Submitted:
29 July 2025
Posted:
30 July 2025
You are already at the latest version
Abstract
In this study, we evaluated the cytotoxicity and antioxidant activity of the soil ciliate Rigidohymena tetracirrata exposed to single and bimetallic mixtures of heavy metals (HMs) for 24 h. Ecotoxicological tests showed LC20 values of 0.51, 0.22 and 23.0 mg l-1 to Cd, Cu, and Zn, respectively, and LC50 values of 1.18, 0.37 and 32.7 mg l-1, respectively. Furthermore, it was observed that the mixture of Cd and Zn exhibited antagonism in comparison to other mixtures, (Cd+Cu and Cu+Zn). In the total phenolic content (TPC) assay, a higher phenolic content was observed for the LC20 of extracellular Cu (p ≤ 0.01) and LC20 of intracellular Cd (p ≤ 0.001). The LC50 values for Cd and Zn in both extracellular and intracellular contents demonstrated increased α,α-diphenyl-β-picrylhydrazyl (DPPH) scavenging activity with significant values of p ≤ 0.05, respectively. Regarding Hydroxyl scavenging activity (HRSA), the LC50 of extracellular Cd (p ≤ 0.001) and LC50 of intracellular Cu (p ≤ 0.001) exhibited higher antioxidant activity. Therefore, the present study suggests that R. tetracirrata holds considerable potential as bioindicators and could be used as a model organism in ecotoxicological studies of soil polluted by HMs.
Keywords:
acute toxicity
; heavy metals
; bimetallic mixtures
; antioxidant
; TPC
; DPPH
1. Introduction
Soil pollution is one of the most significant problems facing the ecosystems and has enormously increased during the last decades due to intensive human activities (use of biocides and fertilizers, industries, urban waste and atmospheric deposition) [1,2]. Soil pollution causes different hardships on soil fertility, soil structure, flora and fauna residing in the soil, crops and groundwater contamination [2]. In soil ecosystems, different types of pollutants exist, with one of the most predominant issues being the contamination of soil by heavy metals [2,3]. Metals are continuously released into the soil by emissions from the rapidly expanding industrial areas, mine tailings, disposal of high metal wastes, leaded gasoline and paints, land application of fertilizers (agriculture), animal manures, sewage sludge, pesticides, wastewater irrigation, spillage of petrochemicals, and atmospheric deposition [1,2,3,4,5].
The heavy metals contaminations may change the soil properties, especially soil biological properties [6]. The toxicity of heavy metals on soil organisms depends on several factors such as soil temperature, pH, clay minerals, organic matter, inorganic anions and cations, and chemical forms of the metal [6,7,8]. Moreover, heavy metals do not degrade but accumulate in the food chain [2] and additionally, some of them produce carcinogenic and toxic effects in humans and animals [2,9]. Heavy metals, such as zinc and copper, play a vital role in organismal metabolism but can be toxic at high concentrations [10,11]. On the other hand, non-essential metals like cadmium (Cd), lead (Pb), and mercury (Hg) have no known biological function and can be toxic even at low concentrations [12]. Over the last few decades, there has been a significant increase in the discharge of anthropogenic heavy metals into ecosystems worldwide.
Nowadays the approach to determining heavy metals soil pollution is still based on the analysis of the concentrations of metals in the soils and comparison with specific threshold values. This approach, however, does not provide indications of their deleterious effects on biota [1,2]. On the other hand, another important aspect to studying the toxicity of heavy metals in soil ecosystems is the evaluation of bioavailability and the interactive effects (synergism and/or antagonisms) with other pollutants or soil components [13]. In ecotoxicology, biomarkers and bioassays are used to evaluate the toxic action of pollutants in the environment and establish contaminant bioavailability. These tests use bioindicator organisms that are ecologically relevant [1,2]. Invertebrates may serve as effective bioindicators of soil pollution as they are in direct contact with soil pore water or food, unlike many vertebrates that are indirectly exposed through the food chain [2,5]. The most utilized bioindicator species are collembola [13,14], earthworms [1,2,5] and snails [15,16].
Another class of bioindicator organisms are the single-celled ciliated protists (ciliates), which are recognised for their vital role in ecosystems. These eukaryotic micro-organisms have been found in almost any habitat where there is sufficient water for their survival, including soils [17,18,19]. Ciliate diversity is largely unknown, with only 8,000 free-living and epibiotic ciliate species identified to date [19]. Ciliates contribute significantly to ecosystem dynamics by decomposing organic matter and transferring energy to higher trophic levels [20,21,22]. They maintain the balance of the ecosystem and play a crucial role in regulating microbial food webs, and this activity appears stimulating mineralization processes in soils [23]. Soil ciliates also have beneficial effects on plant growth through a mechanism known as the “soil microbial loop” [24]. Ciliates are excellent biological indicators of both chemical pollution, particularly heavy metal pollution, and various other types of environmental disturbances [12,22,25,26,27]. Soil ciliate communities provide rapid, unique and essential ecosystem information in comparison with more traditional macroscopic bioindicators [28]. Compared to other microorganisms like bacteria and algae, there are fewer studies on the relationship between ciliates and environmental pollutants. However, ciliates are easy to culture and maintain, making them a good model organism for bioassays [29,30].
Bioassays are widely regarded as effective tools for the assessment of metal pollution, due to their ability to specifically react and detect the available fraction of metal ions. Metal ions have been demonstrated to trigger the formation of reactive oxygen species (ROS), including superoxide anion (O2-), hydroxyl radicals (OH-), hydrogen peroxide (H2O2) and singlet oxygen (1O2), through both normal metabolic activities and external sources [31]. It is notable that high levels of ROS can be generated under conditions of heavy metal exposure, resulting in the overloading of antioxidant mechanisms [12,32]. Ciliates, in particular, experience impacts on their ability to feed and growth rates, as well as changes in respiration and oxidative stress due to the generation of high levels of ROS caused by heavy metal pollution [25,33]. However, to date there are no data on the antioxidant properties of soil ciliate protists. This is a novel study and the first to investigate the antioxidant properties of the soil ciliated protist Rigidohymena tetracirrata in response to heavy metal stress.
The main aim of this study was to assess the cytotoxicity of heavy metals (Cd, Cu and Zn) on the soil ciliate R. tetracirrata. To this end, the ciliate was exposed to a range of concentrations of both single and binary mixtures of heavy metals for a duration of 24 hours. In addition, the antioxidant activities and total phenolic content of intracellular and extracellular extracts of R. tetracirrata were also examined. The obtained results highlight the effectiveness of the antioxidant properties of R. tetracirrata in scavenging free radicals and metal ions. It also provides evidence that ciliates may be an excellent source of natural antioxidants. Overall, this study suggests that R. tetracirrata can be a suitable eukaryotic model organism for the bioremediation of heavy metal-polluted habitats and an effective biological indicator of environmental monitoring.
2. Materials and Methods
2.1. Ciliate Strain and Culture Conditions
The experimental organism used in our study was the ciliated protist, isolated from the soil of an agriculture farm in the Marche region, Italy [34]. Ciliates were grown in Pringsheim’s Medium (PM) [35] at the temperature of 18 ± 1 °C. Cells were kept in the dark and were fed with the green alga Chlorogonium elongatum and analyses were conducted during their logarithmic growth phase.
2.2. Metal Salts (Chemicals)
For the ecotoxicological assays, analytical grade pure chemicals (purity ≥ 99%) were used as a source of metal ions: cadmium chloride (anhydrous CdCl2), zinc sulphate (ZnSO4.7H2O) and copper (II) sulphate (CuSO4.5H2O) from Sigma (Milan, Italy). Stock solutions (0.1M) were made by dissolving each metal salt in PM (pH 7). Working solutions were daily prepared by diluting stock solutions to the following nominal concentrations: 0–2.5 mg Cd/l, 0–0.7 mg Cu/l and 0–45 mg Zn/l. Trypan Blue (TB) was purchased from Merck (Milan, Italy).
2.3. Toxicity Tests with Single Metals (Cd, Cu, and Zn)
To analyse the toxicity of heavy metals Cd, Cu and Zn on the selected soil ciliate species R. tetracirrata, preliminary toxicity range-finding tests using a different set of metal concentrations, were conducted to determine the range of concentrations to be used in the final tests. For each experiment test, a set of 6-12 different concentrations were tested, arranged so that the lowest concentration would have no effect on cell viability and the highest concentration would produce more than 50% loss of viability. Ecotoxicity assays were performed using specific 3-well depression glass slides. One hundred cells were picked from exponentially growing cultures with a micropipette and inoculated into each well in a final volume of 1 ml of PM medium containing the selected metal concentration, covered to avoid evaporation and incubated in a humid chamber at the temperature 18 ± 1 °C for 24 hrs. Ciliates were not fed during the test period. After a 24 hrs period, the cells were checked for accountancy of mortality and survivorship under the different test concentration solutions under a stereoscopic microscope at 20-40x magnifications. Ciliates were accounted as dead when missing due to cell burst or when standing still at the bottom of a well, unable to swim even after gentle mechanical stimulation with the tip of a micropipette. Furthermore, viability tests using the Trypan Blue (TB) exclusion tests were realised according to Strober [36]. As a control, the same number of ciliates was inoculated into wells containing 1 ml of PM medium. Replicates (n=3) for each concentration treatment were averaged. The endpoints utilized for cytotoxicity assessment were the median Lethal Concentration (LC) LC20 (concentration causing mortality in 20% of the cells) and LC50 (concentration causing mortality in 50% of the cells) at 24 hours. Mean mortality values were used to derive a regression equation.
2.4. Cytotoxicity Bioassays with Bimetallic Mixtures (Cd + Zn, Cu + Zn, and Cd + Cu)
The bimetallic cytotoxicity bioassays were designed according to Gallego [37] with slight modifications. This procedure is based on the Concentration Addition (CA) model that uses the individual metal toxicities (LCX) and the Toxic Unit (TU) concept [38]. The dimensionless TU represents the relative contribution to the toxicity of the individual chemical in a mixture of chemicals. In this study, the bimetallic mixtures were prepared by combining the toxicants according to their individual LC50 values. The effects of three different bimetallic mixtures were then analysed (Cd + Zn, Cu + Zn, and Cd + Cu). 1TU corresponds to the LC50 values of single metal toxicities. The total TU of the binary mixture was the sum of its individual metal fractions. In this experiment, we selected and tested four different concentrations of TU (0.5, 0.75, 1, and 1.25 TUs) to cover a wide range of metal concentration mixtures. The expected mortality rates were calculated as the sum of the single metal toxicities obtained for each metal in the mixture, at the same concentration. If the observed mortality rate is higher than that of the expected mortality rate, the interaction is synergistic. At the same time, if the observed mortality is lower than the expected death rate, the interaction is antagonism. Cytotoxicity with bimetallic mixtures was performed using the same test method as defined above for the single metal treatment assay.
2.5. Total Phenolic Content and Antioxidant Activity Assays
Three different types of antioxidant assays, namely Total Phenol Content (TPC), α,α-diphenyl-β-picrylhydrazyl (DPPH), and Hydroxyl Radical Scavenging Assay (HRSA), were applied to evaluate the presence of antioxidant activity in R. tetracirrata exposed to different concentration of the single and bimetallic mixture of heavy metals. To analyse, the antioxidant activity of R. tetracirrata cells (2000 cells/ml) from an exponentially growing culture were transferred into petri plates in the volume of 10 ml and exposed to different concentrations of single and bimetallic mixtures of heavy metals for 24 hrs. In single metal exposure, the LC20 and LC50 concentrations were used to analyse the antioxidant activity. In the bimetallic mixture, Cd + Zn mixtures showed more antagonistic activity. On this basis, eight concentrations were selected for antioxidant analysis, i.e.,: 0.25 + 0.25, 0.5 + 0.25, 0.75 + 0.25, 0.5 + 0.5, 0.25 + 0.5, 1 + 0.25, 0.75 + 0.5, and 0.25 + 1 TUs (Cd + Zn).
Total phenolic content and antioxidant activities were measured in both the extracellular and intracellular fractions of the cell culture. After 24 h exposure to heavy metals, the ciliates were separated from the medium by gentle centrifugation (4000 rpm for 10 – 15 min). The cells pellet was then washed in distilled water and resuspended in 1 ml of 50 mM phosphate buffer (pH 7.0). The cells were then homogenised using a Teflon homogeniser for 4-5 minutes and resuspended in 1 ml of 50 mM phosphate buffer (pH 7.0). The homogenate was then centrifuged at 4000 rpm for 30 minutes and the resulting pellet was collected. This pellet was termed ‘intracellular’. The extracellular fraction was obtained from the ciliate cell culture. Both extracellular and intra-cellular samples were stored at -20°C until analysis. All the antioxidant assays were performed according to the methods by Ravindran, et al. [39].
2.6. Estimation of Total Phenolic Content (TPC)
The analysis of TPC was based on the Folin assay of Vattem and Shetty [40]. In this method, 100 μl of ciliates cell extract was added into the test tube, mixed with 2 ml of sodium bicarbonate, and incubated for 2 min at 18 ± 2 °C. Then, 100 μl of Folin-Ciocalteu reagent was added and incubated in the dark at 18 ± 2 °C for 30 min. After the incubation period, the absorbance was measured at λ= 725 nm using the plate reading spectrophotometer (FLUOstar Omega, BMG LABTECH, Ortenberg, Germany). Gallic acid 1 mg/ml was used as standard and standard curves were obtained using various concentrations of gallic acid. We also used the culture medium (without cells) as a control, as no phenols were observed.
2.7. DPPH Scavenging Assay
We used the DPPH scavenging assay to assess the antioxidant capability in the ciliate extracts. Antioxidants activity was determined by its ability to scavenge 1, l-diphenyl-2-picrylhydrazyl (DPPH) radicals. This analysis is based on the procedure of Yıldırım, et al. [41]. Briefly, 1mM DPPH stock solution was prepared in the 95% of ethanol. 800 μl of 1mM DPPH solution was added to 200 μl of cell extract. The samples were mixed well and incubated in the dark for 30 min at room temperature. After incubation, the samples were transferred to centrifuge tubes and centrifuged at 14,000 rpm for 5 min. Then, the supernatant was collected and the absorbance measured at λ = 517 nm using a spectrophotometer (FLUOstar Omega, BMG LABTECH, Ortenberg, Germany). 200 μl of 95% ethanol was used as a control. Butylated Hydroxy Anisole (BHA) was used as reference compounds. Antioxidant activity is expressed as a percentage (%). The percentage of DPPH scavenging was calculated using the formula below:
2.8. Hydroxyl Radical Scavenging Assay (HRSA)
HRSA was evaluated based on the fenton reaction [42] and followed the procedure of Ravindran et al. [39] with slight modification. The hydroxyl radical is produced by the Fe3+ascorbate EDTA H2O2 system i.e., fenton reaction. To analysis, this reaction, the chemicals of 2–deoxyribose (2.8 mM), FeCl3 (100 µM), H2O2 (1 mM), and EDTA (100 µM) were dissolved in the 20mM phosphate buffer at pH 7.4. From this chemical mixture, 800 µl was transferred to the test tube. Then 10 µl of ascorbic acid (10 mM) and 100 µl sample extract was added. The ascorbic acid was added and incubated at 37 ºC for 1 h. After the incubation, 1 ml of 2.8 % TCA, and 1 ml of 1 % aqueous TBA were added, and the mixture was heated at 90 ºC for 15 min to develop colour. After cooling the absorbance was measured at 532 nm against a suitable blank solution. Mannitol was used as a reference compound. The percentage of scavenging was determined using the formula below.
2.9. Statistical Analysis
Statistical analysis was performed using the InfoStat software ver. 2012; p ≤ 0.05 was regarded as statistically significant. The differences between control and test groups were evaluated using the Student’s t-test. All experiments were performed in triplicate or more, and the results were expressed as mean values ± standard error (SE) values.
3. Results
3.1. Cytotoxicity of Single Metals (Cd, Cu and Zn)
In this study, we have analysed the toxicity of three heavy metals (Cd, Cu, and Zn) on R. tetracirrata. The results obtained indicate that R. tetracirrata exhibits the highest degree of resistance to Zn. In comparison with Cu, the cellular toxicity of Zn and Cd is lower. The most sensitive effect of heavy metals on R. tetracirrata was observed with Cu. However, in comparison to Cu, the ciliate exhibits a heightened susceptibility to the cytotoxic effects of Cd and Zn. The LC20 and LC50 values for Cd are 0.53 and 1.16 mg l-1, and for Zn are 23.0 and 32.7 mg l-1, respectively (Table 1). The LC20 and LC50 values for Cu are 0.22 and 0.37 mg l-1(Table 1) respectively. It is evident that Cu is the most toxic heavy metal for this ciliate species. The decreasing order of toxicity is as follows: Zn > Cd > Cu.
3.2. Cytotoxicity of Bimetallic Mixtures (Cd + Zn, Cu + Zn and Cd + Cu)
In the analyses of bimetallic mixtures, three types of mixtures (Cd + Zn, Cu + Zn, and Cd + Cu) were used. The calculation of the expected level of cytotoxicity was performed based on the percentage graph of the death rate. (Figure 1a–c).
Our study shows that exposure to bimetallic mixtures of Cd + Zn, resulted in a greater degree of antagonistic effects (see Table 2) compared to other bimetallic mixtures. The ciliate exhibited antagonistic activity under concentrations of 0.25 + 0.25, 0.5 + 0.25, 0.75 +0.25, 0.5 + 0.5, 0.25 + 0.75, and 1 + 0.25 (Cd + Zn). However, at higher concentrations exceeding 1 TU (Cd + Zn), the bimetallic mixture interaction changed based on the impact of each metal’s cytotoxicity. For instance, at 0.75 + 0.5 TU (Cd + Zn), the expected value for the median effective concentration (EC50) was 34, but the observed value was 45.83 ± 3.87, indicating a synergistic effect. Conversely at concentrations of 1 + 0.25 TU, an antagonism effect was observed, suggesting that 1 TU of Cd metal was less toxic when mixed with 0.25 TU of Zn metal (Table 2).
The remaining two sets of bimetallic mixtures, Cu + Zn and Cd + Cu, showed similar outcomes with minimal variation, predominantly yielding effects that were not considerably disparate. Cu + Zn bimetallic mixtures exhibited synergetic effects at three different TU concentrations, influenced by the higher impact of Zn compared to Cu (Table 3).
Furthermore Cd + Cu mixtures also showed synergistic interactions, particularly at concentrations 0.75 + 0.25, 0.5 + 0.5, 0.25 + 0.75 and 0.25 + 1.0 TU, while antagonistic interactions at concentrations 0.5 + 0.25 and 0.75 + 0.5 TU. In these mixtures, Cd metal was found to be less toxic to R. tetracirrata than Cu. However, at 1 TU exposure, the interaction became synergistic, indicating that the impact of Cu was higher than that of Cd in terms of its cytotoxic effect (Table 4).
3.3. Antioxidants Properties of R. tetracirrata Extracts Treated with Cd, Cu, Zn, and Cd +Zn
3.3.1. Total Phenolic Content (TPC) from Extracts of R. tetracirrata
The total phenolic contents of ciliate cell extracts treated with single (Cd, Cu, and Zn) and bimetallic mixture (Cd +Zn) were analysed (Figure 2a,b). The highest TPC value of ciliate cell extracts was observed with Cu LC20 value of extracellular and Cd LC20 value of intracellular (p ≤ 0.01) (Figure 2a). In single metal treatments, intracellular TPC was generally higher than that of extracellular content, except for the extract of R. tetracirrata treated with Cu. Conversely in bimetallic mixture treatments, intracellular TPC was found to be lower than that of extracellular content. The TPC of ciliate cell extracts was highest with 0.25 + 0.25 TU (Cd + Zn) of extracellular and 0.75 + 0.25 TU (Cd + Zn) for intracellular (p ≤ 0.001) (Figure 2b). However, the total phenolic contents and the antioxidant capacity of cells exposed to different single heavy metal conditions were found to be significantly increased compared to untreated cells (p ≤ 0.01) and Cd + Zn conditions were also found to be significantly increased compared to the control (p ≤ 0.001) (Figure 2a,b). When the cells were treated with a single metal, phenolic content was released at µg l-1 level, whereas bimetallic mixture-treated cells released phenolic content at mg l-1 level. It is noteworthy, that the TPC released in bimetallic mixture-treated cell extracts was 103 times higher compared to the single metal-treated ciliate extracts.
3.3.2. DPPH Scavenging Activity from Extracts of R. tetracirrata
The free radical scavenging activities of ciliate cell extracts was determined by the DPPH assay. The ciliate cell extracts in this study showed a concentration dependent antiradical activity by inhibiting DPPH radicals (Figure 3a,b). In single metal treatments, the Zn LC50 and Cd LC50 values in intracellular and extracellular contents exhibited a higher DPPH scavenging activity, with a significant value of p ≤ 0.05 (Figure 3a). in summary, DPPH scavenging activity in single metal-treated ciliate extracts was found to be higher in extracellular than intracellular contents. The results of the study demonstrated that both single metals (Cd, Cu, and Zn) and bimetallic mixtures (Cd + Zn) exhibited scavenging activity; with bimetallic mixtures (Cd + Zn) showing more pronounced DPPH activities. The maximal activity was observed with 0.25 + 0.25 TU (Cd + Zn) in intracellular extract and 0.75 + 0.5 TU (Cd + Zn) in extracellular extract (p ≤ 0.001) (Figure 3b). Furthermore, an increase in antioxidant capacity was observed in cells exposed to different heavy metal conditions (single metal, p ≤ 0.05 and bimetallic, p ≤ 0.001) in comparison to untreated cells (control).
3.3.3. Hydroxyl Radical Scavenging Activity of Extracts of R. tetracirrata
The findings of this study demonstrate that the hydroxyl radical scavenging activity (HRSA) of ciliate extracts, when subjected to both single and bimetallic mixtures, exhibited an increase in response to elevated metal concentrations (see Figure 4a,b). In single metal treatments, the LC50 for the extracellular extract of Cd and the LC50 for the intracellular extract of Cu exhibited higher hydroxyl radical scavenging activity (p < 0.001) (Figure 4a). The assay shows that ciliate cell extracts possess the capacity to inhibit hydroxyl radical-mediated deoxyribose degradation in a Fe3+ EDTA ascorbic acid and H2O2 reaction mixture. In the bimetallic mixture treatments, the intracellular HRSA was found to be lower in comparison to the extracellular content. The highest levels of HRSA were observed in ciliate cell extracts with 0.25 + 0.25 TU (Cd + Zn) for extracellular and 0.25 + 1 TU (Cd + Zn) for intracellular conditions (p ≤ 0.001). However, the HRSA of extracellular and intracellular antioxidant capacity of cells exposed to different single and bimetallic mixture (Cd + Zn) heavy metal conditions was significantly increased compared to untreated cells (p ≤ 0.001) (Figure 4b).
4. Discussion
The majority ecotoxicological studies on heavy metal cytotoxicity have focused on freshwater protozoa, those isolated from urban wastewater treatment plants, or standard ciliates like Tetrahymena sp. [25,43,44]. To date, there has been a paucity of research on the toxic effects of heavy metals specifically on soil ciliates [45,46]. In the present study, we have evaluated the cytotoxicity of Cd, Cu, and Zn on the population of the ciliate R. tetracirrata. The outcome of our study indicates that the observed toxicity sequence of metals used is Cu > Cd > Zn. The median lethal concentration (LC50) values for Cd, Cu, and Zn heavy metals were estimated to be 1.16, 0.37, and 32.7 mg/L, respectively. Furthermore R. tetracirrata cells appear to be more resistance to Zn, Cd and Cu than other freshwater ciliate specie studied to date [44,46]. This finding agrees with the observations reported by Madoni and Romeo [47], who found that the freshwater ciliate Colpidium colpoda exhibited higher toxicity to Cu than to Cd. This finding is consistent with the observations of Díaz et al. [45] and Martín-González [48], who found that the order of toxicity was Cd > Cu > Zn in ciliated protists isolated from soils and urban wastewater treatment plants. However, in comparison with the data presented here, these values and the toxicity order rank are lower. On the other hand, the ciliate T. pyriformis is more resistant to these metals [47,48]. Echavez and Leal [49] isolated some ciliates from Lake Maracaibo and discovered that Euplotes sp. (18.5 mg l-1), Oxytricha sp. (9.2 mg l-1), Coleps sp. (53.3 mg l-1), and Chilodonella sp. (19.2 mg l-1) were more resistant to Cd after a one-hour exposure. Meanwhile, Marín-Leal, et al. [50] isolated some ciliates from the same lake and found that Uronema sp. (0.71 mg l-1), Euplotes sp. (0.72 mg l-1), and Loxodes sp. (0.14 mg l-1) were more sensitive to Cd after a one-hour exposure. The LC50 values for these species were found to exceed the values obtained in this study. Gallego et al. [37] reported that the LC50 for Cd, Cu, and Zn in the ciliate T. thermophila were 0.195, 0.47 and 3.58 mg l-1, respectively. These values are lower than those obtained in the present study. Consequently, it can be concluded that our strain of R. tetracirrata generally exhibits increased sensitivity to Zn, Cd, and Cu in comparison with other ciliated protists.
In the context of bimetallic mixtures, Cd + Zn exhibited a greater degree of antagonistic activity in comparison to other metallic mixtures including Cu + Zn and Cd + Cu. Furthermore R. tetracirrata showed that both metallic components within the mixture were significant contributors to biotoxicity. In R. tetracirrata, the combined effects of Cd + Zn at 0.5 to 1 TUs were antagonistic, but under high concentrations of heavy metal (above 1 TUs) the interaction changed to synergistic. Most studies have shown that the toxicity of Cd is reduced in the presence of Zn. However, some studies have reported that the presence of low or moderate Zn concentrations, enhances Cd cytotoxicity in ciliated protists. [45,48]. In contrast to the findings of numerous studies showing a decrease in Cd toxicity in the presence of Zn, our observations indicate that R. tetracirrata exhibits increased resistance to Cd when simultaneously exposed to Zn. In this soil ciliate, the presence of low and moderate concentrations of Zn reduces the biological toxic effect of Cd. This phenomenon has been observed in other organisms, including T. pyriformis, T. thermophila, Daphnia magna and other invertebrates [51].
The combined toxic effects of Cu + Zn and Cd + Cu could be considered either antagonistic or synergistic, depending on the toxicity units (TUs) tested in R. tetracirrata. It was observed that both metallic mixtures (Cu + Zn and Cd + Cu) exhibited a similar behaviour, with case of antagonism, synergism, and, in most concentrations, no significant differences (Table 3 and Table 4). In the Cu + Zn bimetallic mixture, the type of interaction shifts from antagonism to synergism with the addition of Zn. Concurrently, the toxicity exhibited by the Cu + Zn mixture was found to exceed that induced by Cu alone. R. tetracirrata demonstrates variations from antagonism to synergism, depending on the metal concentration. Consistent with these observations, analogous results have been reported in other organisms including the ciliate T. thermophila [37], bacteria [52], microalgae [53], crustaceans [54], insects [51], and plants [55].
However, the antagonism of Cu + Zn, remains ambiguous, and it is conceivable that Cu bioaccumulation is not a substantial process in this ciliate species. In the context of heavy metal toxicity, such as elevated Cu levels within cells, the formation of reactive oxygen species (ROS) has been observed, potentially resulting in damage to lipids, nucleic acids, and proteins [56]. In Cd + Cu interaction, analogous activities have been observed, though it is generally regarded as a more pronounced effect than simple addition [57]. This type of interaction in the binary metal combinations of Cd + Cu has been observed by Gallego [37] in the ciliate T. thermophila.
This study also analysed the total phenolic content (TPC) and antioxidant activity [33] of this soil ciliate species, i.e., R. tetracirrata were analysed. The study examined the TPC, and antioxidant activity of cell extracts from R. tetracirrata, confirming the presence of phenols and antioxidants. Phenols, as primary chemical elements, are responsible for reducing lipid peroxidation and thus function as both primary and secondary antioxidants [58]. Furthermore, it was observed that cell extracts with high phenolic content also exhibited notable antioxidant activity. A strong correlation was observed between total phenol content and antioxidant activity, suggesting the importance of phenols in ciliates, due to their free radical scavenging ability, attributed to their hydroxyl groups. Consequently, the phenolic content of ciliates may directly contribute to their antioxidant activity. The ciliate cell extract in this study demonstrated a higher antioxidant capacity, attributed to its elevated phenolic content, particularly 66.7 mg l-1 in single metal treatments and 114.9 mg l-1 in binary metallic mixtures. These values exceed those previously reported in studies on other organisms. For instance, Ravindran, et al. [39] reported excellent antioxidant capacity in a halophilic pathogenic fungus (NIOCC 1) and the endophytic fungi Nerium oleander L. and liverwort Scapania verrucosa.
The findings of this study demonstrate that the soil-dwelling ciliate R. tetracirrata has antioxidant activity, indicating its potential as a natural antioxidant. This study is the first to investigate and report the total phenolic content (TPC) and antioxidant activity of R. tetracirrata cell extract and in general for ciliates. Since many of the widely recognised strong synthetic antioxidants often contain carcinogenic compounds, the search for potential antioxidants is crucial, particularly in the context of soil ciliates. Phenolic compounds, which are important constituents of plant and fungi, are valued for their hydroxyl groups, which confer scavenging ability [59]. Consequently, these compounds may play an important role in ciliates. In this study, we therefore set out to determine the total phenolic content of ciliate cell extracts, in view for evaluating their antioxidant activity against different concentrations of heavy metal stresses using various antioxidant assays.
Two different assays were employed in this study, with the DPPH free radical scavenging assay being a basic and widely used method. This assay is widely regarded as the most accurate screening method for evaluating antioxidant activity, using DPPH as the substrate. Cysteine, glutathione, ascorbic acid, tocopherol, polyhydroxy aromatic compounds (e.g., hydroquinone, pyrogallol, gallic acid), and aromatic amines (e.g., p-phenylene diamine, p-aminophenol), have been found to reduce and decolorise α,α-diphenyl-β-picrylhydrazyl through their hydrogen-donating ability. Previous studies, including Ravindran, et al. [39] on the marine-derived pathogenic fungus (NIOCC 1), reported DPPH scavenging activity. Further, Uddin, et al. [60] reported DPPH antioxidant activity in the plants Bergeniacaliata, and Horta, et al. [61] reported in the bacteria Bifurcaria bifurcata. The result of the ciliate cell extracts in this study demonstrate their hydrogen- donating capabilities, acting as antioxidants. Our experimental findings indicate that soil ciliate R. tetracirrata produces a higher level of antioxidants under conditions of heavy metals stress.
Hydrogen peroxide itself is not very reactive, but it has been observed to decompose rapidly into oxygen and water, thereby producing hydroxyl radicals (OH-). These radicals have been demonstrated to cause DNA damage [62]. Hydroxyl radicals are the primary active oxygen species responsible for lipid peroxidation and significant biological damage. In this study, hydroxyl radicals were generated by incubating ferric-EDTA with ascorbic acid and H2O2 at pH 7.4, and these radicals reacted with 2-deoxy-2-ribose to generate a malondialdehyde (MDA)-like product. This compound forms a pink chromogen upon heating with TBA at low pH. When ciliate cell extracts were added to the reaction mixture, they were found to remove the hydroxyl radicals from the sugar, thereby preventing the reaction. This outcome is consistent with the observations reported by Ravindran, et al. [39] and Sharma, et al. [63] in their studies of the free radical hydroxyl activity of the green hull of Juglans regia. Consequently, the in vitro study clearly demonstrated that the antioxidants present in the soil ciliate R. tetracirrata can inhibit the process of metal ion-dependent hydroxyl radical formation and mitigate hydroxyl radical induced cell damage under heavy metals stress conditions. Under such conditions, the role of antioxidant enzymes in cells is also significant. Numerous studies have documented the presence of antioxidant enzymes (Superoxide dismutase, Catalase, Glutathione peroxidase and reductase) and stress genes (Heat shock proteins and Metallothionein) in ciliate species of the genera Tetrahymena and Euplotes [25,64,65,66].
5. Conclusions
The soil ciliate R. tetracirrata demonstrates promising potential as a model organism for ecotoxicological studies. It also proves to be an excellent and suitable biosensor for evaluating the toxicity of soils contaminated with heavy metals. The varying concentrations of heavy metals tested in this study confirm the production of higher amounts of antioxidant compounds and scavenging activities in this ciliate. Consequently, R. tetracirrata emerges as an excellent source of natural antioxidant compounds and an excellent bioindicator for heavy metal contaminated soils. Future studies will include analysis of antioxidant enzymes and the expression of potential stress related genes (antioxidant genes) in R. tetracirrata.
Author Contributions
Conceptualization, A.L.T.; Methodology, G.R.V., A.G., S.S., A.L.T.; Software, G.R.V., A.C., F.D.; Validation, G.R.V., A.C., F.D., D.B., S.K., M.C., and A.L.T.; Formal analysis, G.R.V., A.L.T., F.D., A.C.; Investigation, G.R.V., A.L.T., S.K., D.B., A.G., S.S.; Resources, F.D., A.C.K, S.K., A.L.T.; Data Curation, G.R.V., A.L.T., F.D., A.C.; Writing—Original Draft Preparation, G.R.V., A.C., A.L.T.; Writing—Review and Editing A.C., F.D., D.B., S.K., M.C., A.G., A.C.K., S.S. and A.L.T.; Visualization A.C., A.L.T., M.C.; Supervision, A.L.T.; Project Administration, A.L.T.; Funding Acquisition, A.L.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the 2017 FFABR—“ Fund for Financing Basic Research Activities”—of the Italian Ministry of Education (BVI120008) to A.L.T.
Institutional Review Board Statement
This study did not require ethical approval according to the legislation in force—that is, Directive2010/63/EU of the European Parliament and of the Council.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are available when required under the responsibility of the corresponding author. The data are not publicly available due to privacy.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Calisi, A., Cappello, T., Angelelli, M., Maisano, M., Rotondo, D., Gualandris, D., Semeraro, T., Dondero, F. (2024). Non-Destructive Biomarkers in Non-Target Species Earthworm Lumbricus terrestris for Assessment of Different Agrochemicals. Environments, 11, 276. [CrossRef]
- Sanchez-Hernandez, J.C. (2019). Bioremediation of Agricultural Soils, 1st ed.; CRC Press: Boca Raton, FL, USA; p. 296, ISBN 9780367780173.
- Järup, L. (2003) Hazards of heavy metal contamination. British medical bulletin, 68(61), 167-182.
- Wuana, R.A., and Felix E. Okieimen (2011). Heavy metals in contaminated soils: a review of sources, chemistry, risks and best available strategies for remediation. Isrn Ecology, 1-20. [CrossRef]
- Calisi, A., Semeraro, T., Giordano, M.E., Dondero, F., Lionetto, M.G. (2025) Earthworms multi-biomarker approach for ecotoxicological assessment of soils irrigated with reused treated wastewater. Applied Soil Ecology, 206, 105866. [CrossRef]
- Michaela, F. (2010) The influence of heavy metals on soil biological and chemical properties. Soil Water Research, 5, no. 1: 21-27.
- Bååth, E. (1989) Effects of heavy metals in soil on microbial processes and populations (a review). Water, Air, and Soil Pollution, 47,43-44: 335-379. [CrossRef]
- Giller, K.E., Ernst Witter, and Steve P. Mcgrath (1998). Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: a review. Soil Biology and Biochemistry, 30(10): 1389-1414. [CrossRef]
- Chang, L.W.; Magos, L.; Suzuki, T. (1996). Toxicology of metals; CRC Boca Raton, FL.
- Xin, Z.; Wenchao, Z.; Zhenguang, Y.; Yiguo, H.; Zhengtao, L.; Xianliang, Y.; Xiaonan, W.; Tingting, L.; Liming, Z. (2015) Species sensitivity analysis of heavy metals to freshwater organisms. Ecotoxicology, 24, 1621-1631. [CrossRef]
- Schuler, M.S.; Relyea, R.A. (2018). A review of the combined threats of road salts and heavy metals to freshwater systems. BioScience, 68, 327-335. [CrossRef]
- Vilas–Boas, J.A.; Cardoso, S.J.; Senra, M.V.X.; Rico, A.; Dias, R.J.P. (2020). Ciliates as model organisms for the ecotoxicological risk assessment of heavy metals: a meta-analysis. Ecotoxicology and environmental safety, 199, 110669. [CrossRef]
- Fountain, M.T.; Hopkin, S.P. (2005). Folsomia candida (Collembola): a “standard” soil arthropod. Annuals Review of Entomology, 50, 201-222.
- Filser, J.; Wiegmann, S.; Schröder, B. (2014). Collembola in ecotoxicology—Any news or just boring routine? Applied Soil Ecology, 83, 193-199.
- Kammenga, J.E.; Dallinger, R.; Donker, M.H.; Köhler, H.; Simonsen, V.; Triebskorn, R.; Weeks, J.M. (2000) Biomarkers in terrestrial invertebrates for ecotoxicological soil risk assessment. Reviews of Environmental Contamination and Toxicology, 164, 93-147.
- Dallinger, R.; Berger, B.; Triebskorn-Köhler, R.; Köhler, H. (2001) Soil biology and ecotoxicology. In The biology of terrestrial molluscs; CABI Wallingford UK; pp. 489-525.
- Bharti, D.; Kumar, S.; La Terza, A. (2016) Rigidosticha italiensis (Ciliophora, Spirotricha), a novel large hypotrich ciliate from the soil of Lombardia, Italy. European Journal of Protistology, 56, 112-118. [CrossRef]
- Kumar, S.; Kamra, K.; Bharti, D.; La Terza, A.; Sehgal, N.; Warren, A.; Sapra, G.R. (2015) Morphology, morphogenesis, and molecular phylogeny of Sterkiella tetracirrata (Ciliophora, Oxytrichidae), from the Silent Valley National Park, India. European Journal of Protistology, 51,86-97.
- Warren, A.; Patterson, D.J.; Dunthorn, M.; Clamp, J.C.; et al. (2017) Beyond the “Code”: A Guide to the Description and Documentation of Biodiversity in Ciliated Protists (Alveolata, Ciliophora). The journal of eukaryotic microbiology, 64, 539-554.
- Fenchel, T. (2013) Ecology of Protozoa: The biology of free-living phagotropic protists; Springer-Verlag.
- Jiang, J.-G.; Wu, S.-G.; Shen, Y.-F. (2007). Effects of seasonal succession and water pollution on the protozoan community structure in an eutrophic lake. Chemosphere, 66, 523-532. [CrossRef]
- Weisse, T. (2017). Functional diversity of aquatic ciliates. European Journal of Protistology, 61, 331-358. [CrossRef]
- Geisen, S.; Mitchell, E.A.D.; Wilkinson, D.M.; Adl, S.; et al., (2017). Soil protistology rebooted: 30 fundamental questions to start with, Soil Biology and Biochemistry,111, 94-103. [CrossRef]
- Bonkowski, M.; Clarholm, M. (2015). Stimulation of plant growth through interactions of bacteria and protozoa: testing the auxiliary microbial loop hypothesis. Acta Protozool., 51, 237-247.
- Gualandris, D.; Rotondo, D.; Lorusso, C.; La Terza, A.; Calisi, A.; Dondero, F. (2024) The Metallothionein System in Tetrahymena thermophila Is Iron-Inducible. Toxics, 12, 725. [CrossRef]
- Fulgentini, L.; Passini, V.; Colombetti, G.; Miceli, C.; La Terza, A.; Marangoni, R. (2015). UV-radiation and visible light induce hsp70 genes expression in the Antarctic psycrophilic ciliate Euplotes focardii. Microbial Ecology, 70, 372-379. [CrossRef]
- La Terza, A.; Barchetta, S.; Buonanno, F.; Ballarini, P.; Miceli, C. (2008). The protozoan ciliate Tetrahymena thermophila as biosensor of sublethal levels of toxicants in the soil. Fresenius Environmental Bulletin, 17, 1144-1150.
- Bharti, D.; Kumar, S.; Basuri, C.K.; La Terza, A. (2024). Ciliated Protist Communities in Soil: Contrasting Patterns in Natural Sites and Arable Lands across Italy. Soil System, 8, 64. [CrossRef]
- Delmonte Corrado, M.; Trielli, F.; Amaroli, A.; Ognibene, M.; Falugi, C. (2005). Protists as tools for environmental biomonitoring: im-portance of cholinesterase enzyme activities. Water Poll. New Res., 1, 181-200.
- Gilron, G.L.; Lynn, D.H. (2018). Ciliated protozoa as test organisms in toxicity assessments. In Microscale testing in aquatic toxicology; CRC Press; pp. 323-336.
- Halliwell, B.; Gutteridge, J.M. (1986). Oxygen free radicals and iron in relation to biology and medicine: some problems and concepts. Arch. Biochem. Biophys, 246, 501-514. [CrossRef]
- Liu, C.-b.; Qu, G.-b.; Cao, M.-x.; Liang, Y.; Hu, L.-g.; Shi, J.-b.; Cai, Y.; Jiang, G.-b. (2017). Distinct toxicological characteristics and mechanisms of Hg2+ and MeHg in Tetrahymena under low concentration exposure. Aquatic Toxicology, 193, 152-159. [CrossRef]
- Varatharajan, G.R.; Calisi, A.; Kumar, S.; Bharti, D.; Dondero, F.; La Terza, A. (2024) Cytotoxicity and Antioxidant Defences in Euplotes aediculatus Exposed to Single and Binary Mixtures of Heavy Metals and Nanoparticles. Appl. Sci., 14, 5058. [CrossRef]
- Bharti, D.; Kumar, S.; La Terza, A. (2017). Description and molecular phylogeny of a novel hypotrich ciliate from the soil of Marche Region, Italy; including notes on the MOSYSS Project. Journal of Eukariotic Microbiology, 64,678-690. [CrossRef]
- Chapman-Andresen, C. (1958). Pinocytosis of inorganic salts by Amoeba proteus (Chaos diffluens). Comptes rendus des travaux du Laboratoire Carlsberg. Serie chimique, 31, 77.
- Strober, W. (2001). Trypan blue exclusion test of cell viability. Current protocols in immunology, A3. B. 1-A3. B. 3.
- Gallego, A., Martín-González, A., Ortega, R., Gutiérrez, J. C. (2007). Flow cytometry assessment of cytotoxicity and reactive oxygen species generation by single and binary mixtures of cadmium, zinc and copper on populations of the ciliated protozoan Tetrahymena thermophila. Chemosphere 68(64), 647-661. [CrossRef]
- Sprague, J. (1970). Measurement of pollutant toxicity to fish. II. Utilizing and applying bioassay results. Water Research, 4, 3-32. [CrossRef]
- Ravindran, C.; Varatharajan, G.R.; Rajasabapathy, R.; Vijayakanth, S.; Kumar, A.H.; Meena, R.M. (2012). A role for antioxidants in acclimation of marine derived pathogenic fungus (NIOCC 1) to salt stress. Microbial pathogenesis, 53, 168-179. [CrossRef]
- Vattem, D.A.; Shetty, K. (2002). Solid-state production of phenolic antioxidants from cranberry pomace by Rhizopus oligosporus. Food Biotechnology, 16, 189-210. [CrossRef]
- Yıldırım, A.; Mavi, A.; Kara, A.A. (2001). Determination of antioxidant and antimicrobial activities of Rumex crispus L. extracts. Journal of agricultural and food chemistry, 49, 4083-4089.
- Kunchandy, E.; Rao, M. (1990). Oxygen radical scavenging activity of curcumin. International journal of pharmaceutics, 58, 237-240. [CrossRef]
- Lorusso, C.; Calisi, A.; Sanchez-Hernandez, J.C.; Varodi, C.; Pogăcean, F.; Pruneanu, S.; Dondero, F. (2022). Carbon nanomaterial functionalization with pesticide-detoxifying carboxylesterase. Chemosphere, 309, 136594. [CrossRef]
- Kong, R.; Sun, Q.; Cheng, S.; Fu, J.; Liu, W.; Letcher, R.J.; Liu, C. (2021). Uptake, excretion and toxicity of titanate nanotubes in three stains of free-living ciliates of the genus Tetrahymena. Aquatic Toxicology, 233, 105790. [CrossRef]
- Díaz, S.; Martín-González, A.; Gutiérrez, J.C. (2006). Evaluation of heavy metal acute toxicity and bioaccumulation in soil ciliated protozoa. Environment international, 32, 711-717. [CrossRef]
- Luu, H.T.; Esteban, G.F.; Butt, A.A.; Green, I.D. (2022). Effects of Copper and the Insecticide Cypermethrin on a Soil Ciliate (Protozoa: Ciliophora) Community. Protist, 173, 125855. [CrossRef]
- Madoni, P.; Romeo, M.G. (2006). Acute toxicity of heavy metals towards freshwater ciliated protists. Environmental Pollution, 141(141), 141-147. [CrossRef]
- Martín-González, A., Díaz, S., Borniquel, S., Gallego, A., & Gutiérrez, J. C. (2006). Cytotoxicity and bioaccumulation of heavy metals by ciliated protozoa isolated from urban wastewater treatment plants. Research in microbiology, 157(152), 108-118. [CrossRef]
- Echavez, F.L.C.; Leal, J.C.M. (2021). Ecotoxicological effect of heavy metals in free-living ciliate protozoa of Lake Maracaibo, Venezuela. Journal of Water and Land Development, 102-116-102-116. [CrossRef]
- Marín-Leal, J.C.; Rincón-Miquilena, N.J.; Díaz-Borrego, L.C.; Pire-Sierra, M.C. (2022). Acute toxicity of potentially toxic elements on ciliated protozoa from Lake Maracaibo (Venezuela). Acta Limnologica Brasiliensia, 34, e21. [CrossRef]
- Fargašová, A. (2001). Winter third-to fourth-instar larvae of Chironomus plumosus as bioassay tools for assessment of acute toxicity of metals and their binary combinations. Ecotoxicology and Environmental Safety, 48, 1-5. [CrossRef]
- Ince, N.; Dirilgen, N.; Apikyan, I.; Tezcanli, G.; Üstün, B. (1999) Assessment of toxic interactions of heavy metals in binary mixtures: a statistical approach. Archives of Environmental Contamination and Toxicology, 36, 365-372. [CrossRef]
- Wang, J.; Zhang, M.; Xu, J.; Wang, Y. (1995) Reciprocal effect of Cu, Cd, Zn on a kind of marine alga. Water Research, 29, 209-214.
- Negilski, D.; Ahsanullah, M.; Mobley, M. (1981). Toxicity of zinc, cadmium and copper to the shrimp Callianassa australiensis. II. Effects of paired and triad combinations of metals. Marine Biology, 64, 305-309. [CrossRef]
- Otitoloju, A.A. (2002). Evaluation of the joint-action toxicity of binary mixtures of heavy metals against the mangrove periwinkle Tympanotonus fuscatus var radula (L.). Ecotoxicology and Environmental safety, 53, 404-415. [CrossRef]
- Mehta, R.; Templeton, D.M.; O’Brien, P.J. (2006). Mitochondrial involvement in genetically determined transition metal toxicity: II. Copper toxicity. Chemico-biological interactions, 163, 77-85. [CrossRef]
- Norwood, W.; Borgmann, U.; Dixon, D.; Wallace, A. (2003) Effects of metal mixtures on aquatic biota: a review of observations and methods. Human and Ecological Risk Assessment, 9, 795-811. [CrossRef]
- Hajdú, Z.; Hohmann, J.; Forgo, P.; Martinek, T.; Dervarics, M.; Zupkó, I.; Falkay, G.; Cossuta, D.; Máthé, I. (2007). Diterpenoids and flavonoids from the fruits of Vitex agnus-castus and antioxidant activity of the fruit extracts and their constituents. Phytotherapy Research, 21, 391-394. [CrossRef]
- Yıldırım, A.; Mavi, A.; Oktay, M.; Kara, A.A.; Algur, Ö.F.; Bilaloǧlu, V. (2000). Comparison of antioxidant and antimicrobial activities of Tilia (Tilia argentea Desf ex DC), sage (Salvia triloba L.), and Black tea (Camellia sinensis) extracts. Journal of Agricultural and Food Chemistry, 48, 5030-5034.
- Uddin, G.; Rauf, A.; Arfan, M.; Ali, M.; Qaisar, M.; Saadiq, M.; Atif, M. (2012). Preliminary phytochemical screening and antioxidant activity of Bergenia caliata. Middle-East J. Sci. Res, 11, 1140-1142.
- Horta, A.; Pinteus, S.; Alves, C.; Fino, N.; Silva, J.; Fernandez, S.; Rodrigues, A.; Pedrosa, R. (2014). Antioxidant and antimicrobial potential of the Bifurcaria bifurcata epiphytic bacteria. Marine drugs, 12, 1676-1689. [CrossRef]
- Halliwell, B. (1991) Reactive oxygen species in living systems: source, biochemistry, and role in human disease. The American journal of medicine, 91, S14-S22.
- Sharma, P.; Ravikumar, G.; Kalaiselvi, M.; Gomathi, D.; Uma, C. (2013) In vitro antibacterial and free radical scavenging activity of green hull of Juglans regia. Journal of Pharmaceutical Analysis, 3, 298-302. [CrossRef]
- Boldrin, F.; Santovito, G.; Gaertig, J.; Wloga, D.; Cassidy-Hanley, D.; Clark, T.G.; Piccinni, E. (2006). Metallothionein gene from Tetrahymena thermophila with a copper-inducible-repressible promoter. Eukaryotic cell, 5, 422-425. [CrossRef]
- Santovito, G.; Formigari, A.; Boldrin, F.; Piccinni, E. (2007). Molecular and functional evolution of Tetrahymena metallothioneins: New insights into the gene family of Tetrahymena thermophila. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 144, 391-397. [CrossRef]
- Ferro, D.; Bakiu, R.; De Pittà, C.; Boldrin, F.; Cattalini, F.; Pucciarelli, S.; Miceli, C.; Santovito, G. (2015). Cu, Zn superoxide dismutases from Tetrahymena thermophila: molecular evolution and gene expression of the first line of antioxidant defenses. Protist, 166, 131-145. [CrossRef]
Figure 1.
Death rate percentages (each point represents a mean value ± standard deviation) obtained from populations of R. tetracirrata, after 24 hrs exposure at different heavy metal concentrations (mg l-1). (a) Cd, (b) Cu and (c) Zn treatments.
Figure 1.
Death rate percentages (each point represents a mean value ± standard deviation) obtained from populations of R. tetracirrata, after 24 hrs exposure at different heavy metal concentrations (mg l-1). (a) Cd, (b) Cu and (c) Zn treatments.
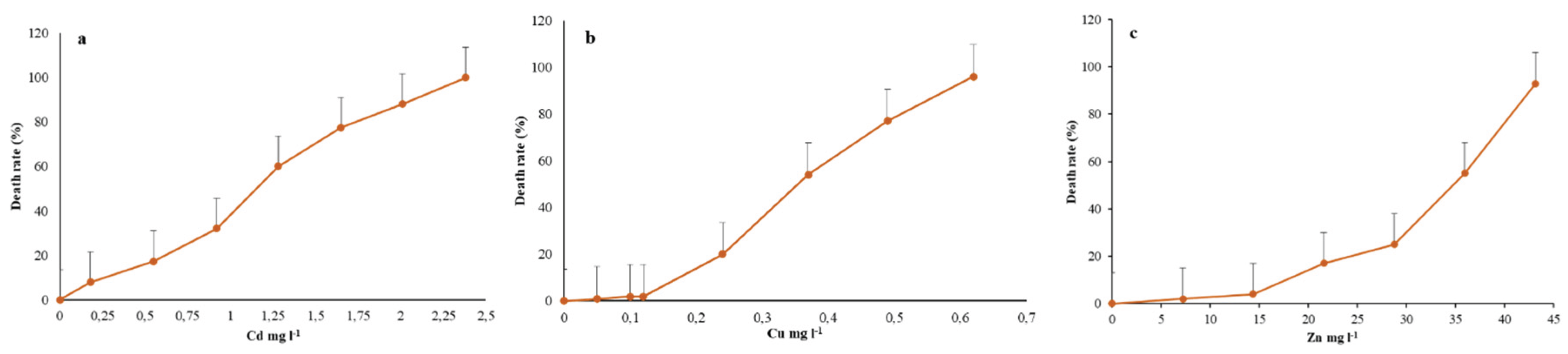
Figure 2.
Total phenolic content measured in the R. tetracirrata cell extracts (a) the exposure to the single metal (Cd, Cu, and Zn) concentrations corresponding to the LC20 and LC50 values (b) the exposure to the bimetallic mixture concentrations (0.25 + 0.25, 0.5 + 0.25, 0.75 + 0.25, 0.5 + 0.5, 0.25 + 0.5, 1 + 0.25, 0.75 + 0.5, and 0.25 + 1 TUs).
Figure 2.
Total phenolic content measured in the R. tetracirrata cell extracts (a) the exposure to the single metal (Cd, Cu, and Zn) concentrations corresponding to the LC20 and LC50 values (b) the exposure to the bimetallic mixture concentrations (0.25 + 0.25, 0.5 + 0.25, 0.75 + 0.25, 0.5 + 0.5, 0.25 + 0.5, 1 + 0.25, 0.75 + 0.5, and 0.25 + 1 TUs).
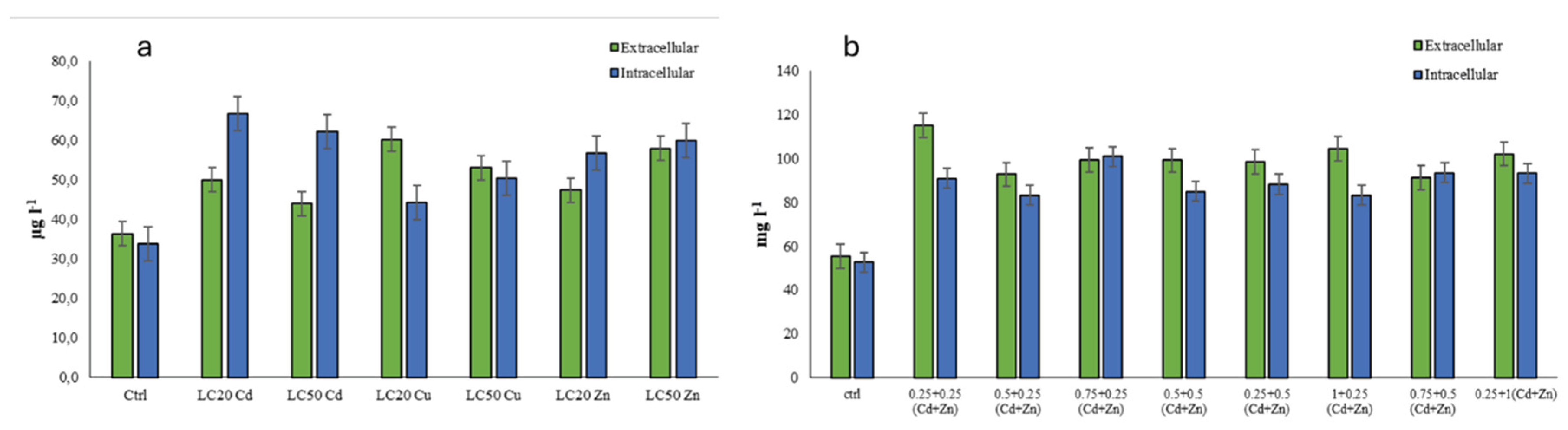
Figure 3.
DPPH scavenging activities measured in the R. tetracirrata cell extracts (a) the exposure to the single metal (Cd, Cu, and Zn) concentrations corresponding to the LC20 and LC50 values (b) the exposure to the bimetallic mixture concentrations (0.25 + 0.25, 0.5 + 0.25, 0.75 + 0.25, 0.5 + 0.5, 0.25 + 0.5, 1 + 0.25, 0.75 + 0.5, and 0.25 + 1 TUs) .
Figure 3.
DPPH scavenging activities measured in the R. tetracirrata cell extracts (a) the exposure to the single metal (Cd, Cu, and Zn) concentrations corresponding to the LC20 and LC50 values (b) the exposure to the bimetallic mixture concentrations (0.25 + 0.25, 0.5 + 0.25, 0.75 + 0.25, 0.5 + 0.5, 0.25 + 0.5, 1 + 0.25, 0.75 + 0.5, and 0.25 + 1 TUs) .
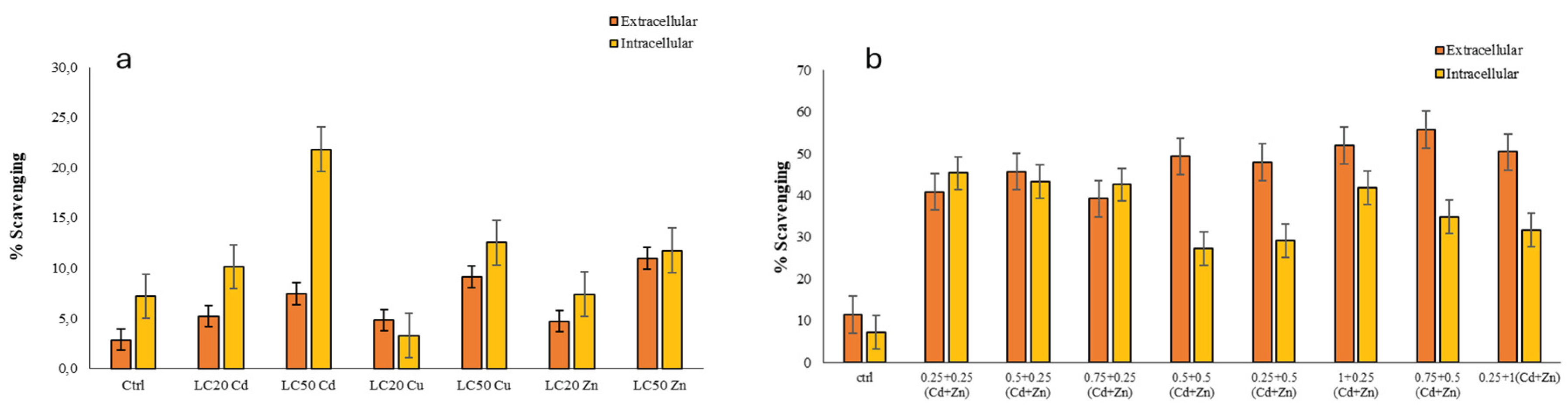
Figure 4.
Hydroxyl radical scavenging activities measured in the R. tetracirrata cell extracts (a) the exposure to the single metal (Cd, Cu, and Zn) concentrations corresponding to the LC20 and LC50 values (b) the exposure to the bimetallic mixture concentrations (0.25 + 0.25, 0.5 + 0.25, 0.75 + 0.25, 0.5 + 0.5, 0.25 + 0.5, 1 + 0.25, 0.75 + 0.5, and 0.25 + 1 TUs).
Figure 4.
Hydroxyl radical scavenging activities measured in the R. tetracirrata cell extracts (a) the exposure to the single metal (Cd, Cu, and Zn) concentrations corresponding to the LC20 and LC50 values (b) the exposure to the bimetallic mixture concentrations (0.25 + 0.25, 0.5 + 0.25, 0.75 + 0.25, 0.5 + 0.5, 0.25 + 0.5, 1 + 0.25, 0.75 + 0.5, and 0.25 + 1 TUs).
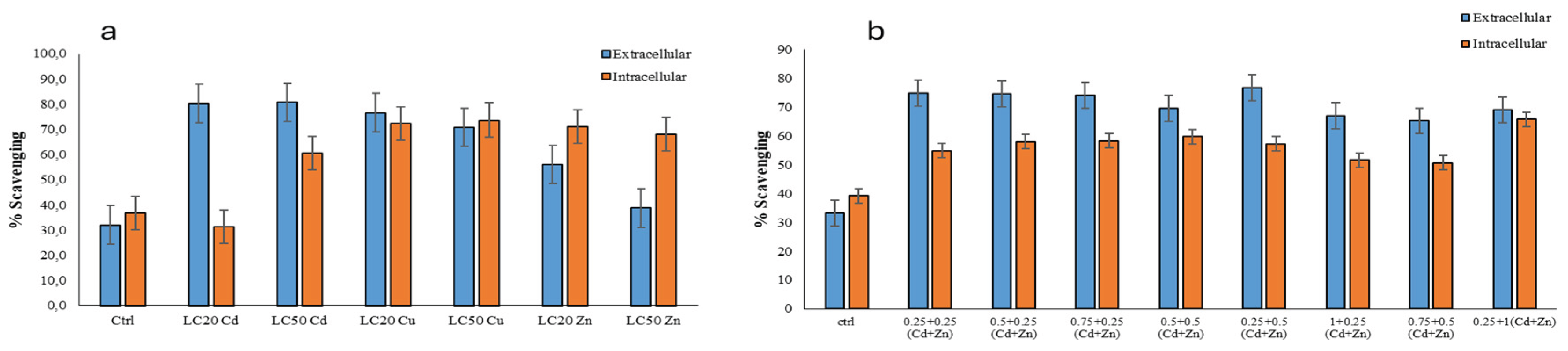

Table 1.
LC20 and LC50 values of R. tetracirrata for Cd, Cu and Zn.
| S. No. | Heavy Metals | LC20 (mg l-1) | LC50 (mg l-1) | R2 |
|---|---|---|---|---|
| 1. | Cd | 0.53 | 1.16 | 0.978 |
| 2 | Cu | 0.22 | 0.37 | 0.990 |
| 3. | Zn | 23.0 | 32.7 | 0.919 |
Table 2.
Observed and predicted cytotoxicity (% mortality) of R. tetracirrata to different mixtures of Cd + Zn after 24 hrs exposition.
Table 2.
Observed and predicted cytotoxicity (% mortality) of R. tetracirrata to different mixtures of Cd + Zn after 24 hrs exposition.
| Cd + Zn Total TUa |
Concentrations (TU)a for each metal Cd Zn |
Obtained Cytotoxicityb | Expected Cytotoxicity | Interaction type | |||
|---|---|---|---|---|---|---|---|
| 0.5 | 0.5 | 0 | 12.77 | ± | 2.54 | 18 | Not significant different |
| 0.25 | 0.25 | 6.07 | ± | 2.54 | 13 | Antagonism | |
| 0 | 0.5 | 3.87 | ± | 0.98 | 4 | Not significant different | |
| 0.75 | 0.75 | 0 | 29.40 | ± | 3.48 | 30 | Not significant different |
| 0.5 | 0.25 | 4.97 | ± | 1.65 | 20 | Antagonism | |
| 0.25 | 0.5 | 18.83 | ± | 2.54 | 15 | Not significant different | |
| 0 | 0.75 | 16.63 | ± | 1.65 | 18 | Not significant different | |
| 1 | 1 | 0 | 49.97 | ± | 1.65 | 50 | Not significant different |
| 0.75 | 0.25 | 22.20 | ± | 2.55 | 32 | Antagonism | |
| 0.5 | 0.5 | 3.87 | ± | 0.98 | 22 | Antagonism | |
| 0.25 | 0.75 | 8.87 | ± | 0.98 | 29 | Antagonism | |
| 0 | 1 | 49.43 | ± | 0.98 | 50 | Not significant different | |
| 1.25 | 1.25 | 0 | 61.07 | ± | 2.54 | 68 | Not significant different |
| 1 | 0.25 | 33.30 | ± | 1.70 | 52 | Antagonism | |
| 0.75 | 0.5 | 45.53 | ± | 3.87 | 34 | Synergism | |
| 0.5 | 0.75 | 32.77 | ± | 2.54 | 36 | Not significant different | |
| 0.25 | 1 | 14.97 | ± | 1.65 | 61 | Antagonism | |
| 0 | 1.25 | 77.77 | ± | 2.54 | 82 | Not significant different | |
Table 3.
Observed and predicted cytotoxicity (% mortality) of R. tetracirrata to different mixtures of Cu + Zn after 24 hrs exposition.
Table 3.
Observed and predicted cytotoxicity (% mortality) of R. tetracirrata to different mixtures of Cu + Zn after 24 hrs exposition.
| Cu + Zn Total TUa |
Concentrations (TU)a for each metal Cu Zn |
Obtained Cytotoxicityb | Expected Cytotoxicity | Interaction type | |||
|---|---|---|---|---|---|---|---|
| 0.5 | 0.5 | 0 | 12.17 | ± | 0.98 | 11 | Not significant different |
| 0.25 | 0.25 | 4.43 | ± | 0.98 | 4 | Not significant different | |
| 0 | 0.5 | 6.07 | ± | 2.54 | 4 | Not significant different | |
| 0.75 | 0.75 | 0 | 28.30 | ± | 1.70 | 30 | Not significant different |
| 0.5 | 0.25 | 6.07 | ± | 2.54 | 13 | Antagonism | |
| 0.25 | 0.5 | 9.97 | ± | 1.65 | 6 | Synergism | |
| 0 | 0.75 | 20.50 | ± | 1.91 | 18 | Not significant different | |
| 1 | 1 | 0 | 48.83 | ± | 2.54 | 50 | Not significant different |
| 0.75 | 0.25 | 34.93 | ± | 2.89 | 32 | Not significant different | |
| 0.5 | 0.5 | 11.63 | ± | 3.35 | 15 | Not significant different | |
| 0.25 | 0.75 | 23.87 | ± | 0.98 | 20 | Synergism | |
| 0 | 1 | 52.20 | ± | 1.91 | 50 | Not significant different | |
| 1.25 | 1.25 | 0 | 69.40 | ± | 2.55 | 71 | Not significant different |
| 1 | 0.25 | 31.63 | ± | 1.65 | 52 | Antagonism | |
| 0.75 | 0.5 | 28.87 | ± | 4.18 | 34 | Not significant different | |
| 0.5 | 0.75 | 32.77 | ± | 2.54 | 29 | Not significant different | |
| 0.25 | 1 | 72.77 | ± | 2.54 | 52 | Synergism | |
| 0 | 1.25 | 80.53 | ± | 2.54 | 82 | Not significant different | |
Table 4.
Observed and predicted cytotoxicity (% mortality) of R. tetracirrata to different mixtures of Cd + Cu after 24 hrs exposition.
Table 4.
Observed and predicted cytotoxicity (% mortality) of R. tetracirrata to different mixtures of Cd + Cu after 24 hrs exposition.
| Cd + Cu Total TUa |
Concentrations (TU)a for each metal Cd Cu |
Obtained Cytotoxicityb | Expected Cytotoxicity | Interaction type | |||
|---|---|---|---|---|---|---|---|
| 0.5 | 0.5 | 0 | 16.10 | ± | 1.91 | 18 | Not significant different |
| 0.25 | 0.25 | 10.53 | ± | 2.54 | 13 | Not significant different | |
| 0 | 0.5 | 13.87 | ± | 0.98 | 11 | Not significant different | |
| 0.75 | 0.75 | 0 | 28.87 | ± | 3.44 | 30 | Not significant different |
| 0.5 | 0.25 | 13.87 | ± | 7.86 | 20 | Antagonism | |
| 0.25 | 0.5 | 20.53 | ± | 0.92 | 22 | Not significant different | |
| 0 | 0.75 | 32.77 | ± | 2.54 | 30 | Not significant different | |
| 1 | 1 | 0 | 49.40 | ± | 3.48 | 50 | Not significant different |
| 0.75 | 0.25 | 34.97 | ± | 1.65 | 32 | Synergism | |
| 0.5 | 0.5 | 38.83 | ± | 2.54 | 29 | Synergism | |
| 0.25 | 0.75 | 44.40 | ± | 2.55 | 41 | Synergism | |
| 0 | 1 | 48.83 | ± | 2.54 | 50 | Not significant different | |
| 1.25 | 1.25 | 0 | 72.20 | ± | 2.55 | 68 | Not significant different |
| 1 | 0.25 | 55.50 | ± | 3.48 | 52 | Not significant different | |
| 0.75 | 0.5 | 34.40 | ± | 3.81 | 41 | Antagonism | |
| 0.5 | 0.75 | 45.53 | ± | 2.54 | 48 | Not significant different | |
| 0.25 | 1 | 84.97 | ± | 1.65 | 61 | Synergism | |
| 0 | 1.25 | 73.87 | ± | 1.96 | 71 | Not significant different | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



