
Submitted:
29 July 2025
Posted:
30 July 2025
You are already at the latest version
Abstract
Escherichia ruysiae is a recently characterized species within the Escherichia genus, often misclassified as E. coli due to limitations in existing operating procedure and diagnostic databases. In this study, we report the first isolation and genomic characterization of E. ruysiae in Togo, from a five-year-old female patient who was hospitalized with gastro-enteritis and bloody diarrhea and subsequently died after eight days. Biochemical tests and MALDI-TOF initially identified the microorganism as E. coli, but phylogenomic and Average Nucleotide Identity (ANI) analysis confirmed it to be E. ruysiae, Clade IV with enteroaggregative associated genes. Whole genome sequencing of the strain FK53-34 enables the identification of resistance genes including blaPBP, blaampH, acrA, emrF, baeS et tolC and various aminoglycoside-modifying enzymes, blaEC-15, eptA and pmrF. The virulence profile of the strain included but not limited to, aap, aatABC, senB, prmF genes which support its pathogenicity and virulence. Multilocus sequence typing (MLST) did not match any known sequence type, which is obvious for a newly characterized mi-croorganism. This study highlights the critical need for enhanced diagnostic tools and surveillance systems to identify emerging pathogens, including Escherichia ruysiae
Keywords:
Escherichia ruysiae
; WGS
; blaEC-15
; senB
; aatABC
; eptA
; pediatric gastroenteritis
; Clade IV Escherichia
; Togo
1. Introduction
The Escherichia genus is one of the most extensively studied bacterial groups in microbiology, largely due to its most prominent species, Escherichia coli. It has served as a model organism for bacterial physiology, genetics, and host–microbe interactions for decades. Importantly, this genus encompasses both commensal and pathogenic members that colonize the gastrointestinal tract of humans and animals [1,2,3,4]. Within the E. coli species itself, a remarkable diversity exists ranging from harmless gut symbionts to strains responsible for severe gastrointestinal and extraintestinal infections, such as enterotoxigenic E. coli (ETEC), enterohemorrhagic E. coli (EHEC), enteroinvasive E. coli (EIEC), enteropathogenic E. coli (EPEC), enteroaggregative E. coli (EAEC), and diffusely adherent E. coli (DAEC). Additionally, uropathogenic E. coli (UPEC) are a major cause of urinary tract infections, and meningitis-associated E. coli (MNEC) cause sepsis and meningitis. This duality makes the Escherichia genus especially compelling from both medical, veterinary and ecological standpoints.
Six species have been described so far, including Escherichia coli, Escherichia hermannii, Escherichia fergusonii, Escherichia albertii, Escherichia marmotae, Escherichia whittamii, and recently E. ruysiae [3,4,5,6,7,8,9,10]. All known Escherichia species are ubiquitous and potentially associated with animal [6] and human disease [5,6,7,8,9,10]. E. ruysiae was first isolated from the faecal sample of an international traveller, and all so far known to belong to two phylogroups (Cryptic Clade III and Clade IV) [9]. Several studies have shown that the phenotypic identification of E. ruysiae is confused with E. coli. This is because there is no standardized protocol in clinical microbiology to phenotypically differentiate E. ruysiae from the other members of the genus. Like other species of the genus Escherichia, E. ruysiae shows a genomic and proteomic signature very close to that of E. coli, but with certain differences that can be subtle and difficult to distinguish without a well-maintained database (for example, the Bruker Biotyper database, VITEK MS, or others) [6]. E ruysiae was spotted in a prevalence study of Escherichia spp., with zoonotic significance in urban crows in Japan, posing a significant risk for their potential transmission to humans [11]. The two cryptic clades described so far, Clades III and IV, are commonly found in animals and the environment. Studies have investigated the ubiquity of E. ruysiae and commensalism in the human intestinal tract [6]. Recently, E. ruysiae was isolated in 2024 from wild boars in Italy, therefore, its prevalence in livestock needs to be further investigated [12]. Also, antimicrobial resistance in E. ruysiae has been shown to be associated with conjugative plasmids carrying clinically relevant antibiotic resistance genes, and it is thought to serve as a reservoir of resistance genes globally [13]. To date, the pathogenicity of E. ruysiae is not known and hasn’t been demonstrated in any clinical or veterinary cases. Also, this bacterium is generally overlooked in clinical and veterinary diagnostics globally because it lacks a standardized differential phenotypic assay in microbiology to delineate it from E. coli. In addition, its spectrum is still not in any of the MALDITOF databases [14]. This makes the majority of E. ruysiae identified as E. coli with a comfortable score.
In Togo, the microbiological diagnosis is usually limited to phenotypic identification assays after the growth and isolation of the microorganism in pure colonies. Using these conventional techniques, a case of gastroenteritis was reported to be caused by Escherichia coli from a bloody stool sample collected from a five-year-old child hospitalized in a district medical center in the maritime region of Togo. For the first time, we performed a thorough genomic description of the strain and demonstrated that this was E. ruysiae instead of E. coli. In this study, we aim to describe the phenotypic and genomic characteristics of this strain and highlight specific genomic characteristics associated with the virulence and antimicrobial resistance.
2. Materials and Methods
2.1. Sample Collection
Two swabs were taken from the stool sample collected in clean containers, equipped with a tight-fitting airtight lid. Each swab was introduced into Cary-Blair medium (OXOID, UK), previously cooled with ice for one to two hours before stool sample collection. The swabs, loaded with stool (by rotating the swab into the stool or anus), were introduced into the transport medium for transportation. The upper part of the stem, touched by fingers, was cut off and discarded. The caps of the Cary-Blair tubes and the lids of the stool containers were screwed back on and the tubes labeled following the protocol described by March and Ratnam [15]. The samples were transported to the laboratory in a refrigerated cooler box to be processed. The remaining waste was destroyed following the guidelines by standards of biohazard type 2 [16,17].
2.2. Isolation and Identification
The physical and microscopic examination of stools was carried out following the standard operational procedure (SOP) of the clinical microbiology laboratory [18,19]. A loopful suspension of the stool sample was inoculated into an enrichment broth (peptone broth) in a 1:10 sample-to-broth ratio and incubated at 37°C overnight. Subsequently, a 10 µl loop-full of the overnight enrichment broth was plated onto SMAC medium (Sorbitol-MacConkey) and incubated at 37°C for 24 h [19]. Pink colonies were further plated on Eosin Methylene Blue (EMB) agar and incubated at 37°C for 24 h [20]. Colony exhibiting a metallic sheen, suggestive of Escherichia coli, were confirmed with phenotypic assay using Api20E read by commercially available Microbact GNB 24E (Oxoid, UK) according to the manufacturer’s instructions [21]. The confirmation Api20E assay was carried on the strain FK53-34 at Institut Pasteur (Paris) using a Biomerieux kit. A 100 µl of 0.5 McFarlan bacterial suspension was inoculated into each well, and the strips were then incubated. After 2 hours of incubation at 37°C, a change in the media (or after adding revelers) was used to identify the profile of the bacterium [15]. In addition, we used MALDI-TOF MS (Matrix Assisted Laser Desorption Ionization – Time Of Flight Mass Spectrometry) to confirm or identify pure growth isolates [22,23].
2.3. Susceptibility Testing
From a suspension of 0.5 McFarland, prepared from 2 to 3 pure colonies of FK53-34 on Mueller Hinton medium [24], we carried out a susceptibility test according to Comite d’antibiogramme de la Societe francaise de microbiologie guideline CA-SFM) V1.0 [25]. Discs of 5µg of Ciprofloxacin(CIP), 30µg of Cefepim (FEP), 30 µg of Aztreonam (ATM), 30µg of Cefotaxim (CTX), 75 µg of Ticarcillin (TIC), 75µg/10 µg of Ticarcillin + clavulanic acid (TCC), 20 µg of Amoxicillin (AMX), 30µg of Cefoxitin (FOX), 30 µg of Cefuroxim (CXM), 10µg of Ceftazidim (CAZ) and 30µg of Nalidixic Acid (NAL) were tested. . The inhibition diameters are interpreted as resistant, intermediate and susceptible according to reference values (diameters in mm) for disc susceptibility testing (Kirby-Bauer method) recommendations [26].
2.4. DNA Extraction and Sequencing
We extracted nucleic acid of FK53-34 at Institut Pasteur from 24-hour culture growth on petri dishes using the KingFisher technique (Themo Scientific , OXQ32853, Oxoid brand, Germany), according to the manufacturer's instructions [21]. After quality control and DNA quantification by Nanodrop spectrophotometry and Qubit fluorometer [27], we denatured. By adding 500 μl prepared 1.4 pM libraries into the reservoir, we diluted and prepared DNA fragment ends for adapter ligation and attached Illumina sequencing adapters to the DNA fragments. We amplified adapter-ligated fragments to enrich the library[28]. Sequencing libraries were constructed, and the libraries were quantified using the 2100 Bioanalyzer System (Agilent) and Kapa Sybr Fast qPCR Kit. We carry on the sequencing on an Illumina Miseq V3 platform using a 2 × 300 paired-end approach (Illumina Inc., San Diego, CA, USA) [29].
2.5. Assembly and Annotation
We assessed the quality of the raw reads of FK53-34 using FastQC v0.11.5 [30] and filtered them using Trimmomatic v3 with cut-off Q28 [29], redundancies or over-represented reads were reduced, sequencing errors corrected, and reads with nucleobase identification quality measures (Phred scores > 28). The high-quality reads of FK53-34 were assembled with a denovo assembly approach using Spades V3.13.1 [31] and checked the final assembly scaffolds using Checkm2 v1.0.2 [32] to provide accurate estimates of genome completeness, contamination and heterogeneity. The taxonomy classification was carried out using GTDB-Tk v2.4.0 (RefDB version r220) [33]. A contig rearrangement was carried on using Mauve progressive alignment and E. ruysiae (strain OPT1704, AB136)). We performed the annotation of the obtained scaffolds using Prokka1.14.5 [32]. We used Get Homologues [35] to compute the coding sequence (CDS) ANI of FK53-34 using OrthoMCL approach with close E. ruysiae genomes available on NCBI. Furthermore, we performed a core alignment of the all available E. ruysiae including FK53-34 using Scapper (https://github.com/tseemann/scapper.git), cloned on March 29th 2025, a reference-based alignment tool composed of MuMmer 4 [36] , and TrimAL [37] . To support the ANI cladogram, we constructed a core genome maximum-likelihood phylogenetic tree using raxml-ng [38] and used FigTree [39] to visualize the tree. Proksee [40] was used to visualize the assembled, the annotation, the virulome and resistome of our genome.
2.6. Resistome, Virulome Genome Sequence Typing
We characterized the resistome and the virulome of FK53-34 using Abricate v1.9.8 [41]. The resistance genes were searched against the Resfinder 4.1 database (367 resistance genes) [42], NCBI (6570 sequences downloaded on June 23, 2023), CARD V3.2.4 (6627 ontology terms, 5010 reference sequences, 1933 mutations, 3004 publications and 5057 AMR detection model [43] , ARG-ANNOT V6 (1749 sequences) [44] and MEGAres V3.0 (9,000 resistance genes antimicrobials) [45]. We identified virulence genes by mapping the genome sequences against the virulence database VFDB V1.0 using default parameters [46]. We performed in silico typing using Ectyper V0.8.1 [47] with default parameters, and sequence types (ST) were determined using MLST 2.23.0 [48] based on seven housekeeping genes (adk , fumC , gryB , icd , mdh , purA and recA ). We finally used the phylogroup classification tool, ClermonTyping [49] to identify the phylogroup of our strain.
3. Results
3.1. Clinical Case Description
A 5-year-old, 18 kg female was examined by a general practitioner for abdominal pain with vomiting and bloody diarrhea without fever. The diagnosis was “gastroenteritis”. The present clinical case was found at Centre Medico-Social de Togblekope (altitude: 71 m above sea level, coordinates: 6°16'24" N and 1°12'49" E), located in the suburban region of Lomé. Clinical symptoms are dominated by vomiting, with bloody diarrhea and absence of fever and abdominal cramps. The patient was first treated with Motilium® (domperidone) oral suspension (2/day), ceftriaxone (500mg) injectable (2/day) and Actapulgite® (activated Mormoiron attapulgite) powder in oral suspension (2 days). The vomiting stopped on the third day; however, the bloody diarrhea and abdominal cramps persisted. On the fourth day, symptoms reappear with a thermal surge up to 40°C in the afternoon. Most of the blood cells counts are within normal range (red count = 4.22x106/µl (4.8-6.2.106), white count = 6200 cells/µl, hemoglobin = 13.6 g/dl (11-15g/dl), hematocrit =43.8 (40-54), platelet = 244,000/µl (150 000-400 000/µl)). On the seventh day, The results of microbiology test and mass spectrometry (MALDI TOF) of FK53-34 showed Escherichia coli with a score of 2.2 and sensitive to all antibiotics tested. The patient was later referred to a gastroenterologist around 10 a.m., but she couldn’t survive. She died after 8 days of hospitalization, despite the care provided. The autopsy concluded at 2 p.m. a massive dehydration from infectious gastroenteritis and weight loss estimated at nearly 14% of total weight.
3.2. Phenotypic, Biochemical Characterization and Resistance Profile
A culture growth of the strain FK53-34 at 37°C in an aerobic atmosphere for 24 hours on the TSA agar as transparent, refractive and translucent colonies of various sizes. The colonies appear creamy to white, with circular shapes and edges ranging from smooth to slightly convex. They vary in size, from 1 to 3 mm in diameter. FK53-34 is Gram-negative (Suppl.Data: Bacteria Cultivation). Biochemical test performed on the API 20E assay showed results in Table 1, the strain FK53-34 shows similar phenotypic metabolism profile with that of the reference AB134 with the exception that the strain FK53-34 metabolise citrate (CIT), tryptophane (TDA) and melibiose (MEL). Compared to E. coli, the latest was positive to Alanine dehydrogenase (ADH), lysine dehydrogenase (LDC) and saccharose (SAC) while E. ruysiae was negative.
3.3. Genomics Features and Challenges in the Strain Identification
We assembled the genomes of FK53-34 into 4785617 bp, comprising 136 contigs with a GC content of 50.55% with an average length of the contigs ≈ 35,190 bp. The N50 is 94629 with a coding density of 0,87. There were 4464 coding sequences (CDS), 4558 predicted genes, 83 tRNA, 1 repeat regions, 1 mmRNA, 10rRNA, 1173 markers and no heterogeneity (Suppl.Data, Genome quality). The ANI (Average Nucleotide Identity) gives an average similarity of nucleotide sequences of 98.78 for Escherichia ruysiae. The MultiLocus Sequence Typing (MLST) match with the following allele: adk(356), gyr(73), icd(670), mdh(56), purA(322), recA(454) and no best match with fumC(~1709). The final ST ID as well as the antigenic profile which could not identify alleles for the genotype ST and OH antigens respectively. However, the serogroup of the strain FK53-34 was identified as Clade IV. The strain FK53-34 is close to the strain C14-1 and S1-IND-07-A with ANI 99.04% and 98.91% respectively (Figure 1). We found two clusters in the ANI dendrogram which was confirmed by the core genome Maximum likelihood phylogenetic tree grouped in Clade III and Clade IV (Figure 2)
3.4. Resistome, Virulome and Mobilome of FK53-34
E. ruysiae have been found to harbor antimicrobial resistance genes, including those encoding extended-spectrum beta-lactamases (ESBLs). FK53-34 carries genes associated with the production of beta-lactamases BlaEC, CPXAR, (Bla)CTX, (Bla)ampH, (Bla)AmpC1, (Bla)AmpC2, (Bla)Penicillin_Binding_Protein, (Bla)ampH. It carries the eptA and pmrF gene, conferring resistance to cationic antimicrobial peptides, such as colistin and multidrug resistance gene (Emr, Mdt, Acr,) (Table 3). Several virulence factor genes were also identified, including those encoding flagellar protein (flgABCDE, filABC, flhABC, motAB), colonization factors (fimA, fimB, fimC, fimD, fimE, fimF, fimH, fimI), fitness (iutA and kpsMII), heat-stable enterotoxin (astA, senB), chemotaxis regulatory protein ( CheR, CheB, CheY et CheZ), AidA-I adhesin-like protein (ehaAB, hofB, hofC, hofq), ompT gene for outer membrane protease and Plasmid R100 complement resistance gene (traJ, traT). We also note the presence of the operon (aatPABC), adaptation gene (cfa), adhesion and biofilm formation gene (csgABCDEFG, agn43) and blood-brain barrier invasion (ibeBC) (Table virulence factor). The virulome shows the absence of the wzx and wzy genes. We noted also the presence of specific adhesion genes (EC958_4611, EC958_4614), immune evasion (ECS88_3547), motility and biofilm (fliC-H56), and pathogenicity islands (EC55989_3335 and Z1307) in the FK53-34 isolate. (Table 4)
4. Discussion
The successful isolation and culturing of Escherichia ruysiae from the fecal sample of a five-year-old patient with gastroenteritis and bloody stools in Togo significantly expands our understanding of its ecology and its pathogenic potential. Previously, E. ruysiae has been detected in diverse hosts and locations, including humans in Europe [50], an urban wild crow in Asia [11], a domestic chicken in North America [51], and domestic dogs in Oakland, USA [52], highlighting its broad ecological niches. The presence of E. ruysiae in humans, animals and the environment confirms its contribution to the One Health concept that recognizes the interconnectedness of human, animal, and environmental health. These findings emphasize the need to investigate the phylogenomic of E. ruysiae to better understand the spray and map the evolutionary relationships as well as the global distribution of this emerging pathogen. Many studies have noted the challenge of correctly identifying E. ruysiae, often misclassifying it as E. coli, particularly when using MALDI-TOF MS [52,53]. This misidentification stems from the lack of E. ruysiae specific spectra in current databases, underscoring the urgent need to update reference libraries. Our phylogenomic analysis of E. ruysiae from various environments reveals notable genetic diversity divided into two branches associated with the Clade III [6] and the clade IV [13]. The FK53-34 strain belongs to the clade IV clusters and some of the strains in that cluster have been described to harbored antimicrobial resistance genes and found in diverse environments. For instance, it shares a close phylogenetic relationship with C14-1, a strain previously isolated from a healthy domestic hen in the UK [51], reinforcing the cross-host adaptability of E. ruysiae. Unlike E. coli, which can typically be classified into phylogroups (A, B1, B2, D, etc.) [51], E. ruysiae does not fit these categories. Furthermore, our isolate could not be assigned to a known sequence type (ST), suggesting either a novel lineage or gaps in current MLST databases for this species. This lack of classification can complicate the epidemiological tracking and the understanding of E. ruysiae's biology and clinical impact. Whether these strains are primarily commensal or pathogenic remains unclear, but their genetic diversity implies complex evolutionary pressures and niche-specific adaptation.
Genomics plays a crucial role in modern clinical microbiology, particularly in taxonomic classification, profiling antimicrobial resistance (AMR) carriage and deciphering their mechanisms. Despite phenotypic susceptibility to all antibiotics tested, genomics analysis shows the presence of common multiple resistance associated genes such as blaPenicillin Binding Protein, blaampH, acrA, emrF, baeS et tolC and many other chromosome-mediated resistance genes (blaEC-15, eptA, pmrF). Among all compared strains only our strain E. ruysiae FK53-34 possessed the gene blaEC-15, a natural non-plasmid transferable chromosomal variant of AmpC-like. This might explain why the strain was sensitive to all cephalosporins tested including Cefepim, Cefotaxim, Cefoxitin, Cefuroxim and Ceftazidim. This has been previously shown with E. coli which harbored genes associated with resistance to antimicrobial but with a susceptible phenotypic profile when tested [54,55,56,57].
The mobilome of FK53-34 shows that this genome is rich in mobile genetic elements (phages, integrons, plasmids). It is a highly diverse and active mobilome, reflecting high genome plasticity. Previous studies have shown the role of mobile elements in the prediction and their contribution to bacterial virulence [58], evolution [59], adaptation [60,61], and transfer within bacteria [62]. Of all E. ruysiae published so far, only the strain FK53-34 carries specific virulence genes namely aap/aspU, aatABCP, senB, traJT. The gene aap/aspU also known as Dispersin is thought to neutralize the bacterial cell surface, repelling and projecting the positively charged AAFs (Aggregative adherence Fimbriae), thereby facilitating the dispersion of bacteria across the intestinal mucosa. It enables the bacteria to disperse after initial aggregation and spread across the intestinal mucosa. This process also supports biofilm formation and chronic colonization, contributing to the persistent diarrhea typical of EAEC infections. This dispersal may allow the bacteria to spread and establish new foci of infection [63]. The AatABCD export system is a critical exporter responsible for secreting specific proteins from the bacterial cytoplasm to the extracellular environment. Its most well-known role in EAEC is the export of the dispersin (Aap/AspU) protein [64]. The senB heat stable enterotoxin gene encodes an enterotoxin that increases fluid and electrolyte secretion in the intestinal mucosa. It contributes to watery diarrhea, complementing other virulence factors such as the type III secretion system and invasions [65]. All these genes, if expressed, may have exacerbated the diarrhea condition of the patient, extensively lost water and inevitably leading to death. In addition, the presence of ibeB and ibeC genes is known to confer to the pathogen the ability to cross the blood-brain barrier, producing the invasion of brain endothelial cells, and promoting neonatal meningitis [66,67]. Although, the gastroenteritis looked mild and should have been treated easily with common antibiotic therapy, the miss identification of the strains, the enclosed antimicrobial resistance profile added to the arsenal of virulence factors have contributed to the overlooking of the diagnosis which let unfortunately to the death of the patient.
5. Conclusions
This study reports the first clinical isolation and genomic characterization of Escherichia ruysiae in Togo, associated with a fatal case of gastroenteritis in a pediatric patient. Our findings reveal that E. ruysiae can harbor a large spectrum of antimicrobial resistance genes not detected in the susceptibility test and a virulome mainly found in pathogenic E. coli. These results underline the diagnostic limitations of conventional methods in accurately identifying cryptic Escherichia species and highlight the potential public health threat posed by overlooked pathogenic strains. The genomic features of FK53-34 reinforce the need for expanding surveillance, updated diagnostic tools, and incorporation of E. ruysiae into routine microbial monitoring. From a One Health perspective, its presence in both clinical, veterinary and environmental settings points to the urgent integrating approaches and controlling antimicrobial resistance of emerging pathogens.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. all raw analysis data are available the the spreadsheet Suppl data.xlsx: Bacterial cultivation: shows groups of E. ruysiae FK53-34 on TSA agar plate, Taxonomy Gtbtk2: shows taxonomy classification using Gtdbtk2. CheckM2: shows assembly quality control metrics, Api20E: phenotypic assay results using Api20e, Susceptibility test: shows phonotypic sensibility to antimicrobial, Resistance genes: table showing overall antimicrobial associated genes with all publicly available genomes, Virulence factor: table showing overall virulence associated genes with all publicly available genomes, MLST: shows MLAST allele found and missing in FK53-34.
Author Contributions
All authors read and approved the final manuscript. We would like to thank the reviewers for their insightful comments on the manuscript, as their remarks led to an improvement of the work. KABO Kossi collected the samples, isolated, and characterized the genome of the strains, performed data analysis, and wrote the manuscript. MLAGA Kodjovi Dodji contributed to the scientific direction of the project, performed the bioinformatics analysis of the data, and revised the manuscript. DIONE Niokhor, prereviewed the manuscript. TCHACONDO Tchadjobo validated the research protocol and supervised the work.
Funding
This research received no external funding, but contribution have been made by LAMICODA Laboratoire de Microbiologie et de Controle qualité des denrées alimentaires for all microbiology procedures and Institut Pasteur, Paris for phenotypic characterization and whole genome sequencing. All other financial contribution has been made by authors.
Institutional Review Board Statement
This study was conducted in accordance with ethical procedures and no personally identifiable data was collected. However, all clinical data were collected with the approval of the regional health director. The study was conducted in accordance with the Declaration of Helsinki, and approved by the review board of Institut national d’hygiène (Ref: 913/2022/MSHPAUS/CAB/SG/INH) and NAGOYA guideline (Ref: 0013/MERF/SG/DRF, 04/01/2024).
Informed Consent Statement
Not applicable.
Data Availability Statement
the genome of the FK53-34 has been submitted to NCBI, Bioproject: PRJNA1223433 with accession number JBLRVR000000000.
Acknowledgments
At the same time, I would like to say a sincere thank you to Olivier Chesneau, Dominique Clermont, Marthe Goly and Fay Betsou (Institut Pasteur), TOUDJI Akouétévi Gérard (LAMICODA/UL) contributed to the microbial culture.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The Population Genetics of Commensal Escherichia Coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia Coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- OMS Le Genre Escherichia Englobe Les Espèces Commensales et Pathogènes 2018.
- Escherich, T. Die Darmbakterien Des Neugeborenen Und Säuglings. 1885.
- Shulman, S.T.; Friedmann, H.C.; Sims, R.H. Theodor Escherich: The First Pediatric Infectious Diseases Physician? Clin. Infect. Dis. 2007, 45, 1025–1029. [Google Scholar] [CrossRef]
- Van Der Putten, B.C.L.; Matamoros, S.; Mende, D.R.; Scholl, E.R.; Consortium†, C.; Schultsz, C. Escherichia Ruysiae Sp. Nov., a Novel Gram-Stain-Negative Bacterium, Isolated from a Faecal Sample of an International Traveller. Int. J. Syst. Evol. Microbiol. 2021, 71. [Google Scholar] [CrossRef]
- Brenner, D.J.; Davis, B.R.; Steigerwalt, A.G.; Riddle, C.F.; McWhorter, A.C.; Allen, S.D.; Farmer, J.J.; Saitoh, Y.; Fanning, G.R. Atypical Biogroups of Escherichia Coli Found in Clinical Specimens and Description of Escherichia Hermannii Sp. Nov. J. Clin. Microbiol. 1982, 15, 703–713. [Google Scholar] [CrossRef]
- Farmer, J.J.; Fanning, G.R.; Davis, B.R.; O’Hara, C.M.; Riddle, C.; Hickman-Brenner, F.W.; Asbury, M.A.; Lowery, V.A.; Brenner, D.J. Escherichia Fergusonii and Enterobacter Taylorae, Two New Species of Enterobacteriaceae Isolated from Clinical Specimens. J. Clin. Microbiol. 1985, 21, 77–81. [Google Scholar] [CrossRef]
- Huys, G.; Cnockaert, M.; Janda, J.M.; Swings, J. Escherichia Albertii Sp. Nov., a Diarrhoeagenic Species Isolated from Stool Specimens of Bangladeshi Children. Int. J. Syst. Evol. Microbiol. 2003, 53, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Jin, D.; Lan, R.; Wang, Y.; Meng, Q.; Dai, H.; Lu, S.; Hu, S.; Xu, J. Escherichia Marmotae Sp. Nov., Isolated from Faeces of Marmota Himalayana. Int. J. Syst. Evol. Microbiol. 2015, 65, 2130–2134. [Google Scholar] [CrossRef]
- Sakaguchi, K.; Tanabe, M.; Takizawa, S.; Kasahara, S.; Denda, T.; Koide, S.; Hayashi, W.; Nagano, Y.; Nagano, N. Zoonotic Potential and Antimicrobial Resistance of Escherichia Spp. in Urban Crows in Japan-First Detection of E. Marmotae and E. Ruysiae. Comp. Immunol. Microbiol. Infect. Dis. 2023, 100, 102040. [Google Scholar] [CrossRef]
- Siddi, G.; Piras, F.; Gymoese, P.; Torpdahl, M.; Meloni, M.P.; Cuccu, M.; Migoni, M.; Cabras, D.; Fredriksson-Ahomaa, M.; De Santis, E.P.L.; et al. Pathogenic Profile and Antimicrobial Resistance of Escherichia Coli, Escherichia Marmotae and Escherichia Ruysiae Detected from Hunted Wild Boars in Sardinia (Italy). Int. J. Food Microbiol. 2024, 421, 110790. [Google Scholar] [CrossRef]
- Campos-Madueno, E.I.; Aldeia, C.; Sendi, P.; Endimiani, A. Escherichia Ruysiae May Serve as a Reservoir of Antibiotic Resistance Genes across Multiple Settings and Regions. Microbiol. Spectr. 2023, 11, e01753–23. [Google Scholar] [CrossRef]
- Karas, Michael. ; Hillenkamp, Franz. Laser Desorption Ionization of Proteins with Molecular Masses Exceeding 10,000 Daltons. Anal. Chem. 1988, 60, 2299–2301. [Google Scholar] [CrossRef]
- Mariani-Kurkdjian, P. Chapitre 15 - Diagnostic bactériologique des infections gastro-intestinales.
- Biosafety in the Laboratory: Prudent Practices for Handling and Disposal of Infectious Materials; National Academies Press: Washington, D.C., 1989; p. 1197; ISBN 978-0-309-03975-8.
- Rutala, W.A.; Weber, D.J. 301 Control of Hospital Waste.
- Avril, J.L. Technique d’une Coproculture. Médecine Mal. Infect. 1979, 9, 478–483. [Google Scholar] [CrossRef]
- March, S.B.; Ratnam, S. Sorbitol-MacConkey Medium for Detection of Escherichia Coli 0157:H7 Associated with Hemorrhagic Colitis. J CLIN MICROBIOL.
- March, S.B.; Ratnam, S. Sorbitol-MacConkey Medium for Detection of Escherichia Coli O157:H7 Associated with Hemorrhagic Colitis. J. Clin. Microbiol. 1986, 23, 869–872. [Google Scholar] [CrossRef] [PubMed]
- Thermo Scientific Kit de d’extraction de l’ADN à Partir de Divers Échantillons Biologiques 2022.
- Emilie Cardot Martin Identification Des Microorganismes Pathogènes Par Spectrométrie de Masse de Type MALDI TOF En Microbiologie Médicale 2020.
- Osthoff, M.; Gürtler, N.; Bassetti, S.; Balestra, G.; Marsch, S.; Pargger, H.; Weisser, M.; Egli, A. Impact of MALDI-TOF-MS-Based Identification Directly from Positive Blood Cultures on Patient Management: A Controlled Clinical Trial. Clin. Microbiol. Infect. 2017, 23, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Stegger, M.; Leihof, R.F.; Baig, S.; Sieber, R.N.; Thingholm, K.R.; Marvig, R.L.; Frimodt-Møller, N.; Nielsen, K.L. A Snapshot of Diversity: Intraclonal Variation of Escherichia Coli Clones as Commensals and Pathogens. Int. J. Med. Microbiol. 2020, 310, 151401. [Google Scholar] [CrossRef] [PubMed]
- EUCAST Comité de l’antibiogramme de la Société Française de Microbiologie 2023.
- EUCAST (European Committee on Antimicrobial Susceptibility Testing) Breakpoint Tables for Interpretation of MICs and Zone Diameters 2024.
- Animal Health Institute, Sebeta, Ethiopia; K, E.; K, L.; Animal Health Institute, Sebeta, Ethiopia Review on Illumina Sequencing Technology. Austin J. Vet. Sci. Anim. Husb. 2022, 9. [CrossRef]
- Illumina MiniSeq System Guide. Retrieved from. 2023.
- Aworh, M.K.; Ekeng, E.; Nilsson, P.; Egyir, B.; Owusu-Nyantakyi, C.; Hendriksen, R.S. Extended-Spectrum ß-Lactamase-Producing Escherichia Coli Among Humans, Beef Cattle, and Abattoir Environments in Nigeria. Front. Cell. Infect. Microbiol. 2022, 12, 869314. [Google Scholar] [CrossRef]
- Liu, N.; Zhang, G.; Fang, L.; Geng, R.; Shi, S.; Li, J.; Wang, W.; Lin, M.; Chen, J.; Si, Y.; et al. The Marine-Origin Exopolysaccharide-Producing Bacteria Micrococcus Antarcticus HZ Inhibits Pb Uptake in Pakchoi (Brassica Chinensis L.) and Affects Rhizosphere Microbial Communities. Microorganisms 2024, 12, 2002. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Chklovski, A.; Parks, D.H.; Woodcroft, B.J.; Tyson, G.W. CheckM2: A Rapid, Scalable and Accurate Tool for Assessing Microbial Genome Quality Using Machine Learning. Nat. Methods 2023, 20, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk v2: Memory Friendly Classification with the Genome Taxonomy Database.
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Contreras-Moreira, B.; Vinuesa, P. GET_HOMOLOGUES, a Versatile Software Package for Scalable and Robust Microbial Pangenome Analysis. Appl. Environ. Microbiol. 2013, 79, 7696–7701. [Google Scholar] [CrossRef]
- Marçais, G.; Delcher, A.L.; Phillippy, A.M.; Coston, R.; Salzberg, S.L.; Zimin, A. MUMmer4: A Fast and Versatile Genome Alignment System. PLOS Comput. Biol. 2018, 14, e1005944. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Lam, T.T.; Max Carvalho, L.; Pybus, O.G. Exploring the Temporal Structure of Heterochronous Sequences Using TempEst (Formerly Path-O-Gen). Virus Evol. 2016, 2, vew007. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-Depth Characterization and Visualization of Bacterial Genomes. Nucleic Acids Res. 2023, 51, W484–W492. [Google Scholar] [CrossRef]
- Seemann, T. ABRicate, Un Logiciel Combinanr Différentes Bases de Données Pour Le Dépistage de Masse Des Contigs Pour La Résistance Aux Antimicrobiens Ou Les Gènes de Virulence 2020.
- Florensa, A.F.; Kaas, R.S.; Clausen, P.T.L.C.; Aytan-Aktug, D.; Aarestrup, F.M. ResFinder – an Open Online Resource for Identification of Antimicrobial Resistance Genes in next-Generation Sequencing Data and Prediction of Phenotypes from Genotypes. Microb. Genomics 2022, 8. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2019, gkz935. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.-M. ARG-ANNOT, a New Bioinformatic Tool To Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef]
- Bonin, N.; Doster, E.; Worley, H.; Pinnell, L.J.; Bravo, J.E.; Ferm, P.; Marini, S.; Prosperi, M.; Noyes, N.; Morley, P.S.; et al. MEGARes and AMR++, v3.0: An Updated Comprehensive Database of Antimicrobial Resistance Determinants and an Improved Software Pipeline for Classification Using High-Throughput Sequencing. Nucleic Acids Res. 2023, 51, D744–D752. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A General Classification Scheme for Bacterial Virulence Factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Bessonov, K.; Laing, C.; Robertson, J.; Yong, I.; Ziebell, K.; Gannon, V.P.J.; Nichani, A.; Arya, G.; Nash, J.H.E.; Christianson, S. ECTyper: In Silico Escherichia Coli Serotype and Species Prediction from Raw and Assembled Whole-Genome Sequence Data. Microb. Genomics 2021, 7. [Google Scholar] [CrossRef]
- GNU Parallel 20230522; charles; Zenodo, 2023;
- Clermont, O.; Gordon, D.; Denamur, E. Guide to the Various Phylogenetic Classification Schemes for Escherichia Coli and the Correspondence among Schemes. Microbiology 2015, 161, 980–988. [Google Scholar] [CrossRef]
- Van Der Putten, B.C.L.; Matamoros, S.; Mende, D.R.; Scholl, E.R.; Consortium†, C.; Schultsz, C. Escherichia Ruysiae Sp. Nov., a Novel Gram-Stain-Negative Bacterium, Isolated from a Faecal Sample of an International Traveller. Int. J. Syst. Evol. Microbiol. 2021, 71. [Google Scholar] [CrossRef] [PubMed]
- Thomson, N.M.; Gilroy, R.; Getino, M.; Foster-Nyarko, E.; Van Vliet, A.H.M.; La Ragione, R.M.; Pallen, M.J. Remarkable Genomic Diversity among Escherichia Isolates Recovered from Healthy Chickens. PeerJ 2022, 10, e12935. [Google Scholar] [CrossRef] [PubMed]
- Dione, N.; Mlaga, K.D.; Liang, S.; Jospin, G.; Marfori, Z.; Alvarado, N.; Scarsella, E.; Uttarwar, R.; Ganz, H.H. Comparative Genomic and Phenotypic Description of Escherichia Ruysiae: A Newly Identified Member of the Gut Microbiome of the Domestic Dog. Front. Microbiol. 2025, 16, 1558802. [Google Scholar] [CrossRef] [PubMed]
- Sauget, M.; Valot, B.; Bertrand, X.; Hocquet, D. Can MALDI-TOF Mass Spectrometry Reasonably Type Bacteria? Trends Microbiol. 2017, 25, 447–455. [Google Scholar] [CrossRef]
- Golden, A.R.; Karlowsky, J.A.; Walkty, A.; Baxter, M.R.; Denisuik, A.J.; McCracken, M.; Mulvey, M.R.; Adam, H.J.; Bay, D.; Zhanel, G.G. Comparison of Phenotypic Antimicrobial Susceptibility Testing Results and WGS-Derived Genotypic Resistance Profiles for a Cohort of ESBL-Producing Escherichia Coli Collected from Canadian Hospitals: CANWARD 2007–18. J. Antimicrob. Chemother. 2021, 76, 2825–2832. [Google Scholar] [CrossRef]
- Ellington, M.J.; Ekelund, O.; Aarestrup, F.M.; Canton, R.; Doumith, M.; Giske, C.; Grundman, H.; Hasman, H.; Holden, M.T.G.; Hopkins, K.L.; et al. The Role of Whole Genome Sequencing in Antimicrobial Susceptibility Testing of Bacteria: Report from the EUCAST Subcommittee. Clin. Microbiol. Infect. 2017, 23, 2–22. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.S.; Jelacic, S.; Habeeb, R.L.; Watkins, S.L.; Tarr, P.I. The Risk of the Hemolytic–Uremic Syndrome after Antibiotic Treatment ofEscherichia ColiO157:H7 Infections. N. Engl. J. Med. 2000, 342, 1930–1936. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, W.; Jiang, X.; Yao, T.; Wang, L.; Yang, B. Virulence-Related O Islands in Enterohemorrhagic Escherichia Coli O157:H7. Gut Microbes 2021, 13, 1992237. [Google Scholar] [CrossRef]
- Shi, Z.; Gewirtz, A. Together Forever: Bacterial–Viral Interactions in Infection and Immunity. Viruses 2018, 10, 122. [Google Scholar] [CrossRef]
- Touchon, M.; Rocha, E.P.C. Coevolution of the Organization and Structure of Prokaryotic Genomes. Cold Spring Harb. Perspect. Biol. 2016, 8, a018168. [Google Scholar] [CrossRef]
- Frost, L.S.; Leplae, R.; Summers, A.O.; Toussaint, A. Mobile Genetic Elements: The Agents of Open Source Evolution. Nat. Rev. Microbiol. 2005, 3, 722–732. [Google Scholar] [CrossRef]
- Casacuberta, E.; González, J. The Impact of Transposable Elements in Environmental Adaptation. Mol. Ecol. 2013, 22, 1503–1517. [Google Scholar] [CrossRef]
- Juhas, M.; Van Der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic Islands: Tools of Bacterial Horizontal Gene Transfer and Evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef]
- Sheikh, J.; Czeczulin, J.R.; Harrington, S.; Hicks, S.; Henderson, I.R.; Le Bouguénec, C.; Gounon, P.; Phillips, A.; Nataro, J.P. A Novel Dispersin Protein in Enteroaggregative Escherichia Coli. J. Clin. Invest. 2002, 110, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Nishi, J.; Sheikh, J.; Mizuguchi, K.; Luisi, B.; Burland, V.; Boutin, A.; Rose, D.J.; Blattner, F.R.; Nataro, J.P. The Export of Coat Protein from Enteroaggregative Escherichia Coli by a Specific ATP-Binding Cassette Transporter System. J. Biol. Chem. 2003, 278, 45680–45689. [Google Scholar] [CrossRef]
- Farfan, M.J.; Torres, A.G. Molecular Mechanisms That Mediate Colonization of Shiga Toxin-Producing Escherichia Coli Strains. Infect. Immun. 2012, 80, 903–913. [Google Scholar] [CrossRef]
- Kim, K.S. Pathogenesis of Bacterial Meningitis: From Bacteraemia to Neuronal Injury. Nat. Rev. Neurosci. 2003, 4, 376–385. [Google Scholar] [CrossRef]
- Wang, Y.; Kim, K.S. Role of OmpA and IbeB in Escherichia Coli K1 Invasion of Brain Microvascular Endothelial Cells In Vitro and In Vivo. Pediatr. Res. 2002, 51, 559–563. [Google Scholar] [CrossRef]
- Cascales, E.; Buchanan, S.K.; Duché, D.; Kleanthous, C.; Lloubès, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin Biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef]
- Touchon, M.; Hoede, C.; Tenaillon, O.; Barbe, V.; Baeriswyl, S.; Bidet, P.; Bingen, E.; Bonacorsi, S.; Bouchier, C.; Bouvet, O.; et al. Organised Genome Dynamics in the Escherichia Coli Species Results in Highly Diverse Adaptive Paths. PLoS Genet. 2009, 5, e1000344. [Google Scholar] [CrossRef]
- Forde, B.M.; Ben Zakour, N.L.; Stanton-Cook, M.; Phan, M.-D.; Totsika, M.; Peters, K.M.; Chan, K.G.; Schembri, M.A.; Upton, M.; Beatson, S.A. The Complete Genome Sequence of Escherichia Coli EC958: A High Quality Reference Sequence for the Globally Disseminated Multidrug Resistant E. Coli O25b:H4-ST131 Clone. PLoS ONE 2014, 9, e104400. [Google Scholar] [CrossRef]
- Ashley, J.W.; Shi, Z.; Zhao, H.; Li, X.; Kesterson, R.A.; Feng, X. Genetic Ablation of CD68 Results in Mice with Increased Bone and Dysfunctional Osteoclasts. PLoS ONE 2011, 6, e25838. [Google Scholar] [CrossRef]
- Holmberg, S.D.; Hamburger, M.E.; Moorman, A.C.; Wood, K.C.; Palella, F.J., Jr. ; HIV Outpatient Study Investigators Factors Associated with Maintenance of Long-Term Plasma Human Immunodeficiency Virus RNA Suppression. Clin. Infect. Dis. 2003, 37, 702–707. [Google Scholar] [CrossRef]
- Perna, N.T.; Plunkett, G.; Burland, V.; Mau, B.; Glasner, J.D.; Rose, D.J.; Mayhew, G.F.; Evans, P.S.; Gregor, J.; Kirkpatrick, H.A.; et al. Genome Sequence of Enterohaemorrhagic Escherichia Coli O157:H7. Nature 2001, 409, 529–533. [Google Scholar] [CrossRef]
- Williams, J.R.; Yang, R.; Clifford, J.L.; Watson, D.; Campbell, R.; Getnet, D.; Kumar, R.; Hammamieh, R.; Jett, M. Functional Heatmap: An Automated and Interactive Pattern Recognition Tool to Integrate Time with Multi-Omics Assays. BMC Bioinformatics 2019, 20, 81. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Heatmap of ortholog CDS Average Nucleotide Identity (ANI) of E. ruysiae FK53-34 compared to other publicly available E. ruysiae as of February 24th 2025. The ANI was computed using the Get Hemologue package with default parameters [35]. The heatmap shows two clusters as demonstrated by Joshua R. Williams, 2009 [74] and the strain FK53-34 belong to the clade IV cluster and is close to the strains C14.1 and S1.IND.07.A confirming the new classification of the FK53-34 as E. ruysiae.
Figure 1.
Heatmap of ortholog CDS Average Nucleotide Identity (ANI) of E. ruysiae FK53-34 compared to other publicly available E. ruysiae as of February 24th 2025. The ANI was computed using the Get Hemologue package with default parameters [35]. The heatmap shows two clusters as demonstrated by Joshua R. Williams, 2009 [74] and the strain FK53-34 belong to the clade IV cluster and is close to the strains C14.1 and S1.IND.07.A confirming the new classification of the FK53-34 as E. ruysiae.
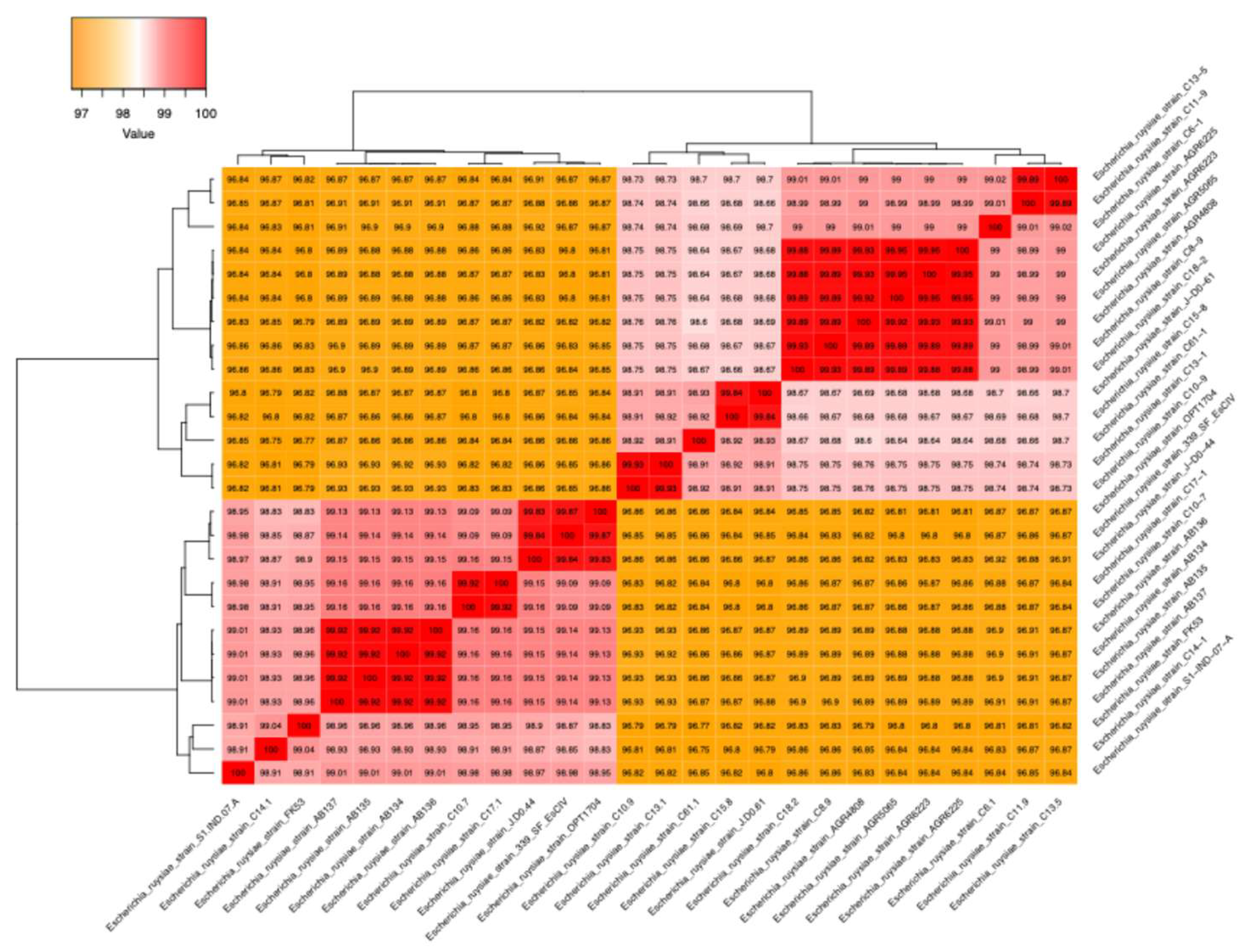
Figure 2.
Maximum likelihood phylogenetic tree of the core genome of all available E. ruysiae, including the strain FK53-34. The tree is mid-rooted in decreasing topology and shows two clades III and IV, the strain Fk53-34 belongs to the clade IV clusters.
Figure 2.
Maximum likelihood phylogenetic tree of the core genome of all available E. ruysiae, including the strain FK53-34. The tree is mid-rooted in decreasing topology and shows two clades III and IV, the strain Fk53-34 belongs to the clade IV clusters.
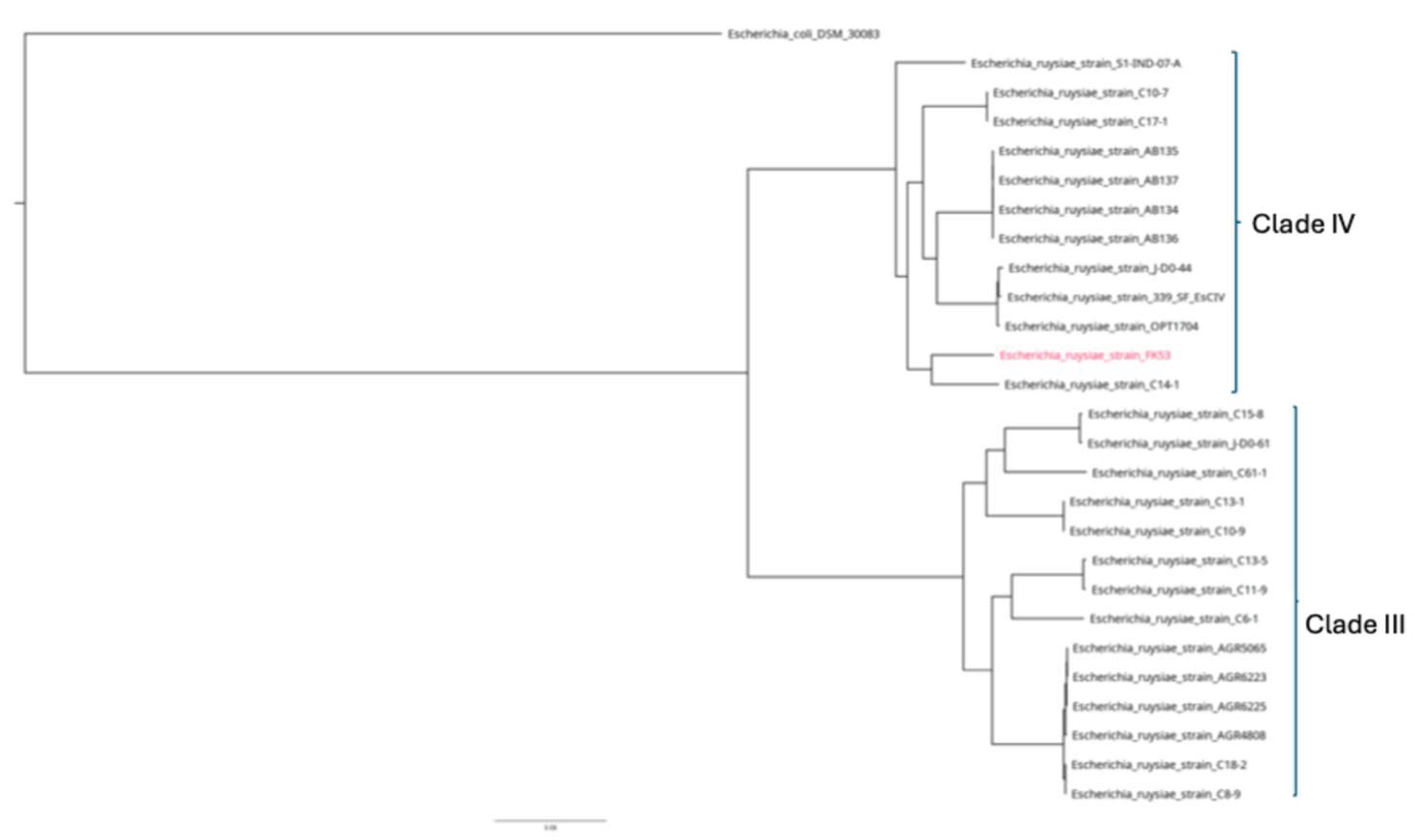

Table 1.
Biochemical reaction assay of the strain FK53-34, E. ruysiae AB134 and E. coli.
| Subtract tested | FK53-34 (this study) | E. ruysiae AB136 [50] | E.coli [11] |
| ONPG | + | + | + |
| ADH | - | - | + |
| LDC | - | - | + |
| ODC | + | + | + |
| CIT | + | - | - |
| H2S | - | - | - |
| URE | - | - | - |
| TDA | + | - | - |
| IND | - | +/- | + |
| VP | - | + | - |
| GEL | - | - | - |
| GLU | + | + | + |
| MAN | + | + | + |
| INO | - | - | - |
| SOR | + | + | + |
| RHA | + | + | + |
| SAC | - | - | + |
| MEL | + | - | - |
| AMY | - | - | - |
| ARA | + | + | + |
ONPG – Ortho-Nitrophenyl-β-galactoside test (tests for β-galactosidase activity) , ADH – Arginine Dihydrolase (tests ability to hydrolyze arginine), LDC – Lysine Decarboxylase (tests ability to decarboxylate lysine), ODC – Ornithine Decarboxylase (tests ability to decarboxylate ornithine), CIT – Citrate utilization test (ability to use citrate as sole carbon source), H2S – Hydrogen Sulfide production, URE – Urease (tests ability to hydrolyze urea), TDA – Tryptophan Deaminase (aDeaminase), IND – Indole production (tests ability to produce indole from tryptophan), VP – Voges-Proskauer test (detects acetoin production), GEL – Gelatinase (tests hydrolysis of gelatin), GLU – Glucose fermentation, MAN – Mannitol fermentation, INO – Inositol fermentation, SOR – Sorbitol fermentation, RHA – Rhamnose fermentation, SAC – Sucrose fermentation, MEL – Melibiose fermentation, AMY – Amygdalin fermentation, ARA – Arabinose fermentation.
Table 2.
Susceptibility test results of E. ruysiae FK53-34.
| Antibiotic | Acronyms | Load (µg) | Interpretation |
| Ciprofloxacin | CIP | 5 | S |
| Cefepime | FEP | 30 | S |
| Aztreonam | ATM | 30 | S |
| Cefotaxime | CTX | 30 | S |
| Ticarcillin-clavulanate | TCC | 75 | S |
| Amoxicillin | AMO | 20 | S |
| Cefoxitin | FOX | 30 | S |
| Cefuroxime | CXM | 30 | S |
| Ceftazidime | CAZ | 10 | S |
| Nalidixic acid | NAL | 30 | S |
Table 3.
Essential Antimicrobial genes found in all E. ruysiae.
| Isolate | (AGly) aadA1-pm | (Bla)PBP_Ecoli | (Bla) ampH_Ecoli | (Bla) blaLAP-2 | (Flq)qnr-S1 | (Sul)sul2 | (Tet)tetA | (Tet)tetR | CTX | blaEC-15 | blaEC-8 |
| ABC134 | - | + | + | - | - | - | - | - | - | + | - |
| ABC135 | - | + | + | - | - | - | - | - | - | + | - |
| ABC136 | - | + | + | - | - | - | - | - | - | + | - |
| ABC137 | - | + | + | - | - | - | - | - | - | + | - |
| C10-7 | - | + | + | - | - | - | - | - | - | + | - |
| C10-9 | - | + | + | - | - | - | - | - | - | + | - |
| C11-9 | - | + | + | - | - | - | - | - | + | + | - |
| C13-1 | - | + | + | - | - | - | - | - | - | + | - |
| C13-5 | - | + | + | - | - | - | - | - | - | + | - |
| C14-1 | - | + | + | - | - | - | - | - | - | + | - |
| C15-8 | - | + | + | - | - | - | - | - | + | + | - |
| C17-1 | - | + | + | - | - | - | - | - | - | + | - |
| C18-2 | - | + | + | - | - | - | - | - | - | + | - |
| C6-1 | - | + | + | - | - | - | - | - | + | - | + |
| C61-1 | - | + | + | - | - | - | - | - | - | + | - |
| C8-9 | - | + | + | - | - | - | - | - | - | + | - |
| OPT1704 | - | + | + | + | + | + | - | - | + | + | - |
| S1-IND-07-A | + | + | + | - | + | + | + | + | + | + | - |
| FK53-34 | - | + | + | - | - | - | - | - | - | + | - |
Table 4.
Distribution of virulence genes found only in strain FK53-34.
| Isolates | aap/aspU | aatA | aatB | aatC | aatP | agn43 | senB | traJ | trat |
| ABC134 | - | - | - | - | - | - | - | - | - |
| ABC135 | - | - | - | - | - | - | - | - | - |
| ABC136 | - | - | - | - | - | - | - | - | - |
| ABC137 | - | - | - | - | - | - | - | - | - |
| C10-7 | - | + | - | - | - | - | - | - | - |
| C10-9 | - | + | - | - | - | - | - | - | - |
| C11-9 | - | + | - | - | - | - | - | - | - |
| C13-1 | - | + | - | - | - | - | - | - | - |
| C13-5 | - | + | - | - | - | - | - | - | - |
| C14-1 | - | - | - | - | - | - | - | - | - |
| C15-8 | - | - | - | - | - | - | - | - | - |
| C17-1 | - | + | - | - | - | - | - | - | - |
| C18-2 | - | - | - | - | - | - | - | - | - |
| C6-1 | - | + | - | - | - | - | - | - | - |
| C61-1 | - | - | - | - | - | - | - | - | - |
| C8-9 | - | - | - | - | - | - | - | - | - |
| OPT1704 | - | - | - | - | - | + | - | - | - |
| S1-IND-07-A | - | - | - | - | - | - | - | - | - |
| FK53-34 | + | + | + | + | + | + | + | + | + |
aap: Anti-aggregation/Dispersin, aatABCP: Autotransporter Adhesin, agn43: Antigen 43 (secretive system), senB: Secreted Enterotoxin (heat stable enterotoxin), traJT: transporter, pili.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



