
Submitted:
21 July 2025
Posted:
22 July 2025
You are already at the latest version
Abstract
Oligosaccharides represent an emerging class of bioactive compounds with diverse and structure-dependent functionalities. While traditionally explored for their prebiotic effects on gut microbiota, growing evidence demonstrates that oligosaccharides can directly interact with host receptors in the intestinal and immune systems, modulating inflammation, mucosal immunity, and epithelial barrier function. In the context of a circular bioeconomy, these molecules offer a sustainable and value-added route for the upcycling of agro-industrial and forestry biomass, particularly through the conversion of polysaccharides into structurally tailored oligosaccharides. This review synthesizes current knowledge on direct effects of plant-derived oligosaccharides on intestinal and immune system receptors. We also explore enzymatic and green chemistry-based strategies for oligosaccharide extraction and production, emphasizing their relevance to developing functional ingredients with targeted functionalities. Furthermore, we critically assess challenges and opportunities for translating this knowledge into practical applications, with a focus on regulatory aspects, outlining a roadmap toward the implementation of receptor-active oligosaccharides in precision nutrition and functional food innovation.
Keywords:
biomass valorisation
; dietary fibre
; intestinal receptors
; pattern recognition receptors (PRRs)
; functional ingredients
; functional foods
; host–glycan interactions
; food legislation
; regulatory frameworks
1. Introduction
1.1. Relevance of Oligosaccharides in Biomass Valorisation
The global food and forestry industries generate vast quantities of organic waste, much of which remains underutilized despite being rich in pectic and hemicellulosic polysaccharides. These are valuable materials that can serve directly as functional food ingredients or be converted into high-value oligosaccharides. These oligosaccharides are increasingly recognised for their potential as functional food ingredients with applications extending far beyond traditional uses. In particular, those derived from agricultural by-products, such as fruit peels [1], vegetable stems [2], cereal husks [3,4], and forestry residues [5], offer a sustainable route to reduce biomass waste and enhance natural resource use. Their recovery not only reduces environmental pressure associated with organic waste disposal but also aligns with principles of the circular bioeconomy.
Like polysaccharides, oligosaccharides can exert their functional effects through three main mechanisms: (i) physicochemical effects, such as binding or sequestration of molecules, which can slow gastric emptying and reduce the absorption of compounds like glucose or cholesterol [6]; (ii) modulation of gut microbiota profile and function through prebiotic activity [7]; and (iii) direct interaction with receptors expressed on intestinal and immune system cells [8,9]. This review focuses primarily on this later mechanism, which is gaining increasing attention due to its structural specificity and potential to influence host physiology beyond well-known physicochemical and prebiotic effects.
1.2. Oligosaccharides in Circular Bioeconomy: Expanding Their Functional Perspectives
The valorisation of agricultural and forestry by-products into oligosaccharides represents a promising strategy within the broader framework of the circular bioeconomy. As the food system continues to deal with resource inefficiencies, waste accumulation, and environmental degradation [10,11], the concept of upcycling biomass into value-added products is gaining momentum. While efforts to convert such biomass into biofuels have historically dominated valorisation approaches [12], the production of functional oligosaccharides offers an alternative that can be aligned with health and sustainable nutrition trends [13]. Green extraction methods, including pressurized hot water extraction, microwave-assisted hydrolysis, and enzymatic depolymerization, which are discussed further in this review, have made it increasingly feasible to obtain structurally diverse oligosaccharides in a sustainable (and scalable) manner. These advances support a shift in biomass utilization strategies [14], moving from energy recovery or bulk feed production to selective conversion into compounds with targeted physiological functions that promote human health.
Traditionally, the functional role of poly- and oligosaccharides has been narrowly interpreted through a prebiotic lens [15,16,17]. Defined as nondigestible carbohydrates that promote the growth of beneficial gut microbes, prebiotic oligosaccharides that can be obtained from biomass processing such as fructooligosaccharides (FOS) and xylooligosaccharides (XOS), have been widely explored for their ability to modulate the gut microbiota [18]. However, this microbiota-cantered paradigm, while valuable, overlooks the growing body of evidence indicating that oligosaccharides can exert direct effects by interacting with human intestinal and immune system receptors.
An increasing amount of evidence highlights that synthetic and naturally occurring oligosaccharide structures can engage with pattern recognition receptors (PRRs), G-protein coupled receptors (GPCRs), and other sensor systems located on the surface of intestinal epithelial and immune cells [8,9,19,20,21,22]. These receptor-mediated interactions have been shown to modulate inflammatory signalling pathways, intestinal permeability, mucosal immunity, and systemic immune responses [23,24]. Notably, these bioactivities are highly structure dependent. Minor changes in oligosaccharide length, degree of acetylation, branching, or linkage type, can influence its biological activity and receptor binding specificity [19,21,25]. This is in clear contrast to the more generalized community-level mode of action associated with physicochemical and prebiotic effects.
Recognising this broader functional potential implies a shift in how oligosaccharides are studied, produced, and not least, regulated. Rather than treating them solely as dietary fibre fractions with fermentability as the main criterion for functionality, there is a need to characterize oligomers based on molecular architecture and their interaction with host physiology. This expanded perspective opens door to precision design of oligosaccharides with targeted immunological or gut barrier-modulating properties.
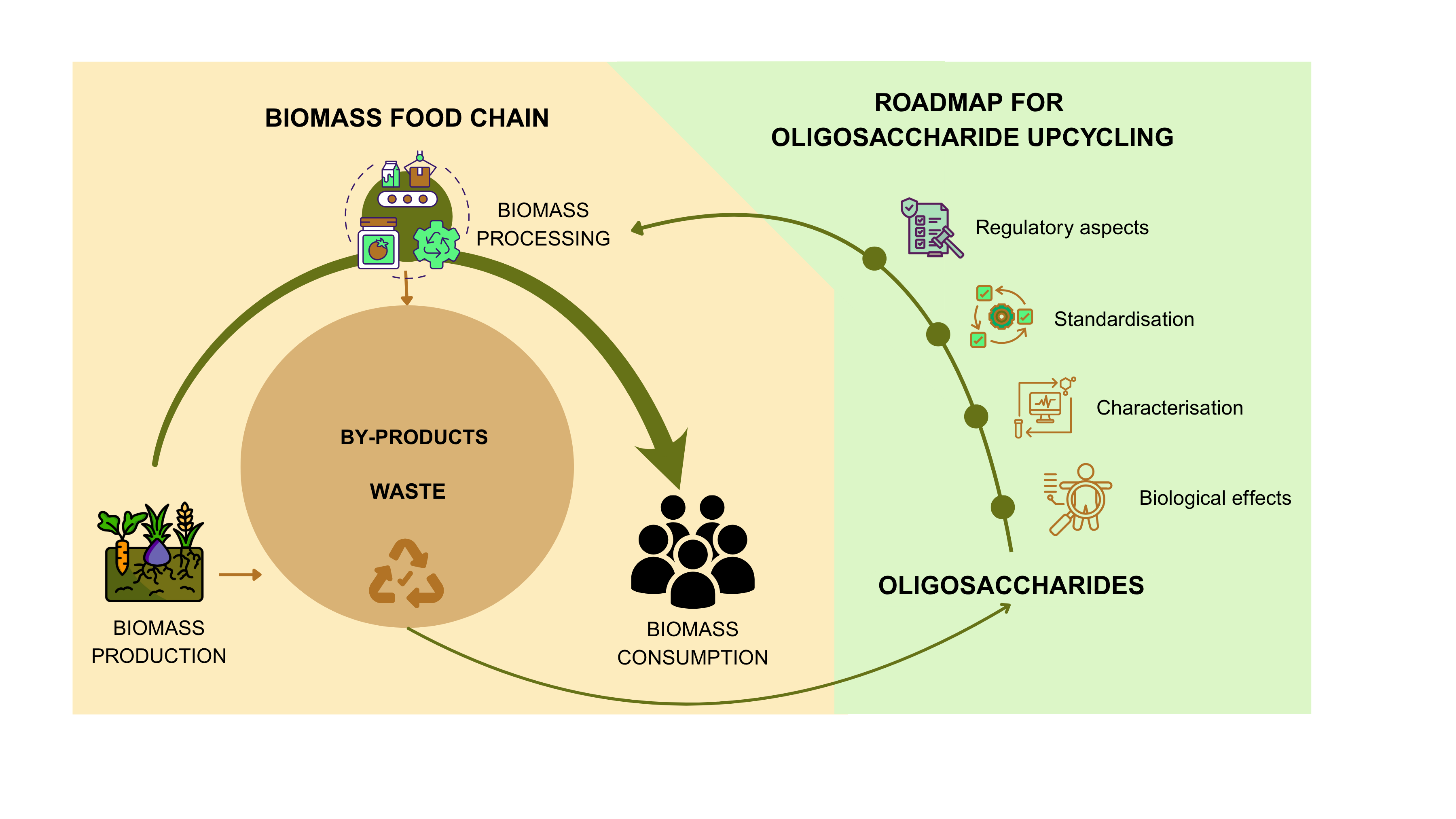
In the context of a circular economy, this approach is particularly powerful. It enables the transformation of varied biomass streams into structurally tailored oligosaccharides with defined health-promoting effects. This not only contributes to waste reduction and resource optimisation but also supports the development of next-generation functional ingredients that are both sustainable and biologically relevant. Following, we build upon this emerging framework, exploring the structural diversity of oligosaccharides, their receptor-mediated mechanisms of action, regulatory considerations, and how these insights can guide the rational design and valorisation of biomass-derived oligomers (Figure 1).
2. Oligosaccharides from Biomass: Sources and Upcycling Pathways
2.1. Structural Diversity of Oligosaccharides
Oligosaccharides derived from biomass exhibit remarkable structural diversity, shaped by the complexity of their parent polysaccharides and the wide range of botanical sources and processing conditions [3,26,27]. Their molecular architecture is defined by several variables, including monosaccharide composition, degree of polymerization (DP), glycosidic linkage type, branching frequency, and the presence of specific substitutions such as acetyl, methyl, or feruloyl groups. These features critically influence their biological functionality.
Forestry industry residues, particularly from softwood and hardwood species, are a rich source of hemicellulosic polysaccharides that can be hydrolysed into functional oligosaccharides [28,29]. In hardwoods, such as birch, glucuronoxylans predominate [30,31]. These consist of a β-(1→4)-linked xylose backbone decorated with α-(1→2)-linked glucuronic or 4-O-methyl-glucuronic acid side chains. The extent of methylation and acetylation differs by species, tissue and processing [31,32]. For example, birch-derived xylans often exhibit higher methylation and acetylation levels than those from eucalyptus [33], possibly resulting in oligosaccharides with distinct structural features and bioactivities. In softwoods like spruce and pine, the dominant hemicellulose is galactoglucomannan [34], composed of a β-(1→4)-linked mannose and glucose backbone with galactose side chains and varying degrees of acetylation. The ratio of mannose to glucose, the degree of galactose substitution, and the acetylation degree, are known to vary significantly between species and even between different parts of the tree, such as bark versus wood [35].
Agro-industrial residues from fruit and vegetable processing are rich in pectins and hemicelluloses that also serve as sources of diverse oligosaccharides [2,36,37]. Citrus peel [38], apple pomace [39,40], and sugar beet pulp [41] contain relatively high levels of homogalacturonan and rhamnogalacturonan-I (RG-I) [42], along with branched arabinans and arabinogalactans. These pectic domains differ considerably depending on the plant source and are often modified during processing. For instance, sugar beet pectin is particularly rich in RG-I with highly branched side chains [43], making it a promising source for arabino- and galactooligosaccharides after enzymatic hydrolysis [44]. Tomato peel and seed residues, commonly subjected to thermal processing during industrial paste production, have shown increased yields of pectic oligosaccharides with higher degrees of methylation and branching, likely due to partial depolymerization and migration of solubilized fragments [45,46].
In addition to hydrolysable polysaccharides, some oligosaccharides are naturally present in plant tissues and can be directly extracted. Fructans, such as inulin-type fructooligosaccharides (FOS), are prevalent in plants like chicory, Jerusalem artichoke, yacon, asparagus, onion, and garlic [47]. These consist of β-(2→1)-linked fructose chains often capped with a glucose moiety. As reviewed previously, their structural features, particularly chain length, degree of branching, and terminal residues, varies by species and maturity stage [48]. Postharvest storage and processing can also lead to inulin depolymerization, shifting the profile toward shorter-chain FOS, which tend to be more soluble and biologically active [49].
One of the most structurally diverse hemicelluloses is arabinoxylan, which is found in cereal brans and hulls such as those from wheat, rye, barley, and corn [50,51]. Arabinoxylans have a β-(1→4)-xylan backbone substituted with α-(1→2)- and/or α-(1→3)-linked arabinose units. The degree of arabinosylation varies by source; wheat bran arabinoxylans exhibit a high arabinose-to-xylose ratio of around 0.6 to 0.7, while those from corn bran are closer to 0.3, reflecting a more linear structure [52,53]. These differences influence not only solubility and fermentability, but also interaction with host receptors. In addition to arabinose substitution, many arabinoxylans are esterified with ferulic acid at the arabinose residues. The degree of feruloylation is also source-dependent [54]; maize and sorghum arabinoxylans tend to be more heavily feruloylated than those from wheat [55], influencing potential biological activity and resistance to enzymatic breakdown.
Other hemicelluloses or storage polymers, including β-glucans, mannans, and xyloglucans, also vary widely depending on the plant source. Oat and barley β-glucans differ in the ratio of β-(1→3) to β-(1→4) linkages [56,57]. Mannans from coffee pulp, ivory nut, and palm kernel can also differ in galactose substitution and molecular size [58,59,60]. Xyloglucans from tamarind seeds are highly branched, whereas those from kiwi are comparatively linear [61,62].
Processing conditions also influences the structural features of pectic and hemicellulosic materials [14]. Thermal treatment such as cooking, extrusion, or drying can cause depolymerization, de-esterification, or the release of branched fragments. In thermally processed carrot pomace and sweet corn husk, studies have reported higher levels of low-DP oligosaccharides with increased branching and methylation, potentially enhancing their functional performance in the gut [63,64]. Thus, the structural variation of oligosaccharides is not only a result of intrinsic plant composition but is also influenced by growing conditions, tissue type, developmental stage [65], and postharvest or industrial processing [66]. In the context of circular bioeconomy and upcycling strategies, understanding this complexity is critical. The traceability of biomass sources, knowledge of their biochemical composition, and awareness of changes induced by processing steps provide essential insight into the types of oligosaccharides that can be recovered or generated. This level of understanding supports more rational design of extraction processes and enhances the ability to produce structurally defined oligosaccharides with tailored functional properties from diverse (and sustainable) biomass streams.
2.2. Extraction Strategies for Obtaining Oligosaccharides
As highlighted above, agro-industrial residues and lignocellulosic materials from forestry operations represent abundant, diverse, and (unfortunately) underutilized sources of polysaccharides and oligosaccharides. For example, by-products from fruit and vegetable processing including peels, pomace, bagasse, and trimmings, are often rich in hemicelluloses and pectins, making them ideal stocks for oligosaccharide production [2,67]. Similarly, agroforestry side-streams such as sawdust, bark, and pruning waste contain valuable lignocellulosic fractions that can potentially be converted into functional oligosaccharides with bioactive potential [68]. The extraction of oligo- and polysaccharides from these biomasses typically follows sequential protocols that begin with mechanical disruption, followed by water or solvent-based solubilization steps, and often incorporate physical, chemical or enzymatic treatments to improve yield and selectivity.
Despite the structural diversity of oligosaccharides, and the well-documented fact that small changes in their fine structure can lead to marked differences in biological activity, many of the extraction workflows are remarkably general. Batch-scale production of oligosaccharides from a range of biomasses can often be achieved using similar unit operations and conditions, particularly during upstream hydrolysis and downstream purification stages. In the case of polysaccharides, the upstream extraction is usually based on hot water or alkaline treatments, occasionally coupled with chelating agents to recover specific fractions (e.g., alkali-soluble hemicelluloses or calcium-bound pectins). These polysaccharide-rich extracts can then serve as substrates for the generation of oligosaccharides through hydrolytic or enzymatic steps. While purification and structural fine-tuning are more complex and typically require tailored approaches, the overall process flow remains broadly applicable across biomass types. Table 1 summarizes common strategies used to process oligosaccharides and polysaccharides from biomass, including key solvents, enzymes, and specific features of each approach.
It is important to emphasize that, in the context of sustainability and circular economy, applying principles of green chemistry to these extraction and transformation processes is not merely beneficial, but essential. The drive to avoid harsh reagents, minimize energy input, and enable solvent recycling or biodegradability becomes more than a scientific priority; it is a prerequisite for economic and ecological viability in the context of circular economy.
2.3. Green Chemistry and Enzymatic Synthesis Approaches
The integration of green chemistry principles into the production of oligosaccharides from biomass is both a scientific imperative and a sustainability-driven necessity. As highlighted earlier, the goal is not merely to recover value from waste but to do so through processes that minimize environmental burden, reduce hazardous chemical use, and promote long-term system resilience. This section outlines the role of green chemistry in both the extraction of oligosaccharides from raw biomass and conversion of polysaccharides into tailored oligosaccharide structures, with emphasis on enzymatic strategies.
Green extraction technologies focus on replacing harsh or non-renewable reagents with safer, recyclable, and biodegradable alternatives [85]. Traditional acid hydrolysis, for example, though effective in breaking down lignocellulosic biomass, often requires neutralization steps that increase waste volume. In contrast, hydrothermal methods offer cleaner profiles while still enabling high solubilization of hemicelluloses or pectins [86,87]. Moreover, solvents such as ethanol and green alternatives like deep eutectic solvents (DES) or ionic liquids (ILs), when designed for recyclability, have shown promise in improving selectivity for certain glycan fractions while enhancing yield and reducing toxicity compared to more traditional approaches [78].
Ultrasound- and microwave-assisted extraction techniques are particularly attractive in green chemistry frameworks. These physical methods enhance mass transfer and accelerate cell wall disruption, shortening processing times [88]. When applied with minimal solvent systems (e.g., water or aqueous ethanol), they support energy-efficient and low-waste oligosaccharide biomass recovery. Notably, ultrasound can be combined with enzymatic pre-treatment to increase enzyme accessibility and facilitate oligosaccharide production in milder conditions [89].
While physical and chemical methods are still used at large scale, enzymatic hydrolysis remains the most structurally selective, environmentally friendly, and biologically compatible method for converting polysaccharides into oligosaccharides [90,91]. Enzymes such as xylanases, β-mannanases, pectinases, and cellulases act under mild aqueous conditions and allow for precise control over the degree of polymerization, substitution pattern, and glycosidic linkage of the resulting oligosaccharides [92,93]. For example, endo-xylanases from fungal sources (e.g., Trichoderma reesei, Aspergillus niger) can be source for a suite of XOS with defined backbone lengths and substitution degrees [94,95]. Similarly, endo-polygalacturonases and pectate lyases can be applied to pectin-rich biomass to generate pectic oligosaccharides (POS) [81,96,97]. Enzyme cascades or blends can be tailored to target side chains or internal bonds, creating structurally diverse mixtures with specific bioactivities [98]. Moreover, enzymatic approaches are not limited to degradation; transglycosylation reactions catalysed by glycoside hydrolases under high substrate concentrations (or by engineered glycosyltransferases) can be used to synthesize oligosaccharides with novel linkages and moieties, as reviewed previously [99]. This approach enables the construction of novel glycosidic backbones or the grafting of functional and side groups such as acetyl, arabinosyl, or galacturonyl residues that can enhance or modify biological activity. Although enzymatic synthesis and modification from waste substrates is still underexplored, recent research shows its potential for tailored design. For example, glucuronyltransferases or sulfatases can be used to introduce uronic acid and sulfate groups, respectively, into oligosaccharides derived from biomass streams [100,101], possibly mimicking the functionality of glycosaminoglycans or mucosal glycans. Importantly, these enzymatic routes are compatible with green chemistry principles: reactions that occur in aqueous media and minimize formation of toxic by-products while generating structurally defined products suitable for functional food applications.
Despite the promise of enzymatic and green strategies, several challenges remain. Enzyme costs, substrate variability, and process scalability can limit industrial adoption. Moreover, enzymatic reactions often produce a distribution of oligomers, requiring downstream separation techniques that are compatible with green processing (e.g., membrane filtration or precipitation). Advances in enzyme engineering, immobilization, and reaction optimisation (e.g., flow reactors, co-factor recycling) can help address these challenges [102]. Regardless of challenges, there is growing interest in coupling green chemistry with precision glycoengineering to produce oligomers with low environmental impact and high biological specificity, targeting intestinal and immune receptors. Synthetic biology platforms may ultimately enable the microbial production of customized oligosaccharides from waste-derived biomass, integrating feedstocks with precision biosynthesis.
3. Oligosaccharides as Bioactive Food Ingredients: Receptor Interactions and Structure-Dependent Host Modulation
3.1. Host Receptor Targets in the Intestinal and Immune System
Emerging evidence has demonstrated that poly- and oligosaccharides can interact directly with receptors on intestinal epithelial and immune cells [103,104], acting beyond their traditional role as prebiotics. These interactions occur at the mucosal interface and can modulate mechanisms such as inflammatory signalling, epithelial integrity, and immune priming. These findings have been validated through in vitro cell models [19,105,106], in vivo studies [105,107], and increasingly through in silico simulations to explore molecular docking and binding affinities [19,108]. Key receptors involved include pattern recognition receptors (PRRs) such as toll-like receptors (TLRs) but also C-type lectin receptors (CLRs), G-protein coupled receptors (GPCRs) and scavenger receptors. For instance, TLR4, a well-studied PRR involved in lipopolysaccharide (LPS) sensing and inflammatory response, has been shown to be downregulated in intestinal epithelial cells of mice with induced colitis when exposed to galacturonic acid-rich pectic oligosaccharides (POS), and to intact and modified (arabinose- and galactose-free) pectin fractions [109]. Pectic-like poly- and oligosaccharides can interfere with LPS-TLR4 interaction, thereby inhibiting the NF-κB pathway, as showed in vitro, explaining the reduction of pro-inflammatory cytokines like IL-6 and TNF-α by immune system cells [103,110,111]. Conversely, arabinoxylooligosaccharides from wheat bran appear to inhibit TLR2-mediated responses and induce TLR4 activation in a structure-dependent manner [112]. The intricate association of receptor-mediated effects with molecule structure is also seen for some pectic material, which have shown potential to inhibit TLR2 induced cytokine secretion [113,114].
CLRs such as DC-SIGN and dectin-1, expressed on dendritic cells and macrophages, also recognise specific glycan motifs on oligosaccharides [115,116]. Although this latter has not yet been thoroughly explored for oligosaccharides obtained from agro-industrial waste biomass, such receptor interactions are known to play a crucial role in maintaining gut barrier integrity, which can potentially be mediated by oligomers from beta-glucans, as proposed in silico [117].
Oligosaccharides may also exert anti-inflammatory effects by activating the nuclear receptor peroxisome proliferator-activated receptor gamma (PPARγ) [106]. Moreover, in vitro studies indicate that dietary fibres, such as inulin, GOS, FOS, and arabinoxylan hydrolysates can modulate cytokine and chemokine secretion by epithelial cells, macrophages, and dendritic cells, partly through PPARγ activation [8,106,118,119,120] . XOS have also been shown to enhance the expression of tight junction proteins claudin-1, occludin, and zonula occludens-1 (ZO-1), contributing to barrier integrity [121]. For intact and modified pectin though, mucin 2 (MUC2) levels were as low as in colitis mice control, showing no positive effect on this marker [122]. Additionally, some oligosaccharides can also interact with MUC-1, which can in turn supress TLR-4 signalling [122,123].
3.2. Structure–Activity Relationships: Fine Structural Features Dictate Bioactivity
Oligosaccharide bioactivity is highly dependent on their fine structural features. As mentioned earlier, variations in linkage type, degree of polymerization (DP), substitution patterns (e.g., acetylation, feruloylation, sulfation), and branching architecture [124] influence receptor binding and downstream effects. One example is the differential immunomodulatory activity observed among pectic oligosaccharides (POS) from distinct regions of the pectin polymer. POS originating from rhamnogalacturonan-I (RG-I) domains, which are rich in arabinose and galactose side chains, have been shown to engage pattern recognition receptors such as TLR2, leading to modulation of cytokine responses in intestinal epithelial and immune cells. In contrast, homogalacturonan (HG)-derived POS, composed primarily of linear galacturonic acid backbones, exhibit lower bioactivity in these contexts. These findings highlight that even subtle differences in structural features, such as size [125], side chain complexity, and the presence of ester-linked modifications [126], can potentially shape the effects of oligosaccharides in host–immune interactions [127].
In studies exploring feruloylated arabinoligosaccharides, typically obtained from rice bran, it was observed modulation of TLR4-mediated pathways in dendritic cells [9] and macrophages [128]. Likewise, feruloylated arabinooligosaccharides extracted from maize bran, a common cereal processing by-product, showed cell mediated responses in vitro [129]. The ratio of sugar residues within an oligosaccharide also matters, as highlighted for wheat arabinoxylooligosaccharides [53]. Recently, it was also found that slightly changes in the structure of arabinoxylooligosaccharides has a significant impact on their interaction with TLR2 and TLR4, as revealed by in vitro and in silico approaches [19]. This suggests that backbone flexibility and steric arrangement of side chains are essential for modulating interaction with receptor binding domains. An overview of the main target receptors and downstream effects in the gut lumen are highlighted in Figure 2.
These insights reinforce that structural tailoring, whether via enzymatic synthesis, selective hydrolysis, or chemical modification, is a powerful strategy to transform oligosaccharides from residues into functional oligosaccharides with specific receptor targets and defined biological effects.
3.3. Translational Applications and the Path Toward Precision Nutrition
Understanding the receptor-mediated mechanisms of oligosaccharides at the cellular level opens possibilities for their application in precision nutrition, clinical nutrition, and therapeutic innovation. This mechanistic specificity makes it possible to design nutritional interventions that are more targeted and adaptable to individual physiological needs. In inflammatory bowel disease (IBD), for instance, a central feature is the disruption of epithelial tight junctions, which compromises gut barrier integrity facilitating translocation of microbial components such as lipopolysaccharides (LPS) leading to chronic immune activation [130,131]. Oligosaccharides that interact with PRR like TLR4 to inhibit LPS binding or modulate downstream signalling can attenuate this inflammatory cascade. Pectic oligosaccharides and arabinooligosaccharides obtained from sugar beet pulp have demonstrated the capacity to interact with TLR4 in epithelial and immune cells, reducing secretion of pro-inflammatory cytokines such as TNF-α and IL-6 [124,125]. Similarly, arabinoxylooligosaccharides from varied cereal sources have been reported to support barrier function, which appears to be linked with enhanced expression of tight junction proteins such as claudin-1 and occludin [132,133].
Metabolic disorders such as obesity, insulin resistance, and type 2 diabetes are also increasingly linked to low-grade systemic inflammation that originates, in part, from altered gut permeability and immune dysfunction [134]. In this context, oligosaccharides that modulate anti-inflammatory pathways through direct receptor engagement, such as TLR2 or DC-SIGN, are of particular interest. Feruloylated arabinooligosaccharides, for example, can increase IL-10 production by activating TLR2, potentially rebalancing immune responses [9]. This receptor-specific bioactivity allows for the development of dietary solutions that target root inflammatory mechanisms rather than merely addressing symptoms or outcomes.
The potential reduction of pro-inflammatory effects lead by oligosaccharides could have a significant clinical relevance in irritable bowel syndrome (IBS). A recent scoping review [135] highlights the central role of TLR4 in the development of IBS, linking its altered expression to symptoms such as abdominal pain and diarrhoea. The authors suggest that TLR4 may be a promising therapeutic target, as multiple evidence indicates that interventions modulating this pathway can help alleviate IBS-related symptoms.
These applications are not limited to clinical settings but can also be extended to personalised nutrition strategies. Individuals with weakened mucosal defences, whether due to genetic predisposition, aging, or environmental exposures, could benefit from oligosaccharides designed to enhance epithelial barrier function or mucin production. Conversely, individuals exhibiting immune hyperresponsiveness or chronic inflammation may respond more favourably to oligosaccharides that promote signalling toward regulatory pathways. This level of dietary precision, informed by both individual physiology and glycan-receptor compatibility, could improve the efficacy of functional food and supplement interventions.
Importantly, these innovations are closely aligned with circular bioeconomy principles. Many of the most promising oligosaccharide candidates for receptor-based applications can be derived from agricultural and forestry by-products that are currently underutilized or discarded. For example, XOS can be extracted from corn cobs, wheat straw, and birch sawdust; galactoglucomannan oligosaccharides from spruce bark and pulp side-streams; arabinooligosaccharides from sugar beet pulp; pectic oligosaccharides from citrus peel; and mannooligosaccharides from spent coffee grounds and palm kernel cake. These residues are often viewed as waste or downcycled into animal feed or energy substrates, but they contain structurally rich polysaccharides that can be selectively hydrolysed and tailored into high-value bioactive food ingredients. Using green extraction methods and enzymatic processing, it is now feasible to isolate and refine oligosaccharides from these biomass sources in a manner that preserves their bioactive features. As discussed earlier, enzymatic treatments not only avoid the use of harsh chemicals but also allow for relatively good control over molecular structure. As such, this integration of sustainable processing with targeted oligomer design maximizes both the functional and ecological value of the biomass.
By advancing our understanding of oligosaccharide structure–function relationships and linking specific structural features to defined biological pathways, it becomes possible to develop functional ingredients that are both scientifically effective and ecologically responsible. This knowledge supports a new generation of foods and supplements tailored to individual needs and derived from renewable resources. In this context, oligosaccharides are not merely fibre fractions; they are programmable biological tools that combine human health and sustainability. The ability to match the right oligosaccharide to the right individual and physiological condition not only enhances health outcomes but also improves resource efficiency in the food system. Valorising side-streams from forestry and agriculture through the selective production of high-impact oligosaccharides contributes to a more sustainable, circular economy (Figure 1). This convergence of nutritional efficacy and biomass optimisation represents a strategic shift toward food systems that are not only health-driven but also waste-aware, climate-conscious.
5. Regulatory Pathways for Functional Oligosaccharides
Oligosaccharides derived from biomass hold promise as next-generation ingredients for nutraceuticals and functional foods, given their capacity to modulate intestinal barrier function, influence immune signalling, and act beyond traditional prebiotic effects. However, translating these findings into practical applications faces several regulatory and technological needs, especially when it comes to defining, standardizing, and approving these compounds for health-related use.
To date, most commercially available oligosaccharides including FOS and XOS, are categorized as dietary fibre and have gained generally recognised as safe (GRAS) status in the United States or been approved as novel food ingredients under EU regulations [136,137]. For example, the European Food Safety Authority (EFSA) permits the use of FOS as fibre ingredients in infant formula and a range of foods [136,138]. However, these approvals are generally based on the prebiotic effect and fermentability of oligosaccharides, rather than direct, receptor-mediated mechanisms in the human host.
This poses a major regulatory bottleneck: the existing definition of dietary fibre is largely functionally narrow, focused on resistance to digestion and partial or full fermentability in the colon. It often fails to account for non-fermentative effects, such as those mediated by interaction with receptors or tight junction proteins, which, as highlighted earlier, are now supported by robust in silico, in vitro and in vivo evidence. Consequently, oligosaccharides that do not conform to the standard model of fermentable fibre, despite having direct immunomodulatory or barrier-enhancing effects, may struggle to obtain regulatory approval or substantiated health claims under current frameworks.
Moreover, novel oligosaccharides derived from agro-industrial or forestry side-streams introduce additional complexity. While some of these molecules may be chemically similar to already approved FOS or XOS, their origin, structural heterogeneity, or extraction methods can classify them as "novel foods" under EU Regulation (EU) 2015/2283 [139], triggering lengthy safety evaluation procedures. In the US, the FDA’s GRAS designation also relies heavily on historical consumption and compositional familiarity [140], potentially excluding structurally unique oligosaccharides derived from unconventional biomass for human nutrition such as spruce sawdust, coffee pulp, or citrus processing residues.
This creates a paradox in which the most sustainable and structurally innovative oligosaccharides, those derived from (re)used biomass and circular bioeconomy models, are often the most difficult to commercialize. Furthermore, there is currently no harmonized international framework for evaluating the health effects of oligosaccharides based on direct receptor interactions, which limits the ability of manufacturers to formulate evidence-based claims for their products in global markets. On the technological side, several challenges must also be addressed to enable reliable application in food systems and health products. One of the key issues is standardisation of structure and composition. As highlighted in this review, the bioactivity of oligosaccharides is highly dependent on fine structural features such as linkage type, branching, and degree of polymerization. Yet, most commercial formulations, and especially potential by-products, contain heterogeneous mixtures, and small batch-to-batch differences can significantly impact efficacy and reproducibility.
Bioavailability is another critical and underexplored aspect. While fermentable oligosaccharides exert effects locally in the colon, receptor-active oligosaccharides may need to interact with epithelial or immune cells in the small intestine or Peyer’s patches. Their stability during digestion, susceptibility to microbial breakdown, and interaction with the mucus layer all influence whether they reach their biological targets. Strategies such as co-formulation with protective matrices, encapsulation, or surface modification may be required to ensure consistent activity in vivo, but these approaches add regulatory and manufacturing complexity.
In addition, the formulation of oligosaccharides into real food matrices presents technical challenges related to solubility, thermal stability, and pH sensitivity. While many oligosaccharides are water-soluble and stable under mild processing conditions, their structural integrity can be compromised by heat or enzymatic degradation in composite formulations [141]. Their interaction with other ingredients, especially proteins, lipids, and minerals, within the food matrix, might also affect bioactivity [142].
Despite these challenges, there are promising paths forward. The increasing acceptance of personalised nutrition as a regulatory and commercial framework may allow for more flexible claims and formulations, especially when linked to biomarkers or companion diagnostics. Progress will also depend on improved analytical tools and reference standards for oligosaccharide characterisation, as well as robust clinical studies demonstrating efficacy based on defined structural profiles and mechanistic endpoints. Collaboration between regulatory agencies, companies and researchers will be critical to bridge the gap between the scientific evidence that is being built and market translation.
In summary, while the application potential of biomass-derived oligosaccharides in health-promoting foods is significant and can clearly contribute to a more sustainable, efficient, and circular food system, the successful application requires (a) modernising the regulatory understanding of dietary fibre, (b) expanding health claim criteria beyond microbiota modulation, and (c) addressing technical barriers related to standardisation, bioavailability, and formulation. Advancing this agenda will not only increase the value of upcycled biomass but also enable the development of more precise, effective, and sustainable dietary strategies promoting human health.
6. Final Remarks
The valorisation of agro-industrial and forestry biomass into structurally defined oligosaccharides offers a compelling opportunity to simultaneously address health, sustainability, and innovation challenges. Moving beyond the traditional prebiotic lens, this review highlights how specific oligosaccharide structures can directly interact with intestinal and immune system receptors, exerting effects on mucosal immunity, inflammation, and gut barrier integrity. These mechanisms highlight novel applications in precision nutrition, where oligosaccharides can be tailored to specific physiological conditions or health needs. Moreover, advances in green chemistry and enzymatic synthesis have made it increasingly feasible to recover or generate bioactive oligosaccharides from side streams in a scalable and environmentally conscious manner. Nonetheless, fully realising their potential as functional foods or nutraceuticals requires updated regulatory frameworks that account for receptor-mediated actions beyond prebiotic potential, and address challenges in standardisation, bioavailability, and claim authentication. By integrating mechanistic insights, sustainable production strategies, and a novel regulatory framework, bioactive oligosaccharides from upcycled biomass can become important ingredients in circular food systems, contributing to both ecological resilience and human health.
Author Contributions
Conceptualization, V.C.; writing—original draft preparation, and writing—review and editing, V.C. and S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Swedish Research Council (VR), grant no. 2021-04937, the Lantmännen Research Foundation, grant no. LM-1013467, and the Swedish Research Council for Sustainable Development (FORMAS), grants no. 2020-02843 and 2024-01022.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| AXOS | Arabinoxylooligosaccharides |
| CLR | C-type lectin receptor |
| DC | Dendritic cell |
| DES | Deep eutectic solvent |
| DP | Degree of polymerization |
| EFSA | European Food Safety Authority |
| EMIM Ac | 1-ethyl-3-methylimidazolium acetate |
| FDA | Food and Drug Administration |
| FFAR2 | Free fatty acid receptor 2 (GPR43) |
| FOS | Fructooligosaccharides |
| GOS | Galactooligosaccharides |
| GPCR | G protein-coupled receptor |
| GRAS | Generally recognised as safe |
| HWE | Hot water extraction |
| IBD | Inflammatory bowel disease |
| IL-10 | Interleukin-10 |
| IL-6 | Interleukin-6 |
| LPS | Lipopolysaccharide |
| MAE | Microwave-assisted extraction |
| MOS | Mannooligosaccharides |
| MUC | Mucin |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| POS | Pectic oligosaccharides |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| PRR | Pattern recognition receptor |
| SCFA | Short-chain fatty acids |
| SIGN | Specific intercellular adhesion molecule-3-grabbing non-integrin |
| SWE | Subcritical water extraction |
| TFF | Tangential flow filtration |
| TLR | Degree of polymerization |
| UAE | Ultrasound-assisted extraction |
| ZO-1 | Zonulla occludens-1 |
References
- Orrego, D.; Olivares-Tenorio, M.L.; Hoyos, L. V.; Alvarez-Vasco, C.; Klotz-Ceberio, B.; Caicedo, N. Towards a Sustainable Circular Bioprocess: Pectic Oligosaccharides (POS) Enzymatic Production Using Passion Fruit Peels. Lwt 2024, 207, 116681. [Google Scholar] [CrossRef]
- Chockchaisawasdee, S.; Stathopoulos, C.E. Functional Oligosaccharides Derived from Fruit-and-Vegetable By-Products and Wastes. Horticulturae 2022, 8. [Google Scholar] [CrossRef]
- Arzami, A.N.; Ho, T.M.; Mikkonen, K.S. Valorization of Cereal By-Product Hemicelluloses: Fractionation and Purity Considerations. Food Res. Int. 2022, 151, 110818. [Google Scholar] [CrossRef] [PubMed]
- Madhukumar, M.S.; Muralikrishna, G. Fermentation of Xylo-Oligosaccharides Obtained from Wheat Bran and Bengal Gram Husk by Lactic Acid Bacteria and Bifidobacteria. J. Food Sci. Technol. 2012, 49, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Karnaouri, A.; Matsakas, L.; Krikigianni, E.; Rova, U.; Christakopoulos, P. Valorization of Waste Forest Biomass toward the Production of Cello-Oligosaccharides with Potential Prebiotic Activity by Utilizing Customized Enzyme Cocktails. Biotechnol. Biofuels 2019, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Chen, Y.; Chen, R.; Wen, Y.; Huang, Q.; Liu, Y.; Zhao, C. Research Status of the Effects of Natural Oligosaccharides on Glucose Metabolism. eFood 2022, 3, 1–12. [Google Scholar] [CrossRef]
- Lee, D.H.; Seong, H.; Chang, D.; Gupta, V.K.; Kim, J.; Cheon, S.; Kim, G.; Sung, J.; Han, N.S. Evaluating the Prebiotic Effect of Oligosaccharides on Gut Microbiome Wellness Using in Vitro Fecal Fermentation. npj Sci. Food 2023, 7. [Google Scholar] [CrossRef] [PubMed]
- Ortega-González, M.; Ocón, B.; Romero-Calvo, I.; Anzola, A.; Guadix, E.; Zarzuelo, A.; Suárez, M.D.; Sánchez de Medina, F.; Martínez-Augustin, O. Nondigestible Oligosaccharides Exert Nonprebiotic Effects on Intestinal Epithelial Cells Enhancing the Immune Response via Activation of TLR4-NFκB. Mol. Nutr. Food Res. 2014, 58, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Chen, H.H.; Chen, Y.K.; Chang, H.C.; Lin, P.Y.; Pan, I.H.; Chen, D.Y.; Chen, C.M.; Lin, S.Y. Rice Bran Feruloylated Oligosaccharides Activate Dendritic Cells via Toll-like Receptor 2 and 4 Signaling. Molecules 2014, 19, 5325–5347. [Google Scholar] [CrossRef] [PubMed]
- Alexander, P.; Brown, C.; Arneth, A.; Finnigan, J.; Moran, D.; Rounsevell, M.D.A. Losses, Inefficiencies and Waste in the Global Food System. Agric. Syst. 2017, 153, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Sarker, A.; Ahmmed, R.; Ahsan, S.M.; Rana, J.; Ghosh, M.K.; Nandi, R. A Comprehensive Review of Food Waste Valorization for the Sustainable Management of Global Food Waste. Sustain. Food Technol. 2023, 2, 48–69. [Google Scholar] [CrossRef]
- Mujtaba, M.; Fernandes Fraceto, L.; Fazeli, M.; Mukherjee, S.; Savassa, S.M.; Araujo de Medeiros, G.; do Espírito Santo Pereira, A.; Mancini, S.D.; Lipponen, J.; Vilaplana, F. Lignocellulosic Biomass from Agricultural Waste to the Circular Economy: A Review with Focus on Biofuels, Biocomposites and Bioplastics. J. Clean. Prod. 2023, 402, 136815. [Google Scholar] [CrossRef]
- Gibas-Dorna, M.; Żukiewicz-Sobczak, W. Sustainable Nutrition and Human Health as Part of Sustainable Development. Nutrients 2024, 16, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Chelliah, R.; Kim, N.H.; Park, S.J.; Park, Y.; Yeon, S.J.; Barathikannan, K.; Vijayalakshmi, S.; Oh, D.H. Revolutionizing Renewable Resources: Cutting-Edge Trends and Future Prospects in the Valorization of Oligosaccharides. Fermentation 2024, 10. [Google Scholar] [CrossRef]
- Liu, N.; Wang, H.; Yang, Z.; Zhao, K.; Li, S.; He, N. The Role of Functional Oligosaccharides as Prebiotics in Ulcerative Colitis. Food Funct. 2022, 13, 6875–6893. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Lu, J.; Li, B.; Lin, W.; Zhang, Z.; Wei, X.; Sun, C.; Chi, M.; Bi, W.; Yang, B.; et al. Effect of Functional Oligosaccharides and Ordinary Dietary Fiber on Intestinal Microbiota Diversity. Front. Microbiol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Gomez, M.X.; Ng, K.M.; Drexler, R.A.; Conner, A.M.S.; Vierra, C.G.; Krishnakumar, N.; Gerber, H.M.; Taylor, Z.R.; Treon, J.L.; Ellis, M.; et al. A Diverse Set of Solubilized Natural Fibers Drives Structure-Dependent Metabolism and Modulation of the Human Gut Microbiota. MBio 2025, 16, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jian, C.; Salonen, A.; Dong, M.; Yang, Z. Designing Healthier Bread through the Lens of the Gut Microbiota. Trends Food Sci. Technol. 2023, 134, 13–28. [Google Scholar] [CrossRef]
- Guerreiro, C. de A.; Andrade, L.A.D.; Fernández-Lainez, C.; Fraga, L.N.; López-Velázquez, G.; Marques, T.M.; Prado, S.B.R.; Brummer, R.J.; Nascimento, J.R.O.; Castro-Alves, V. Bioactive Arabinoxylan Oligomers via Colonic Fermentation and Enzymatic Catalysis: Evidence of Interaction with Toll-like Receptors from in Vitro, in Silico and Functional Analysis. Carbohydr. Polym. 2025, 352. [Google Scholar] [CrossRef]
- Sun, A.; Li, Z.; Zhao, W.; Zheng, J.; Zhang, Y.; Yao, M.; Yao, W.; Zhang, X.; Meng, X.; Li, Z.; et al. Synthesis and Biological Evaluation of Mirror Isomers of β-(1 → 3)-Glucans as Immune Modulators. Carbohydr. Polym. 2025, 357, 123477. [Google Scholar] [CrossRef] [PubMed]
- Castro-Alves, V.C.; Nascimento, J.R.O. do Size Matters: TLR4-Mediated Effects of α-(1,5)-Linear Arabino-Oligosaccharides in Macrophage-like Cells Depend on Their Degree of Polymerization. Food Res. Int. 2021, 141. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Bi, D.; Zheng, R.; Cai, N.; Xu, H.; Zhou, R.; Lu, J.; Wan, M.; Xu, X. Identification and Activation of TLR4-Mediated Signalling Pathways by Alginate-Derived Guluronate Oligosaccharide in RAW264.7 Macrophages. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rose, E.C.; Odle, J.; Blikslager, A.T.; Ziegler, A.L. Probiotics, Prebiotics and Epithelial Tight Junctions: A Promising Approach to Modulate Intestinal Barrier Function. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Semin, I.; Ninnemann, J.; Bondareva, M.; Gimaev, I.; Kruglov, A.A. Interplay Between Microbiota, Toll-Like Receptors and Cytokines for the Maintenance of Epithelial Barrier Integrity. Front. Med. 2021, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Zhu, S.; Yang, H.; Wang, X.; Li, J.; Liu, S.; Xing, R.; Li, P.; Li, K. Impact of O-Acetylation on Chitin Oligosaccharides Modulating Inflammatory Responses in LPS-Induced RAW264.7 Cells and Mice. Carbohydr. Res. 2024, 542, 109177. [Google Scholar] [CrossRef] [PubMed]
- Mathura, S.R.; Landázuri, A.C.; Mathura, F.; Andrade Sosa, A.G.; Orejuela-Escobar, L.M. Hemicelluloses from Bioresidues and Their Applications in the Food Industry – towards an Advanced Bioeconomy and a Sustainable Global Value Chain of Chemicals and Materials. Sustain. Food Technol. 2024, 2, 1183–1205. [Google Scholar] [CrossRef]
- Abik, F.; Palasingh, C.; Bhattarai, M.; Leivers, S.; Ström, A.; Westereng, B.; Mikkonen, K.S.; Nypelö, T. Potential of Wood Hemicelluloses and Their Derivates as Food Ingredients. J. Agric. Food Chem. 2023, 71, 2667–2683. [Google Scholar] [CrossRef] [PubMed]
- Najjoum, N.; Grimi, N.; Benali, M.; Chadni, M.; Castignolles, P. Extraction and Chemical Features of Wood Hemicelluloses: A Review. Int. J. Biol. Macromol. 2025, 311, 143681. [Google Scholar] [CrossRef] [PubMed]
- Gautam, D.; Rana, V.; Sharma, S.; Kumar Walia, Y.; Kumar, K.; Umar, A.; Ibrahim, A.A.; Baskoutas, S. Hemicelluloses: A Review on Extraction and Modification for Various Applications. ChemistrySelect 2025, 10, 1–24. [Google Scholar] [CrossRef]
- Lahtinen, M.H.; Kynkäänniemi, E.; Jian, C.; Salonen, A.; Pajari, A.M.; Mikkonen, K.S. Metabolic Fate of Lignin in Birch Glucuronoxylan Extracts as Dietary Fiber Studied in a Rat Model. Mol. Nutr. Food Res. 2023, 67, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Abad, A.; Giummarella, N.; Lawoko, M.; Vilaplana, F. Differences in Extractability under Subcritical Water Reveal Interconnected Hemicellulose and Lignin Recalcitrance in Birch Hardwoods. Green Chem. 2018, 20, 2534–2546. [Google Scholar] [CrossRef]
- Pinto, P.C.; Evtuguin, D. V.; Neto, C.P. Structure of Hardwood Glucuronoxylans: Modifications and Impact on Pulp Retention during Wood Kraft Pulping. Carbohydr. Polym. 2005, 60, 489–497. [Google Scholar] [CrossRef]
- Morais De Carvalho, D.; Marchand, C.; Berglund, J.; Lindström, M.E.; Vilaplana, F.; Sevastyanova, O. Impact of Birch Xylan Composition and Structure on Film Formation and Properties. Holzforschung 2020, 74, 184–196. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Butler, S.; Al-Rudainy, B.; Wallberg, O.; Stålbrand, H. Enzymatic Conversion of Different Qualities of Refined Softwood Hemicellulose Recovered from Spent Sulfite Liquor. Molecules 2022, 27, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, L.; Chao, Y.; Nawawi, D.S.; Akiyama, T.; Yokoyama, T.; Matsumoto, Y. Relationships between Hemicellulose Composition and Lignin Structure in Woods. J. Wood Chem. Technol. 2016, 36, 9–15. [Google Scholar] [CrossRef]
- Saez-Aguayo, S.; Sanhueza, D.; Fuenzalida, P.; Covarrubias, M.P.; Handford, M.; Herrera, R.; Moya-León, M.A. Back to the Wastes: The Potential of Agri-Food Residues for Extracting Valuable Plant Cell Wall Polysaccharides. Int. J. Mol. Sci. 2025, 26, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Vojvodić, A.; Komes, D.; Vovk, I.; Belščak-Cvitanović, A.; Bušić, A. Compositional Evaluation of Selected Agro-Industrial Wastes as Valuable Sources for the Recovery of Complex Carbohydrates. Food Res. Int. 2016, 89, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Yapo, B.M.; Lerouge, P.; Thibault, J.F.; Ralet, M.C. Pectins from Citrus Peel Cell Walls Contain Homogalacturonans Homogenous with Respect to Molar Mass, Rhamnogalacturonan I and Rhamnogalacturonan II. Carbohydr. Polym. 2007, 69, 426–435. [Google Scholar] [CrossRef]
- Kumari, N.; Kumar, M.; Radha; Rais, N. ; Puri, S.; Sharma, K.; Natta, S.; Dhumal, S.; Damale, R.D.; Kumar, S.; et al. Exploring Apple Pectic Polysaccharides: Extraction, Characterization, and Biological Activities - A Comprehensive Review. Int. J. Biol. Macromol. 2024, 255, 128011. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Fan, R.; Hu, J.; Zhang, X.; Han, L.; Wang, M.; He, C. Valorization of Apple Pomace: Structural and Rheological Characterization of Low-Methoxyl Pectins Extracted with Green Agents of Citric Acid/Sodium Citrate. Food Chem. X 2024, 24, 102010. [Google Scholar] [CrossRef] [PubMed]
- Glaser, S.J.; Abdelaziz, O.Y.; Demoitié, C.; Galbe, M.; Pyo, S.H.; Jensen, J.P.; Hatti-Kaul, R. Fractionation of Sugar Beet Pulp Polysaccharides into Component Sugars and Pre-Feasibility Analysis for Further Valorisation. Biomass Convers. Biorefinery 2024, 14, 3575–3588. [Google Scholar] [CrossRef]
- Kaczmarska, A.; Pieczywek, P.M.; Cybulska, J.; Zdunek, A. A Mini-Review on the Plant Sources and Methods for Extraction of Rhamnogalacturonan I. Food Chem. 2023, 403, 134378. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Lei, R.; Ryan, J.; Arrutia Rodriguez, F.; Rastall, B.; Chatzifragkou, A.; Winkworth-Smith, C.; Harding, S.E.; Ibbett, R.; Binner, E. Understanding the Influence of Processing Conditions on the Extraction of Rhamnogalacturonan-I “Hairy” Pectin from Sugar Beet Pulp. Food Chem. X 2019, 2, 100026. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.; Gullón, B.; Yáñez, R.; Alonso, J.L.; Parajó, J.C. Direct Enzymatic Production of Oligosaccharide Mixtures from Sugar Beet Pulp: Experimental Evaluation and Mathematical Modeling. J. Agric. Food Chem. 2009, 57, 5510–5517. [Google Scholar] [CrossRef] [PubMed]
- Jeddou, K. Ben; Bouaziz, F.; Helbert, C.B.; Nouri-Ellouz, O.; Maktouf, S.; Ellouz-Chaabouni, S.; Ellouz-Ghorbel, R. Structural, Functional, and Biological Properties of Potato Peel Oligosaccharides. Int. J. Biol. Macromol. 2018, 112, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Cámara Hurtado, M.; Greve, L.C.; Labavitch, J.M. Changes in Cell Wall Pectins Accompanying Tomato (Lycopersicon Esculentum Mill.) Paste Manufacture. J. Agric. Food Chem. 2002, 50, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Mudannayake, D.C.; Jayasena, D.D.; Wimalasiri, K.M.S.; Ranadheera, C.S.; Ajlouni, S. Inulin Fructans – Food Applications and Alternative Plant Sources: A Review. Int. J. Food Sci. Technol. 2022, 57, 5764–5780. [Google Scholar] [CrossRef]
- Zhang, Y. yue; Zhuang, D.; Wang, H. yang; Liu, C. yao; Lv, G. ping; Meng, L. juan Preparation, Characterization, and Bioactivity Evaluation of Oligosaccharides from Atractylodes Lancea (Thunb.) DC. Carbohydr. Polym. 2022, 277. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Oku, S.; Shimura, H.; Yoshihira, T.; Onodera, S. Variation of Fructan and Its Metabolizing Enzymes in Onions with Different Storage Characteristics. J. Appl. Glycosci. 2025, 72, n. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Bravo Núñez, Á.; Sahin, A.W.; Arendt, E.K. Arabinoxylans as Functional Food Ingredients: A Review. Foods (Basel, Switzerland) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Roubroeks, J.P.; Andersson, R.; Åman, P. Structural Features of (1 → 3),(1 → 4)-β-D-Glucan and Arabinoxylan Fractions Isolated from Rye Bran. Carbohydr. Polym. 2000, 42, 3–11. [Google Scholar] [CrossRef]
- Marcus Schmidt, B.W. and J.H. Comparison of Alkaline/Oxidative and Hydrothermal. Foods 2021. [Google Scholar]
- Kulathunga, J.; Islam, S. Wheat Arabinoxylans: Insight into Structure-Function Relationships. Carbohydr. Polym. 2025, 348, 122933. [Google Scholar] [CrossRef] [PubMed]
- Schendel, R.R.; Meyer, M.R.; Bunzel, M. Quantitative Profiling of Feruloylated Arabinoxylan Side-Chains from Graminaceous Cell Walls. Front. Plant Sci. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Malunga, L.N.; Beta, T. Isolation and Identification of Feruloylated Arabinoxylan Mono- and Oligosaccharides from Undigested and Digested Maize and Wheat. Heliyon 2016, 2, e00106. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A Concise Review on the Molecular Structure and Function Relationship of β-Glucan. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, M.S.; Jespersen, B.M.; Larsen, F.H.; Blennow, A.; Engelsen, S.B. Molecular Structure of Large-Scale Extracted β-Glucan from Barley and Oat: Identification of a Significantly Changed Block Structure in a High β-Glucan Barley Mutant. Food Chem. 2013, 136, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Reichembach, L.H.; Kaminski, G.K.; Maurer, J.B.B.; de Oliveira Petkowicz, C.L. Fractionation and Characterization of Cell Wall Polysaccharides from Coffee (Coffea Arabica L.) Pulp. Carbohydr. Polym. 2024, 327, 121693. [Google Scholar] [CrossRef] [PubMed]
- Kanai, N.; Yamada, K.; Sumida, C.; Tanzawa, M.; Ito, Y.; Saito, T.; Kimura, R.; Saito-Yamazaki, M.; Oyama, T.; Isogai, A.; et al. Mannan-Rich Holocellulose Nanofibers Mechanically Isolated from Spent Coffee Grounds: Structure and Properties. Carbohydr. Polym. Technol. Appl. 2024, 8, 100539. [Google Scholar] [CrossRef]
- Kalidas, N.R.; Saminathan, M.; Ismail, I.S.; Abas, F.; Maity, P.; Islam, S.S.; Manshoor, N.; Shaari, K. Structural Characterization and Evaluation of Prebiotic Activity of Oil Palm Kernel Cake Mannanoligosaccharides. Food Chem. 2017, 234, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cui, H.; Xie, F.; Song, Z.; Ai, L. Tamarind Seeds Polysaccharide: Structure, Properties, Health Benefits, Modification and Food Applications. Food Hydrocoll. 2024, 155, 110222. [Google Scholar] [CrossRef]
- Li, X.; Sakurai, N.; Nevins, D.J. Characterization of Kiwifruit Xyloglucan. J. Integr. Plant Biol. 2009, 51, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Yue, P.; Hu, Y.; Tian, R.; Bian, J.; Peng, F. Hydrothermal Pretreatment for the Production of Oligosaccharides: A Review. Bioresour. Technol. 2022, 343, 126075. [Google Scholar] [CrossRef] [PubMed]
- Trabert, A.; Schmid, V.; Keller, J.; Emin, M.A.; Bunzel, M. Chemical Composition and Technofunctional Properties of Carrot (Daucus Carota L.) Pomace and Potato (Solanum Tuberosum L.) Pulp as Affected by Thermomechanical Treatment. Eur. Food Res. Technol. 2022, 248, 2451–2470. [Google Scholar] [CrossRef]
- Prado, S.B.R. do; Ferreira, G.F.; Harazono, Y.; Shiga, T.M.; Raz, A.; Carpita, N.C.; Fabi, J.P. Ripening-Induced Chemical Modifications of Papaya Pectin Inhibit Cancer Cell Proliferation. Sci. Rep. 2017, 7, 16564. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, L. de F.; Kouzounis, D.; Schols, H.; de Vos, P.; Fabi, J.P. Assessing High-Temperature and Pressure Extraction of Bioactive Water-Soluble Polysaccharides from Passion Fruit Mesocarp. Carbohydr. Polym. 2024, 335, 122010. [Google Scholar] [CrossRef] [PubMed]
- Narisetty, V.; Parhi, P.; Mohan, B.; Hakkim Hazeena, S.; Naresh Kumar, A.; Gullón, B.; Srivastava, A.; Nair, L.M.; Paul Alphy, M.; Sindhu, R.; et al. Valorization of Renewable Resources to Functional Oligosaccharides: Recent Trends and Future Prospective. Bioresour. Technol. 2022, 346. [Google Scholar] [CrossRef] [PubMed]
- Ávila, P.F.; Goldbeck, R. Fractionating Process of Lignocellulosic Biomass for the Enzymatic Production of Short Chain Cello-Oligosaccharides. Ind. Crops Prod. 2022, 178. [Google Scholar] [CrossRef]
- Klinchongkon, K.; Khuwijitjaru, P.; Wiboonsirikul, J.; Adachi, S. Extraction of Oligosaccharides from Passion Fruit Peel by Subcritical Water Treatment. J. Food Process Eng. 2017, 40. [Google Scholar] [CrossRef]
- Liu, X.; Cao, R.; Xu, Y. Acidic Hydrolyzed Xylo-Oligosaccharides Bioactivity on the Antioxidant and Immune Activities of Macrophage. Food Res. Int. 2023, 163, 112152. [Google Scholar] [CrossRef] [PubMed]
- Fogarin, H.M.; Murillo-Franco, S.L.; Santos, M.C.M.; Silva, D.D.V.; Dussán, K.J. Acid Hydrolysis Pretreatment for Extraction of Oligosaccharides Derived from Spent Coffee Grounds: Valorization of a Promising Biomass. Environ. Sci. Pollut. Res. 2025. [Google Scholar] [CrossRef] [PubMed]
- Komarova, M.I.; Semenova, M. V.; Volkov, P. V.; Shashkov, I.A.; Rozhkova, A.M.; Zorov, I.N.; Kurzeev, S.A.; Satrutdinov, A.D.; Rubtsova, E.A.; Sinitsyn, A.P. Efficient Hydrolysis of Sugar Beet Pulp Using Novel Enzyme Complexes. Agronomy 2025, 15, 1–18. [Google Scholar] [CrossRef]
- Mathew, S.; Karlsson, E.N.; Adlercreutz, P. Extraction of Soluble Arabinoxylan from Enzymatically Pretreated Wheat Bran and Production of Short Xylo-Oligosaccharides and Arabinoxylo-Oligosaccharides from Arabinoxylan by Glycoside Hydrolase Family 10 and 11 Endoxylanases. J. Biotechnol. 2017, 260, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic-Malinovska, R.; Kuzmanova, S.; Winkelhausen, E. Application of Ultrasound for Enhanced Extraction of Prebiotic Oligosaccharides from Selected Fruits and Vegetables. Ultrason. Sonochem. 2015, 22, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Z.; Liu, Y. Ultrasonic-Assisted Extraction and Purification of Xylo-Oligosaccharides from Wheat Bran. J. Food Process Eng. 2022, 45. [Google Scholar] [CrossRef]
- del Amo-Mateos, E.; López-Linares, J.C.; García-Cubero, M.T.; Lucas, S.; Coca, M. Green Biorefinery for Sugar Beet Pulp Valorisation: Microwave Hydrothermal Processing for Pectooligosaccharides Recovery and Biobutanol Production. Ind. Crops Prod. 2022, 184. [Google Scholar] [CrossRef]
- Arrutia, F.; Adam, M.; Calvo-Carrascal, M.Á.; Mao, Y.; Binner, E. Development of a Continuous-Flow System for Microwave-Assisted Extraction of Pectin-Derived Oligosaccharides from Food Waste. Chem. Eng. J. 2020, 395, 125056. [Google Scholar] [CrossRef]
- Morais, E.S.; Da Costa Lopes, A.M.; Freire, M.G.; Freire, C.S.R.; Coutinho, J.A.P.; Silvestre, A.J.D. Use of Ionic Liquids and Deep Eutectic Solvents in Polysaccharides Dissolution and Extraction Processes towards Sustainable Biomass Valorization. Molecules 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Usmani, Z.; Sharma, M.; Gupta, P.; Karpichev, Y.; Gathergood, N.; Bhat, R.; Gupta, V.K. Ionic Liquid Based Pretreatment of Lignocellulosic Biomass for Enhanced Bioconversion. Bioresour. Technol. 2020, 304, 123003. [Google Scholar] [CrossRef] [PubMed]
- Ravn, H.C.; Meyer, A.S. Chelating Agents Improve Enzymatic Solubilization of Pectinaceous Co-Processing Streams. Process Biochem. 2014, 49, 250–257. [Google Scholar] [CrossRef]
- Babbar, N.; Baldassarre, S.; Maesen, M.; Prandi, B.; Dejonghe, W.; Sforza, S.; Elst, K. Enzymatic Production of Pectic Oligosaccharides from Onion Skins. Carbohydr. Polym. 2016, 146, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, P.; Amobonye, A.; Singh, S.; Pillai, S. Deep Eutectic Solvents in the Pretreatment of Feedstock for Efficient Fractionation of Polysaccharides: Current Status and Future Prospects. Biomass Convers. Biorefinery 2022, 12, 171–195. [Google Scholar] [CrossRef]
- Nabarlatz, D.; Torras, C.; Garcia-Valls, R.; Montané, D. Purification of Xylo-Oligosaccharides from Almond Shells by Ultrafiltration. Sep. Purif. Technol. 2007, 53, 235–243. [Google Scholar] [CrossRef]
- Wen, J.; Han, Q.; Qiu, M.; Jiang, L.; Chen, X.; Fan, Y. Membrane Technologies for the Separation and Purification of Functional Oligosaccharides: A Review. Sep. Purif. Technol. 2024, 346, 127463. [Google Scholar] [CrossRef]
- Imamoglu, E. Green Extraction Processes from Renewable Biomass to Sustainable Bioproducts. Bioresour. Technol. Reports 2024, 27, 101952. [Google Scholar] [CrossRef]
- Ramos-Andrés, M.; Aguilera-Torre, B.; García-Serna, J. Hydrothermal Production of High-Molecular Weight Hemicellulose-Pectin, Free Sugars and Residual Cellulose Pulp from Discarded Carrots. J. Clean. Prod. 2021, 290. [Google Scholar] [CrossRef]
- Sato, N.; Takano, Y.; Mizuno, M.; Nozaki, K.; Umemura, S.; Matsuzawa, T.; Amano, Y.; Makishima, S. Production of Feruloylated Arabino-Oligosaccharides (FA-AOs) from Beet Fiber by Hydrothermal Treatment. J. Supercrit. Fluids 2013, 79, 84–91. [Google Scholar] [CrossRef]
- Karbuz, P.; Tugrul, N. Microwave and Ultrasound Assisted Extraction of Pectin from Various Fruits Peel. J. Food Sci. Technol. 2021, 58, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.; Sánchez, J.; Aguilar-Galvez, A.; García-Ríos, D.; Chirinos, R.; Pedreschi, R. Ultrasound-Enhanced Enzymatic Hydrolysis for Efficient Production of Low Molecular Weight Oligogalacturonides from Polygalacturonic Acid. Food Biosci. 2025, 71. [Google Scholar] [CrossRef]
- Martínez Sabajanes, M.; Yáñez, R.; Alonso, J.L.; Parajó, J.C. Pectic Oligosaccharides Production from Orange Peel Waste by Enzymatic Hydrolysis. Int. J. Food Sci. Technol. 2012, 47, 747–754. [Google Scholar] [CrossRef]
- Babbar, N.; Dejonghe, W.; Gatti, M.; Sforza, S.; Elst, K. Pectic Oligosaccharides from Agricultural By-Products: Production, Characterization and Health Benefits. Crit. Rev. Biotechnol. 2016, 36, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.W.; Riaz, T.; Mahmood, S.; Liaqat, H.; Mushtaq, A.; Khan, S.; Amin, S.; Qi, X. Recent Advances in the Production, Analysis, and Application of Galacto-Oligosaccharides. Food Rev. Int. 2023, 39, 5814–5843. [Google Scholar] [CrossRef]
- Garc, A.; Morales, P.C.; Wojtusik, M.; Santos, V.E.; Kovensky, J.; Ladero, M. Production of Oligosaccharides from Agrofood Wastes. Fermentation 2020, 1–27. [Google Scholar]
- Nordberg Karlsson, E.; Schmitz, E.; Linares-Pastén, J.A.; Adlercreutz, P. Endo-Xylanases as Tools for Production of Substituted Xylooligosaccharides with Prebiotic Properties. Appl. Microbiol. Biotechnol. 2018, 102, 9081–9088. [Google Scholar] [CrossRef] [PubMed]
- Linares-Pasten, J.A.; Aronsson, A.; Karlsson, E.N. Structural Considerations on the Use of Endo-Xylanases for the Production of Prebiotic Xylooligosaccharides from Biomass. Curr. Protein Pept. Sci. 2016, 19, 48–67. [Google Scholar] [CrossRef] [PubMed]
- Combo, A.M.M.; Aguedo, M.; Goffin, D.; Wathelet, B.; Paquot, M. Enzymatic Production of Pectic Oligosaccharides from Polygalacturonic Acid with Commercial Pectinase Preparations. Food Bioprod. Process. 2012, 90, 588–596. [Google Scholar] [CrossRef]
- Tang, W.; Han, T.; Liu, W.; He, J.; Liu, J. Pectic Oligosaccharides: Enzymatic Preparation, Structure, Bioactivities and Application. Crit. Rev. Food Sci. Nutr. 2025, 65, 2117–2133. [Google Scholar] [CrossRef] [PubMed]
- Ávila, P.F.; de Mélo, A.H.F.; Goldbeck, R. Cello-Oligosaccharides Production from Multi-Stage Enzymatic Hydrolysis by Lignocellulosic Biomass and Evaluation of Prebiotic Potential. Innov. Food Sci. Emerg. Technol. 2023, 85. [Google Scholar] [CrossRef]
- Vera, C.; Illanes, A.; Guerrero, C. Enzymatic Production of Prebiotic Oligosaccharides. Curr. Opin. Food Sci. 2021, 37, 160–170. [Google Scholar] [CrossRef]
- Lee, C.; Teng, Q.; Zhong, R.; Ye, Z.H. Arabidopsis GUX Proteins Are Glucuronyltransferases Responsible for the Addition of Glucuronic Acid Side Chains onto Xylan. Plant Cell Physiol. 2012, 53, 1204–1216. [Google Scholar] [CrossRef] [PubMed]
- Endo, M.; Kotake, T.; Watanabe, Y.; Kimura, K.; Tsumuraya, Y. Biosynthesis of the Carbohydrate Moieties of Arabinogalactan Proteins by Membrane-Bound β-Glucuronosyltransferases from Radish Primary Roots. Planta 2013, 238, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.C.C.; Dutra, L.; Fé, L.X.S.G.M.; Freire, D.M.G.; Manoel, E.A.; Cipolatti, E.P. Chapter 15 - Immobilized Biocatalysts for Hydrolysis of Polysaccharides. In Foundations and Frontiers in Enzymology; Goldbeck, R., Poletto, P.B.T.-P.-D.B., Eds.; Academic Press, 2023; pp. 385–407 ISBN 978-0-323-99986-1.
- Prado, S.B.R.; Beukema, M.; Jermendi, E.; Schols, H.A.; de Vos, P.; Fabi, J.P. Pectin Interaction with Immune Receptors Is Modulated by Ripening Process in Papayas. Sci. Rep. 2020, 10, 1690. [Google Scholar] [CrossRef] [PubMed]
- Vos, A.P.; M’Rabet, L.; Stahl, B.; Boehm, G.; Garssen, J. Immune-Modulatory Effects and Potential Working Mechanisms of Orally Applied Nondigestible Carbohydrates. Crit. Rev. Immunol. 2007, 27, 97–140. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.Y.; Määttänen, P.; Napper, S.; Scruten, E.; Li, B.; Koike, Y.; Johnson-Henry, K.C.; Pierro, A.; Rossi, L.; Botts, S.R.; et al. Non-Digestible Oligosaccharides Directly Regulate Host Kinome to Modulate Host Inflammatory Responses without Alterations in the Gut Microbiota. Microbiome 2017, 5, 135. [Google Scholar] [CrossRef] [PubMed]
- Zenhom, M.; Hyder, A.; de Vrese, M.; Heller, K.J.; Roeder, T.; Schrezenmeir, J. Prebiotic Oligosaccharides Reduce Proinflammatory Cytokines in Intestinal Caco-2 Cells via Activation of PPARγ and Peptidoglycan Recognition Protein 3. J. Nutr. 2011, 141, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Henry, K.C.; Pinnell, L.J.; Waskow, A.M.; Irrazabal, T.; Martin, A.; Hausner, M.; Sherman, P.M. Short-Chain Fructo-Oligosaccharide and Inulin Modulate Inflammatory Responses and Microbial Communities in Caco2-Bbe Cells and in a Mouse Model of Intestinal Injury. J. Nutr. 2014, 144, 1725–1733. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, L.; Xue, S.; Wang, S.; Zhu, L.; Ma, T.; Liu, H.; Li, R. Molecular Docking and Dynamic Insights on the Adsorption Effects of Soy Hull Polysaccharides on Bile Acids. Int. J. Food Sci. Technol. 2022, 57, 3702–3712. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.H.; Niu, Y.B.; Sun, Y.; Guo, Z.J.; Li, Q.; Li, C.; Feng, J.; Cao, S.S.; Mei, Q.B. An Apple Oligogalactan Prevents against Inflammation and Carcinogenesis by Targeting LPS/TLR4/NF-ΚB Pathway in a Mouse Model of Colitis-Associated Colon Cancer. Carcinogenesis 2010, 31, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Chen, W.; Liu, Q.; Yang, G.; Li, K. Pectin Oligosaccharides Ameliorate Colon Cancer by Regulating Oxidative Stress- and Inflammation-Activated Signaling Pathways. Front. Immunol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pan, X.; Ran, S.; Wang, K. Purification, Structural Elucidation and Anti-Inflammatory Activity in Vitro of Polysaccharides from Smilax China L. Int. J. Biol. Macromol. 2019, 139, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, C. de A.; Andrade, L.A.D.; Fernández-Lainez, C.; Fraga, L.N.; López-Velázquez, G.; Marques, T.M.; Prado, S.B.R.; Brummer, R.J.; Nascimento, J.R.O.; Castro-Alves, V. Bioactive Arabinoxylan Oligomers via Colonic Fermentation and Enzymatic Catalysis: Evidence of Interaction with Toll-like Receptors from in Vitro, in Silico and Functional Analysis. Carbohydr. Polym. 2025, 352. [Google Scholar] [CrossRef]
- Tang, X.; de Vos, P. Structure-Function Effects of Different Pectin Chemistries and Its Impact on the Gastrointestinal Immune Barrier System. Crit. Rev. Food Sci. Nutr. 2025, 65, 1201–1215. [Google Scholar] [CrossRef] [PubMed]
- 114. Sahasrabudhe, N.M.; Beukema, M.; Tian, L.; Troost, B.; Scholte, J.; Bruininx, E.; Bruggeman, G.; van den Berg, M.; Scheurink, A.; Schols, H.A.; et al. Dietary Fiber Pectin Directly Blocks Toll-Like Receptor 2–1 and Prevents Doxorubicin-Induced Ileitis. Front. Immunol. 2018, 9, 383. [Google Scholar]
- Kikete, S.; Luo, L.; Jia, B.; Wang, L.; Ondieki, G.; Bian, Y. Plant-Derived Polysaccharides Activate Dendritic Cell-Based Anti-Cancer Immunity. Cytotechnology 2018, 70, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Fadden, A.J.; Drickamer, K. A Novel Mechanism of Carbohydrate Recognition by the C-Type Lectins DC-SIGN and DC-SIGNR. Subunit Organization and Binding to Multivalent Ligands. J. Biol. Chem. 2001, 276, 28939–28945. [Google Scholar] [CrossRef] [PubMed]
- Huo, R.; Wuhanqimuge; Zhang, M. ; Sun, M.; Miao, Y. Molecular Dynamics Modeling of Different Conformations of Beta-Glucan, Molecular Docking with Dectin-1, and the Effects on Macrophages. Int. J. Biol. Macromol. 2025, 293, 139382. [Google Scholar] [CrossRef] [PubMed]
- Mendis, M.; Leclerc, E.; Simsek, S. Arabinoxylan Hydrolyzates as Immunomodulators in Lipopolysaccharide-Induced RAW264.7 Macrophages. Food Funct. 2016, 7, 3039–3045. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Sahasrabudhe, N.M.; Rösch, C.; Schols, H.A.; Faas, M.M.; de Vos, P. The Impact of Dietary Fibers on Dendritic Cell Responses in Vitro Is Dependent on the Differential Effects of the Fibers on Intestinal Epithelial Cells. Mol. Nutr. Food Res. 2015, 59, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Mendis, M.; Leclerc, E.; Simsek, S. Arabinoxylan Hydrolyzates as Immunomodulators in Caco-2 and HT-29 Colon Cancer Cell Lines. Food Funct. 2017, 8, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Mavrogeni, M.E.; Asadpoor, M.; Henricks, P.A.J.; Keshavarzian, A.; Folkerts, G.; Braber, S. Direct Action of Non-Digestible Oligosaccharides against a Leaky Gut. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Sabater, C.; Molina-Tijeras, J.A.; Vezza, T.; Corzo, N.; Montilla, A.; Utrilla, P. Intestinal Anti-Inflammatory Effects of Artichoke Pectin and Modified Pectin Fractions in the Dextran Sulfate Sodium Model of Mice Colitis. Artificial Neural Network Modelling of Inflammatory Markers. Food Funct. 2019, 10, 7793–7805. [Google Scholar] [CrossRef] [PubMed]
- Sabater, C.; Blanco-Doval, A.; Margolles, A.; Corzo, N.; Montilla, A. Artichoke Pectic Oligosaccharide Characterisation and Virtual Screening of Prebiotic Properties Using in Silico Colonic Fermentation. Carbohydr. Polym. 2021, 255, 117367. [Google Scholar] [CrossRef] [PubMed]
- Meijerink, M.; Rösch, C.; Taverne, N.; Venema, K.; Gruppen, H.; Schols, H.A.; Wells, J.M. Structure Dependent-Immunomodulation by Sugar Beet Arabinans via a SYK Tyrosine Kinase-Dependent Signaling Pathway. Front. Immunol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Castro-Alves, V.C.; Nascimento, J.R.O. do Size Matters: TLR4-Mediated Effects of α-(1,5)-Linear Arabino-Oligosaccharides in Macrophage-like Cells Depend on Their Degree of Polymerization. Food Res. Int. 2021, 141. [Google Scholar] [CrossRef] [PubMed]
- Beukema, M.; Jermendi; van den Berg, M. A.; Faas, M.M.; Schols, H.A.; de Vos, P. The Impact of the Level and Distribution of Methyl-Esters of Pectins on TLR2-1 Dependent Anti-Inflammatory Responses. Carbohydr. Polym. 2021, 251, 117093. [Google Scholar] [CrossRef] [PubMed]
- Fuso, A.; Rosso, F.; Rosso, G.; Risso, D.; Manera, I.; Caligiani, A. Production of Xylo-Oligosaccharides (XOS) of Tailored Degree of Polymerization from Acetylated Xylans through Modelling of Enzymatic Hydrolysis. Food Res. Int. 2022, 162, 112019. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.Y.; Chen, Y.K.; Chen, H.H.; Lin, S.Y.; Fang, Y.T. Immunomodulatory Effects of Feruloylated Oligosaccharides from Rice Bran. Food Chem. 2012, 134, 836–840. [Google Scholar] [CrossRef] [PubMed]
- Akshatha, S.; Gnanesh Kumar, B.S.; Mazumder, K.; Eligar, S.M. Structural Characterization and Bioactivities of Maize Bran Feruloylated Arabinoxylan and Oligosaccharides Obtained by a Combined Hydrothermal and Enzymatic Pre-Treatment. Food Biosci. 2024, 60, 104417. [Google Scholar] [CrossRef]
- Landy, J.; Ronde, E.; English, N.; Clark, S.K.; Hart, A.L.; Knight, S.C.; Ciclitira, P.J.; Al-Hassi, H.O. Tight Junctions in Inflammatory Bowel Diseases and Inflammatory Bowel Disease Associated Colorectal Cancer. World J. Gastroenterol. 2016, 22, 3117–3126. [Google Scholar] [CrossRef] [PubMed]
- Candelli, M.; Franza, L.; Pignataro, G.; Ojetti, V.; Covino, M.; Piccioni, A.; Gasbarrini, A.; Franceschi, F. Interaction between Lipopolysaccharide and Gut Microbiota in Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Van Hée, V.F.; Piront, N.; De Backer, F.; Toussaint, O.; Cani, P.D.; Delzenne, N.M. Wheat-Derived Arabinoxylan Oligosaccharides with Prebiotic Effect Increase Satietogenic Gut Peptides and Reduce Metabolic Endotoxemia in Diet-Induced Obese Mice. Nutr. Diabetes 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, T.; Shikano, M.; Tanaka, M.; Ozeki, K.; Ebi, M.; Katano, T.; Hamano, S.; Nishiwaki, H.; Tsukamoto, H.; Mizoshita, T.; et al. Therapeutic Effects of Biobran, Modified Arabinoxylan Rice Bran, in Improving Symptoms of Diarrhea Predominant or Mixed Type Irritable Bowel Syndrome: A Pilot, Randomized Controlled Study. Evidence-based Complement. Altern. Med. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Rohm, T. V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in Obesity, Diabetes, and Related Disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Wang, L.; Wang, Z.; Wan, C. Toll-like Receptor 4 Plays a Vital Role in Irritable Bowel Syndrome: A Scoping Review. Front. Immunol. 2024, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of the Extension of Use of Galacto-Oligosaccharides (GOS) as a Novel Food in Food for Special Medical Purposes Pursuant to Regulation (EU) 2015/2283. EFSA J. 2022, 20. [Google Scholar] [CrossRef]
- Turck, D.; Bresson, J.L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of Xylo-Oligosaccharides (XOS) as a Novel Food Pursuant to Regulation (EU) 2015/2283. EFSA J. 2018, 16. [Google Scholar] [CrossRef]
- Bührer, C.; Ensenauer, R.; Jochum, F.; Kalhoff, H.; Koletzko, B.; Lawrenz, B.; Mihatsch, W.; Posovszky, C.; Rudloff, S. Infant Formulas with Synthetic Oligosaccharides and Respective Marketing Practices. Mol. Cell. Pediatr. 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Turck, D.; Bohn, T.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pentieva, K.; et al. Guidance on the Scientific Requirements for an Application for Authorisation of a Novel Food in the Context of Regulation (EU) 2015/2283. EFSA J. 2024, 22, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Pomeranz, J.L.; Broad Leib, E.M.; Mozaffarian, D. Regulation of Added Substances in the Food Supply by the Food and Drug Administration Human Foods Program. Am. J. Public Health 2024, 114, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Forgo, P.; Kiss, A.; Korózs, M.; Rapi, S. Thermal Degradation and Consequent Fragmentation of Widely Applied Oligosaccharides. Microchem. J. 2013, 107, 37–46. [Google Scholar] [CrossRef]
- Aguilera, J.M. The Food Matrix: Implications in Processing, Nutrition and Health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic overview of the biomass food chain and roadmap for oligosaccharide upcycling. The left side of the diagram illustrates the biomass food chain, beginning with biomass production (e.g., vegetables and cereals), followed by biomass processing into food products. During this chain, by-products and waste are generated. These underutilized streams are highlighted as valuable sources for oligosaccharide upcycling, as shown in the right side of the figure. The roadmap to valorise these oligosaccharides includes four main steps: assessment of their biological effects, characterisation, standardisation, and alignment with regulatory aspects.
Figure 1.
Schematic overview of the biomass food chain and roadmap for oligosaccharide upcycling. The left side of the diagram illustrates the biomass food chain, beginning with biomass production (e.g., vegetables and cereals), followed by biomass processing into food products. During this chain, by-products and waste are generated. These underutilized streams are highlighted as valuable sources for oligosaccharide upcycling, as shown in the right side of the figure. The roadmap to valorise these oligosaccharides includes four main steps: assessment of their biological effects, characterisation, standardisation, and alignment with regulatory aspects.
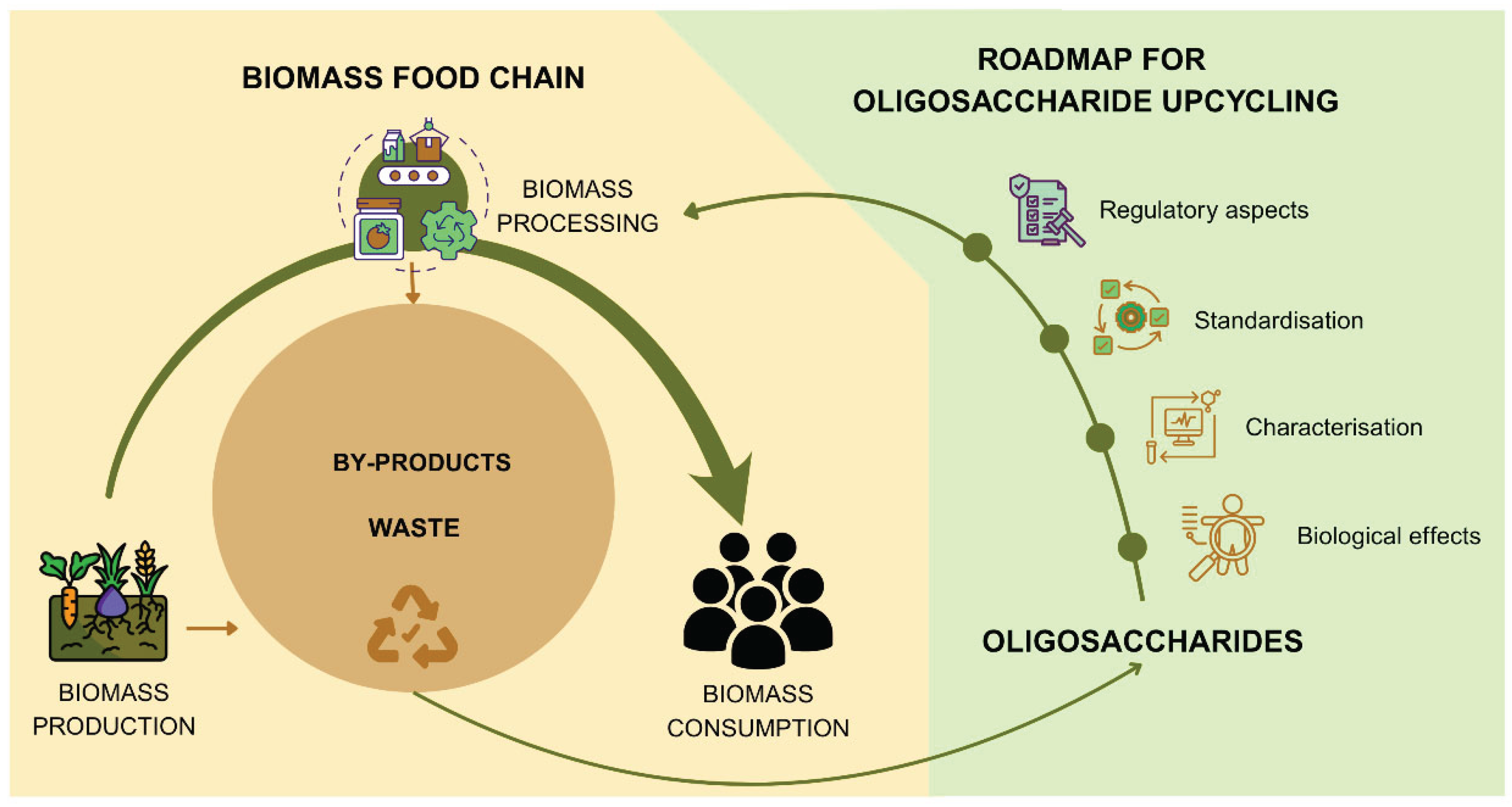
Figure 2.
Direct effects of oligosaccharides from by-products and waste on gut immune signalling and barrier integrity. Different oligosaccharide structures interact with pattern recognition receptors (PRRs) such as TLR2, TLR4, and Dectin-1 at the gut epithelium, leading to either suppression or activation of inflammatory pathways (e.g., NF-κB) and modulation of anti-inflammatory responses (e.g., IL-10). These interactions also impact the expression of genes related to mucus production (MUC2) and tight junction proteins (Claudin-1, Occludin, ZO-1), contributing to epithelial barrier integrity. While general immunomodulatory effects are observed, specific effects are highly dependent on the structural features of the oligosaccharides.
Figure 2.
Direct effects of oligosaccharides from by-products and waste on gut immune signalling and barrier integrity. Different oligosaccharide structures interact with pattern recognition receptors (PRRs) such as TLR2, TLR4, and Dectin-1 at the gut epithelium, leading to either suppression or activation of inflammatory pathways (e.g., NF-κB) and modulation of anti-inflammatory responses (e.g., IL-10). These interactions also impact the expression of genes related to mucus production (MUC2) and tight junction proteins (Claudin-1, Occludin, ZO-1), contributing to epithelial barrier integrity. While general immunomodulatory effects are observed, specific effects are highly dependent on the structural features of the oligosaccharides.
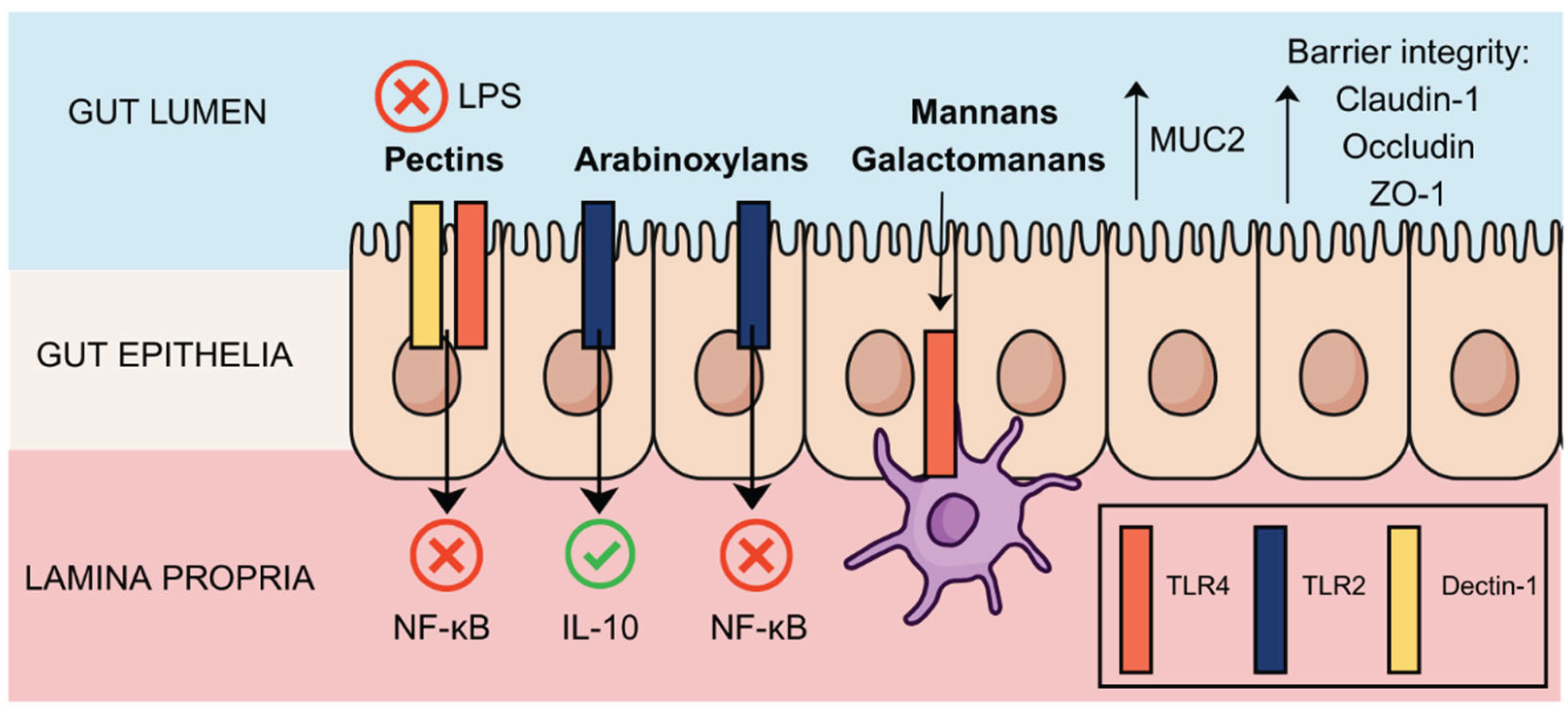

Table 1.
Methods for extraction and recovery of oligosaccharides and polysaccharides from biomass. Overview of common methods used for the recovery and processing of oligosaccharides and polysaccharides. The table highlights target materials, key solvents or enzymes, and notable processing characteristics.
Table 1.
Methods for extraction and recovery of oligosaccharides and polysaccharides from biomass. Overview of common methods used for the recovery and processing of oligosaccharides and polysaccharides. The table highlights target materials, key solvents or enzymes, and notable processing characteristics.
| Extraction Method. | Main processing stage |
Target material |
Main solvents/enzymes used |
Special properties/ Comments |
Examples/Sources |
| Hot water extraction (HWE) | Extraction | Polysaccharides, some oligosaccharides | Water (90–160°C) | Eco-friendly, good for heat-stable material | [52,63] |
| Subcritical water extraction (SWE) | Extraction | Hemicellulosic material (recalcitrant) | Water (>100°C) and pressure (> 10 bar) | Alternative to HWE for recalcitrant material | [31,69] |
| Alkaline extraction | Extraction | Hemicelluloses, lignin removal | NaOH, Ca(OH)₂, others |
Solubilizes hemicellulose, effective lignin disruption | [52,68] |
| Acid hydrolysis | Extraction | Pectic/hemicellulosic oligosaccharides | H₂SO₄, H₃PO₄, others |
Effective but harsh; degradation risk | [70,71] |
| Enzymatic hydrolysis | Hydrolysis or conversion | Oligosaccharides (FOS, XOS, GOS, etc.) | Xylanase, Cellulase, Pectinase, others | Mild, specific, avoids toxic residues | [72,73] |
| Ultrasound-assisted extraction (UAE) | Extraction enhancement | Polysaccharides and oligosaccharides | Water, mechanical agitation | Enhances diffusion, reduces energy/time | [74,75] |
| Microwave-assisted extraction (MAE) | Extraction enhancement | Polysaccharides and oligosaccharides | Water/solvent, microwave radiation | Fast, efficient; high energy input | [76,77] |
| Ionic liquid pretreatment | Pretreatment | Cellulose, lignin, hemicelluloses | EMIM Ac, others | Efficient delignification; recovery needed | [78,79] |
| Chelating agent | Pretreatment | Pectin/rich biomass, crosslinked polysaccharides | EDTA, citric acid, others | Removes metal ions, disrupts Ca2⁺ bridges in pectins | [80,81] |
| Deep eutectic solvent (DES) extraction | Pretreatment | Lignin, hemicelluloses | Choline chloride + lactic acid, urea, others | Biodegradable, tunable solvent properties | [78,82] |
| (Tangential flow) filtration (TFF) | Isolation or fractionation | Fractionation of solubilized saccharides | Membrane-based size separation | Industrial-scale separation of high/low MW fractions | [83,84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



