
Submitted:
09 July 2025
Posted:
11 July 2025
You are already at the latest version
Abstract
Prostate cancer is a major cause of cancer-related deaths among men in Sub-Saharan Africa, where late-stage diagnoses are common due to limited access to affordable and sensitive diagnostic tools. Early detection is essential to improve survival and reduce the disease burden. This review explores the integration of epigenetic biomarkers and CRISPR-Cas12a technology as a transformative approach for early, non-invasive prostate cancer detection in resource-limited settings. Among the many complexities of cancer development, molecular dysregulation plays a remarkable role, and epigenetic modifications such as DNA methylation, histone changes, and non-coding RNA expression have emerged as stable and specific biomarkers with significant potential for the early detection and characterisation of prostate carcinogenesis. However, their low concentration in body fluids presents a detection challenge. CRISPR-Cas12a, known for its high specificity and sensitivity, offers a promising solution. When combined with isothermal amplification and liquid biopsy techniques, it enables rapid, low-cost, and point-of-care diagnostics. This review proposes a low-cost, CRISPR-Cas12a-based diagnostic pipeline for detecting prostate cancer-specific epigenetic markers in liquid biopsies. The implementation of this technology in Sub-Saharan Africa could significantly improve early diagnosis, reduce mortality, and advance health equity.
Keywords:
epigenetic signatures
; CRISPR-Cas12a
; prostate cancer
; early detection
; liquid biopsy
; Sub-Saharan Africa
1. Introduction
Prostate cancer remains one of the leading causes of cancer-related deaths among men globally, with marked disparities in incidence, mortality, and healthcare access between developed and developing regions, particularly Sub-Saharan Africa [1]. In this region, prostate cancer is characterised by alarmingly high incidence and mortality rates, largely due to late-stage diagnoses. Projections indicate that by 2040, the number of prostate cancer cases in Sub-Saharan Africa could more than double, significantly increasing the burden of disease [2,3]. In Nigeria, for instance, prostate cancer is the most frequently diagnosed cancer among men, with an estimated 7.4% of the male population at risk of developing the disease before the age of 75 [4]. The high prevalence of late-stage diagnoses is often attributed to limited access to healthcare, inadequate public awareness, and the absence of widespread screening programs [5,6]. Cultural and socioeconomic barriers, such as stigma and misconceptions about cancer, further delay timely medical intervention, contributing to poorer clinical outcomes [7]. In addition to these systemic challenges, biological factors, including genetic predispositions and environmental influences, are believed to contribute to the more aggressive forms of prostate cancer observed in men of African descent [8]. Among the genetic disorders, epigenetic modifications, such as DNA methylation and histone changes, play a critical role in prostate carcinogenesis and progression. These molecular changes hold potential as stable, specific biomarkers for early detection, particularly in minimally invasive liquid biopsy samples [9]. However, tumour-derived nucleic acids in such samples are often present at low concentrations, necessitating highly sensitive detection platforms. In this context, CRISPR-Cas12a (Cpf1) technology has emerged as a powerful diagnostic tool due to its programmability, high sensitivity, specificity, and suitability for low-resource settings [10]. When combined with the detection of epigenetic signatures, CRISPR-based assays can facilitate the identification of circulating tumour nucleic acids (CTNAs) from minimally invasive samples, such as blood, urine, or saliva [11]. This combination approach offers significant promise for improving early prostate cancer detection while addressing the limitations of prostate-specific antigen (PSA) testing, which often lacks specificity and may result in overdiagnosis and overtreatment [12]. The integration of epigenetic biomarkers with CRISPR-based diagnostics represents a particularly compelling approach for advancing precision oncology [10]. This review examines the synergistic application of epigenetic signatures and CRISPR-Cas12a diagnostics, highlighting their transformative potential for enhancing prostate cancer screening and diagnosis in Sub-Saharan Africa.
2. Epigenetic Signatures in Prostate Cancer
2.1. Overview of Epigenetics
Epigenetics is the study of heritable changes in gene expression that do not involve alterations to the underlying DNA sequence. These modifications include DNA methylation on cytosine bases and post-translational modifications of histone proteins such as methylation, acetylation, phosphorylation, and sumoylation [13]. Among these, DNA methylation is specifically the formation of 5-methylcytosine, a stable epigenetic mark that can persist through cell divisions and influence cellular phenotype across generations [13]. In normal mammalian cells, DNA methylation acts as a gene expression regulator. For instance, promoter regions of tumour suppressor genes such as TP53 and BRCA1 are typically unmethylated, allowing transcription factors and RNA polymerase to access DNA and activate gene expression [14] (Figure 1A). Conversely, hypermethylation of these promoter regions leads to chromatin condensation and the silencing of gene expression, preventing transcription factor binding and facilitating oncogenesis [14] (Figure 1B).
So, in cancer biology, epigenetics plays a crucial role in tumorigenesis, influencing processes such as cell differentiation, proliferation, and apoptosis. Aberrant epigenetic modifications can lead to the silencing of tumour suppressor genes or the activation of oncogenes, thereby contributing to cancer development and progression [15]. The significance of epigenetics in cancer biology is underscored by its potential for therapeutic intervention. Epigenetic alterations, such as DNA methylation and histone modifications, can serve as biomarkers for cancer diagnosis and prognosis. Moreover, the development of epigenetic therapies, which aim to reverse these modifications, has emerged as a promising strategy in oncology [16,17]. These therapies are particularly relevant in the context of drug resistance, where epigenetic changes can confer a survival advantage to cancer cells [18].
2.1.1. Types of Epigenetic Modifications
DNA Methylation
DNA methylation involves the enzymatic addition of methyl groups to the fifth carbon of cytosine residues within CpG dinucleotides, forming 5-methylcytosine [19]. This process is catalysed by DNA methyltransferases (DNMTs), which are classified functionally as "writers". Other associated proteins include "readers" such as methyl-binding domain proteins and zinc finger proteins, which interpret methylation marks to regulate gene expression, and "erasers," including the ten-eleven translocation (TET) enzymes that mediate active DNA demethylation. The TET enzymes oxidise 5-methylcytosine to form 5-hydroxymethylcytosine (5hmC), and further to 5-formylcytosine (5fC) and 5-carboxycytosine (5caC) [20]. These oxidised intermediates can be removed by base excision repair mechanisms involving thymine DNA glycosylase, ultimately restoring unmethylated cytosine (Figure 2).
Histone Modification
In eukaryotic cells, histones are key proteins that package DNA into nucleosomes, the fundamental units of chromatin [22]. Post-translational modifications of histones, including acetylation, methylation, phosphorylation, and ubiquitination, regulate chromatin structure and gene accessibility [23]. These modifications alter the interaction between histones and DNA, affecting the chromatin structure and gene accessibility. Histone acetylation, mediated by histone acetyltransferases (HATs), neutralises positive charges on histones and relaxes chromatin structure, promoting transcription (Figure 3). Histone deacetylases (HDACs) reverse this process, resulting in chromatin condensation and transcriptional repression [24]. Similarly, histone methyltransferases and demethylases regulate the addition and removal of methyl groups, influencing transcription in a position-dependent manner [25]. Histone modifications are dynamically regulated and are crucial for maintaining cellular identity and function [26].
Non-Coding RNAs Mediated Regulation
Non-coding RNAs such as microRNA (miRNA) and long non-coding RNAs (lncRNAs) are functional RNA transcripts which are not translated into proteins [27]. They significantly contribute to the regulation of gene expression at transcription and post-transcriptional levels. miRNAs are 20 – 30 nucleotides long and can bind to complementary sequences on target mRNAs, leading to their degradation or translational inhibition [28]. lncRNAs, which exceed 200 nucleotides, modulate gene expression by interacting with chromatin-modifying complexes, altering chromatin architecture, or recruiting transcriptional co-repressors [29]. These non-coding RNAs are increasingly recognised as important regulators of cancer-related gene expression and potential targets for diagnostic and therapeutic strategies.
2.2. Common Epigenetic Markers Identified in Prostate Cancer
Alterations in epigenetic mechanisms play a significant role in prostate cancer development and progression [30], and many epigenetic aberrations in key genes have been identified (Table 1). One of the most prominent epigenetic markers in prostate cancer is DNA hypermethylation, particularly at tumour suppressor genes. Aberrant hypermethylation of genes involved in several cellular functions, ranging from cell cycle control, DNA repair, apoptosis, tumour invasion, signal transduction, tumour suppression and hormone response, are commonly observed in prostate cancer [31]. The GSTP1 gene, encoding for glutathione-S-transferase that plays a vital role in detoxification, is frequently methylated in over 90% of prostate cancer cases and rarely in normal prostate tissue, making it a highly specific marker for PCa [32,33]. Also, hypermethylation of the RASSF1A gene, a tumour suppressor gene, has been associated with tumour aggressiveness. Pidsley and colleagues [34] reported hypermethylation of the CRACR2A gene in metastatic prostatic tissue. Other frequently reported hypermethylated genes in PCa include the APC, RARβ2, CDH13, RASSF1a, PTGS2, CD44, and E-Cadherin. These epigenetic modifications disrupt normal cellular homeostasis and promote malignant transformation.
In addition to DNA methylation, histone modifications contribute to prostate cancer epigenetics. Increased trimethylation of histone H3 at lysine 27 (H3K27me3) has been associated with more aggressive PCa phenotypes [35], and overexpression of SIRT7, a histone deacetylase, has also been reported in prostate tumours [36]. In addition, reduced acetylation of histone H3 at lysine 9 (H3K9ac) has been linked with tumour development [37]. Although histone modifications offer valuable insights into the underlying mechanisms of prostate cancer development, investigating them remains technically demanding and expensive, limiting their current applicability in clinical settings.
MicroRNAs (miRNAs), on the other hand, represent another class of epigenetic regulators with significant diagnostic potential in PCa. Dysregulated miRNA expression patterns are frequently observed in prostate cancer with diagnostic potential. For instance, miR-21 has shown high diagnostic accuracy, with a sensitivity of 91% and specificity of 89% in distinguishing metastatic prostate cancer, making it a promising serum biomarker [38]. Also, miR-18a-5p, miR-125 b and miR-4534 have been reported to be upregulated in prostate cancer [41].

Table 1.
Common epigenetic marks in prostate cancer.
| Epigenetic Maker | Gene category | Gene | Ref |
|---|---|---|---|
| DNA hypermethylation | DNA damage repair | GSTP 1 | [35] |
| MGMT | [41] | ||
| Cell adhesion | CDH1 | [42] | |
| T IMP3 | [43] | ||
| Tumour suppression and Apoptosis control |
RARβ2 | [44] | |
| APC | [45] | ||
| RASSF1 | [46] | ||
| CRACR2A | [34] | ||
| LGALS3 | [47] | ||
| DNA hypomethylation | Detoxification and hormone response | CYP1B1 | [48] |
| Tumor invasion | HPSE | [49] | |
| Histone modification | Increased methylation | H3K27me3 | [35] |
| Decreased methylation | SIRT7 | [36] | |
| Decreased acetylation | H3K9ac | [37] | |
| miRNAs | Upregulation | MicroRNA-21 | [38] |
| MicroRNA-18a-5p | [39] | ||
| MicroRNA-4534 | [40] | ||
| MicroRNA-375 | [50] |
GSTP 1: Glutathione S-Transferase Pi 1; MGMT: O-6-Methylguanine-DNA Methyltransferase; CDH1: Cadherin 1 (E-cadherin); TIMP3: Tissue Inhibitor of Metalloproteinases 3;RASSF1: Ras Association Domain Family Member 1; RARβ2: Retinoic Acid Receptor Beta 2; APC: Adenomatous Polyposis Coli; CRACR2A: Calcium Release Activated Channel Regulator 2A; LGALS3: Galectin-3 (Lectin, Galactoside-Binding Soluble 3); AR: Androgen Receptor; ER: Estrogen Receptor; CYP1B1: Cytochrome P450 Family 1 Subfamily B Member 1; HPSE: Heparanase; H3K27me3: Trimethylation of Lysine 27 on Histone H3; SIRT7: Sirtuin 7; H3K9ac: Acetylation of Lysine 9 on Histone H3.
Relevance to Early Detection: Stability, Specificity, and Detectability in Biofluids
The gold standard marker for prostate cancer, prostate-specific antigen (PSA), is highly sensitive but less specific and imprecise, as it can’t reliably distinguish between prostate cancer and other conditions affecting the prostate, like prostatitis or benign prostatic hyperplasia [51]. The false positives generated by PSA tests often result in costly and invasive procedures, such as transrectal prostate biopsies and underscore the need for more accurate and non-invasive biomarkers [52]. Epigenetic markers offer a promising alternative due to their high specificity, detectability in liquid biopsies (e.g., urine and blood), and potential for early-stage detection [31]. Unlike protein-based biomarkers, epigenetic changes such as DNA methylation and miRNA expression are stable(58), can be measured quantitatively or qualitatively[55], and remain intact in diverse sample conditions, including EDTA-treated blood stored at 4°C, –20°C, or –80°C, as well as dried blood spots [56]. Aside from the stability of biomarkers in clinical practice, a crucial issue in biomarker assessment is clinical evidence of the test performance and affordability, which are critical to ensure the incorporation of new biomarkers in the clinical setting [57]. Furthermore, despite the emergence of novel epigenetic markers, there is a need to develop and adapt methods for the analysis of specific markers [58]. Several assays have been designed for the detection of epigenetic markers, one of which is the ProCaMTM methylation assay, which quantifies methylation of GSTP1, RARβ2 and APC in urine samples. In a cohort of 320 PCa patients and 384 controls, the assay demonstrated a sensitivity of 60% and specificity of 80% [59]. Notably, the ConfirmMDx assay, developed by MDxHealth, remains the only clinically approved methylation test for PCa. It assesses methylation of GSTP1, RASSF1, and APC in histologically negative prostate biopsies, aiding in the decision-making process regarding repeat biopsies [60].
2.3. Epigenetic Biomarkers in Sub-Saharan Populations
Research on epigenetic alterations that occur in various diseases, including PCa in African populations, is limited. Most studies on epigenetic markers for PCa are carried out on European and Asian populations, and there is a notable lack of data on the epigenetic alterations specific to African populations [61]. Although research on epigenetic markers in African populations remains limited, the field of epigenetics provides crucial insights into the complex interactions between genetic predispositions and environmental factors, particularly in the context of African populations. However, research targeting the unique epigenetic landscapes in these populations remains notably limited. One primary issue within the current landscape of epigenetic research in Africa is the overwhelming lack of studies focusing specifically on African populations. Oladipo et al. emphasise that the majority of epigenetic investigations have predominantly concentrated on individuals of European, Asian, and American backgrounds, leading to a significant oversight of potential epigenetic variations present in African ancestral groups [62]. The specific genetic and epigenetic mechanisms, such as the variation of cytosine modifications, could greatly influence the prevalence of conditions like cancer, which disproportionately affect minority groups in the U.S., including African Americans [63]. This indicates the necessity of expanding research to better understand the unique epigenetic factors at play within African populations. DNA methylation is associated with non-communicable diseases in a black South African cohort, demonstrating how African populations may exhibit unique epigenomic profiles that are not captured in existing global literature [64]. Furthermore, Africa's vast genetic diversity and varying environmental exposures present a unique opportunity to investigate environmental effects on epigenetic profiles; yet this potential remains underexplored due to a lack of localised research initiatives [65]. These findings accentuate an urgent need for comprehensive epigenomic studies designed to reflect the genetic and environmental complexities of African populations.
Additionally, pathogen-induced epigenetic alterations highlight the urgent need for research into how infectious diseases prevalent in Africa may contribute to cancer development through epigenetic mechanisms [66]. Conducting localised studies not only contributes to filling the knowledge void but also aids in developing tailored prevention and treatment strategies that address the unique health challenges faced by these populations.
3. CRISPR-Cas12a Technology for DNA Detection
CRISPR is an isothermal technique first developed for gene editing technology, and coupled with an amplification technique, it can be used for diagnostic purposes. It uses three principal elements, including the endonucleases called “Cas” enzymes (Cas9, Cas12a,b; Cas13;..)[67], the main role is to cut the target sequence at the specific site. The guide RNA (crRNA or gRNA), on the other hand, helps the endonuclease “Cas” enzyme to bind to the target sequence. It has two main parts: the trans-activating CRISPR guide RNA (tracrRNA) that will form a complex with the Cas Enzyme, and the CRISPR guide RNA site (crRNA) that will form a complex with the target sequence. These enzymes have been used to develop novel nucleic acid assays, where the activated Cas-crRNA complex can indiscriminately cleave surrounding non-target single-stranded nucleic acid reporters (labelled probes).
3.1. Mechanism of CRISPR-Cas12a
Cas12 enzymes are used in CRISPR-based diagnostics to target dsDNA and ssDNA, requiring a PAM site for dsDNA cleavage and collateral cleavage of ssDNA. DETECTR, one of the first Cas12-based detection methods, uses Cas12a to target dsDNA and trigger collateral cleavage of short ssDNA reporters carrying a fluorophore and a quencher to reveal the detection as seen in Figure 4 [67].
3.2. Workflow of CRISPR-Cas12a-Based Nucleic Acid Detection in Liquid Biopsies
Practically after Sample Collection (Urine, swab, blood, or other clinical specimen is collected), Nucleic acids are extracted using a rapid method (e.g., magnetic beads or spin columns). For field or low-resource settings, crude extraction buffers may be used. Following this step, the target gene is amplified to Enhance Sensitivity by using isothermal amplification (RPA or LAMP) targeting a specific pathogen gene or biomarker [67]. This step can take 10 to 30 minutes and works at a constant temperature (no thermocycler needed). So, after the pre-amplification of the target gene, the CRISPR reaction is set up by combining in a single tube: Cas12a enzyme, Specific crRNA, ssDNA reporter molecule with fluorophore-quencher or biotin-FAM for lateral flow and the Amplified product (or directly extracted DNA if the target is abundant). For the recognition and the activation, if the target DNA is present, it binds the crRNA-guided Cas12a complex, and Cas12a is activated and starts collateral cleavage of ssDNA reporters. This signal is read out either via fluorometric detection, in which the fluorescence is released and detected using a portable reader or smartphone-based device, or colourimetric via enzyme trans-cleavage and the release of LacZa [68].
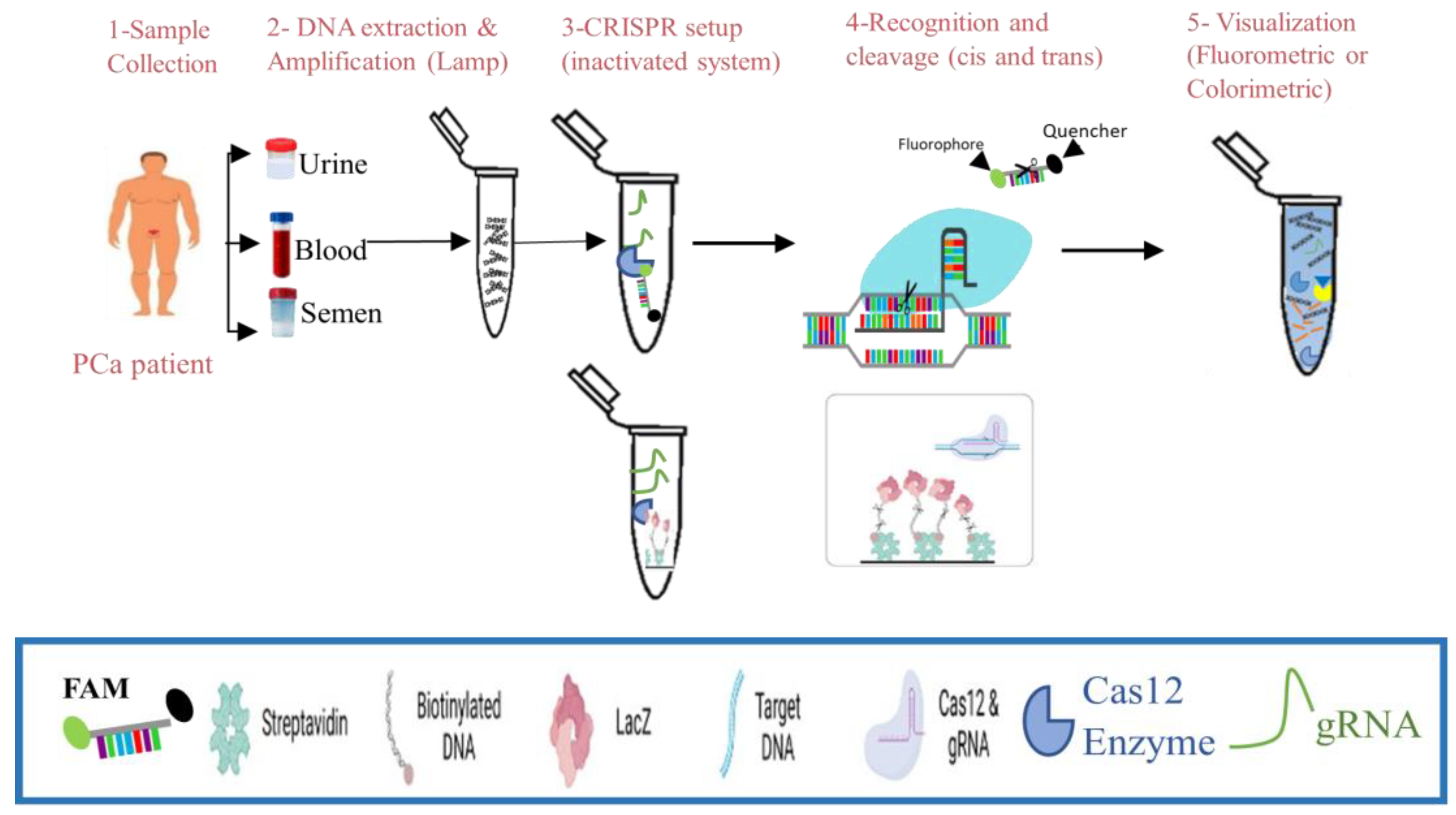
3.3. Advances in CRISPR Diagnostics in Cancer
The advent of CRISPR-Cas12 technology among the different CRISPR systems has revolutionised various fields of molecular diagnostics, particularly in detecting mutations associated with various cancers. For instance, the system's application in detecting circulating EGFR mutations, which are critical in non-small cell lung cancer management, indicates that the CRISPR-Cas12a assay exhibits high sensitivity and could perform rapid evaluations of mutations, suggesting promising implications for real-time monitoring of cancer biomarkers [10]. Similarly, the same approach could be used for visual quantification of single-nucleotide variants (SNVs) associated with cancer, showcasing how the technology's adaptability can maintain high sensitivity and specificity across different cancer types [69]. By targeting various cancer-associated mutations and leveraging the unique properties of the CRISPR system, including its collateral cleavage activity, CRISPR-Cas12 is recognised as a highly reliable tool for diagnosing cancer-related alterations [70]. The system has the potential for detecting epigenetic changes in cancer, such as prostate cancer, particularly DNA methylation, as it is becoming an area of significant interest. Alterations in DNA methylation patterns can serve as early biomarkers for cancer detection [71]. In urologic cancers, including prostate cancer, epigenetic alterations, due to their stability and accessibility in body fluids, have promise as cancer biomarkers [71]. These epigenetic markers could be easily detected via the CRISPR-Cas12a system. For instance, systems integrating CRISPR-Cas12a with techniques like polymerase amplification could significantly enhance the sensitivity of detecting epigenetic modifications, enabling the identification of specific methylation patterns associated with cancer, like prostate cancer [72]. Alongside this, CRISPR technology can be engineered to identify and quantify microRNA expression, which is often altered in cancer states [73]. This dual approach could allow for a comprehensive diagnostic strategy encompassing both genetic and epigenetic alterations. Furthermore, notable advancements in CRISPR diagnostics indicate that localised, efficient detection methods could be developed for point-of-care testing, reducing dependencies on complex laboratory environments[74]. The ability of CRISPR-Cas12 systems to facilitate rapid and accurate detection, especially of epigenetic markers in prostate cancer, represents a significant step forward in personalised medicine and cancer management.
4. Synergistic Application: Epigenetics and CRISPR-Cas12a
4.1. Workflow Integration
Using CRISPR-Cas12a to detect methylation patterns or non-coding RNAs in biofluids.
The integration of CRISPR-Cas12a and methylation-sensitive restriction enzymes (MSREs) provides a precise and amplification-free method to detect DNA methylation at single CpG resolution. The workflow follows a stepwise strategy that leverages the high specificity of MSREs for non-methylated DNA and the collateral cleavage activity of Cas12a for signal amplification [75].
The first step involves identifying CpG sites of clinical interest within the genome. A suitable protospacer adjacent motif (PAM) sequence (typically TTTV) must be present near the CpG site to enable Cas12a binding. crRNAs are then designed to hybridise to a region adjacent to the CpG, ensuring maximum R-loop formation and sensitivity. Optimal designs favour crRNA targeting regions proximal to the CpG site for the highest sensitivity in methylation detection.
Secondly, extracted genomic DNA is then treated with a specific Methylated Sensitive Restriction Enzyme (e.g., AciI, HpaII), which selectively cleaves unmethylated CpG-containing sequences. Methylated sequences remain intact after enzyme digestion, preserving the Cas12a target site.
Thirdly, the Cas12a protein is pre-incubated with the designed crRNA to form a ribonucleoprotein (RNP) complex. This RNP is mixed with the digested DNA sample and a reporter molecule (typically a single-stranded DNA labelled with a fluorophore and quencher pair). If the DNA is methylated and intact (not cleaved by MSRE), it will bind to the crRNA-Cas12a complex, activate trans-cleavage, and cleave the reporter, resulting in a measurable fluorescence signal.
Finally, for the detection and quantification of CpG methylated sites, the fluorescence could be monitored over time using a plate reader. The rate of fluorescence increase corresponds to the degree of Cas12a activation, which reflects the methylation status of the target site. Calibration with known methylated-to-unmethylated DNA ratios allows for quantitative assessment of methylation percentage. This system allows accurate detection of single-site methylation events without bisulfite conversion or PCR amplification. The emergence of CRISPR-Cas12 technology brings a new era in molecular diagnostics, particularly when compared to traditional methods such as quantitative Polymerase Chain Reaction (qPCR) and Next-Generation Sequencing (NGS). CRISPR-Cas systems, especially Cas12, offer notable advantages in terms of sensitivity, specificity, and operational simplicity over these conventional techniques. Traditional nucleic acid detection methods like qPCR and NGS rely heavily on amplification processes to detect target sequences. qPCR is known for its quantitative capabilities but often has limitations regarding sensitivity, particularly at low nucleic acid concentration [76]. In contrast, CRISPR-Cas12 technology exhibits collateral cleavage activity, which can significantly enhance signal detection without necessitating extensive amplification [77,78]. This allows for rapid detection in a simplified format, which is increasingly vital during outbreaks or early diagnosis, where time-to-results is critical. Cas12-based assays have demonstrated limits of detection (LOD) that can surpass those of qPCR, with reports suggesting they can detect nucleic acid at concentrations lower than traditional methods [79].
4.2. Implementation in Resource-Limited Settings
The advent of CRISPR-based diagnostics represents a significant advancement in molecular biology and has the potential to revolutionise cancer (prostate cancer) screening and monitoring, especially in resource-limited settings[80]. The portability, cost-effectiveness, and simplicity of CRISPR-based systems, particularly CRISPR-Cas12 assays, make them attractive candidates for decentralised prostate cancer screening initiatives in rural areas. CRISPR diagnostic technologies, particularly those employing the Cas12 system, demonstrate a range of advantages, including high sensitivity, specificity, and ease of use. CRISPR/Cas systems are noted for their low-cost application and the ability to function effectively in resource-limited environments, making them suitable for widespread use in diagnostics[81]. Furthermore, the CRISPR-Cas12 has the potential to precisely detect mutations, which can be tailored for various applications, enhancing its cost-effectiveness and suitability for routine clinical practices [82]. These attributes position CRISPR-based diagnostics as ideal for settings where traditional laboratory infrastructure is limited, facilitating access to essential cancer screening services. Additionally, portable detection devices leveraging CRISPR technology can significantly reduce logistical challenges associated with cancer diagnostics. Kaminski et al. note that the adaptability of CRISPR assays to point-of-care testing results in quicker diagnostic results compared to established laboratory methods [67]. By employing lateral flow assays or other portable platforms, communities in rural areas can access immediate results, enabling prompt clinical decision-making. Integrating CRISPR-based diagnostics into prostate cancer screening protocols has the potential to decentralise access to healthcare services, particularly in underserved rural populations. The affordability and ease of deploying CRISPR tests can effectively enhance the capacity for early detection of prostate cancer-specific biomarkers, including epigenetic alterations [83]. Given that prostate cancer is prevalent and often asymptomatic in its early stages, implementing readily accessible screening tools could substantially improve early diagnosis rates and patient outcomes.
Moreover, the operational efficiency of CRISPR diagnostics can facilitate extensive screening initiatives without the need for complex laboratory machinery. For instance, lateral flow assays utilising CRISPR technology have shown promise in providing rapid, user-friendly diagnostic results, which can be performed at community health centres or even at home[84]. These programs not only empower individuals to participate in their health monitoring but also serve to raise awareness and education regarding prostate cancer risks in these communities. In addition, the rapid and simplified detection methods can serve as a model for employing CRISPR technology to monitor epigenetic changes within populations at risk for prostate cancer. Epigenetic changes invariably precede genetic alterations in cancer development, and detecting these changes using CRISPR-generated biosensors could represent a novel method of screening that addresses the unique context of the population and could significantly reduce the burden on healthcare systems [85].
5. Challenges and Considerations
Technical Challenges
The deployment of CRISPR-based diagnostics in resource-limited settings is obstructed by infrastructural, technical, and ethical challenges. Many African health facilities lack reliable electricity and molecular biology infrastructure, limiting CRISPR integration [86]. Training healthcare personnel is crucial, especially in cancer and biotechnology centres [67]. Amplification-free CRISPR methods show low sensitivity, and multiplexing remains difficult in simple formats. There is also a trade-off between assay simplicity and sensitivity, often requiring complex multistep devices to achieve accurate results. There is also a trade-off between assay simplicity and sensitivity, often requiring complex multistep devices to achieve accurate results. Innovations like crRNA multiplexing, microfluidics, and smartphone-based readouts aim to improve usability [87].
The regulatory frameworks for CRISPR diagnostics are still evolving. As these tools detect genetic markers without genome editing, regulation must balance safety and accessibility [88]. This distinction may influence the regulatory processes, potentially easing some barriers while imposing rigorous standards in other areas. The WHO supports CRISPR diagnostics under its 2022–2030 STI (Sexually Transmitted Infections) strategy, emphasising ASSURED criteria including accuracy, sensitivity, user-friendliness, and affordability. The WHO also emphasises the development of cost-effective, rapid, and field-deployable diagnostics, particularly in low- and middle-income countries. CRISPR systems (e.g., Cas12, Cas13) are under active evaluation as they can be engineered to meet most or all ASSURED principles [87].
Affordability, reagent access, and trained staff remain concerns [89]. Community engagement and education are vital for building trust and promoting adoption [90]. Inclusive health literacy programs enhance acceptance and align CRISPR use with local needs [91]. In Comparison with existing technologies such as qPCR and Next Generation Sequencing (NGS), CRISPR-Cas12a presents several advantages as point-of-care diagnostics in resource-limited settings, as presented below (Table 2).
6. Future Directions
The integration of CRISPR diagnostics and epigenetic research in Sub-Saharan Africa holds immense promise but requires tailored strategies. Current data on epigenetic markers largely exclude African populations, highlighting the urgent need for epigenome-wide association studies (EWAS) and African ancestry-informative markers (AIMs) to bridge genomic disparities [98]. There is a necessity for large-scale efforts to conduct epigenome-wide association studies (EWAS) that specifically target African populations, facilitating the identification of unique epigenetic markers associated with diseases prevalent in these demographics [98]. CRISPR-based tools must also be adapted to local contexts as portable, cost-effective kits are essential where lab infrastructure is limited [99]. Multidisciplinary collaboration among researchers, policymakers, and healthcare providers is vital to translate innovation into actionable solutions [100]. Stakeholder and community engagement, including education and regulatory harmonisation, will support equitable access and trust in these technologies [91].
7. Conclusion
The synergy between CRISPR-Cas12a and epigenetic markers offers a transformative path for early prostate cancer detection in Africa. Even though challenges remain, a context-driven, collaborative approach can significantly improve diagnostic access and outcomes, advancing health equity across the continent.
Author Contributions
Conceptualization, N.K.N. and O.O.O.; methodology, N.K.N.; software, I.F.K.; validation, N.K.N., E.C.A., and I.F.K.; formal analysis, N.K.N.; investigation, N.K.N.; resources, S.N.C. and O.O.O.; data curation, N.K.N., I.F.K.; writing the original draft preparation, N.K.N.; writing, review and editing, N.K.N., O.O.O., E.C.A., I.F.K., and S.N.C.; visualization, E.C.A.; supervision, O.O.O. and S.N.C.; project administration, N.K.N.; funding acquisition, N.K.N., O.O.O. All authors have read and agreed to the published version of the manuscript.
Funding
This study is funded by Science for Africa Foundation to the Developing Excellence in Leadership, Training and Science in Africa (DELTAS Africa) programme [Afrique One-ASPIRE, Del-15-008 and Afrique One-REACH, Del-22-011] with support from Wellcome Trust and the UK Foreign, Commonwealth & Development Office and is part of the EDCPT2 programme supported by the European Union. For purposes of open access, the author has applied a CC BY public copyright license to any Author Accepted Manuscript version arising from this submission. The funders have no role in the study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Sung H, Ferlay J, Siegel RL, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: A Cancer Journal for Clinicians. 2021;71(3):209-249. [CrossRef]
- Jalloh M, Cassell A, Niang L, Rebbeck T. Global viewpoints: updates on prostate cancer in Sub-Saharan Africa. BJU International. 2024;133(1):6-13. [CrossRef]
- Gumenku L, Sekhoacha M, Abrahams B, Mashele S, Shoko A, Erukainure OL. Genetic Signatures for Distinguishing Chemo-Sensitive from Chemo-Resistant Responders in Prostate Cancer Patients. Current Issues in Molecular Biology. 2024;46(3):2263-2277. [CrossRef]
- Ferlay J, Ervik M, Lam F, Laversanne M, Colombet M, Mery L, Piñeros M, Znaor A, Soerjomataram I, Bray F. Cancer Today. 2024. Accessed April 2, 2024. https://gco.iarc.who.int/today/.
- Bello JO, Buhari T, Mohammed TO, et al. Determinants of prostate-specific antigen screening test uptake in an urban community in North-Central Nigeria. African Health Sciences. 2019;19(1):1665-1670. [CrossRef]
- Bosland MC, Shittu OB, Ikpi EE, Akinloye O. Potential New Approaches for Prostate Cancer Management in Resource-Limited Countries in Africa. Annals of Global Health. 2023;89(1). [CrossRef]
- Mbugua RG, Karanja S, Oluchina S. Effectiveness of a Community Health Worker-Led Intervention on Knowledge, Perception, and Prostate Cancer Screening among Men in Rural Kenya. Advances in Preventive Medicine. 2022;2022(1):4621446. [CrossRef]
- Rotimi SO, Rotimi OA, Salhia B. A Review of Cancer Genetics and Genomics Studies in Africa. Frontiers in Oncology. 2021;10. Accessed November 17, 2023. https://www.frontiersin.org/articles/10.3389/fonc.2020.606400.
- Dumitrescu RG. Early Epigenetic Markers for Precision Medicine. In: Dumitrescu RG, Verma M, eds. Cancer Epigenetics for Precision Medicine : Methods and Protocols. Springer; 2018:3-17. [CrossRef]
- Tsou JH, Leng Q, Jiang F. A CRISPR Test for Rapidly and Sensitively Detecting Circulating EGFR Mutations. Diagnostics. 2020;10(2):114. [CrossRef]
- Zakari S, Niels NK, Olagunju GV, et al. Emerging biomarkers for non-invasive diagnosis and treatment of cancer: a systematic review. Front Oncol. 2024;14. [CrossRef]
- Sakai N. Prostate-Specific Antigen-Based Prostate Cancer Screening. In: Prostate Cancer - Leading-Edge Diagnostic Procedures and Treatments. IntechOpen; 2016. [CrossRef]
- Bae J, Kim Y. Cancer and epigenetics. Animal Cells and Systems. 2008;12(3):117-125. [CrossRef]
- Baylin SB. DNA methylation and gene silencing in cancer. Nat Rev Clin Oncol. 2005;2(1):S4-S11. [CrossRef]
- Ahuja N, Easwaran H, Baylin SB. Harnessing the potential of epigenetic therapy to target solid tumors. J Clin Invest. 2014;124(1):56-63. [CrossRef]
- Dawson MA, Kouzarides T. Cancer Epigenetics: From Mechanism to Therapy. Cell. 2012;150(1):12-27. [CrossRef]
- Nebbioso A, Tambaro FP, Dell’Aversana C, Altucci L. Cancer epigenetics: Moving forward. PLOS Genetics. 2018;14(6):e1007362. [CrossRef]
- Toh TB, Lim JJ, Chow EKH. Epigenetics in cancer stem cells. Mol Cancer. 2017;16(1):29. [CrossRef]
- Jin B, Li Y, Robertson KD. DNA methylation: superior or subordinate in the epigenetic hierarchy? Genes Cancer. 2011;2(6):607-617. [CrossRef]
- Zhang X, Zhang Y, Wang C, Wang X. TET (Ten-eleven translocation) family proteins: structure, biological functions and applications. Signal Transduction and Targeted Therapy. 2023;8(1):297. [CrossRef]
- Puddu F, Johansson A, Modat A, et al. 5-methylcytosine and 5-hydroxymethylcytosine are synergistic biomarkers for early detection of colorectal cancer. bioRxiv. Published online 2024:2024.10.30.621123. [CrossRef]
- Shi X, Zhai Z, Chen Y, Li J, Nordenskiöld L. Recent Advances in Investigating Functional Dynamics of Chromatin. Frontiers in Genetics. 2022;13. [CrossRef]
- Liu R, Wu J, Guo H, et al. Post-translational modifications of histones: Mechanisms, biological functions, and therapeutic targets. MedComm (2020). 2023;4(3):e292. [CrossRef]
- Xu Y, He Z, Du J, et al. Epigenetic modulations of immune cells: from normal development to tumor progression. International Journal of Biological Sciences. 2023;19(16):5120.
- Das R, Thakur K, Shrivastava A, Puri A, Mutsuddi M. IDENTIFYING EPIGENETIC ENDPOINTS OF PESTICIDE EXPOSURE CAN CURTAIL RISK TO DEVELOP CANCER: A REVIEW. International Journal of Advanced Research. 2017;5:1093-1107. [CrossRef]
- López-Hernández L, Toolan-Kerr P, Bannister AJ, Millán-Zambrano G. Dynamic histone modification patterns coordinating DNA processes. Molecular Cell. 2025;85(2):225-237. [CrossRef]
- Nemeth K, Bayraktar R, Ferracin M, Calin GA. Non-coding RNAs in disease: from mechanisms to therapeutics. Nature Reviews Genetics. 2024;25(3):211-232. [CrossRef]
- Baluchamy S, Srijyothi L, Ponne S, Prathama T, Ashok C. Roles of Non-Coding RNAs in Transcriptional Regulation. In: Kais G, ed. Transcriptional and Post-Transcriptional Regulation. IntechOpen; 2018.
- Tsai MC, Manor O, Wan Y, et al. Long noncoding RNA as modular scaffold of histone modification complexes. Science. 2010;329(5992):689-693.
- Conteduca V, Hess J, Yamada Y, Ku SY, Beltran H. Epigenetics in prostate cancer: clinical implications. Transl Androl Urol. 2021;10(7):3104-3116. [CrossRef]
- Majumdar S, Buckles E, Estrada J, Koochekpour S. Aberrant DNA methylation and prostate cancer. Curr Genomics. 2011;12(7):486-505. [CrossRef]
- Nakayama M, Bennett CJ, Hicks JL, et al. Hypermethylation of the human glutathione S-transferase-π gene (GSTP1) CpG island is present in a subset of proliferative inflammatory atrophy lesions but not in normal or hyperplastic epithelium of the prostate: a detailed study using laser-capture microdissection. The American journal of pathology. 2003;163(3):923-933.
- Van Neste L, Herman JG, Otto G, Bigley JW, Epstein JI, Van Criekinge W. The Epigenetic promise for prostate cancer diagnosis. The Prostate. 2012;72(11):1248-1261. [CrossRef]
- Pidsley R, Lam D, Qu W, et al. Comprehensive methylome sequencing reveals prognostic epigenetic biomarkers for prostate cancer mortality. Clinical and Translational Medicine. 2022;12(10):e1030. [CrossRef]
- Ngollo M, Lebert A, Daures M, et al. Global analysis of H3K27me3 as an epigenetic marker in prostate cancer progression. BMC Cancer. 2017;17(1):261. [CrossRef]
- Haider R, Massa F, Kaminski L, et al. Sirtuin 7: a new marker of aggressiveness in prostate cancer. Oncotarget. 2017;8(44):77309.
- Zhen L, Gui-Lan L, Ping Y, Jin H, Ya-Li W. The expression of H3K9Ac, H3K14Ac, and H4K20TriMe in epithelial ovarian tumors and the clinical significance. International journal of gynecological cancer. 2010;20(1):82-86.
- Seputra KP, Purnomo BB, Susianti H, Kalim H, Purnomo AF. miRNA-21 as reliable serum diagnostic biomarker candidate for metastatic progressive prostate cancer: meta-analysis approach. Medical Archives. 2021;75(5):347.
- Ibrahim NH, Abdellateif MS, Kassem SHA, Abd El Salam MA, El Gammal MM. Diagnostic significance of miR-21, miR-141, miR-18a and miR-221 as novel biomarkers in prostate cancer among Egyptian patients. Andrologia. 2019;51(10):e13384.
- Nip H, Dar AA, Saini S, et al. Oncogenic microRNA-4534 regulates PTEN pathway in prostate cancer. Oncotarget. 2016;7(42):68371.
- Sidhu S, Deep J, Sobti R, Sharma V, Thakur H. Methylation pattern of MGMT gene in relation to age, smoking, drinking and dietary habits as epigenetic biomarker in prostate cancer patients. Genetic Engineering and Biotechnology Journal. Published online 01 2010.
- Keil KP, Abler LL, Mehta V, et al. DNA methylation of E-cadherin is a priming mechanism for prostate development. Dev Biol. 2014;387(2):142-153. [CrossRef]
- Deng X, Bhagat S, Dong Z, Mullins C, Chinni SR, Cher M. Tissue inhibitor of metalloproteinase-3 induces apoptosis in prostate cancer cells and confers increased sensitivity to paclitaxel. European Journal of Cancer. 2006;42(18):3267-3273. [CrossRef]
- Jerónimo C, Henrique R, Hoque MO, et al. Quantitative RARβ2 hypermethylation: a promising prostate cancer marker. Clinical Cancer Research. 2004;10(12):4010-4014.
- Rybicki BA, Rundle A, Kryvenko ON, et al. Methylation in benign prostate and risk of disease progression in men subsequently diagnosed with prostate cancer. International journal of cancer. 2016;138(12):2884-2893.
- Daniunaite K, Jarmalaite S, Kalinauskaite N, et al. Prognostic value of RASSF1 promoter methylation in prostate cancer. The Journal of urology. 2014;192(6):1849-1855.
- Abramovic I, Pezelj I, Dumbovic L, et al. LGALS3 cfDNA methylation in seminal fluid as a novel prostate cancer biomarker outperforming PSA. The Prostate. 2024;84(12):1128-1137. [CrossRef]
- Tokizane T, Shiina H, Igawa M, et al. Cytochrome P450 1B1 is overexpressed and regulated by hypomethylation in prostate cancer. Clin Cancer Res. 2005;11(16):5793-5801. [CrossRef]
- Nowacka-Zawisza M, Wiśnik E. DNA methylation and histone modifications as epigenetic regulation in prostate cancer (Review). Oncol Rep. 2017;38(5):2587-2596. [CrossRef]
- Chu M, Chang Y, Li P, Guo Y, Zhang K, Gao W. Androgen receptor is negatively correlated with the methylation-mediated transcriptional repression of miR-375 in human prostate cancer cells. Oncology reports. 2014;31(1):34-40.
- David MK. LS. Prostate-Specific Antigen.
- Tidd-Johnson A, Sebastian SA, Co EL, et al. Prostate cancer screening: Continued controversies and novel biomarker advancements. Curr Urol. 2022;16(4):197-206. [CrossRef]
- Glinge C, Clauss S, Boddum K, et al. Stability of circulating blood-based microRNAs–pre-analytic methodological considerations. PloS one. 2017;12(2):e0167969.
- Zubakov D, Boersma AW, Choi Y, van Kuijk PF, Wiemer EA, Kayser M. MicroRNA markers for forensic body fluid identification obtained from microarray screening and quantitative RT-PCR confirmation. International journal of legal medicine. 2010;124:217-226.
- Halabian R, Valizadeh A, Ahmadi A, Saeedi P, Azimzadeh Jamalkandi S, Alivand MR. Laboratory methods to decipher epigenetic signatures: a comparative review. Cellular & Molecular Biology Letters. 2021;26(1):46. [CrossRef]
- Joo JE, Wong EM, Baglietto L, et al. The use of DNA from archival dried blood spots with the Infinium HumanMethylation450 array. BMC biotechnology. 2013;13:1-6.
- Collinson P. Evidence and cost effectiveness requirements for recommending new biomarkers. Ejifcc. 2015;26(3):183.
- García-Giménez JL, Seco-Cervera M, Tollefsbol TO, et al. Epigenetic biomarkers: Current strategies and future challenges for their use in the clinical laboratory. Crit Rev Clin Lab Sci. 2017;54(7-8):529-550. [CrossRef]
- Baden J, Adams S, Astacio T, et al. Predicting prostate biopsy result in men with prostate specific antigen 2.0 to 10.0 ng/ml using an investigational prostate cancer methylation assay. The Journal of urology. 2011;186(5):2101-2106.
- Van Neste L, Bigley J, Toll A, et al. A tissue biopsy-based epigenetic multiplex PCR assay for prostate cancer detection. BMC Urology. 2012;12(1):16. [CrossRef]
- Djomkam Zune AL, Olwal CO, Tapela K, et al. Pathogen-Induced Epigenetic Modifications in Cancers: Implications for Prevention, Detection and Treatment of Cancers in Africa. Cancers. 2021;13(23):6051.
- Oladipo EK, Olufemi SE, Adediran DA, et al. Epigenetic modifications in solid tumor metastasis in people of African ancestry. Front Oncol. 2024;14. [CrossRef]
- Moen EL, Zhang X, Mu W, et al. Genome-Wide Variation of Cytosine Modifications Between European and African Populations and the Implications for Complex Traits. Genetics. 2013;194(4):987-996. [CrossRef]
- Cronjé HT, Elliott HR, Nienaber-Rousseau C, Pieters M. Replication and expansion of epigenome-wide association literature in a black South African population. Clinical Epigenetics. 2020;12(1):6. [CrossRef]
- Hobbs A, and Ramsay M. Epigenetics and the Burden of Noncommunicable Disease: A Paucity of Research in Africa. Epigenomics. 2015;7(4):627-639. [CrossRef]
- Tristan-Flores FE, de la Rocha C, Pliego-Arreaga R, Cervantes-Montelongo JA, Silva-Martínez GA. Epigenetic Changes Induced by Infectious Agents in Cancer. In: Velázquez-Márquez N, Paredes-Juárez GA, Vallejo-Ruiz V, eds. Pathogens Associated with the Development of Cancer in Humans: OMICs, Immunological, and Pathophysiological Studies. Springer Nature Switzerland; 2024:411-457. [CrossRef]
- Kaminski MM, Abudayyeh OO, Gootenberg JS, Zhang F, Collins JJ. CRISPR-based diagnostics. Nat Biomed Eng. 2021;5(7):643-656. [CrossRef]
- Pascual-Garrigos A, Lozano-Torres B, Das A, Molloy JC. Colorimetric CRISPR Biosensor: A Case Study with Salmonella Typhi. ACS Sens. Published online February 5, 2025. [CrossRef]
- Shao N, Han X, Song Y, Zhang P, Qin L. CRISPR-Cas12a Coupled with Platinum Nanoreporter for Visual Quantification of SNVs on a Volumetric Bar-Chart Chip. Anal Chem. 2019;91(19):12384-12391. [CrossRef]
- Zhou H, Tsou JH, Leng Q, Jiang F. Sensitive Detection of KRAS Mutations by Clustered Regularly Interspaced Short Palindromic Repeats. Diagnostics. 2021;11(1):125. [CrossRef]
- Costa-Pinheiro P, Montezuma ,Diana, Henrique ,Rui, and Jerónimo C. Diagnostic and Prognostic Epigenetic Biomarkers in Cancer. Epigenomics. 2015;7(6):1003-1015. [CrossRef]
- Qu H, Zhang W, Li J, et al. A rapid and sensitive CRISPR-Cas12a for the detection of Fusobacterium nucleatum. Microbiology Spectrum. 2024;12(2):e03629-23. [CrossRef]
- Parveen S, Rizvi SF, Hasan A, Afaq U, Mir SS. Novel Insights into Epigenetic Control of Autophagy in Cancer. OBM Genetics. 2022;6(4):1-45. [CrossRef]
- Tsou JH, Leng Q, Jiang F. A CRISPR Test for Detection of Circulating Nuclei Acids. Translational Oncology. 2019;12(12):1566-1573. [CrossRef]
- Van Dongen JE, Berendsen JTW, Eijkel JCT, Segerink LI. A CRISPR/Cas12a-assisted platform for identification and quantification of single CpG methylation sites. Published online April 6, 2021. [CrossRef]
- Sun Y, Wen T, Zhang P, Wang M, Xu Y. Recent Advances in the CRISPR/Cas-Based Nucleic Acid Biosensor for Food Analysis: A Review. Foods. 2024;13(20):3222. [CrossRef]
- Qiu M, Zhou XM, Liu L. Improved Strategies for CRISPR-Cas12-based Nucleic Acids Detection. J Anal Test. 2022;6(1):44-52. [CrossRef]
- Xiong Y, Zhang J, Yang Z, et al. Functional DNA Regulated CRISPR-Cas12a Sensors for Point-of-Care Diagnostics of Non-Nucleic-Acid Targets. J Am Chem Soc. 2020;142(1):207-213. [CrossRef]
- Lucia C, Federico PB, Alejandra GC. An ultrasensitive, rapid, and portable coronavirus SARS-CoV-2 sequence detection method based on CRISPR-Cas12. Published online March 2, 2020:2020.02.29.971127. [CrossRef]
- Chen W, Wu S, Li G, et al. Accurate diagnosis of prostate cancer with CRISPR-based nucleic acid test strip by simultaneously identifying PCA3 and KLK3 genes. Biosensors and Bioelectronics. 2023;220:114854. [CrossRef]
- Dong J, Feng W, Lin M, et al. Comparative Evaluation of PCR-Based, LAMP and RPA-CRISPR/Cas12a Assays for the Rapid Detection of Diaporthe aspalathi. International Journal of Molecular Sciences. 2024;25(11):5773. [CrossRef]
- Etemadzadeh A, Salehipour P, Motlagh FM, et al. An Optimized CRISPR/Cas12a Assay to Facilitate the BRAF V600E Mutation Detection. Journal of Clinical Laboratory Analysis. 2024;38(21):e25101. [CrossRef]
- Wu P, Cao Z, Wu S. New Progress of Epigenetic Biomarkers in Urological Cancer. Disease Markers. 2016;2016(1):9864047. [CrossRef]
- Liu Y, Zhan L, Qin Z, Sackrison J, Bischof JC. Ultrasensitive and Highly Specific Lateral Flow Assays for Point-of-Care Diagnosis. ACS Nano. 2021;15(3):3593-3611. [CrossRef]
- Hattori N, Ushijima T. Epigenetic impact of infection on carcinogenesis: mechanisms and applications. Genome Med. 2016;8(1):10. [CrossRef]
- Moetlhoa B, Nxele SR, Maluleke K, et al. Barriers and enablers for implementation of digital-linked diagnostics models at point-of-care in South Africa: stakeholder engagement. BMC Health Services Research. 2024;24(1):216. [CrossRef]
- Gul I, Raheem MA, Reyad-ul-Ferdous Md, et al. CRISPR diagnostics for WHO high-priority sexually transmitted infections. TrAC Trends in Analytical Chemistry. 2025;182:118054. [CrossRef]
- Lau A, Ren C, Lee LP. Critical review on where CRISPR meets molecular diagnostics. Prog Biomed Eng. 2020;3(1):012001. [CrossRef]
- Azhar Mohd, Phutela R, Kumar M, et al. Rapid and accurate nucleobase detection using FnCas9 and its application in COVID-19 diagnosis. Biosensors and Bioelectronics. 2021;183:113207. [CrossRef]
- Javalkote VS, Kancharla N, Bhadra B, et al. CRISPR-based assays for rapid detection of SARS-CoV-2. Methods. 2022;203:594-603. [CrossRef]
- Tyumentseva M, Tyumentsev A, Akimkin V. CRISPR/Cas9 Landscape: Current State and Future Perspectives. International Journal of Molecular Sciences. 2023;24(22):16077. [CrossRef]
- Su J, Ke Y, Maboyi N, et al. CRISPR/Cas12a Powered DNA Framework-Supported Electrochemical Biosensing Platform for Ultrasensitive Nucleic Acid Analysis. Small Methods. 2021;5(12). [CrossRef]
- Li C, Lin N, Feng Z, et al. CRISPR/Cas12a Based Rapid Molecular Detection of Acute Hepatopancreatic Necrosis Disease in Shrimp. Frontiers in Veterinary Science. 2022;8. [CrossRef]
- Dai X, Shen L. Advances and Trends in Omics Technology Development. Front Med (Lausanne). 2022;9:911861. [CrossRef]
- Cheng Y, Lyu J, Han J, et al. A specific and ultrasensitive Cas12a/crRNA assay with recombinase polymerase amplification and lateral flow biosensor technology for the rapid detection of Streptococcus pyogenes. Microbiology Spectrum. 2024;12(10). [CrossRef]
- Hu M, Qiu Z, Bi ZR, Tian T, Jiang Y, Zhou X. Photocontrolled crRNA activation enables robust CRISPR-Cas12a diagnostics. Proceedings of the National Academy of Sciences. 2022;119(26). [CrossRef]
- Tan M, Liang L, Liao C, et al. A rapid and ultra-sensitive dual readout platform for Klebsiella pneumoniae detection based on RPA-CRISPR/Cas12a. Frontiers in Cellular and Infection Microbiology. 2024;14. [CrossRef]
- Chilunga FP, Henneman P, Venema A, et al. Genome-wide DNA methylation analysis on C-reactive protein among Ghanaians suggests molecular links to the emerging risk of cardiovascular diseases. npj Genom Med. 2021;6(1):1-9. [CrossRef]
- Harlemon M, Ajayi O, Kachambwa P, et al. A Custom Genotyping Array Reveals Population-Level Heterogeneity for the Genetic Risks of Prostate Cancer and Other Cancers in Africa. Cancer Research. 2020;80(13):2956-2966. [CrossRef]
- Wang S, Qian F, Zheng Y, et al. Genetic variants demonstrating flip-flop phenomenon and breast cancer risk prediction among women of African ancestry. Breast Cancer Res Treat. 2018;168(3):703-712. [CrossRef]
Figure 1.
DNA methylation status in normal (A) and cancer (B) cells [14].
Figure 1.
DNA methylation status in normal (A) and cancer (B) cells [14].
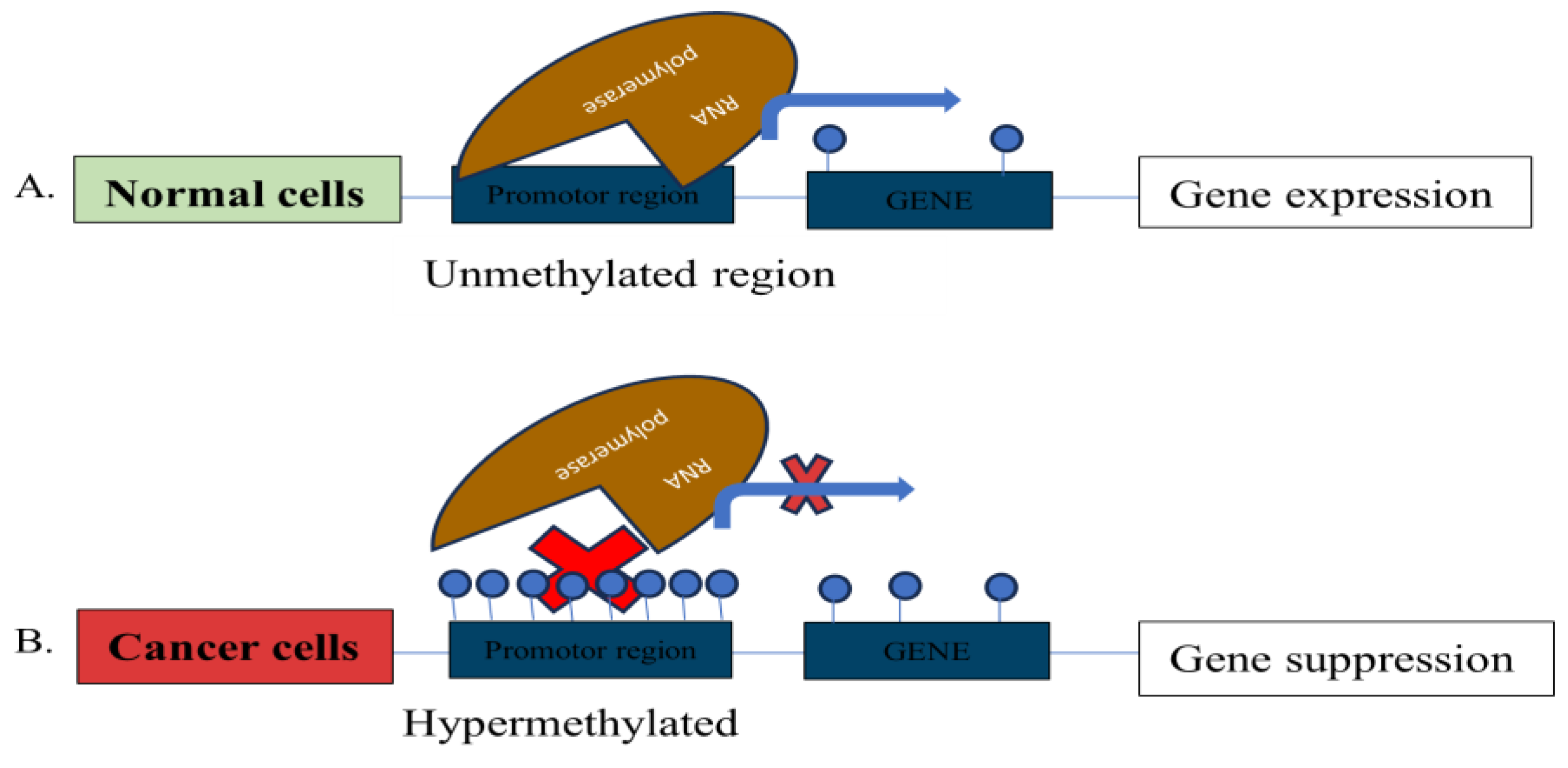
Figure 2.
DNA methylation pathway [21].
Figure 2.
DNA methylation pathway [21].
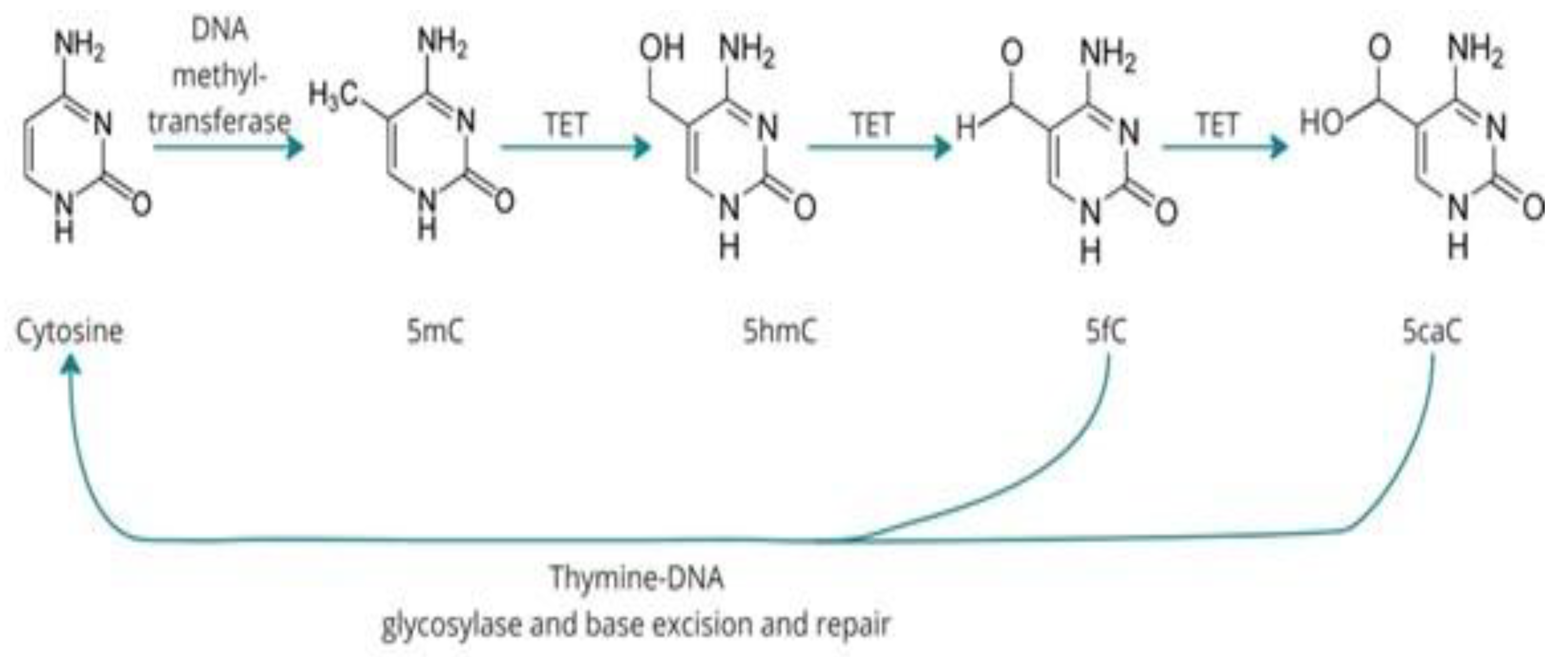
Figure 3.
Histone acetylation leads to relaxed chromatin structure. HATs: Histone acetyltransferase; Ac: acetyl groups [25].
Figure 3.
Histone acetylation leads to relaxed chromatin structure. HATs: Histone acetyltransferase; Ac: acetyl groups [25].
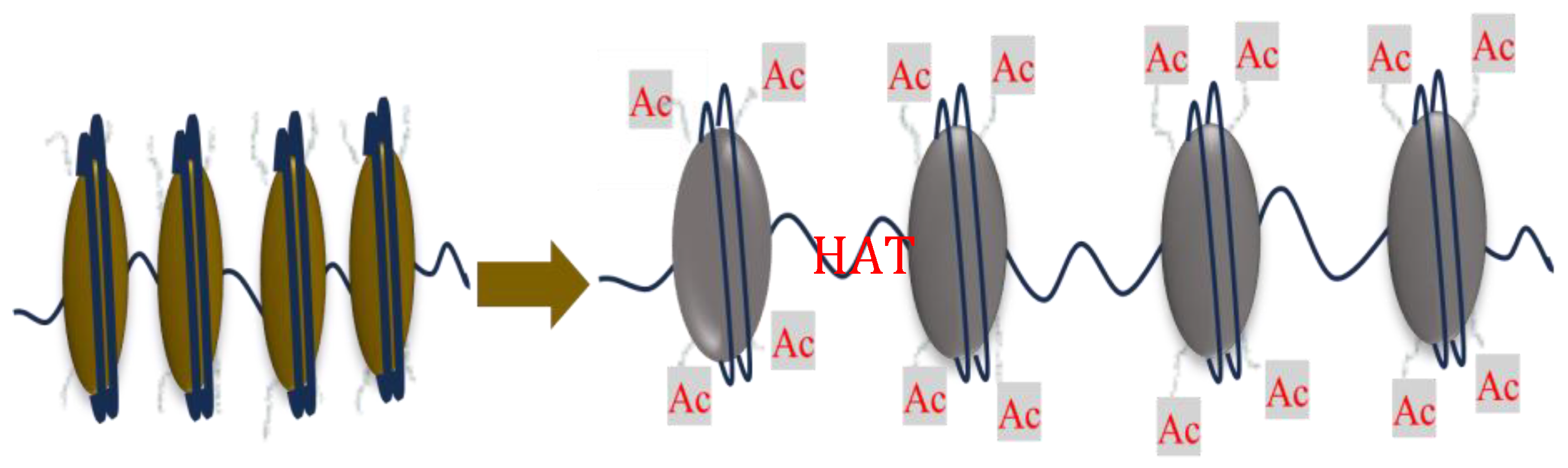
Figure 4.
CRISPR-Cas12 principle for Nucleic Acid detection [67].
Figure 4.
CRISPR-Cas12 principle for Nucleic Acid detection [67].
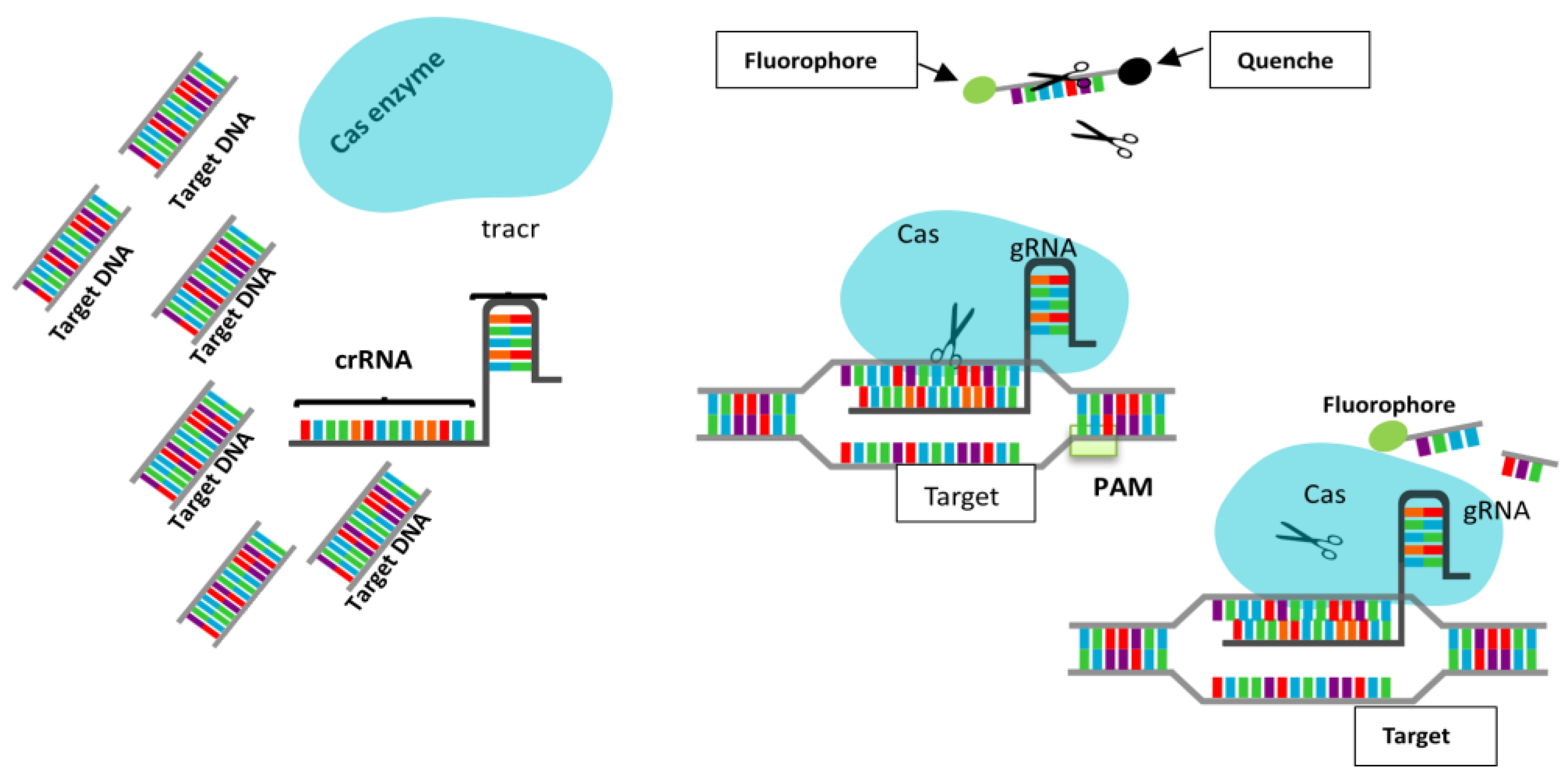
Table 2.
Comparison of CRISPR-Cas12a, qPCR, and NGS for Methylation and miRNA Detection.
| Parameter | CRISPR-Cas12a | qPCR | Next Generation Sequencing (NGS) |
|---|---|---|---|
| Sensitivity | High (as low as 100 fM); enhanced by collateral cleavage [92] | Moderate–High; dependent on primer/probe design and efficiency | Very High; resolution dependent on sequencing depth [94] |
| Cost | Low to moderate; suitable for resource-limited settings [95] | Moderate; less expensive than NGS, but not easily multiplexed [93] | Highly costly reagents, equipment, and bioinformatics [94] |
| Speed | <2 hours ideal for point-of-care [96] | ~1–2 hours real-time readout | Slow (days); long prep and data analysis [94] |
| Scalability | Moderate; being improved for high-throughput screening | Low to moderate; limited multiplexing | Very High; excellent for large sample sets [94] |
| Required Infrastructure | Minimal; no need for complex equipment [97] | Basic molecular lab setup(thermocycler) | High-end sequencing and data analysis infrastructure required [94] |
| Suitability for Resource-Limited Settings | Excellent, portable and cost-effective | Moderate; requires thermal cycler | Poor, impractical without advanced infrastructure |
| Methylation Detection | Emerging; can detect methylation via enzyme-based pre-treatment + CRISPR [75] | Possible with Me-PCR or qMSP; limited to known targets | Comprehensive; can map genome-wide methylation [94] |
| miRNA Detection | Achievable using Cas12a/crRNA targeting mature miRNA; under development | Well established with TaqMan or SYBR Green assays | High-resolution profiling of the entire miRNome |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



