Submitted:
24 July 2025
Posted:
28 July 2025
You are already at the latest version
Abstract
Background/Objectives: Human papillomavirus (HPV) is the main causative agent of cervical cancer and contributes to a significant proportion of other anogenital and oropharyngeal malignancies. The need for better biomarkers and therapeutic approaches in HPV-associated cancers has drawn attention to exosomes, small extracellular vesicles known for their stability, biomolecule transport capabilities, and role in cell-to-cell communication. Methods: This review comprehensively evaluates recent literature on the diagnostic, prognostic, and therapeutic applications of small extracellular vesicles, particularly exosomes, in HPV-related cancers. It analyzes findings on exosomal nucleic acids, proteins, and long non-coding RNAs, as well as engineered exosome-based therapies. Results: Exosomal miRNAs (e.g., miR-204-5p, miR-99a-5p, miR-21), proteins (e.g., glycolytic enzymes, HSP90), and lncRNAs (e.g., HOTAIR, DLEU1) have emerged as promising biomarkers for disease detection and monitoring. In HPV-positive head and neck and cervical cancers, exosomal cargo reflects HPV status, tumor progression, and treatment response. Therapeutic studies demonstrate the utility of exosomes in vaccine delivery, immune modulation, and drug delivery systems, including the use of PROTACs. However, clinical translation faces barriers including isolation protocol standardization, biomarker validation, and scalable production. Conclusions: Exosomes hold great promise for integration into diagnostic and therapeutic workflows for HPV-related cancers. Future research should focus on resolving standardization issues, validating biomarkers in diverse cohorts, and optimizing engineered exosome platforms for targeted therapy.
Keywords:
exosomes
; extracellular vesicles
; Human Papillomavirus (HPV)
; liquid biopsy
; biomarkers
; cervical cancer
; head and neck squamous cell carcinoma (HNSCC)
1. Introduction
Being the primary causative agent for nearly all cervical malignancies and a high fraction of other anogenital and oropharyngeal cancers such as head and neck, vaginal, vulvar, and penile, Human Papillomavirus (HPV) represents a serious global health burden [1]. Globally, cervical cancer presents as the 4th common cancer in women [2] and is estimated to have caused 350 000 deaths with 660 000 new cases in 2022 [3]. It also places 4th in terms of mortality [3]. Low-to-middle income countries are the most affected, as they lack sufficient screening, prevention and treatment options [3]. From data as of August 2022, a total of 21,800 cancer cases for women and 16,000 cancer cases for men each year are thought to be due to HPV [1]. It takes a long time, maybe even decades, for a person infected with HPV to develop cancer, and there is no sure way of knowing who will undergo malignant transformation [1].
In recent years, extracellular vesicles (EVs), particularly exosomes, have gathered attention in the field of oncology as they were demonstrated to play a key role in intercellular signaling and to mirror the functional and disease-related status of their parent cells [4]. Exosomes are vesicles with diameters ranging from 30-150 nm, and they can be secreted by many cell types, including cancer cells. [5] Exosomes can be detected in various bodily fluids like saliva, blood and urine [6,7]. These vesicles can carry a diverse cargo of biomolecules with different functions, including nucleic acids such as DNA, RNA, miRNAs, lncRNAs, proteins, and lipids. The cargo can also be transferred to recipient cells, which influences their function [8]. This mechanism of cargo donation can potentially contribute to cancer progression, metastasis, and even drug resistance [9].
Exosomes have valuable and unique properties that make them promising non-invasive biomarkers such as being stable in the circulation, protecting the cargo from degradation with a lipid bilayer, and being able to pass cellular barriers [5]. Liquid biopsy can be used as a less invasive alternative to the traditional tissue biopsy, and allows to monitor disease progression and response to treatment in real time. It can also be useful for early detection of malignancies [7,10]. It allows clinicians to generate a characteristic molecular picture of the cancer. Exosomes-based liquid biopsy is a candidate with high potential in the field of HPV-related cancers, as exosomal content is shown to change in various ways with respect to the HPV infection status in different types of cancers [11]. The dynamic changes in the tumor microenvironment (TME) can be monitored through the exosomal cargo that closely mirror the ongoing changes [12].
In addition to their potential use in diagnosis and prognostic determination, exosomes are also investigated as novel tools in cancer therapeutics. Their natural compatibility with the body, low potential to trigger immune responses, and ability to transport functional biological cargo make exosomes good vehicles for precision therapies, vaccine platforms, and standalone treatment approaches [13]. This review aims to present a comprehensive overview of the current understanding and future prospects of exosomes in HPV-associated malignancies by exploring their applications as biomarkers in liquid biopsy for detection, prognosis, and monitoring, as well as their promising therapeutic potential.
2. Exosomes and Liquid Biopsy: Diagnostic and Prognostic Applications
2.1. Exosomal Nucleic Acids
Exosome-based liquid biopsy as an emerging field offers a minimally invasive and impactful approach for early detection, prognosis assessment, and therapy monitoring of HPV-associated cancers (Figure 1). Exosomes contain stable cargo such as miRNAs, lncRNAs, mRNAs, proteins, and even viral transcripts that reflect the changing molecular state of their parent cells, making them valuable sources of biomarkers related to diseases. Multiple studies have identified exosomal microRNAs (exo-miRNAs) as promising diagnostic and prognostic tools (Table 1). For instance, serum exosomal miR-204-5p levels progressively increase across cervical lesion severity (healthy < LSIL < HSIL), suggesting applicability in risk stratification for cervical lesions and disease monitoring [14]. Likewise, plasma levels of EV-associated Wnt7b are significantly elevated in cervical cancer patients and correlate with clinical invasiveness and poor outcomes. EV-Wnt7b serves as an independent prognostic marker and is integrated into a validated survival-predictive framework [15].
HPV-specific transcripts have also been detected in exosomes. The spliced E6*I mRNA variant of HPV16, detectable in HPV16-positive cervical cancer patients from plasma-derived exosomal RNA, supports the use of EV-based RNA profiling for detection of viral oncogenes and disease monitoring [16] (Table 2). Additionally, exosomal miR-99a-5p, which is enriched in the plasma of HPV-positive head and neck squamous cell carcinoma (HNSCC) patients but inversely expressed in tissue, could reflect an export mechanism controlled by tumor cell-specific pathways and emerges as a candidate non-invasive diagnostic biomarker [17,18]. In HPV+ HNSCC, 118 exo-miRNAs showed different expression levels compared to HPV- cells and patterns were detectable in the serum of early-stage HNSCC patients. miR-99a-5p, for example, correlated with recurrence-free survival, showing promise as a biomarker in HPV-related HNSCC [19]. In head and neck cancers, a core set of 25 exo-miRNAs were differentially secreted across all HPV-positive and HPV-negative HNSCC cell lines, including consistently upregulated miR-451a and miR-16-2-3p. These miRNAs were also significantly elevated in serum samples from early-stage patients, showing reproducibility and high clinical relevance [19]. An interesting finding is that some miRNAs like miR-125a-5p and miR-3168 were abundant in serum but absent in tumor cells, hinting at a non-tumor source [19]. This further highlights the complexity of EV profiling in search of potential tumor-related biomarkers. In another cancer type, cervical cancer, exosomal miR-125a-5p levels were found to be significantly lower compared to healthy controls. Notably, HPV+ patients showed even lower levels than HPV- ones. This downregulation in miR-125a-5p is likely shaped by the HPV-related suppression of p53, as HPV has been shown to inhibit expression of miR-125a-5p. The reduction in levels of tumor suppressive miR-125a-5p could have a role for cervical carcinogenesis, which could make it a potential biomarker for diagnosis [20]. In cervicovaginal lavage samples, miR-21 and miR-146a were specifically enriched in the exosomal fraction, and not the supernatant and correlated with CD9 (an exosomal marker) levels. The increase was significant in cervical cancer patients compared to both HPV-positive and HPV-negative controls. HPV infection alone was also a contributor to the upregulation. The active secretion and abnormal elevation of exosome-encapsulated miR-21 and miR-146a by the cancer tissue presents other candidates as non-invasive biomarkers to detect cervical cancer [21].
Even though exosome release may be manipulated in the presence of HPV infection, the change in HPV type may not affect the exosomal content. Exosomal miRNAs like let-7d-3p and miR-30d-5p are valuable diagnostic biomarkers for non-invasive screening of cervical cancer and its precursors, irrespective of HPV type. Expression profiles of 8 miRNA signatures targeting the viral carcinogenesis pathway were also not different among various HPV types [22]. Some non-invasive biomarkers could be applicable for multiple HPV types and provide a tool for broad-spectrum screening, as in this example.
2.2. Exosomal Protein Biomarkers
Presence of HPV can also modulate the exosomal protein content, which presents proteins as potential biomarker candidates in discriminating HPV+ patients (Table 2). Proteins like ANXA1, HSP90, and ACTN4 were upregulated in oral cancer derived EVs and have been proposed as markers to assess disease progression [18]. The modified proteins can also be enzymes related to important pathways. In HPV-driven oropharyngeal cancer (OPC) patients, HPV16 E6/7 DNA was detected in salivary exosomes in 80%, but not found in healthy controls. In salivary exosomes that were HPV-modified, six glycolytic enzymes (ALDOA, GAPDH, LDHA, LDHB, PGK1, and PKM) were significantly upregulated. This shows enhanced glycolysis and potentially formation of the Warburg effect in the setting of HPV-driven carcinogenesis. HIF-1α was predicted to get activated and act as an upstream regulator, supporting HPV’s role in metabolic reprogramming. This change in protein profiles of salivary exosomes could be useful in the discrimination of HPV+ OPC patients from healthy individuals [23].
2.3. Exosomal Long Non-Coding RNAs
Exosomal long non-coding RNAs (lncRNAs) also present as changing cargo in HPV+ exosomes (Table 2). Particularly HOTAIR, MALAT1, and MEG3 are highly enriched in cervicovaginal lavage (CVL) samples from HPV-positive patients and indicate potential use for early detection and HPV-related risk stratification [24]. Exosomal DLEU1 (a lncRNA) was also significantly upregulated in cervical cancer patients compared to healthy controls and those with cervical intraepithelial neoplasia (CIN). Its expression was associated with tumor burden and poorer prognosis. It was concluded that HPV infection status did not correlate with exosomal DLEU1 levels [25] but previous studies show that DLEU1 expression in cervical cancer tissues was linked to HPV infection [26]. This shows that while HPV influences DLEU1 expression of tissues, the exosomal release of the lncRNA is reflective of something more than solely the HPV infection status: malignant progression. The presence of increased exosomal DLEU1 could act as a diagnostic and prognostic biomarker in CC [25].
2.4. Monitoring of Treatment Response
EVs also show probability of being real-time indicators of therapy response through longitudinal profiling. In HPV/p16+ HNSCC patients, exosomal miR-21, -let-7a, and -181a were significantly higher than HPV- patients in the initial diagnosis. miR-21 was downregulated in HPV+ tissue but increased in exosomes. In HPV/p16- patients, EV-miR-21 expression was found to significantly increase during the 12 month follow-up. These suggest diagnostic and treatment monitoring utility [27].
Altogether, the stability and accessibility of exosomes, along with their varied content position them as potentially transformative tools in HPV-related oncology. Their cargo provides a molecular picture of tumor activity and host interactions. This could enable sensitive, real-time assessment of disease state and the therapy response. Continued validation across large, prospective cohorts is likely to make their role clearer in clinical practice for HPV-related cancer types like cervical and head and neck cancers.
3. Exosome-Related Therapies
3.1. Exosome-Induced Immune Stimulation
Therapies based on exosomes are emerging as potential tools in the treatment of HPV-associated cancers (Table 3). Multiple studies show their ability to induce antitumor immunity, be used as delivery agents for therapeutic molecules, and help overcome tumor resistance mechanisms. Jin et al [28] demonstrated that the 5-aminolevulinic acid photodynamic therapy (ALA-PDT) could suppress the growth of HPV+ CC cells by promoting apoptosis and downregulating miR-34a. (Figure 2A) This led to increased HMGB1 secretion in exosomes of the CC cells. The change in exosomal content supplemented DC maturation and the production of pro-inflammatory cytokines, leading to strengthened anti-tumor immune response. These effects could be reversed either by silenced HMGB1 or restoration of miR-34a, which confirms that the miR-34a/HMGB1/exosome axis is therapeutically relevant. The ALA-PDT treatment was confirmed to stimulate the immune response through its actions of exosomal mechanisms specifically for HPV+ CC. Additionally, Tong et al [29] showed that HPV+ HNSCC exosomes carry an enriched amount of miR-9. Macrophages in the surrounding TME could import these exosomes which resulted in the downregulation of PPARδ and the subsequent M1 polarization of the macrophages. Interestingly, HPV+ HNSCC cells having a high number of pro-inflammatory M1 macrophages exhibited augmented radiosensitivity when treated with 6 Gy ionizing radiation, according to the increase in γ-H2AX foci. The authors propose miR-9 as a potential treatment element in HPV+ HNSCC. (Figure 2A)
3.2. Exosome-Based Vaccines
Another study related to HPV immunity was from Manfredi et al. [30] who investigated the treatment efficiency of exosomes engineered to carry HPV-E6 tumor-associated antigen (TAA) in combination with the ISCOMATRIX™ adjuvant (containing purified ISCOPREPTM saponin, cholesterol, and phospholipids) (Figure 2B). They found that the co-administration of engineered exosomes carrying HPV-E6 tumor-associated antigen (TAA) and the adjuvant molecule resulted in significantly enhanced in vitro cross-presentation of antigens by B-lymphoblastoid and immature dendritic cells. In in vivo experiments, the CD8+ T cell response against the HPV E6 protein was seen to exceed the one with the presence of exosomes alone. Exosome vaccines enhanced with adjuvant molecules like ISCOMATRIX™ could show promise as immune response-enhancing therapies.
In a study by di Bonito et al. [31], a novel exosome-based vaccination strategy was generated to create CD8+ T lymphocyte (CTL) responses against specific protein antigens. The setup used a mutated HIV-1 Nef protein (Nefmut) as the exosome-anchoring domain. HPV E7 was fused to Nefmut, which enabled efficient loading of the antigen into the exosomes. Even without including the fusogenic protein VSV-G, the engineered exosomes showed immunogenicity same as lentiviral virus-like particles (VLPs). Exosomes carrying HPV E7 could induce E7-targeted CTL responses successfully in mouse models also. When administered prophylactically, this method prevented tumor growth and therapeutically suppressed the established tumors. So, Nefmut-fused E7 loaded onto exosomes were found to be an effective method to elicit CTL immunity. (Figure 2B) In a following study by di Bonito et al. [32] a strong CD8+ T cell immune response against the HPV E7 DNA vector was generated through endogenous production of engineered exosomes, in an effort to create an alternative to in vitro-engineered exosomes that posed problems in terms of clinical applicability. A DNA vector for E7 and Nefmut were fused together and injected intramuscularly, resulting in the production of exosomes and their release into the circulation. In a TC-1 tumor mouse model, antitumor effects were observed as a result of the cytotoxic response (Figure 2B). This cost-effective and scalable method could supplement immunotherapy platforms in HPV-related cancers.
3.3. Exosomes as Drug Delivery Vehicles
Exosomes were also studied as carriers for chemotherapeutic molecules such as natural compounds or synthetic proteins. Abbasifarid et al. [33] investigated the immunological and tumor-therapeutic effects of exosomes loaded with crocin and curcumin compounds administered in combination with a synthetic HPV L1-E7 polypeptide vaccine, in C57BL/6 mouse models bearing TC-1 tumors. In certain doses, the ExoCrocin and ExoCurcumin were not cytotoxic and could enter tumor cells. Cytokine assays showed the secretion of IFN-γ and IL-4 was increased in L1-E7 polypeptide + ExoCurcumin/ExoCrocin groups compared to controls. IFN-γ levels were higher in the L1-E7 polypeptide + ExoCurcumin/ExoCrocin groups compared to both ExoCurcumin/ExoCrocin or the polypeptide alone. The coadministration of the engineered exosomes and the polypeptide vaccine produced a significant level of immunity directed toward Th1 response and CTL activity, demonstrating potential as a successful immune-focused therapy candidate. (Figure 2C)
3.4. Engineered Exosomes: PROTACs and Targeting Strategies
As for synthetic exosomal therapeutic cargo, proteolysis-targeting chimeras (PROTACs) present as new candidates. Although ongoing studies are present in various types of cancers [34] it is an understudied area in HPV-related cancer therapy. Exosomes are derived from body fluids and engineered to carry PROTACs through methods such as liposome fusion and electroporation. The PROTACs facilitate E3 ubiquitin ligase activity to degrade E6 and E7 proteins, which are critical elements of HPV-related carcinogenesis (Figure 2C). This allows for targeting HPV-associated mechanisms and elimination of non-specific toxicity. In addition, exosomes can be specifically targeted to tissues by modifying surface ligands [35]. This strategy aims to strengthen tumor suppressor pathways by eliminating oncogenic driver proteins and may prove to be successful, but as a relatively new area especially in HPV-related cancers, there is much work to be done for clinical translation.
3.5. Exosomes and Therapy Resistance
In addition to being used as therapeutic agents, exosomes produced by cancer cells can contribute to treatment resistance mechanisms (Table 4). In their review on therapy targets for HPV+/- oral and tongue cancers, Gupta et al. [36] present exosomes from cancer stem cells (CSC) as important agents in therapy resistance, recurrence, and metastasis. In tongue squamous cell carcinoma (TSCC), sometimes associated with HPV positivity, one proposed mechanism of mediating tumor progression and relapse is through the CSC-derived exosomes that transmit miRNAs associated with chemoresistance. Targeting of these vesicles is discussed as a route to overcome relapse and resistance in TSCC. In an investigation of the reasons for failure of anti-PD-1 immunotherapy (nivolumab) in HNSCC, Hill et al [37] also point out cargo of small extracellular vesicles (sEV). Non-responders to therapy had high levels of IL-8 and reduced levels of IL-8 targeting miRNAs in their sEVs like miR-146a. Dsg2, an oncoprotein that suppresses miR-146a was found to be elevated in HPV+ tumors and was present in high levels in non-responders post-treatment. The Dsg2/miR-146a/IL-8 axis is defined as a potential contributor to immune checkpoint inhibitor resistance and could be targeted to improve therapy outcomes.
Taken together, these findings demonstrate that exosomes could have various therapeutic applications in HPV-associated cancers, from immune stimulation and tumor antigen presentation to targeted delivery of therapeutic agents and enhancement of peptide-based vaccines. The mentioned studies and reviews each show a unique aspect of exosome biology that can be utilized as a treatment strategy for immunotherapy, combination therapies, or next-generation vaccines. While promising, findings also highlight the need for further validation and optimization to overcome the remaining challenges in clinical translation.
4. Future Directions
Despite the growing interest in exosomes as diagnostic and therapeutic agents in HPV-associated cancers, several important challenges remain. First, there is a need to standardize exosome isolation, quantification, and characterization protocols. [38,39] In the mentioned studies, patient exosomes are derived from different sources: body fluids like blood plasma [6,16,17], saliva [23], urine [6], and from cervicovaginal lavage [21,24] (Figure 1). Blood plasma is the most preferred, but there is no comparison between the effectiveness of different sources and how their utility could differ, and for each study, the most convenient source is chosen for analysis. In addition, no consensus is present on which exosomal markers need to be checked for HPV infection characterization or the related tumor stage/behaviour determination. Because exosomal cargo is very heterogeneous and closely dependent on the cellular context, singular isolation of specific biomolecules is hard. [40,41] Without standardization of the isolation and analysis processes, clinical reproducibility and approval will remain elusive.
Biomarker validation across large, diverse, and longitudinal HPV+ cohorts is also lacking. Many studies report exosomal cargo like miRNAs or proteins as promising biomarkers, but few have demonstrated prognostic or predictive strength in real-world clinical settings. Future efforts should focus on integrating liquid biopsy platforms with multi-omics analysis [40] and clinical outcome data to assess utility in early detection, risk stratification, and real-time therapy monitoring.
In addition, the therapeutic potential of engineered exosomes, especially those delivering HPV oncoprotein-targeting PROTACs or vaccine antigens, remains promising but underexplored [42]. The biocompatibility of engineered exosomes remains questionable, which is why some researchers try to achieve in vivo-production of therapeutic exosomes. Future preclinical work should emphasize in vivo efficacy, immune profiling (to make sure the exosomes do not activate unwanted host immune reactions), biodistribution, and scalability of the manufacturing process. The development of targeted delivery systems, perhaps via ligand engineering or nanomolecule incorporation, may further enhance precision.
In prospective clinical studies, exosome-mediated mechanisms of therapy resistance and immune evasion such as the IL-8/miR-146a axis [37] should be investigated, particularly in the context of immune checkpoint blockade failure. There is also a lack of studies investigating how exosomes are affected during administration of other kinds of mainstream or preclinical chemotherapeutic drugs. Understanding the resistance pathways governed by exosomes could lead to the development of predictive markers using liquid biopsy data or combination strategies to restore immune responsiveness when faced with resistance in cancer treatment. Molecular mechanisms underlying resistance should be further explored in basic studies before clinical application.
5. Conclusions
Even though there are various in vitro studies investigating the relationship between HPV infection, related cancers and the role of extracellular vesicles/exosomes; there is not an extensive amount of research focused on the diagnostic and therapeutic purposes of exosomes in cancers specifically associated with HPV [42]. Because the relationship between EV/exosomal content and HPV infections still remains a mystery, and new findings are coming to light every day, it is not possible to extend the basic research into the clinical application area yet. Much work has to be done to solidify the related molecular pathways in HPV related cancers and how EVs are affected by them, or used as tools by the tumor cells to supplement cancer progression [41]. Then, in addition to introducing engineered exosomes as therapeutic vehicles, the organic EVs can also potentially be manipulated to increase the therapeutic efficiency or as a treatment method itself.
With the rapid advancement of exosome research and biotechnology, the coming decade may see the integration of exosomal diagnostics and therapeutics into the routine clinical framework for HPV-related cancers, provided that these translational and regulatory obstacles are actively addressed [43].
Abbreviations
The following abbreviations are used in this manuscript:
| ALA-PDT CC CSC CTL EV |
5-aminolevulinic acid photodynamic therapy Cervical cancer Cancer stem cell Cytotoxic T lymphocyte Extracellular vesicle |
| HNSCC HPV |
Head and neck squamous cell carcinoma Human papillomavirus |
| OPC | Oropharyngeal carcinoma |
| PROTAC TME TSC |
Proteolysis-targeting chimera Tumor microenvironment Tongue squamous cell carcinoma |
References
- Centers for Disease Control and Prevention. “Cancers Caused by HPV.” Human Papillomavirus (HPV), Available online: www.cdc.gov/hpv/about/cancers-caused-by-hpv.html. (accessed on 15 July 2025).
- Bray, F., Ferlay, J., Soerjomataram, I., Siegel, R. L., Torre, L. A., & Jemal, A. (2018). Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer Journal for Clinicians, 68(6), 394–424. [CrossRef]
- World Health Organization. “Cervical Cancer.” Available online: www.who.int/news-room/fact-sheets/detail/cervical-cancer. (accessed on 15 July 2025).
- Kuang, L., Wu, L., & Li, Y. (2025). Extracellular vesicles in tumor immunity: mechanisms and novel insights. Molecular Cancer, 24(1), BioMed Central Ltd. [CrossRef]
- Chen, Y. F., Luh, F., Ho, Y. S., & Yen, Y. (2024). Exosomes: a review of biologic function, diagnostic and targeted therapy applications, and clinical trials. In Journal of Biomedical Science (Vol. 31, Issue 1). BioMed Central Ltd. [CrossRef]
- Li, M., Zeringer, E., Barta, T., Schageman, J., Cheng, A., & Vlassov, A. v. (2014). Analysis of the RNA content of the exosomes derived from blood serum and urine and its potential as biomarkers. Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1652). [CrossRef]
- Li, S., Yi, M., Dong, B., Tan, X., Luo, S., & Wu, K. (2021). The role of exosomes in liquid biopsy for cancer diagnosis and prognosis prediction. In International Journal of Cancer (Vol. 148, Issue 11, pp. 2640–2651). John Wiley and Sons Inc. [CrossRef]
- Yáñez-Mó, M., Siljander, P. R. M., Andreu, Z., Zavec, A. B., Borràs, F. E., Buzas, E. I., Buzas, K., Casal, E., Cappello, F., Carvalho, J., Colás, E., Cordeiro-Da Silva, A., Fais, S., Falcon-Perez, J. M., Ghobrial, I. M., Giebel, B., Gimona, M., Graner, M., Gursel, I., … de Wever, O. (2015). Biological properties of extracellular vesicles and their physiological functions. In Journal of Extracellular Vesicles (Vol. 4, Issue 2015, pp. 1–60). Co-Action Publishing. [CrossRef]
- Wang, X., Qiao, D., Chen, L., Xu, M., Chen, S., Huang, L., Wang, F., Chen, Z., Cai, J., & Fu, L. (2019). Chemotherapeutic drugs stimulate the release and recycling of extracellular vesicles to assist cancer cells in developing an urgent chemoresistance. Molecular Cancer, 18(1). [CrossRef]
- Nikanjam, M., Kato, S., & Kurzrock, R. (2022). Liquid biopsy: current technology and clinical applications. In Journal of Hematology and Oncology (Vol. 15, Issue 1). BioMed Central Ltd. [CrossRef]
- Sadri Nahand, J., Moghoofei, M., Salmaninejad, A., Bahmanpour, Z., Karimzadeh, M., Nasiri, M., Mirzaei, H. R., Pourhanifeh, M. H., Bokharaei-Salim, F., Mirzaei, H., & Hamblin, M. R. (2020). Pathogenic role of exosomes and microRNAs in HPV-mediated inflammation and cervical cancer: A review. In International Journal of Cancer (Vol. 146, Issue 2, pp. 305–320). Wiley-Liss Inc. [CrossRef]
- Paskeh, M. D. A., Entezari, M., Mirzaei, S., Zabolian, A., Saleki, H., Naghdi, M. J., Sabet, S., Khoshbakht, M. A., Hashemi, M., Hushmandi, K., Sethi, G., Zarrabi, A., Kumar, A. P., Tan, S. C., Papadakis, M., Alexiou, A., Islam, M. A., Mostafavi, E., & Ashrafizadeh, M. (2022). Emerging role of exosomes in cancer progression and tumor microenvironment remodeling. In Journal of Hematology and Oncology (Vol. 15, Issue 1). BioMed Central Ltd. [CrossRef]
- Gabaran, S. G., Ghasemzadeh, N., Rahnama, M., Karatas, E., Akbari, A., & Rezaie, J. (2025). Functionalized exosomes for targeted therapy in cancer and regenerative medicine: genetic, chemical, and physical modifications. In Cell Communication and Signaling (Vol. 23, Issue 1). BioMed Central Ltd. [CrossRef]
- Chen, X., Liu, Y., Luo, X., & et al. (2024). HPV16 E6-induced M2 macrophage polarization in the cervical microenvironment via exosomal miR-204-5p. Scientific Reports, 14(1). [CrossRef]
- Qiu, J., Sun, S., Tang, X., Lin, Y., & Hua, K. (2020). Extracellular vesicular Wnt7b mediates HPV E6-induced cervical cancer angiogenesis by activating the β-catenin signaling pathway. Journal of Experimental & Clinical Cancer Research, 39(1). [CrossRef]
- Bhat, A., Yadav, J., Thakur, K., & et al. (2022). Transcriptome analysis of cervical cancer exosomes and detection of HPVE6*I transcripts in exosomal RNA. BMC Cancer, 22(1). [CrossRef]
- Huang, Q., Shen, Y. J., Hsueh, C. Y., Zhang, Y. F., Yuan, X. H., Zhou, Y. J., Li, J. Y., Lin, L., Wu, C. P., & Hu, C. Y. (2022). Plasma Extracellular Vesicles-Derived miR-99a-5p: A Potential Biomarker to Predict Early Head and Neck Squamous Cell Carcinoma. Pathology and Oncology Research, 28. [CrossRef]
- Leung, L. L., Riaz, M. K., Qu, X., Chan, J., & Meehan, K. (2021). Profiling of extracellular vesicles in oral cancer, from transcriptomics to proteomics. Seminars in Cancer Biology, 74. [CrossRef]
- Galiveti, C. R., Kuhnell, D., Biesiada, J., Zhang, X., Kelsey, K. T., Takiar, V., Tang, A. L., Wise-Draper, T. M., Medvedovic, M., Kasper, S., & Langevin, S. M. (2023). Small extravesicular microRNA in head and neck squamous cell carcinoma and its potential as a liquid biopsy for early detection. Head and Neck, 45(1), 212–224. [CrossRef]
- Lv, A., Tu, Z., Huang, Y., Lu, W., & Xie, B. (2020). Circulating exosomal miR-125a-5p as a novel biomarker for cervical cancer. Oncology Letters, 21(1). [CrossRef]
- Liu, J., Sun, H., Wang, X., Yu, Q., Li, S., Yu, X., & Gong, W. (2014). Increased exosomal microRNA-21 and microRNA-146a levels in the cervicovaginal lavage specimens of patients with cervical cancer. International Journal of Molecular Sciences, 15(1), 758–773. [CrossRef]
- Zheng, M., Hou, L., Ma, Y., Zhou, L., Wang, F., Cheng, B., Wang, W., Lu, B., Liu, P., Lu, W., & Lu, Y. (2019). Exosomal let-7d-3p and miR-30d-5p as diagnostic biomarkers for non-invasive screening of cervical cancer and its precursors. Molecular Cancer, 18(1). [CrossRef]
- Tang, K. D., Wan, Y., Zhang, X., Bozyk, N., Vasani, S., Kenny, L., & Punyadeera, C. (2021). Proteomic Alterations in Salivary Exosomes Derived from Human Papillomavirus-Driven Oropharyngeal Cancer. Molecular Diagnosis and Therapy, 25(4), 505–515. [CrossRef]
- Zhang, J., Liu, S. C., Luo, X. H., Tao, G. X., Guan, M., Yuan, H., & Hu, D. K. (2016). Exosomal Long Noncoding RNAs are Differentially Expressed in the Cervicovaginal Lavage Samples of Cervical Cancer Patients. Journal of Clinical Laboratory Analysis, 30(6), 1116–1121. [CrossRef]
- Chen, Y., Cui, F., Wu, X., Zhao, W., & Xia, Q. (2025). The expression and clinical significance of serum exosomal-long non-coding RNA DLEU1 in patients with cervical cancer. Annals of Medicine, 57(1). [CrossRef]
- Dong, S., Zhang, Y., & Wang, Y. (2023). Role of extracellular vesicle in human papillomavirus-associated cervical cancer. Journal of Cancer Research and Clinical Oncology, 149(17), 16203–16212. [CrossRef]
- Apeltrath, C., Simon, F., Riders, A., Rudack, C., & Oberste, M. (2024). Extracellular Vesicle microRNAs as Possible Liquid Biopsy Markers in HNSCC—A Longitudinal, Monocentric Study. Cancers, 16(22). [CrossRef]
- Jin, Y., Guan, Z., Wang, X., Wang, Z., Zeng, R., Xu, L., & Cao, P. (2018). ALA-PDT promotes HPV-positive cervical cancer cells apoptosis and DCs maturation via miR-34a regulated HMGB1 exosomes secretion. Photodiagnosis and Photodynamic Therapy, 24, 27–35. [CrossRef]
- Tong, F., Mao, X., Zhang, S., Xie, H., Yan, B., Wang, B., Sun, J., & Wei, L. (2020). HPV + HNSCC-derived exosomal miR-9 induces macrophage M1 polarization and increases tumor radiosensitivity. Cancer Letters, 478, 34–44. [CrossRef]
- Manfredi, F., di Bonito, P., Ridolfi, B., Anticoli, S., Arenaccio, C., Chiozzini, C., Morelli, A. B., & Federico, M. (2016). The CD8+ T cell-mediated immunity induced by HPV-E6 uploaded in engineered exosomes is improved by ISCOMATRIXTM adjuvant. Vaccines, 4(4). [CrossRef]
- di Bonito, P., Ridolfi, B., Columba-Cabezas, S., Giovannelli, A., Chiozzini, C., Manfredi, F., Anticoli, S., Arenaccio, C., & Federico, M. (2015). HPV-E7 delivered by engineered exosomes elicits a protective CD8+ T cell-mediated immune response. Viruses, 7(3), 1079–1099. [CrossRef]
- di Bonito, P., Chiozzini, C., Arenaccio, C., Anticoli, S., Manfredi, F., Olivetta, E., Ferrantelli, F., Falcone, E., Ruggieri, A., & Federico, M. (2017). Antitumor HPV E7-specific CTL activity elicited by in vivo engineered exosomes produced through DNA inoculation. International Journal of Nanomedicine, 12, 4579–4591. [CrossRef]
- Abbasifarid, E., Bolhassani, A., Irani, S., & Sotoodehnejadnematalahi, F. (2021). Synergistic effects of exosomal crocin or curcumin compounds and HPV L1-E7 polypeptide vaccine construct on tumor eradication in C57BL/6 mouse model. PLoS ONE, 16(10 October). [CrossRef]
- Wang, C., Zhang, Y., Chen, W., Wu, Y., & Xing, D. (2024). New-generation advanced PROTACs as potential therapeutic agents in cancer therapy. In Molecular Cancer (Vol. 23, Issue 1). BioMed Central Ltd. [CrossRef]
- Mukerjee, N., Maitra, S., & Ghosh, A. (2024). Exosome-based therapy and targeted PROTAC delivery: A new nanomedicine frontier for HPV-mediated cervical cancer treatment. In Clinical and Translational Discovery (Vol. 4, Issue 4). Blackwell Publishing. [CrossRef]
- Gupta, S., Kumar, P., & Das, B. C. (2021). HPV+ve/−ve oral-tongue cancer stem cells: A potential target for relapse-free therapy. In Translational Oncology (Vol. 14, Issue 1). Neoplasia Press, Inc. [CrossRef]
- Hill, B. L., Calder, A. N., Flemming, J. P., Guo, Y., Gilmore, S. L., Trofa, M. A., Daniels, S. K., Nielsen, T. N., Gleason, L. K., Antysheva, Z., Demina, K., Kotlov, N., Davitt, C. J. H., Cognetti, D. M., Prendergast, G. C., Snook, A. E., Johnson, J. M., Kumar, G., Linnenbach, A. J., … Mahoney, M. Ỹ. G. (2023). IL-8 correlates with nonresponse to neoadjuvant nivolumab in HPV positive HNSCC via a potential extracellular vesicle miR-146a mediated mechanism. Molecular Carcinogenesis, 62(9), 1428–1443. [CrossRef]
- Bastón, E., García-Agulló, J., & Peinado, H. (2025). The influence of extracellular vesicles on tumor evolution and resistance to therapy. Physiological Reviews. [CrossRef]
- Mivehchi, H., Eskandari-Yaghbastlo, A., Emrahoglu, S., & et al. (2025). Tiny messengers, big Impact: Exosomes driving EMT in oral cancer. Pathology - Research and Practice, 268, 155873. [CrossRef]
- Shan, Z., Su, X., Liu, L., & Duan, S. (2025). Liquid biopsy based on EV biomarkers: A new frontier for early diagnosis and prognosis assessment of cancer at ESMO 2024. Nano TransMed, 4, 100084. [CrossRef]
- Tang, L., Zhang, W., Qi, T., Jiang, Z., & Tang, D. (2025). Exosomes play a crucial role in remodeling the tumor microenvironment and in the treatment of gastric cancer. Cell Communication and Signaling, 23(1). [CrossRef]
- Owliaee, I., Khaledian, M., Boroujeni, A. K., & Shojaeian, A. (2023). Engineered small extracellular vesicles as a novel platform to suppress human oncovirus-associated cancers. Infectious Agents and Cancer, 18(1). [CrossRef]
- Semeradtova, A., Liegertova, M., Herma, R., & et al. (2025). Extracellular vesicles in cancer´s communication: messages we can read and how to answer. Molecular Cancer, 24(1). [CrossRef]
Figure 1.
Diagnostic Liquid Biopsy Pipeline Using Exosomes.
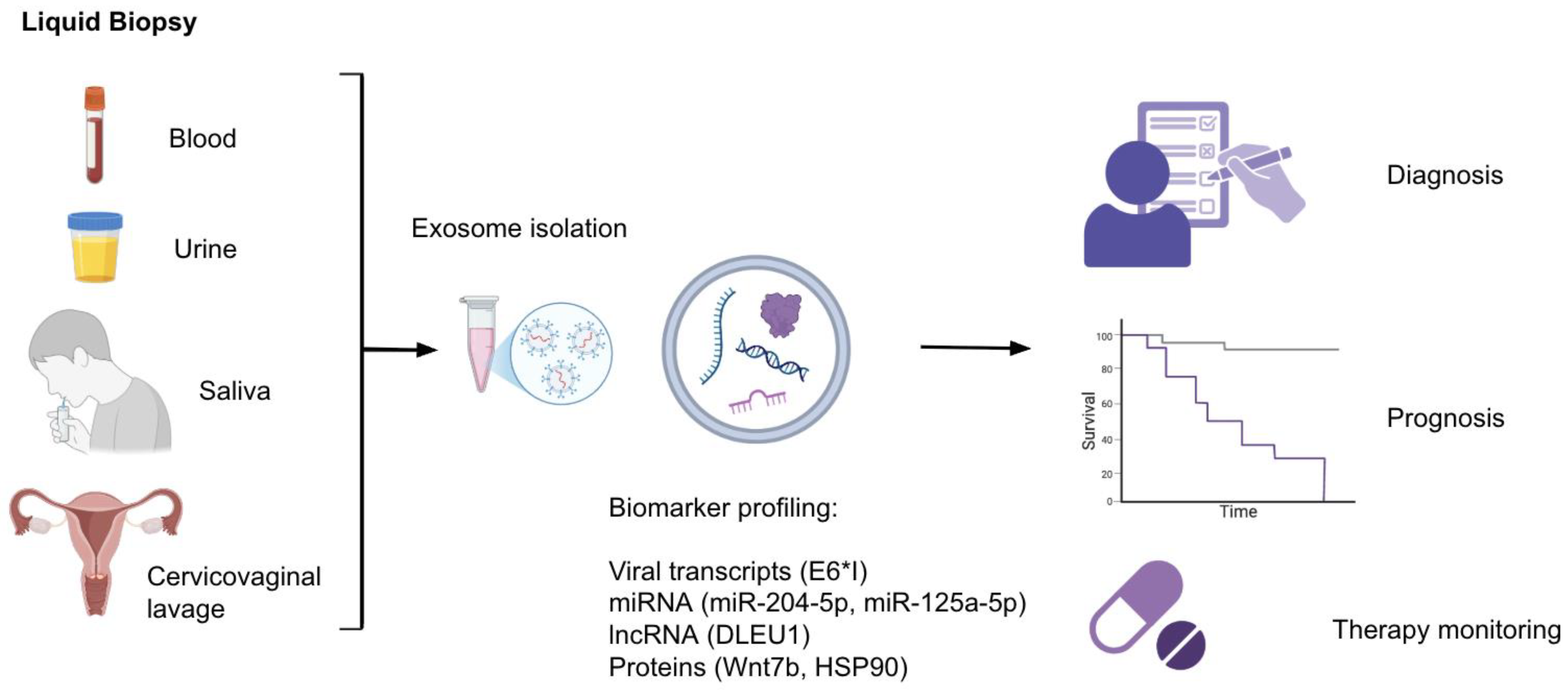
Figure 2.
Therapeutic Roles of Exosomes in HPV-Related Cancers.
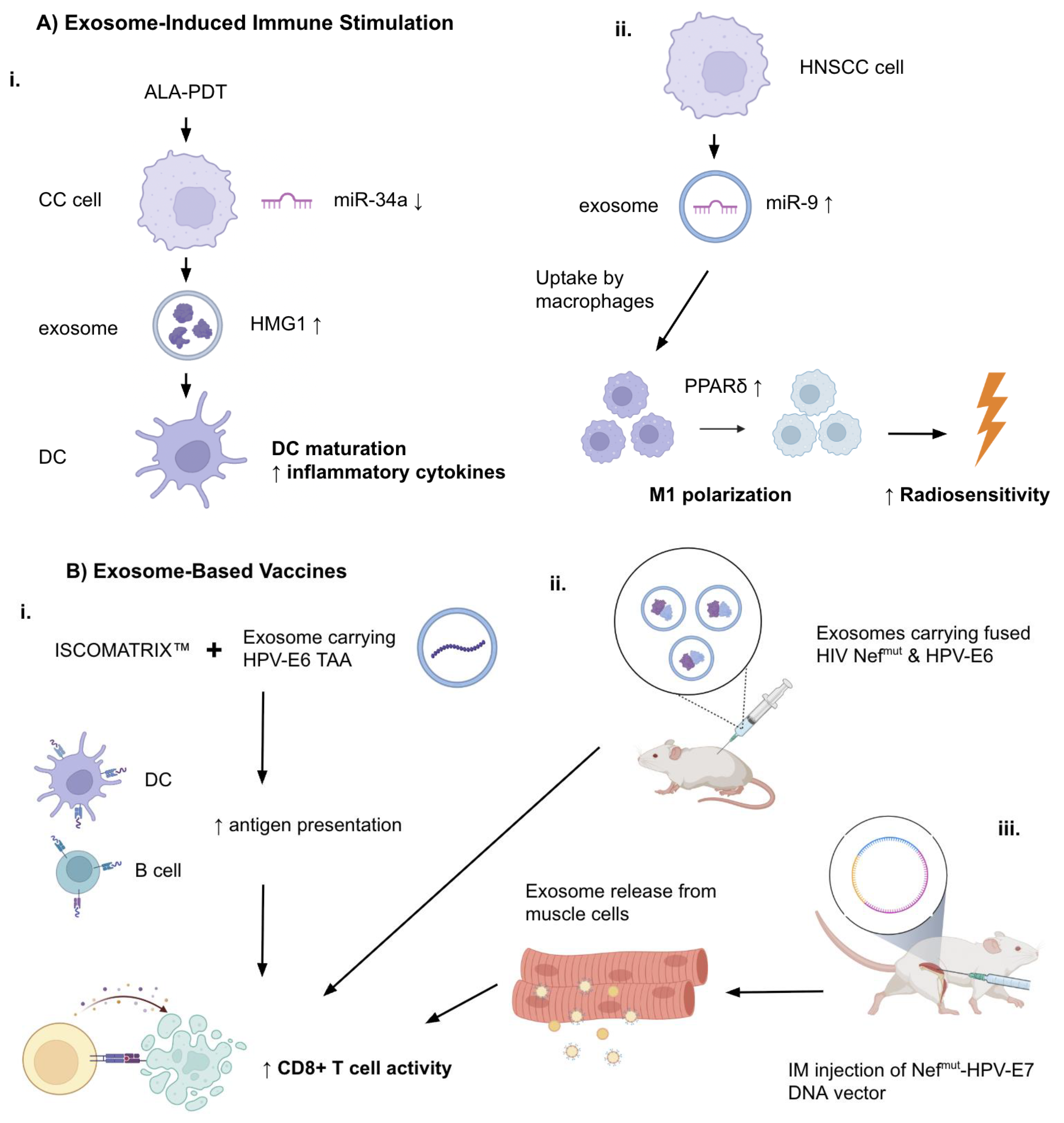
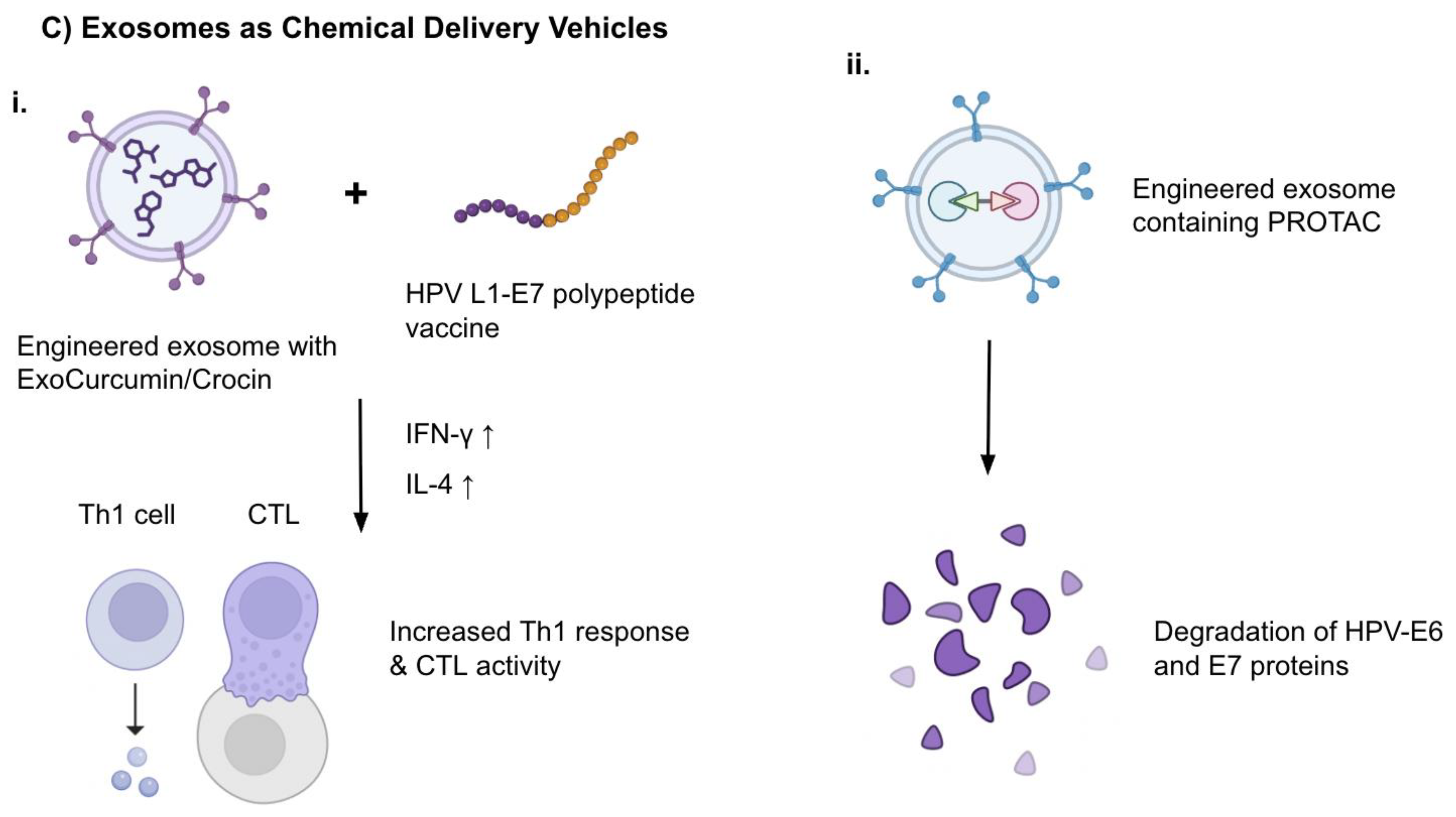

Table 1.
Exosomal miRNAs as Biomarkers in HPV-Associated Cancers.
| Biomarker | Cancer Type | HPV Status Association | Clinical Utility | Reference |
|---|---|---|---|---|
| miR-21 miR-146a |
Cervical Cancer | Upregulated in HPV+ EVs | Diagnosis | Liu et al, 2014 |
| let-7d-3p miR-30d-5p |
Cervical Cancer | Upregulated in HPV+ EVs, regardless of HPV type |
Non-invasive screening of CC, diagnosis | Zheng et al, 2019 |
| miR-125a-5p | Cervical Cancer | Downregulated in HPV+ EVs | Diagnosis | Aixia LV et al, 2021 |
| miR-451a miR-16-2-3p |
HNSCC | Upregulated in HPV+ EVs | Diagnosis, clinical reproducibility | Galiveti et al, 2022 |
| miR-99a-5p | HNSCC | Enriched in HPV+ plasma EVs | Diagnosis, RFS prediction | Huang et al, 2022 Leung et al, 2021 Galiveti et al, 2022 |
| miR-21 miR-let-7a miR-181a |
HNSCC | Upregulated in HPV+ EVs | Diagnosis, follow-up | Apeltrath et al, 2024 |
| miR-204-5p | Cervical Cancer | Upregulated in HPV+ CC EVs | Lesion severity stratification, disease monitoring | Chen et al, 2024 |
Table 2.
Other Exosomal Biomarkers in HPV-Associated Cancers.
| Biomarker | Cancer Type | HPV Status Association | Clinical Utility | Reference |
|---|---|---|---|---|
| Viral RNA (mRNA) | ||||
| HPV16 E6*I | Cervical Cancer | Present in HPV16+ EVs | Viral oncogene detection | Bhat et al, 2022 |
| DNA | ||||
| HPV16 E6/7 DNA | OPC | Present in HPV16+ salivary EVs | Detection of HPV+ OPC patients | Tang et al, 2021 |
| Proteins | ||||
| Wnt7b | Cervical Cancer | Elevated in HPV+ CC | Prognosis (OS, RFS) | Qiu et al, 2020 |
| ANXA1 HSP90 ACTN4 |
Oral Cancer | Upregulated in HPV+ EVs | Disease progression | Leung et al, 2021 |
| Glycolytic enzymes (ALDOA, GAPDH, LDHA, LDHB, PGK1, PKM) | OPC | Present in HPV+ salivary EVs | Detection of HPV+ OPC patients | Tang et al, 2021 |
| lncRNAs | ||||
| HOTAIR MALAT1 MEG3 |
Cervical Cancer | Enriched in exosomes from CVL samples of HPV+ patients | Early detection, risk stratification | Zhang et al, 2016 |
| DLEU1 | Cervical Cancer | Not HPV-type specific | Tumor burden, prognosis | Chen et al, 2025 |
Table 3.
Engineered Exosome-Based Therapeutics.
| Strategy | Cargo | Target Mechanism | Cancer Type/Model | Therapeutic Outcome | Reference |
|---|---|---|---|---|---|
| Nefmut-HPV E7 Exosomes | E7 fusion protein | CTL generation | Mouse TC-1 tumor | Anti-tumor CTL response | Bonito et al, 2015 |
| DNA vector for E7-Nefmut | Endogenous E7 exosome production | Immunization without ex vivo engineering | Mouse TC-1 tumor | Anti-tumor CTL response | Bonito et al, 2017 |
| Exo + ISCOMATRIX™ | HPV E6 protein | Enhance antigen presentation | C57 Bl/6 mice | Anti-tumor CTL response | Manfredi et al, 2016 |
| ExoCurcumin/Crocin + L1-E7 vaccine | Natural compounds + vaccine | Th1/CTL immunity induction | Mouse TC-1 tumor | Increased IFN-γ & IL-4 | Abbasifarid et al, 2021 |
| Exosomal PROTACs | E6/E7 degraders | Oncoprotein elimination | Theoretical model (HPV-related) | Targeted degradation | Mukherjee et al, 2024 |
Table 4.
Exosome-Mediated Therapy Resistance Mechanisms.
| Mechanism | Exosomal Component | Cancer Type | Effect on Therapy | Reference |
|---|---|---|---|---|
| Immune checkpoint failure | miR-146a (↓) | HPV+ HNSCC | Dsg2 (↑) IL-8 (↑) Anti-PD-1 resistance |
Hill et al, 2023 |
| Chemoresistance, relapse | miRNA from CSC-derived exosomes | TSCC (HPV+) | miRNA-driven resistance | Gupta et al, 2021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



