
Submitted:
24 July 2025
Posted:
25 July 2025
You are already at the latest version
Abstract
A growing body of evidence indicates that freshwater bodies, particularly eutrophic systems, can serve as significant sources of the greenhouse gases (GHGs) carbon dioxide (CO₂), methane (CH₄), and nitrous oxide (N₂O). In contrast to marine environments, freshwater systems are typically shallower and more directly influenced by terrestrial inputs, including elevated nutrient loading, increased organic matter deposition, and steeper redox gradients in both the water column and sediments. These conditions foster intensive phytoplankton growth and stimulate microbial processes that drive GHG production and emission. This article explores the biogeochemical mechanisms underlying these emissions and assesses the potential of mitigation treatments to harness phytoplankton populations for carbon sequestration and for reducing CH₄ and N₂O release.
Keywords:
carbon dioxide
; methane
; nitrous oxide
; greenhouse gases
; global warming
; eutrophic waterbodies
; harmful algal blooms
; carbon sequestration
; photosynthesis
; respiration
; sediments
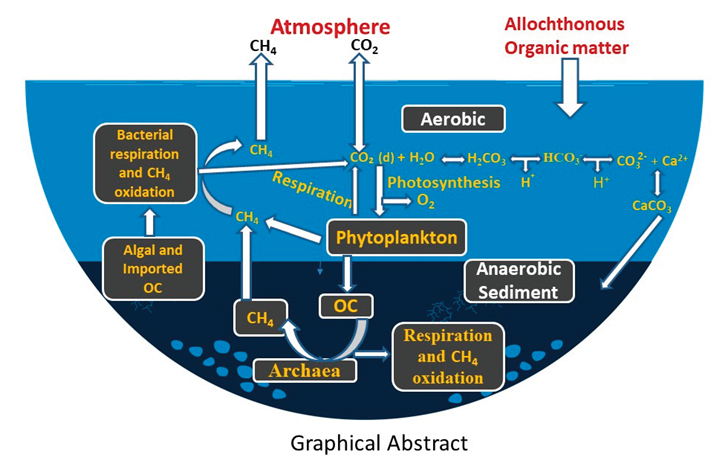
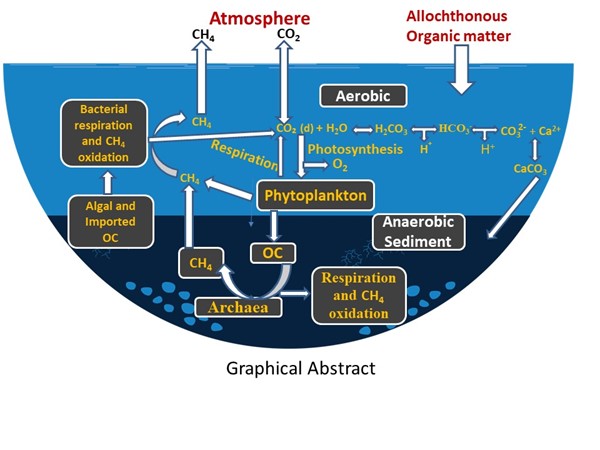
Conceptual schematic of biogeochemical processes in a eutrophic freshwater body during a phytoplankton bloom. Photosynthesis removes CO₂ and produces O₂, supporting aerobic respiration and methane (CH₄) oxidation in surface waters. Anaerobic sediment layers promote CH₄ production by Archaea using organic carbon (OC), with additional CH₄ potentially formed by phytoplankton in oxic zones. Both external and algal-derived OC are subject to mineralization. Dissolved CO₂ equilibrates with HCO₃⁻ and CO₃²⁻, facilitating CaCO₃ precipitation in the presence of Ca²⁺, especially under high pH, with calcite settling to the sediments. CH₄ and CO₂ emissions to the atmosphere are also shown. Local environmental factors govern the magnitude and direction of these fluxes.Note: Nitrous oxide (N₂O) fluxes are omitted for clarity.
1. Introduction
The rising atmospheric CO₂ concentrations from pre-industrial levels of about 280 ppm to over 420 ppm today, primarily due to fossil fuel combustion and deforestation, and that of other (methane and nitrous oxide) greenhouse gases (GHG) is widely recognized as a major driver of global warming. Along with other GHGs, elevated CO₂ levels contribute to a range of adverse effects, including rising global temperatures, ocean acidification, extreme weather events, and loss of biodiversity, all of which have serious implications for both human and ecological systems. International agreements, such as the Paris Agreement, aim to limit the global temperature increase to well below 2°C above pre-industrial levels. Achieving this target requires not only substantial reductions in ongoing emissions but also active removal of existing GHG from the atmosphere. However, despite global efforts and commitments, the concentration of GHG continues to rise and the trend is accelerating highlighting the urgent need for more effective and immediate action.
Various frameworks have been developed to sequester atmospheric CO₂, broadly classified into two main categories: engineered technologies and nature-based solutions. Engineered approaches such as direct air capture, carbon mineralization, and geological storage typically involve substantial energy requirements, complex infrastructure, and extended development timelines. In contrast, nature-based solutions leverage ecological processes like photosynthesis to remove atmospheric CO₂, generally with lower energy demands and a range of ecological co-benefits. Beyond their role in climate mitigation, biologically driven strategies also support biodiversity conservation, water quality improvement, and ecosystem resilience [1]. Nevertheless, despite the progress achieved through existing initiatives, the total volume of CO₂ sequestered to date remains far below the scale needed to significantly alter the atmospheric carbon trajectory, highlighting the magnitude of the challenge that lies ahead.
1.1. Intensification of Harmful Cyanobacteria Blooms (HCB)
One of the consequences of global warming is the intensification of (HCBs) during the last few decades and their spreading to relatively cold regions where such bloom events were previously unheard of. The blooms capitalize on the elevated dissolved CO₂ concentrations, a rising nutrient supply due to leaching from agricultural fields or poorly treated wastewater and the warming lakes temperatures [2,3,4,5,6] enhancing their photosynthetic activity and enabling them to produce massive, and frequently toxic, biomass [4]. These blooms severely affect the water quality in thousands of waterbodies in both developed and underdeveloped countries and hence the wellbeing of people, wildlife and the biodiversity of infected water bodies. It is estimated that 4.4 billion people currently lack access to clean and toxin-free drinking water worldwide [7].
Another consequence of global warming and HCB is a dramatic decline in the biodiversity of infected waterbodies where toxic cyanobacteria represent over 90% of the photosynthetic organisms. It is widely recognized that, in addition to nutrient availability and various abiotic conditions, both inter- and intra-species interactions, particularly allelopathy, play a significant role in shaping the phytoplankton community composition [8,9,10,11,12]. One example is growth experiments that demonstrated interspecies competition between the dinoflagellate Peridinium gatunense and the cyanobacterium Microcystis aeruginosa which strongly inhibit one another, with the final cell abundance and community composition determined by their initial inoculum ratios and the temperature [13,14]. Since, in most cases, the optimal temperature for dinoflagellate, green algae and diatoms growth is lower than that of toxic cyanobacteria strains, the warming winters plays a major role determining the seasonal succession in freshwater systems and in the intensification of HCBs [15]. This is further aggravated by a larger fraction of cyanobacteria cells that overwinter the warmer conditions [16,17,18] and serve as an inoculum for the HCB population in the spring. The competitive advantage of the larger cyanobacteria population is, in our opinion, one of the main reasons for the HCB intensification over the past five decades.
1.2. Shallow Lakes Are Hotspots for HCB
Shallow eutrophic lakes are more vulnerable to massive phytoplankton blooms compared to deeper lakes. Examples include the (many) lakes in the drainage basin of the Yangtze River in China, Lake Okeechobee in Florida, parts of Lake Victoria in Africa and many others. The main reasons being that the nutrients in the sediments are more accessible to the phytoplankton, closer to the photic zone compared with deeper lakes where they may sink to the hypolimnion and become unavailable for the rest of the summer, due to stratification, awaiting for the forthcoming water mixing in the winter. In shallow water bodies, where the smaller volumes warm up more rapidly in the spring [17], cyanobacteria which thrive in warm conditions gain an early-season advantage, allowing them to establish dominance and outcompete other algae.
Given the strong link between rising atmospheric CO₂ and the growth of HCBs, we examine how mitigation treatments that promote biomass sinking may enhance both organic and inorganic carbon sequestration. We also explore strategies to reduce other GHGs emissions from freshwater bodies, focusing on key processes that govern GHG formation and exchange with the atmosphere. Rather than a comprehensive literature review, we highlight mechanisms affected by algal bloom mitigation, as illustrated in the Graphical Abstract. This approach is especially relevant for eutrophic waterbodies burdened by HCBs, offering a potential pathway to reduce their environmental impact while contributing to climate change mitigation.
2. Sequestration of Organic C
In aquatic systems, organic carbon (OC) is produced as part of the “biological CO₂ pump” [19,20,21], whereby atmospheric CO₂, dissolved in the sunlit upper layers of the water column, is converted into organic matter through photosynthesis and associated cellular processes. In addition to this autochthonous production, allochthonous organic matter can also enter the waterbody from the surrounding terrestrial environment. A portion of the OC descends to the sediments, contributing to long-term carbon sequestration, while another portion undergoes microbial decomposition and respiration, releasing CO₂ and methane (see below) completing the carbon cycle (Graphical abstract).
2.1. Are Lakes a Source or a Sink of CO₂?
There are two main sources of CO₂ within the aquatic environment, OC decomposition and calcification (Graphical abstract). Some of the OC produced is buried in the sediment and hence, following thermodynamic principles, lakes should be regarded as a sink for atmospheric CO₂. However, lakes play a complex role in the global carbon cycle, acting as both sources and sinks of CO₂ depending on their location, the season, time of the day and various environmental factors [3,22,23,24,25]. Eutrophic lakes emit significantly more greenhouse gases per unit surface area than oligotrophic lakes, though likely less per cell due to self-shading effects on redox conditions. Model-based estimates [22] suggest lakes globally contribute up to 20% of fossil fuel-equivalent CO₂ emissions. While this may seem to contradict the second law of thermodynamics - given that some photosynthetically fixed carbon is buried rather than respired. The key lies in allochthonous organic carbon input [26,27,28]. Terrestrial carbon, fixed on land and washed into lakes, is degraded and adds to CO₂ and methane emissions, complicating the system’s carbon balance despite signs of in-lake CO₂ drawdown through alkalization and elevated dissolved oxygen (DO) concentration (see below).
2.2. Burial Efficiency (BE) of Organic Carbon
Notably, cyanobacteria produce large quantities and a wide variety of extracellular polysaccharides (EPS), which exert significant influence on their environment, including in biological desert soil crusts, rice paddies, and aquatic systems [29,30,31,32,33]. During intensive blooms, EPS facilitate cell-to-cell adhesion and colony formation, promoting the aggregation of phytoplankton into particulate organic carbon (POC) that sinks toward the sediments as “marine snow” or “flocs.” The size of these particles impacts both their sedimentation rate [34] and internal oxygen dynamics; larger flocs exhibit slower oxygen diffusion, potentially leading to anoxic microenvironments within them. Consequently, anaerobic conditions may develop within the flocs delaying biomass degradation by bacterial respiration. In addition, the EPS also affect anaerobic decomposition, condensation of the sediment and mineralization [35,36] by aiding sediment compaction and maintaining structure. Their decomposition produces extracellular organic compounds that inhibits degrading bacterial communities delaying the breakdown of settled cyanobacterial biomass and methane formation, at least in rice fields [37,38,39].
2.3. Distinguishing Factors in Carbon BE: Lakes vs. Oceans
One of the long-standing dogmas in the field has been that dispersal, grazing by fish and zooplankton and bacterial respiration consumes a large portion of the aggregates, releasing most of the fixed CO₂, while the particles sink to the bottom and during further degradation in the sediments [3,40]. This is indeed the case in the marine environment where only about 1% of the OC produced in the photic zones reach the sediments at >1 km depth [34,41,42] where it remains buried for geological timescale [34,43]. In contrast, detailed studies performed on fresh waterbodies, lakes and reservoirs, spanning various environmental condition, suggested a much larger BE then in the marine environment [26,44,45]. One example is a study performed by the US Geological Survey on 697 waterbodies in continental USA that concluded that the average BE in water reservoirs was 58% [46].
We must conclude that the long-standing dogma that "lakes’ OC is much less permanent than in oceans" doesn’t withstand empirical evaluation. The results obtained, particularly from eutrophic freshwater bodies show just the opposite. Naturally, the question arises - what distinguishes marine systems from freshwater environments as reflected by the much higher BE in the latter. To clarify this important point, it is essential to reveal the processes driving OC degradation within the waterbody which ultimately determine the OC fraction that reach and then buried in sediments. Briefly, the primary factors influencing these processes are the duration of exposure to oxygen, temperature, and the biomass density.
2.3.1. Exposure to Oxygen and the Temperature
Seminal studies by Sebastian Sobek and colleagues [26,40,45] demonstrated that the BE of POC is strongly influenced by duration of exposure to oxygen during descent to the sediments and by temperature. Prolonged exposure to O2 and higher temperatures result in lower BE, primarily due to bacterial respiration which consumes OC and nutrients released from lysing cells. Since the oceans are much deeper than lakes the aggregates sinking to the sediments are subjected to grazing and bacterial respiration for a considerably longer duration than in lakes. Further, in shallow lakes, OC particles often reach anaerobic conditions in the sediments within a few meters from the water surface much faster than in the ocean [40,47]. In addition, the massive biomass often observed in eutrophic freshwater lakes, with substantial amount of EPS, forms larger flocs than marine snow. As mentioned, the larger the flocs are the slower is their bacterial degradation.
Furthermore, the strong currents characteristic of oceanic continental shelves where nutrient inputs fuel intensive algal blooms transport substantial amounts of oxygen to deeper coastal layers, thereby enhancing OC degradation. Ultimately, in addition to the effects of temperature and oxygen [40,41], large spatial variability was observed in the amount of OC reaching specific sediment sites [45] likely reflecting the impact of currents and changing wind directions on local algal abundance. In densely populated waterbodies, the depth of the photic zone (where net oxygen production occurs) is attenuated, particularly in shallow lakes were turbulence by wind or currents may raise sediment particles [48]. Consequently, anoxic condition may develop within a short distance from the water surface [17].
2.3.2. Density of the Phytoplankton Biomass
As indicated, the biomass density is an important parameter affecting the oxygen gradient profile and hence OC degradation. Although blooms intensity increased significantly in the open, oligotrophic, ocean during the last century [49], the overall cell density of phytoplankton biomass is rather low, equivalent to approximately 0.1 - 0.3 mg chlorophyll/m³. It may reach 5-10 mg chlorophyll/m³ during spring-summer blooms mostly near coastal nutrient-rich areas. In contrast, the cell density in eutrophic freshwater bodies can easily exceed 300-500 mg chlorophyll/m³, particularly during HCB that form scums on the water surface [50,51], (https://portal.edirepository.org/nis/mapbrowse?packageid=edi.1756.1). Chlorophyll extraction methodologies, along with recent advances in remote sensing technologies, have made it easier and more accessible to assess chlorophyll concentrations across large areas. Where evaluations of carbon sequestration are required, conversion factors for estimating dry matter from chlorophyll concentrations [52,53] can be employed. However, remote sensing primarily assesses surface phytoplankton populations, while their spatial (including three-dimensional) and temporal distribution is, in most cases, highly variable. Therefore, ground-truthing is essential to accurately calibrate remote sensing measurements. It is timely to develop robust models that integrate remote sensing data with in situ measurements to capture the spatial and 3D distribution of phytoplankton, enabling comprehensive, whole-lake assessments. Alternatively, biomass can be estimated by systematically collecting and analyzing samples from multiple locations and depths. However, this method is labor-intensive, costly, and subject to inherent logistical limitations.
2.4. Sequestration of Organic Carbon in the Marine and Freshwater Bodies
Geologically speaking, it is widely recognized that sediments derived from phytoplankton biomass have played a significant role in the formation of kerogen, a key intermediate in the transformation of organic carbon into natural gas and other fossil fuels. This highlights their crucial contribution to the global carbon cycle over geological timescales. For instance, a recent review [54] proposed a model describing cyanobacterial proliferation and the preservation of organic matter, emphasizing their critical role in organic carbon burial throughout Earth’s history.
This perspective raises a fundamental question: to what extent can the permanence of OC in a given waterbody be predicted based on its environmental conditions and biological productivity? The concept of “burial permanence”- the length of time that OC remains sequestered in sediments is inherently complex as it involves understanding the dynamic processes that govern the stability and longevity of OC [55]. Radiometric dating of sediment cores using isotopes such as ¹⁴C, ¹³⁷Cs, and ²¹⁰Pb is commonly employed to assess the permanence. These analyses provide a detailed reconstruction of the waterbody’s depositional history and have been published in numerous peer-reviewed studies [44,56,57,58,59,60,61], governmental agencies reports (e.g., USGS SIR 2004-5184) and large-scale research initiatives like New Zealand’s Lake380 project. Naturally, the preservation of OC in the sediment is strongly influenced by local environmental conditions. In all cases we are aware of, sediment cores show a pattern of rapid OC degradation during the first few years, followed by a much slower breakdown phase, highlighting a stabilization process in the deeper sediment layers.
The reader is encouraged to explore the Lake380 project available on their website and emerging publications thereof. The data represents one of the most extensive and detailed waterbody analyses, over a range of ambient condition, conducted globally. The sediment cores collected as part of this effort offer invaluable data on both contemporary and historical water quality, encompassing a broad range of biotic and abiotic parameters. In particular, the use of pigment extraction and DNA sequencing along sediment cores serves as a powerful approach for distinguishing between, and reconstructing of algal, shrub and trees-originated biomass over time. Interestingly, the algal originated OC remains fairly constant over tens of thousands of years. This is probably due to various processes taking place in the sediment, where the buried OC undergoes physiochemical processes such as condensation, mineralization, and compaction, ultimately transforming into stable hydrocarbon structures, commonly known as kerogen [37,38,39,54]. Altogether, offering strong evidence for the capacity of freshwater systems to sequester OC over long periods.
2.5. Can Mitigation of Phytoplankton Blooms in Freshwater Bodies Be Used for Substantial Carbon Burial?
Phytoplankton blooms in eutrophic freshwater systems hold promise for OC sequestration [62]; however, their long-term effectiveness and sustainability remain active areas of research and debate. Under business-as-usual conditions, a substantial portion of bloom-derived carbon is mineralized in the water column (see Graphical abstract), thereby limiting BE. In contrast, exposure to mild but prolonged oxidative stress has been shown to trigger extensive programmed cell death (PCD)-like process in cyanobacteria resulting in cell mortality rates as high as 99% [5,63,64,65,66]. This mode of collapse minimizes the release of cyanotoxins into surrounding waters [63,67,68].The resulting biomass forms large aggregates that sink rapidly to the sediment. As reported by Sobek and others [26,40,45], faster sedimentation rates are strongly correlated with increased BE. A key working hypothesis is that PCD-induced massive bloom collapse creates localized anoxia in bottom waters near the sediment, which in turn inhibits microbial OC degradation via respiration. Over the longer term, removal of toxic cyanobacteria enables phytoplankton succession, promoting biodiversity and renewed oxygenic photosynthesis in the water column even in the face of a dramatic decline in cyanobacterial cell abundance. One additional and noticeable benefit of mitigation-induced bloom collapse is a marked improvement in water clarity, enhancing light penetration throughout the water column and thereby photosynthetic oxygen production. Taken together, the BE under these treated conditions may significantly surpass that observed under untreated, baseline scenarios.
Researchers involved in carbon harvesting from phytoplankton blooms may utilize commercial systems that measure Total Organic Carbon (TOC) via combustion and CO₂ detection, followed by acidification to quantify Total Inorganic Carbon (TIC) enabling full TIC/TOC profiling across depths and locations. Remote sensing offers a complementary approach by estimating chlorophyll-a as a proxy for biomass. However, assessing carbon flux to sediments post-mitigation remains challenging due to the spatial and temporal variability in HCB biomass, influenced by factors like wind mixing, stratification, light, and nutrients. Addressing this requires an integrated strategy combining remote sensing, in-situ measurements, and modeling as mentioned in Chapter 2.3.2, above. While chlorophyll-a accounts for 0.6–1.6% of dry biomass [69] under laboratory conditions, field values may vary. It is recommended that chlorophyll extraction be used to assess its level and carbon content assessed by measuring CO₂ released during combustion.
3. The Fate of Inorganic Carbon in the Marine- and Fresh- Waterbodies
3.1. The CO₂-Concentrating Mechanisms (CCMs)
Cyanobacteria and algae have evolved sophisticated CCMs that enable them to accumulate Dissolved Inorganic Carbon (DIC) within their cells, particularly under CO₂-limiting conditions [70,71,72,73,74]. They can thus be regarded as DIC containing bags, intracellular levels as high as 50 mM DIC were recorded [72]. These mechanisms allow the organisms to elevate the internal CO₂ concentration in close proximity to their carboxylating enzyme, ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco), which is predominantly localized within carboxysomes in cyanobacteria and pyrenoids in algae. This adaptation enables them to overcome the large gap between Rubisco’s relatively low affinity for CO₂ and the low concentration of dissolved CO₂ in equilibrium with the atmosphere, particularly in cyanobacteria, where the Km(CO₂) of Rubisco is approximately tenfold higher than ambient levels. The size and dynamics of the internal DIC pool are strongly influenced by environmental factors, especially the CO₂ concentration during growth and light intensity[70,72,75], and they vary among species and even among strains, including Microcystis sp. [76,77].
Seminal studies by Elke Dittmann, Martin Hagemann, and colleagues [78,79,80] revealed a non-canonical CCM in toxic Microcystis sp., where a significant part of Rubisco is located outside the carboxysomes. This distribution results in a less efficient CCM compared to the classical model cyanobacteria studied previously. Additionally, microcystin, a toxic secondary metabolite produced by Microcystis sp., has been shown to bind Rubisco [81]. Fluctuations in extracellular microcystin levels such as those occurring during cell lysis under stress affect the cytoplasmic localization of Rubisco [79]. These findings support earlier hypotheses [82] and provide a mechanistic framework for how cells might sense extracellular microcystin levels and adjust their physiology to cope with environmental stress, thereby enhancing ecological fitness.
A recent study [83] further demonstrated that a similar non-canonical CCM may also operate in the filamentous cyanobacterium Nostoc punctiforme. Here, a poorly functioning CCM necessitates a reliance on heterotrophic bacteria for CO₂ supply under limiting DIC conditions, a surprising dependency that could explain the difficulty in isolating axenic cultures of various cyanobacteria and suggests that elevated ambient CO₂ levels may be critical for their isolation and cultivation. Finally, a quantitative analysis of the internal DIC (and calcite) pool is essential to fully understand the operation and regulation of non-canonical CCMs and their effect on intracellular DIC accumulation [72]. In addition to enhancing the apparent photosynthetic affinity for CO₂, the substantial intracellular accumulation of DIC plays a pivotal role in calcification processes across various algal groups.
3.2. Calcification Processes
The example shown in Figure 1 illustrates that during HCBs, intense photosynthetic activity in the upper layers of the water column, where light is most available, is commonly accompanied by water alkalization. As CO₂ is rapidly drawn down and fixed through photosynthesis, pH levels can exceed 10.0, reflecting a substantial deviation of carbonate chemistry from equilibrium with atmospheric CO₂ (Fig. 1A). Elevated concentrations of dissolved oxygen (DO) in the surface layers (Fig. 1B), exceeding levels expected under equilibrium with air, further support the conclusion that this alkalization is driven by vigorous photosynthetic activity. This alkalization facilitates the precipitation of calcium carbonate, particularly in species with mucilaginous sheaths that promote localized supersaturation, leading to calcite deposition [84,85,86,87,88,89,90].
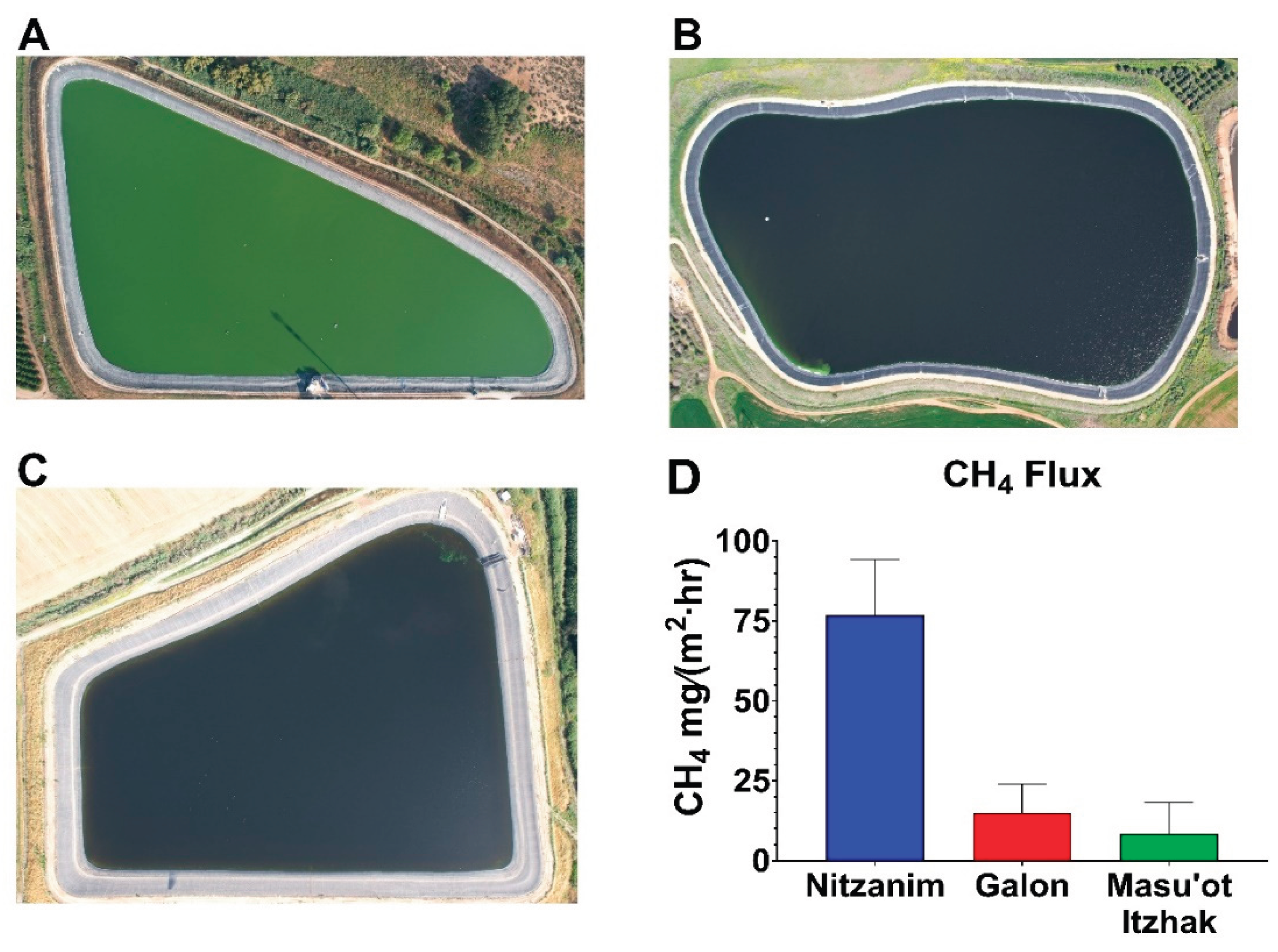
Figure 2.
Aerial photographs of the Nitzanim (A), Galon (B), and Masu’ot Itzhak (C) reservoirs, showing the relative phytoplankton (HCB) intensity, along with corresponding methane emission fluxes (D). Methane emissions were quantified using an ABB GLA131 laser-based gas analyzer connected to a custom-built floating flux chamber of known volume and surface area.
Figure 2.
Aerial photographs of the Nitzanim (A), Galon (B), and Masu’ot Itzhak (C) reservoirs, showing the relative phytoplankton (HCB) intensity, along with corresponding methane emission fluxes (D). Methane emissions were quantified using an ABB GLA131 laser-based gas analyzer connected to a custom-built floating flux chamber of known volume and surface area.
CaCO₃ accumulation in bloom-forming Microcystis sp. has been documented [87,91,92], although quantification of the calcite levels under varying environmental conditions remains incomplete [89,93,94,95,96,97,98]. Given that cyanobacterial cells can be regarded as DIC-bearing bodies, there is potential to employ molecular tools to augment DIC accumulation and, consequently, inorganic carbon sequestration [99,100]. Key targets for such genetic interventions may include bicarbonate transporters (SbtA, BCT1, and BicA), as well as the NDH-1 complexes involved in CO₂ recapture from leaking pools [71,101]. While the environmental release of genetically modified organisms is subject to regulatory constraints, evaluating their capacity for enhanced DIC accumulation and cycling [102] under simulated bloom conditions could provide valuable insights into their potential for carbon capture applications.
Calcification in aquatic systems involves the reaction of Ca²⁺ with HCO₃⁻ to form calcium carbonate (CaCO₃), releasing CO₂ (and water, Graphical abstract). This process can lead to CO₂ emissions from both freshwater and marine environments [89,90,96,103,104]. However, the accumulation of calcium carbonate in sediments suggests a net uptake of DIC over time, as atmospheric CO₂ dissolves in water to replenish bicarbonate consumed during calcification. In freshwater bodies, significant calcification often occurs in areas with high alkalinity, hard water or during substantial phytoplankton blooms. Intensive CO₂ uptake during phytoplankton blooms raises the pH to values where most of the inorganic carbon is in the form of carbonate, promoting calcite precipitation [105]. The Sea of Galilee (Lake Kinneret), a stratified hard-water lake, provides a notable example. It was estimated that over 52,000 tons of calcite settles into the sediment during the spring blooms of the dinoflagellate Peridinium gatunense [93,106]. Mineralization processes within the sediment also lead to carbonate formation particularly during intensive algal blooms [107]. Here, calcification is driven by the high level of dissolved CO₂ consequent on intensive bacterial respiration. It is estimated that carbonate formation through mineralization at the sediment accounts for 3–8% of the deposited OC. However, distinguishing this carbonate from that settling onto the sediments from the top water layer remains challenging [35]. Attempts to resolve this issue using δ¹³C would be complicated by the disequilibrium between CO₂ and bicarbonate species at both the sediment surface and in the photic zone. This disequilibrium, driven by a substantial pH gradient, significantly shifts the δ¹³C values of these species away from the equilibrium value of approximately 7‰ [108].
Naturally, the amount of carbon buried through calcification is strongly influenced by various factors, with calcium, magnesium and proton concentrations playing a particularly significant role. These concentrations vary widely between different lakes and depth in the water column (Fig. 1A) and affects both calcification and dissolution processes. Unfortunately, detailed analyses of the amount of carbonate reaching the sediments, whether due to calcification in the epilimnion or mineralization of OC, and its subsequent fate, including the extent of dissolution [104], remain scarce. Direct measurements of descending carbonates using sediment traps and cores provide valuable insights, though these techniques are labor intensive and costly with substantial variation observed among replicates. Alternatively, an assessment of calcium and magnesium cycles within the waterbody through measurements of their levels in the epilimnion, hypolimnion, as well as inflowing and outgoing streams throughout the year offers a robust approach to evaluate calcification and dissolution dynamics. In the case of Lake Kinneret, analysis [109] revealed that approximately one-third of the calcite formed during the spring bloom undergoes dissolution, while the remaining portion remains buried (Nishri, personal communication). Such assessments are crucial for evaluating carbon fluxes in aquatic environments, understanding their role in the global carbon cycle, and exploring potential pathways for effective carbon sequestration.
A Challenge: Can We Use Changing pH to Assess the Amount of Atmospheric CO₂ Dissolution?
The extent of pH rise during intensive CO₂ consumption by photosynthesis (Fig. 1A) is strongly influenced by the alkalinity of the waterbody and the chemical nature of its buffering components. Higher alkalinity results in a smaller pH increase for a given amount of CO₂ withdrawn. In principle, alkalinity can be used to estimate the amount of CO₂ required to reverse the pH across a given range, such as restoring pre-bloom conditions, assuming equilibrium among DIC species during the process. However, this equilibrium-based assumption can be misleading. Unlike oceanic systems, eutrophic lakes often exhibit significant disequilibrium among DIC species, driven by dynamic biological and chemical processes, most notably the intensive removal of CO₂ during photosynthesis and the precipitation of carbonates at elevated pH. The latter process is especially pronounced when calcium and magnesium ions are sufficiently available leading to effective removal of DIC from the water column and further complicating efforts to model or quantify DIC speciation accurately. To address this complexity, we propose a time-resolved modeling approach based on observed pH dynamics, incorporating non-equilibrium processes that may provide a more accurate framework to estimate CO₂ fluxes.
When a harmful cyanobacterial bloom (HCB) is abruptly terminated—for example, through mitigation treatment—the cessation of photosynthetic CO₂ uptake permits dissolved CO₂ in the water to re-equilibrate with the atmosphere, typically resulting in a gradual decline in pH toward pre-bloom levels. The rate of atmospheric CO₂ dissolution (E) is influenced by abiotic factors such as temperature, wind, turbulence, and vertical mixing. Additional contributors to the rising CO₂ concentration include respiratory CO₂ production (B), the absence of photosynthetic uptake (C), and shifts in the distribution of dissolved inorganic carbon (DIC) species as the system rebalances with changing pH (D).The relationships between these fluxes can be described as:
where (A) is the amount of CO₂ required to reduce the pH over a given range
A = (B – C) + D + E
A key challenge is whether we can quantitatively distinguish between the contributions of each of these processes, in order to extract the net amount of CO₂ dissolved from the atmosphere following bloom collapse. Potential approaches include measuring the initial rate of pH decline in sealed water samples (excluding air exchange) may provide insight into non-atmospheric CO₂ contributions (e.g., from respiration and internal redistribution). Early time points are critical since prolonged incubation can result in substantial pH reduction by microbial respiration alone, especially if oxygen and organic carbon are available. A more robust method involves quantifying DIC directly by acidifying collected samples, which converts all carbonate species to CO₂ for measurement. This bypasses the reliance on pH-alkalinity modeling alone. We acknowledge that spatial heterogeneity, both horizontal and vertical, and temporal fluctuations in pH and alkalinity (see Fig. 1A) are inherent to the dynamics of bloom development and collapse. These variations are also reflected in local dissolved oxygen levels (Fig. 1B), which integrate the combined effects of photosynthesis, respiration, and gas exchange with the atmosphere. To account for these variabilities, representative sampling across depth profiles, followed by volumetric integration over the entire waterbody, is recommended.
4. The Methane Enigma
The escalating methane (CH4) level in the atmosphere is a matter of growing concern due to its projected impact on global warming, surpassing CO₂ (per mol) by approximately 30-fold. It is now firmly established that waterbodies constitute significant contributors to atmospheric methane [110,111,112,113,114,115,116,117,118]. Large spatial and temporal variations in methane emissions were revealed. Using Eddy covariance approach over small waterbodies Hounshell and colleuges [119] and Waldo et. al. [120] showed that under a business-as-usual conditions eutrophic lakes release 1.3-51.4 g CH4 m−2 yr−1, respectively. Unfortunately, this methodology that measures the vertical fluxes of gases (such as CO₂ and CH4) and monitors fluctuations in wind speed and directions above the ecosystem (aquatic or terrestrial), is currently applicable to small waterbodies only. One of the challenges is to capture methane that dissolves in the water-column where a significant portion can escape via ebullition directly into the atmosphere. Buoyant flow chambers enables measurement of the methane emission using its infrared absorption bands or gas chromatography [121].
4.1. Methane Biosynthesis
The mechanisms of methane biosynthesis and degradation are being resolved with recent findings challenging longstanding paradigms and potentially unveiling new strategies to mitigate CH4 emissions. Biogenic CH4 is primarily produced under anaerobic conditions by several archaea groups known as methanogens. They can combine CO2 and hydrogen to form methane in a reaction that consumes the OC produced within the lake, as well as from allochthonous inputs from its surroundings. Additional important methane producing pathways include methylotrophic bacteria that can convert methanol to methane and CH4 production by phytoplankton under oxic condition (discussed below). In addition to O2 level, methane formation is also affected by temperature though in some case less than expected since presence of various methanogens whose optimal activities vary across a wide temperature range.
Biogenic methane has the lowest isotopic carbon ratio compared to other natural sources because it is extremely depleted in 13C [122]. Its δ¹³C value, typically ranges from -50‰ to -110‰ (relative to the Vienna Pee Dee Belemnite standard, VPDB), is the main diagnostic tool that distinguish it from other methane sources. This highly depleted isotopic signature is due to the preferential use of the lighter carbon isotope (¹²C) by methanogenic archaea during microbial methanogenesis, particularly in the reduction of CO₂ or acetate to methane via the two main pathways involved. However, studies utilizing methane to assess the efficacy of CH₄-derived carbon as a substrate for phytoplankton [123,124] should bear in mind that ¹³C discrimination also occurs during CO₂ fixation, and that the CO₂-concentrating mechanism in phytoplankton raises the utilization of ¹³C by the carboxylating enzyme [72].
The Methane Paradox
Until recently, the prevailing belief was that under business-as-usual conditions, most CH₄ produced in anoxic sediments is consumed by methanotrophs in the overlying oxic layers. This paradigm was supported by laboratory experiments and studies in small-scale aquatic systems, but recent research challenges this view. A growing body of evidence indicates that various phytoplankton inhabiting the photic, oxygenated water layers, including cyanobacteria, can produce significant amounts of CH₄ during photosynthesis [112,125,126,127,128,129,130,131,132,133]. This aligns with earlier studies on aerobic CH₄ biosynthesis by bacteria [134].
Collectively, these findings contribute to the recognition of the "methane paradox," wherein CH₄ concentrations in the oxic surface layers of lakes and coastal ocean regions rise toward the top water layer and can surpass levels expected at saturation with air [124,132,135]. The specific organisms involved in freshwater and oceanic methane production are being identified [131,135,136], and the exact metabolic pathways are under intensive investigation. Recent studies have demonstrated that CH₄ formation is light- and photosynthesis-dependent, though the precise mechanisms remain poorly understood [131]. It is hereby hypothesized that CH₄ production may serve as a safety valve to release excess reducing equivalents in the photosynthetic machinery during high illumination or other stress conditions.
Stable carbon and hydrogen isotope analyses suggest that methylated compounds such as methylphosphonate (MP), methylamine, and methionine serve as precursors for oxic CH₄ production. This process involves a gene cluster (phn) that is widespread in filamentous cyanobacteria and also present in certain strains of toxic Microcystis sp., such as those collected during a large bloom in Dianchi Lake, China [137], emphasizing the role of cyanobacteria in oxic methane formation. Complicating matters further, studies suggest that MP released by lysing cyanobacteria is utilized by bacteria as a phosphate source, thereby releasing CH₄ in the photic zone [137]. Mutations in the C–P lyase phosphonate degradation pathway of the marine bacterium Pseudomonas stutzeri disabled its ability to produce methane [135]. Currently, accurate assessments of the relative contributions of cyanobacteria and eukaryotes to methane formation and utilization in the upper aerobic zone are lacking, highlighting the need for genome screenings and identification of potentially involved genes [130].
4.2. Methane Consumption
Methane is consumed by methanotrophic microorganisms, which utilize it as both a carbon and a reducing power source, thereby mitigating its emission to the atmosphere. The activation of the stable C–H bond in methane is thermodynamically unfavorable, requiring a high redox potential and the transfer of eight electrons [138].
Methane oxidation proceeds via two main pathways: aerobic and anaerobic. These pathways differ significantly in their enzymatic mechanisms, the organisms involved, and their sensitivity to environmental factors such as temperature and redox potential. In both systems, methane is oxidized by forms of methane monooxygenase (MMO) or analogous enzymes, each exhibiting distinct kinetic properties. In aerobic methanotrophs, MMOs utilize molecular oxygen as the terminal oxidant. In anaerobic methanotrophs, however, the enzymes rely on alternative oxidants such as Fe³⁺ or copper. In anoxic aquatic environments, particularly in marine systems, sulfate or nitrate can serve as terminal electron acceptors for methanotrophic respiration. These enable energy production that indirectly supports methane oxidation under oxygen-depleted conditions, thereby contributing to CH₄ removal even in anoxic zones.
As mentioned, the eight-electron transfer involved in aerobic methane oxidation is mediated by specific components of the MMO enzyme complex, which catalyzes the conversion of CH₄ to methanol, a key step in the metabolism of methane by aerobic methanotrophs. In anaerobic methane oxidation, CH₄ is ultimately converted to CO₂ and water through the action of microbial consortia, typically involving anaerobic methanotrophic archaea in partnership with sulfate-reducing bacteria. Where sulfate availability is limited, such as in freshwater lakes, alternative electron acceptors like nitrate, iron or manganese can be utilized.
Theoretically, an oxidation-reduction potential (ORP) above approximately +400 mV is likely required to support meaningful anaerobic methanotrophic activity. For reference, the redox potentials of key electron acceptors used in anaerobic processes, Fe³⁺/Fe²⁺ and NO₃⁻/NO₂⁻ are +770 mV and +940 mV, respectively. It is important to note that in most aquatic systems studied to date, nitrate is often depleted in the oxic (photic) zone but remains abundant in deeper, anoxic layers of the water column. Therefore, measuring the redox potential at the sediment-water interface is recommended to assess the potential for anaerobic CH₄ consumption and of the impact of iron fertilization on anaerobic methane oxidation. In contrast, the redox potential required for aerobic CH₄ oxidation is provided directly by oxygen, acting as the terminal electron acceptor during bacterial respiration. Thus, where possible, simultaneous measurements of ORP and dissolved oxygen are highly recommended to better understand the methanotrophic potential and to evaluate strategies for enhancing methane removal in aquatic systems.
As previously noted, cyanobacteria are increasingly recognized as significant contributors to CH₄ production. However, despite CH₄ being a potential source of both carbon and reducing power and despite cyanobacteria’s capacity to support eight electron transfer processes such as nitrogen fixation, they appear to lack the MMO enzyme necessary for cleaving the stable C–H bond in methane. To date, no published studies have demonstrated the growth of axenic cyanobacterial cultures using CH₄ as the sole carbon source. Nevertheless, growing interest in cyanobacteria–methanotroph interactions has led to co-culture studies that reveal enhanced CH₄ oxidation in the presence of cyanobacteria. This enhancement is likely attributable to increased local oxygen availability provided by cyanobacterial photosynthesis. Conversely, methanotroph presence has been associated with accelerated cyanobacterial growth [123,139]. The exact metabolic contribution of methanotrophs to cyanobacterial physiology remains unclear. Given the highly efficient CCMs in cyanobacteria, it is unlikely that these benefits are due solely to respiratory CO₂ release by methanotrophs, as previously proposed.
In the absence of extensive data from freshwater systems, insights may be drawn from studies on cyanobacteria–methanotroph interactions in rice paddy soil microcosms. In such systems, inoculation with cyanobacteria has been shown to reduce CH₄ emissions by up to 90%, with outcomes strongly dependent on the cyanobacterial species used. For instance, Nostoc sp. and Calothrix sp. were found to influence different methanotrophic communities, leading to variable outcomes in methane flux [37].
These emerging studies point to a more complex and nuanced picture of methane dynamics within microbial consortia, highlighting the potential role of species-specific interactions and even secondary metabolites in mediating these effects. Continued exploration of these relationships is essential for understanding their ecological significance and potential applications in methane mitigation strategies.
4.3. Can HCB Mitigation Reduce Methane Emissions?
Naturally, the net flux of CH₄ emissions from waterbodies is governed by the balance between its production and consumption. The significant variability observed in CH₄ emission fluxes across different eutrophic lakes likely reflects differences in local conditions and various parameters affecting this balance. Studies have shown that cyanobacterial blooms can enhance methane emissions through multiple pathways. For instance, research on Lake Tai, China, showed that regions with dense cyanobacterial populations emitted significantly more CH₄ compared to low-density areas, 867 vs. 3.4 μg CH₄ m⁻² min⁻¹, respectively indicating a strong correlation between bloom intensity and methane production [139]. This is an important observation, as it may point to a potential route for decreasing global methane emissions from eutrophic lakes through proper management and control of HCBs.
Three water reservoirs located in the northwestern Negev region of Israel were selected to examine this possibility - Nitzanim (5 ha, Fig. 2A) and Galon (6 ha, Fig. 2B) are filled with tertiary treated recycled nutrient-rich water, whereas Masu’ot Itzhak (8 ha, Fig. 2C) is supplied with water collected from occasional winter floods in a nearby stream. Aerial photographs taken by drone clearly show a dense phytoplankton population in Nitzanim, while phytoplankton presence is barely visible in Galon and Masu’ot Itzhak. Routinely mitigation treatment applied in Galon Reservoir effectively suppressed the HCB that are typically observed in Nitzanim. This difference is further supported by the higher surface pH (Fig. 1A) and dissolved oxygen (DO) concentrations (Figure 1B) in Nitzanim, likely resulting from intensive photosynthetic activity associated with the denser HCB population. Additionally, the steeper vertical decline in both pH and DO in Nitzanim suggests stronger light attenuation, and thus reduced oxygen production at depth, caused by the bloom’s density.
Measurements of methane flux (Fig. 2D) revealed substantially higher CH₄ emissions in Nitzanim compared to the other two reservoirs. It remains to be determined whether the lower emissions observed in Galon and Masu’ot Itzhak are primarily due to reduced CH₄ production by cyanobacteria or to a lower availability of organic carbon in the sediments, which serves as a substrate for methanogenesis. Nevertheless, these preliminary findings are encouraging and underscore the need for further research to elucidate how local environmental factors shape the balance between methane production and consumption, ultimately influencing net emissions. Further, the emerging correlation between eutrophic conditions, bloom intensity, and methane release suggests that bloom-targeted mitigation strategies, such as those employed in Galon Reservoir, may offer an effective approach to reduce CH₄ emissions from eutrophic waterbodies.
5. Nitrous Oxide in Freshwater Bodies
In addition to its detrimental effects on the ozone layer, nitrous oxide (N₂O) is a potent greenhouse gas (GHG), with a global warming potential approximately 270 times greater than that of CO₂. In recent years, freshwater lakes have been recognized as significant sources of atmospheric N₂O emissions [22,140,141,142]. Cyanobacteria and microalgae have been implicated in N₂O biosynthesis in aquatic environments [143,144,145,146].
The primary contributors to N₂O production in waterbodies are microbial processes, particularly ammonia oxidation and denitrification. In oxic waters, ammonia-oxidizing Archaea and Bacteria release N₂O as a byproduct during the oxidation of ammonia to nitrite. Under oxygen-depleted conditions, N₂O also serves as a key intermediate in the denitrification pathway, in which nitrate and nitrite are sequentially reduced to molecular nitrogen (N₂), often coupled with the oxidation of organic matter.
Phytoplankton blooms can significantly influence N₂O dynamics by altering the structure and extent of oxic–anoxic transition zones (Fig. 2D), such as at the sediment–water interface or within stratified water columns. These zones are critical sites for microbial nitrogen transformations, and shifts in redox gradients can determine whether the dominant end-product of nitrogen metabolism is benign N₂ or climate-active N₂O.
Despite its environmental significance, mechanistic understanding of N₂O synthesis and degradation in aquatic systems is poorly understood. Recent research is unfolding the complex interplay of biotic and abiotic pathways that govern N₂O fluxes [147]. Gaining a deeper understanding of these mechanisms is essential for improving nitrogen cycle models in eutrophic waterbodies and developing strategies to mitigate N₂O emissions from inland waters. Interestingly, recent studies have shown that sunlight can drive abiotic N₂O production in both freshwater and marine systems. This occurs primarily through photochemical reduction of nitrate to nitrite, followed by subsequent reactions leading to N₂O formation [148]. However, the precise mechanisms and global relevance of this pathway are still under investigation.
Of note, some methanotrophs are capable of anaerobic methane oxidation using nitrate or nitrite as electron acceptors, effectively reducing one GHG (CH₄) while potentially producing another, far more harmful (N₂O) through a synergistic metabolic process. As with methane, it remains to be determined whether mitigation treatments targeting algal blooms which can significantly reduce organic matter and alter redox dynamics will also lead to measurable reductions in N₂O emissions to the atmosphere.
6. Concluding Remarks
The intensification of HCBs in eutrophic lakes, primarily driven by anthropogenic nutrient enrichment and global warming has been linked to increased GHG emissions, thereby contributing to global warming. While mitigation strategies targeting HCBs hold considerable promise for substantial reduction of these emissions, a better understanding of the mechanisms involved and environmental factors that regulate them is critical. Advancing such knowledge will not only enhance the effectiveness of current mitigation efforts but also support the development of targeted, science-based approaches for minimizing global GHG emissions across a variety of aquatic ecosystems.
Author Contributions
A.K. and M.H. conceived, designed and wrote the paper.
Funding
This project was supported by the Hebrew University of Jerusalem and BlueGreen Water Technologies.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We wish to thank Dr. Ami Nishri of the Israel Oceanographic and Limnological Research Institute, for many fruitful discussions. We are also grateful to Dr. Gad Weiss and Alon Goldman from BlueGreen Water Technologies (https://bluegreenwatertech.com/) for providing the resources and technical support that made the experiments presented in Figures 1 and 2, possible.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Griscom, B.W.; Adams, J.; Ellis, P.W.; Houghton, R.A.; Lomax, G.; Miteva, D.A.; Schlesinger, W.H.; Shoch, D.; Siikamäki, J.V.; Smith, P.; et al. Natural climate solutions. Proc. Natl. Acad.Sci. USA 2017, 114, 11645–11650. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Wang, Y.; Hou, X.; Qin, B.; Kuster, T.; Qu, F.; Chen, N.; Paerl, H.W.; Zheng, C. Harmful algal blooms in inland waters. Nat. Rev. Earth & Environm. [CrossRef]
- Shi, W.; Qin, B.; Zhang, Q.; Paerl, H.W.; Van Dam, B.; Jeppesen, E.; Zeng, C. Global lake phytoplankton proliferation intensifies climate warming. Nat. Commun. 2024, 15, 10572. [Google Scholar] [CrossRef]
- Visser, P.M.; Verspagen, J.M.H.; Sandrini, G.; Stal, L.; Matthijs, H.C.P.; Davis, T.W.; Paerl, H.W.; Huisman, J. How rising CO2 and global warming may stimulate harmful cyanobacterial blooms. Harmful Algae.
- Sukenik, A.; Kaplan, A. Cyanobacterial harmful algal blooms in aquatic ecosystems: A comprehensive outlook on current and emerging mitigation and control approaches. Microorganisms 2021, 9, 1472. [Google Scholar] [CrossRef]
- Qiu, Y.; Chen, J.; Chen, D.; Thiery, W.; Mercado-Bettín, D.; Xiong, L.; Xia, J.; Woolway, R.I. Enhanced heating effect of lakes under global warming. Nat. Commun. 2025, 16, 3954. [Google Scholar] [CrossRef]
- Greenwood, E.E.; Lauber, T.; van den Hoogen, J.; Donmez, A.; Bain, R.E.S.; Johnston, R.; Crowther, T.W.; Julian, T.R. Mapping safe drinking water use in low- and middle-income countries. Science, /: 784–790, https.
- Kaplan, A.; Harel, M.; Kaplan-Levy, R.N.; Hadas, O.; Sukenik, A.; Dittmann, E. The languages spoken in the water body (or the biological role of cyanobacterial toxins). Front aquatic Microbiol, /: https. [CrossRef]
- Gantar, M.; Berry, J.P.; Thomas, S.; Wang, M.; Perez, R.; Rein, K.S. Allelopathic activity among Cyanobacteria and microalgae isolated from Florida freshwater habitats. FEMS Microbiol. Ecol. 2008, 64, 55–64. [Google Scholar] [CrossRef]
- Oberhaus, L.; Briand, J.-F.; Humbert, J.-F. Allelopathic growth inhibition by the toxic, bloom-forming cyanobacterium Planktothrix rubescens. FEMS Microbiol. Ecol. 2008, 66, 243–249. [Google Scholar] [CrossRef]
- Guo, Z.; Li, J.; Hu, J.; An, G.; Wang, C. Deciphering the joint intracellular and extracellular regulatory strategies of toxigenic Microcystis to achieve intraspecific competitive advantage: An integrated multi-omics analysis with novel allelochemicals identified. Water Res. 2025, 283, 123774. [Google Scholar] [CrossRef]
- Harel, M.; Weiss, G.; Lieman-Hurwitz, J.; Gun, J.; Lev, O.; Lebendiker, M.; Temper, V.; Block, C.; Sukenik, A.; Zohary, T.; et al. Interactions between Scenedesmus and Microcystis may be used to clarify the role of secondary metabolites. Environm. Microbiol. Rep. 2013, 5, 97–104. [Google Scholar] [CrossRef]
- Vardi, A.; Schatz, D.; Beeri, K.; Motro, U.; Sukenik, A.; Levine, A.; Kaplan, A. Dinoflagellate-cyanobacterium communication may determine the composition of phytoplankton assemblage in a mesotrophic lake. Curr. Biol. 2002, 12, 1767–1772. [Google Scholar] [CrossRef] [PubMed]
- Sukenik, A.; Eshkol, R.; Livne, A.; Hadas, O.; Rom, M.; Tchernov, D.; Vardi, A.; Kaplan, A. Inhibition of growth and photosynthesis of the dinoflagellate Peridinium gatunense by Microcystis sp. (cyanobacteria): a novel allelopathic mechanism. Limnol. Oceanogr. 2002, 47, 1656–1663. [Google Scholar] [CrossRef]
- Cai, P.; Wu, H.; Pan, M.; Cao, G.; Wu, X.; Tian, C.; Wang, C.; Xiao, B. Eutrophication amplifies Microcystis response to increasing winter temperatures, intensifying winter blooms. J. Appl. Phycol. 2025. [Google Scholar] [CrossRef]
- Cai, P.; Cai, Q.; He, F.; Huang, Y.; Tian, C.; Wu, X.; Wang, C.; Xiao, B. Flexibility of Microcystis overwintering strategy in response to winter temperatures. Microorganisms 2021, 9. [Google Scholar] [CrossRef]
- Brunberg, A.-K.; Blomqvist, P. Benthic overwintering of Microcystis colonies under different environmental conditions. J. Plank. Res. 2002, 24, 1247–1252. [Google Scholar] [CrossRef]
- Reinl, K.L.; Harris, T.D.; North, R.L.; Almela, P.; Berger, S.A.; Bizic, M.; Burnet, S.H.; Grossart, H.-P.; Ibelings, B.W.; Jakobsson, E.; et al. Blooms also like it cold. Limnol. Oceanogr. Lett. 2023, 8, 546–564. [Google Scholar] [CrossRef]
- Bressac, M.; Laurenceau-Cornec, E.C.; Kennedy, F.; Santoro, A.E.; Paul, N.L.; Briggs, N.; Carvalho, F.; Boyd, P.W. Decoding drivers of carbon flux attenuation in the oceanic biological pump. Nature 2024. [Google Scholar] [CrossRef]
- Jiao, N.Z.; Luo, T.W.; Chen, Q.R.; Zhao, Z.; Xiao, X.L.; Liu, J.H.; Jian, Z.M.; Xie, S.C.; Thomas, H.; Herndl, G.J.; et al. The microbial carbon pump and climate change. Nat. Rev. Microbiol. 2024, 22, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.; Scholes, R.J.; Boyle, E.; Canadell, J.; Canfield, D.; Elser, J.; Gruber, N.; Hibbard, K.; Hogberg, P.; Linder, S.; et al. The global carbon cycle: A test of our knowledge of earth as a system. Science 2000, 290, 291–296. [Google Scholar] [CrossRef] [PubMed]
- DelSontro, T.; Beaulieu, J.J.; Downing, J.A. Greenhouse gas emissions from lakes and impoundments: Upscaling in the face of global change. Limnol. Oceanogr. Lett. 2018, 3, 64–75. [Google Scholar] [CrossRef]
- Beaulieu, J.J.; DelSontro, T.; Downing, J.A. Eutrophication will increase methane emissions from lakes and impoundments during the 21st century. Nature Commun. 2019, 10, 1375. [Google Scholar] [CrossRef]
- Beaulieu, J.J.; Waldo, S.; Balz, D.A.; Barnett, W.; Hall, A.; Platz, M.C.; White, K.M. Methane and carbon dioxide emissions from reservoirs: Controls and upscaling. J. Geophys. Res.:Biogeos. [CrossRef]
- Sakaguchi, J.; Nakayama, K.; Komai, K.; Kubo, A.; Shimizu, T.; Omori, J.; Uno, K.; Fujii, T. Carbon dioxide uptake in a eutrophic stratified reservoir: Freshwater carbon sequestration potential. Heliyon 2023, 9, e20322. [Google Scholar] [CrossRef] [PubMed]
- Sobek, S.; Durisch-Kaiser, E.; Zurbrügg, R.; Wongfun, N.; Wessels, M.; Pasche, N.; Wehrli, B. Organic carbon burial efficiency in lake sediments controlled by oxygen exposure time and sediment source. Limnol. Oceanogr. 2243. [Google Scholar]
- Wang, S.; Nie, X.; Li, Z.; Ran, F.; Yang, C.; Xiao, T. Quantification of sedimentary organic carbon sources in a landeriverelake continuum combined with multi-fingerprint and un-mixing models. Intnl. J. Sedim. Res.
- Du, Z.; Wang, L.; Xie, S.; Yang, J.; Yan, F.; Li, C.; Ding, M.; Zhang, Y.; Ding, X.; Xiao, C. Carbon dioxide and methane fluxes in different waterbodies in Inexpressible Island, Ross Sea, East Antarctica. Mar. Pollut. Bull. 2025, 213, 117703. [Google Scholar] [CrossRef]
- Harel, M.; Weiss, G.; Daniel, E.; Wilenz, A.; Hadas, O.; Sukenik, A.; Sedmak, B.; Dittmann, E.; Braun, S.; Kaplan, A. Casting a net: fibres produced by Microcystis sp. in field and laboratory populations. Environ. Microbiol. Rep. 2012, 4, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Franco-Morgado, M.; Amador-Espejo, G.G.; Pérez-Cortés, M.; Gutiérrez-Uribe, J.A. Microalgae and cyanobacteria polysaccharides: Important link for nutrient recycling and revalorization of agro-industrial wastewater. Appl. Food Res. 2023, 3, 100296. [Google Scholar] [CrossRef]
- De Philippis, R.; Vincenzini, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol Rev 1998, 22, 151–175. [Google Scholar] [CrossRef]
- Rossi, F.; De Philippis, R. Role of cyanobacterial exopolysaccharides in phototrophic biofilms and in complex microbial mats. Life (Basel, Switzerland) 2015, 5, 1218–1238. [Google Scholar] [CrossRef]
- Wu, S.; Wang, F.; Wang, H.; Shen, C.; Yu, K. Meta-analysis of abiotic conditions affecting exopolysaccharide production in cyanobacteria. Metabolites 2025, 15, 131. [Google Scholar] [CrossRef]
- Alcolombri, U.; Peaudecerf, F.J.; Fernandez, V.I.; Behrendt, L.; Lee, K.S.; Stocker, R. Sinking enhances the degradation of organic particles by marine bacteria. Nat. Geosci. 2021, 14, 775–780. [Google Scholar] [CrossRef]
- Ma, Y.; Yi, S.; Wang, M. Biomimetic mineralization for carbon capture and sequestration. Carbon Capt. Sci. Technol. 2024, 13, 100257. [Google Scholar] [CrossRef]
- Zhang, W.; Gu, P.; Zhu, W.; Jing, C.; He, J.; Yang, X.; Zhou, L.; Zheng, Z. Effects of cyanobacterial accumulation and decomposition on the microenvironment in water and sediment. J. Soils Sedim. 2020, 20, 2510–2525. [Google Scholar] [CrossRef]
- Pérez, G.; Krause, S.M.B.; Bodelier, P.L.E.; Meima-Franke, M.; Pitombo, L.; Irisarri, P. Interactions between cyanobacteria and methane processing microbes mitigate methane emissions from rice soils. Microorganisms 2023, 11, 2830. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Zhao, M.; Song, L.; Chen, X.; Zhang, Z.; Gao, N. Characteristics of extracellular organic matters and the formation potential of disinfection by-products during the growth phases of M. aeruginosa and Synedra sp. Environm. Sci. Poll. Res. 2022, 29, 14509–14521. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, H.; Yu, J.; Su, Y.; Liu, Z. Geochemical characteristics of n-alkanes in sediments from oligotrophic and eutrophic phases of five lakes and potential use as paleoenvironmental proxies. CATENA 2023, 220, 106682. [Google Scholar] [CrossRef]
- Sobek, S.; Gudasz, C.; Koehler, B.; Tranvik, L.J.; Bastviken, D.; Morales-Pineda, M. Temperature dependence of apparent respiratory quotients and oxygen penetration depth in contrasting lake sediments. Geophys. Res.: Biogeosci. 3076. [Google Scholar]
- Pavia, F.J.; Anderson, R.F.; Lam, P.J.; Cael, B.B.; Vivancos, S.M.; Fleisher, M.Q.; Lu, Y.; Zhang, P.; Cheng, H.; Edwards, R.L. Shallow particulate organic carbon regeneration in the South Pacific Ocean. Proc Natl Acad USA, 9753. [Google Scholar]
- Martin, J.H.; Knauer, G.A.; Karl, D.M.; Broenkow, W.W. ;. VERTEX: carbon cycling in the Northeast Pacific. Deep Sea Res. 1987, 267–285. [Google Scholar] [CrossRef]
- Dunne, J.P.; Sarmiento, J.L.; Gnanadesikan, A. A synthesis of global particle export from the surface ocean and cycling through the ocean interior and on the seafloor. Glob. Biogeochem. Cycl. 2007, 21, GB4006. [Google Scholar] [CrossRef]
- Anderson, N.J.; Heathcote, A.J.; Engstrom, D.R.; contributors, G.d. Anthropogenic alteration of nutrient supply increases the global freshwater carbon sink. Sci Adv 2020, 6. [Google Scholar] [CrossRef]
- Mendonça, R.; Kosten, S.; Sobek, S.; Cardoso, S.J.; Figueiredo-Barros4, M.P.; Estrada, C.H.D.; Roland, F. Organic carbon burial efficiency in a large tropical hydroelectric reservoir. Biogeosci. Dis. 2015, 12, 18513–18540. [Google Scholar]
- Clow, D.W.; Stackpoole, S.M.; Verdin, K.L.; Butman, D.E.; Zhu, Z.; Krabbenhoft, D.P.; Striegl, R.G. Organic carbon burial in lakes and reservoirs of the conterminous United States. Environ. Sci. Technol 2015, https. [Google Scholar] [CrossRef]
- Kallistova, A.Y.; Kosyakova, A.I.; Rusanov, II; Kadnikov, V. V.; Beletsky, A.V.; Koval, D.D.; Yusupov, S.K.; Zekker, I.; Pimenov, N.V. Methane production in a temperate freshwater lake during an Intense cyanobacterial bloom. Microbiology 2023, 92, 638–649. [Google Scholar] [CrossRef]
- Qin, B.; Zhang, Y.; Zhu, G.; Gao, G. Eutrophication control of large shallow lakes in China. Sci. Total Environm. 2023, 881, 163494. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Yang, S.; Zhao, D.; Hu, C.; Xu, W.; Anderson, D.M.; Li, Y.; Song, X.P.; Boyce, D.G.; Gibson, L.; et al. Coastal phytoplankton blooms expand and intensify in the 21st century. Nature 2023, 615, 280–284. [Google Scholar] [CrossRef]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef]
- Phlips, E.J.; Badylak, S.; Milbrandt, E.C.; Stelling, B.; Arias, M.; Armstrong, C.; Behlmer, T.; Chappel, A.; Foss, A.; Kaplan, D.; et al. Fate of a toxic Microcystis aeruginosa bloom introduced into a subtropical estuary from a flow-managed canal and management implications. J. Environm. Manag. 2025, 375, 124362. [Google Scholar] [CrossRef]
- Geider, R.J.; MacIntyre, H.L.; Kana, T.M. Dynamic model of phytoplankton growth and acclimation: Responses of the balanced growth rate and the chlorophyll a:carbon ratio to light, nutrient-limitation and temperature. Mar. Ecol. Prog. Ser. 1997, 148, 187–200. [Google Scholar] [CrossRef]
- Reynolds, C.S. The Ecology of Phytoplankton.; Cambridge University Press. ISBN: 978-0521605199. 2006. [Google Scholar] [CrossRef]
- Wang, X.; Wu, M.; Ma, S.; Su, J.; He, K.; Wang, H.; Zhang, S. From cyanobacteria to kerogen: A model of organic carbon burial. Precam. Res. 2023, 390, 107035–107145. [Google Scholar] [CrossRef]
- Dynarski, K.A.; Bossio, D.A.; Scow, K.M. Dynamic Stability of Soil Carbon: Reassessing the “Permanence” of Soil Carbon Sequestration. Front. Environmen. Sci. /: https. [CrossRef]
- Nakagawa, T.; Gotanda, K.; Haraguchi, T.; Danhara, T.; Yonenobu, H.; Brauer, A.; Yokoyama, Y.; Tada, R.; Takemura, K.; Staff, R.A.; et al. SG06, a fully continuous and varved sediment core from Lake Suigetsu, Japan: Stratigraphy and potential for improving the radiocarbon calibration model and understanding of late Quaternary climate changes. Quat. Sci. Rev. 2012, 36, 164–176. [Google Scholar] [CrossRef]
- Hassan, S.; Bali, B.m.S.; Muneer, W.; Ali, S.N.; Morthekai, P.; Wani, A.H.; Sabreena; Ganai, B. A. Deciphering source, degradation status and temporal trends of organic matter in a himalayan freshwater lake using multiproxy indicators, optically stimulated luminescence dating and time series forecasting. Sci.Total Environm. 2024, 957, 177618. [Google Scholar] [CrossRef] [PubMed]
- Zastepa, A.; Taranu, Z.E.; Kimpe, L.E.; Blais, J.M.; Gregory-Eaves, I.; Zurawell, R.W.; Pick, F.R. Reconstructing a long-term record of microcystins from the analysis of lake sediments. Sci. Total Environm. 2017, 579, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, G.I.; Tani, Y.; Seto, K.; Tazawa, T.; Yamamuro, M.; Watanabe, T.; Nakamura, T.; Takemura, T.; Imura, S.; Kanda, H. Holocene paleolimnological changes in Lake Skallen Oike in the Syowa Station area of Antarctica inferred from organic components in a sediment core (Sk4C-02). J. Paleolimnol. 2010, 44, 677–693. [Google Scholar] [CrossRef]
- Engstrom, D.R.; Schottler, S.P.; Leavitt, P.R.; Havens, K.E. A reevaluation of the cultural eutrophication of Lake Okeechobee using multiproxy sediment records. Ecol. Appl. 2006, 16, 1194–1206. [Google Scholar] [CrossRef]
- Lee, S.; Block, B.; Jessup, B.; Salk, K.; >, P. ; >, P.;. Estimates of sediment accumulation rates and bottom core ages in Northeast Lakes. U.S. Environm. Prot. Agency, Washington, DC, /: https, 3612. [Google Scholar]
- Joarder, M.S.A.; Islam, M.S.; Hasan, M.H.; Sadman- Anjum, J.; Kabir, M.F.; Rashid, F.; Joarder, T.A. A comprehensive review of carbon dioxide capture, transportation, utilization, and storage: a source of future energy. Environ. Sci. Pollut. Res. 2025. [Google Scholar] [CrossRef]
- Zhou, T.R.; Cao, H.S.; Zheng, J.; Teng, F.; Wang, X.J.; Lou, K.; Zhang, X.H.; Tao, Y. Suppression of water-bloom cyanobacterium Microcystis aeruginosa by algaecide hydrogen peroxide maximized through programmed cell death. J. Hazard. Mat. 1016. [Google Scholar]
- Xu, C.C.; Yu, S.M.; Hu, J.; Effiong, K.; Ge, Z.W.; Tang, T.; Xiao, X. Programmed cell death process in freshwater Microcystis aeruginosa and marine Phaeocystis globosa induced by a plant derived allelochemical. Sci.Total Environm. 2022, 838. [Google Scholar] [CrossRef]
- Bai, F.; Li, J.; Li, T.; Sha, J.; Liu, J.; Li, L.; Dai, G.; Jia, Y.; Song, L. Unveiling the susceptibility mechanism of Microcystis to consecutive sub-lethal oxidative stress—Enhancing oxidation technology for cyanobacterial bloom control. Journal of Hazardous Materials 2024, 480, 135993. [Google Scholar] [CrossRef]
- Chen, Y.; Zaman, F.; Jia, Y.; Huang, Y.; Li, T.; Bai, F.; Li, L.; Song, L.; Li, J. Harmful cyanobacterial bloom control with hydrogen peroxide: Mechanism, affecting factors, development, and prospects Curr. Poll. Rep. 2024. [Google Scholar] [CrossRef]
- Kinley-Baird, C.; Calomeni, A.; Berthold, D.E.; Lefler, F.W.; Barbosa, M.; Rodgers, J.H.; Laughinghouse, H.D. Laboratory-scale evaluation of algaecide effectiveness for control of microcystin-producing cyanobacteria from Lake Okeechobee, Florida (USA). Ecotoxicol. Environm. Saf. 2021, 207, 111233. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, G.; Piel, T.; Xu, T.S.; White, E.; Qin, H.J.; Slot, P.C.; Huisman, J.; Visser, P.M. Sensitivity to hydrogen peroxide of the bloom-forming cyanobacterium Microcystis PCC 7806 depends on nutrient availability. Harm. Algae 2020, 99. [Google Scholar] [CrossRef]
- Long, B.M.; Jones, G.J.; Orr, P.T. Cellular microcystin content in N-limited Microcystis aeruginosa can be predicted from growth rate. Appl. Environm. Microbiol. 2001, 67, 278–283. [Google Scholar] [CrossRef]
- Badger, M.R.; Price, G.D. CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution. J. Exp. Bot. 2003, 54, 609–622. [Google Scholar] [CrossRef]
- Burnap, R.L.; Hagemann, M.; Kaplan, A. Regulation of the CO2 concentrating mechanism in cyanobacteria. Life 2015, 5, 348–371. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A.; Reinhold, L. CO2 concentrating mechanisms in photosynthetic microorganisms. Annu. Rev. Plant Physiol. Plant Mol.ar Biol. 1999, 50, 539–570. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A. Inorganic carbon concentrating mechanisms in relation to the biology of algae. Photosynth. Res. 2003, 77, 155–171. [Google Scholar] [CrossRef]
- Lucius, S.; Hagemann, M. The primary carbon metabolism in cyanobacteria and its regulation. Front. Plant Sci. 2024, 15, 1417680. [Google Scholar] [CrossRef]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae:: Mechanisms, environmental modulation, and evolution. Annu. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef]
- Sandrini, G.; Matthijs, H.C.P.; Verspagen, J.M.H.; Muyzer, G.; Huisman, J. Genetic diversity of inorganic carbon uptake systems causes variation in CO2 response of the cyanobacterium Microcystis. ISME J 2014, 8, 589–600. [Google Scholar] [CrossRef]
- Song, Y.F.; Qiu, B.S. The CO2-concentrating mechanism in the bloom-forming cyanobacterium Microcystis aeruginosa (Cyanophyceae) and effects of UVB radiation on its operation. J. Phycol. 2007, 43, 957–964. [Google Scholar] [CrossRef]
- Barchewitz, T.; Guljamow, A.; Meissner, S.; Timm, S.; Henneberg, M.; Baumann, O.; Hagemann, M.; Dittmann, E. Non-canonical localization of RubisCO under high-light conditions in the toxic cyanobacterium Microcystis aeruginosa PCC7806. Environm. Microbiol. 2019, 21, 4836–4851. [Google Scholar] [CrossRef]
- Guljamow, A.; Barchewitz, T.; Grosse, R.; Timm, S.; Hagemann, M.; Dittmann, E. Diel Variations of Extracellular Microcystin Influence the Subcellular Dynamics of RubisCO in Microcystis aeruginosa PCC 7806. Microorganisms 2021, 9, https. [Google Scholar] [CrossRef]
- Roy, S.; Guljamow, A.; Dittmann, E. Impact of temperature on the temporal dynamics of microcystin in Microcystis aeruginosa PCC7806. Front. Microbiol. 2023, 14, 1200816. [Google Scholar] [CrossRef]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Borner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. Plos One 2011, 6, e17615. [Google Scholar] [CrossRef] [PubMed]
- Schatz, D.; Keren, Y.; Vardi, A.; Sukenik, A.; Carmeli, S.; Boerner, T.; Dittmann, E.; Kaplan, A. Towards clarification of the biological role of microcystins, a family of cyanobacterial toxins. Environm. Microbiol. 2007, 9, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Teikari, J.E.; Russo, D.A.; Heuser, M.; Baumann, O.; Zedler, j.A.Z.; Liaimer, A.; Dittmann, E. Competition and interdependence define interactions of Nostoc sp. and Agrobacterium sp. under inorganic carbon limitation. NPJ Biofilms Microbiom. 2025, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A. Distribution and Functions of Calcium Mineral Deposits in Photosynthetic Organisms. In Progress in Botany Vol. 84, Lüttge, U., Cánovas, F.M., Risueño, M.-C., Leuschner, C., Pretzsch, H., Eds.; Springer Nature Switzerland: Cham, 2024; pp. 293–326. [Google Scholar]
- Taylor, A.R.; Brownlee, C.; Wheeler, G. Coccolithophore cell biology: Chalking up progress. Annu. Rev. Mar. Sci. 2017, 9, 283–310. [Google Scholar] [CrossRef]
- Konstantinou, C.; Wang, Y. ;. Unlocking the potential of microbially induced calcium carbonate precipitation (MICP) for hydrological applications: A beview of opportunities, challenges, and environmental considerations. Hydrology. [CrossRef]
- Gaëtan, J.; Halary, S.; Millet, M.; Bernard, C.; Duval, C.; Hamlaoui, S.; Hecquet, A.; Gugger, M.; Marie, B.; Mehta, N.; et al. Widespread formation of intracellular calcium carbonates by the bloom-forming cyanobacterium Microcystis. Environm. Microbiol. 2023, 25, 751–765. [Google Scholar] [CrossRef]
- Zhang, M.L.; Zhao, L.; Li, G.K.; Zhu, C.; Xu, D.; Ji, J.F.; Chen, J. Assessment and selection of cyanobacterial strains for CO2 mineral sequestration: implications for carbonation mechanism. Geomicrobiol. J. 2023, 40, 446–461. [Google Scholar] [CrossRef]
- Khan, H.; Marcé, R.; Laas, A.; Obrador, B. The relevance of pelagic calcification in the global carbon budget of lakes and reservoirs Limnetica. 41, /: https. [CrossRef]
- Riebesell, U.; Zondervan, I.; Rost, B.; Tortell, P.D.; Zeebe, R.E.; Morel, F.M.M. Reduced calcification of marine plankton in response to increased atmospheric CO2. Nature 2000, 407, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Bruley, A.; Gaëtan, J.; Gugger, M.; Pancrace, C.; Millet, M.; Gaschignard, G.; Dezi, M.; Humbert, J.-F.; Leloup, J.; Skouri-Panet, F.; et al. Diel changes in the expression of a marker 1 gene and candidate genes for intracellular 2 amorphous CaCO3 biomineralization in 3 Microcystis Peer Comm. J. 2024, 5, e18. [Google Scholar] [CrossRef]
- Mehta, N.; Gaëtan, J.; Giura, P.; Azaïs, T.; Benzerara, K. Detection of biogenic amorphous calcium carbonate (ACC) formed by bacteria using FTIR spectroscopy. Spect. Acta Part a-Mol. Biomol. Spect. 2022, 278. [Google Scholar] [CrossRef] [PubMed]
- Fruchter, N.; Lazar, B.; Nishri, A.; Almogi-Labin, A.; Eisenhauer, A.; Be'eri Shlevin, Y.; Stein, M. 88Sr/86Sr fractionation and calcite accumulation rate in the Sea of Galilee. Geochim. Cosmochim. Acta 2017, 215, 17–32. [Google Scholar] [CrossRef]
- Escoffier, N.; Perolo, P.; Many, G.; Pasche, N.T.; Perga, M.-E. Fine-scale dynamics of calcite precipitation in a large hardwater lake. Sci. Total Environm. 2023, 864, 160699. [Google Scholar] [CrossRef]
- Leiser, R.; Jongsma, R.; Bakenhus, I.; Möckel, R.; Philipp, B.; Neu, T.R.; Wendt-Potthoff, K. Interaction of cyanobacteria with calcium facilitates the sedimentation of microplastics in a eutrophic reservoir. Water Res. 2021, 189. [Google Scholar] [CrossRef]
- Many, G.; Escoffier, N.; Perolo, P.; Bärenbold, F.; Bouffard, D.; Perga, M.-E. Calcite precipitation: The forgotten piece of lakes’ carbon cycle. Sci. Adv. 2024, 10, eado5924. [Google Scholar] [CrossRef]
- Strong, A.E.; Eadie, B.J. Satellite observations of calcium carbonate precipitations in the Great Lakes. Limnol. Oceanogr. 1978, 23, 877–887. [Google Scholar] [CrossRef]
- Küchler-Krischun, J.; Kleiner, J. Heterogeneously nucleated calcite precipitation in Lake Constance. A short time resolution study. Aquatic Sci. 1990, 52, 176–197. [Google Scholar] [CrossRef]
- Jiang, H.B.; Cheng, H.M.; Gao, K.S.; Qiu, B.S. Inactivation of Ca2+/H+ exchanger in Synechocystis sp. strain PCC 6803 promotes cyanobacterial calcification by upregulating CO2-concentrating mechanisms. Appl. Environ. Microbiol. 2013, 79, 4048–4055. [Google Scholar] [CrossRef] [PubMed]
- Jansson, C.; Northen, T. Calcifying cyanobacteria—the potential of biomineralization for carbon capture and storage. Curr. Opin. Biotechnol. 2010, 21, 365–371. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, M.; Burnap, R.L. Action at a distance: The remarkable coupling of CO2 uptake to electron transfer in specialized cyanobacterial NDH-1 complexes PNAS in press 2025.
- Tchernov, D.; Hassidim, M.; Luz, B.; Sukenik, A.; Reinhold, L.; Kaplan, A. Sustained net CO2 evolution during photosynthesis by marine microorganisms. Curr. Biol. 1997, 7, 723–728. [Google Scholar] [CrossRef] [PubMed]
- McGowan, H.; Lensky, N.; Abir, S.; Shaked, Y.; Wurgaft, E. Direct measurement of CO2 air-sea exchange over a desert Fringing coral rReef, Gulf of Eilat (Aqaba), Israel. J. Geophys. Res.: Oceans 2022, 127, e2022JC018548. [Google Scholar] [CrossRef]
- Dean, W.E. The carbon cycle and biogeochemical dynamics in lake sediments. J. Paleolimnol. 1999, 21, 375–393. [Google Scholar] [CrossRef]
- Zepernick, B.N.; Gann, E.R.; Martin, R.M.; Pound, H.L.; Krausfeldt, L.E.; Chaffin, J.D.; Wilhelm, S.W. Elevated pH Conditions Associated With Microcystis spp. Blooms Decrease Viability of the Cultured Diatom Fragilaria crotonensis and Natural Diatoms in Lake Erie. Front. Microbiol. [CrossRef]
- Nishri, A.; Stiller, M.; Rimmer, A.; Geifman, Y.; Krom, M. Lake Kinneret (The Sea of Galilee): the effects of diversion of external salinity sources and the probable chemical composition of the internal salinity sources. Chem. Geol. 1999, 158, 37–52. [Google Scholar] [CrossRef]
- Liu, C.; Shen, Q.; Gu, X.; Zhang, L.; Han, C.; Wang, Z. Burial or mineralization: Origins and fates of organic matter in the water–suspended particulate matter–sediment of macrophyte- and algae-dominated areas in Lake Taihu. Water Res. 2023, 243, 120414. [Google Scholar] [CrossRef]
- O'Neil, J.R. Theoretical and experimental aspects of isotopic fractionation. In: Stable isotopes in high temperature geological processes (ed. J. W. Valley et al.) Rev. Mineral. 1986, 16, 1–40. [Google Scholar]
- Katz, A.; Nishri, A. Calcium, magnesium and strontium cycling in stratified, hardwater lakes: Lake Kinneret (Sea of Galilee), Israel. Geochim. Cosmochim. Acta 2013, 105, 372–394. [Google Scholar] [CrossRef]
- Xu, X.; Wu, C.; Xie, D.; Ma, J. Sources, migration, transformation, and environmental effects of organic carbon in eutrophic lakes: A critical review. Int. J. Environ. Res. Public Health 2023, 20, 860. [Google Scholar] [CrossRef]
- Zhou, C.; Penga, Y.; Zhou, M.; Jiaa, R.; Bud, H.; Xua, X.; Chen, L.; Mag, J.; Kinouchi, B.; Wang, G. Cyanobacteria decay alters CH4 and CO2 produced hotspots along vertical sediment profiles in eutrophic lakes Water Research 2024, 265, 122319. 265, 122319. [CrossRef]
- Hartmann, I.F.; Günthel, M.; Klintzsch, T.; Kirillin, G.; Grossart, H.-P.; Keppler, F.; Isenbeck-Schröter, M. High spatiotemporal dynamics of methane production and emission in oxic surface water. Environ. Sci. Technol. 1451. [Google Scholar]
- Rocher-Ros, G.; Stanley, E.; Loken, L.; Raymond, P.; Liu, S.; Sponseller, R. Global methane emissions from rivers and streams. Nature 2023, 530–535. [Google Scholar] [CrossRef]
- Rosentreter, J.A.; Borges, A.V.; Deemer, B.R.; Holgerson, M.A.; Liu, S.; Song, C.; Melack, J.; Raymond, P.A.; Duarte, C.M.; Allen, G.H.; et al. Half of global methane emissions come from highly variable aquatic ecosystem sources. Nat. Geosci. 2021, 14, 225–230. [Google Scholar] [CrossRef]
- Holgerson, M.A.; Raymond, P.A. Large contribution to inland water CO2 and CH4 emissions from very small ponds. Nat. Geosci. 2016, 9, 222–226. [Google Scholar] [CrossRef]
- Townsend-Small, A.; Disbennett, D.; Fernandez, J.M.; Ransohoff, R.W.; Mackay, R.; Bourbonniere, R.A. Quantifying emissions of methane derived from anaerobic organic matter respiration and natural gas extraction in Lake Erie. Limnol. Oceanogr. 2016, 61, S356–S366. [Google Scholar] [CrossRef]
- Kh-Hriiziirou, M.; Khoiyangbam, R.S. Strategies for CH4 Emission Reduction from Lakes: Understanding the Key Driving Factors for CH4 Production and Lake Restoration Intnl. j. Lakes Rivers. 2024, 17, 109–124. [Google Scholar]
- Lew, S.; Burandt, P.; Glińska-Lewczuk, K. Microbial Communities Drive Methane Fluxes From Floodplain Lakes—A Hydrological Gradient Perspective. Environm. Microbiol. 2025, 27, e70127. [Google Scholar] [CrossRef] [PubMed]
- Hounshell, A.G.; D’Acunha, B.M.; Breef-Pilz, A.; Johnson, M.S.; Thomas, R.Q.; Carey, C.C. Eddy covariance data reveal that a small freshwater reservoir emits a substantial amount of carbon dioxide and methane. J.Geophys. Res.: Biogeosci. 2022. [Google Scholar] [CrossRef]
- Waldo, S.; Beaulieu, J.J.; Barnett, S.W.; Balz, D.A.; Vanni4, M.J.; Williamson, T.; Walker, J.T. Temporal trends in methane emissions from a small eutrophic reservoir: the key role of a spring burst. Biogeosciences, 2021, 18, 5291–5311. [Google Scholar] [CrossRef] [PubMed]
- Jentzsch, K.; van Delden, L.; Fuchs, M.; Treat, C.C. An expert survey on chamber measurement techniques for methane fluxes. Earth Syst. Sci. Data Discuss. 2024, 2024, 1–59. [Google Scholar] [CrossRef]
- Whiticar, M. Carbon and hydrogen isotope systematics of bacterial formation and oxidation of methane. J. Chem. Geol. 1999, 161, 291–314. [Google Scholar] [CrossRef]
- Cerbin, S.; Pérez, G.; Rybak, M.; Wejnerowski, Ł.; Konowalczyk, A.; Helmsing, N.; Naus-Wiezer, S.; Meima-Franke, M.; Pytlak, Ł.; Raaijmakers, C.; et al. Methane-derived carbon as a driver for cyanobacterial growth. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Schroll, M.; Liu, L.; Einzmann, T.; Keppler, F.; Grossart, H.-P. Methane accumulation and its potential precursor compounds in the oxic surface water layer of two contrasting stratified lakes. Sci. Total Environm. 2023, 903, 166205. [Google Scholar] [CrossRef] [PubMed]
- Bižić, M.; Klintzsch, T.; Ionescu, D.; Hindiyeh, M.Y.; Günthel, M.; Muro-Pastor, M.A.; Eckert, W.; Urich, T.; Keppler, F.; Grossart, H.-P. Aquatic and terrestrial cyanobacteria produce methane. Science Advances, 5343. [Google Scholar] [CrossRef]
- Hu, S.X.; Zhou, B.; Wang, Y.; Wang, Y.; Zhang, X.X.; Zhao, Y.; Zhao, X.Y.; Tang, X.X. Effect of CO2-induced seawater acidification on growth, photosynthesis and inorganic carbon acquisition of the harmful bloom-forming marine microalga, Karenia mikimotoi. Plos One 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Klintzsch, T.; Geisinger, H.; Wieland, A.; Langer, G.; Nehrke, G.; Bizic, M.; Greule, M.; Lenhart, K.; Borsch, C.; Schroll, M.; et al. Stable carbon isotope signature of methane released from phytoplankton. Geophys. Res. Lett. 2023, 50, https. [Google Scholar] [CrossRef]
- Bizic, M. Phytoplankton photosynthesis: an unexplored source of biogenic methane emission from oxic environments. J. Plankton Res. 2021, 43, 822–830. [Google Scholar] [CrossRef]
- Cheng, X.X.; Lian, J.C.; Liu, B.; Zhu, X.W.; Jin, Y.; Zhang, L.J.; Tan, F.X.; Wu, D.J.; Liang, H. Integrated ferrate and calcium sulfite to treat algae-laden water for controlling ultrafiltration membrane fouling: High-efficiency oxidation and simultaneous cell integrity maintaining. Chem. Engin. J. 2023, 461. [Google Scholar] [CrossRef]
- Zepernick, B.N.; Hart, L.N.; Chase, E.E.; Natwora, K.E.; Obuya, J.A.; Olokotum, M.; Houghton, K.A.; Kiledal, E.A.; Achieng, D.; Barker, K.B.; et al. Molecular investigation of harmful cyanobacteria reveals hidden risks and niche partitioning in Kenyan Lakes. Harmful Algae 2024, 140, 102757. [Google Scholar] [CrossRef]
- Rao, Y.; Gao, G.; Berman-Frank, I.; Bizic, M.; Gao, K. Light-dependent methane production by a coccolithophorid may counteract its photosynthetic contribution to carbon dioxide sequestration. Commun. Earth & Environm. 2024, 5, 695. [Google Scholar]
- Khatun, S.; Iwata, T.; Kojima, H.; Fukui, M.; Aoki, T.; Mochizuki, S.; Naito, A.; Kobayashi, A.; Uzawa, R. Aerobic methane production by planktonic microbes in lakes. Sci.Total Environm. 2019, 696, https. [Google Scholar] [CrossRef]
- Fazi, S.; Amalfitano, S.; Venturi, S.; Pacini, N.; Vazquez, E.; Olaka, L.A.; Tassi, F.; Crognale, S.; Herzsprung, P.; Lechtenfeld, O.J.; et al. High concentrations of dissolved biogenic methane associated with cyanobacterial blooms in East African lake surface water. Commun. Biol. 2021, 4, 845. [Google Scholar] [CrossRef]
- Kamat, S.S.; Williams, H.J.; Dangott, L.J.; Chakrabarti, M.; Raushel, F.M. The catalytic mechanism for aerobic formation of methane by bacteria. Nature 2013, https. [Google Scholar] [CrossRef]
- Repeta, D.J.; Ferrón, S.; Sosa, O.A.; Johnson, C.G.; Repeta, L.D.; Acker, M.; DeLong, E.F.; Karl, D.M. Marine methane paradox explained by bacterial degradation of dissolved organic matter. Nat. Geosci. 2016, 9, 884–887. [Google Scholar] [CrossRef]
- Xu, H.L.; Li, H.; Tang, Z.Z.; Liu, Y.; Li, G.; He, Q. Underestimated methane production triggered by phytoplankton succession in river-reservoir systems: Evidence from a microcosm study. Water Res. 2020, 185. [Google Scholar] [CrossRef]
- Zhao, L.; Lin, L.-Z.; Chen, M.-Y.; Teng, W.-K.; Zheng, L.-L.; Peng, L.; Lv, J.; Brand, J.J.; Hu, C.-X.; Han, B.-P.; et al. The widespread capability of methylphosphonate utilization in filamentous cyanobacteria and its ecological significance. Water Res. 2022, 217, 118385. [Google Scholar] [CrossRef]
- Dummer, N.F.; Willock, D.J.; He, Q.; Howard, M.J.; Lewis, R.J.; Qi, G.; Taylor, S.H.; Xu, J.; Bethell, D.; Kiely, C.J.; et al. Methane oxidation to methanol. Chem. Rev. 2023, 123, 6359–6411. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Peng, Y.; Yua, M.; Deng, Y.; Chen, L.; Zhang, L.; Xua, X.; Zhang, S.; Yanb, Y.; Wang, G. Severe cyanobacteria accumulation potentially induces methylotrophic methane producing pathway in eutrophic lakes. Environm. Poll. 2022, 292, 118443. [Google Scholar] [CrossRef]
- Yang, F.; Zheng, X.; Wang, D.; Yao, E.; Li, Y.; Huang, W.; Zhang, L.; Wang, J.; Zhong, J. Significant diurnal variations in nitrous oxide (N2O) emissions from two contrasting habitats in a large eutrophic lake (Lake Taihu, China). Environm. Res. 2024, 261, 119691. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.M.; Townsend-Small, A.; Zastepa, A.; Watson, S.B.; Brandes, J.A. Methane and nitrous oxide measured throughout Lake Erie over all seasons indicate highest emissions from the eutrophic Western Basin. J. Great Lakes Res. 2020, 46, 1604–1614. [Google Scholar] [CrossRef]
- Hu, H.; Zhang, Z.; Chen, B.; Zhang, Q.; Xu, N.; Paerl, H.W.; Wang, T.; Hong, W.; Penuelas, J.; Qian, H. Potential health risk assessment of cyanobacteria across global lakes. Appl. Environ. Microbiol. 2024, e01936–24. [Google Scholar] [CrossRef]
- Weathers, P.J.; Niedzielski, J.J. Nitrous oxide production by cyanobacteria. Arch. Microbiol. 1986, 146, 204–206. [Google Scholar] [CrossRef]
- Fabisik, F.; Guieysse, B.; Procter, J.; Plouviez, M. Nitrous oxide (N2O) synthesis by the freshwater cyanobacterium Microcystis aeruginosa. Biogeosciences 2023, 20, 687–693. [Google Scholar] [CrossRef]
- Cai, S.; Lao, Q.; Chen, C.; Zhu, Q.; Chen, F. The impact of algal blooms on promoting in-situ N2O emissions: A case in Zhanjiang bay, China. J. Environm. Manag. 2024, 358, 120935. [Google Scholar] [CrossRef] [PubMed]
- Plouviez, M.; Shilton, A.; Packer, M.A.; Guieysse, B. Nitrous oxide emissions from microalgae: potential pathways and significance. J. Appl. Phycol. 2019, 31, 1–8. [Google Scholar] [CrossRef]
- Roothans, N.; Pabst, M.; van Diemen, M.; Herrera Mexicano, C.; Zandvoort, M.; Abeel, T.; van Loosdrecht, M.C.M.; Laureni, M. Long-term multi-meta-omics resolves the ecophysiological controls of seasonal N2O emissions during wastewater treatment. Nat. Water 2025, 3, 590–604. [Google Scholar] [CrossRef]
- Leon-Palmero, E.; Morales-Baquero, R.; Thamdrup, B.; Löscher, C.; Reche, I. Sunlight drives the abiotic formation of nitrous oxide in fresh and marine waters. Science 2025, 387, 1198–1203. [Google Scholar] [CrossRef]
Figure 1.
Vertical profiles of pH (A) and dissolved oxygen (DO, B) with water depth in three irrigation reservoirs. Nitzanim and Galon are filled with nutrient-rich, treated recycled water, while Masu’ot Itzhak receives floodwater collected from a nearby stream. Aerial images of the reservoirs (Figure 2A–C) show an extensive bloom in Nitzanim, predominantly Microcystis sp., in contrast to Galon (where routine mitigation treatments effectively limit bloom development) and Masu’ot Itzhak where algal growth is constrained by nutrients availability. The pH and DO measurements were performed using a YSI ProDSS Multi-Parameter Water Quality Meter on June 4th , 2025.
Figure 1.
Vertical profiles of pH (A) and dissolved oxygen (DO, B) with water depth in three irrigation reservoirs. Nitzanim and Galon are filled with nutrient-rich, treated recycled water, while Masu’ot Itzhak receives floodwater collected from a nearby stream. Aerial images of the reservoirs (Figure 2A–C) show an extensive bloom in Nitzanim, predominantly Microcystis sp., in contrast to Galon (where routine mitigation treatments effectively limit bloom development) and Masu’ot Itzhak where algal growth is constrained by nutrients availability. The pH and DO measurements were performed using a YSI ProDSS Multi-Parameter Water Quality Meter on June 4th , 2025.
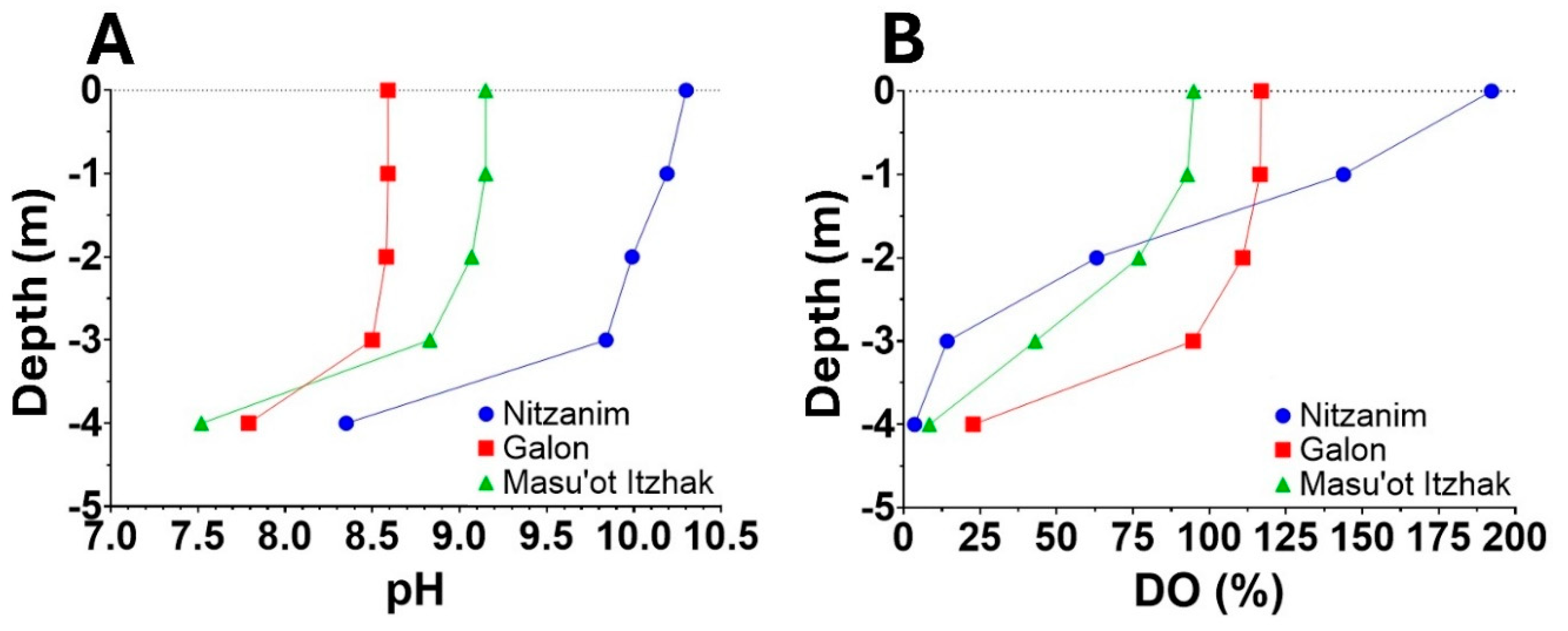
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



