Submitted:
28 July 2025
Posted:
29 July 2025
You are already at the latest version
Abstract
Background: Endoplasmic reticulum (ER) stress occurs when misfolded or unfolded proteins ac-cumulate in the organelle and contributes to the pathogenesis of various diseases. A previous study from our research group showed that aqueous extract from the Sambucus ebulus fruit has anti-inflammatory properties, possibly by reducing ER stress. The extract was found to contain high levels of neochlorogenic acid, chlorogenic acid, idaein, epicatechin, resveratrol, and chry-santhemin. The present review summarizes the effects of these phtyochemicals on ER stress.
Methods: We queried the PubMed and ScienceDirect databases for primary studies discussing ER stress markers influenced by neochlorogenic acid, chlorogenic acid, idaein, epicatechin, resvera-trol, and chrysanthemin.
Results: 43 articles were selected for review. No sufficient data was found regarding neochloro-genic acid and idaine in the context of ER stress. Other polyphenols, at low concentrations, reduce ER stress markers following exposure to stress agents in various experimental models. Interstingly, high doses of resveratrol activate pro-apoptotic signaling in cancer cell lines.
Conclusions: A causal relationship between the polyphenols in the extract and ER stress modu-lation was identified. The PERK pathway was most strongly associated with the effects of the listed compounds. Although further research is needed, recent findings suggest potential thera-peutic applications of these phytochemicals for conditions associated with chronic cellular stress.
Keywords:
endoplasmic reticulum stress
; resveratrol
; chlorogenic acid
; epicatechin
; chrysanthemin
; unfolded protein response
; Sambucus ebulus
; dwarf elderberry
1. Introduction
The endoplasmic reticulum is an organelle characteristic only of eukaryotic cells, that performs various functions, including protein synthesis, post-translational protein modification, participation in lipid metabolism, calcium ion storage, and glycogen production. [1] The lumen of this organelle contains chaperones and enzymes necessary for the proper folding of newly synthesized secretory and membrane proteins, such as PDI (protein disulfide isomerase), GRP78/BiP (glucose-regulated protein 78/binding immunoglobulin protein), GRP94 (glucose-regulated protein 94), etc. [2] Endoplasmic reticulum (ER) stress occurs when misfolded or unfolded proteins accumulate within the ER lumen. It can be caused by a variety of factors, including increased demand for protein synthesis, genetic mutations, oxidative stress, and more. [3] There are special signaling pathways in cells, known as the unfolded protein response (UPR), whose purpose is to rid the cell of ER stress and restore homeostasis. This can be achieved through decreased protein synthesis, increased production of folding-facilitating molecules, or enhanced degradation of misfolded proteins. However, if homeostasis cannot be restored, the UPR may ultimately trigger apoptosis (Figure 1). [4] Chronic inflammation can both induce and be a result of ER stress. Activation of the PERK and IRE1 pathways leads to stimulation of inflammatory signaling. [5,6].
ER stress is involved in the pathogenesis of various diseases, such as cancers, metabolic, and neurodegenerative disorders. The regulation of the adaptive response is an important mechanism guaranteeing the survival and growth of tumor cells, as they grow in an unfavorable environment, such as hypoxia, nutrient deficiencies, and increased demand for protein synthesis. [7] These stressors place a significant burden on the endoplasmic reticulum. For example, in conditions of chronic stress, many tumors upregulate the chaperone GRP78, which promotes cell proliferation, metastasis, and drug resistance. [8,9] The transcription factor XBP1s has been reported to drive the pathogenesis of multiple myeloma. [10] Since the activation of the IRE1-XBP1 pathway is related to the expression of genes involved in endoplasmic reticulum-associated degradation (ERAD), which helps eliminate misfolded proteins, Hepatitis C virus successfully suppresses this pathway, thereby stimulating viral protein synthesis and inducing cellular stress. [11] The accumulation of the large surface protein of Hepatitis B virus also leads to ER stress. [12] Ethyl alcohol has also been shown to induce CHOP expression and trigger hepatocyte apoptosis in the murine model of intragastric ethanol feeding. [13] The role of ER stress in the development of non-alcoholic fatty liver disease (NAFLD) has been demonstrated in an animal model of diet-induced obesity in mice, where overexpression of GRP78 in the liver of mice suppressed insulin signaling and ER stress-dependent activation of the transcription factor SREBP-1c, a key regulator of hepatic lipogenesis. [14] ER stress also contributes to the development of type 2 diabetes and obesity by impairing insulin signaling. This occurs through JNK activation, which phosphorylates IRS1 (insulin receptor substrate 1), thereby preventing proper hormone action. [15] In the context of cardiovascular disease, Tsukano et al. reported that CHOP-mediated apoptosis of foam cells in the vessels of the heart contributes to the rupture of atherosclerotic plaques and the onset of acute coronary syndrome. [16] There is a link between ER stress and some neurological diseases as well. Mutations in the gene encoding presenilin-1 (PS1), associated with familial Alzheimer’s disease, increase cellular sensitivity to ER stress by disrupting the UPR. [17] ER stress increases the expression of the gene encoding parkin, an E3 ubiquitin ligase, involved in the degradation of proteins via ubiquitin-proteasome system. Loss of parkin function removes its neuroprotective effect and makes them susceptible to cellular stress. Mutations in this gene are known to cause juvenile Parkinson’s disease. [18] ER stress has also been implicated in the pathogenesis of amyotrophic lateral sclerosis (ALS). [19].
Polyphenols are found in a wide range of fruits, vegetables, drinks, and cereals. They provide protection from ultraviolet radiation and pathogens, and partly determine flavor profiles. Their potential health benefits have been extensively studied in recent decades. Polyphenols are categorized into four main classes: phenolic acids, flavonoids, stilbenes, and lignans. Flavonoids are further divided into six subclasses based on their specific structural characteristics: flavones, flavanones, flavonols, flavanols, anthocyanins, and isoflavones. [20]
Sambucus ebulus, also known as dwarf elder, is present in the folk medicine of many countries in Europe and the Middle East. Тhe fruits are the most commonly utilized part of the plant, as they contain the highest concentration of polyphenolic compounds [21,22] and have traditionally been used in the form of marmalade for the treatment of tuberculosis and haemorrhoids, as well as in syrup form for appetite-stimulating and tonic action. [23] Our research group recently analyzed the phytochemical composition and biological activity of an aqueous-alcoholic extract of Sambucus ebulus fruits. The results of the chemical analysis revealed a high content of the following compounds: neochlorogenic acid, chlorogenic acid, cyanidin-3-O-galactoside or idaein, epicatechin, resveratrol, cyanidin-3-O-glucoside or chrysanthemin. Using the J774A.1 macrophage cell line, the study demonstrated significant anti-inflammatory properties of the extract, accompanied by a reduction in key endoplasmic reticulum stress markers, including peIF2α, ATF6, and CHOP, under conditions of LPS-induced inflammatory activation. The reported decrease in the expression of these proteins suggests that modulation of ER stress is a likely additional mechanism by which the extract exerts its anti-inflammatory effects. [24] However, the specific phytochemical components responsible for the modulation of ER stress were not identified in the cited study. The present review aims to summarize the effects of the most prevalent polyphenolic compounds identified in the extract on mechanisms related to endoplasmic reticulum stress, based on available data from scientific literature. Thus, the potential causal relationship between the polyphenols contained in the extract and the observed biological effects can be elucidated.
2. Materials and Methods
2.1. Search Strategy
We applied the ’’Advanced search’’ option in two databases, Pubmed and ScienceDirect, where we used the following keywords and Boolean operators: ’’Neochlorogenic acid’’ and ’’ER stress’’, ’’Chlorogenic acid’’ and ’’ER stress’’, ’’Resveratrol’’ and ’’ER stress’’, “Cyanidin-3-o-glucoside” OR “Chrysanthemin” and “ER stress”, “Cyanidin-3-o-galactoside” OR “Idaein” and “ER stress”, ’’Epicatechin’’ and ’’ER stress’’. We refined the results by adding supplementary filters to ensure relevance and quality:
- Text availability – free full texts
- Publication date – from 2000/1/1 to 2025/5/1
- Article type – research articles
- Subject area – Biochemistry, Genetics and Molecular Biology
- Access type – open access and open archive
2.2. Inclusion and Exclusion Criteria
The review included in vivo or in vitro studies that focused on the effects of the individual polyphenolic compounds (resveratrol, cyanidin-3-o-glucoside, cyanidin-3-o-galactoside, epicatechin, neochlorogenic acid, and chlorogenic acid) on ER stress. Full text articles discussing markers and pathways, affected by these phytochemicals, and providing sufficient experimental details (e.g., dosage, model system) for evaluation were also defined as inclusion criteria.
Studies were excluded if they examine plant extracts or analogues of the specific isolated compounds. Articles that didn’t evaluate ER stress key markers or molecular pathways were not considered. Records limited to anti-inflammatory, metabolic, antioxidant or other biological effects of ingredients, without any relation to ER stress, were also excluded.
2.3. Statistical Analyses and Presentation
MS Excel 2016 was used for statistical processing and graphical presentation of the collected data. BioRender was used to build a schematic for the ER stress signaling pathways.
3. Results
3.1. Study Selection
We were able to find 100 records from the PubMed and ScienceDirect databases. The study selection process followed the PRISMA flowchart (Figure 2). First, twenty duplicate records were removed. After screening the remaining 80 articles, 20 titles or abstracts didn’t meet the inclusion and exclusion criteria and were therefore excluded. Sixty full-text articles were assessed for eligibility. Studies discussing biological properties of plant extracts, containing the listed phytochemicals or their analogues, weren’t considered. Ultimately, 42 articles underwent literary analysis.
3.2. Presentation of the Results
Available experimental evidence on the effects of the selected compounds on UPR signaling was found for only 4 of the 6 phytochemicals listed in the inclusion criteria – resveratrol, chlorogenic acid, epicatechin, and cyanidin-3-o-glucoside.
3.2.1. Modulation of ER Stress by Resveratrol
The influence of resveratrol on endoplasmic reticulum stress has been widely investigated by multiple groups. Analysis of the studies revealed that resveratrol modulates the ER stress in different ways depending on the dose and the type of experimental model used, as presented in Table 1. Under conditions of induced ER stress, and when administered at low doses, resveratrol suppresses UPR signaling and reduces the expression of key markers such as GRP78, GRP94, CHOP, p-PERK, IRE1, p-eIF2α and ATF6, thus exhibiting a protective effect. [25,26,27,28,29,30,31,32,33,34,35] Conversely, at higher doses – particularly in tumor cell lines – resveratrol enhances ER stress and apoptosis, which is an indication of its cytotoxic action. [31,36,37,38,39,40] The data show that this polyphenolic compound primarily affects the PERK and IRE1 signaling cascades and modulates the expression of proteins and chaperones involved in the proper folding of newly synthesized polypeptide chains. [27,35,36,37,41,42,43,44,45]
3.2.2. Modulation of ERS by Cyanidin-3-o-Glucoside, Chlorogenic Acid and Epicatechin
According to literature, in experimental models of induced ER stress cyanidin-3-o-glycoside decreases the expression of GRP78, p-PERK, p-eIF2α, ATF4, CHOP, IRE1, XBP1, and ATF6, which indicates its potential to suppress UPR signaling. A similar effect is observed for chlorogenic acid, but its impact on the IRE1 pathway is less characterized. Although data on epicatechin are scarce, it also has the ability to regulate protein homeostasis by acting on IRE1 and PERK signaling pathways. These polyphenols have a protective effect at low doses, similar to the effects of resveratrol.
4. Discussion
Our study investigates and summarizes the known effects of specific polyphenols on endoplasmic reticulum (ER) stress, a process implicated in the development of a wide range of diseases, as mentioned above. This research builds upon our previous work, in which we observed the ER stress-modulating activity of an aqueous extract from the Sambucus ebulus fruit. However, information on the main bioactive components responsible for this effect is still lacking. [24] Future experimental work would enrich our understanding of the mechanisms underlying the biological activity of dwarf elderberry fruits.
In general, the four examined polyphenols act on one or more branches of the UPR signaling. While some studies do not specify the exact branch affected, altered expression levels of certain molecules suggest the pathway involved. Considering Table 2 and Table 3, the polyphenols most frequently influence the PERK signaling cascade, along with a reduction of CHOP, a critical mediator of ER stress-induced apoptosis. Since all four polyphenols affect PERK and IRE1, this may explain the observed anti-inflammatory properties of the dwarf elderberry extract, as these pathways can directly activate inflammatory cascades. [6] Data from two independent studies suggest that epicatechin does not significantly modulate the ATF6 signaling pathway. However, there are some key methodological differences – one study used antibodies against cleaved ATF6 as part of the protocol, while the other did not include such antibodies. [52,53] Thus, the influence of this polyphenol on the ATF6 branch of the UPR is still unexplored, highlighting the need for furhter investigation. Experimental studies are also needed to address the current lack of data on the effects of neochlorogenic acid and idaein on ER stress.
The precise molecular mechanisms by which these phenolic compounds exert their effects on ER stress are not fully understood. It is known, for instance, that the activation of sirtuin 1 (SIRT1) is key to resveratrol’s activity. SIRT1 is an NAD+-dependent protein deacetylase that modulates the activity of molecules involved in the UPR pathway. [25,30,31,47] A study by Luo et al. (2023) clearly highlighted the role of SIRT1 as a mediator of resveratrol’s effects, as inhibition of this enzyme abolished the compound’s protective action against bupivacaine-induced neurotoxicity in PC12 cells. [35] SIRT1 stimulation has also been reported for cyanidin-3-O-glucoside. [63] Resveratrol’s protective effect under induced endoplasmic reticulum (ER) stress may also be mediated by clusterin, a molecular chaperone induced by stress, аlthough its connection to SIRT1 remains unclear. [30] Moreover, a mechanism independent of SIRT1 has been reported by Graham et al., who suggest that resveratrol stimulates the enzyme protein phosphatase 1α (PP1α), which in turn inhibits protein kinase B (Akt), a modulator of protein synthesis and endoplasmic reticulum stress. [45] Furthermore, in prostate cancer cell lines, resveratrol has also been shown to deplete calcium stores in the ER and suppress the store-operated Ca2+ entry (SOCE) mechanism, ultimately leading to ER stress and cell death. [39] Further research should focus on elucidating the mechanisms by which phytochemicals modulate the expression of key markers of UPR signaling, especially in the case of epicatechin and chlorogenic acid.
The effects of polyphenols on endoplasmic reticulum stress in various in vivo and in vitro models suggest that these substances could be used as therapeutic agents. Consequently, plant sources containing these compounds may also be of interest as functional foods. For example, resveratrol, a stilbene, is found in grapes, red wine, berries, and peanuts, while chrysanthemin, an antocyanin, is present in berries, black beans, and black rice. [64,65,66,67] Epicatechin is abundant in green tea. [68] Coffee, apples, berries, and potatoes are sources of chlorogenic acid, a type of phenolic acid. [69] Models of induced ER stress clearly show that, when administered at low doses, chlorogenic acid, chrysanthemin, and resveratrol exert a protective effect by dose-dependently downregulating the expression of molecules associated with UPR signaling or apoptosis. In a pathological context, this protective function may slow disease progression and alleviate symptoms. As shown in Table 1, high doses of resveratrol, on the other hand, induce endoplasmic reticulum stress and activate apoptotic mechanisms, especially in tumor cell lines, which may be therapeutically advantageous. Thus, resveratrol exerts a complex dual effect on ER stress. Some studies further show that combining resveratrol with other chemicals stimulates ER stress in tumor cells, even at lower concentrations. [41,45,49] A study by Wang et al. (2011) reported a paradoxical finding: in multiple myeloma cell lines resveratrol activated the unfolded protein response (UPR) via the IRE1 signaling cascade, promoting XBP1 splicing, while simultaneously suppressing the activity of XBP1s as a transcriptional factor through SIRT1. This suggests that resveratrol may stimulate stress while hindering the cell’s ability to adapt. [37] In another study, low doses of resveratrol were shown to reduce the levels of p-PERK and ATF4, but activated the ATF6 pathway. This suggests that apoptosis-promoting pathways are supressed, while activation of ATF6 may play a protective role, as it is responsible for the production of chaperones and ERAD factors. At high doses, the effect was reversed. [31]
The interpretation of available data has several limitations:
- insufficient information on the effects of some polyphenols across all UPR branches.
- heterogeneity in experimental models limits direct comparisons and generalization.
- dose variation and dual effect of resveratrol on ER stress – determining the precise therapeutic dose and anticipating potential side effects is challenging.
- lack of clinical trials and data on long-term effects of polyphenol intake – it is difficult to determine the therapeutic potential of polyphenols, as well as to assess the long-term safety and efficacy of these compounds.
- although the results highlight the significant contribution of the polyphenols found at the highest concentrations in the aqueous dwarf elder extract, the possible biological activity of other present phytochemicals in lower amounts cannot be excluded.
Other future research should focus on pro-apoptotic effects of other polyphenols, given that high dose resveratrol has been found to have the same effect in cancer cell lines due to the malfunction of ER. More studies investigating tissue-specific and dose-dependent effects of individual polyphenols or combinations in different disease models are essential. Clinical trials and long-term safety studies are also needed to determine their potential as therapeutic agents.
5. Conclusions
The review identifies a possible link between the most represented polyphenols in the dwarf elder fruit aqueous extract and the modulation of ER stress. Among these, only four of the most abundant polyphenols – resveratrol, chrysanthemin, chlorogenic acid, and epicatechin – have been found to influence UPR pathways, with a predominant effect on the PERK/eIF2α/ATF4/CHOP pathway. The data indicate that these phytochemicals reduce ER stress markers in a dose-dependent manner across various in vivo and in vitro experimental models. Notably, resveratrol is the only compound among them that can amplify ER stress and provoke programmed cell death at high doses. This dual role underscores the potential of dwarf elderberry extract as a remedy for conditions associated with chronic cellular stress. Nevertheless, further research is essential to elucidate the molecular mechanisms by which phytochemicals act, determine their optimal dosages, identify potential side effects, and explore their synergistic interactions with other phytochemicals and drugs.
Author Contributions
Writing—original draft, methodology, review and editing S.S. and M.B.; writing— supervision, conceptualization, review and editing O.T. and G.Y. All authors have read and agreed to the published version of the manuscript.
Funding
The European Union-Next Generation EU, through the National Recovery and Resilience Plan of the Republic of Bulgaria, project No. BG-RRP-2.004-0009-C02, supported this work.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schwarz, D.S.; Blower, M.D. The Endoplasmic Reticulum: Structure, Function and Response to Cellular Signaling. Cell Mol Life Sci 2016, 73, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, L.; Tabas, I. Role of Endoplasmic Reticulum Stress in Metabolic Disease and Other Disorders. Annu Rev Med 2012, 63, 317. [Google Scholar] [CrossRef]
- Lin, J.H.; Walter, P.; Yen, T.S.B. Endoplasmic Reticulum Stress in Disease Pathogenesis. Annu Rev Pathol 2008, 3, 399–425. [Google Scholar] [CrossRef]
- Ron, D.; Walter, P. Signal Integration in the Endoplasmic Reticulum Unfolded Protein Response. Nat Rev Mol Cell Biol 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Hasnain, S.Z.; Lourie, R.; Das, I.; Chen, A.C.; McGuckin, M.A. The Interplay between Endoplasmic Reticulum Stress and Inflammation. Immunol Cell Biol 2012, 90, 260–270. [Google Scholar] [CrossRef]
- Sprenkle, N.T.; Sims, S.G.; Sánchez, C.L.; Meares, G.P. Endoplasmic Reticulum Stress and Inflammation in the Central Nervous System. Mol Neurodegener 2017, 12, 42. [Google Scholar] [CrossRef]
- So, A.Y.-L.; de la Fuente, E.; Walter, P.; Shuman, M.; Bernales, S. The Unfolded Protein Response during Prostate Cancer Development. Cancer and Metastasis Reviews 2009, 28, 219–223. [Google Scholar] [CrossRef]
- Lee, A.S. GRP78 Induction in Cancer: Therapeutic and Prognostic Implications. Cancer Res 2007, 67, 3496–3499. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, P.M.; Tabbara, S.O.; Jacobs, L.K.; Manning, F.C.R.; Tsangaris, T.N.; Schwartz, A.M.; Kennedy, K.A.; Patierno, S.R. Overexpression of the Glucose-Regulated Stress Gene GRP78 in Malignant but Not Benign Human Breast Lesions. Breast Cancer Res Treat 2000, 59, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, D.R.; Sukhdeo, K.; Protopopova, M.; Sinha, R.; Enos, M.; Carrasco, D.E.; Zheng, M.; Mani, M.; Henderson, J.; Pinkus, G.S.; et al. The Differentiation and Stress Response Factor XBP-1 Drives Multiple Myeloma Pathogenesis. Cancer Cell 2007, 11, 349–360. [Google Scholar] [CrossRef]
- Tardif, K.D.; Mori, K.; Kaufman, R.J.; Siddiqui, A. Hepatitis C Virus Suppresses the IRE1-XBP1 Pathway of the Unfolded Protein Response. Journal of Biological Chemistry 2004, 279, 17158–17164. [Google Scholar] [CrossRef]
- Xu, Z.; Jensen, G.; Yen, T.S. Activation of Hepatitis B Virus S Promoter by the Viral Large Surface Protein via Induction of Stress in the Endoplasmic Reticulum. J Virol 1997, 71, 7387–7392. [Google Scholar] [CrossRef]
- Ji, C.; Mehrian-Shai, R.; Chan, C.; Hsu, Y.-H.; Kaplowitz, N. Role of CHOP in Hepatic Apoptosis in the Murine Model of Intragastric Ethanol Feeding. Alcohol Clin Exp Res 2005, 29, 1496–1503. [Google Scholar] [CrossRef]
- Kammoun, H.L.; Chabanon, H.; Hainault, I.; Luquet, S.; Magnan, C.; Koike, T.; Ferré, P.; Foufelle, F. GRP78 Expression Inhibits Insulin and ER Stress–Induced SREBP-1c Activation and Reduces Hepatic Steatosis in Mice. Journal of Clinical Investigation 2009, 119, 1201–1215. [Google Scholar] [CrossRef]
- Özcan, U.; Cao, Q.; Yilmaz, E.; Lee, A.-H.; Iwakoshi, N.N.; Özdelen, E.; Tuncman, G.; Görgün, C.; Glimcher, L.H.; Hotamisligil, G.S. Endoplasmic Reticulum Stress Links Obesity, Insulin Action, and Type 2 Diabetes. Science (1979) 2004, 306, 457–461. [Google Scholar] [CrossRef]
- Tsukano, H.; Gotoh, T.; Endo, M.; Miyata, K.; Tazume, H.; Kadomatsu, T.; Yano, M.; Iwawaki, T.; Kohno, K.; Araki, K.; et al. The Endoplasmic Reticulum Stress-C/EBP Homologous Protein Pathway-Mediated Apoptosis in Macrophages Contributes to the Instability of Atherosclerotic Plaques. Arterioscler Thromb Vasc Biol 2010, 30, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Imaizumi, K.; Sato, N.; Miyoshi, K.; Kudo, T.; Hitomi, J.; Morihara, T.; Yoneda, T.; Gomi, F.; Mori, Y.; et al. Presenilin-1 Mutations Downregulate the Signalling Pathway of the Unfolded-Protein Response. Nat Cell Biol 1999, 1, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Soda, M.; Takahashi, R. Parkin Suppresses Unfolded Protein Stress-Induced Cell Death through Its E3 Ubiquitin-Protein Ligase Activity. Journal of Biological Chemistry 2000, 275, 35661–35664. [Google Scholar] [CrossRef]
- Saxena, S.; Cabuy, E.; Caroni, P. A Role for Motoneuron Subtype–Selective ER Stress in Disease Manifestations of FALS Mice. Nat Neurosci 2009, 12, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid Med Cell Longev 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- M. Shokrzadeh; S. S. Saeedi Saravi The Chemistry, Pharmacology and Clinical Properties of Sambucus Ebulus: A Review. Journal of Medicinal Plants Research 2010, 4, 95–103. [CrossRef]
- Jabbari, M.; Daneshfard, B.; Emtiazy, M.; Khiveh, A.; Hashempur, M.H. Biological Effects and Clinical Applications of Dwarf Elder ( Sambucus Ebulus L): A Review. J Evid Based Complementary Altern Med 2017, 22, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Tasinov, O.; Kiselova-Kaneva, Y.; Ivanova, D. Sambucus Ebulus - from Traditional Medicine to Recent Studies. Scripta Scientifica Medica 2013, 45, 36. [Google Scholar] [CrossRef]
- Tasinov, O.; Dincheva, I.; Badjakov, I.; Kiselova-Kaneva, Y.; Galunska, B.; Nogueiras, R.; Ivanova, D. Phytochemical Composition, Anti-Inflammatory and ER Stress-Reducing Potential of Sambucus Ebulus L. Fruit Extract. Plants 2021, 10, 2446. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, S.; Giles, A.; Nakamura, K.; Lee, J.W.; Hou, X.; Donmez, G.; Li, J.; Luo, Z.; Walsh, K.; et al. Hepatic Overexpression of SIRT1 in Mice Attenuates Endoplasmic Reticulum Stress and Insulin Resistance in the Liver. The FASEB Journal 2011, 25, 1664–1679. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, X.; Xie, W.; Li, Y.; Ma, M.; Yuan, T.; Luo, B. Resveratrol Exerts an Anti-Apoptotic Effect on Human Bronchial Epithelial Cells Undergoing Cigarette Smoke Exposure. Mol Med Rep 2015, 11, 1752–1758. [Google Scholar] [CrossRef]
- Lin, Y.; Zhu, J.; Zhang, X.; Wang, J.; Xiao, W.; Li, B.; Jin, L.; Lian, J.; Zhou, L.; Liu, J. Inhibition of Cardiomyocytes Hypertrophy by Resveratrol Is Associated with Amelioration of Endoplasmic Reticulum Stress. Cellular Physiology and Biochemistry 2016, 39, 780–789. [Google Scholar] [CrossRef]
- Cheng, J.; Xia, X.; Rui, Y.; Zhang, Z.; Qin, L.; Han, S.; Wan, Z. The Combination of 1α,25dihydroxyvitaminD3 with Resveratrol Improves Neuronal Degeneration by Regulating Endoplasmic Reticulum Stress, Insulin Signaling and Inhibiting Tau Hyperphosphorylation in SH-SY5Y Cells. Food and Chemical Toxicology 2016, 93, 32–40. [Google Scholar] [CrossRef]
- Yan, W.-J.; Liu, R.-B.; Wang, L.-K.; Ma, Y.-B.; Ding, S.-L.; Deng, F.; Hu, Z.-Y.; Wang, D.-B. Sirt3-Mediated Autophagy Contributes to Resveratrol-Induced Protection against ER Stress in HT22 Cells. Front Neurosci 2018, 12, 116. [Google Scholar] [CrossRef]
- Lee, J.; Hong, S.-W.; Kwon, H.; Park, S.E.; Rhee, E.-J.; Park, C.-Y.; Oh, K.-W.; Park, S.-W.; Lee, W.-Y. Resveratrol, an Activator of SIRT1, Improves ER Stress by Increasing Clusterin Expression in HepG2 Cells. Cell Stress Chaperones 2019, 24, 825–833. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, Y.; Shu, L.; Song, G.; Ma, H. Resveratrol Reduces Liver Endoplasmic Reticulum Stress and Improves Insulin Sensitivity in Vivo and in Vitro. Drug Des Devel Ther 2019, Volume 13, 1473–1485. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, X.; Ding, M.; You, C.; Lin, X.; Wang, Y.; Wu, M.; Xu, G.; Wang, G. Resveratrol Decreases High Glucose induced Apoptosis in Renal Tubular Cells via Suppressing Endoplasmic Reticulum Stress. Mol Med Rep 2020, 22, 4367–4375. [Google Scholar] [CrossRef]
- Neal, S.E.; Buehne, K.L.; Besley, N.A.; Yang, P.; Silinski, P.; Hong, J.; Ryde, I.T.; Meyer, J.N.; Jaffe, G.J. Resveratrol Protects Against Hydroquinone-Induced Oxidative Threat in Retinal Pigment Epithelial Cells. Investigative Opthalmology & Visual Science 2020, 61, 32. [Google Scholar] [CrossRef]
- Yu, X.; Xu, X.; Dong, W.; Yang, C.; Luo, Y.; He, Y.; Jiang, C.; Wu, Y.; Wang, J. DDIT3/CHOP Mediates the Inhibitory Effect of ER Stress on Chondrocyte Differentiation by AMPKα-SIRT1 Pathway. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 2022, 1869, 119265. [Google Scholar] [CrossRef]
- Luo, Y.; Hu, N.; Zhao, Y.; Lai, J.; Luo, X.; Liu, J. Resveratrol mediated Activation of SIRT1 Inhibits the PERK eIF2α ATF4 Pathway and Mitigates Bupivacaine induced Neurotoxicity in PC12 Cells. Exp Ther Med 2023, 26, 433. [Google Scholar] [CrossRef]
- Chinta, S.J.; Poksay, K.S.; Kaundinya, G.; Hart, M.; Bredesen, D.E.; Andersen, J.K.; Rao, R. V. Endoplasmic Reticulum Stress–Induced Cell Death in Dopaminergic Cells: Effect of Resveratrol. Journal of Molecular Neuroscience 2009, 39, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.-M.; Galson, D.L.; Roodman, G.D.; Ouyang, H. Resveratrol Triggers the Pro-Apoptotic Endoplasmic Reticulum Stress Response and Represses pro-Survival XBP1 Signaling in Human Multiple Myeloma Cells. Exp Hematol 2011, 39, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.; Pan-Castillo, B.; Valls, C.; Pujadas, G.; Garcia-Vallve, S.; Arola, L.; Mulero, M. Resveratrol Enhances Palmitate-Induced ER Stress and Apoptosis in Cancer Cells. PLoS One 2014, 9, e113929. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, S.; Sun, Y.; Sukumaran, P.; Singh, B.B. Resveratrol Activates Autophagic Cell Death in Prostate Cancer Cells via Downregulation of STIM1 and the MTOR Pathway. Mol Carcinog 2016, 55, 818–831. [Google Scholar] [CrossRef]
- Heo, J.; Kim, S.; Hwang, K.; Kang, J.; Choi, K. Resveratrol Induced Reactive Oxygen Species and Endoplasmic Reticulum Stress mediated Apoptosis, and Cell Cycle Arrest in the A375SM Malignant Melanoma Cell Line. Int J Mol Med 2018, 42, 1427–1435. [Google Scholar] [CrossRef]
- Ren, M.; Zhou, X.; Gu, M.; Jiao, W.; Yu, M.; Wang, Y.; Liu, S.; Yang, J.; Ji, F. Resveratrol Synergizes with Cisplatin in Antineoplastic Effects against AGS Gastric Cancer Cells by Inducing Endoplasmic Reticulum Stress mediated Apoptosis and G2/M Phase Arrest. Oncol Rep 2020, 44, 1605–1615. [Google Scholar] [CrossRef]
- Pan, P.; Lin, H.; Chuang, C.; Wang, P.; Wan, H.; Lee, M.; Kao, M. Resveratrol Alleviates Nuclear Factor-κB-mediated Neuroinflammation in Vasculitic Peripheral Neuropathy Induced by Ischaemia–Reperfusion via Suppressing Endoplasmic Reticulum Stress. Clin Exp Pharmacol Physiol 2019, 46, 770–779. [Google Scholar] [CrossRef]
- Wang, B.; Ge, S.; Xiong, W.; Xue, Z. Effects of Resveratrol Pretreatment on Endoplasmic Reticulum Stress and Cognitive Function after Surgery in Aged Mice. BMC Anesthesiol 2018, 18, 141. [Google Scholar] [CrossRef]
- Pan, Q.-R.; Ren, Y.-L.; Liu, W.-X.; Hu, Y.-J.; Zheng, J.-S.; Xu, Y.; Wang, G. Resveratrol Prevents Hepatic Steatosis and Endoplasmic Reticulum Stress and Regulates the Expression of Genes Involved in Lipid Metabolism, Insulin Resistance, and Inflammation in Rats. Nutrition Research 2015, 35, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.M.; Hernandez, F.; Puerta, N.; De Angulo, G.; Webster, K.A.; Vanni, S. Resveratrol Augments ER Stress and the Cytotoxic Effects of Glycolytic Inhibition in Neuroblastoma by Downregulating Akt in a Mechanism Independent of SIRT1. Exp Mol Med 2016, 48, e210–e210. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Jiang, J.; Zhang, G.; Bu, Y.; Zhang, G.; Zhao, X. Resveratrol and Caloric Restriction Prevent Hepatic Steatosis by Regulating SIRT1-Autophagy Pathway and Alleviating Endoplasmic Reticulum Stress in High-Fat Diet-Fed Rats. PLoS One 2017, 12, e0183541. [Google Scholar] [CrossRef]
- Ardid-Ruiz, A.; Ibars, M.; Mena, P.; Del Rio, D.; Muguerza, B.; Bladé, C.; Arola, L.; Aragonès, G.; Suárez, M. Potential Involvement of Peripheral Leptin/STAT3 Signaling in the Effects of Resveratrol and Its Metabolites on Reducing Body Fat Accumulation. Nutrients 2018, 10, 1757. [Google Scholar] [CrossRef]
- Lu, J.; Zheng, Y.; Yang, J.; Zhang, J.; Cao, W.; Chen, X.; Fang, S. Resveratrol Alleviates Inflammatory Injury and Enhances the Apoptosis of Fibroblast like Synoviocytes via Mitochondrial Dysfunction and ER Stress in Rats with Adjuvant Arthritis. Mol Med Rep 2019, 20, 463–472. [Google Scholar] [CrossRef]
- Arena, A.; Romeo, M.A.; Benedetti, R.; Masuelli, L.; Bei, R.; Gilardini Montani, M.S.; Cirone, M. New Insights into Curcumin- and Resveratrol-Mediated Anti-Cancer Effects. Pharmaceuticals 2021, 14, 1068. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.T.; Veerisetty, A.C.; Wu, J.; Coustry, F.; Hossain, M.G.; Chiu, F.; Gannon, F.H.; Posey, K.L. Primary Osteoarthritis Early Joint Degeneration Induced by Endoplasmic Reticulum Stress Is Mitigated by Resveratrol. Am J Pathol 2021, 191, 1624–1637. [Google Scholar] [CrossRef]
- Totonchi, H.; Mokarram, P.; Karima, S.; Rezaei, R.; Dastghaib, S.; Koohpeyma, F.; Noori, S.; Azarpira, N. Resveratrol Promotes Liver Cell Survival in Mice Liver-Induced Ischemia-Reperfusion through Unfolded Protein Response: A Possible Approach in Liver Transplantation. BMC Pharmacol Toxicol 2022, 23, 74. [Google Scholar] [CrossRef]
- Bettaieb, A.; Vazquez Prieto, M.A.; Rodriguez Lanzi, C.; Miatello, R.M.; Haj, F.G.; Fraga, C.G.; Oteiza, P.I. (−)-Epicatechin Mitigates High-Fructose-Associated Insulin Resistance by Modulating Redox Signaling and Endoplasmic Reticulum Stress. Free Radic Biol Med 2014, 72, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Lee, J.-H.; Seo, Y.H.; Jang, J.-H.; Jeong, C.-H.; Lee, S.; Jeong, G.-S.; Park, B. Epicatechin Prevents Methamphetamine-Induced Neuronal Cell Death via Inhibition of ER Stress. Biomol Ther (Seoul) 2019, 27, 145–151. [Google Scholar] [CrossRef]
- Ye, H.-Y.; Li, Z.-Y.; Zheng, Y.; Chen, Y.; Zhou, Z.-H.; Jin, J. The Attenuation of Chlorogenic Acid on Oxidative Stress for Renal Injury in Streptozotocin-Induced Diabetic Nephropathy Rats. Arch Pharm Res 2016, 39, 989–997. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Dong, J.; Nie, J.; Zhu, J.-X.; Wang, H.; Chen, Q.; Chen, J.-Y.; Xia, J.-M.; Shuai, W. Amelioration of Bleomycin-Induced Pulmonary Fibrosis by Chlorogenic Acid through Endoplasmic Reticulum Stress Inhibition. Apoptosis 2017, 22, 1147–1156. [Google Scholar] [CrossRef]
- Zhang, Y.; Miao, L.; Zhang, H.; Wu, G.; Zhang, Z.; Lv, J. Chlorogenic Acid against Palmitic Acid in Endoplasmic Reticulum Stress-Mediated Apoptosis Resulting in Protective Effect of Primary Rat Hepatocytes. Lipids Health Dis 2018, 17, 270. [Google Scholar] [CrossRef]
- Kazaz, I.O.; Demir, S.; Kerimoglu, G.; Colak, F.; Turkmen Alemdar, N.; Yilmaz Dogan, S.; Bostan, S.; Mentese, A. Chlorogenic Acid Ameliorates Torsion/Detorsion-Induced Testicular Injury via Decreasing Endoplasmic Reticulum Stress. J Pediatr Urol 2022, 18, 289. [Google Scholar] [CrossRef]
- Preetha Rani, M.R.; Salin Raj, P.; Nair, A.; Ranjith, S.; Rajankutty, K.; Raghu, K.G. In Vitro and in Vivo Studies Reveal the Beneficial Effects of Chlorogenic Acid against ER Stress Mediated ER-Phagy and Associated Apoptosis in the Heart of Diabetic Rat. Chem Biol Interact 2022, 351, 109755. [Google Scholar] [CrossRef] [PubMed]
- Sari, I.; Moch Rizal, D.; Afiyah Syarif, R. The Effect of Chlorogenic Acid on Endoplasmic Reticulum Stress and Steroidogenesis in the Testes of Diabetic Rats: Study of MRNA Expressions of GRP78, XBP1s, 3β-HSD, and 17β-HSD. BIO Web Conf 2022, 49, 01001. [Google Scholar] [CrossRef]
- Azam Moslehi; Tahereh Komeili-Movahhed; Mostafa Ahmadian; Mahdieh Ghoddoosi; Fatemeh Heidari Chlorogenic Acid Attenuates Liver Apoptosis and Inflammation in Endoplasmic Reticulum Stress-Induced Mice. Iran J Basic Med Sci. 2023, 26, 478–485. [CrossRef]
- Boonyong, C.; Angkhasirisap, W.; Kengkoom, K.; Jianmongkol, S. Different Protective Capability of Chlorogenic Acid and Quercetin against Indomethacin-Induced Gastrointestinal Ulceration. Journal of Pharmacy and Pharmacology 2023, 75, 427–436. [Google Scholar] [CrossRef]
- Ping, P.; Yang, T.; Ning, C.; Zhao, Q.; Zhao, Y.; Yang, T.; Gao, Z.; Fu, S. Chlorogenic Acid Attenuates Cardiac Hypertrophy via Up-regulating Sphingosine-1-phosphate Receptor1 to Inhibit Endoplasmic Reticulum Stress. ESC Heart Fail 2024, 11, 1580–1593. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huang, Y.; Luo, G.; Yang, X.; Huang, W. Cyanidin-3-O-Glucoside Attenuates High Glucose–Induced Podocyte Dysfunction by Inhibiting Apoptosis and Promoting Autophagy via Activation of SIRT1/AMPK Pathway. Can J Physiol Pharmacol 2021, 99, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Gambini, J.; Inglés, M.; Olaso, G.; Lopez-Grueso, R.; Bonet-Costa, V.; Gimeno-Mallench, L.; Mas-Bargues, C.; Abdelaziz, K.M.; Gomez-Cabrera, M.C.; Vina, J.; et al. Properties of Resveratrol: In Vitro and In Vivo Studies about Metabolism, Bioavailability, and Biological Effects in Animal Models and Humans. Oxid Med Cell Longev 2015, 2015, 837042. [Google Scholar] [CrossRef]
- Soleas, G.J.; Diamandis, E.P.; Goldberg, D.M. Resveratrol: A Molecule Whose Time Has Come? And Gone? Clin Biochem 1997, 30, 91–113. [Google Scholar] [CrossRef]
- Cheng, Z.; Si, X.; Tan, H.; Zang, Z.; Tian, J.; Shu, C.; Sun, X.; Li, Z.; Jiang, Q.; Meng, X.; et al. Cyanidin-3-O-Glucoside and Its Phenolic Metabolites Ameliorate Intestinal Diseases via Modulating Intestinal Mucosal Immune System: Potential Mechanisms and Therapeutic Strategies. Crit Rev Food Sci Nutr 2023, 63, 1629–1647. [Google Scholar] [CrossRef]
- Frountzas, M.; Karanikki, E.; Toutouza, O.; Sotirakis, D.; Schizas, D.; Theofilis, P.; Tousoulis, D.; Toutouzas, K.G. Exploring the Impact of Cyanidin-3-Glucoside on Inflammatory Bowel Diseases: Investigating New Mechanisms for Emerging Interventions. Int J Mol Sci 2023, 24, 9399. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Saravanan, T.S.; Monteclaro, C.C.; Presser, N.; Ye, X.; Selvan, S.R.; Brosman, S. Epicatechins Purified from Green Tea (Camellia Sinensis) Differentially Suppress Growth of Gender-Dependent Human Cancer Cell Lines. Evid Based Complement Alternat Med 2006, 3, 237–247. [Google Scholar] [CrossRef]
- Shimoyama, A.T.; Santin, J.R.; Machado, I.D.; de Oliveira e Silva, A.M.; de Melo, I.L.P.; Mancini-Filho, J.; Farsky, S.H.P. Antiulcerogenic Activity of Chlorogenic Acid in Different Models of Gastric Ulcer. Naunyn Schmiedebergs Arch Pharmacol 2013, 386, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kaufman, R.J. From Acute ER Stress to Physiological Roles of the Unfolded Protein Response. Cell Death Differ 2006, 13, 374–384. [Google Scholar] [CrossRef]
- Siwecka, N.; Rozpędek-Kamińska, W.; Wawrzynkiewicz, A.; Pytel, D.; Diehl, J.A.; Majsterek, I. The Structure, Activation and Signaling of IRE1 and Its Role in Determining Cell Fate. Biomedicines 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, D.T.; Kaufman, R.J. A Trip to the ER: Coping with Stress. Trends Cell Biol 2004, 14, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Ron, D. Integrating the Mechanisms of Apoptosis Induced by Endoplasmic Reticulum Stress. Nat Cell Biol 2011, 13, 184–190. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Unfolded protein response (UPR) pathways. 1) IRE1 (Inositol-requiring enzyme 1) signaling cascade – IRE1 is associated with the BiP chaperone and remains inactive under normal conditions. Upon accumulation of misfolded proteins, BiP dissociates from the receptor and binds to these proteins. The now-activated IRE1 dimerizes, undergoes autophosphorylation, and initiates splicing of XBP1 (X-box binding protein 1) mRNA by cutting away an intron. This generates XBP1s mRNA, which encodes the transcription factor XBP1s, which translocates into the nucleus and regulates the activity of important genes encoding chaperones, ERAD (endoplasmic reticulum-associated degradation) components, enzymes involved in lipid biosynthesis and oxidative phosphorylation. Additionally, IRE1 degrades selected mRNAs through a mechanism known as regulated IRE1-dependent decay (RIDD) to lower protein production load. Under conditions of severe ER stress, activated IRE1 binds to TRAF2 (TNF receptor-associated factor 2), which activates ASK1 (apoptosis signal-regulating kinase 1). ASK1 then phosphorylates JNK (c-Jun N-terminal kinase), leading to programmed cell death and regulation of genes involved in inflammation, differentiation, and redox homeostasis. Moreover, TRAF2 activates IκB kinase (IKK), which phosphorylates IκB (inhibitor of NF-kB), resulting in the release of NF-κB. NF-κB translocates to the nucleus, where it regulates the expression of genes related to inflammation, immune response, and cell proliferation. [2,4,70,71] 2) ATF6 (Activating Transcription Factor 6) signaling cascade – when misfolded proteins accumulate, BiP dissociates from ATF6 receptor, allowing ATF6 to translocate to the Golgi apparatus. There, it is cleaved by the proteases S1P (site-1 protease) and S2P (site-2 protease), producing its active form as a transcription factor. ATF6 then enters the nucleus and promotes the expression of genes encoding chaperones, ERAD components, and XBP1. [4,72] 3) PERK (Protein kinase RNA-like ER kinase) signaling cascade – under ER stress BiP is released from PERK, leading to its autophosphorylation and activation. Activated PERK phosphorylates the α subunit of eukaryotic initiation factor 2 (eIF2α), which supresses general protein synthesis while promoting the selective translation of ATF4. ATF4 leads to increased expression of genes involved in chaperone production, antioxidant response, amino acid transporters, and proteins necessary for autophagy. If the stress is too severe, ATF4 induces the expression of C/EBP-homologous protein (CHOP), also known as GADD153, a pro-apoptotic factor [2,4,73]. Schematic generated using BioRender.
Figure 1.
Unfolded protein response (UPR) pathways. 1) IRE1 (Inositol-requiring enzyme 1) signaling cascade – IRE1 is associated with the BiP chaperone and remains inactive under normal conditions. Upon accumulation of misfolded proteins, BiP dissociates from the receptor and binds to these proteins. The now-activated IRE1 dimerizes, undergoes autophosphorylation, and initiates splicing of XBP1 (X-box binding protein 1) mRNA by cutting away an intron. This generates XBP1s mRNA, which encodes the transcription factor XBP1s, which translocates into the nucleus and regulates the activity of important genes encoding chaperones, ERAD (endoplasmic reticulum-associated degradation) components, enzymes involved in lipid biosynthesis and oxidative phosphorylation. Additionally, IRE1 degrades selected mRNAs through a mechanism known as regulated IRE1-dependent decay (RIDD) to lower protein production load. Under conditions of severe ER stress, activated IRE1 binds to TRAF2 (TNF receptor-associated factor 2), which activates ASK1 (apoptosis signal-regulating kinase 1). ASK1 then phosphorylates JNK (c-Jun N-terminal kinase), leading to programmed cell death and regulation of genes involved in inflammation, differentiation, and redox homeostasis. Moreover, TRAF2 activates IκB kinase (IKK), which phosphorylates IκB (inhibitor of NF-kB), resulting in the release of NF-κB. NF-κB translocates to the nucleus, where it regulates the expression of genes related to inflammation, immune response, and cell proliferation. [2,4,70,71] 2) ATF6 (Activating Transcription Factor 6) signaling cascade – when misfolded proteins accumulate, BiP dissociates from ATF6 receptor, allowing ATF6 to translocate to the Golgi apparatus. There, it is cleaved by the proteases S1P (site-1 protease) and S2P (site-2 protease), producing its active form as a transcription factor. ATF6 then enters the nucleus and promotes the expression of genes encoding chaperones, ERAD components, and XBP1. [4,72] 3) PERK (Protein kinase RNA-like ER kinase) signaling cascade – under ER stress BiP is released from PERK, leading to its autophosphorylation and activation. Activated PERK phosphorylates the α subunit of eukaryotic initiation factor 2 (eIF2α), which supresses general protein synthesis while promoting the selective translation of ATF4. ATF4 leads to increased expression of genes involved in chaperone production, antioxidant response, amino acid transporters, and proteins necessary for autophagy. If the stress is too severe, ATF4 induces the expression of C/EBP-homologous protein (CHOP), also known as GADD153, a pro-apoptotic factor [2,4,73]. Schematic generated using BioRender.
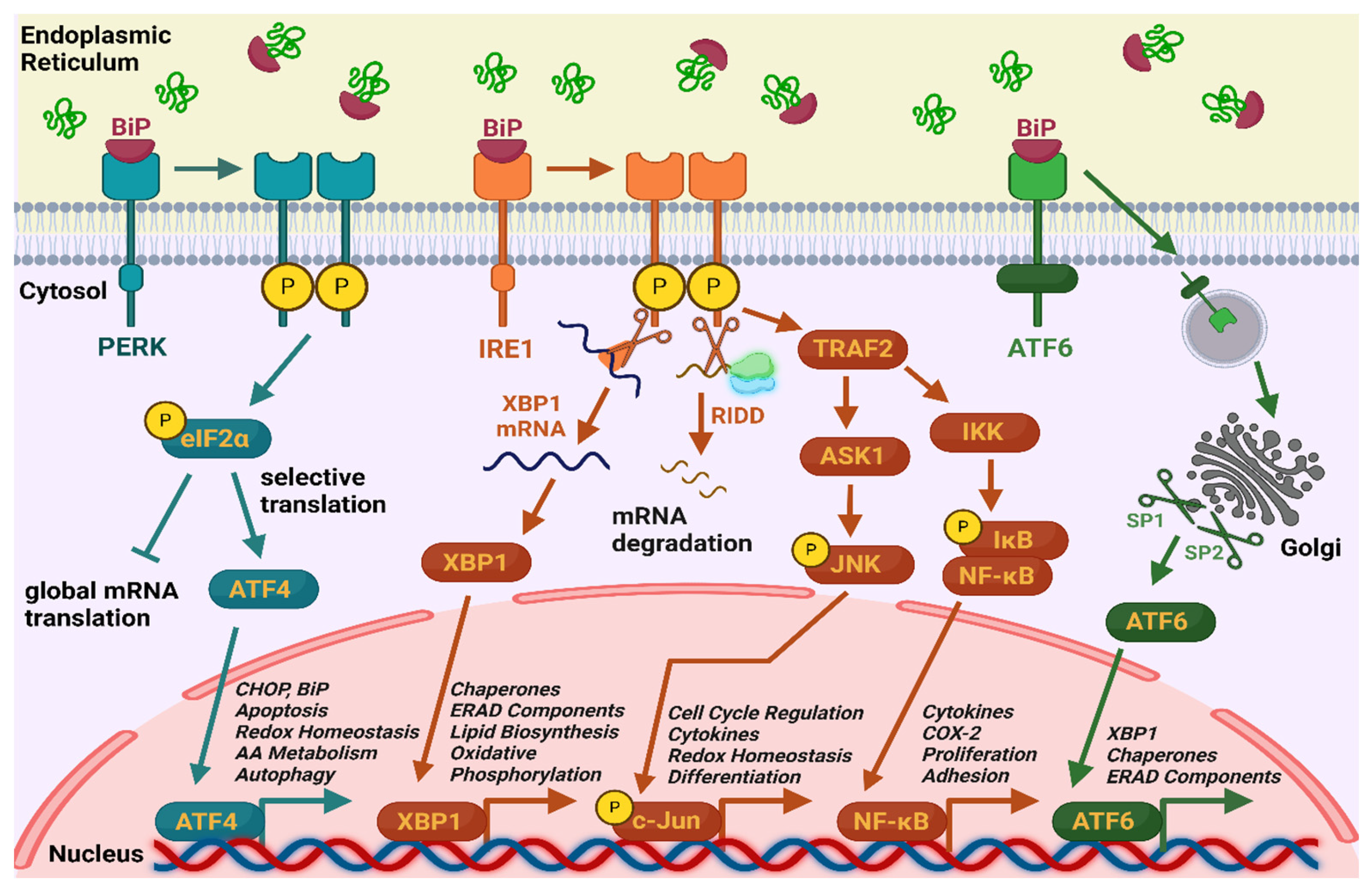
Figure 2.
PRISMA flow diagram of the study selection process.
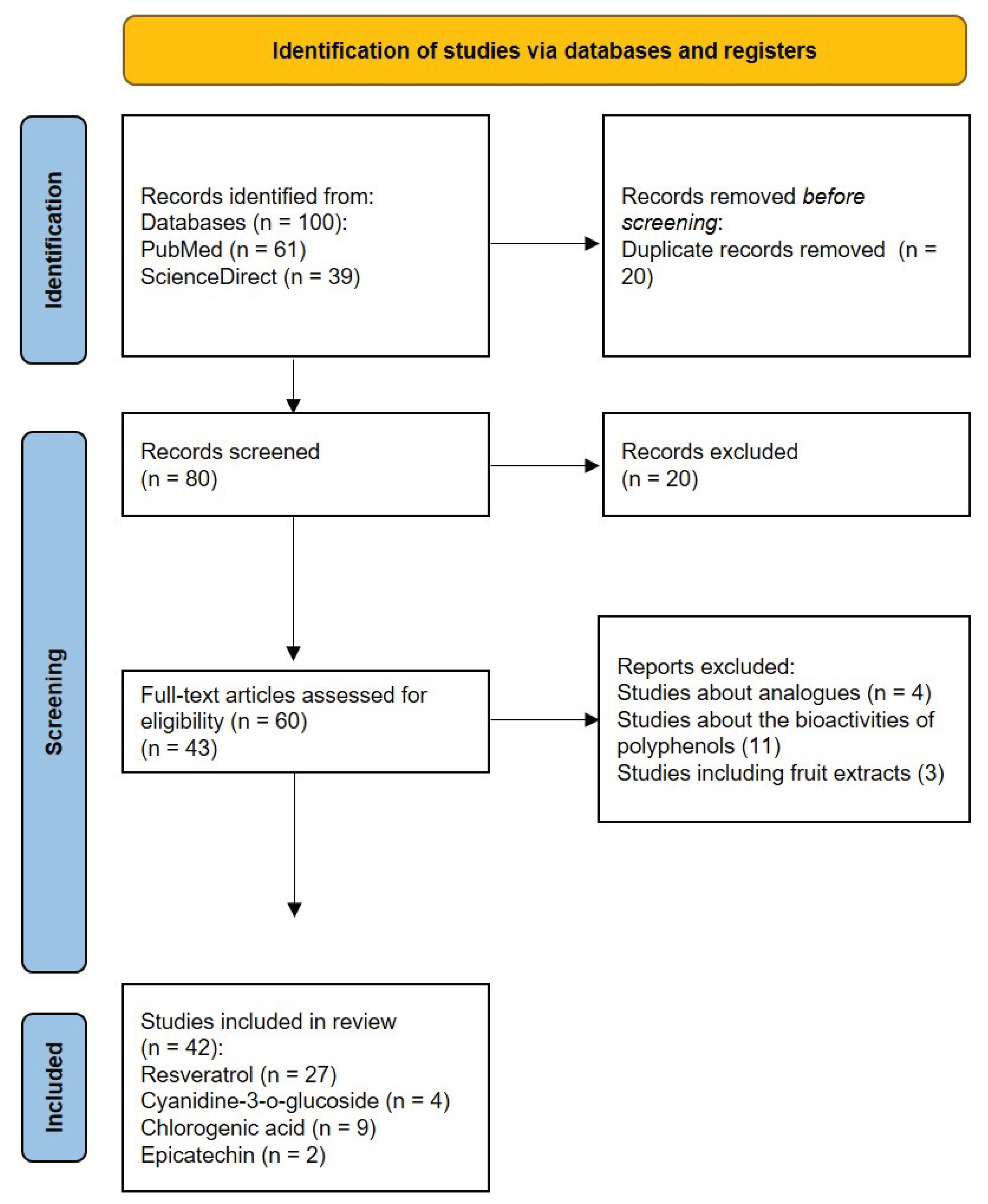

Table 1.
Modulation of ER stress markers by resveratrol.
| Author(s) | Year | Experimental Model | Altered Expression or Activity of ER Stress Markers | Dosage |
|---|---|---|---|---|
| Chinta et al. [36] | 2009 | In vitro (dopaminergic N27 cells) | ↑cleavage of caspases 7 and 3 ↑GRP78 expression ↑GRP94 expression ↑CHOP expression ↑p-eIF2α expression |
50-250 μM |
| Wang et al. [37] | 2011 | In vitro (multiple myeloma cell lines ANBL-6, OPM2, MM.15) | ↑JNK phosphorylation ↑CHOP expression ↑XBP1 mRNA splicing ↓transcription of XBP1s |
100 μM |
| Li et al. [25] | 2011 | In vitro (tunicamycin-induced ER stress in HepG2 cells) | ↓XBP1 mRNA splicing ↓GRP78 expression ↓CHOP expression |
10 μM |
| Rojas et al. [38] | 2014 | In vitro (palmitate-induced ER stress in HepG2 cells) | ↑XBP1 mRNA splicing ↑CHOP expression |
100 μM |
| Zhang et al. [26] | 2015 | In vitro (cigarette smoke extract-induced apoptosis in cultured human bronchial epithelial cells) | ↓CHOP expression ↓caspases 3 and 4 expression |
20 µmol/l |
| Pan et al. [44] | 2015 | In vivo (high-fat diet-fed rats) | ↓ATF4 expression ↓GRP78 expression ↓CHOP expression ↓GRP78 expression ↓p-PERK expression |
100 mg/kg |
| Graham et al. [45] | 2016 | In vitro (2-deoxy-D-glucose inhibition of glycolysis in neuroblastoma cells) | ↑CHOP expression ↓GRP78 expression ↓GRP94 expression |
10 μM |
| Lin et al. [27] | 2016 | In vitro (neonatal rat cardiomyocytes) | ↓GRP78 expression ↓GRP94 expression ↓CHOP expression |
50 μM |
| Cheng et al. [28] | 2016 | In vitro (tunicamycin and Aβ25-35 induced ER stress in SH-SYSY cells) | ↓GRP78 expression ↓CHOP expression ↓p-eIF2α expression |
25 μM |
| Selvaraj et al. [39] | 2016 | In vitro (PC3 and DU145 prostate cancer cell lines) | ↑CHOP expression | 100 μM |
| Ding et al. [46] | 2017 | In vivo (high-fat diet-fed rats) | ↓GRP78 expression ↓CHOP expression |
200 mg/kg |
| Yan et al. [29] | 2018 | In vitro (tunicamycin-induced ER stress in neuronal HT22 cells) | ↓GRP78 expression ↓CHOP expression ↓caspase 12 expression |
50 μM |
| Heo et al. [40] | 2018 | In vitro (A375SM melanoma cells) | ↑p-eIF2α expression ↑CHOP expression |
10 μM |
| Ardid-Ruiz et al. [47] | 2018 | In vivo (diet induced obesity in rats) | ↓XBP1s expression | 200 mg/kg |
| Wang et al. [43] | 2018 | In vivo (surgical mice model) | ↓GRP78 expression ↓XBP1 expression ↓PERK expression ↓IRE1 expression |
100 mg/kg |
| Lee et al. [30] | 2019 | In vitro (tunicamycin-induced ER stress in HepG2 cells) | ↓PERK expression ↓IRE1 expression ↓CHOP expression ↑ERAD factors expression |
10, 50 and 100 μM |
| Zhao et al. [31] | 2019 | In vivo (high-fat diet-fed mice) In vitro (palmitic acid-induced insulin-resistant HepG2 cells) |
in vivo model – ↓p-PERK expression and ↓ATF4 expression in vitro model – ↑p-PERK expression, ↑ATF4 expression, ↓ATF6 expression (at 50 and 100 μM) and ↓ p-PERK expression, ↓ATF4 expression, ↑ATF6 expression (at 20 μM) | 60 mg/kg (in vivo model) 20, 50 and 100 μM (in vitro model) |
| Lu et al. [48] | 2019 | In vitro (fibroblast-like synoviocytes treated with H2O2) | ↑CHOP expression ↑caspase 12 and caspase 3 expression |
50, 100, 200 and 400 μM |
| Pan et al. [42] | 2019 | In vivo (induced vasculitic peripheral neuropathy by ischaemia–reperfusion in rats) | ↓p-PERK expression ↓p-IRE1 expression ↓ATF6 expression |
20 and 40 mg/кg |
| Zhang et al. [32] | 2020 | In vivo (db/db mice) In vitro (high glucose induced ER stress in NRK-52E cells) |
↓GRP78 expression ↓CHOP expression ↓caspase 12 expression |
20 μM (in vitro model) 40 mg/ kg (in vivo model) |
| Ren et al. [41] | 2020 | In vitro (AGS stomach cancer cell line) | ↑GRP78 expression ↑p-eIF2α expression ↑CHOP expression |
20 μM |
| Neal et al. [33] | 2020 | In vitro (retinal pigment cells treated with hydroquinone) | ↑XBP1 expression ↑CHOP expression |
15 and 30 μM |
| Arena et al. [49] | 2021 | In vitro (Her-2 positive breast cancer and salivary gland cancer cell lines) | ↑CHOP expression | 15 μM |
| Hecht et al. [50] | 2021 | In vivo (model of primary osteoarthritis in mice) | ↓CHOP expression | 0.25 g/L |
| Yu et al. [34] | 2022 | In vitro (tunicamycin-induced ER stress in chondrocytes) | ↓CHOP expression | 50 μM |
| Totonchi еt al. [51] | 2022 | In vivo (mice liver-induced ischemia-reperfusion) | ↓GRP78 expression ↓PERK expression ↓IRE1α expression ↓CHOP expression ↓XBP1 expression |
0.02 and 0.2 mg/kg |
| Luo et al. [35] | 2023 | In vitro (bupivacaine-induced cytotoxicity in PC12 rat adrenal pheochromocytoma cells) |
↓p-PERK expression ↓p-eIF2α expression ↓ATF4 expression |
20 µM |
PERK – protein kinase R (PKR)-like endoplasmic reticulum kinase; p-PERK – phosphorylated PERK; eIF2α – eukaryotic translation initiation factor 2α; p-eIF2α – phosphorylated eIF2α; IRE1 – inositol-requiring enzyme 1; p-IRE1 – phosphorylated IRE1; CHOP – C/EBP-homologous protein; XBP1 – X-box binding protein 1; ATF4 – activating transcription factor 4; ATF6 – activating transcription factor 6; GRP78 – glucose-regulated protein 78; GRP94 – glucose-regulated protein 94; ERAD – endoplasmic reticulum-associated protein degradation; JNK – c-Jun N-terminal kinase.
Table 3.
Influence of ER stress markers by cyanidin-3-o-glucoside, epicatechin and chlorogenic acid.
Table 3.
Influence of ER stress markers by cyanidin-3-o-glucoside, epicatechin and chlorogenic acid.
| Author(s) | Year | Experimental Model | ER Stress Modulating Substance |
Altered Expression or Activity of ER Stress Markers | Dosage |
|---|---|---|---|---|---|
| Thummayot et al. [38] | 2016 | In vitro (Aβ 25-35 induced neuronal cell death in SK-N-SH cells) | Cyanidin-3-o-glucoside | ↓GRP78 expression ↓p-PERK expression ↓p-eIF2α expression ↓ IRE1 expression ↓ XBP1 expression ↓ ATF6 expression ↓ CHOP expression |
0.2; 2; 18 and 20 µM |
| Chen et al. [40] | 2022 | In vitro (treated with palmitate isolated mouse pancreatic islets and INS-1E cells) | Cyanidin-3-o-glucoside | ↓ CHOP expression | 12,5; 25 and 50 µM |
| Tu et al. [41] | 2022 | In vivo (induced perodontitis in rats) | Cyanidin-3-o-glucoside | ↓ CHOP expression ↓ JNK and p-JNK expression |
3 or 9 mg/kg |
| Peng et al. [42] | 2022 | In vitro (blue light-irradiated retinal pigment epithelial cells) | Cyanidin-3-o-glucoside | ↓ATF4 expression ↓ CHOP expression |
10 and 25 μM |
| Bettaieb et al. [52] | 2014 | In vivo (high-fructose diet-fed rats) | Epicatechin | ↓p-PERK expression ↓p-IRE1 expression ↓XBP1 splicing |
20 mg/kg |
| Kang et al. [53] |
2019 | In vitro (methamphetamine-induced neurotoxicity in HT22 hippocampal neuronal cells) | Epicatechin | ↓CHOP expression | 10 and 20 μM |
| Ye et al. [54] | 2016 | In vivo (streptozotocin-induced diabetic nephropathy in rats) | Chlorogenic acid | ↓CHOP expression ↓ATF6 expression ↓p-eIF2α expression ↓p- PERK expression |
5 mg/kg, 10 mg/kg, 20 mg/kg |
| Wang et al. [55] | 2017 | In vivo (bleomycin-induced pulmonary fibrosis in mice), in vitro (pulmonary fibroblasts and RLE-6TN cells) | Chlorogenic acid | ↓GRP78 expression ↓CHOP expression ↓p-PERK expression ↓ATF6 expression ↓caspases 9, 3 and 12 expression |
15 mg/kg, 30 mg/kg, 60 mg/kg |
| Zhang et al. [56] | 2018 | In vitro (thapsigargin and palmitic acid-induced ER stress in rat hepatocytes) | Chlorogenic acid | ↓GRP78 expression ↓GRP94 expression ↓CHOP expression |
5 μmol/l |
| Kazaz et al. [57] | 2022 | In vivo (torsion/detorsion-induced testicular injury in rats) | Chlorogenic acid | ↓GRP78 expression ↓ATF6 expression ↓CHOP expression |
100 mg/kg |
| Rani et al. [58] | 2022 | in vitro (model of hyperglycemia in H9c2 embryonic rat heart cells) | Chlorogenic acid | ↓p-PERK expression ↓p-eIF2α expression ↓ATF4 expression ↓p-IRE1 expression ↓TRAF2 expression ↓p-JNK expression ↓XBP1 expression ↓ATF6 expression |
10 and 30 μM |
| Sari et al. [59] | 2022 | In vivo (diabetic model in rats) | Chlorogenic acid | ↓GRP78 expression ↓ XBP1 expression |
12.5 mg/kg, 25 mg/kg and 50 mg/kg |
| Moslehi et al. [60] | 2023 | In vivo (tunicamycin-induced ER stress in mice) | Chlorogenic acid | ↓GRP78 expression ↓PERK expression ↓IRE1 expression ↓caspase 3 expression |
20 and 50 mg/kg |
| Boonyong et al. [61] | 2023 | In vivo (indomethacin-induced gastrointestinal ulcer in rats) | Chlorogenic acid | ↓p-PERK expression ↓p-eIF2α expression ↓ATF-4 expression ↓CHOP expression |
100 mg/kg |
| Ping et al. [62] | 2024 | In vitro (isoproterenol stimulated H9c2 myocardial cells) In vivo (isoproterenol stimulated rats) |
Chlorogenic acid | ↓GRP78 expression ↓p-PERK expression ↓CHOP expression ↓caspases 12, 3 and 9 expression |
90 mg/kg (in vivo model) 50 μM (in vitro model) |
PERK – protein kinase R (PKR)-like endoplasmic reticulum kinase; p-PERK – phosphorylated PERK; eIF2α – eukaryotic translation initiation factor 2α; p-eIF2α – phosphorylated eIF2α; IRE1 – inositol-requiring enzyme 1; p-IRE1 – phosphorylated IRE1; CHOP – C/EBP-homologous protein; XBP1 – X-box binding protein 1; ATF4 – activating transcription factor 4; ATF6 – activating transcription factor 6; GRP78 – glucose-regulated protein 78; GRP94 – glucose-regulated protein 94; TRAF 2 (TNF receptor-associated factor 2); JNK – c-Jun N-terminal kinase; p-JNK – phosphorylated JNK.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



