Submitted:
12 July 2025
Posted:
15 July 2025
You are already at the latest version
Abstract
Entomogenous fungi (EF) are ubiquitous in nature and have demonstrated significant potential for biological pest control and the regulation of ecosystem functions. In recent years, significant progress has been made in research on the role and function of EF in ecosystems. They play a vital role in regulating agricultural pest populations, promoting plant growth, and maintaining ecological balance. This article provides a comprehensive review of the role and significance of EF within ecosystems. Firstly, the biodiversity of EF and the factors affecting the diversity of EF were summarized. The article then systematically examines the intricate interactions between EF and both animals and plants. By taking advantage of the characteristics of EF, they can not only be used for biological pest control but also colonize plants, promote plant growth, and suppress agricultural pests. Additionally, this review explores the ecosystem services provided by EF, including their participation in mycorrhizal networks and their contribution to the nitrogen cycle in soil. In conclusion, this review highlights the immense potential of EF in pest management and ecosystem function regulation, underscores the importance of ecosystem services, and identifies future scientific challenges and opportunities, aiming to inspire further research and practical applications.
Keywords:
entomogenous fungi
; biodiversity
; biological control
; hyphae network
; soil nitrogen cycle
; promote plant growth
1. Introduction
Entomogenous Fungi (EF) are a group of fungi that colonize insects (or other arthropods) and proliferate in them to cause disease or death [1]. EF play a crucial role in biological control, representing over 60% of insect pathogens, making them the largest group of insect pathogenic microorganisms [2]. The definition of EF has a broad sense and a narrow sense. In the narrow sense, EF refers specifically to fungi that infect and multiply within insects or produce toxins that ultimately kill them [3]. In contrast, the generalized definition of EF has more connotations. The generalized EF refers to a class of fungi that can not only parasitize insects, or have other nutritional relationships with insects (such as symbiosis, saprophytic, competition, etc.), but also engage in various nutritional relationships with them (such as spiders, horses, snails, etc.) [4]. Overall, EF is a special class of soil microbes that infect and kill insects and other arthropods by penetrating their exoskeletons, are currently used as biological control agents against plant pests, and play an important role in pest management [5]. About 1000 species of EF have been reported in more than 100 genera [6], among them, Beauveria, Metarhizium, Paecilomyces, Verticillium, Entomophthora and Erynia are common and useful genera, has been well used in the biological control of a variety of insects [7]. However, there are still more new species of entomopathogenic fungi to be discovered [8]. At present, researchers pay more attention to the use of EF to control pests in agriculture, forestry and grass industry. However, comprehensive studies on other roles of EF in ecosystems are still lacking and urgently needed to guide policy and future research activities.
Here, we focus on the role of EF in ecosystems for four main reasons: 1) In recent years, studies on EF diversity mainly focus on ecosystems with rich vegetation species such as forests and nature reserves [9], but there are few studies on EF diversity in different habitats. In fact, the study on the diversity, community structure, ecological distribution, biological characteristics and host interaction of EF in different habitats can provide scientific basis for the rational protection and sustainable utilization of EF resources [10]. 2) EF can provide carbon-nitrogen elements and integrate them into the fungal ecological network system in the soil, which may have a significant impact on biogeochemical cycles [11]. 3) After nitrogen is transferred to the plant, it will be dissolved with plant secretions and dead individuals (including litter). Up to 31% of plant nitrogen in an ecosystem can be lost through herbivorous insects [12]. The plant, in turn, can regain nitrogen previously lost to herbivorous insects through association with the endophytic EF [13]. Then, nitrogen is reduced to the soil through microbial mineralization [14], which will definitely affect the soil nitrogen cycle. Here, we synthesize current knowledge on the importance of EF in natural ecosystems to provide theoretical support for the full exploitation and utilization of EF as a valuable resource in the future.
2. The Diversity of EF and Its Influencing Factors
2.1. Classification of EF
EF are a group of fungi that are specialized in parasitism and death of insects or other arthropods, and can be divided into two categories: cordyceps fungi (CF) and entomophthoralean fungi (EPF) [15].
CF are a group of parasitic fungi that infect insects and other arthropods. Based on their functional roles, these fungi can be categorized into medicinal and edible cordyceps fungi, as well as biocontrol fungi for managing agricultural pests [16]. Morphologically, medicinal and edible cordyceps fungi produce fruiting bodies and release ascospores through ascocarps [17]. In contrast, biocontrol fungi do not form fruiting bodies, instead, they exhibit white or green mycelia with conidia arranged in chains [18], the specific differences between the two are detailed in Table 1. Currently, known CF are classified into three families and over 20 genera [19]. Research indicates that these fungi can parasitize invertebrates, plants, and certain fungi, coordinating with their hosts' life cycles to ensure survival and reproduction [20]. Notable examples include B. bassiana and M. anisopliae as important biocontrol fungi, while Cordyceps sinensis, C. militaris, and C. chanhua possess significant medicinal and edible value. The Metarhizium sp. is a major group within CF, comprising nearly 50 species [21], which parasitize over 200 insect species [7,22]. Specifically, M. anisopliae and M. acridum infect insects across 12 orders and 33 families [7]. In nature, CF often exist as insect-fungus complexes, utilizing the insect body as a nutrient source. They continuously infect and reproduce, gradually converting host tissues into sclerotia, ultimately forming dead insects (Figure 1) [23]. CF are unique EF with both asexual and sexual life stages, each capable of independent growth and spore production [24]. The diversity of hosts has complicated the identification of EF, particularly in distinguishing the asexual forms of species like C. sinensis, leading to long-standing challenges in classification [25,26]. However, advancements in molecular systematics have significantly enhanced the classification of CF. Through the analysis of ribosomal DNA (rDNA) and protein-coding genes, researchers have achieved more accurate classifications. For instance, Sung et al. [27] reclassified the Cordyceps sp. into several independent families, including Cordycipitaceae and Hypocreaceae. Additionally, Kepler et al. [28] used multi-gene phylogenetic analysis to further clarify the taxonomic status of multiple CF species. Recent research has led to the discovery and description of numerous new CF species. Mongkolsamrit et al. [29] identified several new species in Thailand, including novel CF species associated with ants. These discoveries not only expand the know diversity of CF but also highlight their ecological niche differentiation across various ecosystems. Future taxonomic research on CF will continue to benefit from advances in molecular techniques, particularly whole-genome sequencing and multi-gene phylogenetic analysis. These methods will aid in elucidating the evolutionary relationships and species boundaries of CF. Integrating morphological, ecological, and geographical distribution studies will provide a more comprehensive framework for their classification.
Entomophthoralean Fungi (EPF) is an obligate pathogenic fungus. Up to now, more than 300 species of EPF have been discovered, belonging to 4 families and 17 genera [3]. EPF have a narrow host range and use arthropods from groups such as Homoptera and Coleoptera as hosts [63]. Most EPF only target specific pests [64], which is difficult to meet the requirements of integrated pest management. The application scope of biocontrol in integrated pest management can be expanded by optimizing the combination of strains and application strategies [64]. In addition, the CRISPR-Cas9 technology can be utilized to precisely modify virulence genes, and develop broad-spectrum and stress-resistant engineered strains to address this challenge [65,66]. In addition to infecting insects, some strains of EPF can also infect invertebrates such as nematodes [67]. Most EPF infect their hosts by actively ejecting conidia. The infection process begins when the conidia germinate on the insect's surface and penetrate the cuticle [68]. Subsequently, the EPF utilize the nutrients within the host for proliferation [69,70,71]. EPF have high virulence, with just a few spores being sufficient to infect the host and cause its death [7]. Additionally, the conidia of EPF can all produce secondary conidia, and some species can also form chlamydospores to enter a dormant state, which is conducive to wide spread and transmission [72]. Among them, Pandora neoaphidis is a highly representative obligate EPF that often triggers epidemic aphid diseases in agriculture, forestry, and grassland [73]. The Penicillium sp. is a cosmopolitan saprophytic fungus that can also parasitize various substrates and plays a significant ecological role in nature by participating in the decomposition process [74]. Soil contains a large number of Penicillium spores or hyphae, which then participate in the life history of insects and constitute a major group of EPF [75]. It is worth noting that during the application of EPF in the field for biological control, problems such as low survival rate of conidia, short shelf life, high cost of large-scale production, and insufficient stability of existing formulations (such as suspensions) have been discovered. Future research can address these challenges from the following aspects: 1) Developing nano-encapsulation technology or microcapsule formulations to enhance the stability and shelf life of conidia; 2) Developing low-cost culture media (such as fermentation of agricultural waste) to reduce production costs; 3) Exploring compound formulations (such as entomopathogenic fungi + plant essential oils) to enhance the synergistic effect; 4) Utilizing CRISPR-Cas9 technology to directionally modify virulence genes and develop broad-spectrum, stress-resistant engineered strains; 5) Constructing heterologous expression systems to mass-produce highly active toxins.
2.2. Influencing Factors of EF Biodiversity
EF are widely distributed and are regulated by multiple factors in different ecological environments (Figure 2) [22,76,77,78,79], while the community structure was significantly different [69]. In addition to its characteristics, the formation of EF’s biodiversity is closely related to the microenvironment in which EF is located [7]. It is important to study the effects of external environmental conditions on EF to improve their adaptability and production practices [80]. Many factors, such as climate, geographical location, vegetation type, soil, biological and human activities, affect the growth of EF and the successful infection of insects [77]. Soil serves not only as the primary habitat for insects but also as a critical reservoir for EF [81]. Research has demonstrated that the growth and reproduction of these fungi in soil are closely linked to various environmental factors within the soil matrix [82]. Key determinants influencing the structure and diversity of soil fungal communities include nutrient availability, moisture content, pH levels, plant diversity, and temperature [83,84,85,86]. In nature, the resources of EF are extremely rich and widely distributed [77]. Traces of EF have been found from primitive habitats to artificial habitats [87,88]. The existence of EF has been found in soil types such as forests, wastelands, orchards, farmlands and grasslands in different regions [87,88,89]. Notably, forest habitats exhibit higher species diversity and evenness of EF [90,91]. Previous studies have shown that EF can be isolated from a variety of habitats, such as alpine lakes, primary forests, plateau permafrost, and so on [92]. The diversity of these habitats offers a wide range of host options and complex ecological niches for EF, thereby facilitating the diversification of their species and functions [6,93]. In general, areas with high vegetation coverage, high humidity, diverse insect community composition, and little human disturbance were more conducive to EF growth [94,95]. In conclusion, the distribution characteristics of EF in different soil types are different, and these characteristics are important for understanding the ecological functions and biological control potential of EF.
2.2.1. Effect of Temperature and Humidity on the EF
Most EF have specific requirements for the temperature of their survival environment, as temperature can directly affect their growth and development. Both excessively high and low temperatures can directly influence the germination of EF spores, mycelial growth, and sporulation [78]. For instance, when spores attach to the host's surface, if the temperature is not suitable, the spores cannot germinate and penetrate the host's cuticle [96]. Agbessenou et al. [97] found that over 90% of spores germinated at 20 °C, 25 °C, and 30 °C, but only 35% to 50% germinated at 35 °C. At 15 °C, no spore germination was recorded after 18 h of incubation, but a low germination rate was observed after a longer period (delayed germination), which subsequently translated into low mycelial growth and low sporulation [97]. Temperature affects the physiological interactions between the host and the pathogen, and the optimal temperature for EF to infect their hosts is generally between 22 °C and 30 °C [98]. Yeo et al. [99] evaluated the impact of temperature on the germination and growth of EF and their pathogenicity towards two species of aphids. The results showed that the in vitro spore germination rate of all isolates was slower at 10 °C and 15 °C than at 20 °C and 25 °C. Similarly, the in vitro growth of most EF isolates was adversely affected at 10 °C and 15 °C. Moore [100] experimentally determined the virulence of three EF-B. bassiana, Aspergillus flavus, and Fusarium solani - against southern pine beetles at different temperatures. The results showed that southern pine beetles inoculated with the three fungal strains died faster at 30 °C than at other temperatures [100]. Mishra et al. [101] found that the mortality rate of adult and larval houseflies was the highest at 30 °C when treated with B. bassiana.
In addition to temperature, higher humidity is also an important condition required for EF to infect their hosts [102]. Hu et al. [103] reported that humidity is an important factor affecting the germination of EF spores and their infection potential. The spore germination rate of B. bassiana strains increased with increasing humidity. Mishra et al. [101] found that the mortality rate of adults reached 100% at 90% and 100% relative humidity, indicating that humidity is an important factor affecting the virulence of B. bassiana. Maranga et al. [104] applied oil and water formulations of B. bassiana and M. anisopliae spore mixtures to control Haemaphysalis colorata in the rainy and dry seasons. In the rainy season, the mortality rates of H. colorata treated with the oil and water formulations of the spore mixture were 92% and 49%, respectively. However, in the dry season, the mortality rates of the same treatments were 24% and 17%, respectively, which decreased by 68% and 32%, respectively [104]. The above studies indicate that the growth and development of EF are influenced by the temperature and humidity in their growth environment.
2.2.2. Landscape Composition Could Affect the Community Structure of EF
Different landscape compositions significantly influence the spatial distribution of natural enemies [76]. Complex landscapes, compared to simple ones, enhance local agricultural biodiversity more effectively [105]. These complex landscapes support a greater diversity of beneficial species, including pollinators and natural enemies, thereby increasing species richness, abundance, and evenness. They also provide more ecological resources, which is crucial for enhancing regional biodiversity and ecological service functions [106]. Natural or semi-natural habitats are particularly important as reservoirs for natural enemies, contributing to sustainable pest control [107,108]. At the landscape level, permanent grasslands, forest edges, farmland edges, and hedgerows serve as common refuges for EF [109,110,111]. Ground vegetation plays a critical role in determining the presence of EF in soil [112]. Data on isolation rates indicate that well-vegetated soils harbor diverse EF, while no strains were isolated from desert areas [77]. In general, areas with high vegetation coverage, high humidity, diverse insect community composition, and little human disturbance were more conducive to EF growth [94,95]. It has been reported that the abundance of EF is significantly higher in forest areas with good ecological environment and suitable temperature and humidity [113]. Notably, the occurrence of EF is closely linked to insect diversity. In tropical rainforests, where insect diversity is high, EF exhibit corresponding species diversity, influencing both host population dynamics and forest nutrient cycling [114,115]. In temperate forests of North America, there are a variety of EF capable of infecting insects, which can survive in cold and wet environments and effectively control pests [116]. However, EF also plays an important role in pest control in the arid regions of Africa, and although the species of EF in these regions are different from those in humid regions, their functions in the ecosystem are equally critical [117]. EF in soils with strong biodiversity also have strong species diversity. Studies have shown that forest soils contain a richer variety of EF compared to adjacent farmland soils, with six species isolated from forest soils versus three from farmland soils [118]. In different habitat types, M. anisopliae exhibits higher densities in less disturbed permanent grasslands and farmland edges [119]. This indicates that natural or semi-natural habitats can protect microbial natural enemy biodiversity, thereby enhancing their survival rate as natural enemies of pest populations. Habitat management can create microclimates favorable for the spread, survival, and efficacy of natural enemies [120]. For effective pest control, providing non-crop habitats, enhancing plant resistance, and strengthening the role of natural enemies can improve the ecosystem's ability to perform landscape ecological services [121]. Strengthening farmland ecosystem management can promote the distribution and infection spread of pathogens, thereby enhancing control over agricultural pest populations [122], which requires a deeper understanding of the occurrence patterns of microbial natural enemies in these ecosystems.
2.2.3. The Infestation of Host Insects by EF Is Influenced by Different Host Types and Insect States
EF can form mutualistic or neutral relationships with their hosts [26,123], and their colonization and distribution are significantly influenced by the host insects [24]. In nature, CF often form insect-fungus complexes, parasitizing various insect life stages including larvae, pupae, and adults [94]. These fungi typically invade insects through the cuticle, mouth, or other body openings [124]. The susceptibility of host insects to infection by EF varies depending on the host type and developmental stage [22,77]. Aung et al. [125] investigated in the rainforests of Thailand and found that the number of EF isolated from Homoptera was higher than that from Lepidoptera and Hymenoptera. Gindin et al. [126] found that the mortality rate of M. anisopliae on the larvae of three kinds of ticks was above 90%, while the mortality rate on adults was low or even had no effect. Similarly, certain strains of EF, such as M. anisopliae and Isaria fumosorosea, demonstrate high lethality against larvae of Rhipicephalus sanguineus but do not affect adults [127]. Zhou et al. [128] found that V. lecanii interacted with microorganisms in Frankliniella occidentalis and produced sublethal effects, and the pathogenicity degree was adult > pupa > nymphs. Insect diet and environmental factors also influence their susceptibility to EF [129]. Plant nutrients can either enhance or weaken insect resistance to fungal infections [129]. For example, stem borers exhibit varying sensitivities to different EF when feeding on different plants, highlighting the critical role of plant species and nutrient composition in insect-fungal interactions [129]. The incidence, diversity, and host range of EF increase from Arctic to tropical regions [4].
2.2.4. Agricultural Activities Can Affect Soil EF
The microbial community structure varies with organic or conventional production methods [130,131]. Studies have shown that the diversity of soil fungi in natural forest soils is higher compared to cultivated soils [132]. Organic farming practices have been found to enhance the diversity of EF [133], with significantly higher abundance of these fungi in organic farmland soils compared to conventional farmland soils [131]. Klingen et al. [134] reported greater numbers, types, and diversity of EF in organic farmland soils. Similarly, Clifton et al. [131] observed higher abundance of EF in both the soil and surrounding field margins of organic farms compared to conventional farms. Additionally, the diversity of EF in organic farm soils was significantly higher than in conventional farms [134]. However, the impact of farming practices on fungal communities can vary by crop and region. For instance, studies on grape phyllosphere microbial communities found minimal effects of chemical and biological pesticides on fungal richness and diversity [135]. Tkaczuk et al. [118] reported no significant differences in EF between conventional and organic winter cereal fields in Poland. Similarly, there were no notable differences in EF between traditional and organic citrus orchards in Africa [136].
2.2.5. Altitude Is an Important Environmental Factor Affecting EF Community
The distribution of EF was observed in soils of all altitudinal gradients, with higher numbers of species and strains in the middle elevations [137]. The geological movement of the Qinghai-Tibet Plateau during the uplift process not only resulted in increased differences in altitude, climate, and vegetation types [138], but also provided an explanation for the diversity and differentiation of EF [77]. It has been found that the diversity of EF in cropland ecosystems at high altitudes is higher than that in cropland at low altitudes [92], and soil fungal community composition is different at different altitudes, and fungal diversity increases with the increase of altitude [92], the diversity index of EF was also higher at high altitudes [139]. However, altitude has different effects on different species of EF. For instance, B. bassiana increases with altitude, Whereas M. brunneum and M. robertsii decreased with increasing altitude [140].
2.2.6. The Effect of the Niche on EF
Temporal and trophic niches can also affect the diversity of EF. Due to the differences in the occurrence seasons of various fungal species, the degree of overlap in their temporal niches also varies [141,142]. For instance, C. cateniannulata mainly appears in June and September, while I. farinosus is concentrated in March and May, indicating that their temporal niche widths are relatively narrow. Additionally, the width and overlap values of the nutritional niche are closely related to the host range: the broader the host range of a fungus, the wider its nutritional niche, but the smaller the overlap with other fungi [143]. For example, B. bassiana has a wide range of host insect species [144], thus it has less overlap with other EF in terms of nutritional niche, thereby reducing competition for the same host resources [145,146]. In contrast, C. cateniannulata and I. farinosus have a narrower host range, mainly targeting Lepidoptera insects [147], resulting in a smaller nutritional niche width. In summary, EF are not only important tools for natural pest management but also key components of biodiversity and ecological balance. Future research should focus on the protection and rational utilization of EF resources to promote sustainable agricultural development and better maintain environmental and ecological balance.
3. Interactions Between EF and Host Insects
3.1. The Process by Which EF Infects Insects
EF are the only pathogenic microorganisms that can contact infected insects through the body wall under natural conditions and play a unique role in pest control [148], which can be summarized as four stages: 1) The conidium of EF adsorbs to the insect body wall through hydrophobic action [149,150]. Studies have shown that there are two ways of specific adsorption and non-specific adsorption of fungal spores on the body walls of insects [151]. During the process of infecting the body walls of insects, specific adsorption plays a very important role. However, non-specific adsorption can induce specific adsorption to a certain extent [150]; 2) Under suitable conditions, conidia germinate to produce germ tubes, appressoria, invasion nails and other structures, and then directly penetrate the insect's body wall and enter its blood cavity by secreting hydrolases and generating bulging pressure. The germ tube is formed after the spores attach to the insect body wall and finally develop into the attachment spore, which enables the EF to release a series of substances in the insect epidermis. These substances can promote the invasion and growth of hyphae and finally form invasion nails. Infection nails are formed by penetrating the host body wall through multiple actions, including mechanical stress and a range of hydrolases [3,150]. Once the germ tube enters the hemocoel, it will breed a new hypha in it. This hypha will continue to multiply in the hemocoel and gradually form more hyphae. These hyphae will produce toxins that affect the immune function of the insect and eventually lead to the death of the insect [152]; 3) The hyphae that enter the insect's blood lumen secrete various toxins or secondary metabolites to attack or paralyze the insect's immune system. They also differentiate into a cellular structure called the hyphalbody (no cell wall, yeast shape), which rapidly absorbs nutrients from the insect and eventually causes the insect to collapse and die [153,154]; 4) After the invasion of the host insect, EF will carry out large-scale reproduction in the host's nutrients, so as to grow hyphae in every part of the body of the insect. At the same time, under the right environment, the hyphae will gradually grow out of the worm and gradually cover the whole insect body [7,94]. After completing the pathogenic process, new spores leave the insect carcass and enter the soil [155], where they are stored until the next stage of infection (Figure 3). Overall, EF typically enter the host through body surface penetration, with spores adhering to the insect's body wall before entering the host [7,22,24,154].
3.2. Molecular Basis of Evolutionary Adaptation in EF
EF, a class of microorganisms capable of parasitizing insects, have adapted and optimized their ability to infect hosts through a variety of molecular mechanisms [22]. These mechanisms include not only penetration of the host body wall by secreting specific enzymes, but also strategies for colonizing the host and resisting the host immune system [156, 157.] EF secrete a variety of enzymes during the process of penetrating the host body wall [7,158], which is key to successful host invasion [159]. For example, proteases break down host protein structures, while chitinases specialize in breaking down chitin in insect exoskeletons [160,161,162]. In addition, lipases help destroy lipid fractions and relieve their growth inhibition, allowing the EF to successfully penetrate the protective layer of the insect body wall [154], B. bassiana and M. anisopliae extensively use these enzymes to promote their infection processes [148]. During the endocolonization stage of the host blood cavity, the EF further ensure their survival and reproduction in the host by secreting specific metabolites such as bassionin, oosporein, and destruxin [156,157]. These metabolites not only help the fungus maintain a nutrient supply in the host but may also be involved in suppressing host immune responses [153,157,163]. For example, bassionin can inhibit the aggregation of blood cells in host insects and hinder their immune function [164]. In addition, to defend against host immune attack, EF also secrete specific proteins that disrupt host immune recognition mechanisms [153,154]. These secreted proteins include effector proteins and immunosuppressive factors that can directly interfere with host signaling pathways and reduce immune cell activity [153,165]. For example, M. anisopliae is able to secrete an effector protein called DEP1, which increases pathogen survival by inhibiting host cell apoptosis [166]. Through these complex molecular mechanisms, EF can effectively infect and control their insect hosts, showing a high degree of adaptability to the internal environment of host insects during their long-term evolution. The discovery and study of these mechanisms are not only important for understanding the infection strategies of parasitic microorganisms, but also provide the possibility to develop pest management strategies based on biological control.
3.3. Diffusion of EF Spores
Diffusion is a crucial process for the spatial dynamics, evolution and migration of populations [167]. Species populations with poor diffusion ability may thus become isolated and disappear from the entire ecosystem [168]. The spores of CF are mainly spread by air and water, while the spores of EPF often spread by the host's own activities in addition to air and water [4,6], some ascomycetes spread over hundreds of kilometers by forming large numbers of wind-borne conidia (Small asexual propagules: The main differentiating feature of this asexual entomophytic fungus is the formation of conidia on small branches rather than in chains) [169,170]. The fruit-body structure of EF facilitates the spread of spores [171]. For example, the ejection speed of ascospores from maize head mold can reach 34.5 m/s, this means that the asci provide a turgor pressure of at least 870 atmospheres for the spore to achieve this speed [172,173]. The dispersal of such spore propagules has great effects on fungal population growth, spatial distribution patterns, collective population dynamics, and genetic diversity [174]. Among EF, Isaria sp. forms coremium and Cordyceps sp. forms a stroma, which provides a dynamic basis for spore diffusion [175]. The spread method of EF is an important way to control agricultural and forestry pests [176]. EF have evolved unique physical structures to adapt to their transmission dynamics, which can be roughly divided into long-distance dispersal by wind, current, and human activities [177], and short distance transmission by animals (e.g.; Antheraea pernyi, Lasioderma serricorne) [178]. Long-distance propagation on a large scale is a random diffusion process [156]. However, the small-scale and short-distance transmission mechanism closely related to the host is still unknown, and the spore transmission mechanism of EF is worthy of further investigation as the efficiency of biological control.
3.4. EF Completes the Parasitic Life Cycle in the Host
The body wall of an insect is the first barrier that EF needs to break through. With the help of mucus and adhesion proteins, fungal conidia attach to the host surface to germinate and form an infectious structure called appressorium [179]. With the action of mechanical pressure and body wall degrading enzymes, direct infestation by EF destroys the insect cuticle [180]. After entering the hemocoel of insects, EF will face a series of immune responses from insects, including cellular immunity and humoral immunity. During a long period of coevolution, EF have evolved a series of adaptation mechanisms in adapting to physicochemical and biological factors in their hosts [153,156,160]. EF has developed multiple strategies to adapt to host immune defense responses. For example, M. anisopliae prevents insect hemocytes from recognizing its mycelium by expressing MCL1 (collagen-like immune evasion protein) [181] and secretes toxins to inhibit V-ATPase and block the expression of AMPs coding genes, thereby escaping immune phagocytosis [182]. Oosporein produced by B. bassiana can inhibit the ProPO activity of Galleria larvae and down-regulate the expression of gallerimycin gene to resist insect immunity [183]. There are many literatures on the process and mechanism of fungi adapting to the internal environment of insects. For example, Wang et al. [6] summarized the mechanism of interaction between insect pathogenic fungi and insect hosts in detail. Qu et al. [184] found that H. satumaensis can respond to insect ecdysone, and Hsegt (ecdysone-inactivating enzyme) was first identified in EF, which plays an important role in the fungal response to insect immunity. Another noteworthy phenomenon is the ability of EF to interact with insect gut microbes [23,185,186]. EF can establish contact with insect gut microbes when they infect insects. EF perturbs host midgut microbes by regulating antimicrobial peptides and dioxygenase in the midgut [157], which enables opportunistic pathogens Serratia marcescens to enter the hematocoelum and transform into highly virulent pathogens, thereby accelerating host death [187]. The overexpression of Hsegt gene during the infection of G. mellonella by H. satumaensis may aggravate the proliferation of bacteria in the body of G. mellonella, and the increased bacteria would affect the penetration of H. satumaensis mycelium from the epidermis of G. mellonella [188], The interactions between EF and host gut microbes and endosymbionts have attracted researchers' attention, but more detailed microecological mechanisms and processes need to be studied.
3.5. Strategies for Insect Response to EF
Under stress, insects can induce changes in immune activities in vivo and produce a series of defense responses [189,190]. During the growth and development of fungi, a wide variety of volatiles are produced, and these volatiles play important roles in the ability of fungi to communicate with each other, defend themselves against the outside world, or attract other organisms such as insects [191]. When EF infect insects, they exhibit avoidance behaviors toward EF. The insect can identify and distinguish different types of fungi by releasing specific chemicals, which can be detected by the same species and transmitted between individuals as warning signals [23,153,192]. At the same time, the living habits of social insects are very different from those of non-social insects, especially the highly evolved defense capabilities [193], which pose a challenge to the use of microorganisms, including EF, to control pests. In insects, superoxide dismutase, peroxidase and catalase are important protective enzymes to prevent oxidative damage. Through the mutual coordination of the three enzymes, the free radicals in insects can be kept at a low level, so that insects are protected from free radical poisoning [194]. It was found that the activities of superoxide dismutase, catalase and peroxidase in the larvae, infected with B. bassiana increased first and then decreased, and the enzyme activity at 60 h of infection was significantly lower than that of uninfected larvae [195], which indicated that the biocontrol products of EF had inhibitory effects on all three protective enzymes in agricultural pests. The growth, development and behavior of insects are affected by gene expression and regulation [196,197]. Previous studies have shown that cadherin genes, ABC transporter genes and peptidoglycan recognition protein genes in insects can activate the immune response of insects to resist the invasion of pathogens and play an important role in the immune pathway of insects. All these genes were up-regulated after feeding on B. bassiana-maize symbionts, these results indicate that B. bassiana can induce stress responses in insects [198]. Insect immune-related genes such as UDP-glucurono-transferase genes, cytochrome P450 genes, and heat shock protein genes are associated with insect drug resistance and detoxification mechanisms [199], downregulation of related gene expression after feeding on B. bassiana - maize symbionts would indicate reduced insect resistance. Previous studies have also shown that Ostrinia furnacalis can induce the down-regulation of the expression of some resistant-related genes under the stress of EF [200], indicating that B. bassiana can not only infect pests through the body surface, but also form immunosuppressive effects on insects after feeding on them. In conclusion, the spores of EF and symbionts of crop plants can be fed by insects, indirectly affecting the growth and development of insects, and reducing the defense performance of insects by regulating the expression of protective enzymes and resistant-related genes in insects, thereby reducing the damage to plants. It is of great significance to understand the interaction between microorganisms, plants and insects. At the same time, it can provide theoretical basis for the rational use of EF in agricultural ecosystems, and lay the foundation for the development of new pest products for ecological regulation by EF.
4. EF Colonize Plants
4.1. EF Promotes Plant Growth
EF can colonize different host plants in the form of endophytic fungi, thereby promoting plant growth [201,202,203]. Several studies have reported different EF species as natural colonists/endophytes of many economically important crops, including Z. mays, Solanum tuberosum, Gossypium hirsutum, Oryza sativa, S. lycopersicum, and Cicer arietinum [200,202]. In addition to colonizing seedlings, EF spores are also capable of colonizing seeds [110,203]. For instance, by soaking the spores in the suspension, B. bassiana spores were found in S. lycopersicum leaves, stems and roots after 18 weeks of seeding [204]. EF colonize plants and play a significant role in the growth and health of the plants (Figure 4). As endophytic fungi interact with plant hosts, EF can alleviate biotic stress in host plants and play a role in plant health [205,206], which can reduce and eliminate the effects of pathogenic fungi that are harmful to plants [207]. In addition, EF also acts as plant fertilizer and promotes plant growth [208]. A growing body of evidence also suggests that EF can function as plant endophytes, plant pathogen antagonists, and plant growth promoters [13]. Insect-derived nitrogen can be transferred to plants through the interaction between EF and plants [209], which can not only increase plant productivity [31], but also improve the overall performance of plants [32,210]. Owing to their biocontrol properties, EF can colonize plants in the natural state or under different artificial inoculation methods, which can enhance the resistance of host plants to diseases and pests, increase the mortality of herbivorous pests, reduce the probability of pests invading crops, and delay the erosion and reproduction of pests [31]. For example, B. bassiana and M. anisopliae successfully colonized Z. mays and significantly promoted its growth in hydroponic systems [211]. C. cateniannulata can infect and colonized the roots, stems and leaves of S. lycopersicum seedlings by seed soaking, promoting the growth of S. lycopersicum seedlings and improving the activities of antioxidant enzymes [212]. Zheng et al. [213]by evaluating M. flavoviride, C. fumosorosea, B. bassiana, to study the growth promotion and control effects of five mycotic strains of M. anisopliae and M. rileyi on the S. lycopersicum leaf aphid (Phthorimaea absoluta). The results showed that all the five strains could colonize in S. lycopersicum. Both of them can promote the growth of S. lycopersicum and have good control effects on Tuta absoluta. EF can also act as antagonists of plant diseases, they can inhibit the growth of plant pathogens and reduce the occurrence of plant diseases. For instance, B. bassiana, as an endophyte, is able to control pests and plant pathogenic fungi and increase plant resistance to insects [214,215]. The relationship between EF and plants is complex, and further research is needed to fully understand the mechanism of plant growth promotion.
4.2. EF-Insect-Plant Interaction
In the ecosystem, there is a close and complex relationship between plants, microorganisms and insects. EF can change the behavior and ecological adaptability of phytophagous insects by inducing the defense performance and nutrient composition of plants [216]. EF colonizes plants, which can cause changes in the individual behavior of insects, including feeding behavior, spawning behavior, etc.; and can also increase the death rate of herbivorous insects, so as to reduce the damage of pests to crops [7,22,23]. EF has multiple ecological functions, can exchange elements with plants, can establish symbiotic systems with plants, and have ecological values such as improving crop yield and resisting plant diseases (Figure 5). Successfully colonized B. Bassiana can not only spread inside plants, but also provide insecticide protection for the whole plant [212,217], can also improve plant stress resistance [218]. It can be seen that the endophytic colonization of EF in plants can indirectly have adverse effects on multi-planting feeding insects, and further alleviate the harm degree of insects to plants. Therefore, the interaction between EF and plants can affect phytophagous insects from multiple perspectives. The indirect effect of EF on insects has received extensive attention in recent years. However, the current research on the mechanism of EF-plant-insect interaction remains to be further explored.
5. EF Controls Agricultural Pests
In the past few decades, chemical insecticides have been frequently used to control pests all over the world. However, these chemicals are harmful to both human health and ecosystems, and some pests have developed resistance to many of them [7]. Biocontrol is an economical, eco-friendly method for controlling nematodes and pests that have a negative economic impact on agriculture [219]. In recent years, with the application of EF in pest control and the strategy of integrated pet control, EF has been paid more and more attention by researchers. EF is the largest group of entomopathogenic microorganisms with dual biocontrol activities and growth promotion [220]. EF can not only control the natural population growth of agricultural and forestry pests, but also reduce the harm of pathogens to host plants, improve plant disease resistance [220], and promote plant growth [177]. Unlike insect pathogens and viruses that cause disease after oral ingestion, EF infects insects by directly penetrating their cuticle [221]. This makes it ideal for developing contact fungal insecticides that can control not only chewing pests, but also insects with mouthsucking organs, such as aphids and mosquitoes [70]. EF infects the host in a variety of ways, including direct penetration through the exoskeleton of insects, oral infection, and so on [7,222]. Some EF, such as B. bassiana, M. anisoplariae, V. cerococcus, Fusarium, etc.; have been developed as biological control agents and are widely used in pest control [5]. In practical applications, EF not only causes pathogenicity to pests, but also improves disease resistance and promotes plant growth by establishing symbiotic relationships with plants [223]. In agricultural production, pests infected by EF do not occur naturally at an efficient rate of death or on a large scale to prevent crop damage. Because the existing fungal biocontrol agents can infect pests for a long time and are affected by the external environment, the control effect is unstable. The main factors include temperature, humidity, ultraviolet intensity, etc.; which will affect the growth of strains and thus affect the insecticidal efficiency [5,224]. In order to deal with the low efficiency of biological control of EF, some scholars have proposed to improve the control effect of pests through the mixing effect of biological insecticides, chemical pesticides and surface-active substances [224].
EF, as a beneficial biological control measure, has great potential value in pest control, but there are many limitations in practical application. Some entomogenous fungal species, such as I. fumosorosea and I. farinosa, can infect multiple hosts without exhibiting any of the harmful effects associated with chemical pesticides and are therefore considered safe and environmentally friendly [225]. Therefore, the combined application of EF and chemical insecticides can be an effective new strategy for green pest control [195,209]. The combined application of EF and chemical insecticides can significantly reduce the amount of insecticides used, thereby reducing the damage caused by insects to crops, and also greatly improving crop yields [226]. This strategy can also reduce the impact of pesticides on the environment and ecosystems, helping to improve human health. EF and chemical pesticides show synergistic effects due to several factors. First of all, the cleaning ability of the insect body surface was reduced after pesticide stress, and the probability of spores adhesion of pathogenic fungi was increased [23]. Secondly, pesticides can reduce the food intake of pests, which makes them more prone to hunger and thirst, and further lead to the decline of insect pathogen resistance to fungi [226]. At the same time, chemical pesticides can also affect the immunity of pests and resistance to EF [226]. The sensitivity of different strains to chemical pesticides varies greatly, and the sensitivity of the same strain to different chemical pesticides is also different [227]. Selecting excellent strains with high virulence can reduce the use of insecticides and maintain the stability of the ecological environment [228]. Therefore, it is usually necessary to evaluate the compatibility of chemical pesticides and EF when they are used to control pests. Chemical insecticides with good compatibility with EF, combined with EF, can obviously improve the control effect of EF and can play a synergistic role.
6. Mycelia of EF Participated in the Construction of Mycelial Network System
After the mycelium of EF are connected, they form a three-dimensional network structure [229]. Our previous study found that the mycelium of C. chanhua is hydrophobic, which helps the conidium of C. chanhua adhere to the insect epidermis and affects its virulence [42], which is the key to the survival and environment adaptation of C. chanhua. There are "H" type fusion mycelia between the mycelium of C. chanhua, which is conducive to gene exchange and signal transduction between the strains or mycelia. The three-dimensional structure formed between mycelium also has the function of transport, and some soil fungi can also promote the movement of specific bacteria along the mycelium network [230], thus acting as a fungal pipeline or fungal "highway". Fungi can also be used as a carrier to enrich and transport specific bacterial species, for example, Pythium ultimum mycelium in soil can be used as a transport for the soil contaminant enrichment areas of Bacillus, Rhodococcus, and Pseudomonas [231]. Warmink et al. [230] found that Burkholderia terrae strain BS001 not only migrates along fungal mycelia, but also prompts bacteria such as Dugesia japonica BS003 to move along fungal "superhighways". In addition to being able to transport microorganisms, the "fungal network" also serves as a logistics channel for the distribution of material elements, and M. robertsii even receives horizontally transferred DNA information [232]. At the same time, the mycelium formed by EF also has the function of preventing harmful microorganisms from entering the insect body, thus ensuring the normal growth and development of EF [230]. The research of mycelial network system construction model and ecological operation mechanism of EF is of great value in the ecological application of fungi.
7. EF Participate in Soil Nitrogen Cycling
EF not only controls host populations, but also affects nutrient cycling and material flow in ecosystems [114]. It is well known that although nitrogen makes up 78% of the total atmosphere of the Earth, it cannot be directly used as a nitrogen source for plants unless nitrogen can be fixed by microbial symbionts or other nitrogen-fixing microorganisms, and then the available nitrogen-containing compounds can be transferred into the plants [233]. Up to 31% of plant nitrogen in the ecosystem flows to herbivorous insects [12]. Plants can also regain nitrogen previously lost to herbivorous insects through association with endophytic fungi [13]. After nitrogen is transferred to the plant, it will be dissolved with plant secretions and dead individuals (including litter). Subsequently, nitrogen is reduced to the soil through microbial mineralization. Some EF establish a symbiotic relationship with plants, in which EF absorb carbon from host plants while providing host plants with more ways to obtain nitrogen from insect sources, thereby increasing their own biomass [11,233]. This mutually beneficial relationship benefits both parties and helps improve soil fertility. EF can accelerate the nitrogen conversion process in the soil (Figure 6). When insects are infected with EF, they break down and release nitrogen-rich compounds into the soil. This decomposition provides a source of nitrogen that plants and other soil microorganisms can utilize [234]. In our previous study, we found that C. chanhua would discharge nitrogen from its body and absorb nitrogen from the soil environment during its growth and development. The mycelium of EF makes the mycelium in the body of C. chanhua and the soil environment transfer to each other [229]. Some scholars have found that Five species of M. anisopliae, B. bassiana and L. lecanii have demonstrated the ability to transfer nitrogen to plants [13]. Behie et al. [234] used 15N-ammonium sulfate solution to label G. mellonella, and found that M. robertsii not only had a lethal effect on G. mellonella larvae, but also transferred nitrogen from G. mellonella to plants of Panicum virgatum to fix nitrogen after infection [234]. In addition, EF can stimulate the growth of various soil microorganisms, promoting a more diverse microbial network. This diversity can improve the efficiency of nitrogen conversion processes in soil [211]. Studies have shown that EF can stimulate soil ammoniation and nitrification, with maximum increases of 12.4% and 36.8%, respectivel. Low concentrations of EF contribute to denitrification, while high concentrations inhibit denitrification [235]. In contrast, chemical pesticides such as aceamidophos reduce ammoniation, nitrification, and denitrification rates [234]. The change of nitrogen content in soil can lead to the change of community composition of EF [236]. Soil carbon/nitrogen ratio and total nitrogen level are extremely significant environmental factors affecting soil EF community in this region, EF community is positively correlated with carbon/nitrogen ratio and soil total nitrogen content [234,237]. In addition, EF can also survive as saprophytic forms [238], an important member of soil ecosystem [107,108]. EF can decompose organic matter, participating in material cycling in ecosystems and maintaining soil health [112,146].
8. Conclusions and Future Perspectives
EF are rich in resources and various species, and are an important part of biodiversity. Their existence increases the diversity of species in the ecosystem, and plays a great role in the ecosystem, such as controlling the number of pests, promoting plant growth, participating in the construction of mycelium networks, participating in the soil nitrogen cycle, etc.; which is crucial to the health and stability of the ecosystem. EF is a valuable resource the following points should be paid attention to in the future research on EF:
8.1. Fully Develop the Medicinal and Edible Values of CF
CF, as a class of filamentous fungi with dual ecological and medicinal significance, regulate arthropod population dynamics through parasitism within ecosystems [44]. Representative species of CF include C. sinensis, C. chanhua, and C. militaris, are rich in bioactive compounds such as cordyceps polysaccharides [239], cordycepin [32], and cordycepic acid [240]. These species have been demonstrated to exhibit multiple health-promoting properties, including anti-tumor effects38, immune modulation [37], fatigue resistance [34], antioxidant activity [39,62], and intervention in metabolic disorders (e.g.; diabetes and hypertension) [241]. They hold considerable promise for applications in chronic disease management and anti-aging therapies [242]. For instance, cordyceps polysaccharides activate macrophages and promote IL-10 secretion via the gut microbiota-immune axis [243]; Cordycepin inhibits adenosine deaminase to disrupt DNA repair in tumor cells [244]; And cordycepic acid enhances insulin sensitivity by activating the AMPK pathway [241]. With advancements in the health industry, their applications have expanded into functional foods [245], antioxidant skincare products [246], and immune-enhancing animal feed [247]. Nevertheless, wild resources are under threat due to overexploitation [44,248]. Future research should prioritize the integration of multi-omics approaches to decipher synergistic interactions among bioactive components, the development of green extraction methods such as supercritical CO2 extraction [245], and the utilization of artificial intelligence for optimizing metabolic network predictions and fermentation strategies [249], thereby facilitating the transition of CF from resource-dependent to technology-driven systems.
8.2. Make Full Use of the Potential of EF Biological Control Agents
EF, which regulate pest populations via parasitism, antagonism, and competition [250], exhibit both vertical transmission (through eggs) and horizontal dissemination capabilities (via airborne spores) [251]. Their environmentally friendly nature, target specificity, and low likelihood of inducing pest resistance render them valuable for reducing reliance on chemical pesticides and advancing sustainable agriculture [252,253]. For example, the combination of Trichoderma harzianum with low-dose fungicides can enhance the inhibition of tomato gray mold by over 50% [254]. However, their field application is constrained by limited environmental adaptability, formulation stability issues, and weak synergistic mechanisms. Strain infection efficiency is significantly influenced by temperature (25-30 °C), humidity (>92% RH), and pH (5.5-7.0) [251]. For instance, the spore formation of Trichoderma spp. requires high humidity conditions [251]. Additionally, ultraviolet radiation and extreme climatic conditions can easily inactivate spores, necessitating improvements in durability through sodium alginate-chitosan microcapsules or silica-based nanocarriers [255]. Furthermore, the control effect of single bacterial agent is limited, and there is an urgent need to explore the synergistic mechanism between EF and insect pathogenic nematodes or plant resistance substances [31].
8.3. Environmental Driving Mechanisms and Technical Analysis of EF Biodiversity
The biodiversity of EF is shaped by the interplay of host specificity, geographical isolation, and environmental factors such as temperature, humidity, and vegetation type [256]. Investigating the community structure, ecological distribution patterns, and host interaction mechanisms of entomopathogenic fungi across diverse habitats serves as a critical foundation for developing resource protection strategies and sustainable utilization plans. Recent advancements in high-throughput sequencing technologies, such as ITS amplicon sequencing, have markedly enhanced the efficiency and resolution of fungal diversity studies [257]. Moving forward, metagenomics offers the potential to uncover functional genes in EF, hereby elucidating the synthesis and regulation networks of their insecticidal metabolites [258]. Furthermore, the integration of environmental DNA (eDNA) technology enables non-invasive monitoring of the spatio-temporal dynamics of entomopathogenic fungi in forest and agricultural ecosystems. For instance, soil eDNA has been successfully employed to track the population expansion of B. bassiana [259]. Additionally, combining geographic information systems with ecological niche models, such as MaxEnt, allows for quantifying the influence of climatic variables (e.g.; annual average temperature and precipitation), host insect distributions, and vegetation coverage on the suitable habitats for EF [260]. This approach can also facilitate predictions of the potential distribution of EF in tropical rainforests and temperate agricultural regions, optimizing targeted application strategies for these fungi as biocontrol agents [32]. Collectively, the integration of these multi-technology approaches not only provides robust data support for constructing comprehensive EF resource banks but also establishes a solid theoretical framework for the precise management of agricultural and forestry pests and ecological restoration.
8.4. The Multi-Dimensional Ecological Roles and Molecular Regulatory Mechanisms of EF Hyphae
The membrane structure formed by the mycelium of EF during infection, such as the hyphal sheath of B. bassiana, plays a role in nutrient transport, microbial interaction and host defense. Mycelia form a selective barrier through the extracellular matrix, which inhibits the invasion of pathogens (such as P. aeruginosa) while promoting the directional migration of commensal bacteria (such as S. marcescens) [42]. EF hyphal network can also degrade the cuticle of insect body wall by secreting extracellular enzymes and release toxins (such as beauvericin) to damage the host immune system [180,213,216,258]. Studies have shown that secondary metabolites of M. robertsii (such as destruxin A) induce apoptosis by inhibiting the activation of key proteins of the insect Toll pathway (Spatzle) [261]. In addition, the volatile organic compounds emitted by mycelium can regulate the rhizosphere microbial community [11]. In soil ecological restoration, hyphae reduce local pH by secreting organic acids such as oxalic acid and citric acid and activate nitrification gene to enhance nitrogen mineralization efficiency [262]. The residual mycelial network can also adsorb heavy metals such as Cd²⁺ with an enrichment efficiency of 85.3% through extracellular polymers and mediate the enzymatic degradation of pyrethrins [263]. In the future, it is necessary to integrate multi-omics technologies (such as metabolomics and metagenomics) to analyze the hyph-associated metabolite diversity and its environmental response mechanisms. In addition, the construction of hyphomycet-microbial-plant interaction models can promote the precise application of EF in green agriculture and ecological restoration.
8.5. Multi-Functional Ecological Interaction Mechanisms of EF and Their Agricultural Application Prospects
EF not only achieve targeted biological control by parasitizing insects [5], but also colonize plant rhizosphere or endophytic habitats, thereby exerting synergistic growth-promoting and stress-resistance functions [212]. Their metabolites directly promote root differentiation and elongation through the secretion of indole acetic acid and gibberellic acid, as well as the synthesis of catechol siderophores and acid phosphatase [264]. For instance, Aspergillus fumigatus enhances host drought resistance via alkaloid production [265]. The simultaneous activation of jasmonic acid and salicylic acid signaling pathways upregulates disease-associated proteins and induces systemic resistance in plants [266]. T. harzianum increases the activities of superoxide dismutase and peroxidase by 2.3-fold and 1.8-fold, respectively, alleviating oxidative stress damage to chloroplasts [267], while inhibiting F. oxysporum mycelial growth (67% inhibition) through the release of volatile organic compounds such as 6-pentyl-α-pyranone [268]. Furthermore, cyclic peptides and terpenes secreted by hyphae directly disrupt pest midgut mitochondrial function [261]. Strigolactones activate the symbiotic signaling pathway under low phosphorus stress, promoting the secretion of short-chain chitin oligosaccharides by mycorrhizal fungi. This process also suppresses the pathobiologically associated molecular pattern immune response [269]. Future studies should focus on targeted gene improvement, metabolic regulatory network analysis, and ecological security assessment.
8.6. The Mechanism of Soil Carbon and Nitrogen Cycling Mediated by EF
EF regulate soil carbon and nitrogen cycles through unique nutritional and metabolic pathways. In the nitrogen cycle, their mycelial networks can transfer nitrogen elements (such as amino sugars and proteins) produced from the decomposition of insect cuticles to the rhizosphere microdomain, and release excess nitrogen (such as NH₄⁺ and NO₃-) into the soil through excretion, significantly enhancing the nitrogen utilization efficiency of plants (nitrogen transfer efficiency reaching 23-37%) [270]. For instance, after B. bassiana infects Spodoptera litura, the mycelial residues can release 15.8 mg/g of organic nitrogen [262]. In the carbon cycle, EF secrete extracellular enzymes to degrade chitin in insect cuticles (degradation rate 68-92%), releasing dissolved organic carbon into the soil [113]. Their mycelial residues and metabolic products form organic-mineral complexes through mineral adsorption, promoting the long-term sequestration of soil organic carbon [271]. At the same time, mycelia regulate the rhizosphere microbial community (such as a 1.5-fold increase in the abundance of nitrogen-fixing bacteria Azotobacter and a 60% decrease in the abundance of pathogenic bacteria Ralstonia), enhancing the microbial carbon pump effect and increasing the carbon use efficiency to 0.45-0.6248 [272]. However, current research has deficiencies in the electron transfer mechanism at the mycelium-mineral interface, the ecological toxicity of metabolic products, and climate responses. In the future, it is necessary to focus on multi-omics technologies, strain improvement and climate response models, while taking into account ecological security and policy adaptability, in order to achieve efficient application of EF in the carbon neutrality strategy.
Author Contributions
Conceptualization, Y.R.; methodology, G.H. and T.W.; validation, G.H. and H.W.; resources, T.W. and Y.R.; writing—original draft preparation, G.H. and T.W.; writing—review and editing G.P. and Y.R.; visualization, H.L.; supervision, Z.W.; project administration, Y.R.; funding acquisition, T.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Ten Thousand Talents Program Leading Talent Cultivation Fund (2023), the Guizhou Province High-level Innovative Talent Cultivation Fund (2016, 2022), the Science and Technology Plan Project for Large and Medium-sized Industrial Enterprises without R&D Activities in Guizhou Province (the Qianke Unified Research2024 No. 456), the 2025 Guizhou Provincial Science and Technology Commissioner Innovation and Entrepreneurship Service and Training Demonstration Point (Demonstration Base) Construction Project.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| EF | entomogenous Fungi |
| CF | cordyceps fungi |
| EPF | entomophthoralean fungi |
References
- Yan, J. Q.; Bau, T. Cordyceps ningxiaensis sp. nov.; a new species from dipteran pupae in Ningxia Hui Autonomous Region of China. Nova hedwigia 2015, 100, 251–8. [Google Scholar] [CrossRef]
- Saminathan, N.; Subramanian, J.; Sankaran Pagalahalli, S.; Theerthagiri, A.; Mariappan, P. Entomopathogenic fungi: translating research into field applications for crop protection. Arthropod-Plant Interactions 2025, 19, 8. [Google Scholar] [CrossRef]
- Mora, M.A.; Castilho, A.M.; Fraga, M.E. Classification and infection mechanism of entomopathogenic fungi. Arquivos do Instituto Biológico 2017, 84, e0552015. [Google Scholar] [CrossRef]
- De Wint, F.C.; Nicholson, S.; Koid, Q.Q.; Zahra, S.; Chestney-Claassen, G.; Seelan, J.S.; Xie, J.; Xing, S.; Fayle, T.M.; Haelewaters, D. Introducing a global database of entomopathogenic fungi and their host associations. Scientific Data 2024, 11, 1418. [Google Scholar] [CrossRef] [PubMed]
- Mantzoukas, S.; Kitsiou, F.; Natsiopoulos, D.; Eliopoulos, P.A. Entomopathogenic fungi: interactions and applications. Encyclopedia 2022, 2, 646–56. [Google Scholar] [CrossRef]
- Wang, C.; Wang, S. Insect pathogenic fungi, genomics, molecular interactions, and genetic improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; Ali, H. Insect-fungal-interactions: A detailed review on entomopathogenic fungi pathogenicity to combat insect pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Zou, X.; Cao, W.; Xu, Z.; Liang, Z. Two new species of Hirsutella (Ophiocordycipitaceae, Sordariomycetes) that are parasitic on lepidopteran insects from China. MycoKeys 2021, 82, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.A.; González-Olabarria, J.R.; Lyver, P.O.B.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef]
- Liang, Y.; Hong, Y.; Mai, Z.; Zhu, Q.; Guo, L. Internal and external microbial community of the Thitarodes moth, the host of Ophiocordyceps sinensis. Microorganisms 2019, 7, 517. [Google Scholar] [CrossRef] [PubMed]
- Behie, S.W.; Bidochka, M.J. Carbon translocation from a plant to an insect-pathogenic endophytic fungus. Nat. Commun. 2017, 8, 14245. [Google Scholar] [CrossRef] [PubMed]
- Lovett, G.M.; Christenson, L.M.; Groffman, P.M.; Jones, C.G.; Hart, J.E.; Mitchell, M.J. Insect defoliation and nitrogen cycling in forests. BioScience 2002, 52, 335–341. [Google Scholar] [CrossRef]
- Behie, S.W.; Zelisko, P.M.; Bidochka, M.J. Endophytic insect-parasitic fungi translocate nitrogen directly from insects to plants. Science 2012, 336, 1576–1577. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Cheng, X.; Liu, X.; Xiang, M. Genome Studies on Nematophagous and Entomogenous Fungi in China. J. Fungi 2016, 2, 9. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Zou, X.; Yu, L.Q.; Zhang, J.; Han, Y.; Zou, X. A new entomopathogenic fungus, Ophiocordyceps ponerus sp. nov., from China. Phytotaxa 2018, 343, 116–126. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Y.; Zhang, X.; Li, Y.; Zhang, H.; Zhang, Z. Transcriptomic analysis of the orchestrated molecular mechanisms underlying fruiting body initiation in Chinese cordyceps. Gene 2020, 763, 145061. [Google Scholar] [CrossRef] [PubMed]
- Gebremariam, A.; Chekol, Y.; Assefa, F. Phenotypic, molecular, and virulence characterization of entomopathogenic fungi, Beauveria bassiana (Balsam) Vuillemin, and Metarhizium anisopliae (Metschn.) Sorokin from soil samples of Ethiopia for the development of mycoinsecticide. Heliyon 2021, 7, e07091. [Google Scholar] [CrossRef] [PubMed]
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic fungi: unconventional applications. Rev. Environ. Sci. Biotechnol. 2020, 19, 23–42. [Google Scholar] [CrossRef]
- Das, G.; Shin, H.S.; Leyva-Gómez, G.; Del Prado-Audelo, M.L.; Cortés, H.; Singh, Y.D.; Panda, M.K.; Mishra, A.P.; Nigam, M.; Saklani, S.; et al. Cordyceps spp.: A review on its immune-stimulatory and other biological potentials. Front. Pharmacol. 2021, 11, 602364. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, T.; Middelman, A.; Koenraadt, C.J.; Takken, W.; Knols, B.G. Factors affecting fungus-induced larval mortality in Anopheles gambiae and Anopheles stephensi. Malar. J. 2010, 9, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Feng, P.; Wang, C. Fungi that infect insects: altering host behavior and beyond. PLoS Pathog. 2015, 11, e1005037. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Cao, Y.; Li, Y.; Wang, Y.; Guo, K.; Wang, C. A life-and-death struggle: interaction of insects with entomopathogenic fungi across various infection stages. Front. Immunol. 2024, 14, 1329843. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Shang, J.; Sun, Y.; Tang, G.; Wang, C. Fungal infection of insects: molecular insights and prospects. Trends Microbiol. 2024, 32, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Q.; Wang, N.; Qu, L.H.; Li, T.H.; Zhang, W.M. Determination of the anamorph of Cordyceps sinensis inferred from the analysis of the ribosomal DNA internal transcribed spacers and 5.8 S rDNA. Biochem. Syst. Ecol. 2001, 29, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Rehner, S.A.; Aime, M.C.; Smith, K.E.; Rossman, A.Y.; Chaverri, P.; Shrestha, B.; Spatafora, J.W. A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Sung, G.H.; Ban, S.; Nakagiri, A.; Chen, M.; Huang, B.; Li, Z.; Spatafora, J.W. The phylogenetic placement of hypocrealean insect pathogens in the genus Polycephalomyces: an application of One Fungus One Name. Fungal Biol. 2013, 117, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Mongkolsamrit, S.; Noisripoom, W.; Thanakitpipattana, D.; Wutikhun, T.; Spatafora, J.W.; Luangsa-ard, J.J. Molecular phylogeny and morphology reveal cryptic species in Blackwellomyces and Cordyceps (Cordycipitaceae) from Thailand. Mycol. Prog. 2020, 19, 957–983. [Google Scholar] [CrossRef]
- Xia, E.H.; Yang, D.R.; Jiang, J.J.; Zhang, Q.; Liu, Y.; Zhang, Y.; Zhang, Y.; Zhang, H.B.; Yao, N.Y.; Ge, X.J.; et al. The caterpillar fungus, Ophiocordyceps sinensis, genome provides insights into highland adaptation of fungal pathogenicity. Sci. Rep. 2017, 7, 1806. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, S.M.; Rauf, A.; Rehman, S.; Ahmad, P. Plant-Entomopathogenic fungi interaction: recent progress and future prospects on endophytism-mediated growth promotion and biocontrol. Plants 2024, 13, 1420. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Li, J.; Wang, S.; Li, C. Cordycepin extracted from Cordyceps militaris mitigated CUMS-induced depression of rats via targeting GSK3β/β-catenin signaling pathway. J. Ethnopharmacol. 2025, 340, 119249. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, Z.; Malik, K.; Li, C. Comparative metabolite profiling between Cordyceps sinensis and other Cordyceps by untargeted UHPLC-MS/MS. Biology 2025, 14, 118. [Google Scholar] [CrossRef] [PubMed]
- Pei, H.; Zhang, Y.; Li, X.; Guo, M.; Wang, J.; Li, Y. Preparation, characterization, bioactivity, and safety evaluation of PEI-modified PLGA nanoparticles loaded with polysaccharide from Cordyceps militaris. Adv. Compos. Hybrid Mater. 2025, 8, 93. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, Z.; Shi, F.; Wu, W. Application of UHPLC-ESI-MS/MS-based metabonomic techniques to analyze the Cordyceps cicadae metabolic profile changes to the CO(NH₂)₂ response mechanism in the process of ergosterol synthesis. Fermentation 2025, 11, 90. [Google Scholar] [CrossRef]
- Filippou, C.; Coutts, R.H.A.; Kotta-Loizou, I.; El-Kamand, S.; Papanicolaou, A. Transcriptomic analysis reveals molecular mechanisms underpinning mycovirus-mediated hypervirulence in Beauveria bassiana infecting Tenebrio molitor. J. Fungi 2025, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Wang, J.; Wang, C.; Wang, Y. Fungal evasion of Drosophila immunity involves blocking the cathepsin-mediated cleavage maturation of the danger-sensing protease. Proc. Natl. Acad. Sci. USA 2025, 122, e2419343122. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; He, D. Potential geographical distribution of Cordyceps cicadae and its two hosts in China under climate change. Front. Microbiol. 2025, 15, 1519560. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Li, J.; Chen, L.; Zhang, X.; Wang, Y. Antioxidant and anti-aging activities of polysaccharides from Cordyceps cicadae. Int. J. Biol. Macromol. 2020, 157, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Sui, L.; Wang, X.; Zhang, Y.; Li, Y.; Guo, J.; Zou, X. A strain of Talaromyces assiutensis provides multiple protection effects against insect pests and a fungal pathogen after endophytic settlement in soybean plants. Biol. Control 2025, 201, 105703. [Google Scholar] [CrossRef]
- Zhang, D.; Qi, H.; Zhang, F. Parasitism by entomopathogenic fungi and insect host defense strategies. Microorganisms 2025, 13, 283. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Li, Y.; Chen, J.; Wang, H.; Zou, X. Filtration effect of Cordyceps chanhua mycoderm on bacteria and its transport function on nitrogen. Microbiol. Spectr. 2024, 12, e01179–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Z.; Li, X.; Malik, K.; Li, C. Metagenomic analysis: alterations of soil microbial community and function due to the disturbance of collecting Cordyceps sinensis. Int. J. Mol. Sci. 2024, 25, 10961. [Google Scholar] [CrossRef] [PubMed]
- Zou, X. Research and development of Cordyceps resources from an ecological perspective. In Cordyceps and Allied Species; Springer: Singapore, 2025; pp. 27–62. [Google Scholar]
- Scott, M.E. Helminth-host-environment interactions: Looking down from the tip of the iceberg. J. Helminthol. 2023, 97, e59. [Google Scholar] [CrossRef] [PubMed]
- Shashidhar, M.G.; Giridhar, P.; Sankar, K.U.; Manohar, B. Bioactive principles from Cordyceps sinensis: A potent food supplement - A review. J. Funct. Foods 2013, 5, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.T.; Lee, C.H.; Chen, C.C.; Chen, L.C.; Huang, C.Y.; Lee, Y.J. Protective effects of Cordyceps militaris against hepatocyte apoptosis and liver fibrosis induced by high palmitic acid diet. Front. Pharmacol. 2025, 15, 1438997. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhang, Y.; Zhang, X.; Zhang, J.; Li, J. Efficacy of traditional Chinese medicine Cordyceps sinensis as an adjunctive treatment in patients with renal dysfunction: a systematic-review and meta-analysis. Front. Med. 2025, 11, 1477569. [Google Scholar] [CrossRef] [PubMed]
- Pastirčáková, K.; Zemek, R. Biodiversity and ecology of organisms associated with woody plants. Forests 2025, 16, 283. [Google Scholar] [CrossRef]
- Song, P.; Li, Y.; Zhang, H.; Wang, L.; Chen, J. Impact of Cordyceps sinensis on coronary computed tomography angiography image quality and renal function in a beagle model of renal impairment. Front. Pharmacol. 2025, 16, 1538916. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Li, X.; Chen, Y.; Wang, L.; Zhao, Y. Revisiting fermented buckwheat: a comprehensive examination of strains, bioactivities, and applications. Crit. Rev. Food Sci. Nutr. 2025, 15, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Doan, U.V.; Mendez Rojas, B.; Kirby, R. Unintentional ingestion of Cordyceps fungus-infected cicada nymphs causing ibotenic acid poisoning in Southern Vietnam. Clin. Toxicol. 2017, 55, 893–896. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.N.; de Sousa Santos, Y.; de Rezende, R.R.; Alfenas-Zerbini, P. Identification of a novel polymycovirus infecting the entomopathogenic fungus Metarhizium robertsii. Arch. Virol. 2025, 170, 58. [Google Scholar] [CrossRef] [PubMed]
- Mushyakhwo, K.; Maxwell, L.A.; Nai, Y.S.; Srinivasan, R.; Hwang, S.Y. Beauveria bassiana-based management of Thrips palmi in greenhouse. BioControl 2025, 5, 1. [Google Scholar] [CrossRef]
- Guan, J.; Gou, J.; Xu, Z.; Zhou, Y.; Zou, X. Endophytic colonization via Cordyceps cateniannulata induces a growth-enhancing effect and increases the resistance of tomato plants against Tetranychus urticae (Koch). Crop Prot. 2025, 193, 107182. [Google Scholar] [CrossRef]
- Mushtaq, A.R.; Bukhari, S.A.; Mushtaq, M.A.; Hussnain, M.; Rehman, U.U. Entomopathogenic fungi: Revolutionary allies in sustainable pest management and the overcoming of challenges in large-scale field applications. Sci. Res. Timelines J. 2025, 3, 19–23. [Google Scholar]
- Gomaa, E.Z.; Housseiny, M.M.; Omran, A.A. Fungicidal efficiency of silver and copper nanoparticles produced by Pseudomonas fluorescens ATCC 17397 against four Aspergillus species: a molecular study. J. Clust. Sci. 2019, 30, 181–196. [Google Scholar] [CrossRef]
- Lovett, B.; Bilgo, E.; Diabate, A.; St. Leger, R.J. A review of progress toward field application of transgenic mosquitocidal entomopathogenic fungi. Pest Manag. Sci. 2019, 75, 2316–2324. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Cerezo, S.; de Vries, R.P.; Garrigues, S. Strategies for the development of industrial fungal producing strains. J. Fungi 2023, 9, 834. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Chen, Y.; Guo, D.; Li, J. Traditional uses, origins, chemistry and pharmacology of Bombyx batryticatus: A review. Molecules 2017, 22, 1779. [Google Scholar] [CrossRef] [PubMed]
- Insumrong, K.; Chaiyana, W.; Sae-Lim, N.; Chansakaow, S.; Okonogi, S. In vitro skin penetration of 5α-reductase inhibitors from Tectona grandis L.f. leaf extracts. Molecules 2025, 30, 1151. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Tang, J.; Tu, J.; Guo, X. Solvent fractionation and LC-MS profiling, antioxidant properties, and α-glucosidase inhibitory activity of Bombyx batryticatus. Molecules 2025, 30, 1021. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.P.; Wen, T.C.; Chen, C.D.; Hsieh, C.W.; Sung, J.M. Polycephalomycetaceae, a new family of clavicipitoid fungi segregates from Ophiocordycipitaceae. Fungal Divers. 2023, 120, 1–76. [Google Scholar] [CrossRef]
- Saha, N.; Sharma, A.; Bora, P. Expanding the functional landscape of microbial entomopathogens in agriculture beyond pest management. Folia Microbiol. 2025, 70, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sami, A.; Ali, S.; Khan, A.; Khan, S.; Khan, S.; Khan, A.; Khan, A.; Khan, A.; Khan, A.; Khan, A.; et al. CRISPR-Cas9-based genetic engineering for crop improvement under drought stress. Bioengineered 2021, 12, 5814–5829. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, A.; Fourie, G.; van der Nest, M.A.; van der Waals, J.E.; Wingfield, M.J. CRISPR-Cas9 genome editing reveals that the Pgs gene of Fusarium circinatum is involved in pathogenicity, growth and sporulation. Fungal Genet. Biol. 2025, 177, 103970. [Google Scholar] [CrossRef] [PubMed]
- Půža, V.; Tarasco, E. Interactions between entomopathogenic fungi and entomopathogenic nematodes. Microorganisms 2023, 11, 163. [Google Scholar] [CrossRef] [PubMed]
- James, R.R.; Buckner, J.S.; Freeman, T.P. Cuticular lipids and silverleaf whitefly stage affect conidial germination of Beauveria bassiana and Paecilomyces fumosoroseus. J. Invertebr. Pathol. 2003, 84, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.Q.; Zhang, K.Q. Microbial control of plant-parasitic nematodes: a five-party interaction. Plant Soil 2006, 288, 31–45. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, J.; Sit, W.H.; Lee, C.Y.; Wan, J.M. Cordyceps cicadae induces G2/M cell cycle arrest in MHCC97H human hepatocellular carcinoma cells: a proteomic study. Chin. Med. 2014, 9, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Borneman, J.; Ruegger, P.; Liang, L.; Zhang, K.Q. Molecular mechanisms of the interactions between nematodes and nematophagous microorganisms. In Plant Defence: Biological Control; Springer: Cham, 2020; pp. 421–441. [Google Scholar]
- Gotti, I.A.; Ríos-Donato, N.; Pelizza, S.A.; López-Lastra, C.C.; Scorsetti, A.C. Blastospores from Metarhizium anisopliae and Metarhizium rileyi are not always as virulent as conidia are towards Spodoptera frugiperda caterpillars and use different infection mechanisms. Microorganisms 2023, 11, 1594. [Google Scholar] [CrossRef] [PubMed]
- Francis, F.; Fingu-Mabola, J.C.; Ben Fekih, I. Direct and endophytic effects of fungal entomopathogens for sustainable aphid control: a review. Agriculture 2022, 12, 2081. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, Y.; Tang, X.; Li, G.; Qiu, R. Metabolism of Penicillium oxalicum-mediated microbial community reconstructed by nitrogen improves stable aggregates formation in bauxite residue: a field-scale demonstration. J. Clean. Prod. 2025, 493, 144963. [Google Scholar] [CrossRef]
- Ye, R.; Li, Q.; Wang, X.; Li, X.; Guo, S.; Chen, S.; Wang, Y. Genomic, transcriptomic, and ecological diversity of Penicillium species in cheese rind microbiomes. Fungal Genet. Biol. 2024, 171, 103862. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Yang, T.; Zhang, K.; Shen, C.; Chu, H. Fungal communities along a small-scale elevational gradient in an alpine tundra are determined by soil carbon nitrogen ratios. Front. Microbiol. 2018, 9, 1815. [Google Scholar] [CrossRef] [PubMed]
- Veres, A.; Petit, S.; Conord, C.; Lavigne, C. Does landscape composition affect pest abundance and their control by natural enemies? A review. Agric. Ecosyst. Environ. 2013, 166, 110–117. [Google Scholar] [CrossRef]
- Franco, F.P.; Moura, D.S.; Vivanco, J.M.; Silva-Filho, M.C. Plant-insect-pathogen interactions: a naturally complex ménage à trois. Curr. Opin. Microbiol. 2017, 37, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Tkaczuk, C.; Majchrowska-Safaryan, A. Temperature requirements for the colony growth and conidial germination of selected isolates of entomopathogenic fungi of the Cordyceps and Paecilomyces genera. Agriculture 2023, 13, 1989. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, H.; Dong, X.; Xu, S.; Cai, Q. Species diversity and distribution characteristics of entomogenous fungi in Zipeng Mountain National Forest Park. Chin. J. Biol. Control 2018, 34, 568. [Google Scholar]
- Dong, T.; Zhang, B.; Jiang, Y.; Hu, Q. Isolation and classification of fungal whitefly entomopathogens from soils of Qinghai-Tibet Plateau and Gansu Corridor in China. PLoS One 2016, 11, e0156087. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Chenu, C.; Kappler, A.; Rillig, M.C.; Fierer, N. The interplay between microbial communities and soil properties. Nat. Rev. Microbiol. 2024, 22, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Pozo, M.J.; Zabalgogeazcoa, I.; Vázquez de Aldana, B.R.; Martínez-Medina, A. Untapping the potential of plant mycobiomes for applications in agriculture. Curr. Opin. Plant Biol. 2021, 60, 102034. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.; Wang, J.; Zhang, X.; Wang, X.; Li, Y.; Zhang, Y. Soil pH and nutrients shape the vertical distribution of microbial communities in an alpine wetland. Sci. Total Environ. 2021, 774, 145780. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.; Li, Y.; Zhang, Y.; Chen, Y. Assessing the structure and diversity of fungal community in plant soil under different climatic and vegetation conditions. Front. Microbiol. 2023, 14, 1288066. [Google Scholar] [CrossRef] [PubMed]
- Labouyrie, M.; Routh, D.; Taudière, A.; Cébron, A.; Turpault, M.P.; Uroz, S. Patterns in soil microbial diversity across Europe. Nat. Commun. 2023, 14, 3311. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yang, Z.; He, K.; Zhou, W.; Feng, W. Soil fungal community diversity, co-occurrence networks, and assembly processes under diverse forest ecosystems. Microorganisms 2024, 12, 1915. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Chen, Q.; Wang, S.; Li, Y.; Zhang, Y. Soil microbial distribution depends on different types of landscape vegetation in temperate urban forest ecosystems. Front. Ecol. Evol. 2022, 10, 858254. [Google Scholar] [CrossRef]
- Yuan, Y.; Li, Y.; Wang, X.; Zhang, Y.; Chen, Y. Differences in soil microbial communities across soil types in China’s temperate forests. Forests 2024, 15, 1110. [Google Scholar] [CrossRef]
- Fei, G.X.; Wang, Y.; Li, Y.; Zhang, Y.; Chen, Y. A new entomopathogenic fungus, Ophiocordyceps xifengensis sp. nov., from Liaoning, China. Phytotaxa 2024, 642, 211–224. [Google Scholar] [CrossRef]
- Goldmann, K.; Schöning, I.; Buscot, F.; Wubet, T. Forest management type influences diversity and community composition of soil fungi across temperate forest ecosystems. Front. Microbiol. 2015, 6, 1300. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Islam, W.; Gang, L.; Chen, H.Y. Advanced research tools for fungal diversity and its impact on forest ecosystem. Environ. Sci. Pollut. Res. 2022, 29, 45044–45062. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, M.A.; Wakil, W.; Arif, M.J.; Sahi, S.T.; Dunlap, C.A. Diversity and correlation of entomopathogenic and associated fungi with soil factors. J. King Saud Univ. Sci. 2021, 33, 101520. [Google Scholar] [CrossRef]
- Araújo, J.P.M.; Hughes, D.P. Diversity of entomopathogenic fungi: which groups conquered the insect body? Adv. Genet. 2016, 94, 1–39. [Google Scholar] [PubMed]
- Vukicevich, E.; Lowery, D.T.; Bennett, J.A.; Hart, M. Influence of groundcover vegetation, soil physicochemical properties, and irrigation practices on soil fungi in semi-arid vineyards. Front. Ecol. Evol. 2019, 7, 118. [Google Scholar] [CrossRef]
- Sadyś, M.; Kennedy, R.; West, J.S. Potential impact of climate change on fungal distributions: analysis of 2 years of contrasting weather in the UK. Aerobiologia 2016, 32, 127–137. [Google Scholar] [CrossRef]
- Agbessenou, A.; Tounou, A.K.; Dannon, E.A.; Agboton, C.; Srinivasan, R.; Pittendrigh, B.R.; Tamò, M. Temperature-dependent modelling and spatial prediction reveal suitable geographical areas for deployment of two Metarhizium anisopliae isolates for Tuta absoluta management. Sci. Rep. 2021, 11, 23346. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.P.; Sun, M.H.; Wang, Y.; Liu, X.Z.; Chen, S.Y. Development of biphasic culture system for an entomopathogenic fungus Beauveria bassiana PfBb strain and its virulence on a defoliating moth Phauda flammans (Walker). J. Fungi 2025, 11, 202. [Google Scholar] [CrossRef] [PubMed]
- Yeo, H.; Pell, J.K.; Alderson, P.G.; Clark, S.J.; Pye, B.J. Laboratory evaluation of temperature effects on the germination and growth of entomopathogenic fungi and on their pathogenicity to two aphid species. Pest Manag. Sci. 2003, 59, 156–165. [Google Scholar] [CrossRef]
- Moore, G.E. Pathogenicity of three entomopathogenic fungi to the southern pine beetle at various temperatures and humidities. Environ. Entomol. 1973, 2, 54–57. [Google Scholar] [CrossRef]
- Mishra, S.; Kumar, P.; Malik, A. Effect of temperature and humidity on pathogenicity of native Beauveria bassiana isolate against Musca domestica L. J. Parasit. Dis. 2015, 39, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Michereff-Filho, M.; Rezende, J.M.; Venzon, M.; Gondim, M.G.C. The effect of spider mite-pathogenic strains of Beauveria bassiana and humidity on the survival and feeding behavior of Neoseiulus predatory mite species. Biol. Control 2022, 176, 105083. [Google Scholar] [CrossRef]
- Hu, R.; Bai, P.; Liu, B.; Yu, J. On the virulence of two Beauveria bassiana strains against the fall webworm, Hyphantria cunea (Durry) (Lepidoptera: Erebidae), larvae and their biological properties in relation to different abiotic factors. Egypt. J. Biol. Pest Control 2021, 31, 1–7. [Google Scholar] [CrossRef]
- Maranga, R.O.; Kaaya, G.P.; Mueke, J.M.; Hassanali, A. Effects of combining the fungi Beauveria bassiana and Metarhizium anisopliae on the mortality of the tick Amblyomma variegatum (Ixodidae) in relation to seasonal changes. Mycopathologia 2005, 159, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Duan, M.; Yu, Z. Agricultural landscapes and biodiversity in China. Agric. Ecosyst. Environ. 2013, 166, 46–54. [Google Scholar] [CrossRef]
- Estrada-Carmona, N.; Sánchez, A.C.; Remans, R.; Jones, S.K. Complex agricultural landscapes host more biodiversity than simple ones: A global meta-analysis. Proc. Natl. Acad. Sci. USA 2022, 119, e2203385119. [Google Scholar] [CrossRef] [PubMed]
- Bartual, A.M.; Potts, S.G.; Woodcock, B.A.; Dicks, L.V. The potential of different semi-natural habitats to sustain pollinators and natural enemies in European agricultural landscapes. Agric. Ecosyst. Environ. 2019, 279, 43–52. [Google Scholar] [CrossRef]
- Klinnert, A.; Krimmer, E.; Martin, E.A.; Scherber, C.; Steffan-Dewenter, I.; Tscharntke, T. Landscape features support natural pest control and farm income when pesticide application is reduced. Nat. Commun. 2024, 15, 5384. [Google Scholar] [CrossRef] [PubMed]
- Meyling, N.V.; Eilenberg, J. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: potential for conservation biological control. Biol. Control 2007, 43, 145–155. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Aime, M.C.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control 2008, 46, 72–82. [Google Scholar] [CrossRef]
- De Fine Licht, H.H.; Jensen, A.B.; Eilenberg, J. Insect hosts are nutritional landscapes navigated by fungal pathogens. Ecology 2025, 106, e70015. [Google Scholar] [CrossRef] [PubMed]
- Murindangabo, Y.T.; Yobo, K.S.; Laing, M.D. Relevance of entomopathogenic fungi in soil-plant systems. Plant Soil 2024, 495, 287–310. [Google Scholar] [CrossRef]
- Hallsworth, J.E.; Magan, N. Water and temperature relations of growth of the entomogenous fungi Beauveria bassiana, Metarhizium anisopliae, and Paecilomyces farinosus. J. Invertebr. Pathol. 1999, 74, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, R.; Gallery, R.E.; Gripenberg, S.; Gurr, S.J.; Narayan, L.; Addis, C.E.; Freckleton, R.P.; Lewis, O.T. Pathogens and insect herbivores drive rainforest plant diversity and composition. Nature 2014, 506, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, Y.; Wang, X.; Zhang, Y.; Chen, Y. Enhancing soil health and nutrient cycling through soil amendments: Improving the synergy of bacteria and fungi. Sci. Total Environ. 2024, 923, 171332. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Liu, J.; Wang, K.; Li, Y.; Chen, Y. Distributional patterns of soil nematodes in relation to environmental variables in forest ecosystems. Soil Ecol. Lett. 2021, 3, 115–124. [Google Scholar] [CrossRef]
- Kagimu, N.; Ferreira, T.; Malan, A.P. The attributes of survival in the formulation of entomopathogenic nematodes utilised as insect biocontrol agents. Afr. Entomol. 2017, 25, 275–291. [Google Scholar] [CrossRef]
- Tkaczuk, C.; Król, A.; Majchrowska-Safaryan, A.; Nicewicz, Ł. The occurrence of entomopathogenic fungi in soils from fields cultivated in conventional and organic systems. J. Ecol. Eng. 2014, 15, 137–144. [Google Scholar]
- Fernández-Bravo, M.; Garrido-Jurado, I.; Valverde-García, P.; Quesada-Moraga, E. Land-use type drives soil population structures of the entomopathogenic fungal genus Metarhizium. Microorganisms 2021, 9, 1380. [Google Scholar] [CrossRef] [PubMed]
- Tscharntke, T.; Steffan-Dewenter, I.; Kruess, A.; Thies, C. Contribution of small habitat fragments to conservation of insect communities of grassland-cropland landscapes. Ecol. Appl. 2002, 12, 354–363. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Booij, C.J.H.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Duru, M.; Therond, O.; Martin, G.; Martin-Clouaire, R.; Magne, M.A.; Justes, E.; Journet, E.P.; Aubertot, J.N.; Savary, S.; Bergez, J.E.; et al. How to implement biodiversity-based agriculture to enhance ecosystem services: a review. Agron. Sustain. Dev. 2015, 35, 1259–1281. [Google Scholar] [CrossRef]
- Kannan, M.; Opatovsky, I. A review on nutritional and non-nutritional interactions of symbiotic and associated fungi with insects. Symbiosis 2023, 91, 139–149. [Google Scholar] [CrossRef]
- Barcoto, M.O.; de Souza, D.J.; de Queiroz, A.C.; de Azevedo, R.S.; de Paula, M.S.; Pupo, M.T. Fungus-growing insects host a distinctive microbiota apparently adapted to the fungiculture environment. Sci. Rep. 2020, 10, 12384. [Google Scholar] [CrossRef] [PubMed]
- Aung, O.M.; Soytong, K.; Hyde, K.D. Diversity of entomopathogenic fungi in rainforests of Chiang Mai Province, Thailand. Fungal Divers. 2008, 30, 15–22. [Google Scholar]
- Gindin, G.; Samish, M.; Zangi, G.; Mishoutchenko, A.; Glazer, I. The susceptibility of different species and stages of ticks to entomopathogenic fungi. Exp. Appl. Acarol. 2002, 28, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Samish, M.; Gindin, G.; Alekseev, E.; Glazer, I. Pathogenicity of entomopathogenic fungi to different developmental stages of Rhipicephalus sanguineus (Acari: Ixodidae). J. Parasitol. 2001, 87, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.M.; Xie, W.; Zhi, J.R.; Zou, X. Frankliniella occidentalis pathogenic fungus Lecanicillium interacts with internal microbes and produces sublethal effects. Pestic. Biochem. Physiol. 2021, 97, 105679. [Google Scholar] [CrossRef] [PubMed]
- Soth, S.; Lacey, L.A.; Mercadier, G. You are what you eat: fungal metabolites and host plant affect the susceptibility of diamondback moth to entomopathogenic fungi. PeerJ 2022, 10, e14491. [Google Scholar] [CrossRef] [PubMed]
- Mäder, P.; Edenhofer, S.; Boller, T.; Wiemken, A.; Niggli, U. Arbuscular mycorrhizae in a long-term field trial comparing low-input (organic, biological) and high-input (conventional) farming systems in a crop rotation. Biol. Fertil. Soils 2000, 31, 150–156. [Google Scholar] [CrossRef]
- Clifton, E.H.; Jaronski, S.T.; Hodgson, E.W.; Gassmann, A.J. Abundance of soil-borne entomopathogenic fungi in organic and conventional fields in the Midwestern USA with an emphasis on the effect of herbicides and fungicides on fungal persistence. PLoS One 2015, 10, e0133613. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qu, Z.; Zhang, Y.; Ge, Y.; Sun, H. Soil fungal community and potential function in different forest ecosystems. Diversity 2022, 14, 520. [Google Scholar] [CrossRef]
- Ramos, Y.; Portal, O.; Lysøe, E.; Meyling, N.V.; Klingen, I. Diversity and abundance of Beauveria bassiana in soils, stink bugs and plant tissues of common bean from organic and conventional fields. J. Invertebr. Pathol. 2017, 150, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Klingen, I.; Eilenberg, J.; Meadow, R. Effects of farming system, field margins and bait insect on the occurrence of insect pathogenic fungi in soils. Agric. Ecosyst. Environ. 2002, 91, 191–198. [Google Scholar] [CrossRef]
- Perazzolli, M.; Antonielli, L.; Storari, M.; Puopolo, G.; Pancher, M.; Giovannini, O.; Pertot, I. Resilience of the natural phyllosphere microbiota of the grapevine to chemical and biological pesticides. Appl. Environ. Microbiol. 2014, 80, 3585–3596. [Google Scholar] [CrossRef] [PubMed]
- Goble, T.A.; Dames, J.F.P.; Hill, M.; Moore, S.D. The effects of farming system, habitat type and bait type on the isolation of entomopathogenic fungi from citrus soils in the Eastern Cape Province, South Africa. BioControl 2010, 55, 399–412. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, M.; Ma, K.; Li, X.; Zhang, Y. Contrasting floristic diversity of the Hengduan Mountains, the Himalayas and the Qinghai-Tibet Plateau sensu stricto in China. Front. Ecol. Evol. 2020, 8, 136. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Wang, X.; Chen, Y.; Li, J. Diversity and correlation analysis of endophytes and top metabolites in Phlomoides rotata roots from high-altitude habitats. Microorganisms 2025, 13, 503. [Google Scholar] [CrossRef] [PubMed]
- Mao, K.S.; Wang, Y.; Liu, J.Q. Evolutionary origin of species diversity on the Qinghai-Tibet Plateau. J. Syst. Evol. 2021, 59, 1142–1158. [Google Scholar] [CrossRef]
- Inglis, G.D.; Duke, G.M.; Goettel, M.S.; Kabaluk, J.T.; Ortega-Polo, R. Biogeography and genotypic diversity of Metarhizium brunneum and Metarhizium robertsii in northwestern North American soils. Can. J. Microbiol. 2019, 65, 261–281. [Google Scholar] [CrossRef] [PubMed]
- Crowther, T.W.; Maynard, D.S.; Crowther, T.R.; Peccia, J.; Smith, D.P.; Bradford, M.A. Untangling the fungal niche: the trait-based approach. Front. Microbiol. 2014, 5, 579. [Google Scholar] [CrossRef] [PubMed]
- Abrego, N.; Ovaskainen, O.; Somervuo, P.; Huotari, T.; Norberg, A.; Nyman, J.; Roslin, T. Airborne DNA reveals predictable spatial and seasonal dynamics of fungi. Nature 2024, 631, 835–842. [Google Scholar] [CrossRef]
- Chaloner, T.M.; Gurr, S.J.; Bebber, D.P. Geometry and evolution of the ecological niche in plant-associated microbes. Nat. Commun. 2020, 11, 2955. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Urquiza, A.; Keyhani, N.O. Molecular genetics of Beauveria bassiana infection of insects. Adv. Genet. 2016, 94, 165–249. [Google Scholar] [PubMed]
- Rohlfs, C.; Trenkamp, M.; Pötsch, L.; Steinmetz, H.; Dahse, H.M.; Hertweck, C. Variation in physiological host range in three strains of two species of the entomopathogenic fungus Beauveria. PLoS One 2018, 13, e0199199. [Google Scholar]
- McGuire, A.V.; Northfield, T.D. Tropical occurrence and agricultural importance of Beauveria bassiana and Metarhizium anisopliae. Front. Sustain. Food Syst. 2020, 4, 6. [Google Scholar] [CrossRef]
- Hurst, C.J. Dirt and disease: the ecology of soil fungi and plant fungi that are infectious for vertebrates. In Understanding Terrestrial Microbial Communities; Springer: Cham, Switzerland, 2019; pp. 289–405. [Google Scholar]
- Masiulionis, V.E.; Samuels, R.I. Investigating the biology of leaf-cutting ants to support the development of alternative methods for the control and management of these agricultural pests. Agriculture 2025, 15, 642. [Google Scholar] [CrossRef]
- Xu, J.N.; Feng, G.M. The time-dose-mortality modeling and virulence indices for the two entomophthoralean species Pandora delphacis and P. neoaphidis against the green peach aphid Myzus persicae. Biol. Control 2000, 17, 29–34. [Google Scholar] [CrossRef]
- Holder, D.J.; Keyhani, N.O. Adhesion of the entomopathogenic fungus Beauveria (Cordyceps) bassiana to substrata. Appl. Environ. Microbiol. 2005, 71, 5260–5266. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yao, Y.; Xu, B.; Li, Y.; Chen, J.; Zhang, X.; Wang, C. Infection process and genome assembly provide insights into the pathogenic mechanism of destructive mycoparasite Calcarisporium cordycipiticola with host specificity. J. Fungi 2021, 7, 918. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; He, K.; Zhang, X.; Wang, C. An entomopathogenic fungus exploits its host humoral antibacterial immunity to minimize bacterial competition in the hemolymph. Microbiome 2023, 11, 116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, X.; Wang, C.; Wang, S. Cross-talk between immunity and behavior: insights from entomopathogenic fungi and their insect hosts. FEMS Microbiol. Rev. 2024, 48, fuae003. [Google Scholar] [CrossRef] [PubMed]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies. Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Meyling, N.V.; Eilenberg, J. Isolation and characterisation of Beauveria bassiana isolates from phylloplanes of hedgerow vegetation. Mycol. Res. 2006, 110, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, X.; Wang, C.; Wang, S. Entomopathogenic fungal-derived metabolites alter innate immunity and gut microbiota in the migratory locust. J. Pest Sci. 2024, 97, 853–872. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Liu, X.; Chen, X.; Wang, C. An iron-binding protein of entomopathogenic fungus suppresses the proliferation of host symbiotic bacteria. Microbiome 2024, 12, 202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wei, X.; Fang, W.; Huang, X.; Zhang, X. The secretory protein COA1 enables Metarhizium robertsii to evade insect immune recognition during cuticle penetration. Commun. Biol. 2024, 7, 1220. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Chávez, M.J.; Pérez-García, L.A.; Niño-Vega, G.A.; Mora-Montes, H.M. Fungal strategies to evade the host immune recognition. J. Fungi 2017, 3, 51. [Google Scholar] [CrossRef] [PubMed]
- Vidhate, R.P.; Dawkar, V.V.; Punekar, S.A.; Giri, A.P. Genomic determinants of entomopathogenic fungi and their involvement in pathogenesis. Microb. Ecol. 2023, 85, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Unuofin, J.O.; Okaiyeto, K.; Oluwaseun, O.; Oguntibeju, O.O.; Nwonuma, C.O.; Lebelo, S.L. Chitinases: expanding the boundaries of knowledge beyond routinized chitin degradation. Environ. Sci. Pollut. Res. 2024, 31, 38045–38060. [Google Scholar] [CrossRef] [PubMed]
- Rohlfs, M.; Churchill, A.C.L. Fungal secondary metabolites as modulators of interactions with insects and other arthropods. Fungal Genet. Biol. 2011, 48, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Orozco, R.; Wijeratne, E.M.K.; Gunatilaka, A.A.L.; Stock, S.P.; Molnár, I. Biosynthesis of the cyclooligomer depsipeptide beauvericin, a virulence factor of the entomopathogenic fungus Beauveria bassiana. Chem. Biol. 2008, 15, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, Y.; Liu, X.; Wang, C. A secreted effector with a dual role as a toxin and as a transcriptional factor. Nat. Commun. 2022, 13, 7779. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; St Leger, R.J. The Metarhizium anisopliae perilipin homolog MPL1 regulates lipid metabolism, appressorial turgor pressure, and virulence. J. Biol. Chem. 2007, 282, 21110–21115. [Google Scholar] [CrossRef] [PubMed]
- Becklin, S.; Jin, Y.; Rebarber, R.; Tenhumberg, B. Spatial dynamics of a pest population with stage-structure and control. J. Math. Biol. 2025, 90, 48. [Google Scholar] [CrossRef] [PubMed]
- Serva, D.; Krofel, M.; Cerasoli, F.; Biondi, M.; Iannella, M. Supporting reintroduction planning: a framework integrating habitat suitability, connectivity and individual-based modelling. A case study with the Eurasian lynx in the Apennines. Divers. Distrib. 2025, 31, e70024. [Google Scholar] [CrossRef]
- Qu, J.; Yu, L.Q.; Zhang, J.; Han, Y.; Zou, X. A new entomopathogenic fungus, Ophiocordyceps ponerus sp. nov., from China. Phytotaxa 2018, 343, 116–126. [Google Scholar] [CrossRef]
- Lacey, J. Spore dispersal—its role in ecology and disease, the British contribution to fungal aerobiology. Mycol. Res. 1996, 100, 641–660. [Google Scholar] [CrossRef]
- Custer, G. F. , Bresciani, L., Dini-Andreote, F. Ecological and Evolutionary Implications of Microbial Dispersal. Front. Microbiol. 2022, 13, 855859. [Google Scholar] [CrossRef] [PubMed]
- Trail, F. , Gaffoor, I., Vogel, S. Ejection Mechanics and Trajectory of the Ascospores of Gibberella zeae (Anamorph Fusarium graminearum). Fungal Genet. Biol. 2005, 42, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Snyder, R. E. , Chesson, P. Local Dispersal Can Facilitate Coexistence in the Presence of Permanent Spatial Heterogeneity. Ecol. Lett. 2010, 6, 301–309. [Google Scholar] [CrossRef]
- Johansson, V. , Ranius, T., Snäll, T. Epiphyte Metapopulation Dynamics Are Explained by Species Traits, Connectivity, and Patch Dynamics. Ecology 2012, 93, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Udayababu, P. , Sunil, Z. Evaluation of Entomopathogenic Fungi for the Management of Tobacco Caterpillar Spodoptera litura (Fabricius). Indian J. Plant Prot. 2012, 40, 214–220. [Google Scholar]
- Muok, A. R. , Claessen, D., Briegel, A. Microbial Hitchhiking: How Streptomyces Spores Are Transported by Motile Soil Bacteria. ISME J. 2021, 15, 2591–2600. [Google Scholar] [CrossRef] [PubMed]
- Golan, J. J. , Pringle, A. Long-Distance Dispersal of Fungi. Microbiol. Spectr. 2017, 5, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Nitayavardhana, S. , Rakshit, S. K., Grewell, D., Van Leeuwen, J., Khanal, S. K. Ultrasound Improved Ethanol Fermentation from Cassava Chips in Cassava-Based Ethanol Plants. Bioresour. Technol. 2010, 101, 2741–2747. [Google Scholar] [CrossRef] [PubMed]
- Santi, L. , Silva, W. O. B., Silva, C. P., Beys da Silva, W. O., Schrank, A., Vainstein, M. H. The Entomopathogen Metarhizium anisopliae Can Modulate the Secretion of Lipolytic Enzymes in Response to Different Substrates Including Components of Arthropod Cuticle. Fungal Biol. 2010, 114, 911–916. [Google Scholar]
- Lopes, F. C. , Martinelli, A. H., John, E. B., Ligabue-Braun, R. Microbial Hydrolytic Enzymes: Powerful Weapons Against Insect Pests. Microbes for Sustainable Insect Pest Management: Hydrolytic Enzyme; Secondary Metabolite 2021, 2, 1–31. [Google Scholar]
- Amiri-Besheli, B. , Khambay, B., Cameron, S., Deadman, M. L., Butt, T. M. Inter- and Intra-Specific Variation in Destruxin Production by Insect Pathogenic Metarhizium spp. and Its Significance to Pathogenesis. Mycol. Res. 2000, 104, 447–452. [Google Scholar] [CrossRef]
- Chen, X. R. , Hu, Q. B., Yu, X. Q., Ren, S. X. Effects of Destruxins on Free Calcium and Hydrogen Ions in Insect Hemocytes. Insect Sci. 2014, 21, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Zdybicka-Barabas, A. , Stączek, S., Kunat-Budzyńska, M., Cytryńska, M. Innate Immunity in Insects: The Lights and Shadows of Phenoloxidase System Activation. Int. J. Mol. Sci. 2025, 26, 1320. [Google Scholar] [CrossRef] [PubMed]
- Qu, J. , Feng, Y., Zou, X., Zhou, Y., Cao, W. Transcriptome and Proteome Analyses Reveal Genes and Signaling Pathways Involved in the Response to Two Insect Hormones in the Insect-Fungal Pathogen Hirsutella satumaensis. mSystems 2024, 9, e00166–24. [Google Scholar] [CrossRef] [PubMed]
- Ost, K. S. , Round, J. L. Commensal Fungi in Intestinal Health and Disease. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Tang, C. , Zhang, L., Li, Y., Wang, Y., Li, J., Zhang, X. The Symbiont Acinetobacter baumannii Enhances the Insect Host Resistance to Entomopathogenic Fungus Metarhizium anisopliae. Commun. Biol. 2024, 7, 1184. [Google Scholar] [CrossRef] [PubMed]
- Wei, G. , Lai, Y., Wang, G., Chen, X., Li, F., Wang, S., Jin, D., Peng, Y., Lin, Z. Insect Pathogenic Fungus Interacts with the Gut Microbiota to Accelerate Mosquito Mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef] [PubMed]
- Qu, J. , Zou, X., Yu, J., Zhou, Y. The Conidial Mucilage, Natural Film Coatings, Is Involved in Environmental Adaptability and Pathogenicity of Hirsutella satumaensis Aoki. Sci. Rep. 2017, 7, 1301. [Google Scholar] [CrossRef] [PubMed]
- Kong, H. , Wang, C., Zhang, S., Lin, Z., Wang, L., Luo, L. Altered Immunity in Crowded Mythimna separata Is Mediated by Octopamine and Dopamine. Sci. Rep. 2018, 8, 3215. [Google Scholar] [CrossRef] [PubMed]
- Franco, F. P. , Fávero, A. C., de Castro, É. C., de Moraes, L. A., Rodrigues, F. Å., Lourenção, A. L., Furtado, E. L., Quecine, M. C., Azevedo, J. L., Pizzirani-Kleiner, A. A. Fungal Phytopathogen Modulates Plant and Insect Responses to Promote Its Dissemination. ISME J. 2021, 15, 3522–3533. [Google Scholar] [CrossRef] [PubMed]
- Ai, S. , Wang, M., Li, H., Guo, X., Zhang, Y., Chen, X. Insect-Microorganism Interaction Has Implicates on Insect Olfactory Systems. Insects 2022, 13, 1094. [Google Scholar] [CrossRef] [PubMed]
- Hong, S. , Sun, Y., Chen, H., Wang, C. Suppression of the Insect Cuticular Microbiomes by a Fungal Defensin to Facilitate Parasite Infection. ISME J. 2023, 17, 1. [Google Scholar] [CrossRef] [PubMed]
- Abbot, P. Defense in Social Insects, Diversity, Division of Labor, and Evolution. Annu. Rev. Entomol. 2022, 67, 407–436. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y. , Nakamura, Y., Yoshida, M., Sakamoto, K., Matsumoto, S., Imanishi, S., Yamakawa, M. Comparative Analysis of Seven Types of Superoxide Dismutases for Their Ability to Respond to Oxidative Stress in Bombyx mori. Sci. Rep. 2019, 9, 2170. [Google Scholar]
- Zhang, C. , Wang, L., Li, Y., Wang, M., Yang, Y., Wang, Y., Zhang, X., Cuthbertson, A. G. S., Qiu, B. Impact of Beauveria bassiana on Antioxidant Enzyme Activities and Metabolomic Profiles of Spodoptera frugiperda. J. Invertebr. Pathol. 2023, 198, 107929. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A. , Nair, S. Epigenetic Processes in Insect Adaptation to Environmental Stress. Curr. Opin. Insect Sci. 2024, 7, 101294. [Google Scholar]
- Heidel-Fischer, H. M. , Vogel, H. Molecular Mechanisms of Insect Adaptation to Plant Secondary Compounds. Curr. Opin. Insect Sci. 2015, 8, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q. , Ren, M., Liu, X., Xia, H., Chen, K. Peptidoglycan Recognition Proteins in Insect Immunity. Mol. Immunol. 2019, 106, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Shahrestani, P. , Müller, P., Wertheim, B., van Doorn, G. S. Sexual Dimorphism in Drosophila melanogaster Survival of Beauveria bassiana Infection Depends on Core Immune Signaling. Sci. Rep. 2018, 8, 12501. [Google Scholar] [CrossRef] [PubMed]
- Jin, T. , Chang, X., Sen, S., Ye, G. Downregulation and Mutation of a Cadherin Gene Associated with Cry1Ac Resistance in the Asian Corn Borer, Ostrinia furnacalis (Guenée). Toxins 2014, 6, 2676–2693. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, M. A. , Wakil, W., Arif, M. J., Sahi, S. T., Dunlap, C. A. Infection of Helicoverpa armigera by Endophytic Beauveria bassiana Colonizing Tomato Plants. Biol. Control 2015, 90, 200–207. [Google Scholar] [CrossRef]
- Bamisile, B. S. , Dash, C. K., Akutse, K. S., Keppanan, R., Wang, L. Fungal Endophytes: Beyond Herbivore Management. Front. Microbiol. 2018, 9, 544. [Google Scholar] [CrossRef] [PubMed]
- Wakil, W. , Tahir, M., Al-Sadi, A. M., Shapiro-Ilan, D. Interactions between Two Invertebrate Pathogens: An Endophytic Fungus and an Externally Applied Bacterium. Front. Microbiol. 2020, 11, 522368. [Google Scholar] [CrossRef] [PubMed]
- Guo, W. , Zhang, J., Zhang, X., Li, C., Wang, L., Li, D., Li, T., Wang, M. Endophytic Colonization of Beauveria bassiana Enhances Drought Stress Tolerance in Tomato via “Water Spender” Pathway. Int. J. Mol. Sci. 2024, 25, 11949. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. J. , Goettel, M. S., Gillespie, D. R. Evaluation of Lecanicillium longisporum, Vertalec® for Simultaneous Suppression of Cotton Aphid, Aphis gossypii, and Cucumber Powdery Mildew, Sphaerotheca fuliginea, on Potted Cucumbers. Biol. Control 2008, 45, 404–409. [Google Scholar] [CrossRef]
- Jaber, L. R. , Ownley, B. H. Can We Use Entomopathogenic Fungi as Endophytes for Dual Biological Control of Insect Pests and Plant Pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Waqar, S. , Bhat, A. A., Khan, A. A. Endophytic Fungi: Unravelling Plant-Endophyte Interaction and the Multifaceted Role of Fungal Endophytes in Stress Amelioration. Plant Physiol. Biochem. 2024, 206, 108174. [Google Scholar] [CrossRef] [PubMed]
- Bamisile, B. S. , Akutse, K. S., Siddiqui, J. A., Xu, Y. Model Application of Entomopathogenic Fungi as Alternatives to Chemical Pesticides: Prospects, Challenges, and Insights for Next-Generation Sustainable Agriculture. Front. Plant Sci. 2021, 12, 741804. [Google Scholar] [CrossRef] [PubMed]
- Leger, R. J. The Evolution of Complex Metarhizium-Insect-Plant Interactions. Fungal Biol. 2024, 128, 2513–2528. [Google Scholar] [CrossRef] [PubMed]
- Alquichire-Rojas, S. , Escobar, E., Bascuñán-Godoy, L., González-Teuber, M. Root Symbiotic Fungi Improve Nitrogen Transfer and Morpho-Physiological Performance in Chenopodium quinoa. Front. Plant Sci. 2024, 15, 1386234. [Google Scholar]
- Liu, Y. , Yang, Y., Wang, B. Entomopathogenic Fungi Beauveria bassiana and Metarhizium anisopliae Play Roles of Maize (Zea mays) Growth Promoter. Sci. Rep. 2022, 12, 15706. [Google Scholar]
- Imoulan, A. , El Meziane, A., Alaoui, A., El Aouami, A., El Khourassi, A., El Harti, A., El Alaoui, A. Entomopathogenic Fungus Beauveria: Host Specificity, Ecology and Significance of Morpho-Molecular Characterization in Accurate Taxonomic Classification. J. Asia-Pac. Entomol. 2017, 20, 1204–1212. [Google Scholar] [CrossRef]
- Barelli, L. , Moonjely, S., Behie, S. W., Bidochka, M. J. Fungi with Multifunctional Lifestyles: Endophytic Insect Pathogenic Fungi. Plant Mol. Biol. 2016, 90, 657–664. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, A. C. , Saari, S., Moran-Diez, M. E., Meyling, N. V., Raad, M., Glare, T. R. Beauveria bassiana as an Endophyte: A Critical Review on Associated Methodology and Biocontrol Potential. BioControl 2017, 62, 1–7. [Google Scholar]
- Sui, L. , Li, Y., Li, J., Zhang, X., Wang, C. Endophytic Beauveria bassiana Promotes Plant Biomass Growth and Suppresses Pathogen Damage by Directional Recruitment. Front. Microbiol. 2023, 14, 1227269. [Google Scholar] [CrossRef] [PubMed]
- Noman, A. , Aqeel, M., Qasim, M., Haider, I., Lou, Y. Plant-Insect-Microbe Interaction: A Love Triangle between Enemies in Ecosystem. Sci. Total Environ. 2020, 699, 134181. [Google Scholar] [CrossRef] [PubMed]
- Feng, M. , Chen, Q., Guo, Q., Liu, B., Ying, S. Assessment of Beauveria bassiana for the Biological Control of Corn Borer, Ostrinia furnacalis, in Sweet Maize by Irrigation Application. BioControl 2023, 68, 49–60. [Google Scholar] [CrossRef]
- Kuzhuppillymyal-Prabhakarankutty, L. , Rajendran, G., Karthikeyan, G., Senthil-Nathan, S., Al-Dhabi, N. A., Arasu, M. V., Duraipandiyan, V., Samiyappan, R. Endophytic Beauveria bassiana Promotes Drought Tolerance and Early Flowering in Corn. World J. Microbiol. Biotechnol. 2020, 36, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Eilenberg, J. , Hajek, A., Lomer, C. Suggestions for Unifying the Terminology in Biological Control. BioControl 2001, 46, 387–400. [Google Scholar] [CrossRef]
- He, D. C. , Zhan, J. S., Xie, L. H. Problems, Challenges and Future of Plant Disease Management: From an Ecological Point of View. J. Integr. Agric. 2016, 15, 705–715. [Google Scholar] [CrossRef]
- Wang, G. , Li, Y., Zhang, Y., Dong, S., Chen, S., Wang, M., Sun, C., Khashnobish, A., Li, Y., Wang, C. Biosynthesis of Antibiotic Leucinostatins in Bio-Control Fungus Purpureocillium lilacinum and Their Inhibition on Phytophthora Revealed by Genome Mining. PLoS Pathog. 2016, 12, e1005685. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F. M. , Silva, M., Molina-Cruz, A., Barillas-Mury, C. Molecular Mechanisms of Insect Immune Memory and Pathogen Transmission. PLoS Pathog. 2022, 18, e1010939. [Google Scholar] [CrossRef] [PubMed]
- De Ligne, L. , Van den Bulcke, J., Van Acker, J., De Baets, B. Analysis of Spatio-Temporal Fungal Growth Dynamics under Different Environmental Conditions. IMA Fungus 2019, 10, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L. , Fasoyin, O. E., Molnár, I., Xu, Y. Secondary Metabolites from Hypocrealean Entomopathogenic Fungi: Novel Bioactive Compounds. Nat. Prod. Rep. 2020, 37, 1181–1206. [Google Scholar] [CrossRef] [PubMed]
- Parveen, S. S. , Rashtrapal, P. S. Integrated Pest Management Strategies Using Endophytic Entomopathogenic Fungi. Plant Sci. Today 2024, 11, 568–574. [Google Scholar]
- Bartling, M. T. , Brandt, A., Hollert, H., Vilcinskas, A. Current Insights into Sublethal Effects of Pesticides on Insects. Int. J. Mol. Sci. 2024, 25, 6007. [Google Scholar] [CrossRef] [PubMed]
- Herrick, N. J. , Cloyd, R. A. Direct and Indirect Effects of Pesticides on the Insidious Flower Bug (Hemiptera: Anthocoridae) under Laboratory Conditions. J. Econ. Entomol. 2017, 110, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Moraga, E. , Landa, B. B., Muñoz-Ledesma, F., Santiago-Álvarez, C. Key Role of Environmental Competence in Successful Use of Entomopathogenic Fungi in Microbial Pest Control. J. Pest Sci. 2024, 97, 1–5. [Google Scholar] [CrossRef]
- Lehmann, C.; Schmidt, B.; Stephan, D.; Meyer, V. Investigation of the interface of fungal mycelium composite building materials by means of low-vacuum scanning electron microscopy. J. Microsc. 2024, 294, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Warmink, J.A.; Elsas, J.D. van. Migratory response of soil bacteria to Lyophyllum sp. strain Karsten in soil microcosms. Appl. Environ. Microbiol. 2009, 75, 2820–2830. [Google Scholar] [CrossRef] [PubMed]
- Furuno, S.; Remer, R.; Chatzinotas, A.; Harms, H.; Wick, L.Y. Use of mycelia as paths for the isolation of contaminant-degrading bacteria from soil. Microb. Biotechnol. 2012, 5, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Zhang, J.; Wang, D.; Zhang, T.; Yang, H.; Luo, Y.; Li, J.; Zhang, G.; Liu, X.; Zhang, Y.; Zheng, X.; Zhang, L.; Zhang, Y.; Zhang, X. Alternative transcription start site selection in Mr-OPY2 controls lifestyle transitions in the fungus Metarhizium robertsii. Nat. Commun. 2017, 8, 1565. [Google Scholar] [CrossRef] [PubMed]
- Cocking, E.C. Endophytic colonization of plant roots by nitrogen-fixing bacteria. Plant Soil 2003, 252, 169–175. [Google Scholar] [CrossRef]
- Behie, S.W.; Bidochka, M.J. Ubiquity of insect-derived nitrogen transfer to plants by endophytic insect-pathogenic fungi: an additional branch of the soil nitrogen cycle. Appl. Environ. Microbiol. 2014, 80, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Guilger-Casagrande, M.; Lima, R.; Andrade, G.; Ribeiro, A.; Campos, E.V.R.; Fraceto, L.F.; Oliveira, H.C.; de Oliveira, J.L. Beauveria bassiana biogenic nanoparticles for the control of Noctuidae pests. Pest Manag. Sci. 2024, 80, 1325–1337. [Google Scholar] [CrossRef] [PubMed]
- Sarathchandra, S.U.; Ghani, A.; Yeates, G.W.; Burch, G.; Cox, N.R. Effect of nitrogen and phosphate fertilisers on microbial and nematode diversity in pasture soils. Soil Biol. Biochem. 2001, 33, 953–964. [Google Scholar] [CrossRef]
- Gao, L.; Sun, M.H.; Liu, X.Z.; Che, Y.S. Effects of carbon concentration and carbon to nitrogen ratio on the growth and sporulation of several biocontrol fungi. Mycol. Res. 2007, 111, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Reingold, V.; Halperin, O.; Shalaby, S.; Dubov, D.; Freilich, S.; Zakin, V.; Golani, Y.; Dubey, A.; Prasad, R.; Bhat, B.N.; Rav-David, D.; Sharon, A.; Freeman, S. Transcriptional reprogramming in the entomopathogenic fungus Metarhizium brunneum and its aphid host Myzus persicae during the switch between saprophytic and parasitic lifestyles. BMC Genomics 2024, 25, 917. [Google Scholar] [PubMed]
- Gu, F.T.; Li, H.; Zhao, H.; Liu, Q.; Liu, H.; Zhang, C.; Zhang, H.; Zhang, J. Metabolic outcomes of Cordyceps fungus and Goji plant polysaccharides during in vitro human fecal fermentation. Carbohydr. Polym. 2025, 350, 123019. [Google Scholar] [CrossRef] [PubMed]
- Booi, H.N.; Zhang, T.; Li, L.; Zhang, Y.; Zhang, Y.; Zhang, X.; Zhang, L. Evidence to support cultivated fruiting body of Ophiocordyceps sinensis (Ascomycota)’s role in relaxing airway smooth muscle. J. Ethnopharmacol. 2025, 336, 118727. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Y.; Li, J.; Zhang, H.; Zhang, L.; Zhang, X.; Zhang, Y. Innovative mycelium-based food: Advancing one health through nutritional insights and environmental sustainability. Compr. Rev. Food Sci. Food Saf. 2025, 24, e70166. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Li, S. Cordyceps as an herbal drug for anti-aging and neurodegenerative diseases. Biomed. Pharmacother. 2021, 134, 111142. [Google Scholar]
- Wu, Z.; Zhang, Y.; Zhang, X.; Zhang, L.; Zhang, H.; Zhang, J.; Zhang, C. Agrocybe cylindracea polysaccharides repair the intestinal mucosal barrier in DSS-induced colitis by modulating the gut microbiota-macrophage axis. Food Sci. Hum. Wellness 2025, 15, in, in press. [Google Scholar] [CrossRef]
- Shao, Y.; Zhang, X.; Zhang, L.; Zhang, Y.; Zhang, H.; Zhang, J. Cordycepin affects Streptococcus mutans biofilm and interferes with its metabolism. BMC Oral Health 2025, 25, 25. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.M.; Nguyen, T.T.; Nguyen, T.H.; Nguyen, T.T.; Nguyen, T.T.; Nguyen, T.T. Low-glycemic index cookies supplemented with Cordyceps militaris substrate: nutritional values, physicochemical properties, antioxidant activity, bioactive constituents, and bioaccessibility. Food Chem. X 2025, 23, 102494. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.Q.; Van Pham, T.; Andriana, Y.; Truong, M.N. Cordyceps militaris-derived bioactive gels: therapeutic and anti-aging applications in dermatology. Gels 2025, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Devi, W.M.; Saha, H.; Irungbam, S.; Saha, R.K. A novel approach in valorization of spent mushroom substrate of Cordyceps militaris as in-feed antibiotics in Labeo rohita against Aeromonas hydrophila infection. Environ. Sci. Pollut. Res. 2024, 31, 62305–62314. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.M.; Zhang, X.; Zhang, L.; Zhang, H.; Zhang, J.; Zhang, C.; Zhang, Y. The composition and function of bacterial communities in Bombyx mori (Lepidoptera: Bombycidae) changed dramatically with infected fungi: a new potential to culture Cordyceps cicadae. Insect Mol. Biol. 2024, 33, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, Y.; Zhang, L.; Zhang, H.; Zhang, J.; Zhang, C. Fermentation design and process optimization strategy based on machine learning. BioDesign Res. 2025, 7, 100002. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A.K.; Singh, P.K.; Singh, V.; Singh, R.; Singh, R. Physio-biochemical insights of endophytic microbial community for crop stress resilience: an updated overview. J. Plant Growth Regul. 2025, 5, 1–24. [Google Scholar] [CrossRef]
- Vega, F.E.; Meyling, N.V.; Luangsa-ard, J.J.; Blackwell, M. Fungal entomopathogens: new insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef]
- Sparks, T.C.; Nauen, R. IRAC: Mode of action classification and insecticide resistance management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, X.; Zhang, L.; Zhang, H.; Zhang, J.; Zhang, C. Defense and inhibition integrated mesoporous nanoselenium delivery system against tomato gray mold. Environ. Sci. Nano 2020, 7, 210–227. [Google Scholar] [CrossRef]
- de Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: a comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Abdelaziz, A.M.; Hashem, A.H.; Abd-Elsalam, K.A.; Attia, M.S. Biodiversity of fungal endophytes. In Fungal Endophytes Volume I: Biodiversity and Bioactive Materials; Springer: Singapore, 2025; pp. 43–61. [Google Scholar]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: high-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Arias-Aravena, M.; Altimira, F.; Gutiérrez, D.; Ling, J.; Tapia, E. Identification of exoenzymes secreted by entomopathogenic fungus Beauveria pseudobassiana RGM 2184 and their effect on the degradation of cocoons and pupae of quarantine pest Lobesia botrana. J. Fungi 2022, 8, 1083. [Google Scholar] [CrossRef] [PubMed]
- Borsato, N.D.; Morais, D.K.; Silva, J.C.; Silva, D.B. Identification of potential insect ecological interactions using a metabarcoding approach. PeerJ 2025, 13, e18906. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zhang, X.; Zhang, L.; Zhang, H.; Zhang, J.; Zhang, C. Quantifying potentially suitable geographical habitat changes in chinese caterpillar fungus with enhanced MaxEnt model. Insects 2025, 16, 262. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xia, Y. Host-pathogen interactions between Metarhizium spp. and locusts. J. Fungi 2022, 8, 602. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Gahtyari, N.C.; Chhabra, R.; Kumar, D. Role of microbes in improving plant growth and soil health for sustainable agriculture. Diversity 2020, 12, 207–256. [Google Scholar]
- Tarfeen, N.; Zhang, X.; Zhang, L.; Zhang, H.; Zhang, J.; Zhang, C. Microbial remediation: a promising tool for reclamation of contaminated sites with special emphasis on heavy metal and pesticide pollution: a review. Processes 2022, 10, 1358. [Google Scholar] [CrossRef]
- Thanuja, A.; Rahman, S.J.; Rajanikanth, P.; Bhat, B.N. Combined effect of entomopathogenic fungus Metarhizium anisopliae (Metschinkoff) and novel insecticides on the cellular and humoral immune mechanisms of Spodoptera litura (Fabricius). Plant Arch. 2025, 25, 1234–1244. [Google Scholar]
- Yildiz, M.; Romanazzi, G.; Oguz, M.C. Bio-based strategies for biotic and abiotic stress management in sustainable agriculture. Front. Plant Sci. 2025, 15, 1536460. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, X.; Zhang, L.; Zhang, H.; Zhang, J.; Zhang, C. Beauveria bassiana induces strong defense and increases resistance in tomato to Bemisia tabaci. J. Fungi 2025, 11, 141. [Google Scholar] [CrossRef] [PubMed]
- Sorahinobar, M.; Eslami, S.; Shahbazi, S.; Najafi, J. A mutant Trichoderma harzianum improves tomato growth and defense against Fusarium wilt. Eur. J. Plant Pathol. 2025, 172, 1–6. [Google Scholar] [CrossRef]
- Ahluwalia, V.; Kumar, A.; Sharma, D.; Chhabra, R.; Kumar, D. Isolation, characterisation of major secondary metabolites of the Himalayan Trichoderma koningii and their antifungal activity. Arch. Phytopathol. Plant Prot. 2014, 47, 1063–1071. [Google Scholar] [CrossRef]
- Lanfranco, L.; Fiorilli, V.; Venice, F.; Bonfante, P. Strigolactones cross the kingdoms: plants, fungi, and bacteria in the arbuscular mycorrhizal symbiosis. J. Exp. Bot. 2018, 69, 2175–2188. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Singh, A.K.; Singh, P.K.; Singh, V.; Singh, R.; Singh, R. Integrative approaches to improve litchi (Litchi chinensis Sonn.) plant health using bio-transformations and entomopathogenic fungi. BMC Plant Biol. 2024, 24, 902. [Google Scholar] [CrossRef] [PubMed]
- Kleber, M.; Sollins, P.; Sutton-Grier, A. Dynamic interactions at the mineral-organic matter interface. Nat. Rev. Earth Environ. 2021, 2, 402–421. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, L.; Zhang, H.; Zhang, J.; Zhang, C. Species diversity and vertical distribution characteristics of entomogenous fungi in soils of northern Gaoligong Mountains. J. Yunnan Agric. Univ. (Nat. Sci.) 2024, 39, 1–9. [Google Scholar]
Figure 1.
Schematic diagram of the process of transforming the tissues in the body of an insect infected by cordyceps fungi into internal sclerotium (taking C. chanhua infected A. pernyi pupa as an example). In the figure: (1) Pupae of A. pernyi contact C. chanhua conidium; (2) C. chanhua conidium infects the tissues of A. pernyi pupae; (3) The tissues in the pupae of A. pernyi were completely infected as sclerotium; (4) The surface of A. pernyi pupae was covered by the mycelium of C. chanhua; (5) C. chanhua gradually mature, grow fruiting bodies and conidia; (6) A new round of infection. The Black straight line in the figure indicates the corresponding external structure of A. pernyi pupae at a certain period.
Figure 1.
Schematic diagram of the process of transforming the tissues in the body of an insect infected by cordyceps fungi into internal sclerotium (taking C. chanhua infected A. pernyi pupa as an example). In the figure: (1) Pupae of A. pernyi contact C. chanhua conidium; (2) C. chanhua conidium infects the tissues of A. pernyi pupae; (3) The tissues in the pupae of A. pernyi were completely infected as sclerotium; (4) The surface of A. pernyi pupae was covered by the mycelium of C. chanhua; (5) C. chanhua gradually mature, grow fruiting bodies and conidia; (6) A new round of infection. The Black straight line in the figure indicates the corresponding external structure of A. pernyi pupae at a certain period.
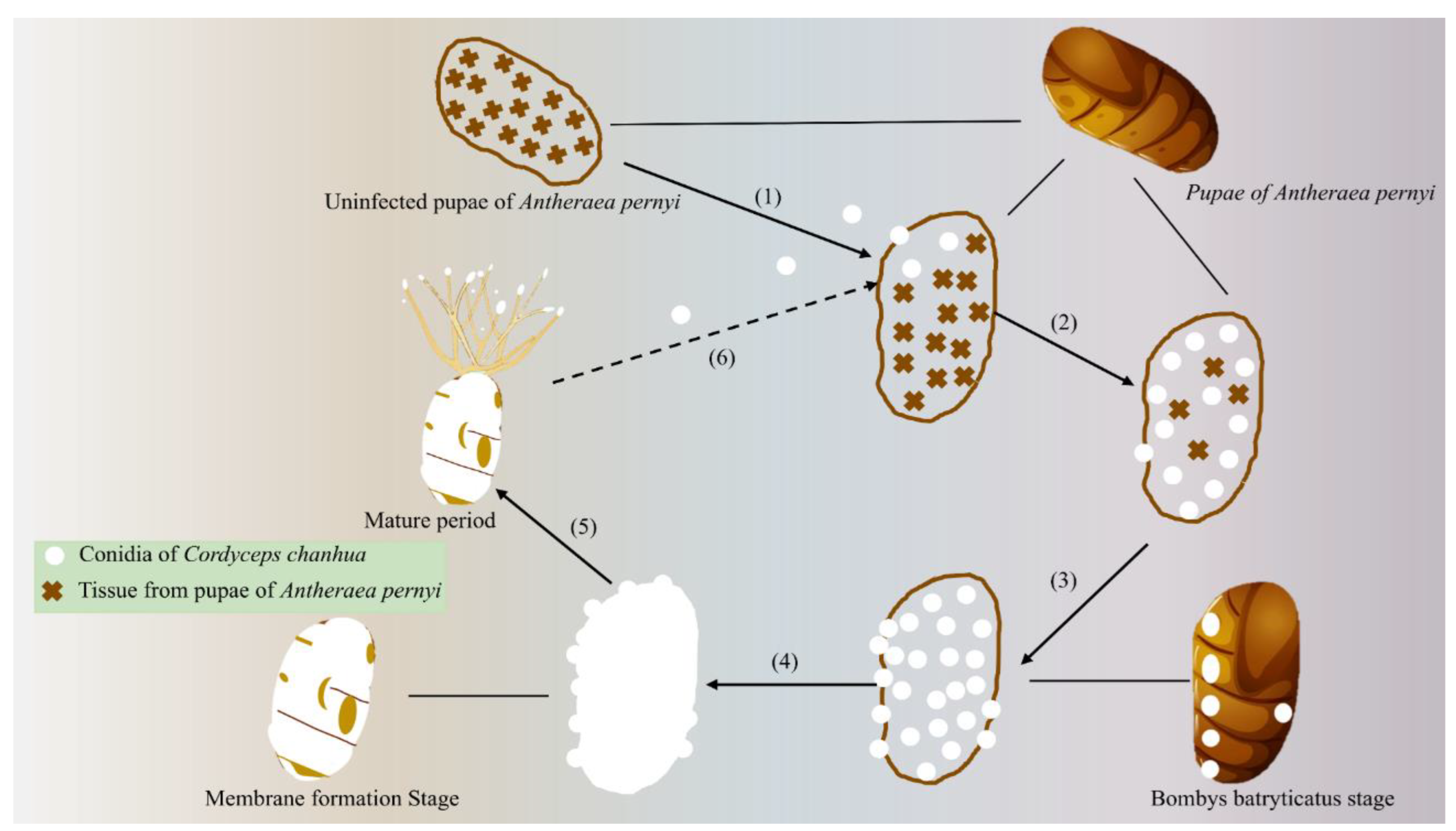
Figure 2.
EF biodiversity is affected by temperature, humidity, landscape, elevation, host type, agricultural activities, and ecological niche. a) The temperature at which the mycotic fungi grow is between 15 °C and 40 °C [97] and is optimal between 22 °C and 30 °C [98]. b) Humidity is also an important factor affecting the growth and virulence of EF [101,102], and the germination and virulence of EF increase with the increase of humidity [101,103]. The virulence of EF to host insects was increased by 68% and 32% during the rainy season compared to the dry season [104]. c) Altitude is an important environmental factor affecting the community of EF. The distribution of EF was found in soils of all altitudinal gradients, with higher numbers of species and strains in the middle elevations [137,272]. d) Different host insect types can also affect the diversity of EF, and some EF strains have different lethality to different parasitoids of the same insect [127]. e) Landscape composition could affect the occurrence of EF. Compared with simple landscapes, complex landscapes have a wide variety of insects, and EF also show a high species diversity [114,115]. Because ground vegetation serves as a shelter for host insects and EF [112], EF abundance is significantly higher in forest areas with a favorable ecological environment and appropriate temperature and humidity. f) Agricultural activities can affect the occurrence of EF in soil. The abundance of EF in the soil of organic cropland and its surrounding ridges was significantly higher than that of conventional cropland [131]. The abundance, species, and diversity of EF in soil of organic cropland were greater than those of conventional chemical cropland [134]. g) In addition, the ecological niche could also affect the EF biodiversity. Because of seasonal differences in species occurrence, temporal niche overlap can vary [141,142]. Trophic niche breadth and overlap values are related to host range, with a wider fungal host range leading to greater trophic niche breadth and relatively smaller overlap [143].
Figure 2.
EF biodiversity is affected by temperature, humidity, landscape, elevation, host type, agricultural activities, and ecological niche. a) The temperature at which the mycotic fungi grow is between 15 °C and 40 °C [97] and is optimal between 22 °C and 30 °C [98]. b) Humidity is also an important factor affecting the growth and virulence of EF [101,102], and the germination and virulence of EF increase with the increase of humidity [101,103]. The virulence of EF to host insects was increased by 68% and 32% during the rainy season compared to the dry season [104]. c) Altitude is an important environmental factor affecting the community of EF. The distribution of EF was found in soils of all altitudinal gradients, with higher numbers of species and strains in the middle elevations [137,272]. d) Different host insect types can also affect the diversity of EF, and some EF strains have different lethality to different parasitoids of the same insect [127]. e) Landscape composition could affect the occurrence of EF. Compared with simple landscapes, complex landscapes have a wide variety of insects, and EF also show a high species diversity [114,115]. Because ground vegetation serves as a shelter for host insects and EF [112], EF abundance is significantly higher in forest areas with a favorable ecological environment and appropriate temperature and humidity. f) Agricultural activities can affect the occurrence of EF in soil. The abundance of EF in the soil of organic cropland and its surrounding ridges was significantly higher than that of conventional cropland [131]. The abundance, species, and diversity of EF in soil of organic cropland were greater than those of conventional chemical cropland [134]. g) In addition, the ecological niche could also affect the EF biodiversity. Because of seasonal differences in species occurrence, temporal niche overlap can vary [141,142]. Trophic niche breadth and overlap values are related to host range, with a wider fungal host range leading to greater trophic niche breadth and relatively smaller overlap [143].
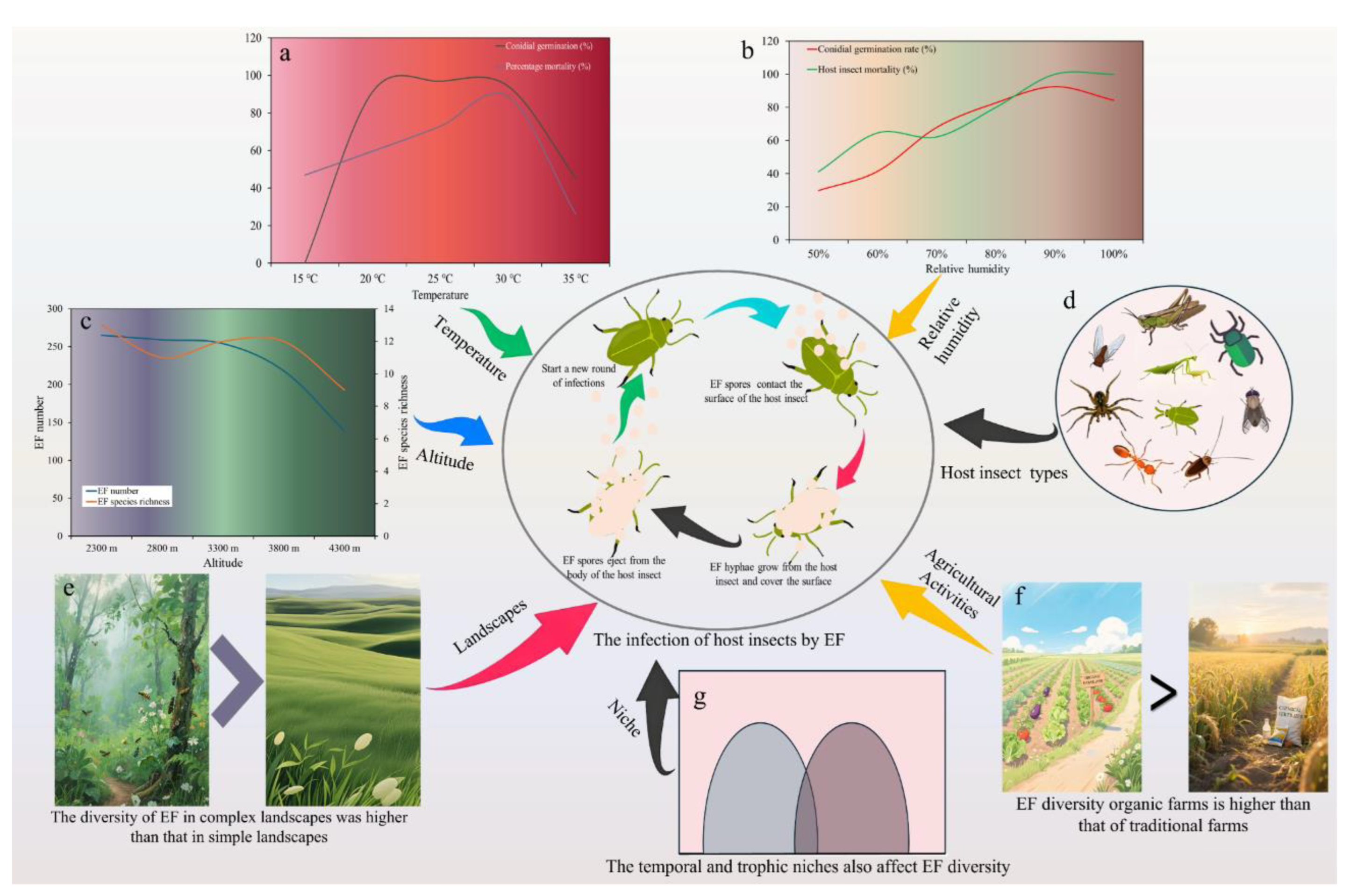
Figure 3.
Schematic diagram of insect infestation by EF in nature. (1) The process of egg hatching into larva; (2) The process in which insect larvae are infected by EF spores and grow into fungus-worm complex; (3) The process by which insect larvae develop into adults; (4) The process in which adult worms are infected by EF spores and grow into fungus-worm complex; (5~8): Schematic diagram of the process of infestation of EF from the host insect epidermis, in which: (5) A spore adheres to the insect cuticle, germinates, and forms the infection structure-the appressorium [179]; (6) In addition to the secretion of protease and chitinase, lipid droplets are transferred from the mother conidium to the appressor for hydrolysis, producing high concentration of glycerol, forming swelling pressure and destroying the stratum corneum [160,161,162]; (7) During this process and after reaching the host blood cavity, fungal cells change their cell wall structure during blood cell recognition, encapsulation, and blackening [3]. In addition, EF cells secrete effector proteins and secondary metabolites that evade host immunity by fighting host receptors (resistant proteins) [153,154]; (8) To successfully colonize the hemocoel and kill the insect, escaped filamentous cells switch to a yeast-type propagation strategy to form hyphal bodies (also called blastospores) for quick growth and host nutrient deprivation [153,154]. The red gradient arrow in the image indicates that the conidium of the EF touches and infects the insect larva or adult, and the black dotted arrow indicates that the contents of the black circle are enlarged.
Figure 3.
Schematic diagram of insect infestation by EF in nature. (1) The process of egg hatching into larva; (2) The process in which insect larvae are infected by EF spores and grow into fungus-worm complex; (3) The process by which insect larvae develop into adults; (4) The process in which adult worms are infected by EF spores and grow into fungus-worm complex; (5~8): Schematic diagram of the process of infestation of EF from the host insect epidermis, in which: (5) A spore adheres to the insect cuticle, germinates, and forms the infection structure-the appressorium [179]; (6) In addition to the secretion of protease and chitinase, lipid droplets are transferred from the mother conidium to the appressor for hydrolysis, producing high concentration of glycerol, forming swelling pressure and destroying the stratum corneum [160,161,162]; (7) During this process and after reaching the host blood cavity, fungal cells change their cell wall structure during blood cell recognition, encapsulation, and blackening [3]. In addition, EF cells secrete effector proteins and secondary metabolites that evade host immunity by fighting host receptors (resistant proteins) [153,154]; (8) To successfully colonize the hemocoel and kill the insect, escaped filamentous cells switch to a yeast-type propagation strategy to form hyphal bodies (also called blastospores) for quick growth and host nutrient deprivation [153,154]. The red gradient arrow in the image indicates that the conidium of the EF touches and infects the insect larva or adult, and the black dotted arrow indicates that the contents of the black circle are enlarged.
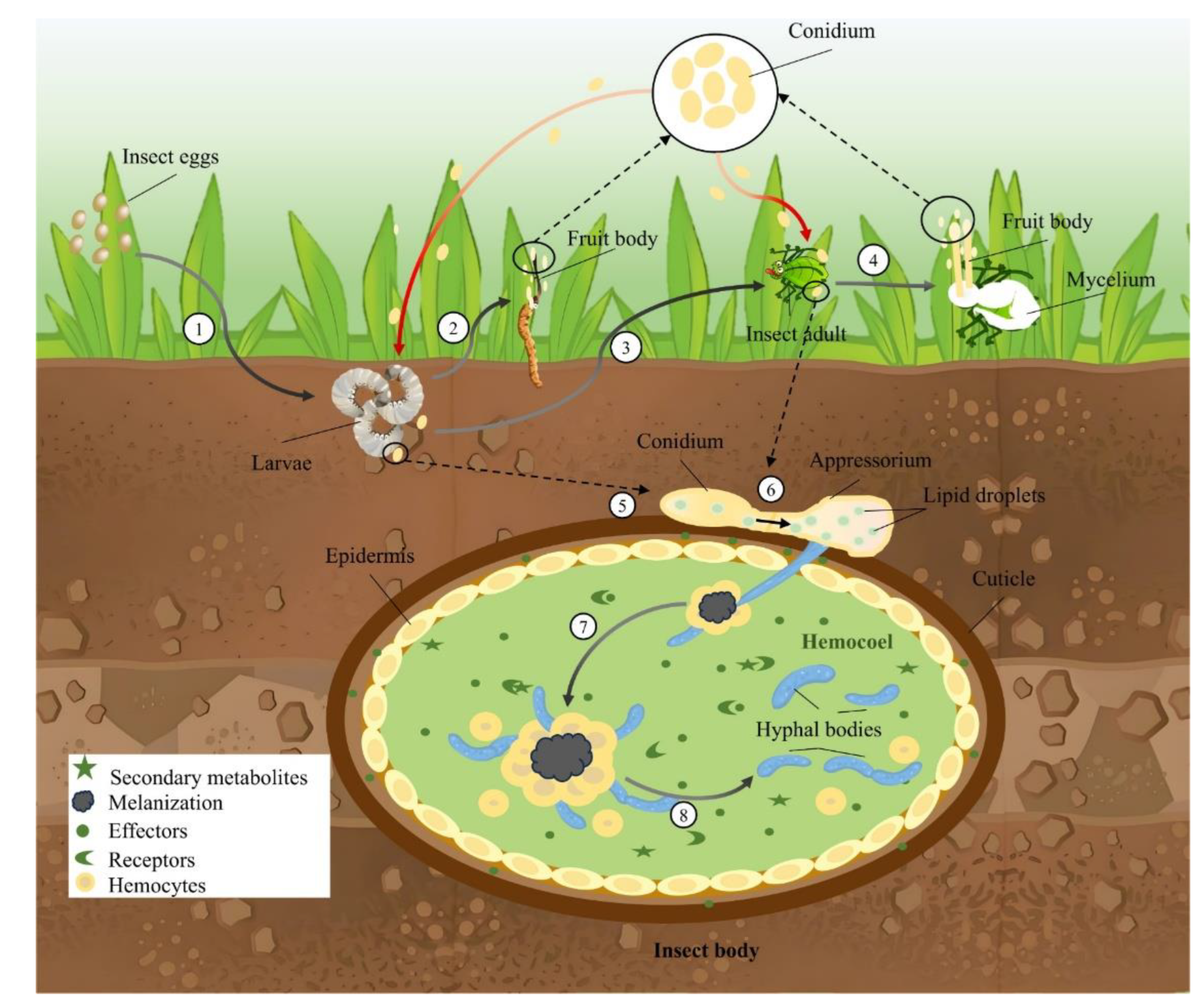
Figure 4.
Schematic diagram of EF colonizing plants and their effects on plants. On the left side of the double dotted lines in the picture, there is the colonization of EF; While on the right side, there is no colonization of EF.
Figure 4.
Schematic diagram of EF colonizing plants and their effects on plants. On the left side of the double dotted lines in the picture, there is the colonization of EF; While on the right side, there is no colonization of EF.
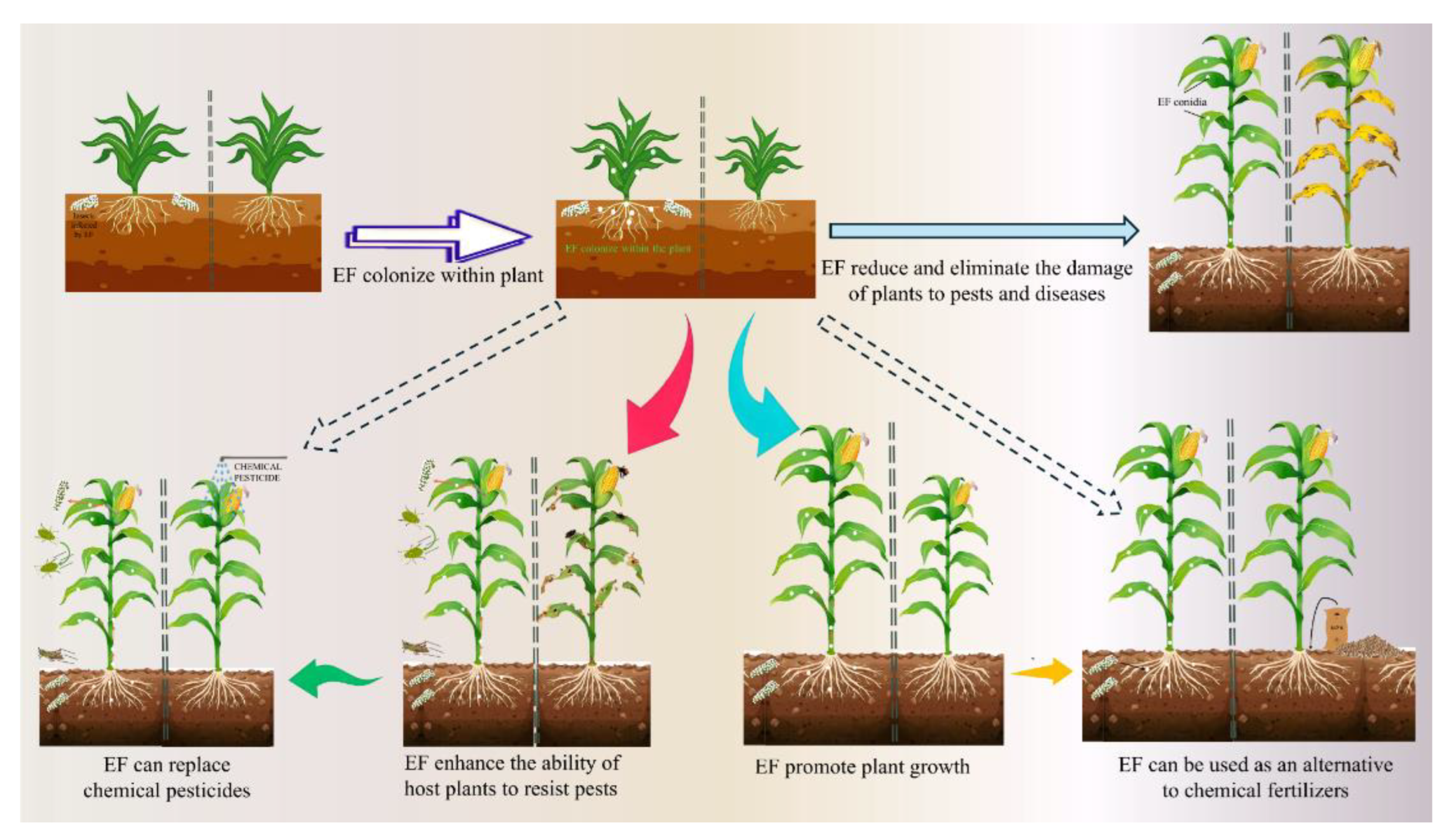
Figure 5.
EF-plant-insect interaction. (1) The process of insect gnawing on plants; (2) The process of insect infection and death by EF conidium in soil; (3) EF colonize plants; (4) EF act as endophytic fungi to protect plants from insect pests.
Figure 5.
EF-plant-insect interaction. (1) The process of insect gnawing on plants; (2) The process of insect infection and death by EF conidium in soil; (3) EF colonize plants; (4) EF act as endophytic fungi to protect plants from insect pests.
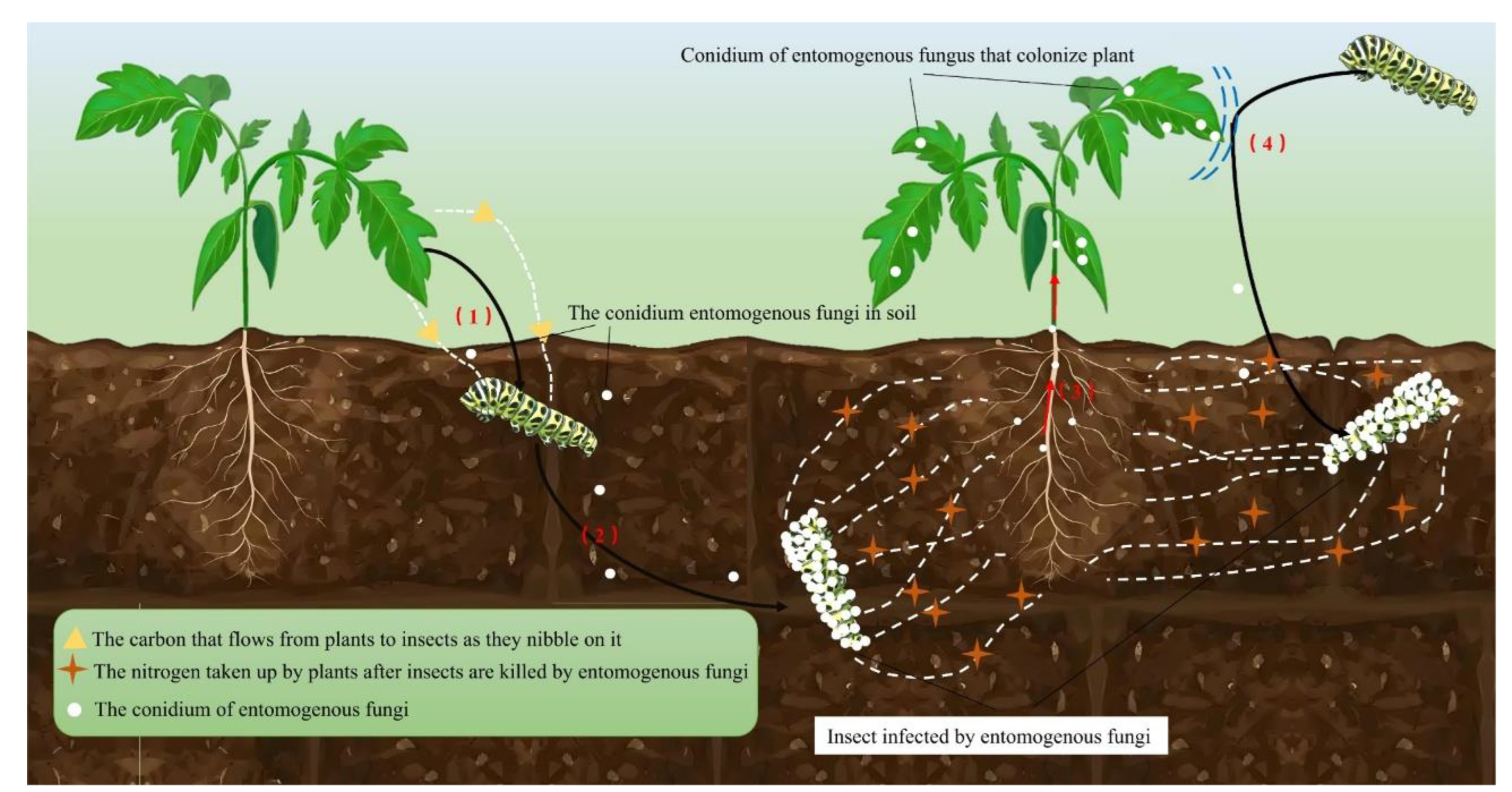
Figure 6.
Schematic diagram of the involvement of EF in soil N conversion.
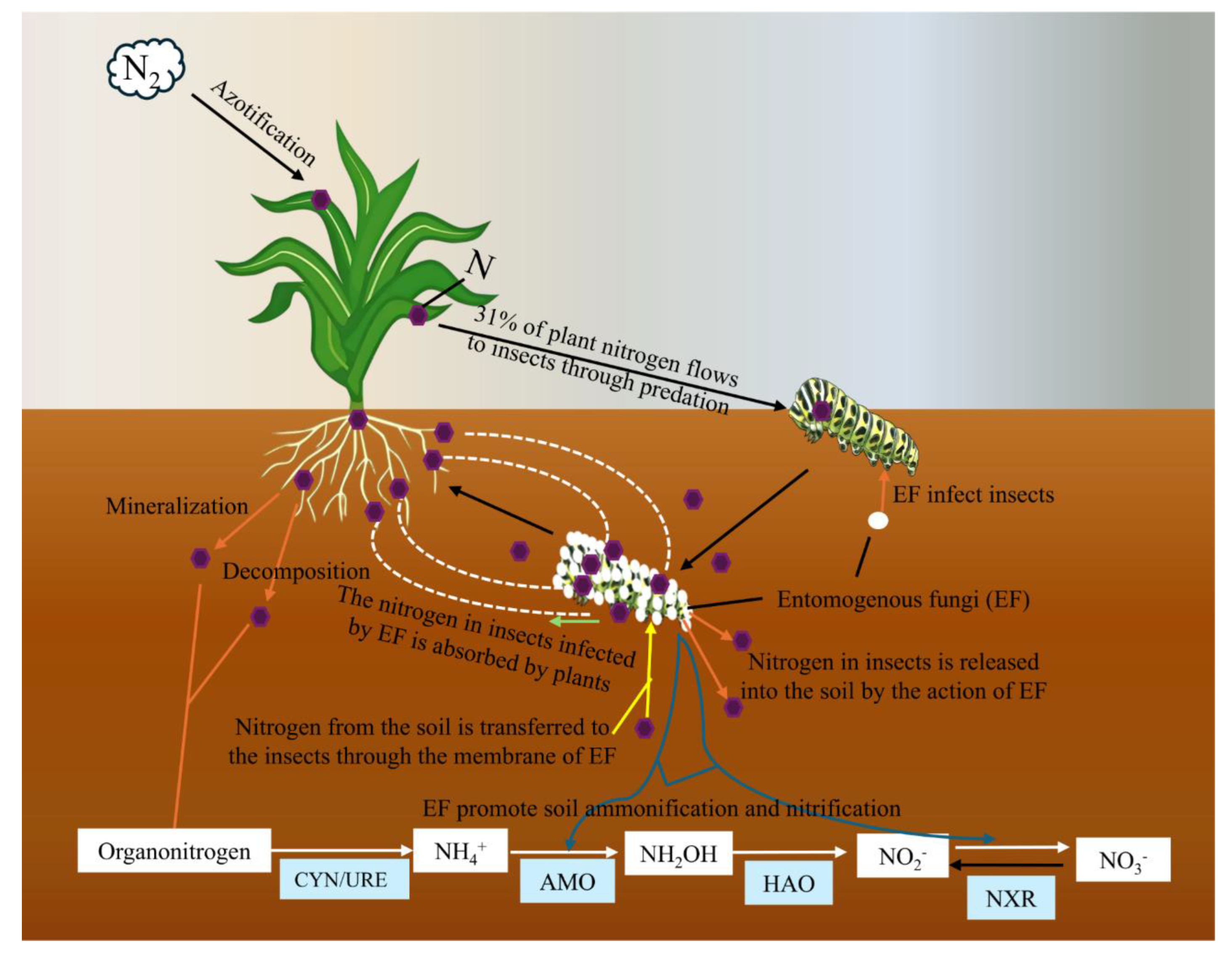

Table 1.
Differences, similarities and relationships between cordyceps fungi and biocontrol fungi.
| Cordyceps fungi | Biocontrol fungi | |
|---|---|---|
| Morphological characteristics | Fruiting bodies are formed, and ascospores are released through the perithecia [17]. | The mycelium is white or green, and the conidia are arranged in chains [18]. |
| Infection process | Spores invade the host → Hyphae slowly occupy the body cavity → Fruiting body formation after host death (weeks to months) [30]. | Spore attachment host → Secretase breaks down the body wall → Hyphae proliferate to kill the host (3-7 d) → Secondary transmission of the spores is facilitated through insect carcasses [31]. |
| Major metabolites | Cordycepin [32], adenosine [33], polysaccharide [34], ergosterol [35], etc | Beauvericin [36], cyclic peptide toxins [36], etc |
| Core function | Edible and medicinal dual-purpose: immunoregulation [37], antineoplastic [38], antioxidant [39], etc. | Agriculture and forest pest control: broad-spectrum insecticidal and fungicidal activity [40]. |
| Host specificity | Highly specialized (e.g.; C. sinensis only parasites bat moth larvae) [40]. | Broad spectrum (Lepidoptera, Coleoptera, Homoptera, etc.) [41]. |
| Ecological function | By modulating the populations of specific insects, insect carcasses contribute to soil nitrogen cycling, while decomposed fruiting bodies participate in soil carbon cycling [42,43,44]. | The capacity for cross-host transmission is robust [45], making it highly suitable for extensive agricultural and forestry pest management programs. |
| Primary application domains | Health supplements [46,47], medicines [48], food processing [46], etc. | Agricultural pest control [41], forest pest management [41,49], etc. |
| Typical products | C. sinensis capsule [50], C. militaris functional beverage [51], C. chanhua wine [52], etc. | M. anisopliae granule [53], B. bassiana agent [54], C. cateniannulata agent [55], etc. |
| Similarities | 1) Both cordyceps fungi and biocontrol fungi participate in ecological balance by parasitizing or inhibiting insects [40,41,44]; 2) Both of them can replace chemical pesticides to reduce environmental pollution [2,41,56]; 3) Genetic engineering, nano-preparation and other technologies can improve the application performance of the two fungi [57,58,59]. |
|
| Relationships | Metabolite complementation: 1) Extracts from cordyceps fungi, including polysaccharides, can serve as synergistic agents for biocontrol agents. 2) Certain biocontrol fungi also contain active compounds with medicinal and edible value. For instance, Bombyx batryticatus, formed by the infection of B. mori L. larvae with B. bassiana, is a rare traditional Chinese medicine [60]. It contains various bioactive components such as proteins, peptides, fatty acids, flavonoids, nucleosides, steroids, legumes, and polysaccharides [61,62]. Both cordyceps fungi and biocontrol fungi target insects and contribute to sustainable agriculture. Future research should focus on enhancing comprehensive benefits through interdisciplinary approaches, such as gene editing and drug combination. |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



