Submitted:
16 July 2025
Posted:
17 July 2025
You are already at the latest version
Abstract
Using a systematic approach, this review investigates whether the adipostatic signalling function of leptin is conserved across the vertebrate taxa (mammals, birds, reptiles and fish), with a focus on seasonally reproducing species. Of the 49 studies analysed, only nine investigated sub-eutherian mammals (monotreme, n = 1; marsupials, n = 0 birds, n = 4; reptiles n = 2 and fish, n = 2); therefore, it was not possible to draw solid conclusions for these taxa. Nevertheless, the evidence collated in this review appears to suggest that an adipostatic function of leptin is absent in avian fish species. Further investigation is required for sub-eutherian and reptilian species as the presented results were inconclusive. Twenty-five species of eutherian mammals were investigated across 40 studies. The adipostatic signalling function of leptin was not observed in all eutherian species, leading to the suggestion of species-specific functionality which may extend to sub-eutherian mammals and reptiles. Further research is necessary across a variety of species of all taxa to confirm whether or not the adipostatic function of leptin is confined to therian mammals.
Keywords:
leptin
; adiposity
; adipostat
; seasonal vertebrate
; systematic review
1. Introduction
Discovered in mice in 1994, the protein hormone, leptin, has been described as a satiety signal, synthesised and secreted by adipose tissue [1]. Leptin has since been identified in a wide range of vertebrate taxa including mammals, amphibians, fish, reptiles [2], and more recently, birds [3]. Further research conducted in eutherian mammals has shown leptin to have significant functional diversity with roles ranging from the regulation of reproduction and immune function to operating as an adipostat (regulating body fat within a narrow range based on energy intake and expenditure; [4]. This adipostatic function of leptin has been observed in a number of eutherian mammals, where an increase in adiposity is generally associated with increased circulating leptin concentrations [2]. Systemic leptin binds to receptors expressed in intracellular compartments of the hypothalamus (e.g., endoplasmic reticulum and endosomes)[5], regulating the production of anorexigenic neuropeptides. These anorexigenic neuropeptides reduce appetite and increase metabolic rate, resulting in a loss of adipose tissue; thus, regulating fat reserves [5].
While this adipostatic function would appear adaptive across species for keeping individuals in optimal body condition, many seasonally reproducing and seasonally hibernating eutherian mammals demonstrate circannual patterns of adiposity, purposely accumulating in large fat depots in preparation for periods of reproductive activity or hibernation [6,7]; clearly an adipostatic function of leptin in these species would be problematic. Consequently, many seasonally reproducing and hibernating eutherian mammals prevent loss of adiposity during these periods in one of two ways: (1) leptin resistance, where circulating leptin concentrations correlate with adiposity but hypothalamic receptors are resistant to the effects of elevated leptin concentrations [6]; or (2) leptin decoupling, whereby circulating leptin concentrations no longer correlate with adiposity [7].
As most studies have focussed on eutherian mammals, it is unclear if the adipostatic function of leptin is also conserved in sub-eutherian mammals or across the different vertebrate taxa. In the only study of the Monotremata [8] investigated the adipostatic function of leptin in the seasonally reproducing short-beaked echidna (Tachyglossus aculeatus) and reported a weak negative relationship between circulating leptin levels and body mass. While the short-beaked echidna lays eggs, they also possess a yolk sac placenta in utero for the transport of nutrients during the early stages of foetal development in a similar manner to viviparous therian mammals [9]. The echidna, therefore, represents an intermediate stage between oviparous reptiles and viviparous therian mammals. Given their unique biology, and noting a similar relationship between leptin and adiposity in a small selection of studies on reptiles [10], birds [11,12]; and fish [13], some authors speculated that the adipostatic function of leptin may be restricted to therian (Metatheria and Eutheria) mammals which separated from the Prototheria approximately 166 million years ago, or eutherian mammals which diverged from the metatherians approximately 148 million years ago [8].
In the 30 years since its discovery, there has been no review that has collated studies reporting the adipostatic function of leptin in mammalian and non-mammalian vertebrates. Using a systematic approach, this review will focus on seasonally reproducing and hibernating vertebrates from a range of taxa, including mammals, birds, reptiles and fish, in order to further investigate the hypothesis proposed by Sprent [8], that the adipostatic signalling function of leptin is confined to therian mammals.
2. Materials and Methods
The search engines used to identify papers were PubMed advanced search, using medical subject headings (MeSH) (1951– January 2021; and Web of Science, Core Collection, advanced search (1990–January 2019). Database specific search terms were created to ensure the database search contained literature relevant to the topic. The reference lists of included studies were also searched as an additional means of identifying appropriate studies. The inclusion criteria for studies identified in the searches were (1) peer-reviewed, (2) written in the English language, (3) empirically conducted research and (4) research which investigated the role of naturally circulating leptin levels in regulating adiposity in seasonally reproducing (where reproductive activity only occurs during certain times of the year) or seasonally hibernating (where species enter a state of inactivity and metabolic depression at certain times of the year), non-domesticated mammals, birds, reptiles or fish.
Titles and abstracts of all identified papers were examined against the pre-defined inclusion criteria. Where an article appeared to meet the inclusion criteria, the full text was obtained and then subjected to a second phase of screening to ensure compliance with the inclusion criteria. Each publication was then analysed for publication date, species data (e.g. Taxa, Order, species name (common and scientific), diet, gender, and whether the species is a long- or short-day breeder, captive or wild, mature or immature and whether or not the species engages in hibernation or migratory behaviour, number of animals included in the study, method of body fat determination and whether this method had been validated, duration of study and finally, a description of how leptin interacts with changes in adiposity in the species studied.
In order to determine if the function of leptin as an adipostat may be confined to specific groups of species, species were subsequently sorted into the appropriate categories including Order, diet, timing of seasonal breeding strategy (long-day or short-day) and whether the species hibernates or migrates.
3. Results
3.1. Database Search
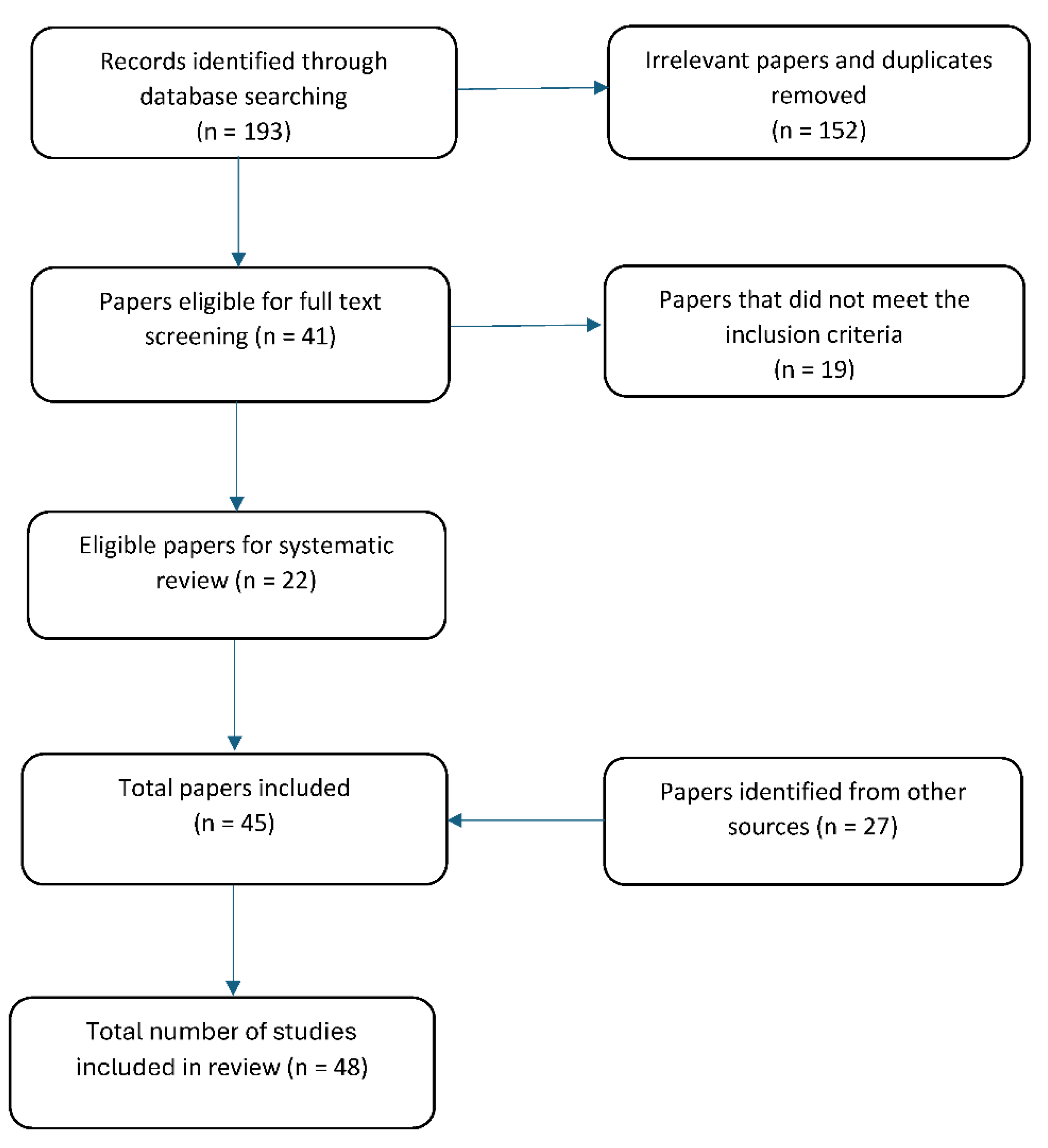
A total of 193 publications were identified from electronic searching. Following the initial screening of titles and abstracts and after removal of duplicates, 41 publications were found to be eligible for full text screening, after which, 22 met the inclusion criteria. Discarded publications (n = 19) assessed exogenous leptin, were based on domesticated species, investigated other aspects of leptin, or the full publication was unable to be retrieved. An additional 27 publications were identified after cross-checking the references of those meeting the inclusion criteria, resulting in a total of 46 publications for review (Figure 1). These 46 publications referred to 48 separate studies as three publications investigated more than one species.
3.2. Leptin and Adiposity in Mammals, Birds, Reptiles and Fish
The identified studies represented all taxa of interest including 25 species of mammals (24 eutherians - placental mammals and 1 prototherian - monotreme mammal), four species of birds, two species of reptiles and two species of fish (Table 1); surprisingly there has yet to be any study that has investigated the relationship between adiposity and circulating leptin levels in metatherian mammals (i.e., Marsupials). Leptin was found to function as an adipostat in 18 species (72%) of eutherians; of these, six (33%) demonstrated leptin decoupling or leptin sensitivity during periods of hibernation or migration. In two separate studies conducted on the American mink (Mustela vison), Nieminen et al. (2000) reported that leptin did not function as an adipostat, whereas Tauson and Forsberg [14] suggested that it does. In the single prototherian species, the short-beaked echidna [8], and in two of the four bird species (European starlings, Sturnus vulgaris, [12] and thin billed prions, Pachyptila belcheri [11], the authors determined that leptin did not function as an adipostat. In the remaining two bird species (Adele penguins, Pygoscelis adeliae, and Bartailed Godwits, Limosa lapponica) [15], while seasonal changes in body mass were observed, leptin was not biochemically detected in either species. In the Eastern fence lizard (Sceloporus undulates), while Spanovich [10] observed that leptin concentrations were lowest when fat stores were at their highest in the pre-hibernation period (suggestive of a decoupling mechanism), data were unavailable to determine the relationship between fat levels and leptin at other times. Paolucci [16] observed leptin concentration in the Italian wall lizard (Podarcis sicula) to be highest during the reproductive quiescent period, however, adiposity was not monitored in the study. In both fish species that were reviewed (Artic Charr, Salvelinus alpinus) [13] and Rainbow trout (Oncorhynchus mykiss) [17], leptin levels were not positively correlated with body mass.
When the eutherian mammal species were categorised based on Order, diet, whether or not the species hibernates, and whether or not the species is a long- or short-day breeder, there was no grouping where leptin functioned as an adipostat in all species of that particular category (Table 2 & Table 3). As the adipostatic function of leptin was not observed in non-eutherian mammals, reptiles, birds and fish, these groups were not included in this categorisation.
4. Discussion
The aim of this systematic review was to determine if the adipostatic function of leptin is conserved across taxa, or if as speculated by Sprent [8], it is restricted to therian or eutherian mammals. This review identified numerous studies investigating eutherian mammals, however, there was a paucity of literature investigating sub-eutherian mammals, birds, reptiles and fish. While the few studies investigating these less represented groups failed to identify a relationship between leptin and adiposity, it is important to consider the limitations of these studies before concluding that no relationship exists. Surprisingly, the adipostatic function of leptin was not observed in all eutherian species investigated, suggesting species-specific functionality. These findings are discussed here, highlighting limitations of the included studies.
4.1. Sub-Eutherian Mammals and Reptiles—Further Research Required
While Sprent [8] did not observe a relationship between leptin and adiposity in the short-beaked echidna, there are two major caveats to consider. First, their study was conducted on the Tasmanian sub-species of short-beaked echidna which demonstrates unique peculiarities compared to the other four sub-species. For example, due to the cooler Tasmanian climate, this sub-species maintains hibernation for extended periods in comparison to those from milder climates (e.g., South-East Queensland and Kangaroo Island populations), so much so, that hibernation and reproductive activity reportedly overlap, a phenomenon not witnessed in the other sub-species [18]. Although a eutherian species, Weitten, Robin [19] has reported that the Siberian hamster showed decreased levels of circulating leptin concentrations during hibernation. Consequently, the extended hibernation period observed in the Tasmanian sub-species of short-beaked echidna could potentially confound the correct interpretation of the relationship between adiposity and leptin in this sub-species, such that extrapolation to the remaining sub-species may be problematic.
Secondly, Sprent [8] did not measure adiposity directly, but instead used body mass as a proxy measure. While body mass is a reasonable alternative given the cost associated with specialised equipment (e.g., duel-energy x-ray absorptiometry (DEXA) scans) or ethical concerns associated with sacrificing study animals to determine their fat composition [20], the literature suggests that body mass does not always reflect body fat levels [21]. For example, Zhao [22] reported that striped hamsters maintained constant body mass between seasons while body fat composition fluctuated. Similarly, Spady [23] showed that body mass and body condition index were not as accurate as leptin itself in monitoring adiposity in the American Black Bear. Consequently, the results reported by Sprent [8] need to be interpreted with caution and further investigation into the adipostatic function of leptin in echidnas and other sub-eutherian mammals is required.
Since the publication of the Sprent [8] study, new evidence has emerged supporting an adipostatic role of leptin in Tachyglossus aculeatus. In a study of captive short-beaked echidnas housed under non-breeding conditions, Dutton-Regester [24] demonstrated a significant positive correlation between circulating leptin levels and directly measured body fat percentage using dual-energy X-ray absorptiometry (DEXA). This represents the first robust evidence that leptin may function as an adipostat in a monotreme species, suggesting that this mechanism could be evolutionarily conserved beyond eutherian mammals. Notably, no sex differences in leptin concentrations were observed despite females having higher body fat, indicating a possible divergence from patterns seen in some therian species. Furthermore, no seasonal variation in leptin levels was detected in the study cohort, though this may reflect the buffered environmental conditions of captivity. These findings warrant further investigation in wild echidna populations and other sub-eutherian taxa to assess seasonal and environmental influences on leptin signalling.
In reptiles, Spanovich, Niewiarowski [10] and Paolucci, Rocco [25] have provided evidence that leptin may function as an adipostat in the Eastern Fence lizard and the Italian Wall lizard, respectively. While there was insufficient information presented to determine the relationship between circulating leptin concentrations and adiposity outside of the hibernation period in the Eastern Fence lizard, Spanovich [10] did observe leptin decoupling during the hibernation period. Therefore, it is quite possible that at other times of the year, leptin may signal adiposity as it does in many hibernating eutherian mammals that demonstrate this phenomenon [26,27]. In the Italian wall lizard, while adiposity was not monitored by Paolucci, Rocco [25], the authors refer to a previous study [28] where fat bodies were largest during the quiescent period of the reproductive cycle, corresponding to the period of high leptin concentration reported by Paolucci, Rocco [25].
4.2. Birds and Fish – Evidence for No Adipostatic Function
Prior to 2014 there had been long-standing contention surrounding the presence of leptin in birds. However, since 2014 avian leptin has been identified in the genomes of various avian species including falcons (Falco peregrinus and Falco cherrug), Tibetan ground tit (Pseudopodoces humilis), zebra finch (Taeniopygia guttata), rock dove (Columba livia), bald eagle (Haliaeetus leucocephalus), downy woodpecker (Picoides pubescens), budgerigar (Melopsittacus undulatus), duck (Anas platyrhynchos) and chicken (Gallus gallus) [3]. These avian leptin sequences have significantly higher guanine-cytosine content compared to other vertebrates, low sequence conservation with mammalian leptin and low level of expression; all of which precluded the identification of avian leptin for so long [3]. Given this lack of conservation, it is not surprising that Yosefi, Hen [15] were unable to detect biologically relevant levels of leptin in the Adele penguin and the Bar-tailed godwit. While Kordonowy [12] and Quillfeldt [11] have reported leptin concentrations for the European starling and the thin-billed prion, respectively, it is likely that these studies were based on reagents that were developed using leptin sequences with more than 95% similarilty to mouse leptin [3]. Consequently, it would be interesting to repeat these studies using the appropriate reagent to ascertain the true relationship, if any, between leptin and adiposity in these species.
Despite these findings, there is also evidence to suggest that leptin may not function as an adipostat in birds. For example, there is high correlation between leptin and leptin receptor transcript in birds, suggesting that leptin may not circulate but rather function as an autocrine or paracrine factor [29]. Further, while leptin is almost exclusively expressed in the adipose tissue of mammals, it is almost undetectable in the adipose tissue of avian species [30,31]. Instead, it is largely expressed in the liver. This hepatic expression is likely associated with the primary role that the liver plays in vitellogenesis in avian species [30].
Similar to birds, there is strong evidence that leptin does not signal adiposity in fish. First, as is the case for birds, the liver is the primary tissue of leptin expression in fish [32]. Second, many observations in fish species run counter to the adipostatic model proposed in mammals. For example, plasma leptin concentrations increase with fasting in salmonids [17] and flounder [33], and even when offered food, Arctic Charr stop feeding when leptin titres fall but eat during periods of rising leptin concentrations. Finally, the ancestral leptin giving rise to leptins in birds, reptiles, and mammals is more closely related to coelacanth and shark (Callorhinchus milii) leptins than leptins from bony fish, i.e., fish leptins appear to have diverged along their own lineage independent of leptins in higher mammals [34].
4.3. Eutherian Mammals – Evidence for Species-Specific Functionality
While leptin is commonly referred to as an adipostat in eutherian mammalian species [2], the results of this systematic review have revealed that this is not a universal phenomenon, with several species including the Arctic Fox, Antarctic Fur seal, Northern elephant seal, Little brown bat, Japanese macaque, Vervet monkey, European beaver, Striped mouse and the Striped hamster all failing to demonstrate a relationship between adiposity and circulating leptin levels. In some cases, the lack of this relationship may be attributed to deficiencies in study design. For example, studies on the Northern elephant seal [35], Vervet monkey [21] and the Striped mouse [36] all failed to use a validated proxy measure for body fat, while the study on the Antarctic fur seal [37] suggested that suckling of young or a post exercise response may have confounded their results. Nevertheless, it is unclear why the adipostatic function of leptin was not observed in the remaining species.
Some authors have suggested that the adipostatic function of leptin is absent in carnivorous species [38,39]. However, this was not evident in this review, as four of the eight carnivorous species identified demonstrated a positive correlation between leptin and adiposity. Additionally, in two separate studies on the mink, while Niemenen [39] found leptin did not signal adiposity, a later study by Tauson [14] contradicted this finding. While the reason for this difference is unclear, Tauson [14] exposed minks to a variable food supply with plasma leptin concentrations mirroring body weight lost and gain as food was restricted or increased, respectively. Further, having previously demonstrated that plasma insulin levels change in response to energy supply, Tauson [14] demonstrated plasma leptin and insulin concentrations followed the same pattern, providing further evidence that leptin signals adiposity in the carnivorous mink.
Irrespective of whether the eutherian mammal species identified in this review were grouped according to Order, diet, long- or short- day breeder, and whether they hibernate or migrate, there was no single group category whereby leptin functioned as an adipostat (or vice versa) in all species. Thus, as non-seasonal mammals have not been included in this review, it is hypothesised that in seasonal eutherian mammals, the adipostatic function of leptin is species-specific. As Zhao [22] explains, there may be species-specific physiological and hormonal control of body mass and adiposity which may be linked to species-specific responses to seasonal changes in temperature, photoperiod or food availability.
5. Conclusions
The objective of this review was to determine if the adipostatic function of leptin was conserved across the vertebrate taxa, with a focus on seasonally reproducing vertebrates. Few studies have investigated sub-eutherian mammals (with a complete lack of research on marsupial mammals), birds, reptiles or fish; therefore, it was not possible to reach a definitive conclusion regards our original question. Nevertheless, current evidence would suggest that the adipostatic function of leptin is absent in avian and fish species. More research is necessary before conclusions can be drawn for sub-eutherian and reptilian species as the presented results were inconclusive. Surprisingly, the adipostatic function of leptin was also not observed in all eutherian species investigated, leading to a conclusion of species-specific functionality which we suggest may extend to sub-eutherian mammals and reptiles.
Funding
This research received no external funding.
Data Availability Statement
Dataset available on request from the authors
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| DEXA | Duel-Energy X-ray Absorptiometry |
References
- Zhang, Y.M. , et al., Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Denver, R.J., R. M. Bonett, and G.C. Boorse, Evolution of leptin structure and function. Neuroendocrinology 2011, 94, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Prokop, J.W. , et al., Discovery of the elusive leptin in birds: identification of several 'missing links' in the evolution of leptin and its receptor. PLoS One 2014, 9, e92751. [Google Scholar] [CrossRef] [PubMed]
- Friedman-Einat, M. , et al., Discovery and characterization of the first genuine avian leptin gene in the rock dove (Columba livia). Endocrinology 2014, 155, 3376–84. [Google Scholar] [CrossRef] [PubMed]
- Wauman, J., L. Zabeau, and J. Tavernier, The Leptin Receptor Complex: Heavier Than Expected? Front Endocrinol (Lausanne) 2017, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Rosseau, K. , et al., Photoperiodic Regulation of Leptin Resistance in the Seasonally Breeding Siberian Hamster (Phodopus sungorus). Endocrinology 2002, 148, 3083–95. [Google Scholar] [CrossRef] [PubMed]
- Townsend, K.L., T. H. Kunz, and E.P. Widmaier, Changes in body mass, serum leptin, and mRNA levels of leptin receptor isoforms during the premigratory period in Myotis lucifugus. J Comp Physiol B 2008, 178, 217–23. [Google Scholar] [CrossRef] [PubMed]
- Sprent, J., S. M. Jones, and S.C. Nicol, Does leptin signal adiposity in the egg-laying mammal, Tachyglossus aculeatus? Gen Comp Endocrinol 2012, 178, 372–9. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, M. , The Biology of Monotremes, ed. A. Press. 1978, New York.
- Spanovich, S., P. H. Niewiarowski, and R.L. Londraville, Seasonal effects on circulating leptin in the lizard Sceloporus undulatus from two populations. Comp Biochem Physiol B Biochem Mol Biol 2006, 143, 507–13. [Google Scholar] [CrossRef] [PubMed]
- Quillfeldt, P. , et al., Relationship between plasma leptin-like protein levels, begging and provisioning in nestling thin-billed prions Pachyptila belcheri. Gen Comp Endocrinol 2009, 161, 171–8. [Google Scholar] [CrossRef] [PubMed]
- Kordonowy, L.L., J. P. McMurtry, and T.D. Williams, Variation in plasma leptin-like immunoreactivity in free-living European starlings (Sturnus vulgaris). Gen Comp Endocrinol 2010, 166, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Froiland, E. , et al., Seasonal appetite regulation in the anadromous Arctic charr: evidence for a role of adiposity in the regulation of appetite but not for leptin in signalling adiposity. Gen Comp Endocrinol 2012, 178, 330–7. [Google Scholar] [CrossRef] [PubMed]
- Tauson, A.H. and M. Forsberg, Body-weight changes are clearly reflected in plasma concentrations of leptin in female mink (Mustela vison). Br J Nutr 2002, 87, 101–5. [Google Scholar] [CrossRef] [PubMed]
- Yosefi, S. , et al., Lack of leptin activity in blood samples of Adelie penguin and bar-tailed godwit. J Endocrinol 2010, 207, 113–22. [Google Scholar] [CrossRef] [PubMed]
- Paolucci, M., M. Rocco, and E. Varricchio, Leptin presence in plasma, liver and fat bodies in the lizard Podarcis sicula- fluctuations throughout the reproductive cycle. Life Sciences 2001, 143, 3083–95. [Google Scholar]
- Salmeron, C. , et al., Effects of nutritional status on plasma leptin levels and in vitro regulation of adipocyte leptin expression and secretion in rainbow trout. Gen Comp Endocrinol 2015, 210, 114–23. [Google Scholar] [CrossRef] [PubMed]
- Morrow, G.E., S. M. Jones, and S.C. Nicol, Frozen embryos? Torpor during pregnancy in the Tasmanian short-beaked echidna Tachyglossus aculeatus setosus. General and Comparative Endocrinology 2017, 244, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Weitten, M. , et al., Hormonal changes and energy substrate availability during the hibernation cycle of Syrian hamsters. Horm Behav 2013, 64, 611–7. [Google Scholar] [CrossRef] [PubMed]
- LeeHong, P.A. , et al., Dual-energy X-ray absorptiometry (DXA) and chemical composition as measures of body composition of the short-beaked echidna (Tachyglossus aculeatus aculeatus). Australian Journal of Zoology 2019, 67. [Google Scholar] [CrossRef]
- Garcia, C. , et al., Energetic consequences of seasonal breeding in female Japanese macaques (Macaca fuscata). Am J Phys Anthropol 2011, 146, 161–70. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.J. , et al., Seasonal changes of body mass and energy budget in striped hamsters: the role of leptin. Physiol Biochem Zool 2014, 87, 245–56. [Google Scholar] [CrossRef] [PubMed]
- Spady, T.J. , et al., Leptin as a surrogate indicator of body fat in the American black bear. Ursus 2009, 20, 120–130. [Google Scholar] [CrossRef]
- Dutton-Regester, K.J. , et al., Body fat and circulating leptin levels in the captive short-beaked echidna (Tachyglossus aculeatus). Journal of Comparative Physiology B 2024, 194, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Paolucci, M., M. Rocco, and E. Vachio, Leptin presence in plasma, liver and fat bodies in the lizard Podarcis sicula- fluctuations throughout the reproductive cycle. Life Sciences 2001, 69, 2399–2408. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, P. , et al., Endocrine response to fasting in the overwintering captive raccoon dog (Nyctereutes procyonoides). J Exp Zool A Comp Exp Biol 2004, 301, 919–29. [Google Scholar] [CrossRef] [PubMed]
- Xing, X., M. Yang, and D.H. Wang, The expression of leptin, hypothalamic neuropeptides and UCP1 before, during and after fattening in the Daurian ground squirrel (Spermophilus dauricus). Comp Biochem Physiol A Mol Integr Physiol 2015, 184, 105–12. [Google Scholar] [CrossRef] [PubMed]
- Filosa, S. , Biological and cytological aspects of the ovarian cycle in Lacerta s.sicula. Monitore Zoologico Italiano 1973, 7, 151–165. [Google Scholar]
- Seroussi, E. , et al., Identification of the long-sought leptin in chicken and duck: expression pattern of the highly GC-rich avian leptin fits an autocrine/Paracrine rather than endocrine function. Endocrinology 2016, 157, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Taouis, M. , et al., Cloning the chicken leptin gene. Gene 1998, 208, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Ashwell, C.M. , et al., Hormonal regulation of leptin expression in broiler chickens. Am. J. Physiol. Integr. Comp. Physiol. 1999, 276, R226–R232. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, T., S. Uji, and T. Suzuki, Identification of cDNA coding for a homologue to mammalian leptin from pufferfish, Takifugu rubripes. Peptides 2005, 26, 45–750. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, E.N. , et al., Plasma leptin and growth hormone levels in the fine flounder (Paralichthys adspersus) increase gradually during fasting and decline rapidly after refeeding. General and Comparative Endocrinology 2012, 177, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Londraville, R.L. , et al., Comparative endocrinology of leptin: assessing function in a phylogenetic context. Gen Comp Endocrinol 2014, 203, 146–57. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, R.M., C. E. Wade, and C.L. Ortiz, Effects of prolonged fasting on plasma cortisol and TH in postweaned northern elephant seal pups. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 2001, 280, R790–5. [Google Scholar] [CrossRef] [PubMed]
- Schradin, C. , et al., Leptin levels in free ranging striped mice (Rhabdomys pumilio) increase when food decreases: the ecological leptin hypothesis. Gen Comp Endocrinol 2014, 206, 139–45. [Google Scholar] [CrossRef] [PubMed]
- Arnould, J.P. , et al., Variation in plasma leptin levels in response to fasting in Antarctic fur seals (Arctocephalus gazella). J Comp Physiol B 2002, 172, 27–34. [Google Scholar] [PubMed]
- Ahima, R.S. and J.S. Flier, Leptin. Review Annu Rev Physiol 2000, 62, 413–37. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, N. , et al., Plasma leptin and thyroxine of mink (Mustela vison) vary with gender, diet and subchronic exposure to PCBs. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 2000, 127, 515–522. [Google Scholar]

Table 1.
Data extracted from each study including species taxonomic information, number of animals included in the study and sex, whether or not there was evidence that leptin functions as an adipostat, whether there was evidence for leptin decoupling or sensitivity, the method of fat determination used and whether this was validated (if necessary), and study duration.
Table 1.
Data extracted from each study including species taxonomic information, number of animals included in the study and sex, whether or not there was evidence that leptin functions as an adipostat, whether there was evidence for leptin decoupling or sensitivity, the method of fat determination used and whether this was validated (if necessary), and study duration.
| Author (year) | Taxa | Order | Species (Scientific name) | No. of animals | Sex | Evidence that leptin indicates adiposity? | Evidence of leptin decoupling or sensitivity? | Fat determination method (validated? Y/N) | Study duration | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soppela et al. (2008) | Mammal (eutherian) | Artiodactyla | Reindeer (Rangifer tarandus) | 16 | M | Yes | Yes | Body mass (N) | 4.5 months | ||||
| (Spady et al., 2009) | Mammal (eutherian) | Carnivora | American Black Bear (Ursus americanus) | 20 | M & F | Yes | No | Body fat % (NA) | 6 months | ||||
| Fuglei et al. (2004) | Mammal (Placental) | Carnivora | Arctic fox (Alopex lagopus) | 8 | M | No | No | Body mass (N) | 6 months | ||||
| Arnould et al. (2002) | Mammal (Placental) | Carnivora | Antarctic fur seal (Arctocephalus gazelle) | 28 | M & F | No | No | Body mass (Y) | 3-5 days | ||||
| (Nieminen et al., 2001) | Mammal (Placental) | Carnivora | Blue fox (Vulpes lagopus) | 11 | M & F | Yes | Yes | BMI (N) | 6 months | ||||
| Mustonen et al. (2005) | Mammal (Placental) | Carnivora | Blue fox (Vulpes lagopus) | 48 | M & F | Yes | Yes | BMI (Y) | 12 months | ||||
| Hissa et al. (1998) | Mammal (Placental) | Carnivora | European Brown bear (Ursus arctos arctos) | 6 | M & F | Yes | No | Fat reserves (NA) | 12 months | ||||
| Tauson et al. (2002) | Mammal (Placental) | Carnivora | Mink (Neovison vison) | 6 | F | Yes | No | Body mass (N) | 2 months | ||||
| Nieminen et al. (2000) | Mammal (eutherian) | Carnivora | Mink (Neovison vison) | 53 | F | No | No | BMI (N) | 5 months | ||||
| (Ortiz, Noren, et al., 2001) | Mammal (Placental) | Carnivora | Northern Elephant Seal (Mirounga angustirostris) | 40 | M & F | No | No | Fat mass (NA) | Unclear | ||||
| (Ortiz, Wade, et al., 2001) | Mammal (Placental) | Carnivora | Northern Elephant Seal (Mirounga angustirostris) | 15 | M & F | No | No | Body mass (N) | 7 weeks | ||||
| Kitao et al. (2011) | Mammal (Placental) | Carnivora | Racoon dog (Nyctereutes procyonoides) |
9 | M & F | Yes | Yes | Body fat % (NA) | 8 months | ||||
| Nieminen et al. (2004) | Mammal (Placental) | Carnivora | Racoon dog (Nyctereutes procyonoides) | 11 | M & F | Yes | Yes | BMI (N) | 6 months | ||||
| Nieminen et al. (2002) | Mammal (Placental) | Carnivora | Racoon dog (Nyctereutes procyonoides) | 33 | M & F | Yes | Yes | Body mass (Y) | 6 months | ||||
| Srivastava and Krishna (2008) | Mammal (Placental) | Chiroptera | Greater Asiatic yellow bat (Scotophilus heathi) | 120 | F | Yes | No | Fat Content & body mass (NA) | 12 months | ||||
| Banerjee et al. (2011) | Mammal (Placental) | Chiroptera | Indian short-nosed fruit bat (Cynopterus sphinx) | 72 | F | Yes | No | Body fat content & body mass (NA) | 12 months | ||||
| Banerjee et al. (2010) | Mammal (Placental) | Chiroptera | Indian short-nosed fruit bat (Cynopterus sphinx) | 76 | F | Yes | No | Body fat content & body mass (NA) | 10 months | ||||
| Widmaier et al. (1997) | Mammal (Placental) | Chiroptera | Little Brown Bat (Myotis lucifugus) | Unclear | F | No | No | Fat index (fat mass/lean dry mass) (NA) | 1 month | ||||
| Kronfeld-Schor et al. (2000) | Mammal (Placental) | Chiroptera | Little Brown Bat (Myotis lucifugus) | Unclear | F | Unclear | Yes | Body fat & body mass (NA) | 2 months | ||||
| Roy and Krishna (2010) | Mammal (Placental) | Chiroptera | Greater asiatic yellow bat (Scotophilus heathi) | Unclear | M | Yes | No | Body fat content & body mass (NA) | 12 months | ||||
| Srivastava and Krishna (2007) | Mammal (Placental) | Chiroptera | Greater asiatic yellow bat (Scotophilus heathi) | Unclear | F | Yes | No | Body fat content & body mass (NA) | 6 months | ||||
| Wang et al. (2006c) | Mammal (Placental) | Lagomorphs | Pikas (Ochotona curzoniae) | 40 | M & F | Yes | NR | Body fat content & body mass (NA) | 9 months | ||||
| Garcia et al. (2011) | Mammal (Placental) | Primate | Japanese macaques (Macaca fuscata) | 14 | F | No | No | BMI (N) | 2 months | ||||
| Whitten and Turner (2008) | Mammal (Placental) | Primate | Vervet Monkeys (Chlorocebus pygerythrus) | 116 | M & F | No | No | BMI (Y) | 12 months | ||||
| Li and Wang (2005) | Mammal (Placental) | Rodent | Brandt's Voles Microtis Brandti | Unclear | Unclear | Yes | No | Body fat content & body mass (NA) | 5 months | ||||
| Zhang and Wang (2006) | Mammal (Placental) | Rodent | Brandts voles (Microtis Brandti) | 16 | M & F | Yes | No | Body fat content & body mass (NA) | 4 weeks | ||||
| Zhang and Wang (2006) | Mammal (Placental) | Rodent | Brandts voles (Microtis Brandti) | 50 | M | Yes | No | Body fat content & body mass (NA) | 8 weeks | ||||
| Xing et al. (2015) | Mammal (Placental) | Rodent | Daurian Ground Squirrel (Spermophilus dauricus) | Unclear | F | Yes | Yes | Body fat content & body mass (NA) | 8 months | ||||
| Chen et al. (2012) | Mammal (Placental) | Rodent | Maximowiczi’s voles (Microtus maximowiczii) | Unclear | Unclear | Yes | No | Body fat content & body mass (NA) | 8 months | ||||
| Zhang and Wang (2007) | Mammal (Placental) | Rodent | Mongolian gerbils (Meriones unguiculatus) | 75 | M & F | Yes | Yes | Body fat content & body mass (NA) | 12 months | ||||
| Wang et al. (2006b) | Mammal (Placental) | Rodent | Root voles (Microtus oeconomu) | 10 | M & F | Yes | No | Body fat content & body mass (NA) | 10 months | ||||
| Wang et al. (2006a) | Mammal (Placental) | Rodent | Root voles (Microtus oeconomu) | 20 | M & F | Yes | No | Body fat content & body mass (NA) | 4 weeks | ||||
| Schradin et al. (2014) | Mammal (Placental) | Rodent | Striped mice (Rhabdomys pumilio) | Unclear | M & F | No | Yes | Body mass (N) | 12 months | ||||
| Zhao (2011) | Mammal (Placental) | Rodent | Striped hampsters (Cricetulus barabensis) | 64 | Unclear | No | No | Body fat content & body mass (NA) | 3 months | ||||
| Zhao et al. (2014) | Mammal (Placental) | Rodent | Striped hampsters (Cricetulus barabensis) | 52 | M | No | No | Body fat | 3 months | ||||
| Florant et al. (2004) | Mammal (Placental) | Rodent | Yellow bellied marmot (Marmota flaviventris) | 7 | M & F | Yes | Yes | Body fat & body mass (NA) | 12 months | ||||
| Sprent et al. (2012) | Mammal (prototherian) | Monotremata | Short beaked echidna (Tachyglossus aculeatus) | 34 | M & F | No | Yes | Lean body mass (N) | 36 months | ||||
| Yosefi et al. (2010) | Avian | Charadriiformes | Bar-tailed Godwit (Limosa lapponica) | Unclear | M & F | No | NA | Correlation of body fat mass, body mass & wing length (Y) | On arrival at landing site | ||||
| Yosefi et al. (2010) | Avian | Sphenisciformes | Adele penguin (Pygoscelis adeliae) | Unclear | M & F | No | NA | Isotope dilution approach (NA) | Pre-incubation and 45 days post egg laying | ||||
| Kordonowy et al. (2010) | Avian | Passiferormes | European starling (Sturnus vulgaris) | 57 | F | No | No | Body fat & body mass (NA | 4 months | ||||
| Quillfeldt et al. (2009) | Avian | Procellariiformes | Thin billed prion (Pachyptila belcheri) | Unclear | Unclear | No | No | Body condition (N) | 5 months | ||||
| Spanovich et al. (2006) | Reptile | Squamata | Eastern fence lizard (Sceloporus undulatus) | 60-180 | M & F | Unclear | Yes | Fat Stores (NA) | 12 months | ||||
| Paolucci et al. (2001) | Reptile | Squamata | Italian wall lizard (Podarcis sicula) | Unclear | F | Unclear | NA | Fat body mass (NA) | 9 months | ||||
| Froiland et al. (2012) | Fish | Salmoniformes | Arctic charr (Salvelinus alpinus) | 230 | M & F | No | No | Body fat & body mass (NA | 9 months | ||||
| (Salmeron et al., 2015) | Fish | Salmoniformes | Rainbow trout (Oncorhynchus mykiss) | 92 | Unclear | No | No | Adipose Tissue mass (NA) | 8 WEEKS | ||||
Table 2.
Species specific data collected from each study including species diet category, whether wild or captive, mature or immature, long- or short- day breeders and whether the species hibernates or migrates.
Table 2.
Species specific data collected from each study including species diet category, whether wild or captive, mature or immature, long- or short- day breeders and whether the species hibernates or migrates.
|
Author (year) |
Species | Diet | Mature or immature | Long- or short-day breeder | Hibernate or migrate |
|---|---|---|---|---|---|
| Soppela et al. (2008) | Reindeer | Herbivore | Immature | Long | No |
| Spady et al. (2009) | American Black Bear | Omnivore | Mature | Long | Yes |
| Fuglei et al. (2004) | Arctic fox | Carnivore | Mature | Long | No |
| Arnould et al. (2002) | Antarctic fur seal | Carnivore | Both | Long | No |
| Nieminen et al. (2001) | Blue fox | Omnivore | Mature | Short | No |
| Mustonen et al. (2005) | Blue fox | Omnivore | Mature | Short | No |
| Hissa et al. (1998) | European Brown bear | Omnivore | Mature | Long | Yes |
| Tauson et al. (2002) | Mink | Carnivore | Mature | Short | No |
| Nieminen et al. (2000) | Mink | Carnivore | Mature | Short | No |
| Ortiz et al. (2001) | Northern Elephant Seal | Carnivore | Immature | Short | No |
| Kitao et al. (2011) | Racoon dog | Omnivore | Mature | Long | Yes |
| Nieminen et al. (2004) | Racoon dog | Omnivore | Immature | Long | Yes |
| Nieminen et al. (2002) | Racoon dog | Omnivore | Immature | Long | Yes |
| Srivastava et al. (2008) | Greater Asiatic yellow bat | Insectivorous | Mature | Long | Yes |
| Banerjee et al. (2011) | Indian short-nosed fruit bat | Frugivore | Mature | Long | Yes |
| Banerjee et al. (2010) | Indian short-nosed fruit bat | Frugivore | Mature | Long | Yes |
| Widmaier et al. (1997) | Little Brown Bat | Insectivorous | Mature | Long | Yes |
| Townsend et al. (2008) | Little Brown Bat | Insectivorous | Mature | Long | Yes |
| Kronfeld-Schor et al. (2000) | Little Brown Bat | Insectivore | Mature | Long | Yes |
| Roy et al. (2010) | Greater asiatic yellow bat | Insectivorous | Mature | Long | Yes |
| Srivastava et al. (2007) | Greater asiatic yellow bat | Insectivorous | Mature | Long | Yes |
| Wang et al. (2006) | Pikas | Herbivore | Mature | Long | No |
| Garcia et al. (2011) | Japanese macaques | Omnivore | Mature | Short | No |
| Garcia et al. (2010) | Japanese macaques | Omnivore | Mature | Short | No |
| Whitten et al. (2008) | Vervet Monkeys | Herbivore | Mature | Long | No |
| Li et al. (2005) | Brandt's Voles | Omnivore | Unclear | Long | No |
| Zhang et al. (2006) | Brandts voles | Omnivore | Mature | Long | No |
| Xing et al. (2015) | Daurian Ground Squirrel | Herbivore | Mature | Long | Yes |
| Chen et al. (2012) | Maximowiczi’s voles | Herbivore | Mature | Long | No |
| Zhang et al. (2007) | Mongolian gerbils | Herbivore | Mature | Long | No |
| Wang et al. (2006) | Root voles | Omnivore | Mature | Long | No |
| Wang et al. (2006) | Root voles | Omnivore | Mature | Long | No |
| Schradin, et al. (2014) | Striped mice | Omnivore | Mature | Long | No |
| Zhao et al. (2011) | Striped hamsters | Omnivore | Mature | Long | Yes |
| Zhao et al. (2014) | Striped hamsters | Omnivore | Mature | Long | Yes |
| Florant et al. (2004) | Yellow bellied marmot | Herbivore | Mature | Long | Yes |
| Sprent et al. (2012) | Short beaked echidna | Insectivorous | Mature | Short | Yes |
| Yosefi et al. (2010) | Bar-tailed Godwit | Omnivore | Mature | Unclear | Yes |
| Yosefi et al. (2010) | Adele penguin | Carnivore | Mature | Long | Yes |
| Kordonowy et al. (2010) | European starlings | Omnivore | Immature | Long | Yes |
| Quillfeldt et al. (2009) | Thin billed prions | Carnivore | Immature | Long | Yes |
| Spanovich, et al. (2006) | Eastern fence lizard | Insectivorous | Mature | Long | Yes |
| Paolucci et al. (2001) | Italian wall lizard | Insectivorous | Mature | Long | Yes |
| Froiland et al. (2012) | Arctic charr | Insectivore | Immature | Long | No |
| Salmeron, et al. (2015) | Rainbow trout | Carnivore | Mature | Long | No |
Table 3.
Eutherian mammal species identified in the literature search categorised by Order, diet, whether or not the species hibernates and whether or not the species is a long- or short-day breeder.
Table 3.
Eutherian mammal species identified in the literature search categorised by Order, diet, whether or not the species hibernates and whether or not the species is a long- or short-day breeder.
| Does leptin signal adiposity? | ||||
|---|---|---|---|---|
| Category | No. species in group | Yes (%) | No (%) | Unclear (%) |
| Order | ||||
| Carnivora | 8 | 4 (50) | 3 (37) | 1 (12) |
| Chiprotera | 4 | 3 (75) | 1 (25) | 0 |
| Rodent | 8 | 5 (63) | 3 (37) | 0 |
| Primate | 4 | 2 (50) | 2 (50) | 0 |
| Diet | ||||
| Herbivore | 6 | 4 (67) | 2 (33) | 0 |
| Insectivore | 2 | 1 (50) | 1 (50) | 0 |
| Carnivore | 4 | 0 | 3 (75) | 1 (25) |
| Omnivore | 9 | 6 (67) | 3 (33) | 0 |
| Hibernating species | 9 | 7 (78) | 2 (22) | 0 |
| Timing of breeding season | ||||
| Long-day | 19 | 13 (68) | 6 (32) | 0 |
| Short-day | 5 | 1 (20) | 3 (60) | 1(20) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



