
Submitted:
17 July 2025
Posted:
18 July 2025
You are already at the latest version
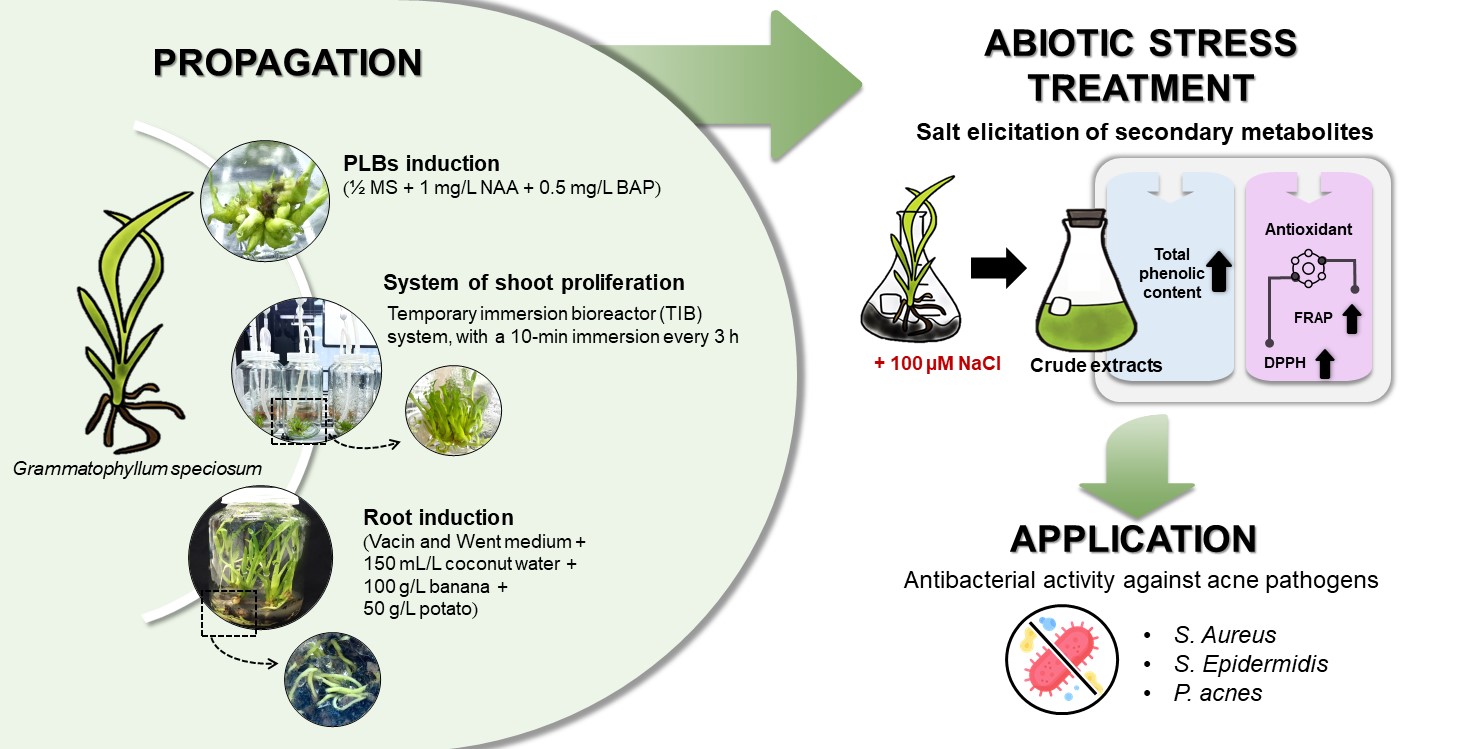
Abstract
Grammatophyllum speciosum is an endangered wild orchid with medicinal properties. This research propagated G. speciosum from vegetative organs grown under different NaCl-stressed conditions. Half-strength Murashige and Skoog (½ MS) medium supplemented with a combination of 1 mg L–1 1-naphthaleneacetic acid (NAA) and 0.5 mg L–1 6-benzylaminopurine (BAP) proved to be a suitable medium for shoot formation (32.33 ± 2.52 shoots per explant). The protocorm-like bodies were transferred into a temporary immersion bioreactor (TIB) system. A 10-min immersion time every 3 h enhanced the maximum number of shoots, the shoot height, and the fresh growth index (127.00 ± 2.16, 5.00 ± 0.51 cm and 4.26 ± 0.52, respectively). Rooting of proliferated plantlets from the TIB system was successfully achieved in Vacin and Went medium. The plantlets were maintained in ½ MS medium supplemented with Sodium chloride (NaCl) (0, 50, 100 or 200 µM) under white light-emitting diode light to induce the total phenolic content (TPC) from in vitro culture for 72 h. The TPC was highest in the medium with 100 µM NaCl (111.06 ± 2.24 mg gallic acid equivalents g-1 dry weight). The diphenyl picrylhydrazyl antioxidant activity was 24.50 ± 0.76% and ferric reducing antioxidant power values were in the range 2,441.79 ± 1.21 to 2,491.96 ± 3.23 µM ascorbic acid equivalent g-1 dry weight. The G. speciosum extracts showed antibacterial activity against acne pathogens, with minimum inhibitory concentration and minimum bactericidal concentration values in the ranges 6.4–12.8 mg mL–1 and 12.8–25.6 mg mL–1, respectively.
Keywords:
antioxidant activity
; biological activity
; bioactive compounds
; medicinal plant
; micropropagation
; temporary immersion bioreactors
1. Introduction
Grammatophyllum speciosum is an endangered wild orchid with various medicinal properties. G. speciosum distributes across the southern islands of the Pacific Ocean and the tropical regions of Southeast Asia, including Thailand [1]. It is commonly known as the Giant orchid or Tiger orchid. In Thailand, it is known as ‘waan phet cha hueng’. G. speciosum is used as a Thai traditional medicine for the treatment of fever, insect bites, skin rashes, and as an anti-inflammatory elixir [2]. Currently, G. speciosum pseudobulb ethanolic extract exhibits various promising pharmacological activities to treat breast cancer [3]. In addition, this extract has been utilized in the cosmetics industry for its antioxidant, anti-aging, and anti-collagenase activities [4], as well as for its skin-whitening efficacy [5]. These therapeutic effects result from the biological compounds produced through the plant’s metabolism.
The cultivation of plant cells, tissue, and organs serves as a crucial tool for investigating the processes associated with plant growth and development [6,7]. Such cultivation produces disease-free plants that are used for breeding, propagation, and the conservation of endangered plant species. In vitro plant propagation presents a viable alternative for bioactive compound production [8]. A temporary immersion bioreactor (TIB) is an innovative system designed for plant tissue culture using a liquid medium. This system excels in simultaneously and efficiently producing large quantities of plants by providing a suitable medium and controlling the timing of nutrient delivery. An essential function of the TIB system is ensuring that plant tissues are not continuously submerged in the liquid medium, which is especially valuable in preventing them from becoming overly succulent [9]. The first TIB system for banana propagation was reported by Alvard [10]. Subsequently, the TIB system has been used globally for the large-scale micropropagation of crops [11,12,13,14], medicinal plants [15,16], and other economic plants [11,17]. The shoot multiplication rate was significantly enhanced by two-fold to more than ten-fold in a TIB, compared to semi-solid culture methods [18,19,20]. In addition, micropropagation using the TIB system has been reported for several orchid species, such as Cattleya tigrine [21], Dendrobium sp. [22,23], Phalaenopsis sp. [24], Vanilla planifolia [25], and Vanda tricolor [9].
Therefore, this could be an interesting approach to cultivating G. speciosum because this plant reproduces by seed propagation or the separation of pups (suckers). Nevertheless, the conventional seeding method is difficult, as many seeds tend to disperse naturally, resulting in a low survival rate due to insufficient nutrients for germination [26]. In addition, the suckers process is protracted, requiring a substantial period to establish robust mother plants [27]. Importantly, the propagation of G. speciosum using aseptic techniques is highly advantageous because it enables rapid production of high-quality plants within a short timeframe. This not only ensures a sufficient supply for immediate use but also facilitates the conservation and preservation of the mother plant for future applications and sustainability. Consequently, the current research evaluated the efficiency of cultivating G. speciosum from vegetative organs grown in various media for shoot and root inductions. In addition, the study determined suitable culture conditions for enhancement of bioactive compound production under NaCl stress and the optimal immersion time in the TIB system to efficiently stimulate shoot multiplication. Furthermore, this research examined the biological properties of G. speciosum crude extracts.
2. Results and Discussion
2.1. Induction of Protocorm-like Bodies
The shoot apex and nodal segments of G. speciosum cultivated in various media induced PLBs (Figure 1a). The formation of PLBs visibly appeared after 21 days of culture. However, the explants of G. speciosum showed different growth and development. Some explants formed friable calluses that were clearly visible on the node of the explant after 14 days of culture. The friable calluses covered the node surface and varied from white to milky white (Figure 1b). Shoot formation was observed on the surface of the shoot apex and was clearly detected and showed good growth after 28 days of culture. The occurrence of these PLBs reached its highest point at 35 days and remained consistent thereafter until 42 days. The PLBs displayed vigorous growth and demonstrated the capacity to develop into mature shoots (Figure 1c-j).
The greatest number of shoots from the induction of PLBs was observed in the T2 medium ( and Table 1). Consequently, T2 was adopted as the suitable medium for shoot formation, and it had the greatest number of shoots compared to the other media. The sample with the greatest number of shoots had the highest fresh growth index. The T4 medium had the lowest shoot number; however, the shoots appeared to be fully tillering and had large stems. Therefore, its fresh growth index was not different from the control (T1), while the T3, T5, and T6 media formed similar shoot numbers to T1 but had different fresh growth index values. Samala [28] investigated the PLBs formation of G. speciosum in a conventional solid medium (½ MS + BA + NAA); they reported an increase in the number of PLBs to 6.87 shoots explant-1 after 70 days of cultivation. Notably, the current work successfully induced the formation of PLBs in a liquid medium (½ MS + 1 mg L-1 BAP + 0.5 mg L-1 NAA), resulting in 35.00 ± 3.00 shoots per explant within 42 days (Table 1).
The formation of PLBs in liquid medium with a shorter cultivation time was compared to that in the solid medium. In addition, the optimal proportion of BAP and NAA considerably promoted shoot generation in G. speciosum. The interaction between auxin and cytokinin regulates meristem development, which is important in creating the whole plant body [29]. The presence of elevated levels of cytokinin, such as BAP, exerts an influence on apical dominance by promoting the formation of axillary and adventitious shoots; conversely, higher levels of auxin have been found to activate the formation of adventitious roots [30,31]. Other studies have demonstrated that optimal shoot proliferation in different cultivars is achieved by manipulating the cytokinin-to-auxin ratio. In a study conducted by Madhulatha [32], a pulse treatment involving a combination of 6-benzylaminopurine (BAP) and kinetin in a 1:1 ratio increased shoot formation in banana (Musa spp.). Jafari [33] revealed that a concentration of 33 μM BAP stimulated the multiplication of abnormal shoots in banana (Musa acuminata cv. Berangan), while shoot elongation was supported by indole-3-acetic acid. Additionally, Rahman [34] reported that the combination of 4.0 mg L-1 BAP and 1.5 g L-1 NAA activated shoot formation in banana (Musa sapientum). The current study showed that the ½ MS medium supplemented with 1 mg L-1 BAP and 0.5 mg L-1 NAA was appropriate for shoot multiplication in G. speciosum.
Figure 1.
Morphological development of G. speciosum. (a) Nodal segment of explant. (b) Formation of friable calli from nodal segment after culture in T2 medium. (c–d) Top view of explant showing formation of PLBs from shoot apex after 14 days of culture in T2 medium. (e–f) PLB formations were clearly detected and showed well-grown explant surface after 21 days of culture in T2 medium. (g–i) Shoot development at different growth stages on explant after 28 days (g), after 35 days (h), after 42 days (i) of culture in T2 medium. (j) Shoot with different growth stages from 7 days to 42 days after subculture. (k) Plantlet with different growth stages showing root development in root induction medium (RT5) from 7 days to 28 days of culture. (l) Plantlets of G. speciosum after transplantation for 3 months.
Figure 1.
Morphological development of G. speciosum. (a) Nodal segment of explant. (b) Formation of friable calli from nodal segment after culture in T2 medium. (c–d) Top view of explant showing formation of PLBs from shoot apex after 14 days of culture in T2 medium. (e–f) PLB formations were clearly detected and showed well-grown explant surface after 21 days of culture in T2 medium. (g–i) Shoot development at different growth stages on explant after 28 days (g), after 35 days (h), after 42 days (i) of culture in T2 medium. (j) Shoot with different growth stages from 7 days to 42 days after subculture. (k) Plantlet with different growth stages showing root development in root induction medium (RT5) from 7 days to 28 days of culture. (l) Plantlets of G. speciosum after transplantation for 3 months.

Figure 2.
Effect of plant growth regulator on shoot proliferation of G. speciosum after 42 days of culture. (a) T1 medium, ½ MS. (b) T2 medium, ½ MS supplemented with 1 mg L-1 NAA and 0.5 mg L-1 BAP. (c) T3 medium, ½ MS supplemented with 1 mg L-1 NAA and 1 mg L-1 BAP. (d) T4 medium, ½ MS supplemented with 2 mg L-1 NAA and 0.5 mg L-1 BAP. (e) T5 medium, ½ MS supplemented with 2 mg L-1 NAA and 1 mg L-1 BAP. (f) T6 medium, ½ MS supplemented with 2 mg L-1 NAA and 2 mg L-1 BAP.
Figure 2.
Effect of plant growth regulator on shoot proliferation of G. speciosum after 42 days of culture. (a) T1 medium, ½ MS. (b) T2 medium, ½ MS supplemented with 1 mg L-1 NAA and 0.5 mg L-1 BAP. (c) T3 medium, ½ MS supplemented with 1 mg L-1 NAA and 1 mg L-1 BAP. (d) T4 medium, ½ MS supplemented with 2 mg L-1 NAA and 0.5 mg L-1 BAP. (e) T5 medium, ½ MS supplemented with 2 mg L-1 NAA and 1 mg L-1 BAP. (f) T6 medium, ½ MS supplemented with 2 mg L-1 NAA and 2 mg L-1 BAP.

2.2. Effect of Different Immersion Times and Frequencies on Shoot Multiplication
In the TIB system, both the immersion time and frequency were identified as crucial parameters affecting the shoot multiplication efficiency of G. speciosum (Figure 3). Notably, there were significant differences in growth parameters between immersion times of 5 min and 10 min. Immersion for 10 min every 3 h (IF4) enhanced the maximum number of shoots per explant and greatest shoot height, as well as the fresh growth index (127.00 ± 2.16 shoots per explant, 5.00 ± 0.51 cm, and 4.26 ± 0.52, respectively). According to the data in Table 2, the growth parameters of 10-min immersion decreased significantly when the immersion times reached 6 h (IF5) and 12 h (IF6), respectively. With immersion for 5 min, there were no observable differences in the number of shoots per explant or shoot height between 3, 6, and 12 h (IF1, IF2, and IF3, respectively). Nevertheless, it is worth noting that the immersion for 5 min every 3 h (IF1) resulted in the lowest number of shoots per explant and the lowest fresh growth index compared to all other immersion treatments. Ekmekçigil [35] conducted a study that showcased the effectiveness of utilizing the RITA® temporary immersion bioreactor for efficient in vitro mass propagation of Cattleya forbesii Lindl. through PLBs. Their study revealed that immersion for 1 min every 4 h produced the highest number of shoots as 111.9 shoots per explant from inoculum densities 20 explant in 250 mL medium [35]. Based on these findings, the optimal system for growth and development to increase the number of shoots of G. speciosum was immersion for 10 mins every 3 h (IF4).
2.3. Root Induction
The G. speciosum multiple shoots cultured in the TIB system were subsequently transferred to six different rooting media to induce roots (Figure 4). All media supported the growth of G. speciosum (Figure 5). Notably, successful root germination occurred in the RT1, RT2, RT5, and RT6 media. The RT5 medium promoted the highest root number and longest root length (Figure 1k, Figure 8 and Table 3). Conversely, root germination was not observed in the RT3 and RT4 media, both of which were based on the full MS medium. Additionally, the plantlets exhibited atypical growth characterized by abnormal elongated stems and unhealthy plantlet, indicating possible symptoms of excessive nutrient exposure in the G. speciosum culture. The composition of the MS medium consisted of both macro- and micro-nutrients, as outlined by Murashige and Skoog[36]. On the other hand, the VW medium comprised three macronutrients (potassium nitrate, ammonium sulfate, and monopotassium phosphate) along with essential micronutrients (magnesium sulfate, manganese sulfate, and ferric sulfate), according to the report by Vacin and Went [37]. The current results were consistent with their findings, which indicated that the VW medium provided suitable nutrients for promoting the root growth of G. speciosum (Figure 5d and Figure 8e). Therefore, nutrient uptake could potentially play a significant role in physiology [38] and morphological development. In addition, the RT5 medium contained coconut water and bananas; another study highlighted the growth-enhancing effects of coconut water on orchids, leading to elevated propagation rates [39]. Furthermore, the combined application of coconut water and banana additives reportedly enhanced root regeneration in Florida’s native orchid species [40]. Therefore, supplemented coconut water and banana in VW medium played an important role to root induction without negative effect on shoot growth of G. speciosum.
2.4. Effect of NaCl Stress on Total Phenolic Content
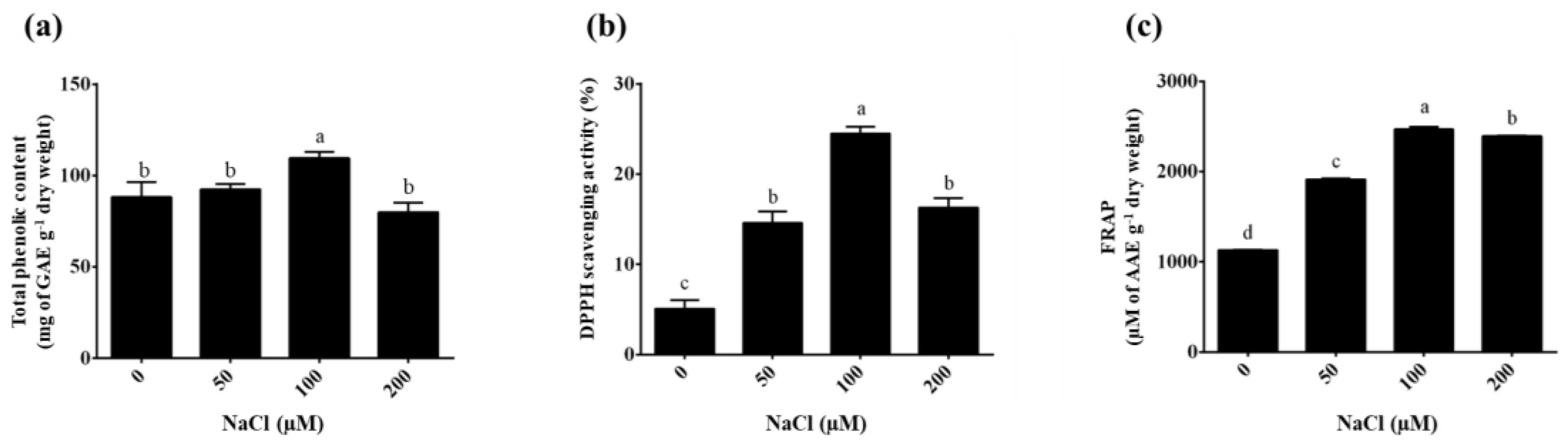
The total phenolic content of the plantlets extracted with ethyl alcohol was determined using the regression equation determined for gallic acid (y = 0.0034x + 0.0005; R² = 0.999). The highest total phenolic content was in the 100 µM NaCl treatment (111.06 ± 2.24 mg GAE g-1 DW) compared to the 0 µM NaCl treatment (control = 83.71 ± 2.47 mg GAE g-1 DW), as shown in Figure 6a. The total phenolic contents were 92.39 ± 2.86 and 82.50 ± 2.63 mg mg GAE g-1 DW in the 50 and 200 µM of NaCl treatments, respectively. Chowjarean [41] reported that the phenolic compounds analyzed from naturally grown G. speciosum ethanolic extract had a total phenolic content of 48.19 ± 0.39 mg EGCG equivalent g-1 extract. Cultivating G. speciosum using aseptic techniques can significantly reduce the required growth period to produce high-quality biological extracts. Importantly, this method does not reduce the total quantity of phenolic compounds. Furthermore, this approach has the potential to regulate the increase in total phenolic content by incorporating nutrients (NaCl) into the cultivation medium. This approach aligns with research by Nag & Kumaria [42] which indicated that NaCl can stimulate the production of phenolic compounds in Vanda coerulea Griff. ex Lindl. Similarly, the current findings showed a significant increase in the total phenolic content after NaCl treatment, which is responsible for protecting the plant from the harmful influences of salt stress [43,44,45,46]. However, notably, in some plants, exposure to high salinity can lead to ion imbalances, hyper-osmotic stress, and subsequent osmotic and oxidative stresses [47]. In addition, salinity stress stimulates the phenylpropanoid biosynthetic pathway; accordingly, it supports phenolic compound accumulation [48]. According to these reports, salinity acts as an elicitor to induce the accumulation of phenolic compounds in plants. The study of environmental stress combinations is an interesting concept that has led to high levels of secondary metabolite production in G. speciosum.
2.5. Antioxidant Activity
The antioxidant activity of G. speciosum extract based on DPPH and FRAP assays is shown in . The G. speciosum extracts treated with 100 µM NaCl showed the highest potential DPPH scavenging activity compared with the untreated condition (0 µM NaCl) (24.50 ± 0.76% and 3.93 ± 0.36%, respectively), as shown in Figure 6b. The DPPH scavenging activity levels in 50 and 200 µM NaCl were 14.59 ± 1.04% and 16.29 ± 0.84%, respectively. Previously, the percentage of DPPH scavenging activity in Cottonia peduncularis and Dendrobium spp. orchids were 29.83% and 32.58%, respectively [49,50]. The FRAP assay is based on the capacity of antioxidants to reduce the [Fe(III)(TPTZ)2]3+ complexes to [Fe(II)(TPTZ)2]2+. The FRAP value was calculated using a standard curve of ascorbic acid concentrations ranging from 0 to 1,000 µM (y = 0.0012x + 0.0022; R² = 0.999). The G. speciosum extracts treated with 100 µM NaCl had reducing activity within the range 2441.79 ± 1.21 to 2491.96 ± 3.23 µM AAE g-1 DW (Figure 6c). The FRAP values were within the range 1129.48 ± 0.57 to 1132.62 ± 1.00 for the control (0 µM NaCl). The G. speciosum extracts treated with 50 and 200 µM NaCl have a reducing activity within the range 1904.17 ± 3.09 to 1927.74 ± 2.69 µM AAE g-1 DW and 2385.85 ± 3.35 to 2397.88 ± 2.68 µM AAE g-1 DW, respectively. In addition, these results presented a positive correlation between the total phenolic content and the antioxidant capacity in the G. speciosum plantlets treated with 100 µM NaCl. Elsewhere, the radical scavenging activity in Luisia zeylanica [51] and Nepal wild orchids [52] had antioxidant activity levels that were correlated positively with total polyphenolics. The results indicated that salt stress could stimulate antioxidant activity in the plantlets of G. speciosum under TIB system.
Figure 6.
Total phenolic content and antioxidant activity levels of G. speciosum extracts under salinity treatments for 72 h. (a) Total phenolic content. (b) Percentage of DPPH scavenging activity. (c) FRAP values. Data represent mean ± SD values of ten replications. Different lowercase letters indicate significant (p<0.05) differences among NaCl concentrations based on Tukey's multiple comparisons test.
Figure 6.
Total phenolic content and antioxidant activity levels of G. speciosum extracts under salinity treatments for 72 h. (a) Total phenolic content. (b) Percentage of DPPH scavenging activity. (c) FRAP values. Data represent mean ± SD values of ten replications. Different lowercase letters indicate significant (p<0.05) differences among NaCl concentrations based on Tukey's multiple comparisons test.
2.6. Antibacterial Activity
From the above results, the highest total phenolic content was observed in the G. speciosum plantlets treated with 100 µM NaCl. Consequently, they were selected for the antibacterial activity evaluation. The G. speciosum crude extracts showed antibacterial action with the minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) values (Table 4). The different bacteria were inhibited growth and eliminated at different concentration of the extract. P. acnes (6.4 mg mL-1) had the lowest MIC, followed by S.aureus (12.8 mg mL-1) and S. epidermidis (25.6 mg mL-1). Each bacteria species died when it was treated with 2-fold extract concentration of MIC that noticed from MBC in Table 4. Other studies reported that the bioactive compounds isolated from G. speciosum extract included gastrodin [41,53] and vitexin and orientin [4]. Gastrodin is a phenolic glycoside [54], whereas vitexin and orientin are flavonoid derivatives [4] belonging to a group of phenolic compounds that can be found in plants. Polyphenols exhibit antimicrobial activity through interactions with bacterial cell surfaces and disruption of the membrane integrity [55,56]. González-Cortazar [57] reported that gastrodin was active against S. aureus (MIC> 0.1 mg mL-1). Likewise, the MIC values of vitexin (> 1.0 mg mL-1) and orientin (0.5 mg mL-1) showed they have antibacterial activity against S. aureus; however, the mechanism of action is still unknown [58]. In addition, other orchid extracts have reported activity against acne bacteria. Irimescu [59] and Olivares [60] reported that different parts of Phalaenopsis orchids (roots, leaves, and stems) extracts had inhibitory activity against different bacteria. They found that the stem and root extracts could be antimicrobial agents for S. aureus and S. epidermidis, respectively [59]. The mixture of root and leave extracts showed the antimicrobial activity against P. acnes [60]. The inhibitory activity could be linked to the presence of the bioactive compound in the orchid extracts as the occurrence of phenolic in G. speciosum plantlets revealed a potential of antimicrobial activity against pathogens causing acne.
3. Materials and Methods
Plant Materials and Induction of Protocorm-like Bodies
The shoot apex and nodal segment of Grammatophyllum speciosum were cultured in half-strength Murashige and Skoog (½ MS) liquid medium [36] supplemented with 20 g L–1 sucrose and various concentrations of 6-benzylaminopurine (BAP) at 0.5, 1.0 or 2.0 mg L–1 and 1-naphthaleneacetic acid (NAA) at 1.0 and 2.0 mg L–1.
The pH of each liquid medium was adjusted to 5.7. Then, 20 mL of each medium was dispersed into a 250 mL tissue culture bottle. Subsequently, all media were sterilized using an autoclave (Zealway, Germany) at 121 °C and 105 kPa pressure for 20 mins. The cultures were shaken at 110 revolutions per min at 25 ± 1 °C and humidity of 51 ± 2% under white light-emitting diode (LED) light at an intensity of 37 µmol m-2 s-1 for 16 h day-1 for 42 days.
Optimization of High Efficiency Shoot Multiplication
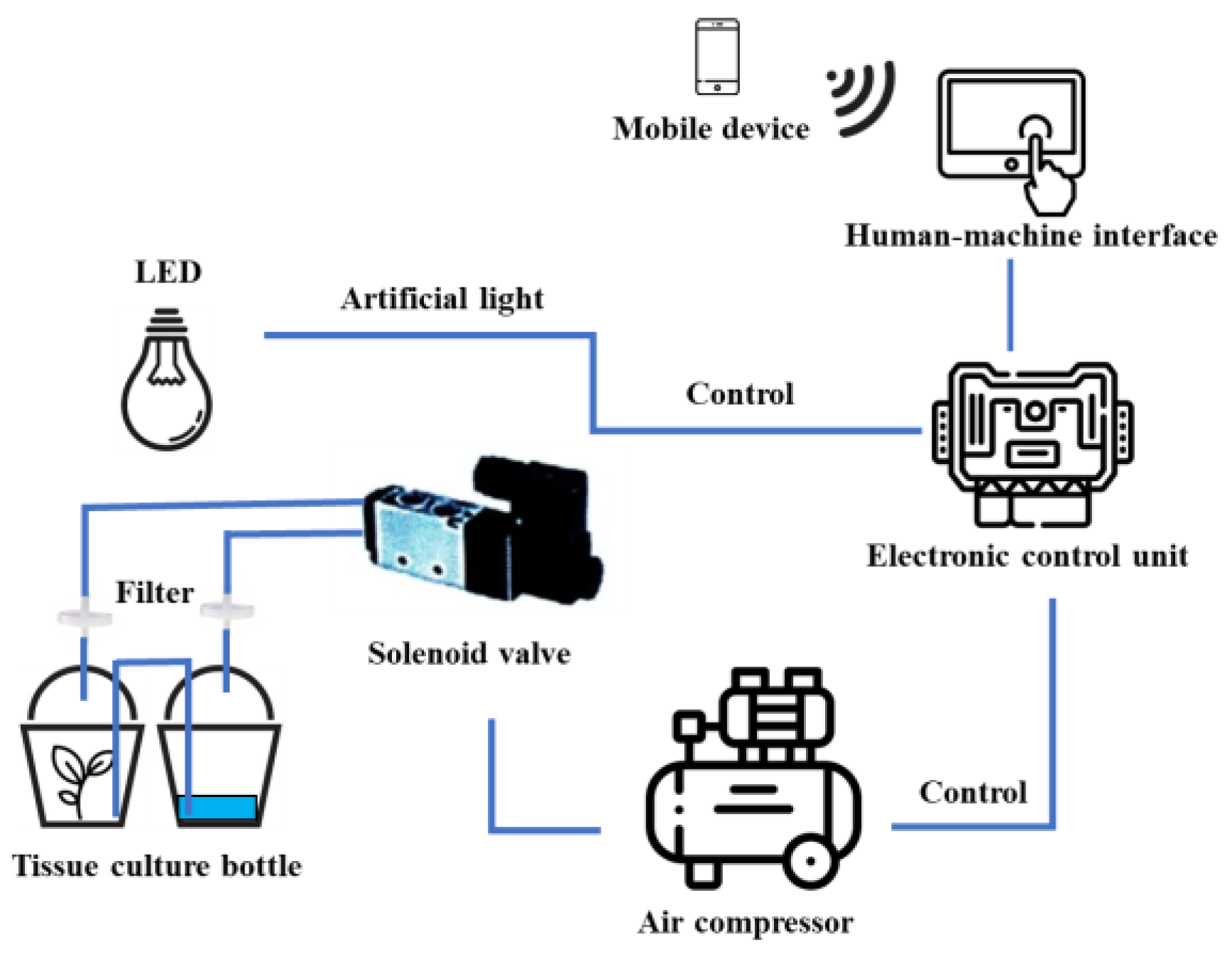
A sample of 0.5 cm protocorm-like bodies (PLBs) was inoculated into a 720 mL glass chamber of a temporary immersion bioreactor (TIB). Another chamber contained 100 mL of ½ MS liquid medium supplemented with 20 g L-1 sucrose, 1 mg L-1 6-benzylaminopurine (BAP), and 0.5 mg L-1 1-naphthaleneacetic acid (NAA), as shown in Figure 7. The growth chamber was equipped with an air compressor and a controller box programmed to regulate the frequency and duration of medium supply (Figure 8). The immersion frequency was compared at intervals of 3, 6, or 12 h. In each case, the PLBs were immersed for 5 or 10 min. The experimental setup allowed for the evaluation of how different immersion intervals impacted the growth process of the PLBs. The TIB system was maintained at 25 ± 1 °C and humidity of 51 ± 2% under white LED light at an intensity of 40 µmol m-2 s-1 for 16 h day-1 for 1 month.
Figure 7.
Shoot proliferation in glass chamber of temporary immersion bioreactor with twin flask system.
Figure 7.
Shoot proliferation in glass chamber of temporary immersion bioreactor with twin flask system.

Figure 8.
Schematic diagram of temporary immersion bioreactor system integrated with Internet of Things (IoT) technology.
Figure 8.
Schematic diagram of temporary immersion bioreactor system integrated with Internet of Things (IoT) technology.
Root Induction
The sample of 2.0 cm shoot length of G. speciosum was cultured in six different rooting media. RT1: ½ MS medium supplemented with 0.5 mg L-1 NAA and 1 mg L-1 BAP, RT2: ½ MS medium supplemented with 0.5 mg L-1 NAA, RT3: MS medium supplemented with 0.5 mg L-1 NAA and 1 mg L-1 BAP, RT4: MS medium supplemented with 0.5 mg L-1 NAA, RT5: VW medium [37] supplemented with 150 mL L-1 coconut water, 100 g L-1 banana and 50 g L-1 potato, and RT6: VW medium. The culture media contained 20 g L-1 sucrose and 1 g L-1 charcoal and was solidified using 7.2 g L-1 agar. The pH of the media was adjusted to 5 using 1 M HCl and NaOH. The cultures were maintained at 25 ± 1 °C and humidity of 51 ± 2% under white LED light at an intensity of 40 µmol m-2 s-1 for 16 h day-1 for 1 month.
Morphological Evaluation
Different stages of development in G. speciosum were observed every 7 days of culture. Fresh explants were selected and observed under a stereo microscope with camera (Olympus SZ 40) for morphological studies. The morphological parameters of the explant, including the number of shoots and roots as well as the maximum length of shoots, roots, and leaves, were recorded following cultivation in different culture media. The fresh growth index was assessed after culture using the following equation.
Sodium Chloride Stress Treatments
The 1-year-old plantlets of G. speciosum were cultured in ½ MS liquid medium supplemented with 20 g L-1 sucrose, 1 g L-1 activated charcoal, and sodium chloride (NaCl) at 0, 50, 100 or 200 µM to induce the total phenolic content from in vitro culture for 72 h. The cultures were maintained at 25 ± 1 °C and humidity of 51 ± 2% under white LED light at an intensity of 40 µmol m-2 s-1 for 16 h day-1 using the TIB system (IF4, 10 min immersion time every 3 h).
Preparation of Crude Extract
The NaCl-treated plantlets were collected and dried in a hot-air oven at 60 °C for 2 days. The dried samples (each 100 mg) were ground separately to a fine powder and soaked in 100 mL of ethyl alcohol. The extracts were incubated at room temperature for 3 days and shaken occasionally. After incubation, the extracts passed through WhatmanTM No.1 qualitative filter paper (Merck, Germany) and concentrated at 40 °C using a speed vacuum concentrator (Eppendorf, Germany). The obtained semisolid extracts were stored at -20 °C.
Quantification of Total Phenolic Content
The total phenolic content was determined using the Folin-Ciocalteu colorimetric method [61]. The reaction mixture was created in a 96-well plate by combining 20 µL of each extract (dissolved in ethyl alcohol) and 100 µL of 10% (v/v) Folin-Ciocalteu reagent (VWR Chemicals, France). The reaction mixture was incubated at room temperature for 3 min. Then, 80 µL of 1 M sodium carbonate (Na2CO3) was added and incubated at room temperature in the dark for 20 min. The absorbance was measured at 765 nm using a microplate reader (Tecan, Switzerland). Gallic acid (Sigma-Aldrich, USA) at 0, 0.0125, 0.025, 0.05, 0.1, or 0.2 mg mL-1 was used as a calibrated standard. The results were expressed in milligrams gallic acid equivalent per gram of dry weight (mg GAE g-1 DW).
DPPH Radical Scavenging Assay
The antioxidant activity of extracts was determined using 2,2-diphenyl-2-picrylhydrazyl (DPPH), according to the method of Herald [62]. A sample (20 µL) of the extract was mixed with 200 µL of 1.5 mM DPPH solution in ethyl alcohol and incubated at room temperature in the dark for 30 min. The absorbance was measured at 517 nm using a microplate reader. Ascorbic acid (Fisher Scientific, UK) at 0, 2.5, 5, 10, 20, 40, 80, or 160 µg mL-1 was used as a calibrated standard. The results were expressed as the percentage of DPPH scavenging activity using the following equation.
where, A0 is the absorbance of the control and A1 is the absorbance of the extract.
DPPH scavenging activity (%) = [(A0 - A1) / A0] × 100
Ferric Reducing Antioxidant Power Assay
The ferric reducing antioxidant power (FRAP) assay was carried out using the method described by Benzie and Strain [63], with minor modifications for use in a 96-well microplate. The FRAP reagent was prepared by mixing 300 mM sodium acetate buffer (pH 3.6), 10 mM 2,4,6-tripyridyl-s-triazine (TPTZ) in 40 mM hydrochloric acid, and 20 mM ferric chloride hexahydrate (FeCl3·6H2O) solution in the ratio 10:1:1 at 37 °C in the dark. Then, 20 µL of each extract was mixed with 280 µL of freshly prepared working FRAP reagent. The reaction mixture was incubated at 37 °C in the dark for 30 min. The absorbance of the ferrous-tripyridyltriazine (Fe (II)-TPTZ) complex blue color was measured at 593 nm using a microplate reader. Ascorbic acid (at 0, 62.5, 125, 250, 500, or 1,000 µM was used to build up a calibration curve. The FRAP value was expressed in micromoles of ascorbic acid equivalent per gram of dry weight (µM AAE g-1 DW).
Antibacterial Activity
Staphylococcus aureus (DMST 8840), Staphylococcus epidermidis (DMST 15505), and Propionibacterium acnes (DMST 14916) were obtained from the Herbal and Bioactive Substances Technology Laboratory, Kasetsart Agricultural and Agro-Industrial Product Improvement Institute (KAPI), Bangkok, Thailand. The two aerobic bacteria (S. aureus and S. epidermidis) were cultured on tryptone soy agar at 37 °C for 24 h. The aero-tolerant anaerobic bacterium (P. acnes) was cultured on brain heart infusion agar at 37 °C for 72 h under anaerobic conditions. The colonies were evenly suspended in 0.85% NaCl and diluted to a turbidity equivalent to McFarland No. 0.5 (108 colony forming units mL-1).
The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) were determined using the two-fold serial microdilution method [64]. The diluted extracts (in dimethyl sulfoxide) were prepared at concentrations in the range 0.05–102.4 mg mL-1 with sterile Mueller-Hinton broth (for S. aureus and S. epidermidis) and brain heart infusion broth (for P. acnes) before adding the bacterial suspensions (each 100 µL) into the 96-well microtiter plates. S. aureus and S. epidermidis were incubated at 37 °C for 24 h, while P. acnes was incubated at 37 °C under anaerobic conditions for 72 h. The bacterial suspensions were used as positive controls and the crude extracts in broth were used as negative controls. The MIC values were defined as the lowest concentration of the crude extracts to inhibit the growth of microorganisms. MBC was determined by inoculating a broth with no visible bacterial growth onto agar plates. The MBC values were defined as the lowest concentration of the extracts needed to kill each bacterium.
Statistical Analysis
Comparisons of the data between the treatments were carried out based on Tukey's multiple comparisons test, with significance tested at p≤0.05 using GraphPad Prism 6 software (GraphPad Software, Inc., USA). Results were expressed as mean ± standard deviation (SD) values.
4. Conclusions
The current effectively identified an appropriate medium for both shoot and root induction. In addition, the application of the TIB system was successful in enhancing the micro-propagation of G. speciosum under controlled and sterile conditions. The efficient TIB technology described herein was the first reported for cultivating G. speciosum. This methodology not only ensures heightened quality but also offers remarkable efficiency in the production of bioactive compounds. These findings demonstrated that NaCl stress played a role in promoting the accumulation of phenolic compounds in G. speciosum. This intensified presence of phenolic compounds was correlated directly with antioxidant capacity. Furthermore, the extracted compounds exhibited activity against bacteria responsible for causing acne. In Thailand, G. speciosum extract is utilized in various treatments. Upcoming studies will focus on evaluating the biological activities of G. speciosum extracts, particularly their anti-skin cancer and anti-bacterial properties, aiming to promote the medical applications of Thai medicinal herbs. Overall, this TIB process should provide a strategic advantage, offering a promising avenue for producing high-quality secondary metabolites that hold great potential for pharmaceutical industries.
Author Contributions
P. Tongkok and W. Phonphoem designed the experimental setup. G. Matmarurat performed experiments. J. Chusrisom and N.P. T-Tienprasert analysed data and biochemical analyses results. J. Chusrisom and P. Tongkok wrote the manuscript. All authors have read and approved the last draft of the manuscript.
Funding
This work was funded and supported by the Kasetsart University Research and Development Institute (KURDI), Bangkok, Thailand under project FF (KU) 32.66.
Acknowledgments
The authors would like to thank Dr. Andrew John Warner (KURDI, KU) for improving the English of this manuscript.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- Yam, T.W.; Chua, J.; Tay, F.; Ang, P. Conservation of the Native Orchids Through Seedling Culture and Reintroduction—A Singapore Experience. Bot. Rev. 2010, 76, 263–274. [Google Scholar] [CrossRef]
- Teoh, E.S. Medicinal orchids of Asia; Springer, Switzerland: 2016.
- Chowjarean, V.; Sadabpod, K.; Mejía-Aranguré, J. Antiproliferative Effect of Grammatophyllum speciosum Ethanolic Extract and Its Bioactive Compound on Human Breast Cancer Cells. Sci. World J. 2021, 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yingchutrakul, Y.; Sittisaree, W.; Mahatnirunkul, T.; Chomtong, T.; Tulyananda, T.; Krobthong, S. Cosmeceutical Potentials of Grammatophyllum speciosum Extracts: Anti-Inflammations and Anti-Collagenase Activities with Phytochemical Profile Analysis Using an Untargeted Metabolomics Approach. Cosmetics 2021, 8, 116. [Google Scholar] [CrossRef]
- Chowjarean V, Phiboonchaiyanan PP, Harikarnpakdee S. Skin brightening efficacy of grammatophyllum speciosum: a prospective, split-face, randomized placebo-controlled study. Sustainability [Internet]. 2022; 14(24).
- Etienne, H.; Berthouly, M. Temporary immersion systems in plant micropropagation. Plant Cell, Tissue Organ Cult. (PCTOC) 2002, 69, 215–231. [Google Scholar] [CrossRef]
- Rathore J, Rathore V, Shekhawat N, Singh R, Liler G, Phulwaria M, et al. Micropropagation of woody plants. Plant biotechnology and molecular markers: Springer; 2004. p. 195-205.
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; García-Lara, S. In vitro plant tissue culture: means for production of biological active compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Esyanti, R.R.; Adhitama, N.; Manurung, R. Efficiency Evaluation of Vanda Tricolor Growth in Temporary Immerse System Bioreactor and Thin Layer Culture System. J. Adv. Agric. Technol. 2016, 3. [Google Scholar] [CrossRef]
- Alvard D, Cote F, Teisson C. Comparison of methods of liquid medium culture for banana micropropagation: Effects of temporary immersion of explants. Plant cell, tissue and organ culture. 1993;32:55-60.
- Lorenzo, J.C.; Ojeda, E.; Espinosa, A.; Borroto, C. Field performance of temporary immersion bioreactor-derived sugarcane plants. Vitr. Cell. Dev. Biol. - Plant 2001, 37, 803–806. [Google Scholar] [CrossRef]
- Reis COD, Silva ABD, Landgraf PRC, Batista JA, Jacome GAR. Bioreactor in the micropropagation of ornamental pineapple. Ornamental Horticulture. 2018;24:182-7.
- Bello-Bello, J.J.; Schettino-Salomón, S.; Ortega-Espinoza, J.; Spinoso-Castillo, J.L. A temporary immersion system for mass micropropagation of pitahaya (Hylocereus undatus). 3 Biotech 2021, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S.; Solis-Gracia, N.; Teale, M.K.; Mandadi, K.; da Silva, J.A.; Vales, M.I. Development of an in vitro Microtuberization and Temporary Immersion Bioreactor System to Evaluate Heat Stress Tolerance in Potatoes (Solanum tuberosum L.). Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- De Carlo, A.; Tarraf, W.; Lambardi, M.; Benelli, C. Temporary Immersion System for Production of Biomass and Bioactive Compounds from Medicinal Plants. Agronomy 2021, 11, 2414. [Google Scholar] [CrossRef]
- Rout, G.; Samantaray, S.; Das, P. In vitro manipulation and propagation of medicinal plants. Biotechnol. Adv. 2000, 18, 91–120. [Google Scholar] [CrossRef] [PubMed]
- Le, K.-C.; Dedicova, B.; Johansson, S.; Lelu-Walter, M.-A.; Egertsdotter, U. Temporary immersion bioreactor system for propagation by somatic embryogenesis of hybrid larch (Larix × eurolepis Henry). Biotechnol. Rep. 2021, 32, e00684. [Google Scholar] [CrossRef] [PubMed]
- Farahani F, Majd A. Comparison of liquid culture methods and effect of temporary immersion bioreactor on growth and multiplication of banana (Musa, cv. Dwarf Cavendish). African Journal of Biotechnology. 2012;11(33):8302-8.
- Ahmadian, M.; Babaei, A.; Shokri, S.; Hessami, S. Micropropagation of carnation ( Dianthus caryophyllus L. ) in liquid medium by temporary immersion bioreactor in comparison with solid culture. J. Genet. Eng. Biotechnol. 2017, 15, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Vendrame, W.A.; Xu, J.; Beleski, D.G. Micropropagation of Brassavola nodosa (L.) Lindl. using SETIS™ bioreactor. Plant Cell, Tissue Organ Cult. (PCTOC) 2023, 153, 67–76. [Google Scholar] [CrossRef]
- Fritsche, Y.; Deola, F.; da Silva, D.A.; Holderbaum, D.F.; Guerra, M.P. Cattleya tigrina (Orchidaceae) in vitro regeneration: Main factors for optimal protocorm-like body induction and multiplication, plantlet regeneration, and cytogenetic stability. South Afr. J. Bot. 2022, 149, 96–108. [Google Scholar] [CrossRef]
- Cui, H.-Y.; Murthy, H.N.; Moh, S.H.; Cui, Y.-Y.; Lee, E.-J.; Paek, K.-Y. Production of biomass and bioactive compounds in protocorm cultures of Dendrobium candidum Wall ex Lindl. using balloon type bubble bioreactors. Ind. Crop. Prod. 2014, 53, 28–33. [Google Scholar] [CrossRef]
- Zhang B, Niu Z, Zhou A, Zhang D, Xue Q, Liu W, et al. Micropropagation of Dendrobium nobile Lindl. plantlets by temporary immersion bioreactor. Journal of Biobased Materials and Bioenergy. 2019;13(3):395-400.
- Kang F, Hsu S, Shen R. Virus elimination through meristem culture and rapid clonal propagation by temporary immersion system in Phalaenopsis. Journal of the Taiwan Society for Horticultural Science. 2011;57(3):207-18.
- Spinoso-Castillo, J.L.; Chavez-Santoscoy, R.A.; Bogdanchikova, N.; Pérez-Sato, J.A.; Morales-Ramos, V.; Bello-Bello, J.J. Antimicrobial and hormetic effects of silver nanoparticles on in vitro regeneration of vanilla (Vanilla planifolia Jacks. ex Andrews) using a temporary immersion system. Plant Cell, Tissue Organ Cult. (PCTOC) 2017, 129, 195–207. [Google Scholar] [CrossRef]
- Obchant, T. Orchids of Thailand. Baanlaesuan. 2003.
- Nitcha, L. Academic documents "Grammatophyllum speciosum". 2016.
- Samala, S.; Te-Chato, S.; Yenchon, S.; Thammasiri, K. Protocorm-like body proliferation of Grammatophyllum speciosum through asymbiotic seed germination. ScienceAsia 2014, 40. [Google Scholar] [CrossRef]
- Su, Y.-H.; Liu, Y.-B.; Zhang, X.-S. Auxin–Cytokinin Interaction Regulates Meristem Development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Mangena, P. Benzyl adenine in plant tissue culture-succinct analysis of the overall influence in soybean [Glycine max (L.) Merrill.] seed and shoot culture establishment. Journal of Biotech Research. 2020;11:23-34.
- Moubayidin, L.; Di Mambro, R.; Sabatini, S. Cytokinin–auxin crosstalk. Trends Plant Sci. 2009, 14, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Madhulatha, P.; Anbalagan, M.; Jayachandran, S.; Sakthivel, N. Influence of Liquid Pulse Treatment with Growth Regulators on in vitro Propagation of Banana (Musa spp. AAA). Plant Cell, Tissue Organ Cult. (PCTOC) 2004, 76, 189–192. [Google Scholar] [CrossRef]
- Jafari N, Othman RY, Khalid N. Effect of benzylaminopurine (BAP) pulsing on in vitro shoot multiplication of Musa acuminata (banana) cv. Berangan. African Journal of Biotechnology. 2011;10:2446-50.
- Rahman, M. In vitro Response and Shoot Multiplication of Banana with BAP and NAA. Asian J. Plant Sci. 2004, 3, 406–409. [Google Scholar] [CrossRef]
- Ekmekçigil, M.; Bayraktar, M.; Akkuş, Ö.; Gürel, A. High-frequency protocorm-like bodies and shoot regeneration through a combination of thin cell layer and RITA® temporary immersion bioreactor in Cattleya forbesii Lindl. Plant Cell, Tissue Organ Cult. (PCTOC) 2018, 136, 451–464. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Vacin, E.F.; Went, F.W. Some pH Changes in Nutrient Solutions. Bot. Gaz. 1949, 110, 605–613. [Google Scholar] [CrossRef]
- Adelberg, J.W.; Delgado, M.P.; Tomkins, J.T. Spent medium analysis for liquid culture micropropagation of Hemerocallis on Murashige and Skoog medium. Vitr. Cell. Dev. Biol. - Plant 2009, 46, 95–107. [Google Scholar] [CrossRef]
- Salsabila, S.N.; Fatimah, K.; Noorhazira, S.; Halimatun, T.S.T.A.B.; Aurifullah, M.; Suhana, Z. Effect of Coconut Water and Peptone in Micropropagation of Phalaenopsis amabilis (L.) Blume Orchid.CONFERENCE NAME, LOCATION OF CONFERENCE, COUNTRYDATE OF CONFERENCE; p. 012002.
- De Stefano, D.; Costa, B.N.S.; Downing, J.; Fallahi, E.; Khoddamzadeh, A.A. In-Vitro Micropropagation and Acclimatization of an Endangered Native Orchid Using Organic Supplements. Am. J. Plant Sci. 2022, 13, 380–393. [Google Scholar] [CrossRef]
- Chowjarean V, Sucontphunt A, Vchirawongkwin S, Charoonratana T, Songsak T, Harikarnpakdee S, et al. Validated RP-HPLC method for quantification of gastrodin in ethanolic extract from the pseudobulbs of grammatophyllum speciosum blume. Malays. J. Anal. Sci. 2018, 22. [Google Scholar] [CrossRef]
- Nag, S.; Kumaria, S. In silico characterization and transcriptional modulation of phenylalanine ammonia lyase (PAL) by abiotic stresses in the medicinal orchid Vanda coerulea Griff. ex Lindl. Phytochemistry 2018, 156, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Khenifi; M, L. ; Boudjeniba; Kameli Effects of salt stress on micropropagation of potato (Solanum tuberosum L.). Afr. J. Biotechnol. 2011, 10, 7840–7845. [Google Scholar] [CrossRef]
- Joshi, M.; Mishra, A.; Jha, B. NaCl plays a key role for in vitro micropropagation of Salicornia brachiata, an extreme halophyte. Ind. Crop. Prod. 2012, 35, 313–316. [Google Scholar] [CrossRef]
- Alturki, S. Effect of NaCl on Growth and Development of in vitro Plants of Date Palm (Phoenix dactylifera L.) ‘Khainazi’ Cultivar. Asian J. Plant Sci. 2018, 17, 120–128. [Google Scholar] [CrossRef]
- Dogan, M. Effect of salt stress on in vitro organogenesis from nodal explant of Limnophila aromatica (Lamk.) Merr. and Bacopa monnieri (L.) Wettst. and their physio-morphological and biochemical responses. Physiol. Mol. Biol. Plants 2020, 26, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Li Y, Zhang Y, Feng F, Liang D, Cheng L, Ma F, et al. Overexpression of a Malus vacuolar Na+/H+ antiporter gene (MdNHX1) in apple rootstock M. 26 and its influence on salt tolerance. Plant Cell, Tissue and Organ Culture (PCTOC). 2010;102:337-45.
- Kumar, K.; Debnath, P.; Singh, S.; Kumar, N. An Overview of Plant Phenolics and Their Involvement in Abiotic Stress Tolerance. Stresses 2023, 3, 570–585. [Google Scholar] [CrossRef]
- Nagananda G, Rajath S, Shankar PA, Rajani ML. Phytochemical evaluation and in vitro free radical scavenging activity of successive whole plant extract of orchid Cottonia Peduncularis. Res Art Biol Sci. 2013;3:91.
- Athipornchai, A.; Jullapo, N. Tyrosinase inhibitory and antioxidant activities of Orchid (Dendrobium spp.). South Afr. J. Bot. 2018, 119, 188–192. [Google Scholar] [CrossRef]
- Rashmi, K.; Shweta, S.; Sudeshna, C.; Vrushala, P.; Kekuda, T.; Raghavendra, H. Antibacterial and Radical Scavenging Activity of Selected Orchids of Karnataka, India. Sci. Technol. Arts Res. J. 2015, 4, 160. [Google Scholar] [CrossRef]
- Chand MB, Paudel MR, Pant B. The antioxidant activity of selected wild orchids of Nepal. Journal of Coastal Life Medicine. 2016;4(9):731-6.
- Sahakitpichan, P.; Mahidol, C.; Disadee, W.; Chimnoi, N.; Ruchirawat, S.; Kanchanapoom, T. Glucopyranosyloxybenzyl derivatives of (R)-2-benzylmalic acid and (R)-eucomic acid, and an aromatic glucoside from the pseudobulbs of Grammatophyllum speciosum. Tetrahedron 2013, 69, 1031–1037. [Google Scholar] [CrossRef]
- 54. Duan X-H, Li Z-L, Yang D-S, Zhang F-L, Lin Q, Dai R. Study on the chemical constituents of Gastrodia elata. Journal of Chinese medicinal materials 2013, 36, 1608–11.
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial Activity and Mode of Action of Ferulic and Gallic Acids Against Pathogenic Bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) Models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- González-Cortazar, M.; López-Gayou, V.; Tortoriello, J.; Domínguez-Mendoza, B.E.; Ríos-Cortes, A.M.; Delgado-Macuil, R.; Hernández-Beteta, E.E.; Blé-González, E.A.; Zamilpa, A. Antimicrobial gastrodin derivatives isolated from Bacopa procumbens. Phytochem. Lett. 2019, 31, 33–38. [Google Scholar] [CrossRef]
- Adamczak, A.; Ożarowski, M.; Karpiński, T.M. Antibacterial Activity of Some Flavonoids and Organic Acids Widely Distributed in Plants. J. Clin. Med. 2019, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Irimescu LS, Digută CF, Encea RŞ, Matei F. Preliminary study on the antimicrobial potential of phalaenopsis orchids methanolic extracts. Scientific Bulletin Series F Biotechnologies. 2020;24(2):149-53.
- Olivares CG, editor final degree project of biotechnology studies on the inhibitory potential of orchid extracts against acne associated bacteria 2020.
- Attard, E. A rapid microtitre plate Folin-Ciocalteu method for the assessment of polyphenols. Open Life Sci. 2013, 8, 48–53. [Google Scholar] [CrossRef]
- Herald, T.J.; Gadgil, P.; Tilley, M. High-throughput micro plate assays for screening flavonoid content and DPPH-scavenging activity in sorghum bran and flour. J. Sci. Food Agric. 2012, 92, 2326–2331. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. http://linkinghub.elsevier.com/retrieve/pii/S0003269796902924.
- Basri, D.; Fan, S. The potential of aqueous and acetone extracts of galls ofQuercus infectoriaas antibacterial agents. Indian J. Pharmacol. 2005, 37, 26. [Google Scholar] [CrossRef]
Figure 3.
Shoot multiplication from protocorm-like bodies under different immersion times and frequencies within TIB system after 1 month. (a) IF1: 5 min immersion time every 3 h. (b) IF2: 5 min immersion time every 6 h. (c) IF3: 5 min immersion time every 12 h. (d) IF4: 10 min immersion time every 3 h. (e) IF5: 10 min immersion time every 6 h. (f) IF6: 10 min immersion time every 12 h.
Figure 3.
Shoot multiplication from protocorm-like bodies under different immersion times and frequencies within TIB system after 1 month. (a) IF1: 5 min immersion time every 3 h. (b) IF2: 5 min immersion time every 6 h. (c) IF3: 5 min immersion time every 12 h. (d) IF4: 10 min immersion time every 3 h. (e) IF5: 10 min immersion time every 6 h. (f) IF6: 10 min immersion time every 12 h.

Figure 4.
Overview of G. speciosum propagation. (a) Development of PLBs for shoot induction in TIB system (IF4) after 7 days. (b) Shooting in TIB system after 1 month. (c) Multiplication of shoots within TIB system after 3 months. (d) Plantlets on RT5 medium after 3 months.
Figure 4.
Overview of G. speciosum propagation. (a) Development of PLBs for shoot induction in TIB system (IF4) after 7 days. (b) Shooting in TIB system after 1 month. (c) Multiplication of shoots within TIB system after 3 months. (d) Plantlets on RT5 medium after 3 months.

Figure 5.
Overview of G. speciosum propagation. (a) Development of PLBs for shoot induction in TIB system (IF4) after 7 days. (b) Shooting in TIB system after 1 month. (c) Multiplication of shoots within TIB system after 3 months. (d) Plantlets on RT5 medium after 3 months.
Figure 5.
Overview of G. speciosum propagation. (a) Development of PLBs for shoot induction in TIB system (IF4) after 7 days. (b) Shooting in TIB system after 1 month. (c) Multiplication of shoots within TIB system after 3 months. (d) Plantlets on RT5 medium after 3 months.


Table 1.
Effect of PLB induction media on morphological development of G. speciosum. Values are the means of 10 replicates ± SD. Different letters indicate significant difference between treatments based on Tukey’s multiple comparisons test (p≤0.05).
Table 1.
Effect of PLB induction media on morphological development of G. speciosum. Values are the means of 10 replicates ± SD. Different letters indicate significant difference between treatments based on Tukey’s multiple comparisons test (p≤0.05).
| Treatment |
Hormone (mg/L) |
Number of shoots explant-1 | Fresh growth index | |||||
| BAP | NAA | 28 days | 35 days | 42 days | 28 days | 35 days | 42 days | |
| T1 | - | - | 9.00 ± 1.73b | 18.33 ± 1.53b | 19.33 ± 1.15b | 0.63 ± 0.11 | 5.44 ± 0.46b | 6.69 ± 0.40b |
| T2 | 1.0 | 0.5 | 16.00 ± 1.73a | 32.33 ± 2.52a | 35.00 ± 3.00a | 1.00 ± 0.34 | 8.78 ± 2.09a | 9.92 ± 1.31a |
| T3 | 1.0 | 1.0 | 7.67 ± 0.58bc | 18.00 ± 2.00b | 18.67 ± 2.08b | 0.55 ± 0.15 | 5.14 ± 0.77b | 6.41 ± 0.77bc |
| T4 | 2.0 | 0.5 | 4.33 ± 2.31c | 10.67 ± 1.15c | 12.33 ± 1.53c | 0.23 ± 0.07 | 4.44 ± 1.07bc | 4.88 ± 0.60cd |
| T5 | 2.0 | 1.0 | 8.00 ± 1.00bc | 12.67 ± 2.08c | 13.67 ± 1.53c | 0.66 ± 0.12 | 4.89 ± 0.19bc | 5.83 ± 0.06bc |
| T6 | 2.0 | 2.0 | 5.67 ± 1.15bc | 6.33 ± 1.15d | 6.67 ±0.58d | 0.38 ± 0.06 | 3.17 ± 1.11c | 3.82 ± 0.77d |
Table 2.
Effect of different immersion times and frequencies on shoot multiplication of G. speciosum after 1 month in TIB system. Values are the means of 6 replicates ± SD. Different letters indicate significant difference between treatments based on Tukey’s multiple comparisons test (p≤0.05).
Table 2.
Effect of different immersion times and frequencies on shoot multiplication of G. speciosum after 1 month in TIB system. Values are the means of 6 replicates ± SD. Different letters indicate significant difference between treatments based on Tukey’s multiple comparisons test (p≤0.05).
| Treatment | Immersion frequency | No. shoots/explant |
Shoot height (cm) |
Fresh growth index | |
| Time day-1 | Immersion | ||||
| IF1 | 8 | 5 min, every 3 h | 20.00 ± 0.82d | 2.77 ± 0.31bc | 0.23 ± 0.02c |
| IF2 | 4 | 5 min, every 6 h | 38.00 ± 2.94c | 1.93 ± 0.12bc | 2.12 ± 0.63b |
| IF3 | 2 | 5 min, every 12 h | 33.00 ± 3.27cd | 1.10 ± 0.33cd | 2.32 ± 0.50b |
| IF4 | 8 | 10 min, every 3 h | 127.00 ± 2.16a | 5.00 ± 0.51a | 4.26 ± 0.52a |
| IF5 | 4 | 10 min, every 6 h | 95.33 ± 7.59b | 1.27 ± 0.12d | 3.21 ± 0.59ab |
| IF6 | 2 | 10 min, every 12 h | 47.33 ± 5.56c | 3.70 ± 0.45b | 3.05 ± 0.50ab |
Table 3.
Effect of root induction media on morphological characteristic of G. speciosum after 1 month. Values are the means of 10 replicates ± SD. Different letters indicate significant difference between treatments based on Tukey’s multiple comparisons test (p≤0.05). nd means ‘not detected’. * means VW medium supplemented with 150 mL L-1 coconut water, 100 g L-1 banana and 50 g L-1 potato.
Table 3.
Effect of root induction media on morphological characteristic of G. speciosum after 1 month. Values are the means of 10 replicates ± SD. Different letters indicate significant difference between treatments based on Tukey’s multiple comparisons test (p≤0.05). nd means ‘not detected’. * means VW medium supplemented with 150 mL L-1 coconut water, 100 g L-1 banana and 50 g L-1 potato.
| Treatment | Medium |
Hormone (mg L-1) |
No. roots /explant |
Length of root (cm) |
Fresh growth index |
Shoot Height (cm) |
No. Leaves explant-1 |
Length of leaf (cm) |
|
| ½ MS | BAP | NAA | |||||||
| RT1 | ½ MS ½ MS |
1.0 | 0.5 | 1.00 ± 0.00 | 0.40 ± 0.10 b | 1.17 ± 0.50 c | 2.35 ± 0.15 | 4.50 ± 0.50 | 1.25 ± 0.05 |
| RT2 | - | 0.5 | 0.50 ± 0.50b | 0.20 ± 0.20 b | 2.42 ± 0.08 | 3.45 ± 0.35 | 3.50 ± 0.50 | 2.20 ± 0.30 | |
| RT3 | MS | 1.0 | 0.5 | nd | nd | 1.75 ± 0.08 b | 2.65 ± 0.15 | 4.50 ± 0.50 | 0.95 ± 0.15 |
| RT4 | MS | - | 0.5 | nd | nd | 1.83 ± 0.17 b | 2.75 ± 0.25 | 3.50 ± 0.50 | 1.75 ± 0.25 |
| RT5 | VW* | - | - | 2.50 ± 0.50a | 1.35 ± 0.15 a | 3.92 ± 0.25 a | 2.85 ± 0.05 | 4.50 ± 0.50 | 2.25 ± 0.15 |
| RT6 | VW* | - | - | 0.50 ± 0.50b | 0.50 ± 0.50 b | 3.00 ± 0.33 ab | 3.35 ± 0.15 | 3.50 ± 0.50 | 0.95 ± 0.25 |
Table 4.
Antibacterial activity of G. speciosum extracts.
| Bacterium | MIC(mg mL-1) | MBC(mg mL-1) |
| S. aureus | 12.8 | 25.6 |
| S. epidermidis | 25.6 | 51.2 |
| P. acnes | 6.4 | 12.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



