 2000s and has allowed for high-sensitivity, high-resolution studies of biological samples[37,38]. The acquired datasets can be analysed downstream using bioinformatics tools such as STRING-db and KEGG, offering novel biological insights based on the proteomic profile of the sample in question[39,40]. These bioinformatics data can then be pursued further by classical molecular biology and biochemical experimentation.
Here we measured changes in skeletal muscle mitochondria in response to cycling. We quantified protein level differences of mitochondrial fractions in response to i) eccentric or concentric exercise, and ii) sex of the participant. Label-free mass spectrometry and western blot were used to determine the proteome of isolated mitochondria. Biochemical assays, including ATPase activity and lipid peroxidation (MDA), were used to further characterise the mitochondrial changes in response to the exercise.
2. Methods
2.1. COPD Patient Exercise and Tissue Sampling
COPD quadriceps biopsy samples were collected as part of a previous study[15]. Full details of participants, exercise regime and sampling methodology can be found in the earlier study.
The protocol was approved by the Leicestershire South Regional Ethics Committee (Identifier: IRAS 214536). All participants gave informed written consent before taking part. Separate informed written consent was obtained for biopsy sampling.
Local area ethical approval was applied for and approved by the University of Nottingham approval number 3628 220728 HTA.
In brief, participants performed 20-minute bouts of eccentric and concentric exercise at 65% peak workload. Biopsies were obtained at rest and 60 s after exercise cessation from incisions at least 2 cm apart. Biopsies for eccentric exercise and concentric exercise were obtained from opposite legs. Approximately 100 mg of tissue was removed under local anaesthetic using the modified Bergstrom technique. Tissue was frozen immediately in liquid nitrogen and stored at -80°C until analysis.
2.2. Mitochondrial Isolation
Mitochondria were isolated from 25 mg of tissue per biopsy sample via differential centrifugation, using previously described standard methods[41]. Male and female samples for pre- and post-exercise condition were chosen (n=3 biological replicates, 24 samples in total). In brief, tissue was lysed in mitochondrial isolation buffer with a Dounce homogeniser. The lysate was centrifuged at 850 g for 10 mins to remove insoluble material. The supernatant was centrifuged at 1050 g for 10 mins, and the supernatant collected before centrifugation at 10,000 g for 15 mins to generate the mitochondrial pellet, and subsequent supernatant was discarded.
2.3. SDS-PAGE
The mitochondrial pellet generated from each 25 mg tissue sample was subject to SDS-PAGE. The pellet was resuspended in 5 μL PBS, 3 μL DTT, and 3 μL LDS. The resuspended sample was loaded onto polyacrylamide pre-cast gels (NuPAGE™ 4 to 12%, Bis-Tris, 1.0 mm, Mini Protein Gel, 12-well). A fixed voltage (200 V) was applied to the gel until the dye front reached 2 cm into the gel and the gel was then stained with ProtoBlue Safe Coomassie G-250. The gels were de-stained with deionised water, and stained protein bands were excised from the gel and added to 1.5 mL microfuge tubes.
2.4. Label-Free Mass Spectrometry
The mitochondrial protein samples were sent to the Centre of Excellence for Mass Spectrometry at the University of York for analysis. Protein was in-gel digested overnight at 37oC with 0.2 μg sequencing grade trypsin and Lys-C protease mixture (Promega V5073), following reduction 10 mM dithioerythritol and S-carbamidomethylation with 50 mM iodoacetamide.
Extracted peptides were loaded onto an mClass nanoflow UPLC system (Waters) equipped with a nanoEaze M/Z Symmetry 100 Å C18, 5 µm trap column (180 µm x 20 mm, Waters) and a PepMap, 2 µm, 100 Å, C18 EasyNano nanocapillary column (75 μm x 500 mm, Thermo). The trap wash solvent was aqueous 0.05% (v:v) trifluoroacetic acid and the trapping flow rate was 15 µL/min. The trap was washed for 5 min before switching flow to the capillary column. Separation used gradient elution of two solvents: solvent A, aqueous 0.1% (v:v) formic acid; solvent B, acetonitrile containing 0.1% (v:v) formic acid. The flow rate for the capillary column was 330 nL/min and the column temperature was 40°C. The linear multi-step gradient profile was: 3-10% B over 7 mins, 10-35% B over 30 mins, 35-99% B over 5 mins and then proceeded to wash with 99% solvent B for 4 min. The column was returned to initial conditions and re-equilibrated for 15 min before subsequent injections.
The nanoLC system was interfaced with an Orbitrap Fusion Tribrid mass spectrometer (Thermo) with an EasyNano ionisation source (Thermo). Positive ESI-MS and MS2 spectra were acquired using Xcalibur software (version 4.0, Thermo). Instrument source settings were: ion spray voltage, 1,900 V; sweep gas, 0 Arb; ion transfer tube temperature; 275°C. MS1 spectra were acquired in the Orbitrap with: 120,000 resolution, scan range: m/z 375-1,500; AGC target, 4e5; max fill time, 100 ms. Data dependant acquisition was performed in top speed mode using a 1 s cycle, selecting the most intense precursors with charge states >1. Easy-IC was used for internal calibration. Dynamic exclusion was performed for 50 s post precursor selection and a minimum threshold for fragmentation was set at 5e3. MS2 spectra were acquired in the linear ion trap with: scan rate, turbo; quadrupole isolation, 1.6 m/z; activation type, HCD; activation energy: 32%; AGC target, 5e3; first mass, 110 m/z; max fill time, 100 ms. Acquisitions were arranged by Xcalibur to inject ions for all available parallelizable time.
Peaklists in .raw format were loaded into PEAKS Studio X Pro (Version 10.6, Bioinformatics Solutions Inc.) for peptide identification and peak area-based label-free relative quantification. Database searching was performed against the human subset of the UniProt database (20476 sequences), specifying: Enzyme, trypsin; Max missed cleavages, 2; Fixed modifications, Carbamidomethyl (C); Variable modifications, Oxidation (M); Peptide tolerance, 3 ppm; MS/MS tolerance, 0.5 Da. Subsequent PEAKS-PTM searching was used to consider all Unimod listed modifications for PEAKS-DB identified proteins. Peptide identifications were filtered to 1% false discovery rate as assessed empirically against a decoy database. Protein quantification was performed using MS1 precursor ion areas following LC-MS chromatogram alignment. Protein quantifications were set to require a minimum of two unique peptides to be considered quantifiable. 727 proteins were accepted as quantifiable among samples. MS intensity was normalized between samples by total MS1 ion signal. An ANOVA comparison was applied for significance testing with the null hypothesis that proteins are of equal abundance between sample groups tested. The Hochberg and Benjamini multiple test correction was used to convert p-values to q-values. For pairwise comparisons multiple unpaired t tests were applied in GraphPad Prism.
2.5. Enrichment Analysis and Hierarchical Clustering
Protein accession numbers were uploaded to STRING-db for enrichment analysis, and additional analysis of data was completed in GraphPad Prism. Hierarchical clustering analysis was completed using Morpheus (https://software.broadinstitute.org/morpheus). Data are presented with average-linkage and one minus Pearson correlation distance.
2.6. ATPase Assay
The ATPase activity of tissue lysates from patient tissue samples was measured using the ab234055 ATPase Assay Kit – Colorimetric (Abcam), according to the manufacturer’s instructions. Briefly, 10 mg of quadriceps tissue sample was homogenised with a Dounce homogeniser in ATPase assay buffer, before centrifugation (10,000 g, 4°C, 10 mins) to pellet insoluble material. Ammonium sulphate method was used to remove endogenous phosphate. Samples were added to a 96 well plate and activity was measured in endpoint mode at wavelength 650 nm.
2.7. Lipid Peroxidation Assay
Lipid peroxidation in tissue samples was quantified using the Lipid Peroxidation ‘malondialdehyde’ (MDA) Assay Kit, ab118970 (Abcam) according to the manufacturer’s instructions. Briefly, 10 mg of tissue was washed with cold PBS, homogenised in MDA lysis buffer with a Dounce homogeniser, then centrifuged (13,000 g, 4°C, 10 mins). The supernatants were mixed with thio-barbituric acid (TBA) suspended in glacial acetic acid, then incubated at 95°C (10 mins). Samples were added to a 96 well plate and MDA content was measured in endpoint mode at wavelength 532 nm.
2.8. Western Blot
Gel immobilised proteins were transferred to a nitrocellulose membrane at 30 V for 60 minutes. The membrane was blocked with 3% (w/v) milk powder in TBS-T for 60 minutes with gentle agitation (RT). The membranes were then probed overnight (18 hours, 4°C) with the primary antibody - anti-plectin, ab229467; anti-annexin 6, ab196942; anti-annexin 11, ab137424; anti-GLO2, ab154108; anti-ALDH4A1, ab181256 (Abcam), at a 1:5000 dilution in 3% (w/v) milk powder in TBS-T. Membranes were washed three times with TBS-T before incubation with secondary antibody, goat anti-rabbit HRP conjugate, (ab6721, Abcam), at a 1:5000 dilution in 5% (w/v) milk powder in TBS-T. Membranes were then washed three times with TBS-T before a five-minute incubation with ECL substrate and chemiluminescence measurement. Band densities were measured using Image J (NIH) and samples were normalised to appropriate loading controls[42].
Statistical analysis was performed using GraphPad Prism version 9.3.1.
2.9. P5cDH Gene Methylation
Pre-concentric and post-concentric exercise samples were assessed for methylation of the P5CDH encoding gene ALDH4A1 by the Genome Centre (Blizard Institute, Queen Mary University of London). Briefly, DNA was extracted from the tissue using the Qiagen DNeasy Blood & Tissue kit. DNA was quantified with NanoDrop QC, and double stranded DNA was then quantified with Qubit 2.0 Fluorometer. Agilent 4200 Tapestation was then used to assess the quantity and quality of the extracted DNA.
Primers were designed to cover 500 bp upstream and 400 bp downstream of exon 1, ALDH4A1 (chr1:18,901,960-18,902,956 997 bp). PCR optimisation was completed with Roche High-Fidelity PCR system. The amplified PCR products were barcoded and then validated with Agilent 4200 Tapestation. Bisulphite conversion of the DNA (200 – 500 ng) was completed with the Zymo EZ DNA Methylation Kit, and samples quantified with the Roche High-Fidelity PCR System.
DNA pools were quantified with Qubit 2.0 Fluorometer and determination of average product size with Agilent 4200 Tapestation. Illumina MiSeq was used for DNA denaturation and loading before Illumina sequencing. Bismarck mapping was used to determine methylated and unmethylated DNA[43].
3. Results
3.1. Isolated Mitochondrial Fractions Were Enriched with Mitochondrial Proteins
We first confirmed the mitochondrial-enriched fractions had been isolated successfully for analysis. This could be verified with a high proportion of mitochondrial proteins in the list of ‘hits’. A cellular compartment (GO) enrichment analysis as performed (Figure 1) and the returned terms, were ranked by lowest false discovery rate (FDR), and the five terms with the lowest FDR were then presented based on strength of the enrichment effect. The term ‘mitochondrion’ was highly enriched in both the total protein list and in the list of proteins with significant variance (ANOVA) across the eight conditions (pre-/post-eccentric/concentric, male/female), indicating that our method for mitochondrial enrichment had been successful with this tissue samples set.
3.2. Changes to Protein Levels in Response to Concentric and Eccentric Exercise
We used hierarchical clustering to compare the individual samples of males and females for pre- and post- concentric and eccentric exercise (Figure 2). There were 36 proteins identified with significant changes in abundance across the four pairwise comparisons (Table 1). As might be expected with older human tissue samples, there was significant heterogeneity in these data. However, there were some consistently changing proteins that could be considered as a ‘rapid signature’ for each condition.
In response to concentric exercise, plectin relative quantity was upregulated in both, male and female participants, while annexin A11 was upregulated in males and annexin A6 was upregulated in females. Canonical mitochondrial proteins with altered expression in the mitochondrial fractions include MICOS complex subunit MIC27 and delta-1-pyrroline-5-carboxylate dehydrogenase mitochondrial (P5cDH), also known as aldehyde dehydrogenase 4 family member A1 (ALDH4A1); relative levels for these proteins were decreased with concentric exercise in males. Post-eccentric exercise, hydroxy-acyl-glutathione hydrolase mitochondrial (HAGH) increased in males, and MICOS complex subunit MIC13 decreased in females.
We had selected a sub-set of available samples, which could be matched with controls, for proteomic analysis. Therefore, it was possible to interrogate the proteins of interest arising from this work in the remaining samples set. This was done to increase the number of replicates measured for the most interesting hits. We proceeded with western blot, to measure the relative quantity of some of the proteins in additional samples that had not been analysed by mass spectrometry. Included for western blot analyses were plectin in eccentric exercise males and concentric exercise males and females, GLO2 in eccentric males, annexin A11 and P5cDH in concentric males, and annexin A6 in concentric females. The relative quantity of P5cDH in concentric exercise males and plectin in concentric exercise females, matched the initial observations within the mass spectrometry proteomic data (Figure 3), while heterogenous levels of protein (using GAPDH as reference) were observed for the other proteins (Supplementary Figure S1).
3.3. P5cDH Gene Methylation Is not the Mechanism for Decreased Abundance of this Protein Immediately After Concentric Exercise
Post-exercise alterations of epigenetic states in skeletal muscle have been documented[44,45]. As P5cDH was decreased following concentric exercise, we measured DNA methylation levels within a regulatory region of this gene. The P5cDH promoter is located within a CpG island containing 75 CpG sites on chromosome 1p36.13. Bisulfite sequencing showed that ALDH4A1 is almost completely unmethylated in all human skeletal muscle tissue samples tested (N=3), and that concentric exercise does not influence this epigenetic state at the time-point when the tissue samples were collected (Figure 4).
3.4. Concentric Exercise in COPD Males: Enrichment for Translation Elongation Factors
Enrichment analysis of proteins with altered abundance in each of the four pairwise comparisons (Table 1) revealed that the proteins associated with male concentric exercise were significantly enriched for factors operating in the ‘eukaryotic translation elongation’ network (Figure 5). For example, the tRNA delivery proteins elongation factor 1-delta (EEF1D) and elongation factor 1-gamma (EEF1G), are both present in the enrichment cluster and are upregulated. In contrast, five ribosomal subunit proteins, also present in this cluster, are downregulated (Figure 5).
3.5. Carbonic Anhydrases I-III, Haemoglobin Subunits, and Haemoglobin Associated Proteins Are Present in COPD Skeletal Muscle Mitochondria
We have previously found certain proteins associated with mitochondria that are interesting in ageing, particularly haemoglobin and carbonic anhydrase II (CA-II) [41,46,47]. Haemoglobin subunit α and haemoglobin subunit β were found in all the samples we measured, as was haemoglobin subunit δ. We didn’t find significant variation (Figure 6) when comparing these proteins between sexes and exercise conditions. Hemopexin and haptoglobin were also found in these mitochondrial samples, as were CA-I, CA-II, and CA-III.
3.6. Tissue ATPase Activity and Levels of Lipid Peroxidation Did not Change Significantly in Response to Concentric or Eccentric Exercise
A clear, yet not significant decrease in ATPase activity of the post-concentric exercise samples (unpaired t test, P = 0.095) was observed (Figure 7). Lipid peroxidation (MDA) levels in the same quadriceps samples were also measured; neither concentric nor eccentric exercise resulted in a significant difference in lipid peroxidation (MDA) levels.
4. Discussion
This is the first study in which sex-specific alterations to the mitochondrial proteome in response to eccentric and concentric exercise have been explored and identified in the context of COPD. We also investigated potential epigenetic modulation of the P5cDH gene expression as an explanation for the reduction of this protein after exercise. We confirmed the presence of proteins interesting in the context of ageing, in the mitochondrial proteome of individuals with COPD. We measured ATPase activity and lipid peroxidation levels in the skeletal muscle samples studied.
Our observations are limited in the way many physiology studies involving human participants are. The heterogeneity of human samples after over half a century of life with different environments, including diet and activity is difficult to overcome, without large sample sets. In this experiment, we were also limited by the previous study design which provided us with the muscle biopsies. We were only able to measure the immediate changes in protein expression with exercise. Nevertheless, we report some interesting pathways and proteins that might be ‘first responders’ in the skeletal muscle mitochondrial milieu. We have also discovered sex specific differences at this molecular level in COPD skeletal muscle mitochondria.
4.1. Concentric Exercise Upregulates Plectin and Annexin Proteins in Male and Female Participants
Sex specific adaptations of the mitochondrial proteome in response to the concentric and eccentric cycling is one of the key findings of this study. The differences between male and female samples include the numbers of proteins with altered abundance, and the specific proteins that were identified (Table 1). Exercise type revealed uniform responses; concentric cycling induced upregulation of plectin and annexin proteins (annexin A11 in males, annexin A6 in females) in male and female participants. Plectin is a member of the plakin family of cytolinker proteins, and is a major component of intermediate filament networks of muscle tissue[48,49]. Beyond its role in cellular structure, multiple studies have shown plectin to be an important modulator of mitochondrial morphology and function, in particular in the structural organisation of mitochondria within muscle tissue[50,51].
The plectin 1b isoform has been shown to bind to the outer mitochondrial membrane, where it links mitochondria to intermediate filaments, and deficiency of the plectin 1b isoform results in elongated mitochondria in primary fibroblast cells[52]. In addition, plectin 1b isoform deficiency in mice has been shown to reduce the mitochondrial respiratory capacity in heart, soleus, and gastrocnemius tissue[53]. It is therefore possible that the upregulation of plectin in the mitochondrial fraction of skeletal muscle in response to concentric exercise is taking place to better provide for the bioenergetic demands of exercise. In COPD this upregulation could make it easier for individuals to maintain physical activity, which would lead to further gains.
Annexin A6 has been shown to be an important calcium channel in the inner mitochondrial membrane[54]. Fibroblast mitochondria from annexin A6 knockout mice were highly fragmented, and it was shown that annexin A6 inhibits Drp1 mediated mitochondrial fission[55,56]. Drp1 has already been implicated in the pathology of skeletal muscle in COPD and is linked to cigarette smoke exposure which is known to be the greatest risk factor for developing COPD.
4.2. Mitochondrial Protein Abundance Alterations Are Sex-Specific in Response to Concentric and Eccentric Exercise
We set up the proteomics analyses so that we could identify potential sex-specific responses to the two different exercise conditions. We found that in male skeletal muscle fractions, concentric exercise led to decreased expression of several proteins (Table 1). Known mitochondrial proteins that decreased in expression were the MICOS complex subunit MIC27 (MIC27) and delta-1-pyrroline-5-carboxylate dehydrogenase (P5CDH). The mitochondrial contact site and cristae organizing system (MICOS) complex is one of the key inner membrane organisation structures, as well as being a hub for protein-protein interactions[57]. A double knockout study of MIC26 and MIC27 in HAP1 cells reported that the two subunits have a cooperative role in maintaining the integrity of respiratory super-complex and ATP synthase structures, through modulation of cardiolipin levels[58]. Lysocardiolipin acyltransferase (LYCAT) regulates the fatty acid composition of cardiolipin and is expressed at high levels in lung epithelial cells exposed to cigarette smoke[59]. It could be that cardiolipin regulation is a key factor in COPD pathology.
P5CDH is responsible for the second step in the degradation of proline, and a study of the D. melanogaster orthologue reported that a homozygous mutant (CG7145f04633/CG7145f04633) had elevated proline levels and a swollen mitochondrial morphology [60]. There is not that much known about the P5CDH protein but a transcriptomics study of the effect of cigarette smoke on human bronchial epithelial cells, as a model of COPD, highlighted pathways associated with its transcript[61].
Male participants had a significant increase in the expression of mitochondrial hydroxyacylglutathione hydrolase (GLO2) protein, while in female participants the expression of the MICOS complex subunit MIC13 could no longer be detected post-eccentric exercise. GLO2 is part of a defence mechanism that is used against dicarbonyl stress which is reduced in skeletal muscle with exercise[62]. MIC13 knockout cells have been shown to have impaired cristae junction formation and a decreased basal and maximal respiratory capacity, with MIC13 also shown to be necessary for the integration of MIC26 and MIC27 into the MICOS complex structure[63].
4.3. Western Blotting Shows that P5cDH Is Decreased in Samples Not Analysed by Mass Spectrometry
We completed analyses of all the samples we had not used for proteomics by western blot, to complete the dataset. Western blots also showed there was decreased expression of the P5cDH protein (Figure 3). We measured methylation of the gene that encodes P5cDH, to check whether this mechanism of regulation was applicable in this case[44]. We used targeted bisulfite sequencing but did not find any significant levels of methylation, at the CpG island we had identified upstream of the gene (Figure 4). Pathways that could downregulate this protein at the post-translational level are the next point of interrogation.
4.4. Protein Translation and Elongation Factors Changed with Concentric Exercise in Males
Proteins that had significantly altered abundance in male skeletal muscle fractions, post-concentric exercise, had local network enrichment for the term ‘eukaryotic translation elongation’ (Figure 5). Specifically, the tRNA delivery proteins elongation factor 1-gamma and elongation factor 1-delta were upregulated in expression. The ribosomal proteins 60S ribosomal protein L6, 60S ribosomal protein L7, 60S ribosomal protein L18, 60S acidic ribosomal protein P0, and 60S acidic ribosomal protein P0 were all downregulated in expression (Table 1).
It has previously been reported that the expression of ribosomal proteins is highly differentiated across different tissue types[64], and that the elongation factor 1 complex (comprised of alpha, beta, gamma and delta) has also been shown to have a range of functions beyond protein translation[65]. Eukaryotic translation elongation factor 1-delta is already implicated to interact with a long coding RNA NONMMUT033452.2 in an airway inflammation paradigm[66].
The presence of cytosolic ribosomal subunits in the mitochondrial fraction of skeletal muscle may support the observation by Gold et al. that cytosolic ribosomes can localise to the surface of the mitochondria through the TOM complex[67]. A decrease in the levels of association between the cytosolic ribosomes associated with the surface of the mitochondria may indicate a (possibly short-term) decreased rate of import, of nuclear-encoded mitochondrial proteins, in males doing concentric exercise.
4.5. Haemoglobin Subunits, Haptoglobin, Hemopexin, and Carbonic Anhydrases I, II and III Are Part of the Mitochondrial Proteome of COPD Skeletal Muscle
We have localised haemoglobin and carbonic anhydrase II to the mitochondria and connected these proteins to Parkinson’s disease and mitochondrial ageing[41,46,47]. One of the many questions that remain about non-erythroid haemoglobin is its potential cytotoxicity[68]. We found cell-free haemoglobin detoxifying protein haptoglobin together with the haem detoxifying protein hemopexin in the mitochondrial fraction of muscle tissue offering a potential solution to this conundrum (Figure 6).
While carbonic anhydrases VA and VB are the isozymes known to be associated with mitochondria[69,70], we have made repeated observations of carbonic anhydrases II and III in the mitochondrial proteome. We previously showed increased levels of mitochondrial carbonic anhydrase II is associated with decreased lifespan[47].
Here we can see that carbonic anhydrase I is present in the mitochondrial fraction of COPD skeletal muscle. This isozyme has previously been found in the cytosol, and it was reported that its increased expression in the ischemic myocardium of type 2 diabetes increases rates of endothelial cell death[71]. Without further work it is not possible to say whether this is an association with the outer mitochondrial membrane or if the protein is located within the organelle. However, considering the role of carbonic anhydrases in maintaining homeostasis it may be useful to revisit the potential and dynamic localisations of this group of proteins.
4.6. ATPase Activity and Levels of Lipid Peroxidation Are Unchanged in Response to Concentric and Eccentric Exercise
To better understand changes in mitochondrial physiology in response to concentric and eccentric cycling, we measured ATPase activity from pre- and post-concentric and eccentric cycling tissue lysates and observed a trend toward decreased ATPase activity, but no statistically significant changes (Figure 7A). Mitochondria are a major source of endogenous ROS and COPD is associated with oxidative stress[72], while exercise is also recognised as promoting ‘healthy levels’ of oxidative stress that ameliorate insulin resistance[73]. However, a lipid peroxidation assay revealed that there were no changes in the MDA content of the samples pre- and post-concentric and eccentric cycling (Figure 7B), which suggests the exercise did not promote significant measurable oxidative stress in skeletal muscle.
5. Conclusions
Eccentric and concentric exercise in COPD induced some rapid changes in the expression profile of the mitochondrial proteome. Proteins with altered abundance in response to both exercise conditions include those that have previously been associated with regulating mitochondrial morphology and ultrastructure. In this study, we have been able to identify proteomic profile changes that correlate with exercise and the type of muscle action elicited, eccentric or concentric. Most interestingly, we find clear indications that the proteome response to exercise is influenced by the sex of the study participant. It is important to address sex differences in the physiology of disease as this reflects the observed sex specific clinical presentation of COPD. This should be considered when trialling new therapies for COPD and could be important for the design of treatment for people with this condition. Changes to mitochondrial morphology may be a positive adaptation to exercise in COPD and the pathways that are implicated could represent new targets for pharmaceutical development.
Supplementary Materials
The following supporting information can be downloaded at website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Lisa Chakrabarti; Data curation, Brad Ebanks and Reinhard Stöger; Formal analysis, Adam Dowle and Reinhard Stöger; Investigation, Brad Ebanks, Adam Dowle and Thomas Ward; Methodology, Brad Ebanks, Adam Dowle, Reinhard Stöger and Lisa Chakrabarti; Project administration, Lisa Chakrabarti; Resources, Lisa Chakrabarti; Supervision, Lisa Chakrabarti; Validation, Adam Dowle; Writing – original draft, Brad Ebanks, Adam Dowle, Reinhard Stöger and Lisa Chakrabarti; Writing – review & editing, Jacob Reed, Katherine Mortimer and Lisa Chakrabarti.
Funding
BE and KRHM are funded by the Biotechnology and Biological Sciences Research Council (grant number BB/J014508/1).
Data Availability
Complete mass spectrometry data sets and proteomic identifications are available to download from MassIVE (MSV000091127), [doi:10.25345/C5PK07B6V] and ProteomeXchange (PXD039636). Openly accessible upon publication.
Acknowledgments
The York Centre of Excellence in Mass Spectrometry was created thanks to a major capital investment through Science City York, supported by Yorkshire Forward with funds from the Northern Way Initiative, and subsequent support from EPSRC (EP/K039660/1; EP/M028127/1).
Conflicts of Interest
none of the authors has any conflicts of interests.
References
- Adeloye, D.; Song, P.; Zhu, Y.; Campbell, H.; Sheikh, A.; Rudan, I.; Unit, N.R.G.R.H. Global, regional, and national prevalence of, and risk factors for, chronic obstructive pulmonary disease (COPD) in 2019: a systematic review and modelling analysis. Lancet Respir Med 2022, 10, 447-458. [CrossRef]
- GOLD. Global Strategy for Prevention, Diagnosis and Management of COPD.; 2022.
- Buist, A.S.; McBurnie, M.A.; Vollmer, W.M.; Gillespie, S.; Burney, P.; Mannino, D.M.; Menezes, A.M.; Sullivan, S.D.; Lee, T.A.; Weiss, K.B.; et al. International variation in the prevalence of COPD (the BOLD Study): a population-based prevalence study. Lancet 2007, 370, 741-750. [CrossRef]
- Barnes, P.J. Chronic obstructive pulmonary disease. N Engl J Med 2000, 343, 269-280. [CrossRef]
- Calverley, P.M. Respiratory failure in chronic obstructive pulmonary disease. Eur Respir J Suppl 2003, 47, 26s-30s. [CrossRef]
- Kent, B.D.; Mitchell, P.D.; McNicholas, W.T. Hypoxemia in patients with COPD: cause, effects, and disease progression. Int J Chron Obstruct Pulmon Dis 2011, 6, 199-208. [CrossRef]
- Wust, R.C.; Degens, H. Factors contributing to muscle wasting and dysfunction in COPD patients. Int J Chron Obstruct Pulmon Dis 2007, 2, 289-300.
- Rabinovich, R.A.; Bastos, R.; Ardite, E.; Llinas, L.; Orozco-Levi, M.; Gea, J.; Vilaro, J.; Barbera, J.A.; Rodriguez-Roisin, R.; Fernandez-Checa, J.C.; et al. Mitochondrial dysfunction in COPD patients with low body mass index. Eur Respir J 2007, 29, 643-650. [CrossRef]
- Puente-Maestu, L.; Perez-Parra, J.; Godoy, R.; Moreno, N.; Tejedor, A.; Gonzalez-Aragoneses, F.; Bravo, J.L.; Alvarez, F.V.; Camano, S.; Agusti, A. Abnormal mitochondrial function in locomotor and respiratory muscles of COPD patients. Eur Respir J 2009, 33, 1045-1052. [CrossRef]
- Haji, G.; Wiegman, C.H.; Michaeloudes, C.; Patel, M.S.; Curtis, K.; Bhavsar, P.; Polkey, M.I.; Adcock, I.M.; Chung, K.F.; consortium, C. Mitochondrial dysfunction in airways and quadriceps muscle of patients with chronic obstructive pulmonary disease. Respir Res 2020, 21, 262. [CrossRef]
- Maremanda, K.P.; Sundar, I.K.; Rahman, I. Role of inner mitochondrial protein OPA1 in mitochondrial dysfunction by tobacco smoking and in the pathogenesis of COPD. Redox Biol 2021, 45, 102055. [CrossRef]
- Lee, H.; Smith, S.B.; Yoon, Y. The short variant of the mitochondrial dynamin OPA1 maintains mitochondrial energetics and cristae structure. J Biol Chem 2017, 292, 7115-7130. [CrossRef]
- Calverley, P.M.A. Exercise and dyspnoea in COPD. European Respiratory Review 15, 72-79. [CrossRef]
- Nicholas, J.J.; Gilbert, R.; Gabe, R.; Auchincloss, J.H., Jr. Evaluation of an exercise therapy program for patients with chronic obstructive pulmonary disease. Am Rev Respir Dis 1970, 102, 1-9. [CrossRef]
- Ward, T.J.C.; Lindley, M.R.; Ferguson, R.A.; Constantin, D.; Singh, S.J.; Bolton, C.E.; Evans, R.A.; Greenhaff, P.L.; Steiner, M.C. Submaximal Eccentric Cycling in People With COPD: Acute Whole-Body Cardiopulmonary and Muscle Metabolic Responses. Chest 2021, 159, 564-574. [CrossRef]
- Joschtel, B.; Gomersall, S.R.; Tweedy, S.; Petsky, H.; Chang, A.B.; Trost, S.G. Effects of exercise training on physical and psychosocial health in children with chronic respiratory disease: a systematic review and meta-analysis. BMJ Open Sport Exerc Med 2018, 4, e000409. [CrossRef]
- Padulo, J.; Laffaye, G.; Chamari, K.; Concu, A. Concentric and eccentric: muscle contraction or exercise? Sports Health 2013, 5, 306. [CrossRef]
- Dufour, S.P.; Lampert, E.; Doutreleau, S.; Lonsdorfer-Wolf, E.; Billat, V.L.; Piquard, F.; Richard, R. Eccentric cycle exercise: training application of specific circulatory adjustments. Med Sci Sports Exerc 2004, 36, 1900-1906. [CrossRef]
- Rocha Vieira, D.S.; Baril, J.; Richard, R.; Perrault, H.; Bourbeau, J.; Taivassalo, T. Eccentric cycle exercise in severe COPD: feasibility of application. COPD 2011, 8, 270-274. [CrossRef]
- Hody, S.; Croisier, J.L.; Bury, T.; Rogister, B.; Leprince, P. Eccentric Muscle Contractions: Risks and Benefits. Front Physiol 2019, 10, 536. [CrossRef]
- Rattray, B.; Caillaud, C.; Ruell, P.A.; Thompson, M.W. Heat exposure does not alter eccentric exercise-induced increases in mitochondrial calcium and respiratory dysfunction. Eur J Appl Physiol 2011, 111, 2813-2821. [CrossRef]
- MacMillan, N.J.; Kapchinsky, S.; Konokhova, Y.; Gouspillou, G.; de Sousa Sena, R.; Jagoe, R.T.; Baril, J.; Carver, T.E.; Andersen, R.E.; Richard, R.; et al. Eccentric Ergometer Training Promotes Locomotor Muscle Strength but Not Mitochondrial Adaptation in Patients with Severe Chronic Obstructive Pulmonary Disease. Front Physiol 2017, 8, 114. [CrossRef]
- Touron, J.; Costes, F.; Coudeyre, E.; Perrault, H.; Richard, R. Aerobic Metabolic Adaptations in Endurance Eccentric Exercise and Training: From Whole Body to Mitochondria. Front Physiol 2020, 11, 596351. [CrossRef]
- Isner-Horobeti, M.E.; Rasseneur, L.; Lonsdorfer-Wolf, E.; Dufour, S.P.; Doutreleau, S.; Bouitbir, J.; Zoll, J.; Kapchinsky, S.; Geny, B.; Daussin, F.N.; et al. Effect of eccentric versus concentric exercise training on mitochondrial function. Muscle Nerve 2014, 50, 803-811. [CrossRef]
- Yang, M.; Kohler, M.; Heyder, T.; Forsslund, H.; Garberg, H.K.; Karimi, R.; Grunewald, J.; Berven, F.S.; Nyrén, S.; Magnus Sköld, C.; et al. Proteomic profiling of lung immune cells reveals dysregulation of phagocytotic pathways in female-dominated molecular COPD phenotype. Respiratory Research 2018, 19, 39. [CrossRef]
- Zhang, Y.H.; Hoopmann, M.R.; Castaldi, P.J.; Simonsen, K.A.; Midha, M.K.; Cho, M.H.; Criner, G.J.; Bueno, R.; Liu, J.; Moritz, R.L.; et al. Lung proteomic biomarkers associated with chronic obstructive pulmonary disease. Am J Physiol Lung Cell Mol Physiol 2021, 321, L1119-l1130. [CrossRef]
- Bai, S.; Ye, R.; Wang, C.; Sun, P.; Wang, D.; Yue, Y.; Wang, H.; Wu, S.; Yu, M.; Xi, S.; et al. Identification of Proteomic Signatures in Chronic Obstructive Pulmonary Disease Emphysematous Phenotype. Front Mol Biosci 2021, 8, 650604. [CrossRef]
- Kim, S.H.; Ahn, H.S.; Park, J.S.; Yeom, J.; Yu, J.; Kim, K.; Oh, Y.M. A Proteomics-Based Analysis of Blood Biomarkers for the Diagnosis of COPD Acute Exacerbation. Int J Chron Obstruct Pulmon Dis 2021, 16, 1497-1508. [CrossRef]
- Ebanks, B.; Wang, Y.; Katyal, G.; Sargent, C.; Ingram, T.L.; Bowman, A.; Moisoi, N.; Chakrabarti, L. Exercising D. melanogaster Modulates the Mitochondrial Proteome and Physiology. The Effect on Lifespan Depends upon Age and Sex. Int J Mol Sci 2021, 22. [CrossRef]
- Ebanks, B.; Ingram, T.L.; Katyal, G.; Ingram, J.R.; Moisoi, N.; Chakrabarti, L. The dysregulated Pink1(-) Drosophila mitochondrial proteome is partially corrected with exercise. Aging (Albany NY) 2021, 13, 14709-14728. [CrossRef]
- Hoene, M.; Kappler, L.; Kollipara, L.; Hu, C.; Irmler, M.; Bleher, D.; Hoffmann, C.; Beckers, J.; Hrabe de Angelis, M.; Haring, H.U.; et al. Exercise prevents fatty liver by modifying the compensatory response of mitochondrial metabolism to excess substrate availability. Mol Metab 2021, 54, 101359. [CrossRef]
- David, D.C.; Hauptmann, S.; Scherping, I.; Schuessel, K.; Keil, U.; Rizzu, P.; Ravid, R.; Drose, S.; Brandt, U.; Muller, W.E.; et al. Proteomic and functional analyses reveal a mitochondrial dysfunction in P301L tau transgenic mice. J Biol Chem 2005, 280, 23802-23814. [CrossRef]
- Granata, C.; Caruana, N.J.; Botella, J.; Jamnick, N.A.; Huynh, K.; Kuang, J.; Janssen, H.A.; Reljic, B.; Mellett, N.A.; Laskowski, A.; et al. High-intensity training induces non-stoichiometric changes in the mitochondrial proteome of human skeletal muscle without reorganisation of respiratory chain content. Nat Commun 2021, 12, 7056. [CrossRef]
- Walls, K.C.; Coskun, P.; Gallegos-Perez, J.L.; Zadourian, N.; Freude, K.; Rasool, S.; Blurton-Jones, M.; Green, K.N.; LaFerla, F.M. Swedish Alzheimer mutation induces mitochondrial dysfunction mediated by HSP60 mislocalization of amyloid precursor protein (APP) and beta-amyloid. J Biol Chem 2012, 287, 30317-30327. [CrossRef]
- Broadwater, L.; Pandit, A.; Clements, R.; Azzam, S.; Vadnal, J.; Sulak, M.; Yong, V.W.; Freeman, E.J.; Gregory, R.B.; McDonough, J. Analysis of the mitochondrial proteome in multiple sclerosis cortex. Biochim Biophys Acta 2011, 1812, 630-641. [CrossRef]
- Neilson, K.A.; Ali, N.A.; Muralidharan, S.; Mirzaei, M.; Mariani, M.; Assadourian, G.; Lee, A.; van Sluyter, S.C.; Haynes, P.A. Less label, more free: approaches in label-free quantitative mass spectrometry. Proteomics 2011, 11, 535-553. [CrossRef]
- Geiger, T.; Cox, J.; Mann, M. Proteomics on an Orbitrap benchtop mass spectrometer using all-ion fragmentation. Mol Cell Proteomics 2010, 9, 2252-2261. [CrossRef]
- Zubarev, R.A.; Makarov, A. Orbitrap mass spectrometry. Anal Chem 2013, 85, 5288-5296. [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res 2019, 47, D607-D613. [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res 2017, 45, D353-D361. [CrossRef]
- Shephard, F.; Greville-Heygate, O.; Marsh, O.; Anderson, S.; Chakrabarti, L. A mitochondrial location for haemoglobins--dynamic distribution in ageing and Parkinson's disease. Mitochondrion 2014, 14, 64-72. [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat Methods 2012, 9, 671-675. [CrossRef]
- Krueger, F.; Andrews, S.R. Bismark: a flexible aligner and methylation caller for Bisulfite-Seq applications. Bioinformatics 2011, 27, 1571-1572. [CrossRef]
- Voisin, S.; Eynon, N.; Yan, X.; Bishop, D.J. Exercise training and DNA methylation in humans. Acta Physiol (Oxf) 2015, 213, 39-59. [CrossRef]
- Plaza-Diaz, J.; Izquierdo, D.; Torres-Martos, A.; Baig, A.T.; Aguilera, C.M.; Ruiz-Ojeda, F.J. Impact of Physical Activity and Exercise on the Epigenome in Skeletal Muscle and Effects on Systemic Metabolism. Biomedicines 2022, 10. [CrossRef]
- Shephard, F.; Greville-Heygate, O.; Liddell, S.; Emes, R.; Chakrabarti, L. Analysis of Mitochondrial haemoglobin in Parkinson's disease brain. Mitochondrion 2016, 29, 45-52. [CrossRef]
- Pollard, A.; Shephard, F.; Freed, J.; Liddell, S.; Chakrabarti, L. Mitochondrial proteomic profiling reveals increased carbonic anhydrase II in aging and neurodegeneration. Aging (Albany NY) 2016, 8, 2425-2436. [CrossRef]
- Leung, C.L.; Green, K.J.; Liem, R.K. Plakins: a family of versatile cytolinker proteins. Trends Cell Biol 2002, 12, 37-45. [CrossRef]
- Winter, L.; Wiche, G. The many faces of plectin and plectinopathies: pathology and mechanisms. Acta Neuropathol 2013, 125, 77-93. [CrossRef]
- Reipert, S.; Steinbock, F.; Fischer, I.; Bittner, R.E.; Zeold, A.; Wiche, G. Association of mitochondria with plectin and desmin intermediate filaments in striated muscle. Exp Cell Res 1999, 252, 479-491. [CrossRef]
- Appaix, F.; Kuznetsov, A.V.; Usson, Y.; Kay, L.; Andrienko, T.; Olivares, J.; Kaambre, T.; Sikk, P.; Margreiter, R.; Saks, V. Possible role of cytoskeleton in intracellular arrangement and regulation of mitochondria. Exp Physiol 2003, 88, 175-190. [CrossRef]
- Winter, L.; Abrahamsberg, C.; Wiche, G. Plectin isoform 1b mediates mitochondrion-intermediate filament network linkage and controls organelle shape. J Cell Biol 2008, 181, 903-911. [CrossRef]
- Winter, L.; Kuznetsov, A.V.; Grimm, M.; Zeold, A.; Fischer, I.; Wiche, G. Plectin isoform P1b and P1d deficiencies differentially affect mitochondrial morphology and function in skeletal muscle. Hum Mol Genet 2015, 24, 4530-4544. [CrossRef]
- Rainteau, D.; Mansuelle, P.; Rochat, H.; Weinman, S. Characterization and ultrastructural localization of annexin VI from mitochondria. FEBS Lett 1995, 360, 80-84. [CrossRef]
- Chlystun, M.; Campanella, M.; Law, A.L.; Duchen, M.R.; Fatimathas, L.; Levine, T.P.; Gerke, V.; Moss, S.E. Regulation of mitochondrial morphogenesis by annexin A6. PLoS One 2013, 8, e53774. [CrossRef]
- Tan, Z.; Zhao, M.; Li, J.; Li, S.; Zhu, S.; Yao, X.; Gao, X.; Yang, S. Myostatin is involved in skeletal muscle dysfunction in chronic obstructive pulmonary disease via Drp-1 mediated abnormal mitochondrial division. Ann Transl Med 2022, 10, 162. [CrossRef]
- Mukherjee, I.; Ghosh, M.; Meinecke, M. MICOS and the mitochondrial inner membrane morphology - when things get out of shape. FEBS Lett 2021, 595, 1159-1183. [CrossRef]
- Anand, R.; Kondadi, A.K.; Meisterknecht, J.; Golombek, M.; Nortmann, O.; Riedel, J.; Peifer-Weiß, L.; Brocke-Ahmadinejad, N.; Schlütermann, D.; Stork, B.; et al. MIC26 and MIC27 cooperate to regulate cardiolipin levels and the landscape of OXPHOS complexes. Life Sci Alliance 2020, 3. [CrossRef]
- Bandela, M.; Suryadevara, V.; Fu, P.; Reddy, S.P.; Bikkavilli, K.; Huang, L.S.; Dhavamani, S.; Subbaiah, P.V.; Singla, S.; Dudek, S.M.; et al. Role of Lysocardiolipin Acyltransferase in Cigarette Smoke-Induced Lung Epithelial Cell Mitochondrial ROS, Mitochondrial Dynamics, and Apoptosis. Cell Biochem Biophys 2022, 80, 203-216. [CrossRef]
- He, F.; DiMario, P.J. Drosophila delta-1-pyrroline-5-carboxylate dehydrogenase (P5CDh) is required for proline breakdown and mitochondrial integrity-Establishing a fly model for human type II hyperprolinemia. Mitochondrion 2011, 11, 397-404. [CrossRef]
- Zhang, Y.; Xue, X.; Meng, L.; Li, D.; Qiao, W.; Wang, J.; Xie, D. Roles of autophagy-related genes in the therapeutic effects of Xuanfei Pingchuan capsules on chronic obstructive pulmonary disease based on transcriptome sequencing analysis. Front Pharmacol 2023, 14, 1123882. [CrossRef]
- Maessen, M.F.H.; Schalkwijk, C.G.; Verheggen, R.; Aengevaeren, V.L.; Hopman, M.T.E.; Eijsvogels, T.M.H. A comparison of dicarbonyl stress and advanced glycation endproducts in lifelong endurance athletes vs. sedentary controls. J Sci Med Sport 2017, 20, 921-926. [CrossRef]
- Anand, R.; Strecker, V.; Urbach, J.; Wittig, I.; Reichert, A.S. Mic13 Is Essential for Formation of Crista Junctions in Mammalian Cells. PLoS One 2016, 11, e0160258. [CrossRef]
- Bortoluzzi, S.; d'Alessi, F.; Romualdi, C.; Danieli, G.A. Differential expression of genes coding for ribosomal proteins in different human tissues. Bioinformatics 2001, 17, 1152-1157. [CrossRef]
- Sasikumar, A.N.; Perez, W.B.; Kinzy, T.G. The many roles of the eukaryotic elongation factor 1 complex. Wiley Interdiscip Rev RNA 2012, 3, 543-555. [CrossRef]
- Zhao, J.; Zhang, Q.; Yao, D.; Wang, T.; Ni, M.; Xu, Y.; Tang, Z.; Liu, Z. Prenatal LPS Exposure Promotes Allergic Airway Inflammation via Long Coding RNA NONMMUT033452.2, and Protein Binding Partner, Eef1D. Am J Respir Cell Mol Biol 2023, 68, 610-624. [CrossRef]
- Gold, V.A.; Chroscicki, P.; Bragoszewski, P.; Chacinska, A. Visualization of cytosolic ribosomes on the surface of mitochondria by electron cryo-tomography. EMBO Rep 2017, 18, 1786-1800. [CrossRef]
- Saha, D.; Patgaonkar, M.; Shroff, A.; Ayyar, K.; Bashir, T.; Reddy, K.V. Hemoglobin expression in nonerythroid cells: novel or ubiquitous? Int J Inflam 2014, 2014, 803237. [CrossRef]
- Shah, G.N.; Hewett-Emmett, D.; Grubb, J.H.; Migas, M.C.; Fleming, R.E.; Waheed, A.; Sly, W.S. Mitochondrial carbonic anhydrase CA VB: differences in tissue distribution and pattern of evolution from those of CA VA suggest distinct physiological roles. Proc Natl Acad Sci U S A 2000, 97, 1677-1682. [CrossRef]
- Vullo, D.; Nishimori, I.; Innocenti, A.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase activators: an activation study of the human mitochondrial isoforms VA and VB with amino acids and amines. Bioorg Med Chem Lett 2007, 17, 1336-1340. [CrossRef]
- Torella, D.; Ellison, G.M.; Torella, M.; Vicinanza, C.; Aquila, I.; Iaconetti, C.; Scalise, M.; Marino, F.; Henning, B.J.; Lewis, F.C.; et al. Carbonic anhydrase activation is associated with worsened pathological remodeling in human ischemic diabetic cardiomyopathy. J Am Heart Assoc 2014, 3, e000434. [CrossRef]
- Antunes, M.A.; Lopes-Pacheco, M.; Rocco, P.R.M. Oxidative Stress-Derived Mitochondrial Dysfunction in Chronic Obstructive Pulmonary Disease: A Concise Review. Oxid Med Cell Longev 2021, 2021, 6644002. [CrossRef]
- Ristow, M.; Zarse, K.; Oberbach, A.; Kloting, N.; Birringer, M.; Kiehntopf, M.; Stumvoll, M.; Kahn, C.R.; Bluher, M. Antioxidants prevent health-promoting effects of physical exercise in humans. Proc Natl Acad Sci U S A 2009, 106, 8665-8670. [CrossRef]
Figure 1.
Cellular compartment (GO) enriched terms for mitochondrial isolates. A) Cellular compartment (GO) enriched terms for all proteins identified across both sexes and exercise conditions; B) all proteins with significant ANOVA variation identified across both sexes and conditions. Enrichment analysis in STRING-db. The five pathways with the lowest false discovery rates (FDR) are scored according to strength of the enrichment effect (Log10(observed/expected)).
Figure 1.
Cellular compartment (GO) enriched terms for mitochondrial isolates. A) Cellular compartment (GO) enriched terms for all proteins identified across both sexes and exercise conditions; B) all proteins with significant ANOVA variation identified across both sexes and conditions. Enrichment analysis in STRING-db. The five pathways with the lowest false discovery rates (FDR) are scored according to strength of the enrichment effect (Log10(observed/expected)).
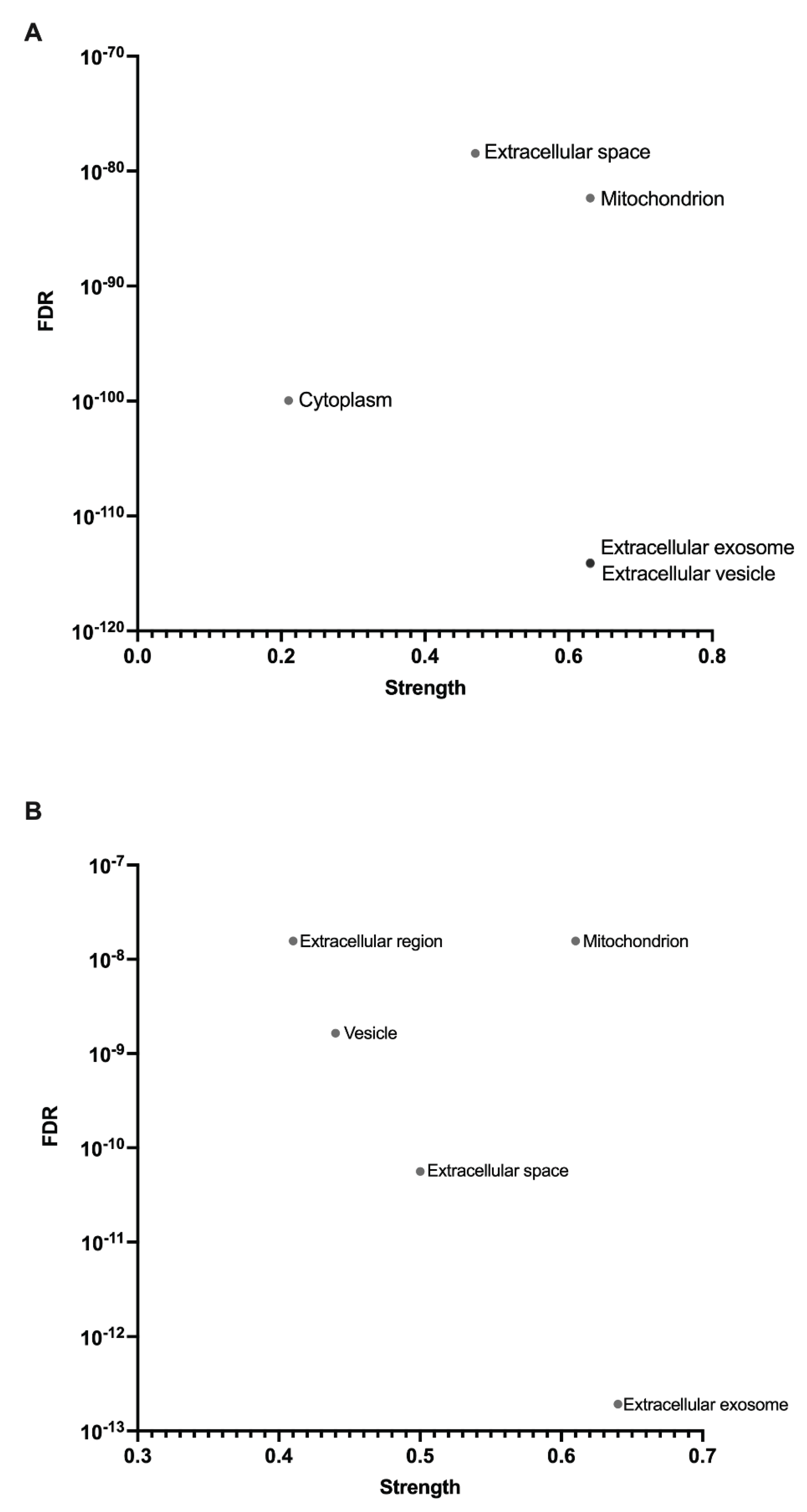
Figure 2.
Hierarchical clustering heatmap of protein expression for concentric and eccentric exercise. M = male, F = female, C = concentric, E = eccentric. A) Hierarchical clustering of identified protein expression in pre-concentric and post-concentric exercise for male and female participants, measured by relative peak areas; B) Hierarchical clustering of identified protein expression in pre-eccentric and post-eccentric exercise for male and female participants, measured by relative peak areas. Clustering performed with Morpheus (https://software.broadinstitute.org/morpheus), average linkage, one minus Pearson correlation. Red = increased expression, blue = decreased expression.
Figure 2.
Hierarchical clustering heatmap of protein expression for concentric and eccentric exercise. M = male, F = female, C = concentric, E = eccentric. A) Hierarchical clustering of identified protein expression in pre-concentric and post-concentric exercise for male and female participants, measured by relative peak areas; B) Hierarchical clustering of identified protein expression in pre-eccentric and post-eccentric exercise for male and female participants, measured by relative peak areas. Clustering performed with Morpheus (https://software.broadinstitute.org/morpheus), average linkage, one minus Pearson correlation. Red = increased expression, blue = decreased expression.
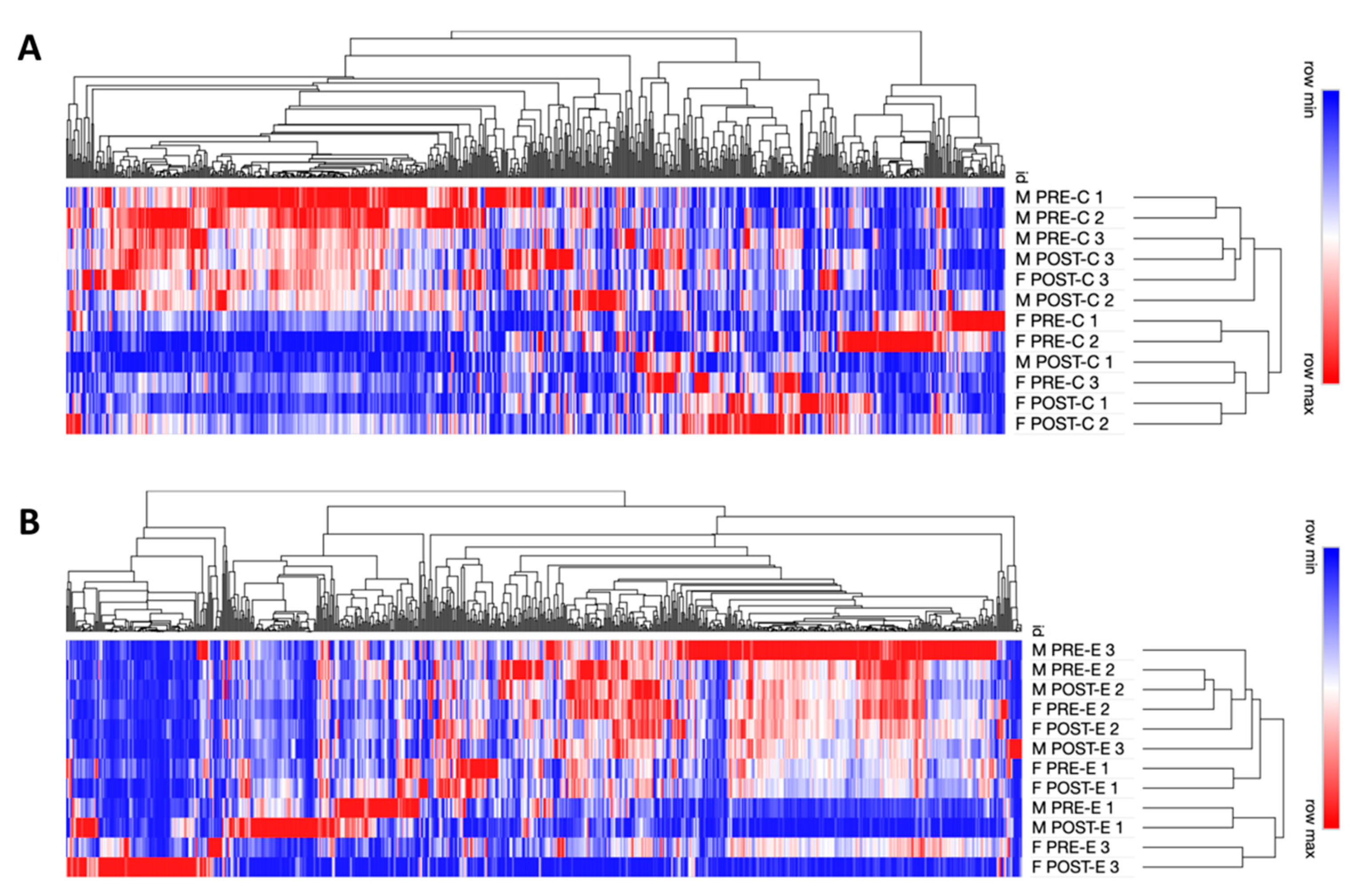
Figure 3.
Western blot of P5cDH in male (concentric exercise) muscle mitochondria fractions and of Plectin in female (concentric exercise) skeletal muscle mitochondria fractions matches the proteomics findings. A) relative amount of P5cDH in concentric male skeletal muscle mitochondrial fraction, normalised to GAPDH (pre-concentric exercise n=3, post concentric exercise n=1); B) relative quantity of Plectin in concentric female skeletal muscle mitochondrial fraction, normalised to GAPDH (n=1).
Figure 3.
Western blot of P5cDH in male (concentric exercise) muscle mitochondria fractions and of Plectin in female (concentric exercise) skeletal muscle mitochondria fractions matches the proteomics findings. A) relative amount of P5cDH in concentric male skeletal muscle mitochondrial fraction, normalised to GAPDH (pre-concentric exercise n=3, post concentric exercise n=1); B) relative quantity of Plectin in concentric female skeletal muscle mitochondrial fraction, normalised to GAPDH (n=1).
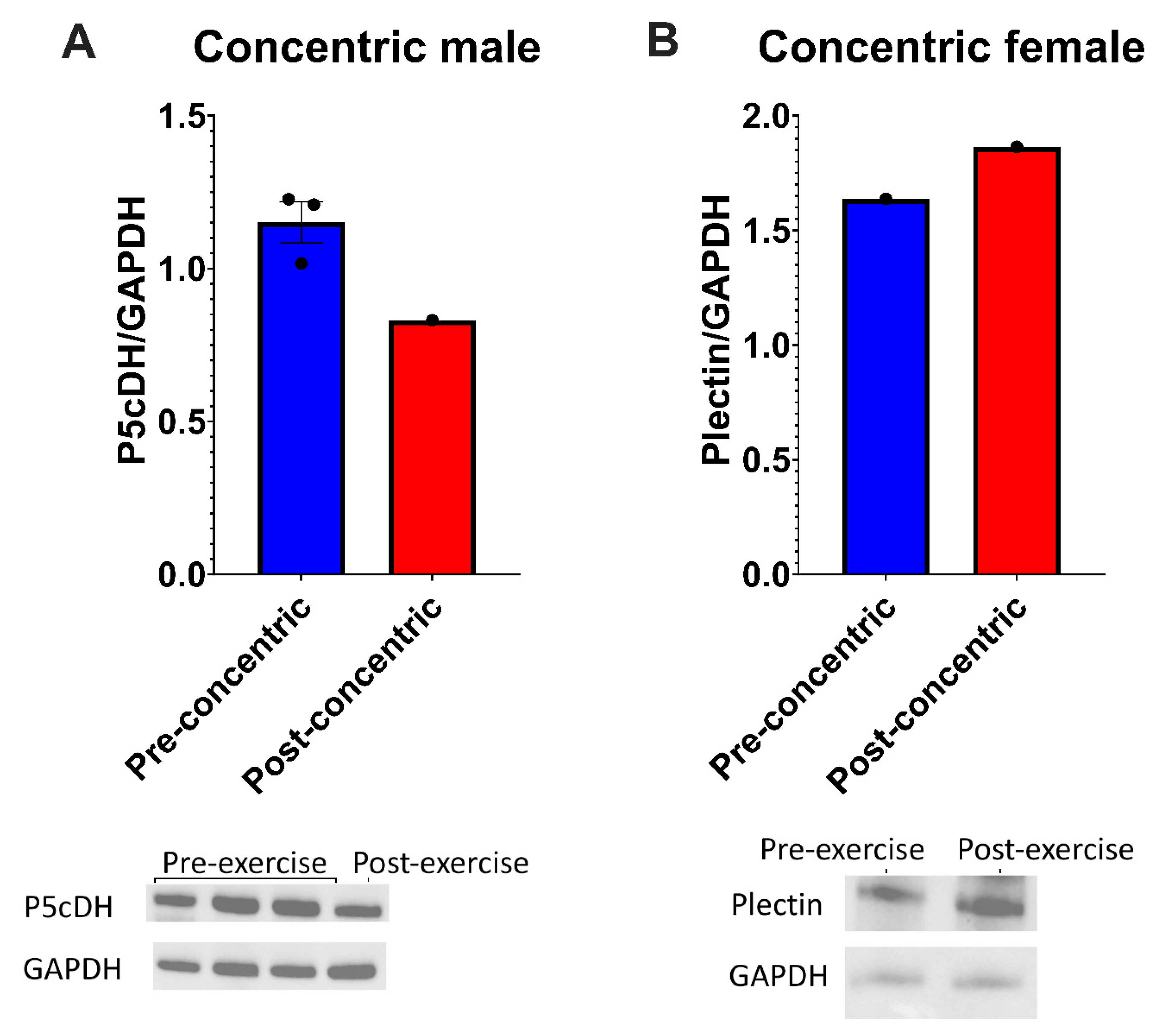
Figure 4.
the CpG island upstream of the P5cDH gene (ALDH4A1) is unmethylated in human skeletal muscle and remains unmethylated post-concentric exercise. The beta value (number of methylated reads/total number of reads) was not significantly different for any of the CpG sites (numbered on x-axis) located on chromosome 1: 18902000 – 18903000 (multiple unpaired t tests, error bars = SEM, N=3 biological replicates).
Figure 4.
the CpG island upstream of the P5cDH gene (ALDH4A1) is unmethylated in human skeletal muscle and remains unmethylated post-concentric exercise. The beta value (number of methylated reads/total number of reads) was not significantly different for any of the CpG sites (numbered on x-axis) located on chromosome 1: 18902000 – 18903000 (multiple unpaired t tests, error bars = SEM, N=3 biological replicates).
Figure 5.
Protein-protein interaction network of proteins with significantly altered expression in concentric exercise male participants. Nodes are proteins, and edges indicate functional and physical interactions, with edge thickness indicative of the strength of supporting data. Red nodes show local network cluster enrichment for the term ‘eukaryotic translation elongation’. The minimum required interaction score was 0.700 (high confidence), and the PPI enrichment p-value < 1.0e-16.
Figure 5.
Protein-protein interaction network of proteins with significantly altered expression in concentric exercise male participants. Nodes are proteins, and edges indicate functional and physical interactions, with edge thickness indicative of the strength of supporting data. Red nodes show local network cluster enrichment for the term ‘eukaryotic translation elongation’. The minimum required interaction score was 0.700 (high confidence), and the PPI enrichment p-value < 1.0e-16.
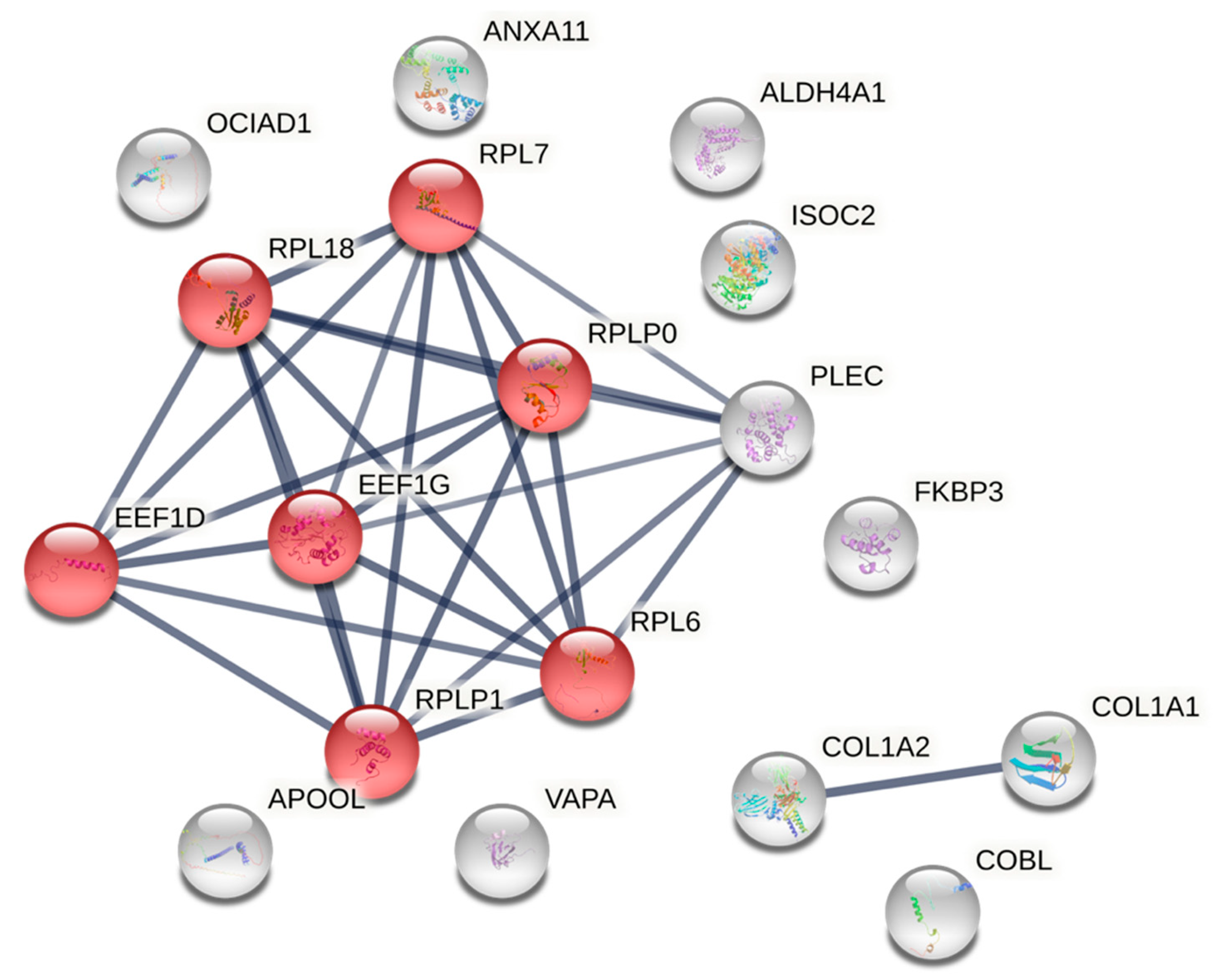
Figure 6.
Expression of haemoglobin subunits, haptoglobin, hemopexin, and carbonic anhydrase I, II and III across both sexes and exercise conditions. M = male, F = female, C = concentric, E = eccentric. Relative percentage abundance of A) haemoglobin subunit α; B) haemoglobin subunit β; C) haemoglobin subunit δ; D) haptoglobin; E) hemopexin; F) carbonic anhydrase I; G) carbonic anhydrase II; H) carbonic anhydrase III. Data presented as mean of relative percentage abundance within concentric exercise samples and eccentric exercise samples, n=3 biological replicates, SEM, one-way ANOVA P * < 0.05.
Figure 6.
Expression of haemoglobin subunits, haptoglobin, hemopexin, and carbonic anhydrase I, II and III across both sexes and exercise conditions. M = male, F = female, C = concentric, E = eccentric. Relative percentage abundance of A) haemoglobin subunit α; B) haemoglobin subunit β; C) haemoglobin subunit δ; D) haptoglobin; E) hemopexin; F) carbonic anhydrase I; G) carbonic anhydrase II; H) carbonic anhydrase III. Data presented as mean of relative percentage abundance within concentric exercise samples and eccentric exercise samples, n=3 biological replicates, SEM, one-way ANOVA P * < 0.05.
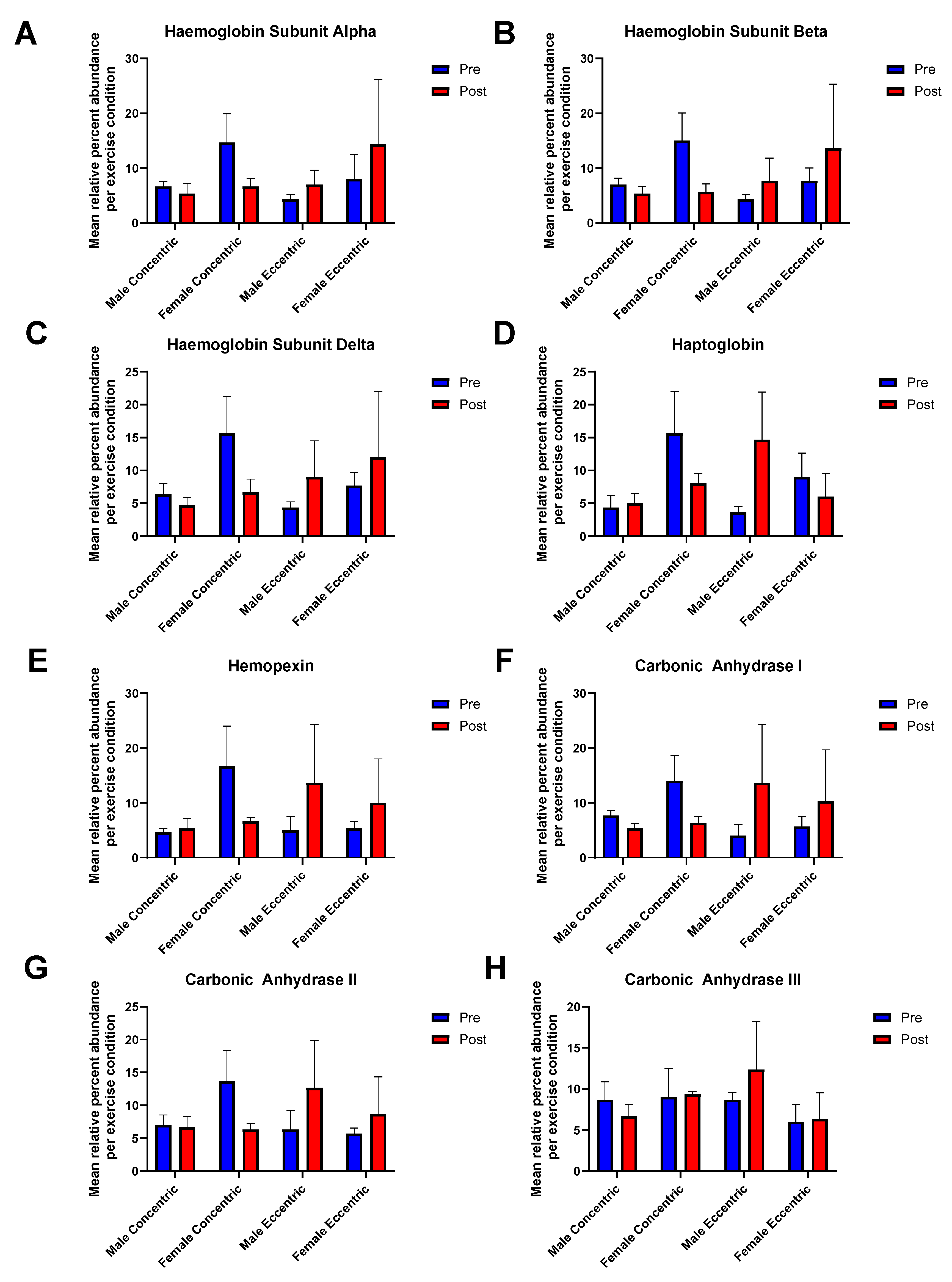
Figure 7.
ATPase activity and lipid peroxidation levels of male quadriceps samples pre- and post-concentric and eccentric exercise. A) Exercise resulted in a slight, but not significant decrease in ATPase activity (U/mL) of COPD patient quadriceps tissue; B) Exercise resulted in no significant difference in the level of lipid peroxidation (MDA) in the tissue samples post-concentric and eccentric exercise. n=3 biological replicates, error bars = SEM, unpaired t test *p = 0.05.
Figure 7.
ATPase activity and lipid peroxidation levels of male quadriceps samples pre- and post-concentric and eccentric exercise. A) Exercise resulted in a slight, but not significant decrease in ATPase activity (U/mL) of COPD patient quadriceps tissue; B) Exercise resulted in no significant difference in the level of lipid peroxidation (MDA) in the tissue samples post-concentric and eccentric exercise. n=3 biological replicates, error bars = SEM, unpaired t test *p = 0.05.

Table 1.
Proteins with significantly altered quantity in pairwise comparisons of concentric and eccentric exercise conditions for male and female participants. 18 proteins had altered abundance in response to concentric exercise in males, and 8 proteins had altered abundance in response to concentric exercise in females. 7 proteins had altered abundance in response to eccentric exercise in males, and 3 proteins had altered abundance in response to eccentric exercise in females. M= male, F = female. Mean values of relative peak areas, unpaired t test, * P < 0.05. Blue = increased expression, red = decreased expression. Protein names in bold were considered interesting for further study.
Table 1.
Proteins with significantly altered quantity in pairwise comparisons of concentric and eccentric exercise conditions for male and female participants. 18 proteins had altered abundance in response to concentric exercise in males, and 8 proteins had altered abundance in response to concentric exercise in females. 7 proteins had altered abundance in response to eccentric exercise in males, and 3 proteins had altered abundance in response to eccentric exercise in females. M= male, F = female. Mean values of relative peak areas, unpaired t test, * P < 0.05. Blue = increased expression, red = decreased expression. Protein names in bold were considered interesting for further study.
| Sex | Exercise condition | Protein | Fold change | P value |
|---|---|---|---|---|
| Male | Concentric | Elongation factor 1-delta | 5.666 | 0.039 |
| Annexin A11 | 3.166 | 0.045 | ||
| Plectin | 1.971 | 0.000 | ||
| Elongation factor 1-gamma | 1.59 | 0.030 | ||
| MICOS complex subunit MIC27 | 0.515 | 0.042 | ||
| 60S acidic ribosomal protein P1 | 0.449 | 0.015 | ||
| OCIA domain-containing protein 1 | 0.449 | 0.048 | ||
| Protein cordon-bleu | 0.394 | 0.038 | ||
| Delta-1-pyrroline-5-carboxylate dehydrogenase mitochondrial (P5cDH) | 0.375 | 0.042 | ||
| 60S acidic ribosomal protein P0 | 0.370 | 0.010 | ||
| Isochorismatase domain-containing protein 2 | 0.333 | 0.026 | ||
| Peptidyl-prolyl cis-trans isomerase FKBP3 | 0.316 | 0.012 | ||
| Vesicle-associated membrane protein-associated protein A | 0.316 | 0.037 | ||
| 60S ribosomal protein L18 | 0.253 | 0.038 | ||
| 60S ribosomal protein L6 | 0.250 | 0.032 | ||
| Collagen alpha-2(I) chain | 0.176 | 0.049 | ||
| 60S ribosomal protein L7 | 0.099 | 0.002 | ||
| Collagen alpha-1(I) chain | 0.000 | 0.018 | ||
| Eccentric | Alpha-1-acid glycoprotein 1 | 3.166 | 0.012 | |
| Hydroxyacylglutathione hydrolase mitochondrial (HAGH/GLO2) | 1.631 | 0.033 | ||
| Kelch-like protein 41 | 0.597 | 0.046 | ||
| Calpain-1 catalytic subunit | 0.578 | 0.015 | ||
| Myosin-9 | 0.429 | 0.043 | ||
| Complement C4-B | 0.408 | 0.033 | ||
| Cytosol aminopeptidase | 0.282 | 0.024 | ||
| Female | Concentric | Collagen alpha-1(VI) chain | 4.263 | 0.013 |
| Alpha-1-syntrophin | 3.949 | 0.022 | ||
| Collagen alpha-3(VI) chain | 3.166 | 0.000 | ||
| Myosin regulatory light chain 2 ventricular/cardiac muscle isoform | 2.961 | 0.028 | ||
| Heat shock protein beta-7 | 2.667 | 0.049 | ||
| Plectin | 2.333 | 0.041 | ||
| Annexin A6 | 1.564 | 0.038 | ||
| Vitronectin | 0.282 | 0.032 | ||
| Eccentric | T-complex protein 1 subunit delta | 0.754 | 0.049 | |
| Ryanodine receptor 1 | 0.190 | 0.023 | ||
| MICOS complex subunit MIC13 | 0.000 | 0.033 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



