
Submitted:
28 July 2025
Posted:
30 July 2025
You are already at the latest version
Abstract
This study uncovered a significant correlation between interferon-gamma (IFN-γ) secretion from immune cells and bone quality alterations in pancreatic cancer-bearing humanized-BLT (hu-BLT) mice. Reduced bone formation and IFN‐γ inhibition were observed in tumor mice versus those without tumors. When AJ2 probiotic bacteria were fed to MP2 pancreatic tumor-bearing mice, it increased bone integrity compared to tumor-bearing mice not fed with AJ2, correlating with increased BV/TV, Tb/Th, Tb/n, and decreased Tb/Sp. Most significantly, pancreatic tumor-bearing hu-BLT mice treated with a combination of supercharged NK cells (sNK) injections and AJ2 supplementation exhibited elevated levels of bone formation and volume, surpassing untreated or solely AJ2-fed tumor mice. Combination of sNK cell infusion and AJ2 feeding further augmented the levels of Tb/Th and Tb/n and decreased the level of Tb/Sp. We also have observed a great correlation between data obtained by micro-CT-generated 3D images, bone thickness and morphology, and secretion of IFN‐γ from various tissues of hu-BLT mice. Increased IFN‐γ secretion was evident in cell cultures of isolated single cells of pancreas, spleen, PBMCs, splenocyte-derived NK cells, and bone marrow of mice receiving combined treatment of sNK cells and AJ2 compared to untreated tumor-bearing mice. TRAP staining data highlighted heightened osteoclastic activity and bone resorption in untreated tumor mice compared to those receiving sNK cell injections and AJ2 feeding or AJ2 feeding alone. This research unveils new insights into cancer-related bone quality changes in hu-BLT mice, emphasizing the potential of immune cell-derived IFN‐γ in counteracting tumor-induced bone loss and fostering bone formation.
Keywords:
supercharged NK cells
; probiotic bacteria
; pancreatic cancer
; humanized-BLT mice
; osteoclasts
; IFN‐γ
1. Introduction
For the past four decades, pancreatic cancer has remained the leading cause of cancer-related mortality worldwide, with limited advancement in the 5-year survival rate [1,2]. Pancreatic cancer commonly spreads to the liver, peritoneal cavity, lung, brain, and bone, with the first documented case of pancreatic cancer metastasizing to bone dating back to 1963 in Russian literature[3]. Patients with pancreatic cancer often suffer from severe nutritional deficiencies, muscle wasting, and bone mass loss, leading to cachexia-induced bone deterioration and an increased risk of bone fractures [4]. Studies have shown the dynamic interplay between NK and osteoclasts. NK cells were found to exhibit positive expression of RANKL and M-CSF; therefore, NK cells, when interacting with monocytes, could induce the generation of osteoclasts, as RANKL and M-CSF are two crucial factors required for osteoclast formation [1]. Interestingly, osteoclasts express ligands activating NK cell receptors and release cytokines, fostering NK cell activation and expansion. [2].
We have previously demonstrated that our innovative approach leverages osteoclasts and probiotic bacteria AJ2 to synergistically activate NK cells, enhancing IFN-γ release [3,4]. AJ2 comprises seven gram-positive strains. NK cells expanded using osteoclasts and AJ2 were named as supercharged NK (sNK) cells, as they exhibit superior anti-cancer efficacy compared to traditionally activated NK cells and other expansion methods [3,5,6,7,8,9,10,11,12]. The process of sNK generation involved co-culturing NK cells with osteoclasts (OCs) and seven strains of sonicated probiotic bacteria [6]. The interaction between surface markers of NK cell receptor ligands and secreted factors of OCs played a pivotal role in expanding and activating NK cells [6,13]. Treatment with selected probiotic bacteria strains significantly enhanced NK cell activation and cytokine secretion [6,14]. The combination of OCs and probiotics triggered signals promoting NK cell expansion and functional activation.
Our previous studies have shown that sNK cells exhibited substantial improvements in cell expansion, cytotoxicity, and cytokine secretion. These cells also demonstrated an extended lifespan and the ability to induce tumor differentiation both in vivo and in vitro. Notably, sNK cells displayed enhanced survival in the tumor microenvironment due to elevated levels of anti-apoptotic proteins and reduced levels of pro-apoptotic proteins, enabling them to resist tumor-induced cell death [15]. Moreover, sNK cells exhibited increased expression levels of cytotoxic-associated granules and Trail, contributing to their enhanced cytotoxic function [15]. sNK cells also displayed enhanced regulatory function, with a significant portion of the sNK cell population in an active cycling phase. The cells expressed higher levels of proliferation-associated genes and proteins, along with memory-associated genes [15]. Most importantly, sNK cells showed elevated expression of activating receptors such as CD16, CD56, Nkp30, Nkp44, Nkp46, NKG2D, CD94, CD54, KIR2, KIR3, and 4-1BBL, while demonstrating decreased expression of inhibitory receptors like NKG2A, PD-1, and TIGIT [6,15]. Our previously published work highlights the exceptional anti-cancer efficacy of sNK cells compared to various other NK cell treatments. In direct comparisons with IL-2, IL-2 + anti-CD16 mAbs, IL-2 + anti-CD16 mAbs + sAJ2, IL-12, IL-15, IL-18, OSCSCs, and K562 expanded NK cells, sNK cells demonstrate outstanding anti-cancer capabilities [6,9,16,17]. Furthermore, when compared with cord blood-derived NK cells, iPSC-derived NK cells, NK92, and several other NK cell types, sNK cells showcase significantly heightened levels of cytotoxicity and cytokine secretion [18,19]. Our in vivo studies highlighted the therapeutic potential of sNK cells alone or in conjunction with AJ2, in bolstering the immune function and reducing tumor burden. This intervention shows promise in combating pancreatic tumor cancer stem-like cells in hu-BLT mice, highlighting its immunotherapeutic impact [10].
When a pancreatic cancer patient orally consumed AJ2, increased levels of IFN-γ and enhanced cytotoxic potential were seen in the patient’s peripheral blood NK cells [20]. FN-γ, primarily released by NK cells, CD4+ T cells, and CD8+ T cells, was found to enhance the immune response against pathogens and tumors, inhibiting osteoclastogenesis [21,22,23]. To date, limited knowledge is available on in vivo effects of IFN-γ on bone tissue [24,25]. IFN-γ's role in regulating RANKL signaling and bone destruction has been noted, as it binds to osteoclasts, degrading RANKL signaling and inhibiting bone resorption [39]. To explore the impact of IFN-γ on bone health in cancer patients, we used the therapeutic approach of sNK cells and probiotic bacteria AJ2 in pancreatic tumor-bearing hu-BLT mice. Using a humanized pancreatic tumor bone marrow/liver/thymus (hu-BLT) mice model infused with sNK cells and fed with AJ2, we aim to investigate therapeutic effects on bone remodeling. This research sheds light on the potential of IFN-γ in maintaining bone quality in cancer patients.
The modulation of bone turnover during tumorigenesis and the impact of secreted IFN-γ on bone remodeling are the focus of this study. Findings of this study highlight the potential role of sNK cells in activating immune cells within pancreatic tumor-bearing hu-BLT mice. Data shown in the study indicate that treating pancreatic tumor hu-BLT mice with sNK cells, either alone or in conjunction with AJ2, leads to heightened IFN-γ levels across different tissue compartments. Interestingly, a strong correlation emerged between IFN-γ secretion and bone formation, emphasizing this cytokine's ability to impede tumor-induced bone loss and promote bone regeneration.
2. Materials and Methods
2.1. Ethics Approval Number and Name of the Committee
All procedures performed using human peripheral blood were approved by the UCLA Institutional Review Board (IRB#11-00781). Written informed consent was obtained by all human donor participants. Studies performed on hu-BLT mice were approved by the UCLA Animal Research Committee (ARC protocol # 2012-101-13).
2.2. Cell Lines and Reagents
DMEM (Life Technologies, CA, USA) media supplemented with 10% Fetal Bovine Serum (FBS) (Gemini Bio-Products, CA) was used for cell cultures of MiaPaCa-2 (MP2) tumor cells. Single cells isolated from hu-BLT mice tissues were cultured using RPMI 1640 (Life Technologies, CA, USA) supplemented with 10% FBS. Recombinant IL-2 (rhIL-2) was obtained from Peprotech (NJ, USA). To generate osteoclasts, monocytes were cultured using alpha-MEM (Life Technologies, CA, USA) supplemented with 10% FBS, M-CSF (Biolegend ,CA, USA), and RANKL (PeproTech,NJ, USA).
2.3. Probiotic Bacteria AJ2 Sonication
A patented combination of seven probiotic strains was weighed and resuspended at a concentration of 10 mg/ml using RPMI 1640 supplemented with 10% FBS. Five pulses of sonication were conducted, each pulse consisted of a thorough vortex of bacterial strains and incubation of sonicated samples on ice for 15 seconds at amplitudes of 6 to 8. Samples obtained after sonication were incubated on ice for 30 seconds. Post five pulses, samples were observed using a microscope to ensure minimal 80 percent lysis of bacteria cell walls. On average 20 rounds of sonication and ice incubation were performed to obtain complete sonication. Sonicated probiotic samples were aliquoted using RPMI 1640 media and were frozen at -80 degrees.
2.4. Purification of Human NK Cells and Monocytes
PBMCs were used to isolate NK cells and monocytes. Negative isolation of NK cells and monocytes was performed using the EasySep® Human NK cell enrichment kit and monocyte isolation kit, respectively. Cell isolation kits were purchased from Stem Cell Technologies, (Vancouver, BC, Canada). NK cells and monocytes expressing greater than 94% purity were used for experiments.
2.5. Generation of Osteoclasts and Supercharging of NK Cells
Monocytes purified from human peripheral blood were cultured using an alpha-MEM medium containing M-CSF (25 ng/mL) for the first 3 days. On day 3 onwards, monocytes were cultured using alpha-MEM medium containing M-CSF (25 ng/mL) and RANKL (25 ng/mL) for 18 days. Media was 70% replaced with fresh alpha-MEM medium containing M-CSF (25 ng/mL) and RANKL (25 ng/mL) every 3 days. To supercharge NK cells, freshly isolated NK cells were treated with a combination of rh-IL-2 (1000 U/mL) and anti-CD16 mAb (3 μg/mL) overnight and were co-cultured with osteoclasts in the presence of sonicated AJ2 (sAJ2). RMPI 1640 supplemented with rh-IL-2 (1500 U/ml) was used to refresh culture media every 3 days, and sNK cells were used for hu-BLT injection on day 15.
2.6. Tumor Implantation, sNK Cells Infusions, and AJ2 Fed in hu-BLT Mice
All of the procedures performed in hu-BLT mice were approved by the UCLA Animal Research Committee (ARC) in accordance with all federal, state, and local guidelines. Hu-BLT mice were generated using NSG mice purchased from Jackson Laboratory [26,27] (Figure 1A). For pancreatic tumor hu-BLT mice experiments, mice were anesthetized using a combination of isoflurane and oxygen, and 1 x 106 human MP2 pancreatic cancer stem-like cells suspended in 10 μl HC Matrigel were implanted directly in the pancreas. Post one to two weeks of pancreatic tumor implantation, 1 x 106 sNK cells were injected into mice via the tail vein. For the AJ2 group, oral AJ2 feeding was started one to two weeks prior to pancreatic tumor implantation and feeding was done every 48 hours until the mice were euthanized (Figure 3A). When the signs of morbidity were evident, the mice were euthanized followed by harvesting of pancreatic tumor, pancreas, bone marrow, spleen, and peripheral blood.
2.7. Cell Isolation and Cultures of hu-BLT Mice Tissue Samples
Single-cell suspensions of BM were obtained by cutting femurs at both ends, followed by flushing with RPMI 1640 media. Recovered BM cells were filtered using a 40 µm cell strainer. Single-cell suspensions of the spleen and liver were obtained by mincing the tissues, followed by filtering of samples using a 40 µm cell strainer. Recovered samples were centrifuged at 1500 rpm for 5 minutes at 4˚C, and pellets were re-suspended in ACK buffer for an average of 5 minutes to get rid of red blood cells. Final samples were re-suspended in RPMI media and centrifugation at 1500 rpm for 5 minutes at 4˚C. Peripheral blood was harvested in heparin, and Ficoll-Hypaque centrifugation was used to obtain single-cell isolation or PBMCs from peripheral blood, and samples were re-suspended in RPMI 1640 medium. Cell culture of each tissue was performed for 7 days by treating the samples with IL-2 (1000 U/ml) in RPMI 1640 medium.
2.8. Isolation of hu-BLT Mice Spleen NK Cells
Splenocytes of hu-BLT mice were used to isolate NK cells using a human CD56+CD3+ selection kit (Stem Cells Technologies, Vancouver, BC, Canada). Cell culture of NK cells was performed for 7 days by treating the samples with IL-2 (1000 U/ml) in RPMI 1640 medium.
Enzyme-linked immunosorbent assay (ELISA)
IFN-γ secretion analysis of hu-BLT tissue cultures was performed using a human ELISA kit (Biolegend, San Diego, CA, USA). Assays were conducted as recommended by the manufacturer.
2.9. Bone Analysis
Micro-computed tomography (micro-CT) was used to assess bone architecture. Harvested samples were fixed using formalin and images were taken at a resolution of 10 u pixels using high-resolution microCT (Skyscan 1275, Skyscan, Belgium). Images were analyzed using Data Viewer, Recon, CTAn and CTVol software provided by the manufacturer. For specimen preparation, the lumbar vertebrae (L3) were dissected from hu-BLT mice and were fixed using 70% ethanol. We have scanned vertebras using Skyscan 1275 (Bruker microCT N.V., Belgium), equipped with a 5-μm focal spot micro-focus x-ray tube at the resolution of 10μm (60 kVp, 166 mA, and 1mm Al Filter). Specimens were aligned with the vertical axis of the scanner, and low-density foam (a non-attenuating material) was used to stabilize the specimens firmly into a 0.25-diameter tube. Phantom calibration was performed to relate the micro-CT values to a mineral-equivalent value (g/cm3) of calcium hydroxyapatite. For imaging, scanned images were reconstructed with NRecon (Bruker microCT N.V., Belgium) for attenuation correction, ring artifact reduction, and beam hardening. After data acquisition, images were aligned in 3D view for vertical orientation with Data Viewer software for accuracy. Segmentation of the images was completed manually by comparing the binarized image with the unsegmented image, and a single global threshold of 60 was applied. An irregular ROI selection was manually drawn parallel and close to the endocortical surface. For ex vivo assessment of bone architecture by micro-CT analysis, the length of the ROI was adjusted in proportion to the total vertebral height. For the third lumbar vertebrae, transverse micro-CT slices were acquired for the entire vertebral body, and trabecular bone was evaluated within the region of 0.5 mm away from the growth plate. To ensure accuracy, each ROI was drawn manually in a sequential manner for each trans-axial micro-CT slice. Morphometric parameters were computed from the binarized images using a direct three-dimensional approach that does not rely on any prior assumptions about the underlying structure. For trabecular morphology, assessment of bone volume fraction (BV/TV %), trabecular thickness (Tb. Th, mm), trabecular number (Tb. N, mm), and trabecular separation (Tb.Sp, mm) were used. All analyses were performed with CTAn software (Bruker microCT N.V., Belgium). The 3D-rendered model of lumbar vertebrae was constructed by CTVol software (Bruker microCT N.V., Belgium). A comparison of the 3D-rendered volume was performed to show differences in the trabecular structure of the study groups.
2.10. Histology and Quantitative Histomorphometry
Static histomorphometry was carried out on hu-BLT mice. Third lumbar vertebras (L3) were dissected, fixed in 70% ethanol, dehydrated, and embedded undecalcified in methyl methacrylate. Frontal sections, 5 µm thick and stained with 0.1% toluidine blue, pH 6.4. The static parameters of bone formation (OB) and resorption (OC) were measured in a defined area between 0.25mm from both growth plates and endochondral bone surfaces. Additional histochemical stain tartrate-resistant acid phosphatase (TRAP) was performed to identify osteoclasts.
2.11. Statistical Analysis
Linear Mixed Effects Models were used to analyze micro-CT data to determine the differences between the comparisons (as differences in means between the groups). The linear model was used for the outcome on an interval scale (as opposed to a categorical scale) and a mixed-effects model was used to account for the correlation between the different outcomes within each mouse. For ex vivo data, for two group comparisons an unpaired two-tailed Student t-test and for multiple groups one-way ANOVA using Prism-10 software (Graphpad Prism, San Diego, CA, USA) were performed. “n” denotes the number of mice used for each group. The following symbols represent the levels of statistical significance within each analysis, *** (p-value <0.001), ** (p-value 0.001–0.01), * (p-value 0.01–0.05).
3. Results
3.1. Successful Reconstitution of Human CD45+ Immune Cells of hu-BLT Mice Tissues
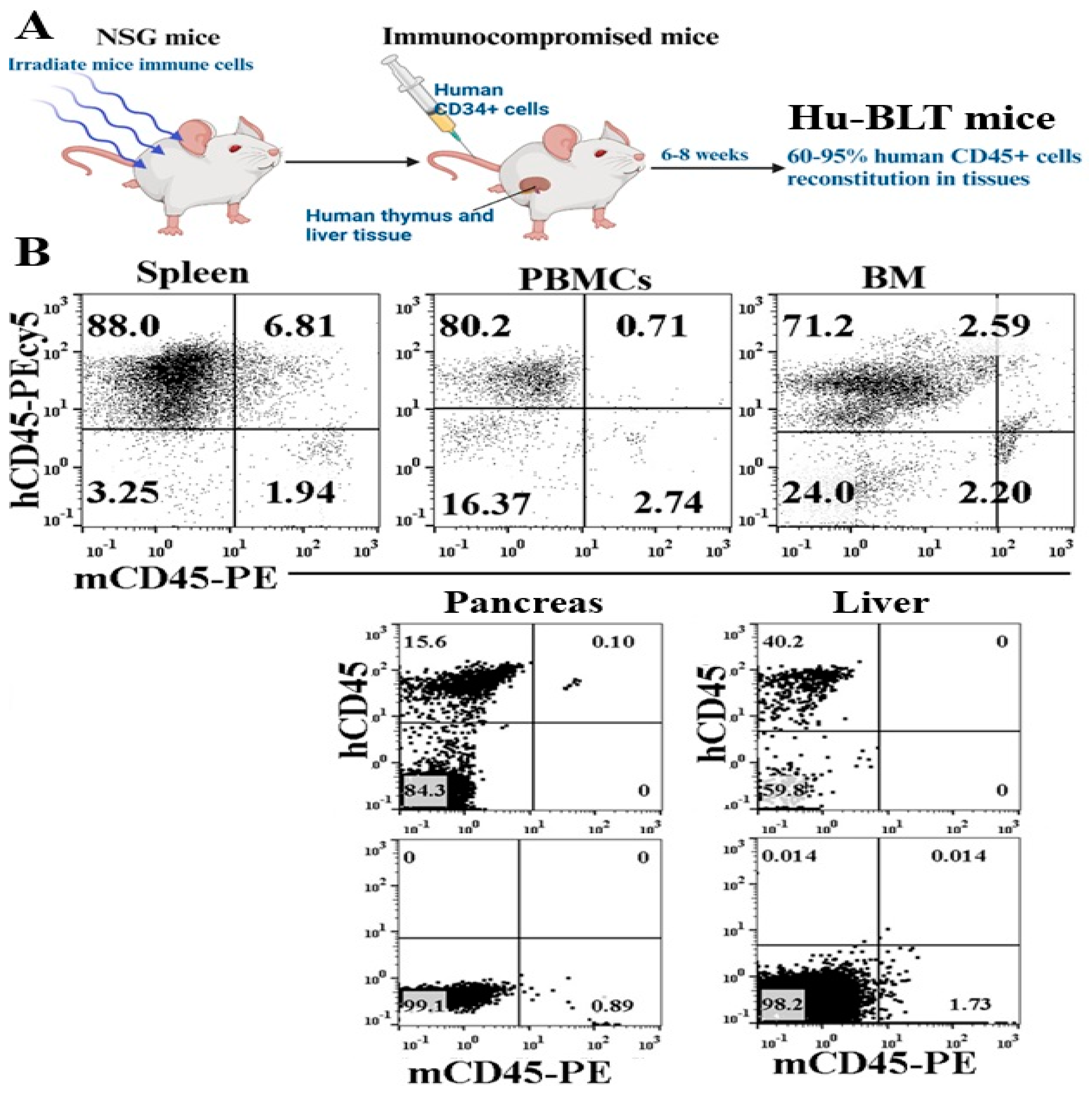
The generation procedure of hu-BLT mice, as illustrated in Figure 1A, involved detailed steps. To evaluate the reconstitution of human and mouse immune cells in hu-BLT mice, flow cytometric analysis was conducted on single cells extracted from various tissues. These tissues included the peripheral blood, spleen, BM cells, pancreas, and liver. The assessment focused on murine and hu-CD45 expression levels. Greater than 71% expression of human CD45 was observed in cells obtained from hu-BLT mice peripheral blood, spleen, and BM (Figure 1B). Human CD45+ immune cell expression was 15.6% in the pancreas and 40.2% in the liver of hu-BLT mice (Figure 1B). Across all tissues examined, murine CD45 levels ranged between 0.89% and 2.74%. These results reveal the successful reconstitution of human immune cells in hu-BLT mice across various tissues, with distinct levels of human CD45 expression observed.
Figure 1.
Reconstitution of human CD45+ immune cells in humanized-BLT (hu-BLT) mice tissues. Hu-BLT mice were generated as described in Materials and Methods and shown in Figure (A). Reconstitution of human immune cells was determined in hu-BLT mice spleen, PBMCs, BM, pancreas, and liver using anti-human and anti-mouse CD45 and analyzed by flow cytometry (B).
Figure 1.
Reconstitution of human CD45+ immune cells in humanized-BLT (hu-BLT) mice tissues. Hu-BLT mice were generated as described in Materials and Methods and shown in Figure (A). Reconstitution of human immune cells was determined in hu-BLT mice spleen, PBMCs, BM, pancreas, and liver using anti-human and anti-mouse CD45 and analyzed by flow cytometry (B).
3.2. Increased Bone Formation Was Seen in Probiotic Bacteria-Fed hu-BLT Mice Compared to the Control Group
To investigate the role of probiotic bacteria on bone health, hu-BLT mice were orally fed with 5 billion probiotic bacteria AJ2 every 48 hours for 4-5 weeks before mice were euthanized and bone analysis was performed (Figure S1A). leading to enhanced bone formation compared to control mice. Analysis showed increased bone volume and trabecular number in the fed group. While comparisons of bone volume proportion, trabecular bone thickness, trabecular number, and trabecular bone spacing did not yield statistically significant differences between the two groups, higher values of Tb.Th or Tb.N suggest elevated bone formation (Figure S1B). Additionally, a smaller Tb.Sp indicates denser bone structure. Remarkably, 3D images showcased greater trabecular bone formation in AJ2-fed mice, reinforcing the overall trend of increased bone development in this group (Figure S1C). Even though no statistical significance was seen, overall, this data indicates increased bone formation in AJ2-fed hu-BLT mice. These findings shed light on the potential benefits of AJ2 probiotic supplementation in promoting bone health.
3.3. Adoptively Transferred sNK Cells Circulate to Tissue Compartments of Pancreatic Tumor-Bearing Hu-BLT Mice
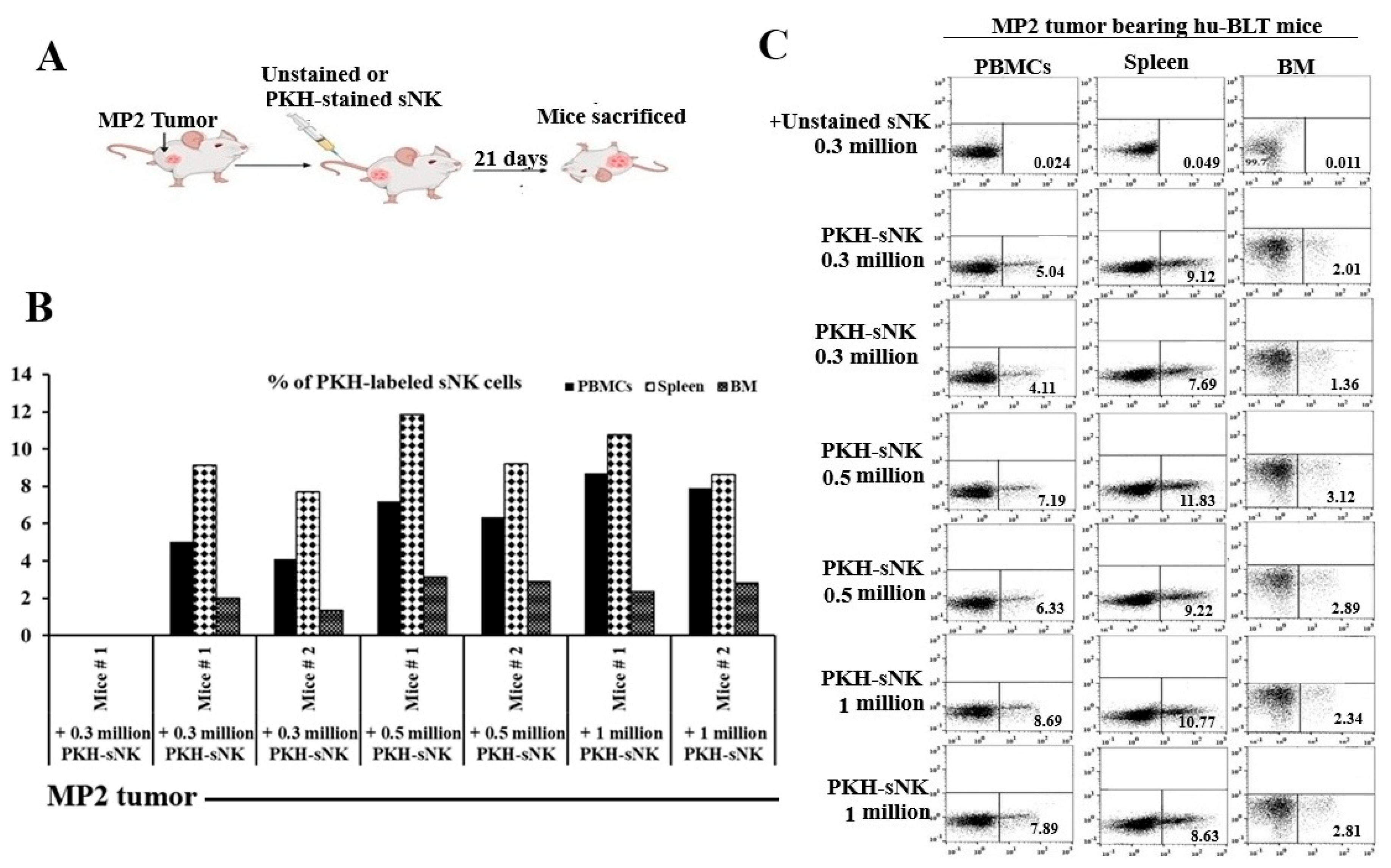
Our investigations also focused on tracking the biodistribution of adoptively transferred sNK cells within different tissue compartments of the pancreatic tumor-bearing hu-BLT mice. The experiment involved MP2 pancreatic cancer stem cells (CSCs) tumor implantation in the pancreas of mice, followed by infusions of PKH-labeled sNK cells as depicted in Figure 2A. We found that the infused sNK cells navigated through the peripheral blood, spleen, and bone marrow of the mice, with their presence and distribution levels varying based on the dosage administered (Figure 2B,C).
Figure 2.
sNK cells persist and circulate in vivo in pancreatic tumor-bearing hu-BLT mice. Hu-BLT mice were implanted with 1 x 106 MP2 in the pancreas, one week later the group 1 (n=1) was adoptively transferred unstained 0.3 x 106 sNK cells, group 2 (n=2) was transferred 0.3 x 106 PKH-sNK cells, group 3 (n=2) was transferred PKH-0.5 x 106 sNK cells, and group 4 (n=2) was transferred 1 x 106 PKH-sNK cells (A). sNK cells labeled with the fluorochrome PKH-26 were monitored by flow cytometry on day 21 in peripheral blood-derived PBMCs, spleen, and BM (B, C).
Figure 2.
sNK cells persist and circulate in vivo in pancreatic tumor-bearing hu-BLT mice. Hu-BLT mice were implanted with 1 x 106 MP2 in the pancreas, one week later the group 1 (n=1) was adoptively transferred unstained 0.3 x 106 sNK cells, group 2 (n=2) was transferred 0.3 x 106 PKH-sNK cells, group 3 (n=2) was transferred PKH-0.5 x 106 sNK cells, and group 4 (n=2) was transferred 1 x 106 PKH-sNK cells (A). sNK cells labeled with the fluorochrome PKH-26 were monitored by flow cytometry on day 21 in peripheral blood-derived PBMCs, spleen, and BM (B, C).
3.4. A Remarkable Increase in IFN-γ Secretion Was Seen in Mice Tissues When Mice Were Infused with sNK Cells and Fed with Probiotic Bacteria
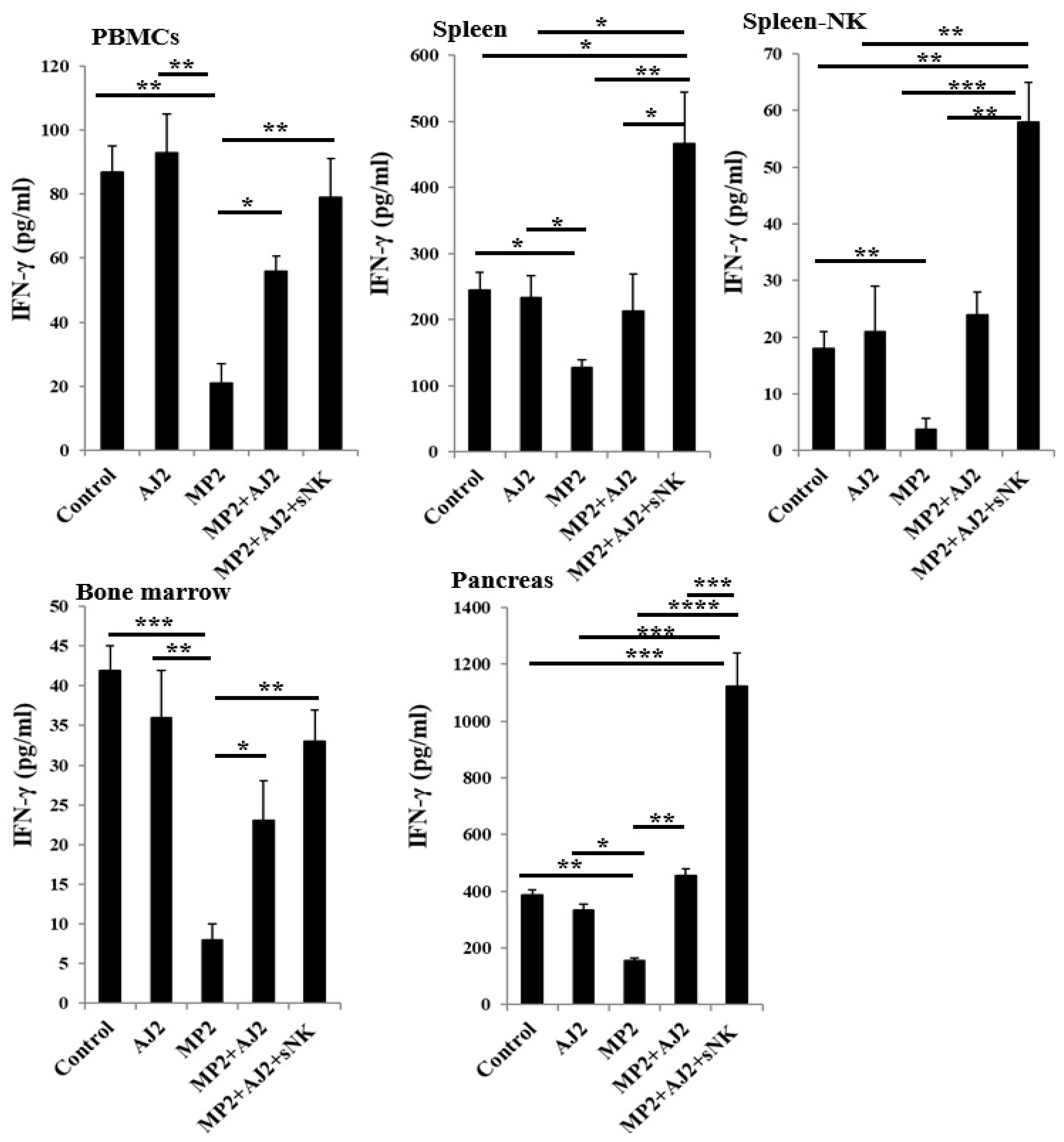
When immune cells from various tissue compartments of hu-BLT mice were activated with IL-2 for seven days, the induced IFN-γ secretion was examined. The findings indicated a significant decrease in IFN-γ levels in MP2 tumor-bearing mice compared to the control group (Figure 3 and Figure S2). Conversely, a similar IFN-γ secretion level was observed in both the control group and hu-BLT mice fed only AJ2 (Figure 3). Remarkably, administering AJ2 to MP2 tumor-bearing mice led to the restoration of IFN-γ levels. Furthermore, the infusion of sNK cells alongside AJ2 feeding demonstrated a notable recovery in IFN-γ secretion in peripheral blood-derived PBMCs and bone marrow. This approach also resulted in elevated IFN-γ levels in peripheral blood-derived serum, spleen, spleen-derived NK cells, and pancreas (Figure 3 and Figure S2).
Figure 3.
Combination of sNK cells and AJ2 increased IFN-γ secretion by PBMCs, splenocytes, and bone marrow-derived immune cells in hu-BLT mice. Hu-BLT mice were orthotopically injected with MP2 tumors, injected with sNK cells, and fed with AJ2 as described in Figure 3A. At the end of the experiment, hu-BLT mice were sacrificed, and the spleens, peripheral blood, and bone marrow were harvested, and single-cell suspensions were prepared. NK cells were isolated from splenocytes. Peripheral blood-derived PBMCs, splenocytes, spleen-derived NK cells, and bone marrow-derived cells were cultured in the presence of IL-2 (1000 U/ml) for 7 days. On day 7, the supernatants were harvested, and the secretion of IFN-γ was determined using a single ELISA. A representative experiment is shown as a bar graph and data are presented as Mean±SD. ****(p value <0.0001), ***(p value 0.0001-0.001), **(p value 0.001-0.01), *(p value 0.01-0.05).
Figure 3.
Combination of sNK cells and AJ2 increased IFN-γ secretion by PBMCs, splenocytes, and bone marrow-derived immune cells in hu-BLT mice. Hu-BLT mice were orthotopically injected with MP2 tumors, injected with sNK cells, and fed with AJ2 as described in Figure 3A. At the end of the experiment, hu-BLT mice were sacrificed, and the spleens, peripheral blood, and bone marrow were harvested, and single-cell suspensions were prepared. NK cells were isolated from splenocytes. Peripheral blood-derived PBMCs, splenocytes, spleen-derived NK cells, and bone marrow-derived cells were cultured in the presence of IL-2 (1000 U/ml) for 7 days. On day 7, the supernatants were harvested, and the secretion of IFN-γ was determined using a single ELISA. A representative experiment is shown as a bar graph and data are presented as Mean±SD. ****(p value <0.0001), ***(p value 0.0001-0.001), **(p value 0.001-0.01), *(p value 0.01-0.05).
3.5. A Significant Reduction in Tumor Weight Was Seen When Pancreatic Tumor-Bearing hu-BLT Mice Were Infused with sNK Cells and Fed with Probiotic Bacteria
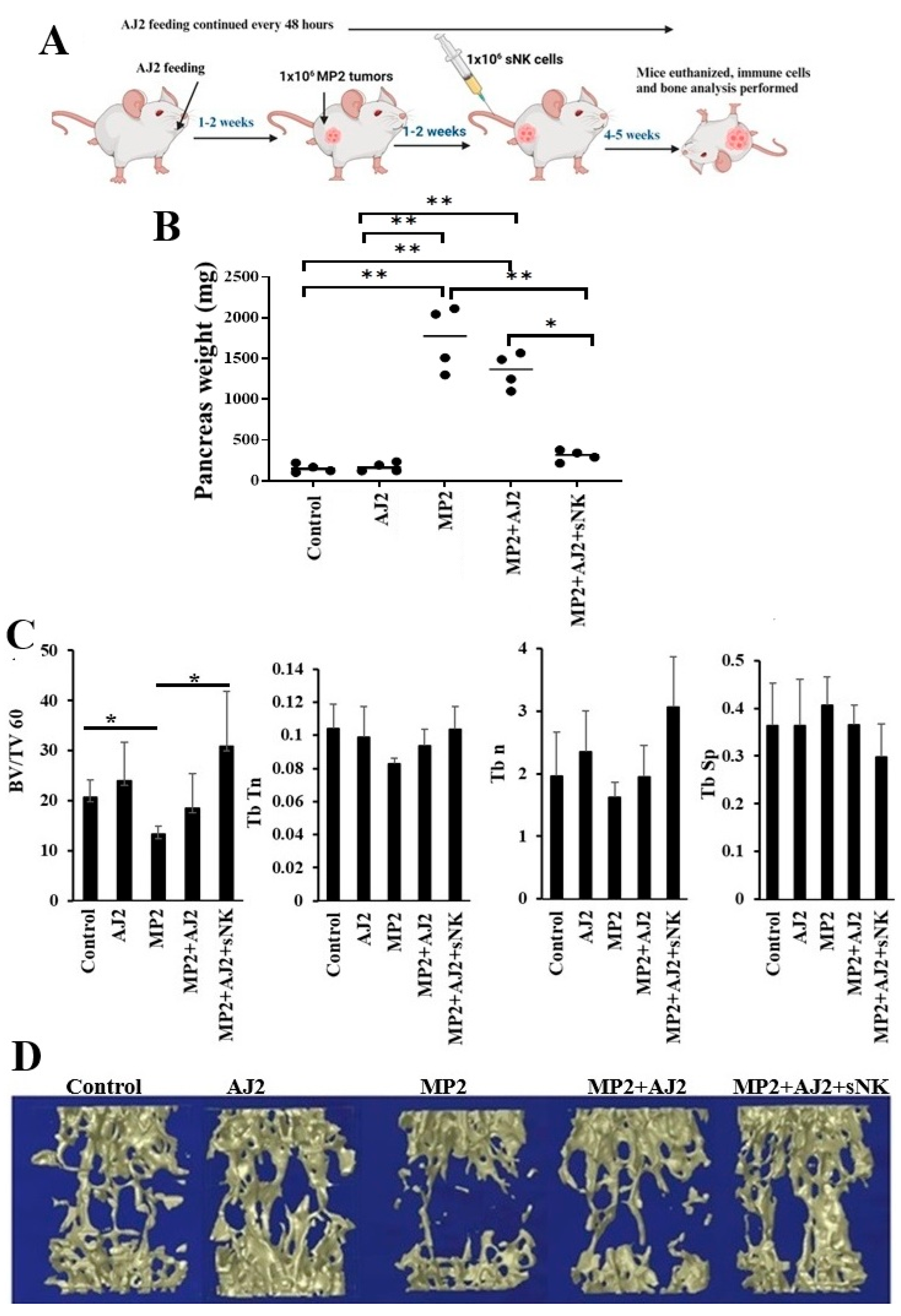
The efficacy of sNK cells to inhibit tumor growth and to restore immune cell function in tumor-bearing hu-BLT mice has been previously published [28,29,30,31,32,33]. In this study, MP2 pancreatic CSCs tumors were implanted in the pancreas of hu-BLT mice, followed by sNK cell injections, and feeding of probiotic bacteria AJ2, as depicted in Figure 4A. Upon sacrifice, the pancreas weight was measured, showing no difference in mice fed with AJ2 alone without tumors (Figure 4B). However, mice with MP2 tumors exhibited increased pancreatic weight due to the tumor growth. Interestingly, tumor weight slightly decreased with AJ2 feeding, but significantly decreased when combined with sNK cell treatment (Figure 4B).
3.6. An Increase in Bone Formation Was Seen When Pancreatic Tumor-Bearing hu-BLT Mice Were Infused with sNK Cells and Fed with Probiotic Bacteria
MP2 pancreatic tumors led to a notable decrease in bone volume fraction (BV/TV), trabecular thickness (Tb.Th), trabecular number (Tb.n), and an increase in trabecular spacing (Tb.Sp) compared to the control group (Figure 4C). Conversely, both healthy and tumor-bearing mice fed with AJ2 showed improvements in BV/TV, Tb.Th, Tb.n, and Tb.Sp. Furthermore, mice injected with sNK cells and fed with AJ2 exhibited enhanced BV/TV, Tb.Th, Tb.n, and decreased Tb.Sp compared to AJ2-fed mice (Figure 4B, Figures S3A and S4A). The findings were reinforced by 3D images, highlighting the impact of the interventions on the pancreatic tumor environment (Figure 4C, Figures S3B and S4B). The data on IFN-γ secretion aligned with bone analysis results, showing reduced bone formation levels and decreased IFN-γ secretion in the corresponding compartments of MP2 tumor-bearing mice (Figure 3 and Figure 4B,C). Overall, the introduction of AJ2 feeding or a combination of sNK cells and AJ2 feeding not only reinstated IFN-γ secretion but also effectively hindered tumor-induced bone loss in the experimental model (Figure 3 and Figure 4B,C).
Figure 4.
MP2 tumor-bearing mice injected with sNK cells and fed with AJ2, presented statistically significantly higher trabecular bone volume when compared to the MP2 tumor and MP2+AJ2 group, respectively. Hu-BLT mice were orthotopically injected with 1 x 106 human MP2 tumors in the pancreas. One week after the tumor implantation, mice received supercharged NK (sNK) cells via tail-vein injection. Mice were orally fed with 5 billion AJ2 every 48 hours (n=5) (A). Bone analysis is performed as described in the Materials and Methods section (n=5) (B-C). Tb.Th, or trabecular bone thickness, is a parameter used to measure the microarchitecture of trabecular bone. It's one of several parameters used in histomorphometric studies, along with Tb.N, The number of trabeculae per unit length, Tb.Sp, The average distance between trabecular bones, BV/TV, The proportion of bone volume in the total tissue volume. A higher value of Tb.Th or Tb.N indicates a higher level of bone formation. A narrower gap between trabecular bones is indicated by a smaller Tb.Sp, indicates higher bone density. **(p value 0.001-0.01), *(p value 0.01-0.05).
Figure 4.
MP2 tumor-bearing mice injected with sNK cells and fed with AJ2, presented statistically significantly higher trabecular bone volume when compared to the MP2 tumor and MP2+AJ2 group, respectively. Hu-BLT mice were orthotopically injected with 1 x 106 human MP2 tumors in the pancreas. One week after the tumor implantation, mice received supercharged NK (sNK) cells via tail-vein injection. Mice were orally fed with 5 billion AJ2 every 48 hours (n=5) (A). Bone analysis is performed as described in the Materials and Methods section (n=5) (B-C). Tb.Th, or trabecular bone thickness, is a parameter used to measure the microarchitecture of trabecular bone. It's one of several parameters used in histomorphometric studies, along with Tb.N, The number of trabeculae per unit length, Tb.Sp, The average distance between trabecular bones, BV/TV, The proportion of bone volume in the total tissue volume. A higher value of Tb.Th or Tb.N indicates a higher level of bone formation. A narrower gap between trabecular bones is indicated by a smaller Tb.Sp, indicates higher bone density. **(p value 0.001-0.01), *(p value 0.01-0.05).
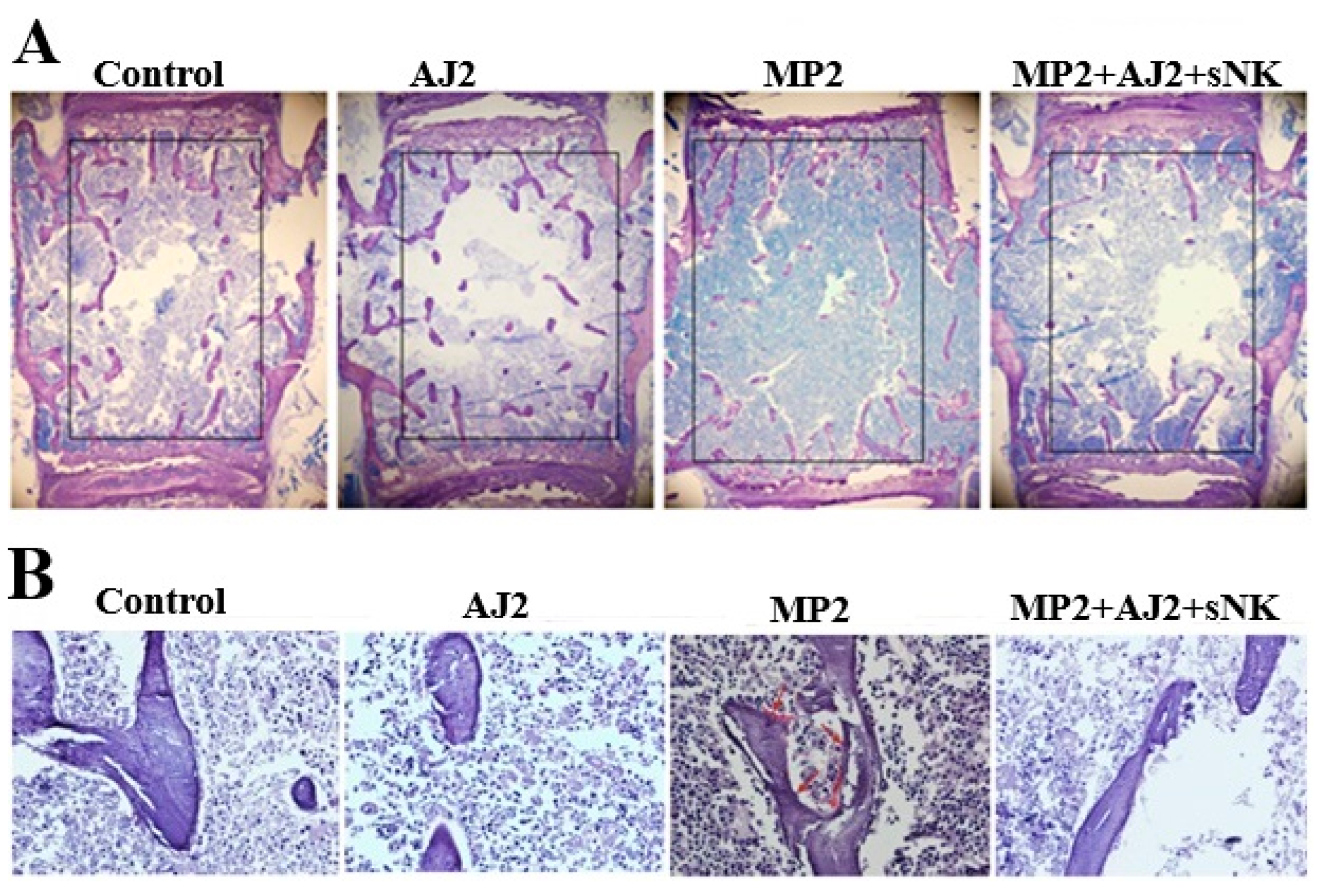
3.7. Increased Trabecular Bone Formation Was Observed When Pancreatic Tumor-Bearing Mice Were Fed Probiotic Bacteria Alone or Combined with sNK Cells Infusion
To further understand the underlying cellular process of bone remodeling, we performed the histological analysis and static indices assessment at the third lumbar vertebra. In parallel to micro-CT findings and IFN-γ secretion, immunohistochemistry assay showed increased bone formation in AJ2 fed mice in contrast to control mice (Figure 5A). Also, increased bone formation was observed in MP2 tumor-bearing mice treated with a combination of sNK cells injection alongside AJ2 consumption, as opposed to untreated MP2 tumor-bearing mice (Figure 5A). Next to micro-CT observations and IFN-γ secretion analyses, our exploration extended to assessing static parameters of bone resorption, particularly osteoclast activity, through TRAP staining. Remarkably, TRAP-positive outcomes were evident in samples from MP2 tumor-bearing subjects, while samples from the control group, AJ2-fed mice, and MP2-implanted mice treated with sNK cells and AJ2 diet exhibited TRAP-negative results (Figure 5B). Overall, these findings revealed higher osteoclastic activity and bone resorption in the presence of tumors compared to the control group or tumor-bearing mice that received treatment (Figure 5B).
4. Discussion
In our previous studies, we have discovered that sNK cells play a crucial role in inhibiting pancreatic tumor growth and metastasis through tumor lysis and differentiation mechanisms. Pancreatic tumor samples collected from the pancreatic tumor-bearing hu-BLT mice infused with sNK cells alongside AJ2 feedings expressed a rise in IFN-γ secretion and a decline in IL-6 secretion [10]. In pancreatic tumor hu-BLT mice, treatment with sNK cells, with or without AJ2-fed, resulted in increased secretion levels of IFN-γ in the serum (Figure S1) [33]. This effect was consistent with previous findings, showing elevated IFN-γ secretion in pancreas cell cultures of pancreatic tumor-bearing hu-BLT mice treated with sNK cells and AJ2 feeding compared to untreated tumor-bearing and control mice [10] (Figure 3). Furthermore, the restoration of immune cell cytotoxic function in various tissue compartments of pancreatic tumor-bearing hu-BLT mice treated with sNK cells was observed [10]. In the current study, we have observed a strong correlation between the 3D images generated by micro-CT analysis, bone morphology, and the levels of IFN-γ secretion.
The overall effect of decreased IFN-γ secretion in the MP2 alone implanted hu-BLT group was a significant reduction in bone volume, suggesting that the deficiency in IFN-γ may result in decreased levels of bone formation. The deficiency in IFN-γ was associated with a notable decrease in bone volume, indicating a potential link to reduced bone formation. This connection was further supported by findings showing reduced trabecular bone volume as observed through 3D micro-CT analysis, histology, and histomorphometry. When assessing the bone parameters, we have discovered a strong link between bone integrity loss in 3D microCT analysis of the tumor alone implanted mouse group and notable changes in BV/TV, Tb/Th, Tb/n, and Tb/Sp. These alterations were reversed when tumor-bearing mice were infused with sNK cells and fed with AJ2. These findings highlight the crucial role of NK cells in maintaining bone integrity during pathological conditions. Moreover, the bacterial strains in AJ2 were specifically chosen to enhance NK cell function and boost their IFN-γ production levels, indicating a promising avenue for treating osteoporosis and bone loss [34]. Data shown in this study highlighted the role of IFN-γ in modulating osteoblastic and osteoclastic activities. Lower levels of IFN-γ appeared to hinder osteoblastic function and potentially enhance osteoclastic activity, ultimately leading to a significant reduction in bone volume. Furthermore, IFN-γ's direct anti-tumor effects were noted to counteract tumor-induced bone loss, emphasizing its dual role in bone health and tumor suppression. Moreover, this research illustrated a positive correlation between immune cells' IFN-γ secretion and enhanced bone formation, as evidenced by micro-CT analysis. Introducing AJ2 feeding in both control and tumor-bearing mice groups resulted in increased bone formation compared to their respective counterparts, underlining the potential of IFN-γ in promoting bone health.
Additionally, observations from TRAP staining indicated heightened osteoclast activity and bone resorption in tumor-bearing mice, emphasizing the detrimental impact of tumors on bone health. However, increased IFN-γ levels appeared to mitigate tumor-induced bone loss, highlighting the therapeutic potential of IFN-γ in combating cancer-related bone complications. The data obtained in this study emphasize the intricate relationship between the immune and skeletal systems, emphasizing the crucial role of IFN-γ in maintaining skeletal integrity. The study suggests that targeting the IFN-γ signaling pathway could offer promising avenues for enhancing bone strength and combating bone-related disorders.
5. Conclusion
Overall, this study emphasizes the crucial role of sNK cells and AJ2 in impeding tumor growth and metastasis. It reveals that AJ2 fed alone or in combination with sNK cell infusions triggers heightened IFN-γ secretion in hu-BLT mice, effectively curbing tumor-induced bone deterioration. The report suggests the therapeutic potential of sNK cells, prompting further investigation into innovative remedies for individuals with osteolytic cancers. It underscores the necessity for a comprehensive comprehension of the mechanisms governing tumor progression within bone, advocating for a deeper exploration of the bone marrow microenvironment to address cancer-related bone impairments effectively
Author Contributions
KK and AJ were the co-principal investigators, obtained the funding, and wrote the manuscript. KK, PR, and JC performed experiments. KK analyzed all the results, prepared figures, and wrote the manuscript.
Data Availability Statement
The data that supports the findings of this study are available within the article and the supplementary file.
Acknowledgments
The authors are grateful to funding agencies and donors for supporting the work.
Conflict of Interest Statement
The authors declare that this research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
| NK cells | Natural killer cells |
| sNK cells | Supercharged NK cells |
| CSCs | Cancer-stem-like-cells |
| Hu-BLT | Humanized-bone marrow/liver/thymus |
| IFN-γ | Interferon-gamma |
| IL-2 | Interleukin 2 |
| OCs | Osteoclasts |
| PBMCs | Peripheral blood-derived mononuclear cells |
| MP2 | MiaPaCa-2 |
| ELISAs | Enzyme-Linked Immunosorbent Assays |
References
- Soderstrom K, et al. Natural killer cells trigger osteoclastogenesis and bone destruction in arthritis. Proceedings of the National Academy of Sciences of the United States of America2010. p. 107(29): p. 13028-33.
- Tseng HC, Kanayama K, Kaur K, Park SH, Park S, Kozlowska A, et al. Bisphosphonate-induced differential modulation of immune cell function in gingiva and bone marrow in vivo: role in osteoclast-mediated NK cell activation. Oncotarget. 2015;6(24):20002-25.
- Ryschich EN, T.; Hinz, U.; Autschbach, F.; Ferguson, J.; Simon, I.; Weitz, J.; Fröhlich, B.; Klar, E.; Büchler, M.W.; et al. . Control of T-Cell-mediated immune response by HLA class I in human pancreatic carcinoma. Clin. Cancer Res. 2005, 11, 498–504.
- Pandha HR, A.; John, J.; Lemoine, N. . Loss of expression of antigen-presenting molecules in human pancreatic cancer and pancreatic cancer cell lines. Clin. Exp. Immunol. 2007, 148, 127–135.
- Bui VTT, H.C.; Kozlowska, A.; Maung, P.O.; Kaur, K.; Topchyan, P.; Jewett, A. . Augmented IFNgamma and TNF-alpha Induced by Probiotic Bacteria in NK Cells Mediate Differentiation of Stem-Like Tumors Leading to Inhibition of Tumor Growth and Reduction in Inflammatory Cytokine Release; Regulation by IL-10. Front. Immunol. 2015, 6, 576.
- Kaur K, Cook J, Park SH, Topchyan P, Kozlowska A, Ohanian N, et al. Novel Strategy to Expand Super-Charged NK Cells with Significant Potential to Lyse and Differentiate Cancer Stem Cells: Differences in NK Expansion and Function between Healthy and Cancer Patients. Front Immunol. 2017;8:297. [CrossRef]
- Kaur K, Jewett A. Supercharged NK Cell-Based Immuotherapy in Humanized Bone Marrow Liver and Thymus (Hu-BLT) Mice Model of Oral, Pancreatic, Glioblastoma, Hepatic, Melanoma and Ovarian Cancers. Crit Rev Immunol. 2023;43(2):13-25. [CrossRef]
- Chiang J, Chen PC, Pham J, Nguyen CQ, Kaur K, Raman SS, et al. Characterizing hepatocellular carcinoma stem markers and their corresponding susceptibility to NK-cell based immunotherapy. Front Immunol. 2023;14:1284669. [CrossRef]
- Kaur K, Chen PC, Ko MW, Mei A, Senjor E, Malarkannan S, et al. Sequential therapy with supercharged NK cells with either chemotherapy drug cisplatin or anti-PD-1 antibody decreases the tumor size and significantly enhances the NK function in Hu-BLT mice. Front Immunol. 2023;14:1132807. [CrossRef]
- Kaur K, Kozlowska AK, Topchyan P, Ko MW, Ohanian N, Chiang J, et al. Probiotic-Treated Super-Charged NK Cells Efficiently Clear Poorly Differentiated Pancreatic Tumors in Hu-BLT Mice. Cancers (Basel). 2019;12(1). [CrossRef]
- Kaur K, Safaie T, Ko MW, Wang Y, Jewett A. ADCC against MICA/B Is Mediated against Differentiated Oral and Pancreatic and Not Stem-Like/Poorly Differentiated Tumors by the NK Cells; Loss in Cancer Patients due to Down-Modulation of CD16 Receptor. Cancers (Basel). 2021;13(2). [CrossRef]
- Kaur KC, J.; Park, S.H.; Topchyan, P.; Kozlowska, A.; Ohanian, N.; Fang, C.; Nishimura, I.; Jewett, A. Novel Strategy to Expand Super-Charged NK Cells with Significant Potential to Lyse and Differentiate Cancer Stem Cells: Differences in NK Expansion and Function between Healthy and Cancer Patients. Front. Immunol. 2017, 8, 297.
- Li H, Hong S, Qian J, Zheng Y, Yang J, Yi Q. Cross talk between the bone and immune systems: osteoclasts function as antigen-presenting cells and activate CD4+ and CD8+ T cells. Blood. 2010;116(2):210-7. [CrossRef]
- Dong H, Rowland I, Yaqoob P. Comparative effects of six probiotic strains on immune function in vitro. Br J Nutr. 2012;108(3):459-70. [CrossRef]
- Ko MW, Mei A, Senjor E, Nanut MP, Gao LW, Wong P, et al. Osteoclast-expanded supercharged NK cells perform superior antitumour effector functions. BMJ Oncol. 2025;4(1):e000676. [CrossRef]
- Romee R, Rosario M, Berrien-Elliott MM, Wagner JA, Jewell BA, Schappe T, et al. Cytokine-induced memory-like natural killer cells exhibit enhanced responses against myeloid leukemia. Sci Transl Med. 2016;8(357):357ra123. [CrossRef]
- Terrén I, Orrantia A, Astarloa-Pando G, Amarilla-Irusta A, Zenarruzabeitia O, Borrego F. Cytokine-Induced Memory-Like NK Cells: From the Basics to Clinical Applications. Front Immunol. 2022;13:884648. [CrossRef]
- Kaur K, Safaie T, Ko M-W, Wang Y, Jewett A. ADCC against MICA/B Is Mediated against Differentiated Oral and Pancreatic and Not Stem-Like/Poorly Differentiated Tumors by the NK Cells; Loss in Cancer Patients due to Down-Modulation of CD16 Receptor. Cancers. 2021;13(2):239. [CrossRef]
- Huerta-Yepez S, Chen PC, Kaur K, Jain Y, Singh T, Esedebe F, et al. Supercharged NK cells, unlike primary activated NK cells, effectively target ovarian cancer cells irrespective of MHC-class I expression. BMJ Oncol. 2025;4(1):e000618. [CrossRef]
- Ko M-W, Kaur K, Safaei T, Chen W, Sutanto C, Wong P, et al. Defective Patient NK Function Is Reversed by AJ2 Probiotic Bacteria or Addition of Allogeneic Healthy Monocytes. Cells. 2022;11(4):697. [CrossRef]
- Takayanagi H, et al. T-cell-mediated regulation of osteoclastogenesis by signalling cross-talk between RANKL and IFN-gamma. nature2000. p. 408(6812): p. 600-5. [CrossRef]
- Takayanagi H, J. Mechanistic insight into osteoclast differentiation in osteoimmunology. . Mol Med (Berl)2005. p. 83(3): p. 170-9. [CrossRef]
- Delves PJaIMR. The immune system. Second of two parts. . N Engl J Med; 2000. p. 343(2): p. 108-17.
- Gao Y, et al. IFN-gamma stimulates osteoclast formation and bone loss in vivo via antigen-driven T cell activation. The Journal of clinical investigation2007. p. 117(1): p. 22-32. [CrossRef]
- Cenci S, et al. Estrogen deficiency induces bone loss by increasing T cell proliferation and lifespan through IFN-gamma-induced class II transactivator. Proceedings of the National Academy of Sciences of the United States of America2003. p. 100(18): p. 10405-10. [CrossRef]
- Shimizu S, Hong P, Arumugam B, Pokomo L, Boyer J, Koizumi N, et al. A highly efficient short hairpin RNA potently down-regulates CCR5 expression in systemic lymphoid organs in the hu-BLT mouse model. Blood. 2010;115(8):1534-44. [CrossRef]
- Vatakis DN, Koya RC, Nixon CC, Wei L, Kim SG, Avancena P, et al. Antitumor activity from antigen-specific CD8 T cells generated in vivo from genetically engineered human hematopoietic stem cells. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(51):E1408-16. [CrossRef]
- Chennamadhavuni A, Iyengar V, Mukkamalla SKR, Shimanovsky A. Leukemia. StatPearls. Treasure Island (FL) with ineligible companies. Disclosure: Varun Iyengar declares no relevant financial relationships with ineligible companies. Disclosure: Shiva Kumar Mukkamalla declares no relevant financial relationships with ineligible companies. Disclosure: Alex Shimanovsky declares no relevant financial relationships with ineligible companies.: StatPearls Publishing Copyright © 2025, StatPearls Publishing LLC.; 2025.
- Kaur K, Chen P-C, Ko M-W, Mei A, Senjor E, Malarkannan S, et al. Sequential therapy with supercharged NK cells with either chemotherapy drug cisplatin or anti-PD-1 antibody decreases the tumor size and significantly enhances the NK function in Hu-BLT mice. Frontiers in Immunology. 2023;Volume 14 - 2023. [CrossRef]
- Kaur K, Jewett A. Supercharged NK Cell-Based Immuotherapy in Humanized Bone Marrow Liver and Thymus (Hu-BLT) Mice Model of Oral, Pancreatic, Glioblastoma, Hepatic, Melanoma and Ovarian Cancers. 2023;43(2):13-25. [CrossRef]
- Kaur K, Ko M-W, Ohanian N, Cook J, Jewett A. Osteoclast-expanded super-charged NK-cells preferentially select and expand CD8+ T cells. Scientific Reports. 2020;10(1):20363. [CrossRef]
- Kaur K, Paytsar T, Karolina KA, Nick O, Jessica C, Ou MP, et al. Super-charged NK cells inhibit growth and progression of stem-like/poorly differentiated oral tumors in vivo in humanized BLT mice; effect on tumor differentiation and response to chemotherapeutic drugs. OncoImmunology. 2018;7(5):e1426518.
- Kaur K, Kozlowska AK, Topchyan P, Ko M-W, Ohanian N, Chiang J, et al. Probiotic-Treated Super-Charged NK Cells Efficiently Clear Poorly Differentiated Pancreatic Tumors in Hu-BLT Mice. Cancers. 2020;12(1):63. [CrossRef]
- Bui VT, Tseng HC, Kozlowska A, Maung PO, Kaur K, Topchyan P, et al. Augmented IFN-γ and TNF-α Induced by Probiotic Bacteria in NK Cells Mediate Differentiation of Stem-Like Tumors Leading to Inhibition of Tumor Growth and Reduction in Inflammatory Cytokine Release; Regulation by IL-10. Front Immunol. 2015;6:576. [CrossRef]
Figure 5.
The AJ2-fed group exhibited increased bone formation when compared to the control and tumor-bearing groups. Hu-BLT mice were orthotopically injected with MP2 tumors, injected with sNK cells, and fed with AJ2 as described in Figure 3A. Histology and quantitative histomorphometry were performed as described in the Materials and Methods section (n=5) (A-B).
Figure 5.
The AJ2-fed group exhibited increased bone formation when compared to the control and tumor-bearing groups. Hu-BLT mice were orthotopically injected with MP2 tumors, injected with sNK cells, and fed with AJ2 as described in Figure 3A. Histology and quantitative histomorphometry were performed as described in the Materials and Methods section (n=5) (A-B).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



