
Submitted:
11 July 2025
Posted:
15 July 2025
You are already at the latest version
Abstract
Carbapenem-resistant Klebsiella pneumoniae (CR‑Kp) is eroding therapeutic options for urinary-tract infections. We isolated a multidrug-resistant strain from the urine of a chronically bacteriuric patient and confirmed its identity by Vitek-2 and MALDI-TOF MS. Initial disk-diffusion profiling against 48 antibiotics revealed susceptibility to only five agents. One month later, repeat testing showed that tetracycline alone remained active, highlighting the strain’s rapidly evolving resistome. Given the scarcity of drug options, we performed an “aromatogram” with six pure essential oils and two commercial phytotherapeutic blends. Biomicin Forte® produced a 30 mm bactericidal halo, while thyme, tea-tree, laurel and palmarosa oils yielded clear inhibition zones of 11–22 mm. These in-vitro data demonstrate that carefully selected plant-derived products can target CR-Kp where conventional antibiotics fail. Integrating aromatogram results into One-Health stewardship plans may therefore help preserve last-line antibiotics and provide adjunctive options for persistent urinary infections.
Keywords:
Klebsiella pneumoniae
; multidrug resistance
; urinary tract infection
; essential oils
; One Health
1. Introduction
As a foremost cause of hospital acquired infections worldwide, Klebsiella pneumoniae (K. pneumoniae) exemplifies a One Health threat that bridges human, animal, and environmental reservoirs. Contemporary phylogenomic analyses recognise at least seven species or subspecies: K. pneumoniae subsp. pneumoniae, K. quasipneumoniae subsp. quasipneumoniae, K. quasipneumoniae subsp. similipneumoniae, K. variicola subsp. variicola, K. variicola subsp. tropica, K. quasivariicola, and K. africana [1,2]. Accurate species delineation can be achieved using a seven-gene multilocus sequence typing (MLST) barcode that combines single-linkage clustering at multiple thresholds with life identification numbers (LIN) [3].
K. pneumoniae is ubiquitous, occurring in soil, water, and the gastrointestinal tracts of humans and diverse animal species, which facilitates zoonotic transmission [4]. After faecal contamination, it adapts and multiplies in varied biotopes such as tree surfaces, plant phyllospheres, and the interior walls of wooden water tanks. Other reported habitats include sewage, drinking water, industrial effluents, and vegetation. In humans, the bacterium is common in hospital environments, colonising the urinary and respiratory tracts or the intestine, particularly following prolonged antibiotic therapy [5].
Clinically, K. pneumoniae is a prominent opportunistic pathogen in both nosocomial and community settings, renowned for its ability to develop multidrug resistance and for its pronounced genotypic and phenotypic diversity [1,6,7]. Companion animals (dogs and cats) with urinary-tract infections have been shown to carry K. pneumoniae strains harboring resistance and virulence genes that mirror those found in human isolates [8].
The emergence of multidrug-resistant (MDR) and extensively drug-resistant (XDR) clones represents a serious public-health threat, limiting therapeutic options and increasing both morbidity and mortality [5,9]. Within this genetically diverse species, it remains difficult to recognize or predict the rise of clinically significant clones. K. pneumoniae possesses a large accessory genome, comprising approximately 30,000 protein-coding genes, many of which encode virulence functions. The convergence of virulence and resistance loci may ultimately yield untreatable invasive infections [1]. Nosocomial outbreaks with wide geographic amplitude have been attributed to K. pneumoniae, causing severe infections for which effective treatments are lacking owing to antibiotic resistance [10,11]. The organism is now considered an urgent global health threat because of the emergence of MDR and hypervirulent strains capable of causing severe disease worldwide [1,12].
K. pneumoniae isolates are broadly divided into two major pathotypes: classical (cKP) and hypervirulent (hvKP) [13,14]. Both can lead to bacteraemia with substantial morbidity and mortality, but hvKP strains are more likely to provoke severe, disseminated disease than cKP [15,16]. These groups are genetically distinct, occupy non-overlapping ecological niches, cause different clinical syndromes, and interact differently with host immunity [14]. The species has thus evolved from a common opportunist into one of the most dangerous pathogens, with hvKP causing life-threatening, invasive disease [17,18]. Hypervirulent strains are globally disseminated, but accurate discrimination from classical strains may require animal models or molecular biomarkers such as the murine sepsis assay [19,20]. In addition to animal lethality tests, molecular detection offers a practical alternative [21]. A hypermucoviscous colony phenotype is a strong laboratory indicator of hvKP [13]. MDR, hvKP clones are emerging pathogens of global concern [22]. hvKP has spread from the Asia-Pacific region worldwide, causes highly invasive infections, and is progressively acquiring antimicrobial resistance [20,21,23]. The simultaneous presence of resistance and hypervirulence has raised major concern [2,12,24]. The rapid geographic expansion of MDR-hvKP clones is alarming [25], with hvKP ST23 now documented in at least one country in each of the six WHO regions [5]. HvKP strains are more likely to cause severe, disseminated infection than classical strains; their colonies appear hypermucoviscous on agar, and the simple string test aids early laboratory recognition [16].
K. pneumoniae harbours multiple potent virulence factors, including capsular exopolysaccharides, lipopolysaccharide (LPS), adhesive fimbriae (type 1 and type 3), and iron-scavenging siderophores. The polysaccharide capsule is a pivotal determinant that forms the bacterium’s outermost layer and shields it against host defenses such as phagocytosis, complement, opsonophagocytosis, oxidative killing, and antimicrobial peptides [24,26]. Capsule and fimbriae also cooperate in biofilm formation. More than 80 capsular serotypes are recognized among pathogenic strains, yet only a subset predominates in invasive disease [27]. Determination of anti-Klebsiella antibodies (IgG, IgM, and IgA) is recommended in urinary-tract infections. LPS acts as an endotoxin and, together with the capsule, provides an effective barrier to serum complement [24]. In uropathogenic strains, fimbriae are critical for adherence to uroepithelial cells, preventing washout during micturition and allowing persistence in the host [18]. The frequency, clinical significance, and morbidity of K. pneumoniae UTIs have risen steadily [25,28,29]. Beyond pneumonia and other syndromes, the organism is a leading and often recalcitrant uropathogen [18,25,30]. It is also among the most common UTI agents that produce carbapenemases (KPCp isolates), creating major therapeutic challenges [29]. Given that UTIs rank among the world’s most prevalent bacterial infections, the emergence of multidrug-resistant K. pneumoniae within this niche is of particular concern [6,24,31].
Accordingly, this study aimed to (i) identify and characterize a K. pneumoniae strain isolated from the urine of a human patient, (ii) describe its antimicrobial-resistance mechanisms, and (iii) evaluate the in-vitro activity of 56 antibiotics, six essential oils, a propolis tincture and two commercial phytotherapeutic blends as potential adjuncts or alternatives for urinary-tract infection management.
2. Materials and Methods
2.1. Isolation of the Strain
The strain was isolated from an 84-year-old male patient with a history of bacteriuria. The K. pneumoniae isolate was obtained by streaking routine media - nutrient broth and nutrient agar - together with selective media (MacConkey agar and UriSelect™, Bio-Rad Laboratories Inc., Hercules, CA, USA) that facilitate the preliminary recognition of Enterobacteriaceae. After inoculation, tubes and Petri plates were aerobically incubated at 37°C in a thermostatic chamber and examined after 24 and 48 h.
2.2. Phenotypic and Morphological Characterization
Presumptive identification relied on colony appearance on the various media and cellular morphology revealed by Gram-stained smears.
2.3. Definitive Identification
Species confirmation was achieved with two automated systems, Vitek® 2 Compact 15 (bioMérieux, Craponne, France), which uses an array of biochemical reactions to generate species-level identifications, and MALDI Biotyper® Sirius System (Bruker, Ettlingen, Germany), which applies matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry (MALDI-TOF MS) to analyse the bacterial protein profile (CLSI, 2023).
2.4. Antimicrobial Susceptibility Testing
Susceptibility was evaluated by the standard Kirby-Bauer agar-disk diffusion method, interpreted according to EUCAST breakpoints, but cross-checked with CLSI zone-diameter tables. A 0.5 McFarland suspension of the isolate was swab-spread onto Mueller-Hinton agar (Merck, Darmstadt, Germany) plates; antibiotic disks (commercially coded micro-tablets) were then placed on the surface. Plates were incubated at 35±1°C for 18±2 hours. Inhibition zones were measured to the nearest millimeter with a digital vernier caliper and categorized as susceptible (S), intermediate (I), or resistant (R). Two evaluations were performed using the same antibiotics at one month apart. For the second susceptibility testing, the K. pneumoniae strain was once again isolated from the urine sample of the same patient.
The tested antimicrobials were the following (drug/code/content): Penicillin (PEN, 1 unit), Oxacillin (OX, 1 unit), Mecillinam (MEC, 10 µg), Piperacillin (PRL, 30 µg), Ampicillin + Sulbactam (SAM, 10/10 µg, Amoxicillin + Clavulanic Acid (AMC, 20/10), Cefadroxil (CFR, 30 µg), Cephradine (CE, 30 µg), Cefacetrile (CEF, 30 µg), Cephalothin (CH, 30 µg), Cefaclor (CEC, 30 µg), Cefotetan (CTT, 30 µg), Cefuroxime (CXM, 30 µg), Cefamandole (MA, 30 µg), Ceftriaxone (CRO, 30 µg), Ceftazidime (CAZ, 10 µg), ), Ceftazidime + avibactam (CZA, 10/4 µg), Cefoperazone (CEP, 75 µg), Ceftiofur (FUR, 30 µg), Cefixime (CFM, 5 µg), Cefquinome (CFQ, 30 µg), Meropenem (MEM, 10 µg), Imipenem (IMI, 10 µg), Amikacin (AK, 30 µg), Streptomycin (S, 10 µg), Gentamicin (GME, 10 µg), Azithromycin (AZM, 15 µg), Clarithromycin (CLR, 15 µg), Tetracycline (TE, 30 µg), Doxycycline (DOX, 30 µg), Tulathromycin (TUL, 30 µg), Spiramycin (SP, 30 µg), Bacitracin (DTD, 10 units), Colistin (CT, 25 µg), Fosfomycin (FF, 200 µg), Pristinamycin (PT, 15 µg), Novobiocin (NV, 30 µg), Norfloxacin (NX, 10 µg), Marbofloxacin (MAR 5 µg), Ofloxacin (OFX, 5 µg), Chloramphenicol (C, 30 µg), Rifampicin (RD, 5 µg), Nitrofurantoin (F, 100 µg), Metronidazole (MET, 5 µg), Furazolidone (FX, 50 µg), Fusidic Acid (FA, 10 µg), Compound Sulphonamides (S3, 300 µg), Trimethoprim + Sulfamethoxazole (SXT, 1.25/23.75 µg). All commercial antibiotic disks were purchased from Liofilchem®, Roseto degli Abruzzi, Italy, and Bio-Rad Laboratories Inc., Hercules, CA, USA.
2.5. Essential Oil and Antiseptic Susceptibility Testing
Sterile blank filter-paper disks (⌀ 6 mm) were impregnated with 10 µL of each test solution - neat essential oil, tincture, or antiseptic - then placed on Mueller–Hinton agar plates that had been lawn-inoculated with the K. pneumoniae 0.5 McFarland suspension, following the same layout as the antibiotic disk-diffusion assay. Plates were incubated at 37°C for 24 h and re-read at 48 h; inhibition halos were measured in millimeters. Complete absence of growth within the halo was interpreted as bactericidal activity, whereas satellite colonies were recorded as evidence of partial resistance. Essential oils screened were represented by palmarosa (Cymbopogon martini), geranium (Pelargonium graveolens), frankincense (Boswellia carteri), laurel (Laurus nobilis), tea tree (Melaleuca alternifolia), and thyme (Thymus vulgaris), all purchased from Elemental SRL, Oradea, Romania. Natural remedy formulations were represented by propolis tincture, Biomicin Urinar® (A20, Fares, Romania), Biomicin Forte® (A3, Fares, Romania). An antiseptic comparator represented by 1 % methylene-blue solution was also used. Moreover, a standard antibiotic disk containing amoxicillin + clavulanic acid (AMC) was included on each plate as a broad-spectrum reference control.
3. Results
3.1. Cultural Characterization
Regarding growth in nutrient broth, after 24 h of incubation, the broth became markedly turbid, with a thick surface pellicle that later sedimented to the bottom of the tube. When the tubes were tilted or gently rotated, the culture adhered to the glass walls, indicating abundant extracellular mucus. With aging (48–72 h), the broth viscosity increased further because of copious capsular-mucus production. The hyper-mucoviscous phenotype was most obvious when the culture was withdrawn with a loop or Pasteur pipette, producing long, filamentous strings (“string test” positive).
After 24 h on solid media, the isolate formed colonies whose size, surface appearance, pigmentation and consistency evolved over time (48–72 h), as follows: on nutrient and Mueller-Hinton agar: large (3-5 mm diameter), convex, opaque, glossy, non-pigmented colonies (Supplementary material). On UriSelect™ chromogenic agar, colonies were similar in size and shape, but black in color, consistent with the primary differentiation of Enterobacteriaceae. On blood agar, intensely shiny, non-hemolytic colonies were observed that clearly exhibited the mucoid character. Upon prolonged incubation, the colonies enlarged, acquired a faint pink hue, and tended to coalesce (Supplementary material).
3.2. Antibiotic Susceptibility Testing
Regarding the first evaluation, out of the 48 antibiotics and chemotherapeutics agents tested, susceptibility was demonstrated for only 5 (10.42%, CI 95% 3.47-22.66), as follows: streptomycin, tetracycline, doxycycline, chloramphenicol, and tulathromycin. Resistance was recorded to a large number of the agents tested, 43 in total (89.58%, CI 95% 77.34-96.53) (Table 1).
On the other hand, the second evaluation revealed a higher resistance rate, with only 1 efficient antibiotic (2.08%, CI 95% 0.05-11.07), namely tetracycline, the bacterial strain being resistant to 47 antimicrobials (97.92%, CI 95% 88.93-99.95). Results are presented in Table 1.
Serial testing revealed a precipitous loss of activity, so that streptomycin, tulathromycin, chloramphenicol and doxycycline all flipped from susceptible to resistant between evaluations, leaving tetracycline as the sole agent retaining efficacy (Figure 1).
Periodic evaluations consisting of urine culture with susceptibility testing (interpreted according to EUCAST guidelines), that were performed in a private laboratory (Synevo, Romania), with the identification of the bacterial species and quantification of the bacterial load (CFU mL−1). Results are summarized in Table 2.
Notably, the susceptibility profile shifted during bacteriuria for several agents, such as gentamicin, amikacin, amoxicillin–clavulanic acid, ceftazidime, cefepime, and trimethoprim–sulfamethoxazole, indicating acquisition or loss of resistance over time. Accordingly, we recommend periodic urine cultures with a full antibiogram, even when the bacterial count is below the usual detection threshold (<1,000 CFU mL−1), to ensure that therapy remains effective. Multiple intrinsic and extrinsic factors can alter the susceptibility/resistance status of the Klebsiella strain in question, and timely monitoring allows treatment to be adjusted before clinical failure occurs.
3.3. Essential oils susceptibility testing
The antimicrobial activity of the tested essential oils and natural products against K. pneumoniae varied significantly. The largest inhibition zones were observed for Biomicin forte® (A3) and Thymus vulgaris (thyme), with diameters of 30 mm and 22 mm, respectively, indicating strong antibacterial properties. Melaleuca alternifolia (tea tree oil) showed moderate efficacy, with inhibition zones of 20 mm. Moderate activity was also recorded for Laurus nobilis (laurel) and Biomicin urinar® (A20), both yielding zones around 12 mm, although the latter showed the presence of resistant colonies. In contrast, several oils, including Pelargonium graveolens (geranium), Boswellia carteri (frankincense), Cymbopogon nardus (citronella), and propolis, exhibited no inhibitory activity, as indicated by resistant growth. These findings highlight considerable variability in the susceptibility of K. pneumoniae to different natural products, suggesting potential for selective use in antimicrobial strategies. The results are presented in Table 3.
4. Discussion
Klebsiella pneumoniae remains a leading cause of healthcare-associated infections worldwide, driven by a highly diverse population structure that complicates both genomic surveillance and clinical management. Kleborate, a recently developed analytic pipeline, now streamlines genotype to phenotype prediction directly from intestinal metagenomes and cultured isolates, offering a practical answer to this complexity [32]. MDR in K. pneumoniae arises through four, often co-occurring, mechanisms: (i) production of extended-spectrum β-lactamases (ESBLs), (ii) decreased outer-membrane permeability via porin loss (e.g., OmpK35/OmpK36), (iii) over-expression of efflux pumps such as the intrinsic MFS pump KpnGH, and (iv) modification of antimicrobial targets [10,33]. Most resistance determinants reside on mobile genetic elements such as plasmids, transposons, and integrons, facilitating both vertical inheritance and horizontal exchange within and across species [2,34].
Plasmids are typically circular, autonomously replicating DNA molecules, although linear variants have been documented. Standardized multiplex PCR enables rapid plasmid typing, a prerequisite for tracking dissemination of drug-resistance cassettes in K. pneumoniae populations [35]. Conjugative plasmids, in particular, encode the full complement of transfer machinery and therefore mediate inter-strain spread over large taxonomic distances [36]. Specific β-lactamase and carbapenemase genes often segregate with discrete plasmid backbones, and copy-number amplification can further boost resistance levels. Porin loss, particularly of OmpK36, synergizes with plasmid-borne ESBLs and carbapenemases to confer pan-β-lactam resistance [37].
MDR K. pneumoniae lineages are genomically plastic, engaging in frequent chromosomal recombination that reshuffles capsule (K-locus) and O-antigen loci [7]. By contrast, hyper-virulent clones tend to recombine less but acquire large virulence plasmids, siderophore systems and regulators of the hyper-mucoid phenotype [38,39]. Alarmingly, recent reports describe pathotypes that unite extensive drug resistance with hyper-virulence, creating both untreatable and highly invasive strains [11,40]. Global travel and healthcare tourism accelerate their dispersal, highlighting the need for continuous genomic surveillance.
Our isolate displayed broad resistance encompassing aminoglycosides, β-lactams (penicillins, cephalosporins, carbapenems), macrolides, fluoroquinolones, and several second-line classes. Serial susceptibility testing revealed a precipitous loss of activity: streptomycin, tulathromycin, chloramphenicol and doxycycline transitioned from susceptible to resistant between two evaluations, leaving tetracycline as the sole agent retaining efficacy. The phenotype aligns with the presence of transferable ESBL/carbapenemase genes combined with porin down-regulation and active efflux.
Class-by-class overview
Aminoglycosides: only streptomycin remained initially active; aminoglycosides act by irreversible binding to the 30S ribosomal subunit [34].
Penicillins & β-lactams: universal resistance likely reflects high-level ESBL and carbapenemase activity [34,43,44,45,46].
Macrolides: intrinsic resistance predominated, although veterinary tulathromycin was transiently effective. Efflux appears contributory [34].
Other classes: resistance to polypeptides, streptogramins, aminocoumarins, fluoroquinolones, rifamycins, sulfonamides, and nitrofuran derivatives was universal. Tetracycline and chloramphenicol showed the only measurable activity, but both were undermined by rapid resistance emergence.
With conventional options exhausted, monoclonal antibodies targeting pili and outer-membrane adhesins show promise in animal models of urinary-tract infection [9,26,48]. Bacteriophage therapy is garnering renewed interest for MDR K. pneumoniae infections [49].
The worldwide rise in microbial resistance to conventional chemicals and drugs has spurred intensive searches for new broad-spectrum biocides and alternative therapeutic strategies [50,51]. Essential oils (EOs) possess notable antibacterial activity: their constituents disrupt the bacterial cell membrane, lower its permeability, and ultimately kill the microorganism. Moreover, pathogens appear unable to develop resistance to EOs because the oils contain such a wide variety of components that adaptive mutation is virtually impossible.
Natural products such as EOs are promising because of their complex composition; they have already proved effective against drug-resistant K. pneumoniae strains, although their overall mechanisms are not yet fully elucidated [45,52]. EOs contain diverse secondary metabolites capable of inhibiting or slowing the growth of bacteria, yeasts, and molds, mainly by targeting the cell membrane, cytoplasm and, in some cases, profoundly altering cell morphology [50]. Plant-derived EOs from oregano (Origanum vulgare), sage (Salvia officinalis), and thyme (Thymus vulgaris) have attracted attention because they may substitute for failing traditional antibiotics against pathogens, including Klebsiella spp. [53].
Representative single-compound and mechanistic findings include:
- Eugenol - a guaiacol derivative in clove, nutmeg, basil, and bay oils—acts as a bactericidal antiseptic. At 0.2 mg mL−1 it damages the cell membrane of carbapenem-resistant K. pneumoniae and blocks biofilm formation [54].
- Cinnamon bark oil (Cinnamomum verum) induces oxidative stress in KPC-producing K. pneumoniae, disrupts the phospholipid bilayer, and leads to loss of major outer-membrane proteins and cell viability [45].
- Thymol (from thyme) displays strong synergy with streptomycin or kanamycin, both inhibiting biofilm formation and destroying pre-formed K. pneumoniae biofilm [55].
- Purified EO from hibiscus rosa-sinensis inhibits biofilm formation; scanning electron microscopy shows marked morphological alteration [56].
- Ylang-ylang and vanilla/patchouli oils stabilized on iron-oxide nanostructures prevent K. pneumoniae adhesion and biofilm development [57].
- Tea tree and thyme EOs exhibit potent activity against MDR K. pneumoniae and cause severe loss of cellular integrity under electron microscopy [51].
Although in-vitro EO activity is encouraging, pharmacokinetic and toxicity profiles in vivo remain under-studied. Future work should integrate whole-genome sequencing with plasmid typing to map resistance determinants, while controlled clinical trials are required to validate EO-based adjuncts.
The convergence of multidrug resistance and hyper-virulence in K. pneumoniae heightens the urgency for innovative countermeasures. Continuous genomic surveillance, strict antimicrobial stewardship and the development of adjunctive therapies, ranging from monoclonal antibodies to tailored essential-oil formulations, are pivotal to preserving treatment efficacy and safeguarding public health.
5. Conclusions
The urinary K. pneumoniae isolate displayed the classic hyper-mucoid phenotype and was confirmed by Vitek-2 and MALDI-TOF analyses. Kirby-Bauer testing showed that susceptibility was limited to a small fraction of the antimicrobial panel, whereas three-quarters of the agents were categorized as resistant. Follow-up cultures, performed at intervals and interpreted with the same standard, reproduced this predominantly resistant profile and revealed that the strain’s response to gentamicin, amikacin, amoxicillin–clavulanate, ceftazidime, cefepime, and trimethoprim-sulfamethoxazole fluctuated over time. These shifts highlight the importance of repeating the antibiogram, even when bacteriuria falls below 103 CFU mL−1, to keep treatment aligned with the current susceptibility pattern.
Essential-oil screening indicated reproducible inhibition by palmarosa (Cymbopogon martini), laurel (Laurus nobilis), tea tree (Melaleuca alternifolia), and thyme (Thymus vulgaris) oils; geranium, frankincense, citronella oil, and propolis tincture were inactive. The commercial blends Biomicin Urinar and, in particular, Biomicin Forte produced clear, stable halos, with Biomicin Forte exerting a bactericidal effect. Given that resistant subcolonies were absent within these zones, selected essential oils could serve as adjuncts for K. pneumoniae urinary infections, provided they are chosen on the basis of a laboratory “aromatogram” performed in parallel with the routine antibiogram.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, S.R. and M.L.M.; methodology, G.C.N.; software, C.Ș.N.; validation, S.R., G.C.N. and M.L.M.; formal analysis, S.C.; investigation, M.L.M. and S.R.; resources, N.F.; data curation, C.B.; writing—original draft preparation, M.L.M., G.C.N. and S.R.; writing—review and editing, G.C.N. and C.Ș.N.; visualization, S.C.; supervision, S.R.; project administration, C.M.B.; funding acquisition, C.M.B. and N.I.F. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by funds from Romania’s Ministry of Education, CNFIS-FDI-2025-F-0579.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
No new data were created.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proceedings of the National Academy of Sciences. 2015, 112, 27–E3574. [Google Scholar] [CrossRef] [PubMed]
- Karampatakis, T.; Tsergouli, K.; Behzadi, P. Carbapenem-Resistant Klebsiella pneumoniae: Virulence Factors, Molecular Epidemiology and Latest Updates in Treatment Options. Antibiotics. 2023, 12, 234. [Google Scholar] [CrossRef] [PubMed]
- Hennart, M.; Guglielmini, J.; Bridel, S.; Maiden, M.C.; Jolley, K.A.; Criscuolo, A.; Brisse, S. A dual barcoding approach to bacterial strain nomenclature: genomic taxonomy of Klebsiella pneumoniae strains. Molecular Biology and Evolution. 2022, 39, 7–msac135. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.C.; Tang, H.L.; Chiou, C.S.; Lin, Y.C.; Chiang, M.K.; Tung, K.C.; Lai, Y.C.; Lu, M.C. Prevalence and Virulence Profiles of Klebsiella pneumoniae Isolated From Different Animals. Veterinary Medicine and Science. 2025, 11, 2–e70243. [Google Scholar] [CrossRef] [PubMed]
- WHO Antimicrobial Resistance, Hypervirulent Klebsiella pneumoniae—Global Situation. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2024-DON527 (accessed on 20 June 2025).
- Blin, C.; Passet, V.; Touchon, M.; Rocha, E.P.; Brisse, S. Metabolic diversity of the emerging pathogenic lineages of Klebsiella pneumoniae. Environmental microbiology. 2017, 19, 5–1881. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Wick, R.R.; Judd, L.M.; Froumine, R.; Tokolyi, A.; Gorrie, C.L.; Lam, M.M.; Duchêne, S.; Jenney, A.; Holt, K.E. Distinct evolutionary dynamics of horizontal gene transfer in drug resistant and virulent clones of Klebsiella pneumoniae. PLoS genetics. 2019, 15, 4–e1008114. [Google Scholar] [CrossRef] [PubMed]
- Marques, C.; Menezes, J.; Belas, A.; Aboim, C.; Cavaco-Silva, P.; Trigueiro, G.; Telo Gama, L.; Pomba, C. Klebsiella pneumoniae causing urinary tract infections in companion animals and humans: population structure, antimicrobial resistance and virulence genes. Journal of Antimicrobial Chemotherapy. 2019, 74, 3–594. [Google Scholar] [CrossRef] [PubMed]
- Petrosillo, N.; Taglietti, F.; Granata, G. Treatment options for colistin resistant Klebsiella pneumoniae: present and future. Journal of clinical medicine. 2019, 8, 7–934. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.H.; Porto, W.F.; de Faria Jr, C.; Dias, S.C.; Alencar, S.A.; Pickard, D.J.; Hancock, R.E.; Franco, O.L. Genomic insights into the diversity, virulence and resistance of Klebsiella pneumoniae extensively drug resistant clinical isolates. Microbial Genomics. 2021, 7, 8–000613. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Yang, X.; Chan, E.W.; Zhang, R.; Chen, S. Klebsiella species: Taxonomy, hypervirulence and multidrug resistance. EBioMedicine. 2022, 79. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae population genomics and antimicrobial-resistant clones. Trends in microbiology. 2016, 24, 12–944. [Google Scholar] [CrossRef] [PubMed]
- Walker, K.A.; Miller, V.L. The intersection of capsule gene expression, hypermucoviscosity and hypervirulence in Klebsiella pneumoniae. Current opinion in microbiology. 2020, 54, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ferrer, S.; Peñaloza, H.F.; Budnick, J.A.; Bain, W.G.; Nordstrom, H.R.; Lee, J.S.; Van Tyne, D. Finding order in the chaos: outstanding questions in Klebsiella pneumoniae pathogenesis. Infection and immunity. 2021, 89, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Kochan, T.J.; Nozick, S.H.; Medernach, R.L.; Cheung, B.H.; Gatesy, S.W.; Lebrun-Corbin, M.; Mitra, S.D.; Khalatyan, N.; Krapp, F.; Qi, C.; Ozer, E.A. Genomic surveillance for multidrug-resistant or hypervirulent Klebsiella pneumoniae among United States bloodstream isolates. BMC infectious diseases. 2022, 22, 1–603. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Ong, E.L. Positive string test in hypervirulent Klebsiella pneumoniae liver abscess. Oxford Medical Case Reports. 2022, 4, omac035. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Lam, M.M.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nature Reviews Microbiology. 2020, 18, 6–344. [Google Scholar] [CrossRef] [PubMed]
- Ochońska, D.; Brzychczy-Włoch, M. Klebsiella Pneumoniae–Taxonomy, Occurrence, Identification, Virulence Factors and Pathogenicity. Advancements of Microbiology. 2024, 63, 3. [Google Scholar] [CrossRef]
- Russo, T.A.; Olson, R.; Fang, C.T.; Stoesser, N.; Miller, M.; MacDonald, U.; Hutson, A.; Barker, J.H.; La Hoz, R.M.; Johnson, J.R. Identification of biomarkers for differentiation of hypervirulent Klebsiella pneumoniae from classical K. pneumoniae. Journal of clinical microbiology. 2018, 56, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Marr, C.M. Hypervirulent klebsiella pneumoniae. Clinical microbiology reviews. 2019, 32, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Hu, D. The making of hypervirulent Klebsiella pneumoniae. Journal of Clinical Laboratory Analysis. 2022, 36, 12–e24743. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.M.; Wyres, K.L.; Duchêne, S.; Wick, R.R.; Judd, L.M.; Gan, Y.H.; Hoh, C.H.; Archuleta, S.; Molton, J.S.; Kalimuddin, S.; Koh, T.H. Population genomics of hypervirulent Klebsiella pneumoniae clonal-group 23 reveals early emergence and rapid global dissemination. Nature communications. 2018, 9, 2703. [Google Scholar] [CrossRef] [PubMed]
- Lan, P.; Jiang, Y.; Zhou, J.; Yu, Y. A global perspective on the convergence of hypervirulence and carbapenem resistance in Klebsiella pneumoniae. Journal of global antimicrobial resistance. 2021, 25, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.S.; Opstrup, K.V.; Christiansen, G.; Rasmussen, P.V.; Thomsen, M.E.; Justesen, D.L.; Schønheyder, H.C.; Lausen, M.; Birkelund, S. Complement mediated Klebsiella pneumoniae capsule changes. Microbes and infection. 2020, 22, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, G.; Chao, X.; Xie, L.; Wang, H. The characteristic of virulence, biofilm and antibiotic resistance of Klebsiella pneumoniae. International journal of environmental research and public health. 2020, 17, 17–6278. [Google Scholar] [CrossRef] [PubMed]
- Diago-Navarro, E.; Calatayud-Baselga, I.; Sun, D.; Khairallah, C.; Mann, I.; Ulacia-Hernando, A.; Sheridan, B.; Shi, M.; Fries, B.C. Antibody-based immunotherapy to treat and prevent infection with hypervirulent Klebsiella pneumoniae. Clinical and Vaccine Immunology. 2017, 24, 1–e00456. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, X.; An, H.; Wang, J.; Ding, M.; Wang, L.; Li, L.; Ji, Q.; Qu, F.; Wang, H.; Xu, Y. Capsule type defines the capability of Klebsiella pneumoniae in evading Kupffer cell capture in the liver. PLoS Pathogens. 2022, 18, 8–e1010693. [Google Scholar] [CrossRef] [PubMed]
- Clegg, S.; Murphy, C.N. Epidemiology and virulence of Klebsiella pneumoniae. Urinary Tract Infections: Molecular Pathogenesis and Clinical Management, 2017; 435-57. [Google Scholar]
- Chapelle, C.; Gaborit, B.; Dumont, R.; Dinh, A.; Vallee, M. Treatment of UTIs due to Klebsiella pneumoniae carbapenemase-producers: how to use new antibiotic drugs? A narrative review. Antibiotics. 2021, 10, 1332. [Google Scholar] [CrossRef] [PubMed]
- Tarnagda, H.; Sagna, T.; Nadembega, W.M.; Ouattara, A.K.; Traoré, L.; Ouedraogo, R.A.; Bado, P.; Bazie, B.V.; Bouda, N.; Zongo, L.; Djigma, F.W. Prevalence and Antibiotic Resistance of Urinary Tract Pathogens, with Molecular Identification of Klebsiella pneumoniae, Klebsiella oxytoca, and Acinetobacter spp., Using Multiplex Real-Time PCR. American Journal of Molecular Biology. 2024, 14, 4–245. [Google Scholar] [CrossRef]
- Filev, R.; Lyubomirova, M.; Bogov, B.; Kolevski, A.; Pencheva, V.; Kalinov, K.; Rostaing, L. Urinary Tract Infections Caused by Klebsiella pneumoniae and Prolonged Treatment with Trimethoprim/Sulfamethoxazole. Microorganisms 2025, 13, 422. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.M.; Wick, R.R.; Watts, S.C.; Cerdeira, L.T.; Wyres, K.L.; Holt, K.E. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nature communications. 2021, 12, 1–4188. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.B.; Singh, B.B.; Priyadarshi, N.; Chauhan, N.K.; Rajamohan, G. Role of novel multidrug efflux pump involved in drug resistance in Klebsiella pneumoniae. PLoS One. 2014, 9, 5–e96288. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, Garima; Saigal, Saurabh; Elongavan, Ashok. Action and resistance mechanisms of antibiotics: A guide for clinicians. Journal of Anaesthesiology Clinical Pharmacology. 2017, 33, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, A.; Henquet, S.; Compain, F.; Genel, N.; Arlet, G.; Decré, D. Partition locus-based classification of selected plasmids in Klebsiella pneumoniae, Escherichia coli and Salmonella enterica spp.: An additional tool. Journal of Microbiological Methods. 2015, 110, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.; Hansen, L.H.; Sørensen, S.J. Conjugative plasmids: vessels of the communal gene pool. Philosophical Transactions of the Royal Society B: Biological Sciences. 2009, 364, 1527–2275. [Google Scholar] [CrossRef] [PubMed]
- Doménech-Sánchez, A.; Martínez-Martínez, L.; Hernández-Allés, S.; del Carmen Conejo, M.; Pascual, A.; Tomás, J.M.; Albertí, S.; Benedí, V.J. Role of Klebsiella pneumoniae OmpK35 porin in antimicrobial resistance. Antimicrobial agents and chemotherapy. 2003, 47, 3332–5. [Google Scholar] [CrossRef] [PubMed]
- Doorduijn, D.J.; Rooijakkers, S.H.; van Schaik, W.; Bardoel, B.W. Complement resistance mechanisms of Klebsiella pneumoniae. Immunobiology. 2016, 221, 1102–9. [Google Scholar] [CrossRef] [PubMed]
- Sedrakyan, A.; Gevorgyan, Z.; Zakharyan, M.; Arakelova, K.; Hakobyan, S.; Hovhannisyan, A.; Aminov, R. Molecular Epidemiology and In-Depth Characterization of Klebsiella pneumoniae Clinical Isolates from Armenia. International Journal of Molecular Sciences. 2025, 26, 504. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Li, Y.; Ren, P.; Tian, D.; Chen, W.; Fu, P.; Wang, W.; Li, X.; Jiang, X. Molecular epidemiology of hypervirulent carbapenemase-producing Klebsiella pneumoniae. Frontiers in Cellular and Infection Microbiology. 2021, 11, 661218. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, R.; Khanna, N.R.; Safadi, A.O.; et al. Bacitracin Topical. In: StatPearls. Treasure Island (FL): StatPearls Publishing. 2025. Available online: https://www.ncbi.nlm.nih.gov/books/NBK536993/ (accessed on 20 June 2025).
- Mezzatesta, M.L.; La Rosa, G.; Maugeri, G.; Zingali, T.; Caio, C.; Novelli, A.; Stefani, S. In vitro activity of fosfomycin trometamol and other oral antibiotics against multidrug-resistant uropathogens. International Journal of Antimicrobial Agents. 2017, 49, 763–6. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Héritier, C.; Tolün, V.; Nordmann, P. Emergence of oxacillinase-mediated resistance to imipenem in Klebsiella pneumoniae. Antimicrobial agents and chemotherapy. 2004, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Popescu, G.-A.; Codiță, I.; Szekely, E.; Șerban, R.; Ruja, G.; Tălăpan, D. Ghid privind Enterobacteriaceele producătoare de carbapenemaze. 2016. ISBN 978-973-0-22283-8.
- Yang, S.K.; Yusoff, K.; Ajat, M.; Thomas, W.; Abushelaibi, A.; Akseer, R.; Lim, S.H.; Lai, K.S. Disruption of KPC-producing Klebsiella pneumoniae membrane via induction of oxidative stress by cinnamon bark (Cinnamomum verum J. Presl) essential oil. PloS one. 2019, 14, e0214326. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Li, K.; Ji, Y.; Jiang, Q.; Wang, Y.; Shi, M.; Mi, Z. Carbapenem and cefoxitin resistance of Klebsiella pneumoniae strains associated with porin OmpK36 loss and DHA-1 β-lactamase production. Brazilian Journal of Microbiology. 2013, 44, 435–42. [Google Scholar] [CrossRef] [PubMed]
- Murri, R.; Fiori, B.; Spanu, T.; Mastrorosa, I.; Giovannenze, F.; Taccari, F.; Palazzolo, C.; Scoppettuolo, G.; Ventura, G.; Sanguinetti, M.; Cauda, R. Trimethoprim–sulfamethoxazole therapy for patients with carbapenemase-producing Klebsiella pneumoniae infections: retrospective single-center case series. Infection. 2017, 45, 209–13. [Google Scholar] [CrossRef] [PubMed]
- Motley, M.P.; Fries, B.C. A new take on an old remedy: generating antibodies against multidrug-resistant Gram-negative bacteria in a postantibiotic world. MSphere. 2017, 2, 10–128. [Google Scholar] [CrossRef] [PubMed]
- Zaki, B.M.; Hussein, A.H.; Hakim, T.A.; Fayez, M.S.; El-Shibiny, A. Phages for treatment of Klebsiella pneumoniae infections. Progress in Molecular Biology and Translational Science. 2023, 200, 207–39. [Google Scholar] [PubMed]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals. 2013, 6, 1451–74. [Google Scholar] [CrossRef] [PubMed]
- El-Demerdash, A.S.; Alfaraj, R.; Farid, F.A.; Yassin, M.H.; Saleh, A.M.; Dawwam, G.E. Essential oils as capsule disruptors: enhancing antibiotic efficacy against multidrug-resistant Klebsiella pneumoniae. Frontiers in Microbiology. 2024, 15, 1467460. [Google Scholar] [CrossRef] [PubMed]
- Baptista, C.T.; Cerveira, M.M.; Barboza, V.; Maria, E.; Ferrer, K.; Miller, R.G.; Souza, T.D.; Zank, P.D.; Blanke, A.D.; Klein, V.P.; Rosado, R.P. A Systematic Review of Essential Oils’ Antimicrobial and Antibiofilm Activity against Klebsiella pneumoniae. Current Research in Complementary & Alternative Medicine, 2022; 6, 2577–201. [Google Scholar]
- Fournomiti, M.; Kimbaris, A.; Mantzourani, I.; Plessas, S.; Theodoridou, I.; Papaemmanouil, V.; Kapsiotis, I.; Panopoulou, M.; Stavropoulou, E.; Bezirtzoglou, E.E.; Alexopoulos, A. Antimicrobial activity of essential oils of cultivated oregano (Origanum vulgare), sage (Salvia officinalis), and thyme (Thymus vulgaris) against clinical isolates of Escherichia coli, Klebsiella oxytoca, and Klebsiella pneumoniae. Microbial ecology in health and disease. 2015, 26, 23289. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Sun, Z.; Wang, T.; Yang, M.; Liu, M.; Zhang, J.; Li, Y. Antimicrobial activity of eugenol against carbapenem-resistant Klebsiella pneumoniae and its effect on biofilms. Microbial pathogenesis. 2020, 139, 103924. [Google Scholar] [CrossRef] [PubMed]
- Bisso Ndezo, B.; Tokam Kuaté, C.R.; Dzoyem, J.P. Synergistic antibiofilm efficacy of thymol and piperine in combination with three aminoglycoside antibiotics against Klebsiella pneumoniae biofilms. Canadian Journal of Infectious Diseases and Medical Microbiology. 2021, 1, 7029944. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Rajivgandhi, G.N.; Ramachandran, G.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Almanaa, T.N.; Manoharan, N. Preparative HPLC fraction of Hibiscus rosa-sinensis essential oil against biofilm forming Klebsiella pneumoniae. Saudi Journal of Biological Sciences. 2020, 27, 2853–62. [Google Scholar] [CrossRef] [PubMed]
- Bilcu, M.; Grumezescu, A.M.; Oprea, A.E.; Popescu, R.C.; Mogoșanu, G.D.; Hristu, R.; Stanciu, G.A.; Mihailescu, D.F.; Lazar, V.; Bezirtzoglou, E.; et al. Efficiency of Vanilla, Patchouli and Ylang Ylang Essential Oils Stabilized by Iron Oxide@C14 Nanostructures against Bacterial Adherence and Biofilms Formed by Staphylococcus aureus and Klebsiella pneumoniae Clinical Strains. Molecules. 2014, 19, 17943–17956. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
K. pneumoniae strain’s antibiotic susceptibility variation.
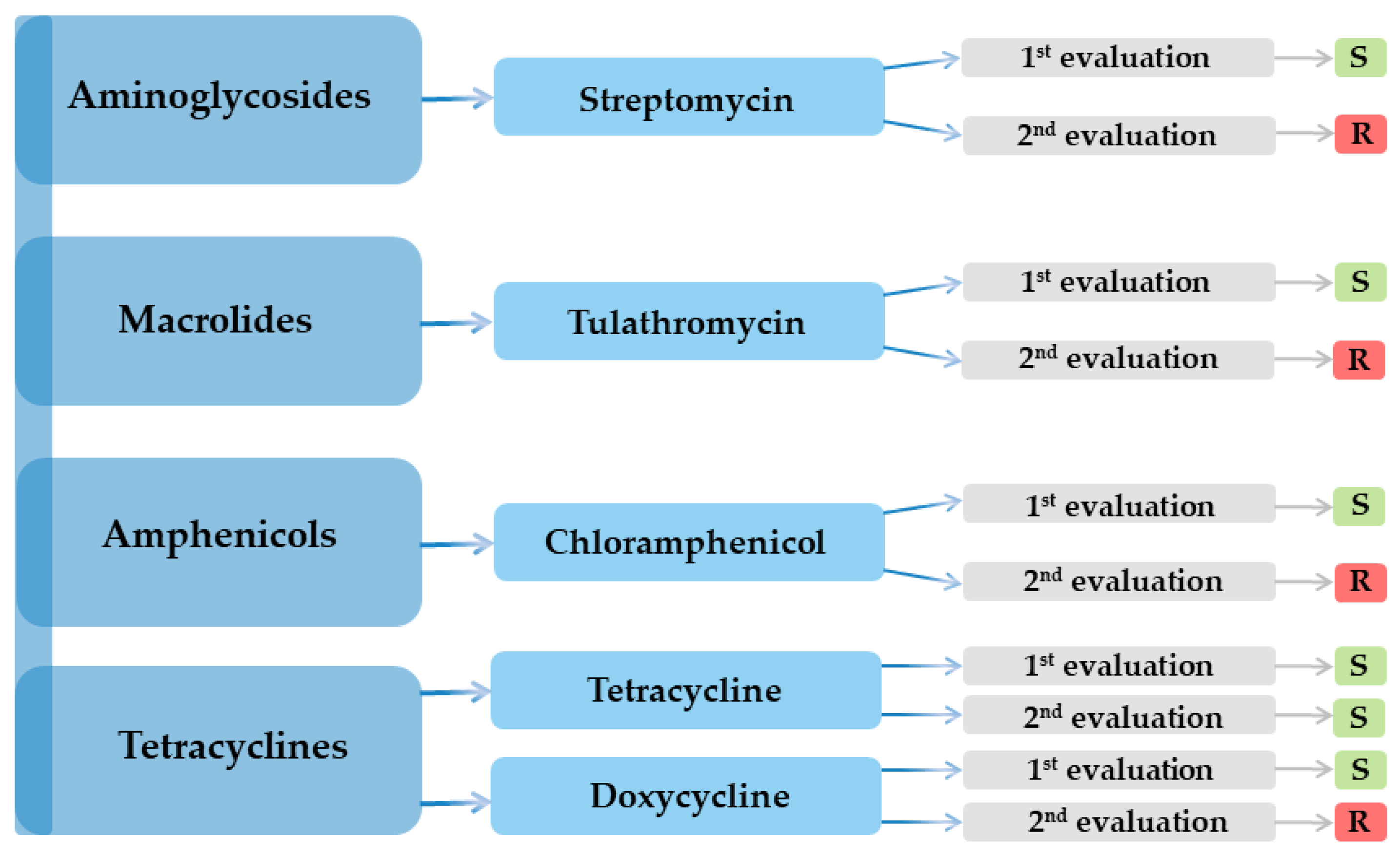

Table 1.
Antibiotic susceptibility profile of the K. pneumoniae isolate.
| Antimicrobial class | Nr. | Antibiotic | Result | |
|---|---|---|---|---|
| 1st evaluation | 2nd evaluation | |||
| Aminoglycosides | 1 | Amikacin (AK) | R | R |
| 2 | Streptomycin (S) | S | R | |
| 3 | Gentamicin (GME) | R | R | |
| Penicillins | 4 | Penicillin (PEN) | R | R |
| 5 | Oxacillin (OX) | R | R | |
| 6 | Ampicillin–sulbactam (SAM) | R | R | |
| 7 | Piperacillin (PRL) | R | R | |
| 8 | Mecillinam (MEC) | R | R | |
| 9 | Amoxicillin–clavulanic acid (AMC) | R | R | |
| Macrolides | 10 | Azithromycin (AZM) | R | R |
| 11 | Clarithromycin (CLR) | R | R | |
| 12 | Tulathromycin (TUL) | S | R | |
| 13 | Spiramycin (SP) | R | R | |
| Polypeptides | 14 | Bacitracin (DTD) | R | R |
| Polymyxins | 15 | Colistin (CT) | R | R |
| Phosphonic acid derivatives | 16 | Fosfomycin (FF) | R | R |
| Cephalosporins ⮚ 1st generation |
17 | Cefadroxil (CFR) | R | R |
| 18 | Cephradine (CE) | R | R | |
| 19 | Cefacetrile (CEF) | R | R | |
| 20 | Cephalothin (CH) | R | R | |
| ⮚ 2nd generation | 21 | Cefaclor (CEC) | R | R |
| 22 | Cefotetan (CTT) | R | R | |
| 23 | Cefuroxime (CXM) | R | R | |
| 24 | Cefamandole (MA) | R | R | |
| ⮚ 3rd generation | 25 | Ceftriaxone (CRO) | R | R |
| 26 | Ceftazidime (CAZ) | R | R | |
| 27 | Cefoperazone (CEP) | R | R | |
| 28 | Ceftiofur (FUR) | R | R | |
| 29 | Cefixime (CFM) | R | R | |
| 30 | Ceftazidime-avibactam (CZA) | R | R | |
| ⮚ 4th generation | 31 | Cefquinome (CFQ) | R | R |
| Carbapenems | 32 | Meropenem (MEM) | R | R |
| 33 | Imipenem (IMI) | R | R | |
| Streptogramins | 34 | Pristinamycin (PT) | R | R |
| Aminocoumarins | 35 | Novobiocin (NV) | R | R |
| Fluoroquinolones | 36 | Norfloxacin (NX) | R | R |
| 37 | Ofloxacin (OFX) | R | R | |
| 38 | Marbofloxacin (MAR) | R | R | |
| Tetracyclines | 39 | Tetracycline (TET) | S | S |
| 40 | Doxycycline (DOX) | S | R | |
| Rifamycins | 41 | Rifampicin (RD) | R | R |
| Amphenicols | 42 | Chloramphenicol (C) | S | R |
| Fusidic acid | 43 | Fusidic acid (FA) | R | R |
| Chemotherapeutic agents | 44 | Furazolidone (FX) | R | R |
| 45 | Nitrofurantoin (F) | R | R | |
| 46 | Metronidazole (MET) | R | R | |
| Sulfonamides | 47 | Sulfonamide compound (S3) | R | R |
| 48 | Trimethoprim–sulfamethoxazole (SXT) | R | R | |
R - resistant; S – susceptible.
Table 2.
Evolution of K. pneumoniae CFU mL−1.
| 06 Feb 2024 – Urine culture with antibiogram: > 100,000 CFU mL−1, K. pneumoniae | ||
|---|---|---|
| Susceptible to three agents: colistin, gentamicin, and fosfomycin. | Resistance to 17 agents: amikacin, ampicillin, amoxicillin–clavulanic acid, aztreonam, ceftazidime, cefuroxime, cefepime, cefotaxime, ciprofloxacin, imipenem, nitrofurantoin, piperacillin, tobramycin, trimethoprim–sulfamethoxazole, piperacillin–tazobactam, meropenem, and ertapenem. |
Observations Treatment with fosfomycin Over time, the strain became resistant to fosfomycin. |
| 05 Mar 2024 – Urine culture: < 1,000 CFU mL−1; not clinically significant. | ||
| 23 Sep 2024 – Urine culture: < 1 000 CFU mL−1; not clinically significant. | ||
| 22 Nov 2024 – Urine culture with antibiogram: > 100,000 CFU mL−1, K. pneumoniae | ||
|
Susceptible to four agents: amikacin, amoxicillin–clavulanic acid, gentamicin, and trimethoprim–sulfamethoxazole Intermediate susceptibility to two agents: ceftazidime and cefepime |
Resistant to two agents: piperacillin–tazobactam and levofloxacin | Treatment with trimethoprim–sulfamethoxazole |
| 13 Dec 2024 – Urine culture with antibiogram: > 100,000 CFU mL−1, K. pneumoniae | ||
| Susceptible to two agents: colistin and tigecycline | Resistant to 14 agents: amikacin, aztreonam, ceftazidime, cefepime, cefotaxime, ciprofloxacin, gentamicin, imipenem, tobramycin, trimethoprim–sulfamethoxazole, piperacillin–tazobactam, meropenem, levofloxacin, and ertapenem | Over time, the strain became resistant to trimethoprim–sulfamethoxazole |
| 26 Feb 2025 – Urine culture: < 1 000 CFU mL−1; not clinically significant. | ||
| 25 Apr 2025 – Urine culture with antibiogram: 10,000–100,000 CFU mL−1, K. pneumoniae | ||
| Susceptible to two agents: gentamicin and trimethoprim–sulfamethoxazole | Resistant to ten agents: ampicillin, amoxicillin–clavulanic acid, ceftazidime, ceftriaxone, cefpodoxime, ciprofloxacin, meropenem, levofloxacin, ertapenem, and cefixime | Susceptibility to trimethoprim–sulfamethoxazole is once again observed. |
Table 3.
Susceptibility/resistance of the K. pneumoniae isolate to essential oils.
| International name | Latin name/composition | Inhibition area diameter (mm) |
|---|---|---|
| Palmarosa | Cymbopogon martini | 11 |
| Geranium | Pelargonium graveolens | R |
| Frankincense | Boswellia carteri | R |
| Laurel | Laurus nobilis | 12 |
| Tea tree | Melaleluca alternifolia | 20 |
| Citronella | Cymbopogon nardus | R |
| Thyme | Thymus vulgaris | 22 |
| Propolis | Apis mellifera propolis | R |
| Biomicin urinar® (A20) |
Origani aetheroleum + Cinnamomun verum + Salvia officinalis + Thymi aetheroleum |
12 (resistant colonies) |
| Biomicin forte® (A3) |
Thymi aetheroleum + Caryophylli floris aetheroleum |
30 |
| Methylene blue 3% | - | 13 |
R-resistant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



