Submitted:
07 July 2025
Posted:
08 July 2025
You are already at the latest version
Abstract
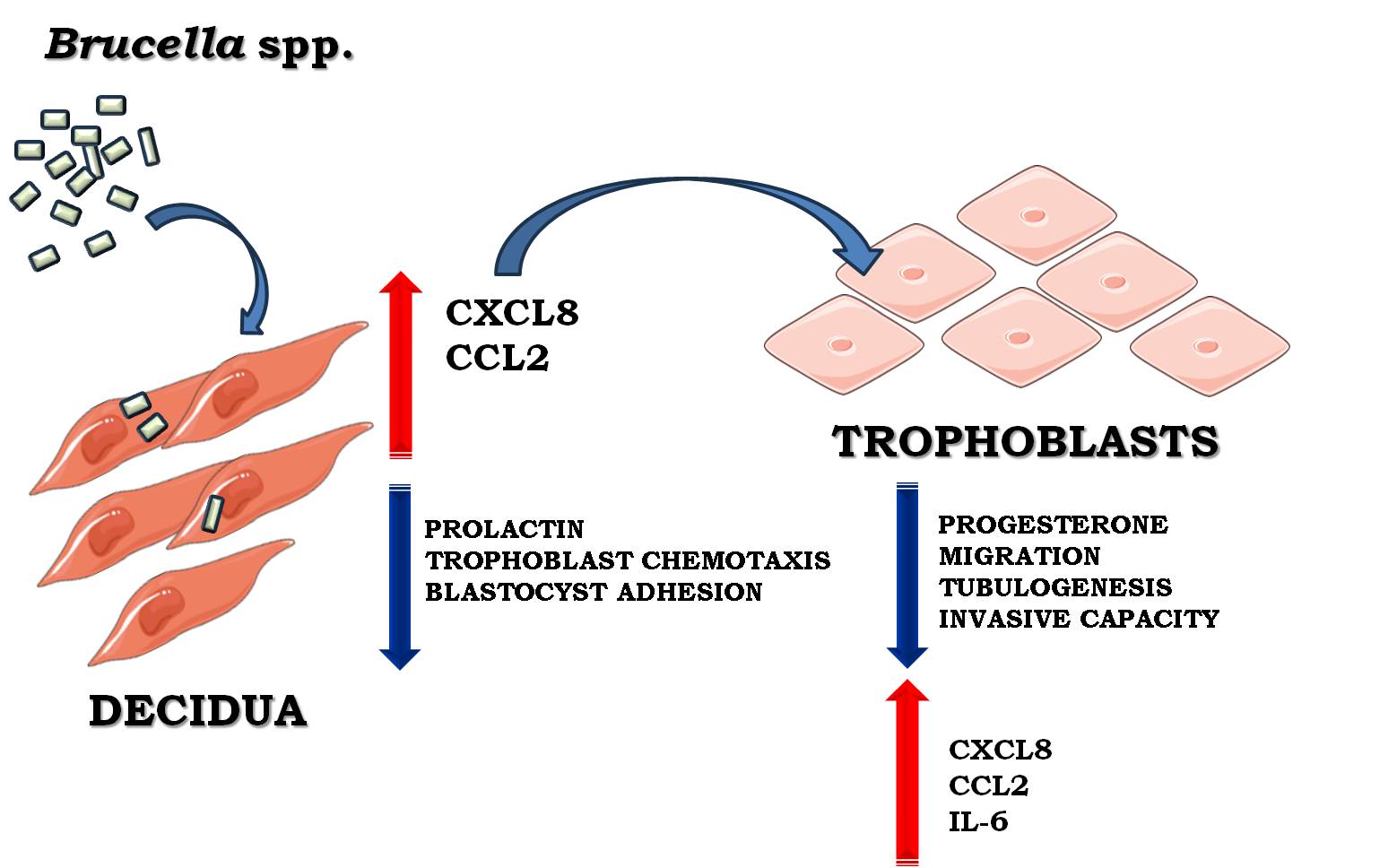
Brucella infection is associated with an increased risk of adverse obstetric outcomes in humans and animals. Decidualization, a process involving structural and functional changes in endometrial stromal cells, is essential for proper trophoblast implantation and placental development. Here we demonstrate that infection of human endometrial stromal cells (T-HESC cell line) with either B. abortus, B. suis or B. melitensis results in deficient decidualization (as revealed by reduced prolactin levels) and an increased production of proinflammatory chemokines (CXCL8 and CCL2) as compared to uninfected cells subjected to decidualization stimuli. In addition, conditioned media (CM) from infected decidualized T-HESC induced an inflammatory response (CXCL8, CCL2 and IL-6) in human trophoblasts (Swan-71 cell line) but reduced their ability to produce progesterone. Trophoblasts migration and their ability to invade the decidua and to undergo tubulogenesis, critical for proper implantation and placental development, are normally promoted by decidual cells. In contrast, we found that all these abilities were impaired when trophoblasts were preincubated with CM from Brucella-infected decidualized T-HESC cells, and this impairment was mediated, at least in part, by CXCL8 and CCL2. Moreover, infection of decidual stromal cells impaired the adhesion and spreading of blastocyst-like spheroids formed by Swan-71 cells. Brucella infection also affected the chemotactic capacity of decidual stromal cells for trophoblasts. Overall, these results suggest that Brucella infection of endometrial stromal cells impairs key processes required for successful implantation and placental development.
Keywords:
endometrial stromal cells
; decidua
; trophoblasts
; Brucella
; chemokines
; pregnancy complications
; infertility
1. Introduction
Brucellosis is a globally distributed zoonotic disease caused by Gram-negative bacteria of the Brucella genus. The disease primarily affects domestic and wild animals, which act as reservoirs, facilitating transmission to humans [1]. Among the species most pathogenic to humans are B. melitensis, B. suis, and B. abortus. In animals, brucellosis is closely linked to reproductive disorders, including abortion, premature birth, orchitis, epididymitis, and infertility. In pregnant women, numerous studies have reported an increased risk of adverse obstetric outcomes such as preterm delivery, spontaneous abortion, fetal death, and low birthweight [2,3,4,5,6,7]. Notably, infection can be transmitted to the fetus, primarily through transplacental spread [8,9]. Given the association between Brucella infection and reproductive complications, understanding its impact on key processes at the maternal-fetal interface is crucial.
Establishing a successful pregnancy relies on maternal endometrial receptivity during the implantation window. A pivotal event in this process is decidualization, which involves structural and functional changes in endometrial stromal cells, including cell enlargement, spiral artery remodeling, and immune cell infiltration by macrophages and uterine natural killer cells [10]. Endometrial stromal cells, the predominant cell type in the endometrium, undergo cyclic differentiation during the luteal phase in response to rising levels of estrogen and progesterone [11]. This differentiation transforms stromal cells from a fibroblast-like morphology into epithelioid-like decidual cells, a process that in humans occurs even in the absence of implantation and reaches full maturity at the onset of gestation. Decidualization not only supports implantation by promoting the secretion of key factors such as prolactin but also facilitates immune tolerance by attracting predominantly anti-inflammatory leukocytes, preventing excessive recruitment of cytotoxic T cells [12]. Additionally, decidual cells secrete chemokines that promote trophoblast invasion and angiogenesis, essential for early pregnancy development[13].
At the maternal-fetal interface, the decidua and the placenta form a highly specialized structure critical for pregnancy maintenance. The placenta contains three types of trophoblasts: cytotrophoblasts, which are mononuclear proliferative cells; syncytiotrophoblasts, which form a multinucleated layer covering the chorionic villi; and extravillous trophoblasts (EVTs). Among these, EVTs invade the decidua to anchor the placenta and remodel maternal spiral arteries, ensuring adequate blood flow to the fetus. Together, the decidua and trophoblasts contribute to the production of essential hormones and nutrients required for fetal growth. Proper decidualization and trophoblast invasion are, therefore, fundamental processes that, if disrupted, may lead to pregnancy complications.
Although the uterus provides a unique immune environment that supports the development of the semi-allogeneic fetus, it remains susceptible to infections. Pathogens may access the endometrium via ascending routes from the vagina or through hematogenous circulation [14], potentially altering its function and triggering inflammatory responses that can compromise pregnancy [15]. Infections have been implicated in up to 15% of early pregnancy losses [16,17]. Decidual and non-decidual stromal cells express various pathogen recognition receptors, including Toll-like receptors (TLRs) and Nod-like receptors (NLRs), which detect microbial components and initiate immune responses through the secretion of inflammatory cytokines, chemokines, and metalloproteinases [18,19]. This innate immune defense is essential for limiting infection but can also lead to tissue damage and impaired decidual function when dysregulated.
Altered decidualization, measured by reduced expression of decidual markers such as prolactin, has been linked to pregnancy complications, including miscarriage, preeclampsia, and intrauterine growth restriction[20,21,22]. Moreover, infections by bacteria such as Chlamydia trachomatis have been shown to impair decidualization and decidual function in uterine stromal cells [23]. Despite these findings, little is known about how bacterial infections, such as those caused by Brucella, affect the interaction between endometrial stromal cells and EVTs at the maternal-fetal interface.
We previously demonstrated that B. abortus infects fully decidualized and non-decidualized endometrial stromal cells, inducing a proinflammatory response [24]. Moreover, a recent study in mice showed that lipopolysaccharide from B. suis S2 disrupts decidualization and reduces implantation rates[25]. In this study, we aimed to investigate the effects of infection with highly virulent Brucella species on stromal cell decidualization, as well as its subsequent effects on trophoblast migration, adhesion, invasion and immune responses in the context of trophoblast-decidua interactions.
2. Results
2.1. Brucella spp. Infection of Human Endometrial Stromal Cells Results in Deficient Decidualization
Brucella infections are associated with infertility. Like other hematogenously disseminated pathogens, Brucella can colonize the uterus even in the absence of pregnancy [26,27,28]. Therefore, we evaluated whether infection with virulent Brucella species affects the subsequent decidualization process of uterine endometrial stromal cells.
To investigate this, endometrial stromal cells from the T-HESC line were infected with B. abortus, B. melitensis, and B. suis at different multiplicities of infection (MI: 50, 250, 500) or were left uninfected as controls. At 24 h post-infection, both infected and non-infected control cells were subjected to the classical decidualization protocol as described in Methods. Culture supernatants were collected at different times post-decidualization and prolactin (PRL) levels were quantified by ELISA. As shown in Figure 1, prior infection with Brucella significantly reduced PRL production in an MI-dependent manner at 6 days post-decidualization compared to non-infected cells. No changes in PRL secretion were observed at earlier time points (Figure S1). These findings indicate that prior Brucella spp. infection impairs the ability of endometrial stromal cells to undergo proper decidualization, which could contribute to the implantation and pregnancy maintenance disorders associated with Brucella infections.
2.2. Brucella spp. Infection of Human Endometrial Stromal Cells Induces a Proinflammatory Chemokine Response
We previously demonstrated that B. abortus can induce the production of the inflammatory chemokines CXCL8 (IL-8, neutrophil chemoattractant) and CCL2 (MCP-1, monocyte chemoattractant) in both fully decidualized and non-decidualized stromal cells, which may contribute to the inflammation-driven gestational complications of brucellosis [24]. To further investigate the impact of prior Brucella infection on the production of inflammatory factors by decidualized T-HESC cells, CXCL8, lL-6 and CCL2 levels were measured in culture supernatants. The results show that prior B. abortus, B. melitensis, and B. suis infection significantly increased the secretion of both CXCL8 and CCL2 compared to non-infected but decidualized cells (Figure 2). In contrast, infection with all tested Brucella species did not increase IL-6 production (data not shown). Taken together, these findings indicate that infection with virulent Brucella strains alters the secretome of decidualized stromal cells, affecting not only prolactin production but also enhancing the secretion of proinflammatory chemokines. This dysregulation may compromise trophoblast functionality and, consequently, impair placental development.
2.3. Factors Released by Infected Decidualized Stromal Cells Induce an Inflammatory Response in Trophoblasts
The maintenance of a regulated inflammatory microenvironment during the gestational period is crucial to ensure a successful pregnancy. Whereas decidualized stromal cells support pregnancy through their interactions with trophoblasts, in the context of infection they may contribute to an altered secretome that affects trophoblast functionality and inflammatory profile. To investigate this, we analyzed the effects of conditioned medium (CM) from B. abortus–infected and later decidualized stromal cells (CM Ba) on the immune response of trophoblasts from the Swan-71 cell line. These cells were stimulated with CM Ba or CM from non-infected stromal cells (CM NI) for 24 or 48 hours, and CXCL8, IL-6, and CCL2 levels were quantified by ELISA. Cytokine levels present in the CMs were subtracted from those detected in the supernatants of stimulated trophoblasts to account for baseline cytokine presence.
Our results indicate that exposure to CM from Brucella-infected decidual cells significantly increased the production of IL-6 and CXCL8 in trophoblasts compared to non-stimulated controls (Basal) at 48 h post-stimulation (Figures 3A and 3B). Additionally, CCL2 secretion was elevated at both 24 and 48 h post-stimulation (Figure 3C). In contrast, trophoblasts stimulated with CM from non-infected but decidualized THESCs (CM NI) did not show increased cytokine or chemokine production compared to the basal control, suggesting that the inflammatory response of trophoblasts is specifically triggered by infection-induced changes in the decidual secretome. Overall, these results highlight a potential mechanism by which infected maternal tissues influence placental immunity.
2.4. Brucella Infection Reduces the Ability of Decidualized T-HESC Cells to Stimulate Progesterone Production in Trophoblasts
Progesterone has a crucial role in pregnancy maintenance through modulation of maternal immune responses, reduction of uterine contractility, improvement of utero-placental circulation and promotion of extravillous trophoblasts invasion to the decidua [29]. Progesterone also stimulates the proliferation and differentiation of endometrial stromal cells [11]. Given the extensive cross-talk between trophoblasts and decidual cells during pregnancy [30,31] and that some factors produced by decidual cells can stimulate progesterone secretion in trophoblasts [32], we evaluated whether CM from T-HESC decidual cells may induce progesterone production in Swan-71 trophoblasts and whether this effect may be altered by Brucella infection. Trophoblasts were stimulated or not (basal) with CM from infected decidualized T-HESC cells (infection with B. abortus, B. suis or B. melitensis) or from non-infected cells. Progesterone levels were measured 48 h later in the supernatants of the stimulated cultures. As shown in Figure 4, trophoblasts stimulated with CM from non-infected T-HESC cells significantly increased their secretion of progesterone as compared to the basal control. In contrast, progesterone levels induced by stimulation with CM from decidualized T-HESC previously infected with any of the Brucella species tested were significantly lower and this reduction was proportional to the MI used for infection. These results suggest that Brucella infection may induce changes in the decidual soluble factors involved in progesterone secretion by trophoblasts.
2.5. Brucella Infection of Decidualized Stromal Cells Impairs Outgrowth of Trophoblast Spheroid in Co-Culture Models
Proper implantation requires dynamic interactions between invading trophoblast cells and receptive decidual stromal cells, which undergo extensive remodeling during early pregnancy. Decidualization promotes a supportive environment for embryo implantation [33]. Given that Brucella can infect and alter stromal cell function, we hypothesized that infection of decidual stromal cells may impair their interaction with trophoblasts, thereby disrupting blastocyst-like spheroid adhesion (BLS) and spreading (a surrogate of impaired implantation).
To assess whether Brucella infection of decidual stromal cells suppresses the outgrowth of Swan-71 spheroids, stromal cells were first infected and then decidualized. Non-infected Swan-71 BLS were subsequently placed directly on top of the decidualized stromal cell layer and incubated for 24 hours. Images were taken at 0 and 24 hours to quantify changes in spheroid area.
As shown in Figure 5, previous infection of stromal cells with any of the Brucella species tested reduced the spreading of spheroids over decidual monolayers in an MI-dependent manner, compared to the uninfected control, as indicated by a significant decrease in BLS area. These findings suggest that Brucella infection of decidual stromal cells negatively affects trophoblasts adhesion to the decidua, potentially disrupting the decidua-trophoblast interaction required for proper implantation and placental development.
2.6. Conditioned Medium from Infected Decidualized Stromal Cells Affects the Migration of Trophoblast
Trophoblast migration is a critical process for proper placental development and remodeling of the maternal-fetal interface. Disruptions in this process can contribute to pregnancy complications. Taking into account that decidual infection by Brucella reduced the spreading of BLS over decidual monolayers, and that the CMs of infected decidualized stromal cells modulated trophoblast immune responses, we next evaluated, using a wound healing assay, whether trophoblast migration, a key process for BLS spreading, may be also influenced by these CMs. To this end, a wound was created on monolayers of Swan-71 cells, which were subsequently stimulated for 18 hours with either CM Ba, CM Bs, CM Bm or CM NI. Images of the wound were taken at 0 hours post-stimulation and at the end of the assay. As shown in Figure 6, stimulation with CM Ba obtained at two different MI significantly inhibited trophoblast wound closure compared to stimulation with CM NI. No significant differences in wound closure were detected between cells stimulated with CM NI and those treated with culture medium supplemented with 10% FBS (positive control). A similar inhibition was observed for CM Bs and CM Bm as depicted in Figures S2 and S3, respectively.
To assess whether the chemokines CXCL8 and CCL2, induced by Brucella spp. infection and present in the CM Ba, CM Bs and CM Bm, might be involved in wound closure inhibition, we preincubated CM from infected cells and CM NI for 1 hour with two concentrations of neutralizing antibodies against CXCL8 and CCL2 (0.5 and 1 µg/ml) before repeating the wound healing assay. As shown in Figure 6A, neutralization of CXCL8 with the lower antibody concentration (0.5 µg/ml) fully restored the migration of CM Ba MI 50, with no significant differences compared to the untreated control (CM NI). However, in CM Ba MI 500, low-dose CXCL8 neutralization only partially restored migration, as significant differences remained compared to the untreated CM Ba MI 500. Notably, high-dose CXCL8 neutralization (1 µg/ml) completely reversed the migration inhibition induced by CM Ba MI 500, restoring it to untreated CM NI levels (Figure 6B). Interestingly, high-dose CXCL8 neutralization also partially reduced trophoblast migration in both CM NI and CM Ba MI 50, compared to untreated CM NI (Figure 6B).
Similar to the effects observed with CXCL8 neutralization, blocking CCL2 in CM Ba MI 50 at the lower tested antibody dose restored migration, achieving wound closure levels comparable to CM NI (Figure 6C). Although no significant differences were observed in wound closure between CM Ba MI 500 treated with the lower dose of neutralizing anti-CCL2, high-dose CCL2 neutralization further enhanced CM-induced migration (Figure 6D). Consistent with the findings for CXCL8, treatment with neutralizing anti-CCL2 at 1 µg/ml significantly reduced migration in both CM NI and CM Ba MI 50 compared to untreated CM NI. Furthermore, pre-treatment of both CM Ba MI 50 and MI 500 with low-dose neutralizing antibodies against CXCL8 and CCL2 restored wound closure to levels comparable to untreated CM NI (Figure 6E). In contrast, treatment with both neutralizing antibodies at the higher dose not only failed to improve wound closure in CM Ba MI 50 and MI 500 but also reduced CM NI-induced wound closure compared to untreated CM NI (Figure 6F).
Similar results were observed in the trophoblast wound closure induced by CM Bs following neutralization of CXCL8, CCL2, or both (Figure S2). Regarding B. melitensis infections, contrast to what was observed for CM from THESC infected with B. abortus and B. suis, treatment with the lower doses of neutralizing antibodies against CXCL8 or CCL2 failed to reverse the inhibition of wound closure induced by CM Bm MI 50 and MI 500 (Figure S3). Only the highest dose of individual neutralizing antibodies could counteract the inhibitory effect on wound closure induced by CM Bm MI 500. However, this dose of neutralizing antibody also inhibited migration induced by CM NI, compared to the respective untreated condition (Figure S3). Notably, pretreatment of CM Bm with both neutralizing antibodies at the lowest dose, but not at the highest dose, fully reversed the wound closure inhibition induced by CM Bm MI 50 and MI 500, compared to CM NI. In addition, high doses of both neutralizing antibodies strongly inhibited wound closure induced by CM NI.
Overall, these findings suggest that a delicate balance between CXCL8 and CCL2 levels regulates trophoblast migration, either promoting or inhibiting it. Brucella infection in the decidua disrupts this equilibrium, impairing migratory capacity. While low-dose neutralization of CXCL8 and CCL2 restores migration, high-dose blockade further reduces it, even under non-infected conditions. These results underscore the critical role of finely tuned chemokine signaling in trophoblast migration and suggest that infection-induced imbalances may contribute to placental dysfunction.
2.7. Conditioned Medium from Infected Decidualized Stromal Cells Impairs Trophoblast Invasion
Trophoblast invasion into the decidua is a critical process for proper placental development and pregnancy progression, as it ensures adequate maternal-fetal exchange and vascular remodeling. Disruptions in this process are associated with pregnancy complications. Given that the secretome of Brucella-infected decidual stromal cells induces a proinflammatory response in trophoblasts and impairs their migration, we next evaluated whether it also affects their invasive capacity.
Swan-71 cells were incubated for 1 hour with CM from infected or non-infected decidual cells. Following incubation, the cells were seeded onto a protein matrix-coated membrane placed in the upper compartment of a Transwell system. The cultures were maintained for 18 hours with continuous exposure to CM in the upper compartment, while the lower compartment was supplemented with DMEM/F12 containing 10% FBS to promote invasion. After stimulation, cells retained on the membrane were stained and quantified using microscopy-based image analysis.
As shown in Figures 7A and B, stimulation with CM from Brucella spp.-infected decidual stromal cells (MI 250) significantly reduced the number of invading trophoblasts compared to CM from non-infected cells (CM NI). Among the three Brucella species tested, CM from B. melitensis-infected decidual cells had the strongest inhibitory effect on trophoblast invasion. Moreover, pre-treatment of CM from both infected and non-infected cells with neutralizing antibodies against CXCL8 or CCL2 significantly increased trophoblast invasion compared to their respective untreated CM (Figure 7A and B). This suggests that physiological levels of CXCL8 and CCL2 are required to support proper trophoblast invasion, whereas an excessive production of these chemokines induced by Brucella infection of decidua cells disrupts this process. These findings indicate that Brucella spp. infection alters the secretome of decidual stromal cells, leading to an overproduction of CXCL8 and CCL2, which in turn impairs trophoblast invasive capacity.
2.8. Brucella Infection of Decidualized Stromal Cells Impairs Their Chemotactic Capacity for Trophoblasts
The above results demonstrated that exposure to CM from Brucella-infected decidual cells induces a proinflammatory response in trophoblasts, impairing both their migration and invasive capacity. Given the essential role of chemotactic signals in directing trophoblast movement, we next evaluated whether Brucella infection affects the chemotactic capacity of decidual stromal cells, potentially contributing to defective trophoblast invasion.
Swan-71 cells were suspended in DMEM/F12 with 2% FBS and were seeded onto a protein matrix-coated membrane placed in the upper compartment of a Transwell system. CM from infected or non-infected decidual stromal cells was placed in the lower compartment to evaluate chemotactic activity. After 6 hours of incubation, cells retained on the membrane were stained and quantified using microscopy-based image analysis. As shown in Figure 8A and B, stimulation with CM from Brucella-infected decidual stromal cells (MI 250) significantly reduced trophoblasts migration compared to CM NI. Among the three Brucella species tested, CM from B. melitensis- and B. suis-infected cells (CM Bm and CM Bs) exhibited the strongest inhibitory effect on trophoblast chemotaxis (Figure 8).
To further assess the role of specific chemokines in this process, CM were pre-treated with neutralizing antibodies against CXCL8 or CCL2. CXCL8 neutralization in CM Bm significantly increased trophoblast migration compared to the respective untreated CM. However, blocking CXCL8 in CM Ba or CM Bs did not significantly alter trophoblast migration (Figure 8A). Interestingly, CXCL8 neutralization in CM NI led to a reduction in trophoblast migration compared to its untreated control, suggesting that physiological levels of CXCL8 are necessary for optimal chemotaxis.
Similarly, CCL2 neutralization in CM Ba and CM Bm resulted in increased trophoblast migration but had no significant effect on migration when applied to CM Bs (Figure 8B). Notably, as observed with CXCL8 blockade, CCL2 neutralization also reduced trophoblast migration induced by CM NI compared to the untreated control (Figure 8B), further supporting the idea that physiological levels of these chemokines are required for proper trophoblast chemotaxis. However, an excess of either CXCL8 or CCL2, as may be induced by Brucella infection, may result in impaired trophoblasts migration. In addition, these results suggest that, particularly in the case of CM Bs, whose inhibitory effect on trophoblast chemotaxis was not reversed by CXCL8 or CCL2 neutralization, other infection-induced mediators not evaluated in this study may be involved in the modulation of this chemotactic process.
2.9. Conditioned Medium from Infected Decidualized Stromal Cells Impairs Trophoblast Tubulogenesis
After initial implantation, extravillous cytotrophoblasts invade the uterine wall and maternal spiral arteries, undergoing a specialized epithelial-to-endothelial transition known as pseudo-vasculogenesis [34], which is a form of tubulogenesis. This process is essential for placental development and successful pregnancy progression. As the secretome of Brucella-infected decidualized stromal cells was found to modify several trophoblasts functions, including migration and invasion capacity, we hypothesized that it may also disrupt trophoblast tubulogenesis by dysregulating key signaling molecules. To assess this, we evaluated the ability of trophoblasts to form tubular structures in response to stimulation with CM from infected and non-infected decidual cells.
Swan-71 cells were preincubated with CM Ba, CM Bm, CM Bs or CM NI for 1 h and were then seeded on a protein matrix (Geltrex) in Transwell filters (8 µm pore size). Cells were then incubated for a further 6 hours in the presence of the respective CM. After incubation, images were captured using a microscope to assess the number of master segments, master junctions, and meshes, which are key parameters of tubulogenesis.
As expected, stimulation of trophoblasts with CM NI increased the number of master segments and meshes compared to the unstimulated control (Basal) (Figures 9A and 9B). Similarly, stimulation with CM from decidual cells infected at the lowest MI (MI 50) of the tested Brucella species significantly increased the number of master segments and meshes compared to the basal control, although to a significantly lower extent than CM NI. In contrast, CM from decidual cells infected at the higher MIs (MI 250 and 500) did not induce this increase, suggesting an MI-dependent inhibitory effect of Brucella on tubulogenesis (Figures 9A and 9B). Stimulation with CM NI also increased the number of master junctions compared to the unstimulated control (Figure 9C). In contrast, CM from cells infected with Brucella species at all tested MIs did not increase the number of master junctions compared to the basal control. Representative images are shown in Figure 9D. These findings suggest that CM from Brucella-infected decidualized stromal cells impairs trophoblast tubulogenesis in an MI-dependent manner. While CMs from low MI-infected decidual cells still promote some tubulogenesis, higher MIs do not enhance this process and may even inhibit it. These results highlight the potential impact of Brucella-induced dysregulation of stromal signaling on placental development.
To assess whether proinflammatory chemokines induced by decidual cell infection contribute to the reduced ability of trophoblasts to form tubular structures, we repeated the tubulogenesis assays by pre-incubating the CM with neutralizing antibodies against CXCL8 and CCL2. As shown in Figure 10, treatment with anti-CXCL8 or anti-CCL2 antibodies increased the number of master segments, meshes, and junctions in cells stimulated with CM from Brucella-infected cells (MI 250) compared to the respective untreated CM. Notably, neutralizing these chemokines in CM NI also significantly reduced all tubulogenesis parameters compared to the untreated CM NI, highlighting the dual role of inflammatory signaling in trophoblast tubulogenesis.
3. Discussion
Trophoblast invasion of the maternal endometrial stroma and the remodeling of spiral arteries mediated by these cells are fundamental processes in early placental development. Disruptions in any of these steps can compromise pregnancy progression and lead to severe gestational pathologies. Decidual stromal cells play a pivotal role in modulating trophoblast function during implantation. These maternal cells create a finely tuned microenvironment that supports trophoblast migration, invasion, and differentiation by secreting cytokines, chemokines, growth factors, and extracellular matrix components. For example, factors secreted by decidualized endometrial stromal cells enhance trophoblast invasion by upregulating matrix metalloproteinases (MMPs) and integrins [35]. Moreover, decidual stromal cells actively shape the local immune landscape, contributing to immune tolerance and tissue remodeling, both essential for successful placentation.
Using the first-trimester placental trophoblast cell line Swan-71 and the telomerase-immortalized human endometrial stromal cell line T-HESC as experimental models, we demonstrated that infection with virulent Brucella species impairs endometrial decidualization and induces an exacerbated inflammatory microenvironment, ultimately disrupting trophoblast functionality.
We had previously reported that B. abortus infection of already decidualized stromal cells did not alter their decidual phenotype [24]. However, the present study shows that prior Brucella infection of stromal endometrial cells attenuates the subsequent decidualization process, reflected in reduced PRL levels, in a MI-dependent manner across all virulent Brucella species tested.
This observation is particularly relevant because animal models show that the uterus can serve as a reservoir for Brucella persistence and replication even outside pregnancy, and chronic Brucella infection has been linked to infertility [26]. Thus, previously infected stromal cells may undergo incomplete or defective decidualization, impairing crosstalk with trophoblasts and contributing to gestational complications associated with brucellosis. Interestingly, Lei et al. [25] recently demonstrated that purified LPS from B. suis S2 also disrupts decidualization, leading to implantation failure.
Importantly, we found that the secretome of infected and decidualized T-HESCs showed increased levels of CXCL8 and CCL2 but not IL-6. In turn, CM from these cells stimulated Swan-71 trophoblasts to secrete IL-6, CXCL8, and CCL2. Notably, these infection-driven changes in the decidual microenvironment correlated with reduced acquisition of invasive phenotypes by trophoblasts, suggesting that an altered cytokine milieu negatively affects trophoblast behavior.
A key step in implantation is thus the coordinated migration and invasion of trophoblast cells through the maternal tissue. Among the various maternal-derived factors modulating this process, CXCL8 and CCL2 play a critical role. Decidual stromal cells and uterine natural killer cells are prominent sources of CXCL8 within the maternal-fetal interface, and its receptors are highly expressed on extravillous trophoblasts (EVTs) [36]. Proper CXCL8 signaling is considered essential for a successful pregnancy, as diminished levels of CXCL8 or its receptors have been linked to recurrent pregnancy loss [37]. Our group [38] and others [39,40] have shown that exogenous CXCL8 stimulates MMP secretion, thereby promoting the migration and invasion of HTR-8 trophoblast cells. However, under pathological conditions, excessive CXCL8 levels may exert detrimental effects, shifting from a stimulatory to an inhibitory role on trophoblast behavior [38,39]. Consistent with these previous observations, we demonstrate here that neutralization of CXCL8 partially reverses the ability of CM from Brucella-infected and decidualized T-HESCs to inhibit trophoblast migration and invasion. Interestingly, the highest concentration of neutralizing anti-CXCL8 antibody also reduced migration induced by CM from non-infected decidualized T-HESCs, reinforcing the idea that a fine-tuned balance of CXCL8 is critical for optimal trophoblast function.
Moreover, our study shows that trophoblast migration was also restored when CCL2 in the CM from infected and decidualized T-HESCs was neutralized. CCL2 is secreted by decidual tissue during early pregnancy, where its production is induced by cytokines, estrogens, progesterone, and hCG, and is regulated in an autocrine manner via the ERK/MAPK pathway through its receptor CCR2 [41,42,43]. Variations in CCL2 levels can influence both normal pregnancy progression and pathological outcomes [43,44]. Although CCR2 mRNA has been detected in first-trimester bovine trophoblasts [45], the expression of CCR2 in human trophoblasts remains uncertain. Nevertheless, CCL2 can promote cell migration through the CCR4 receptor [46]. CCR4 expression has been specifically detected in invasive EVTs where it mediates migration [47]. Thus, the CCL2/CCR4 axis could partially mediate the migratory response seen upon treatment with CM from Brucella-infected decidual cells.
Beyond direct effects, CCL2 exerts indirect trophoblast regulation by recruiting macrophages expressing G-CSF, which promote EVT growth and invasion [48], guiding Th17 cells that produce IL-17 [45], and suppressing COX-2-associated oxidative stress, creating a favorable implantation environment. Additionally, since Swan-71 cells stimulated with CM from infected decidual cells also secreted CCL2 and CXCL8, we cannot exclude autocrine effects of these chemokines on trophoblast migration.
Although IL-6 has been shown to enhance trophoblast migration and invasion [49], we observed no significant increase in IL-6 levels following decidual infection. However, excessive IL-6 produced by Swan-71 cells in response to CM from infected dedicual cells could still contribute to the inhibition of migratory and invasive capacity.
Our data further show that previous Brucella infection of stromal endometrial cells not only modified the cytokine profile but also affected the chemotactic response of trophoblast cells, reducing their directed migration toward the CM. These findings highlight that beyond impairing intrinsic trophoblast invasive capacity, infection-driven alterations in the decidual secretome compromise its chemotactic and supportive functions towards trophoblasts, further disrupting the finely orchestrated maternal-trophoblast interactions essential for successful implantation and placentation.
Following the initial stages of implantation, extravillous cytotrophoblasts progressively invade the uterine wall and maternal spiral arteries, undergoing a specialized epithelial-to-endothelial transition known as vascular mimicry or pseudo-vasculogenesis, which is essential for proper placental vascular remodeling [34]. The remodeling process can be seen as a form of tubulogenesis, where new tubular structures (the remodeled spiral arteries) are formed. Our results demonstrate that CM from Brucella-infected decidualized T-HESC significantly impairs the tubulogenic capacity of Swan-71 trophoblast cells, whereas CM from non-infected THESC promotes tubule formation. This inhibitory effect was MI-dependent across all Brucella species tested. Therefore, Brucella infection may also hamper a proper vascular remodeling of the placenta, leading to pregnancy complications.
Although numerous studies have shown that CCL2 promotes angiogenesis in endothelial cells [50], a recent report demonstrated that thrombin can inhibit tube formation by HTR-8 trophoblast cells through the induction of CCL2 production [51]. Consistent with these findings, our study shows that CCL2 present in CM from infected decidualized T-HESC negatively and partially regulates the tube-forming capacity of Swan-71 cells. While CCL2 is essential for maintaining a successful pregnancy, elevated CCL2 levels have been associated with pathological conditions such as preeclampsia [43]. Therefore, we speculate that the negative impact of CM from the infected decidua on Swan-71 tubulogenesis and spiral artery remodeling may critically depend on the local concentration of CCL2.
Another key pro-angiogenic factor involved in trophoblast function is CXCL8. Our results reveal that CXCL8 present in the CM of infected decidualized T-HESC plays a central role in regulating tubulogenesis, as neutralization of CXCL8 in this CM partially restores tubule formation to levels comparable to those observed with CM from non-infected controls. These findings suggest that, although physiological levels of CXCL8 promote trophoblast migration, invasion, and tubulogenesis, consistent with its well-established pro-angiogenic effects in both endothelial and trophoblast cells [52,53], excessive or deregulated CXCL8 under pathological conditions may disrupt these processes, contributing to defective spiral artery remodeling and compromised placental development.
Overall, our study highlights the dual and context-dependent roles of CXCL8 and CCL2 signaling in trophoblast biology, underscoring the importance of maintaining a finely tuned balance of pro-angiogenic factors at the maternal-fetal interface to ensure successful placentation.
Previous studies have established that the decidua plays a pivotal role in supporting implantation by facilitating trophoblast outgrowth. Decidualized human endometrial stromal cells are notably more effective in this role compared to their non-decidualized counterparts, largely due to their capacity to secrete matrix metalloproteinases (MMPs), migrate, and encase the blastocyst, thereby regulating the extent of its invasion [33,54]. To investigate how Brucella infection affects endometrial receptivity, we developed an in vitro implantation model by laying blastocyst-like spheroids derived from Swan-71 trophoblasts on either Brucella-infected or uninfected decidualized stromal cells. Our results revealed that Brucella infection significantly impaired trophoblast spreading over the stromal monolayer, with B. melitensis exerting the most pronounced inhibitory effect. These results suggest that Brucella infection of the decidua may also lead to implantation failures.
Progesterone has several key roles in pregnancy maintenance, including modulation of maternal immune responses, reduction of uterine contractility, improvement of utero-placental circulation, promotion of extravillous trophoblasts invasion to the decidua, and stimulation of proliferation and differentiation of endometrial stromal cells [11,29]. After placental development, progesterone is produced by trophoblasts under the control of several factors including estrogen, hCG, and corticotropin-releasing hormone (CRH). GlycodelinA, a glycoprotein secreted by decidual cells, has been shown to stimulate progesterone and hCG production in different trophoblast populations[32,55]. Here we found that CM from decidualized T-HESC cells stimulates progesterone production in trophoblasts, but this stimulatory effect is significantly reduced when T-HESC are infected with Brucella prior to decidualization. While the mechanisms involved in this reduced progesterone production remain to be established, these results suggest that Brucella infection may negatively impact the ability of decidual cells to stimulate progesterone secretion by trophoblasts thus reducing the gestation-promoting effects of this hormone.
4. Materials and Methods
4.1. Brucella spp. Growth Conditions
B. abortus 2308, B. suis 1330 and B. melitensis 16M (wild type strains, obtained from our collection) were grown in tryptic soy broth at 37 °C with agitation. Bacterial cells were washed twice with sterile phosphate buffer saline (PBS), and the respective inocula were adjusted to the desired concentration based on optical density (OD) readings. Aliquots of each inoculum were plated on tryptic soy agar (TSA) and incubated at 37 °C to determine the actual number of colony-forming units (CFU) per mL. All live Brucella manipulations were performed in biosafety level 3 facilities.
4.2. Cell Lines
The human endometrial stromal cell line used in this study (T-HESC, kindly provided by Dr. Andrea Randi, University of Buenos Aires) was derived from normal stromal cells through immortalization by telomerase (hTERT) transfection, and conserved the characteristics of the regular endometrial cells [56]. T-HESC were maintained in DMEM-F12 (without phenol red) supplemented with 10% fetal bovine serum, 100 U/ml penicillin, 50 µg/ml streptomycin, 2 mM glutamine, 50 mM sodium pyruvate and 500 ng/mL puromycin. For infection assays, cells were cultured for 24 h in antibiotic-free culture medium.
The human trophoblastic cell line Swan-71, obtained from normal first trimester trophoblasts through immortalization by hTERT transfection, was kindly provided by Dr. Gil Mor (Wayne State University). These cells conserve most phenotypical characteristics of primary trophoblasts as well as their biological responses[57]. Swan-71 cells were cultured in DMEM/F-12 supplemented with 10% fetal bovine serum (FBS), 100 U/ml penicillin, 50 µg/ml streptomycin, 2 mM glutamine, 50 mM sodium pyruvate.
4.3. Cellular Infections
T-HESC cells were dispensed at 5x104 cells/well in 24-well plates, and were cultured for 24 h in antibiotic-free medium at 37 °C in a 5% CO2 atmosphere. Cells were infected with B. abortus, B. melitensis or B. suis at a multiplicity of infection (MI) of 50, 250 or 500 for 2 h. After dispensing the bacterial suspension, the plates were centrifuged (10 min at 400 xg) and then incubated for 2 h at 37 °C in a 5% CO2 atmosphere. Non-internalized bacteria were eliminated by several washes with medium alone followed by incubation in medium supplemented with 100 µg/ml gentamicin and 50 µg/ml streptomycin. After that, cells were washed and then incubated with culture medium without antibiotics.
4.4. Decidualization of T-HESC Endometrial Cells
T-HESC were infected or not with B. abortus, B. suis or B. melitensis at different MIs (50, 250, 500) as described above, and 24 h later cells were subjected to decidualization treatment as described previously [58], with minor modifications. Briefly, T-HESC (5x104 cells/well) were cultured in DMEM/F12 2% FBS with medroxyprogesterone acetate (MPA, 1 µM) and dibutyryl cAMP (0.5 mM). The culture medium was replaced every 48 hours to maintain hormonal stimulation. Cells were maintained at 37°C in a humidified atmosphere with 5% CO2 throughout the experiment. Decidualization status was evaluated at 1, 2, 4, and 6 days after the initiation of treatment by measuring prolactin levels in the culture supernatants using a sandwich ELISA kit (R&D Systems), following the manufacturer’s instructions.
4.5. Preparation of Conditioned Media from Infected and Decidualized T-HESC
Conditioned media (CM) were obtained from T-HESC that had undergone sequentially the infection and decidualization procedures described above. After 8 days of decidualization treatment, culture supernatants were collected and subsequently filtered through 0.22 µm pore-size filters to ensure sterility. To obtain control CM, mock-infected T-HESC were subjected to the same decidualization protocol to simulate the infection and decidualization conditions without bacterial exposure. The levels of CXCL8 (interleukin-8, IL-8) and CCL2 (monocyte chemoattractant protein-1, MCP-1) were measured in these CM using commercial ELISA kits (R&D Systems).
4.6. Cytokine Response of Trophoblasts Stimulated with T-HESC Conditioned Media
To evaluate the impact of a Brucella-infected decidua on the cytokine response of adjacent trophoblasts, Swan-71 cells (5x104 cells/well) were stimulated with CM from infected and non-infected decidualized stromal cells. The CM was applied at 1:2 and 1:5 dilution. After 24 and 48 h of stimulation, culture supernatants were collected to measure interleukin-6 (IL-6), CXCL8, and CCL2 levels using commercial ELISA kits (R&D Systems). To determine the cytokine production specifically attributable to the stimulated trophoblasts, baseline cytokine levels present in the corresponding CM were subtracted from the measured levels in the stimulated cultures.
4.7. Progesterone Response of Trophoblasts Stimulated with T-HESC Conditioned Media
Swan-71 trophoblasts (5x104 cells/well) were stimulated for 48 h with CM from infected and decidualized T-HESC (B. abortus, B. suis and B. melitensis infections) or from non-infected decidualized cells. Culture supernatants of the stimulated trophoblasts were harvested to determine progesterone levels using a commercial ELISA kit (Cayman Chemical Company, USA) following the instructions of the manufacturer.
4.8. Functional Response of Trophoblasts Stimulated with T-HESC Conditioned Media
a) Migration. The effect of CM from infected and non-infected decidualized stromal cells on the migration ability of trophoblasts was evaluated using the scratch test essentially as described by Rattila et al. [59]. Swan-71 cells were plated at 5x104 cells/well in culture medium without antibiotics to allow the formation of a confluent layer. The following day, the culture medium was carefully removed and cells were gently washed with DMEM-F12. A vertical scratch was performed through the monolayer using a pipette tip, and cells were cultured in the presence of CM from infected and non-infected decidualized stromal cells. Swan-71 cells cultured in DMEM/F-12 supplemented with 10% FBS were used as a positive control of migration. Pictures were taken at time 0 and later on the same microscopic field at 18 h post-stimulation. Each wound area in duplicate wells was measured with ImageJ software. The degree of wound closure was calculated as: [(area time 0 h - area time = 18h) / area time = 0 h] x 100. The independent experiments were repeated three times.
b) Invasion. Trophoblast invasive capacity was evaluated as described by Rattila et al. [60]. A Geltrex matrix (0.6 mg/mL in DMEM-F12) was applied to Transwell inserts (8 µm pore size) in 24-well plates to simulate the basal membrane. Swan-71 cells (3×104) were pre- treated for 1 hour with CM (1:2 dilution) from non-infected or Brucella-infected decidualized stromal cells (B. abortus, B. suis, B. melitensis, MI 250), then seeded in the upper chamber on top of the Geltrex matrix. CM was maintained throughout the assay in the upper chamber.
In the lower chamber of the insert 500 µl of DMEM-F12 with 10% FCS were dispensed. After 18 h of culture the medium was removed and the matrix was cleaned with a cotton swab. Transwells were very carefully washed with PBS and were fixed with PFA 4% for 15 min. After a further wash, the transwells were stained with crystal violet for 15 min. The excess of colorant was removed with distilled water and the membrane was carefully cleaned with a swab on the upper side, without touching the lower chamber to avoid the removal of cells that crossed the matrix and the membrane. The membranes of inserts were analyzed using an EVOS microscope (Thermo Fisher) to determine the number of cells that crossed the membranes. The positive control wells included 10% FBS as the attractive stimulus to check the functionality of cells.
c) Tubulogenesis. This assay was performed essentially as described by Rattila et al.[59]. A Geltrex matrix (9 mg/ml in DMEM-F12) was dispensed in a 96-well plate. When gelification was complete, Swan-71 cells (2x104/well) previously stimulated for 1 h with CM from infected (B. abortus, B. suis or B. melitensis, MI 50, 250 or 500) or non-infected stromal decidualized cells were dispensed. After 6 h the wells were imaged using an EVOS microscope (Thermo Fisher) to evaluate the number of master junctions, master segments and meshes using using ImageJ software and the Angiogenesis Analyser plugin. CM were kept during the whole assay.
4.9. Evaluation of the Chemotactic Effect of Conditioned Media from Decidualized Stromal Cells on Trophoblasts
To assess the chemotactic effect of CM from decidualized stromal cells, either infected with Brucella or not, a Transwell-based migration assay was performed. This assay aims to determine whether soluble factors present in the CM can promote the directed migration of trophoblast cells. A similar protocol to the invasion assay described above was used, with key modifications to specifically evaluate chemotaxis. CM from infected (MI 250, three Brucella species) or non-infected decidualized stromal cells was placed in the lower chamber of the transwell inserts, while Swan-71 cells were seeded in the upper chamber on a Geltrex matrix without prior CM treatment. The assay was conducted for 6 hours, with 2% FBS present in both chambers to control for serum-driven migration. The membranes of inserts were analyzed using an EVOS microscope (Thermo Fisher) to determine the number of cells retained in the membranes.
4.10. Neutralization of CXCL8 and CCL2 in Conditioned Media
To test the role of CXCL8 and CCL2 in the effects produced by CM from infected or non-infected decidualized stromal cells, in some experiments these CM were preincubated for 1 h with neutralizing antibodies against these chemokine (R&D Systems) in two different concentrations (0.5 µg/ml and 1 µg/ml). The neutralizing antibodies in CM were maintained for the duration required for each functional assay in which they were used. In some experiments CM were preincubated with mixtures of both antibodies.
4.11. Spheroids Adhesion to the Infected Decidua
In order to form blastocyst-like spheroids[61], Swan-71 cells were stained with VybrantTM DiD (Thermo-Fisher Scientific) and were dispensed in a low-binding Petri dish at 2x104 cells per 20 ul drop of medium (DMEM/F-12 supplemented with 2% FCS). Cells were incubated at 37 °C with agitation for 48 h. At this time cells were observed under the optical microscope to check whether the spheroids had formed correctly.
T-HESC cells were plated at 1x104 cells/well in 96-well plates, were infected (or not) with the tested Brucella species (MI 50, 250, 500) and were later decidualized as described above. Trophoblasts spheroids were carefully laid on the decidualized cells using a pipette with a blunt end tip to avoid damaging the spheroids. Images were taken at time 0 (T0) and 24 h later (T24) using an EVOS microscope (Thermo Fisher), and were analyzed using ImageJ software. Spheroids areas were measured at both time points and results were expressed as relative spheroid outgrowth calculated as: area T24/area T0.
4.12. Statistical Analysis
Each experiment was performed in duplicate on three separate occasions. The data are presented as the mean ± standard deviation (SD). Statistical analyses were performed using one-way ANOVA, followed by either Tukey’s post hoc test or Dunnett’s post hoc test, with GraphPad Prism 8.0 software.
5. Conclusions
Our findings demonstrate that infection of the stromal cells with virulent Brucella species exerts profound negative effects on the cellular or molecular dialogue in the maternal-fetal interface, impairing key processes required for successful implantation and placental development. By disrupting endometrial stromal cell decidualization, altering the local cytokine and chemokine milieu, and inhibiting trophoblast migration, invasion, and tubulogenesis, Brucella infection compromises the coordinated cellular interactions necessary for proper trophoblast function and spiral artery remodeling, as shown in this in vitro model of maternal-fetal interaction.
Importantly, our data reveal that while maternal-derived factors such as CXCL8 and CCL2 play essential roles in promoting trophoblast differentiation and vascular remodeling under physiological conditions, their excessive or dysregulated production in the context of infection paradoxically inhibits these processes, potentially contributing to defective placentation.
Overall, this study highlights the decidua as a critical target of Brucella-induced pathology and positions infection-driven defects in maternal stromal cells as key contributors to gestational complications. By elucidating the molecular and cellular mechanisms underlying these interactions, our work provides new insights into how Brucella infections compromise fertility and pregnancy outcomes, while also opening avenues for the development of targeted interventions to restore decidual-trophoblast crosstalk and improve reproductive health in infected individuals.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Brucella pre-infection does not impact on prolactin production by human endometrial stromal cells at early time points after decidualization stimuli.; Figure S2: Conditioned medium from Brucella suis-infected decidual cells impairs trophoblast migration.; Figure S3: Conditioned medium from Brucella melitensis-infected decidual cells impairs trophoblast migration.
Author Contributions
Conceptualization, L.Z., M.F. and P.B; Methodology, L.Z, R.M., M.P.; D.P.; M.F., A.C. and P.B; Formal Analysis, L.Z., M.F; Investigation, L.Z., M.F., M.P.; D.P.; R.M.; Writing – Original Draft Preparation, L.Z., M.F. and P.B.; Writing – Review & Editing, P.B., M.F. and A.C.; Supervision, M.F. and P.B.; Project Administration, M.F. and P.B.; Funding Acquisition, P.B.
Funding
This research was funded by grants PICT-2021-I-A-00680 from Agencia Nacional de Promoción Científica, and UBACYT 20020220100012BA from Universidad de Buenos Aires.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
We thank the staff of the INBIRS institute (UBA-CONICET) for expert technical assistance with BSL3 facilities.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pappas, G.; Papadimitriou, P.; Akritidis, N.; Christou, L.; Tsianos, E. V The New Global Map of Human Brucellosis. Lancet Infect. Dis. 2006, 6, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Bosilkovski, M.; Arapović, J.; Keramat, F. Human Brucellosis in Pregnancy – an Overview. Bosn. J. Basic Med. Sci. 2020, 20, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Elshamy, M.; Ahmed, A.I. The Effects of Maternal Brucellosis on Pregnancy Outcome. J. Infect. Dev. Ctries. 2008, 2, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Kurdoglu, M.; Adali, E.; Kurdoglu, Z.; Karahocagil, M.K.; Kolusari, A.; Yildizhan, R.; Kucukaydin, Z.; Sahin, H.G.; Kamaci, M.; Akdeniz, H. Brucellosis in Pregnancy: A 6-Year Clinical Analysis. Arch. Gynecol. Obstet. 2010, 281, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Makala, R.; Majigo, M. V.; Bwire, G.M.; Kibwana, U.; Mirambo, M.M.; Joachim, A. Seroprevalence of Brucella Infection and Associated Factors among Pregnant Women Receiving Antenatal Care around Human, Wildlife and Livestock Interface in Ngorongoro Ecosystem, Northern Tanzania. A Cross-Sectional Study. BMC Infect. Dis. 2020, 20. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Akhter, S.; Neubauer, H.; Scherag, A.; Kesselmeier, M.; Melzer, F.; Khan, I.; El-Adawy, H.; Azam, A.; Qadeer, S.; et al. Brucellosis in Pregnant Women from Pakistan: An Observational Study. BMC Infect. Dis. 2016, 16, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kledmanee, K.; Liabsuetrakul, T.; Sretrirutchai, S. Seropositivities against Brucellosis, Coxiellosis, and Toxoplasmosis and Associated Factors in Pregnant Women with Adverse Pregnancy Outcomes: A Cross-Sectional Study. PLoS One 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Giannacopoulos, I.; Eliopoulou, M.I.; Ziambaras, T.; Papanastasiou, D.A. Transplacentally Transmitted Congenital Brucellosis Due to Brucella Abortus [2]. J. Infect. 2002, 45, 209–210. [Google Scholar] [CrossRef] [PubMed]
- Alsaif, M.; Dabelah, K.; Girim, H.; Featherstone, R.; Robinson, J.L. Congenital Brucellosis: A Systematic Review of the Literature. Vector-Borne Zoonotic Dis. 2018, 18, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Benirschke, P.; Kaufmann, K. Pathology of the Human Placenta; Third.; Springer Verlag, 1995.
- Gellersen, B.; Brosens, J.J. Cyclic Decidualization of the Human Endometrium in Reproductive Health and Failure. Endocr. Rev. 2014, 35, 851–905. [Google Scholar] [CrossRef] [PubMed]
- Nancy, P.; Tagliani, E.; Tay, C.S.; Asp, P.; Levy, D.E.; Erlebacher, A. Chemokine Gene Silencing in Decidual Stromal Cells Limits T Cell Access to the Maternal-Fetal Interface. Science 2012, 336, 1317–1321. [Google Scholar] [CrossRef] [PubMed]
- Du, M.R.; Wang, S.C.; Li, D.J. The Integrative Roles of Chemokines at the Maternal-Fetal Interface in Early Pregnancy. Cell. Mol. Immunol. 2014, 11, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.R.; Bakardjiev, A.I. Pathogens and the Placental Fortress. Curr. Opin. Microbiol. 2012, 15, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Cappelletti, M.; Presicce, P.; Feiyang, M.; Senthamaraikannan, P.; Miller, L.A.; Pellegrini, M.; Sim, M.S.; Jobe, A.H.; Divanovic, S.; Way, S.S.; et al. The Induction of Preterm Labor in Rhesus Macaques Is Determined by the Strength of Immune Response to Intrauterine Infection. PLoS Biol. 2021, 19. [Google Scholar] [CrossRef] [PubMed]
- Giakoumelou, S.; Wheelhouse, N.; Cuschieri, K.; Entrican, G.; Howie, S.E.M.; Horne, A.W. The Role of Infection in Miscarriage. 2016, 22, 116–133. [CrossRef]
- Benedetto, C.; Tibaldi, C.; Marozio, L.; Marini, S.; Masuelli, G.; Pelissetto, S.; Sozzani, P.; Latino, M.A. Cervicovaginal Infections during Pregnancy: Epidemiological and Microbiological Aspects. J. Matern. Fetal. Neonatal Med. 2004, 16 Suppl 2, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Aflatoonian, R.; Tuckerman, E.; Elliott, S.L.; Bruce, C.; Aflatoonian, A.; Li, T.C.; Fazeli, A. Menstrual Cycle-Dependent Changes of Toll-like Receptors in Endometrium. Hum. Reprod. 2007, 22, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Hirata, T.; Osuga, Y.; Hamasaki, K.; Hirota, Y.; Nose, E.; Morimoto, C.; Harada, M.; Takemura, Y.; Koga, K.; Yoshino, O.; et al. Expression of Toll-like Receptors 2, 3, 4, and 9 Genes in the Human Endometrium during the Menstrual Cycle. J. Reprod. Immunol. 2007, 74, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Salker, M.; Teklenburg, G.; Molokhia, M.; Lavery, S.; Trew, G.; Aojanepong, T.; Mardon, H.J.; Lokugamage, A.U.; Rai, R.; Landles, C.; et al. Natural Selection of Human Embryos: Impaired Decidualization of Endometrium Disables Embryo-Maternal Interactions and Causes Recurrent Pregnancy Loss. PLoS One 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Tessier, C.; Prigent-Tessier, A.; Li, F.; Buzzio, O.L.; Callegari, E.A.; Horseman, N.D.; Gibori, G. Decidual Prolactin Silences the Expression of Genes Detrimental to Pregnancy. Endocrinology 2007, 148, 2326–2334. [Google Scholar] [CrossRef] [PubMed]
- Durairaj, R.R.P.; Aberkane, A.; Polanski, L.; Maruyama, Y.; Baumgarten, M.; Lucas, E.S.; Quenby, S.; Chan, J.K.Y.; Raine-Fenning, N.; Brosens, J.J.; et al. Deregulation of the Endometrial Stromal Cell Secretome Precedes Embryo Implantation Failure. Mol. Hum. Reprod. 2017, 23, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Giakoumelou, S.; Wheelhouse, N.; Brown, J.; Wade, J.; Simitsidellis, I.; Gibson, D.; Saunders, P.T.K.; Horner, P.; Entrican, G.; Howie, S.E.M.; et al. Chlamydia Trachomatis Infection of Human Endometrial Stromal Cells Induces Defective Decidualisation and Chemokine Release. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Zavattieri, L.; Ferrero, M.C.; Alonso Paiva, I.M.; Sotelo, A.D.; Canellada, A.M.; Baldi, P.C. Brucella Abortus Proliferates in Decidualized and Non-Decidualized Human Endometrial Cells Inducing a Proinflammatory Response. Pathogens 2020, 9, 369. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Wang, X.; Zhang, J.; Yin, J.; Xu, Q.; Wang, T.; Jin, Y.; Wang, A. Lipopolysaccharides of Brucella Suis S2 Impaired the Process of Decidualization in Early Pregnancy in Mice. Toxins (Basel). 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Adetunji, S.A.; Faustman, D.L.; Adams, L.G.; Garcia-Gonzalez, D.G.; Hensel, M.E.; Khalaf, O.H.; Arenas-Gamboa, A.M. Brucella Abortus and Pregnancy in Mice: Impact of Chronic Infection on Fertility and the Role of Regulatory T Cells in Tissue Colonization. Infect. Immun. 2020, 88. [Google Scholar] [CrossRef] [PubMed]
- Foster, G.; Jahans, K.L.; Reid, R.J.; Ross, H.M. Isolation of Brucella Species from Cetaceans, Seals and an Otter. Vet. Rec. 1996, 138, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Smither, S.J.; Perkins, S.D.; Davies, C.; Stagg, A.J.; Nelson, M.; Atkins, H.S. Development and Characterization of Mouse Models of Infection with Aerosolized Brucella Melitensis and Brucella Suis. Clin. Vaccine Immunol. 2009, 16, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, G.C.; Giardina, I.; Clerici, G.; Brillo, E.; Gerli, S. Progesterone in Normal and Pathological Pregnancy. Horm. Mol. Biol. Clin. Investig. 2016, 27, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Oreshkova, T.; Dimitrov, R.; Mourdjeva, M. A Cross-Talk of Decidual Stromal Cells, Trophoblast, and Immune Cells: A Prerequisite for the Success of Pregnancy; 2012; Vol. 68, pp. 366–373.
- Xu, L.; Li, Y.; Sang, Y.; Li, D.J.; Du, M. Crosstalk Between Trophoblasts and Decidual Immune Cells: The Cornerstone of Maternal-Fetal Immunotolerance. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.L.; Lam, K.K.W.; Koistinen, H.; Seppala, M.; Kurpisz, M.; Fernandez, N.; Pang, R.T.K.; Yeung, W.S.B.; Chiu, P.C.N. Glycodelin-A as a Paracrine Regulator in Early Pregnancy. J. Reprod. Immunol. 2011, 90, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Gellersen, B.; Reimann, K.; Samalecos, A.; Aupers, S.; Bamberger, A.M. Invasiveness of Human Endometrial Stromal Cells Is Promoted by Decidualization and by Trophoblast-Derived Signals. Hum. Reprod. 2010, 25, 862–873. [Google Scholar] [CrossRef] [PubMed]
- McMaster, M.T.; Zhou, Y.; Fisher, S.J. Abnormal Placentation and the Syndrome of Preeclampsia. Semin. Nephrol. 2004, 24, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Godbole, G.; Modi, D. Decidual Control of Trophoblast Invasion. Am. J. Reprod. Immunol. 2016, 75, 341–350. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.G.; Lash, G.E.; Murray-Dunning, C.; Bulmer, J.N.; Innes, B.A.; Searle, R.F.; Sass, N.; Robson, S.C. Role of Interleukin 8 in Uterine Natural Killer Cell Regulation of Extravillous Trophoblast Cell Invasion. Placenta 2010, 31, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Pitman, H.; Innes, B.A.; Robson, S.C.; Bulmer, J.N.; Lash, G.E. Altered Expression of Interleukin-6, Interleukin-8 and Their Receptors in Decidua of Women with Sporadic Miscarriage. Hum. Reprod. 2013, 28, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Macchi, R.; Sotelo, A.D.; Parrado, A.C.; Salaverry, L.S.; Blanco, G.A.; Castro, M.S.; Rey-Roldán, E.B.; Canellada, A.M. Losartan Impairs HTR-8/SVneo Trophoblast Migration through Inhibition of Angiotensin II-Induced pro-Inflammatory Profile in Human Endometrial Stromal Cells. Toxicol. Appl. Pharmacol. 2023, 461. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, M.; Stefanoska, I.; Radojčić, L.; Vićovac, L. Interleukin-8 (CXCL8) Stimulates Trophoblast Cell Migration and Invasion by Increasing Levels of Matrix Metalloproteinase (MMP)2 and MMP9 and Integrins A5 and Β1. Reproduction 2010, 139, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.J.; Han, J.W.; Kim, R.H.; Yun, S.; Kim, T.H.; Hur, S.E.; Kim, C.J.; Lee, S.K. Activation of NOD-1/JNK/IL-8 Signal Axis in Decidual Stromal Cells Facilitates Trophoblast Invasion. Am. J. Reprod. Immunol. 2017, 78. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Y.; Du, M.R.; Guo, P.F.; He, X.J.; Zhou, W.H.; Zhu, X.Y.; Li, D.J. Regulation of C-C Motif Chemokine Ligand 2 and Its Receptor in Human Decidual Stromal Cells by Pregnancy-Associated Hormones in Early Gestation. Hum. Reprod. 2007, 22, 2733–2742. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.X.; Jin, L.P.; Xu, B.; Liang, S.S.; Li, D.J. Decidual Stromal Cells Recruit Th17 Cells into Decidua to Promote Proliferation and Invasion of Human Trophoblast Cells by Secreting IL-17. Cell. Mol. Immunol. 2014, 11, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Shi, J.L.; Chen, M.; Zheng, Z.M.; Li, M.Q.; Shao, J. CCL2: An Important Cytokine in Normal and Pathological Pregnancies: A Review. Front. Immunol. 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Weng, Y.; Liu, W.; Chen, L.; Iqbal, F.; Yin, Z.; He, Y.; Wang, Y. TLRs Induce Th1/Th2 Responses by Affecting the Secretion of CCL2 at the Maternal-Foetal Interface. Int. Immunopharmacol. 2021, 100. [Google Scholar] [CrossRef] [PubMed]
- Sakumoto, R.; Hayashi, K.G.; Fujii, S.; Kanahara, H.; Hosoe, M.; Furusawa, T.; Kizaki, K. Possible Roles of CC- and CXC-Chemokines in Regulating Bovine Endometrial Function during Early Pregnancy. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Li, W.; Hu, J.; Li, Y.; Deng, M.; Zhang, S.; Ren, X.; Wu, T.; Xia, J.; Cheng, B.; et al. Targeting CCL2-CCR4 Axis Suppress Cell Migration of Head and Neck Squamous Cell Carcinoma. Cell Death Dis. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, C.M.; Hou, L.; Zhang, H.; Zhang, W.Y. CCL17 Induces Trophoblast Migration and Invasion by Regulating Matrix Metalloproteinase and Integrin Expression in Human First-Trimester Placenta. Reprod. Sci. 2014, 193371911351917. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Yang, C.; Zhang, Y.; Wang, J.; Zhang, S.; Guo, D.; Yin, T.; Yang, J. M2 Macrophage-Derived G-CSF Promotes Trophoblasts EMT, Invasion and Migration via Activating PI3K/Akt/Erk1/2 Pathway to Mediate Normal Pregnancy. J. Cell. Mol. Med. 2021, 25, 2136–2147. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, M.; Vićovac, L. Interleukin-6 Stimulates Cell Migration, Invasion and Integrin Expression in HTR-8/SVneo Cell Line. Placenta 2009, 30, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, R.; Ponce, M.L.; Young, H.A.; Wasserman, K.; Ward, J.M.; Kleinman, H.K.; Oppenheim, J.J.; Murphy, W.J. Human Endothelial Cells Express CCR2 and Respond to MCP-1: Direct Role of MCP-1 in Angiogenesis and Tumor Progression. Blood 2000, 96, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Brünnert, D.; Kumar, V.; Kaushik, V.; Ehrhardt, J.; Chahar, K.R.; Sharma, P.K.; Zygmunt, M.; Goyal, P. Thrombin Impairs the Angiogenic Activity of Extravillous Trophoblast Cells via Monocyte Chemotactic Protein-1 (MCP-1): A Possible Link with Preeclampsia. Reprod. Biol. 2021, 21. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Dubey, S.; Varney, M.L.; Dave, B.J.; Singh, R.K. IL-8 Directly Enhanced Endothelial Cell Survival, Proliferation, and Matrix Metalloproteinases Production and Regulated Angiogenesis. J. Immunol. 2003, 170, 3369–3376. [Google Scholar] [CrossRef] [PubMed]
- Das, M.K.; Basak, S.; Ahmed, M.S.; Attramadal, H.; Duttaroy, A.K. Connective Tissue Growth Factor Induces Tube Formation and IL-8 Production in First Trimester Human Placental Trophoblast Cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 181, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Grewal, S.; Carver, J.G.; Ridley, A.J.; Mardon, H.J. Implantation of the Human Embryo Requires Rac1-Dependent Endometrial Stromal Cell Migration. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 16189–16194. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, U.; Richter, D.U.; Möbius, B.M.; Briese, V.; Mylonas, I.; Friese, K. Stimulation of Progesterone, Estradiol and Cortisol in Trophoblast Tumor BeWo Cells by Glycodelin A N-Glycans. Anticancer Res. 2007, 27, 2101–2108. [Google Scholar] [PubMed]
- Krikun, G.; Mor, G.; Alvero, A.; Guller, S.; Schatz, F.; Sapi, E.; Rahman, M.; Caze, R.; Qumsiyeh, M.; Lockwood, C.J. A Novel Immortalized Human Endometrial Stromal Cell Line with Normal Progestational Response. Endocrinology 2004, 145, 2291–2296. [Google Scholar] [CrossRef] [PubMed]
- Straszewski-Chavez, S.L.; Abrahams, V.M.; Alvero, A.B.; Aldo, P.B.; Ma, Y.; Guller, S.; Romero, R.; Mor, G. The Isolation and Characterization of a Novel Telomerase Immortalized First Trimester Trophoblast Cell Line, Swan 71. Placenta 2009, 30, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Grasso, E.; Gori, S.; Paparini, D.; Soczewski, E.; Fernández, L.; Gallino, L.; Salamone, G.; Martinez, G.; Irigoyen, M.; Ruhlmann, C.; et al. VIP Induces the Decidualization Program and Conditions the Immunoregulation of the Implantation Process. Mol. Cell. Endocrinol. 2018, 460, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Rattila, S.; Kleefeldt, F.; Ballesteros, A.; Beltrame, J.S.; Ribeiro, M.L.; Ergün, S.; Dveksler, G. Pro-Angiogenic Effects of Pregnancy-Specific Glycoproteins in Endothelial and Extravillous Trophoblast Cells. Reproduction 2020, 160, 737–750. [Google Scholar] [CrossRef] [PubMed]
- Rattila, S.; Dunk, C.E.; Im, M.; Grichenko, O.; Zhou, Y.; Cohen, M.; Yanez-Mo, M.; Blois, S.M.; Yamada, K.M.; Erez, O.; et al. Interaction of Pregnancy-Specific Glycoprotein 1 with Integrin A5β1 Is a Modulator of Extravillous Trophoblast Functions. Cells 2019, 8, 1369. [Google Scholar] [CrossRef] [PubMed]
- Holmberg, J.C.; Haddad, S.; Wünsche, V.; Yang, Y.; Aldo, P.B.; Gnainsky, Y.; Granot, I.; Dekel, N.; Mor, G. An in Vitro Model for the Study of Human Implantation. Am. J. Reprod. Immunol. 2012, 67, 169–178. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Brucella pre-infection impairs the decidualization of human endometrial stromal cells. Endometrial stromal cells from the T-HESC line were infected with B. abortus (Ba), B. melitensis (Bm), or B. suis (Bs) at different multiplicities of infection (MI) or were left uninfected (NI) as controls. At 24 h post-infection, both infected or non-infected cells were subjected to the decidualization protocol. Culture supernatants were collected at day 6 post-decidualization, and prolactin (PRL) levels were quantified by ELISA. Results are expressed as mean ± SD of three independent experiments performed in duplicate. *** p<0.0001 versus NI.
Figure 1.
Brucella pre-infection impairs the decidualization of human endometrial stromal cells. Endometrial stromal cells from the T-HESC line were infected with B. abortus (Ba), B. melitensis (Bm), or B. suis (Bs) at different multiplicities of infection (MI) or were left uninfected (NI) as controls. At 24 h post-infection, both infected or non-infected cells were subjected to the decidualization protocol. Culture supernatants were collected at day 6 post-decidualization, and prolactin (PRL) levels were quantified by ELISA. Results are expressed as mean ± SD of three independent experiments performed in duplicate. *** p<0.0001 versus NI.
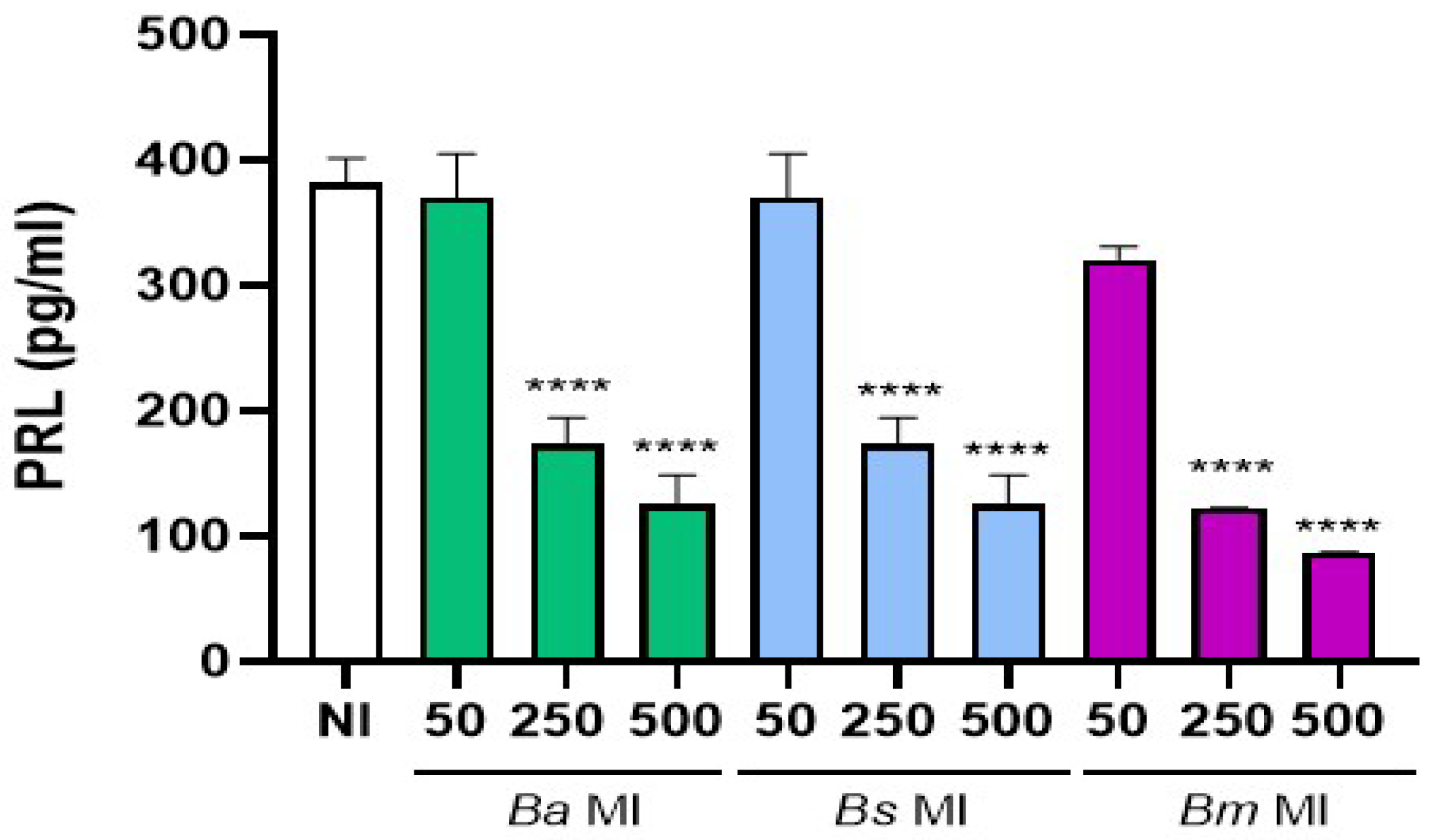
Figure 2.
Brucella preinfection induces a proinflammatory chemokine response in decidualized human endometrial cells. T-HESC cells were infected with the different Brucella species (MI 250) or were left uninfected (NI) as controls. At 24 h post-infection, both infected and non-infected cells were subjected to the decidualization protocol for 8 days. Cells were incubated for a further 24 or 48 h, and at culture supernatants were harvested for measuring CXCL8 and CCL2 using commercial ELISA kits. Results are expressed as mean ± SD of three independent experiments performed in duplicate. ns: non-significant; ***p<0.001 versus NI at the corresponding time point.
Figure 2.
Brucella preinfection induces a proinflammatory chemokine response in decidualized human endometrial cells. T-HESC cells were infected with the different Brucella species (MI 250) or were left uninfected (NI) as controls. At 24 h post-infection, both infected and non-infected cells were subjected to the decidualization protocol for 8 days. Cells were incubated for a further 24 or 48 h, and at culture supernatants were harvested for measuring CXCL8 and CCL2 using commercial ELISA kits. Results are expressed as mean ± SD of three independent experiments performed in duplicate. ns: non-significant; ***p<0.001 versus NI at the corresponding time point.
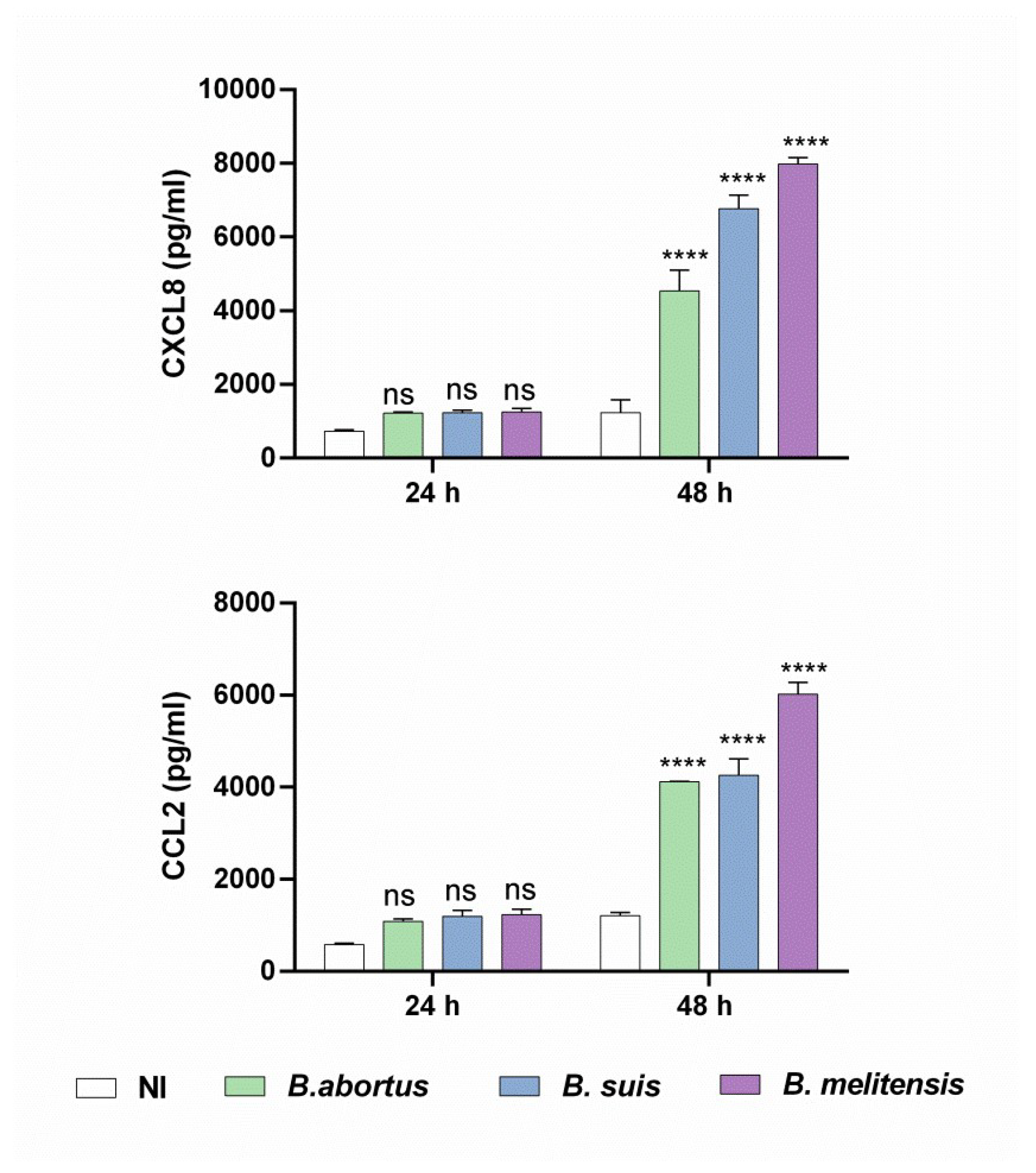
Figure 3.
Factors secreted by Brucella-infected decidualized stromal cells stimulate a proinflammatory response in trophoblasts. Trophoblasts from the Swan-71 cell line were stimulated for 24 or 48 h with conditioned medium from B. abortus–infected and later decidualized THESC cells (CM Ba) or from non-infected but decidualized cells (CM NI). After stimulation, CXCL8, IL-6, and CCL2 levels in the culture supernatants were quantified by ELISA. Cytokine levels present in the CMs were subtracted from those detected in the supernatants of stimulated trophoblast to account for baseline cytokine presence. Results are expressed as mean ± SD of three independent experiments performed in duplicate. NS: non-stimulated control. ** p<0.01; *** p<0.001; ****p<0.0001 versus NS.
Figure 3.
Factors secreted by Brucella-infected decidualized stromal cells stimulate a proinflammatory response in trophoblasts. Trophoblasts from the Swan-71 cell line were stimulated for 24 or 48 h with conditioned medium from B. abortus–infected and later decidualized THESC cells (CM Ba) or from non-infected but decidualized cells (CM NI). After stimulation, CXCL8, IL-6, and CCL2 levels in the culture supernatants were quantified by ELISA. Cytokine levels present in the CMs were subtracted from those detected in the supernatants of stimulated trophoblast to account for baseline cytokine presence. Results are expressed as mean ± SD of three independent experiments performed in duplicate. NS: non-stimulated control. ** p<0.01; *** p<0.001; ****p<0.0001 versus NS.
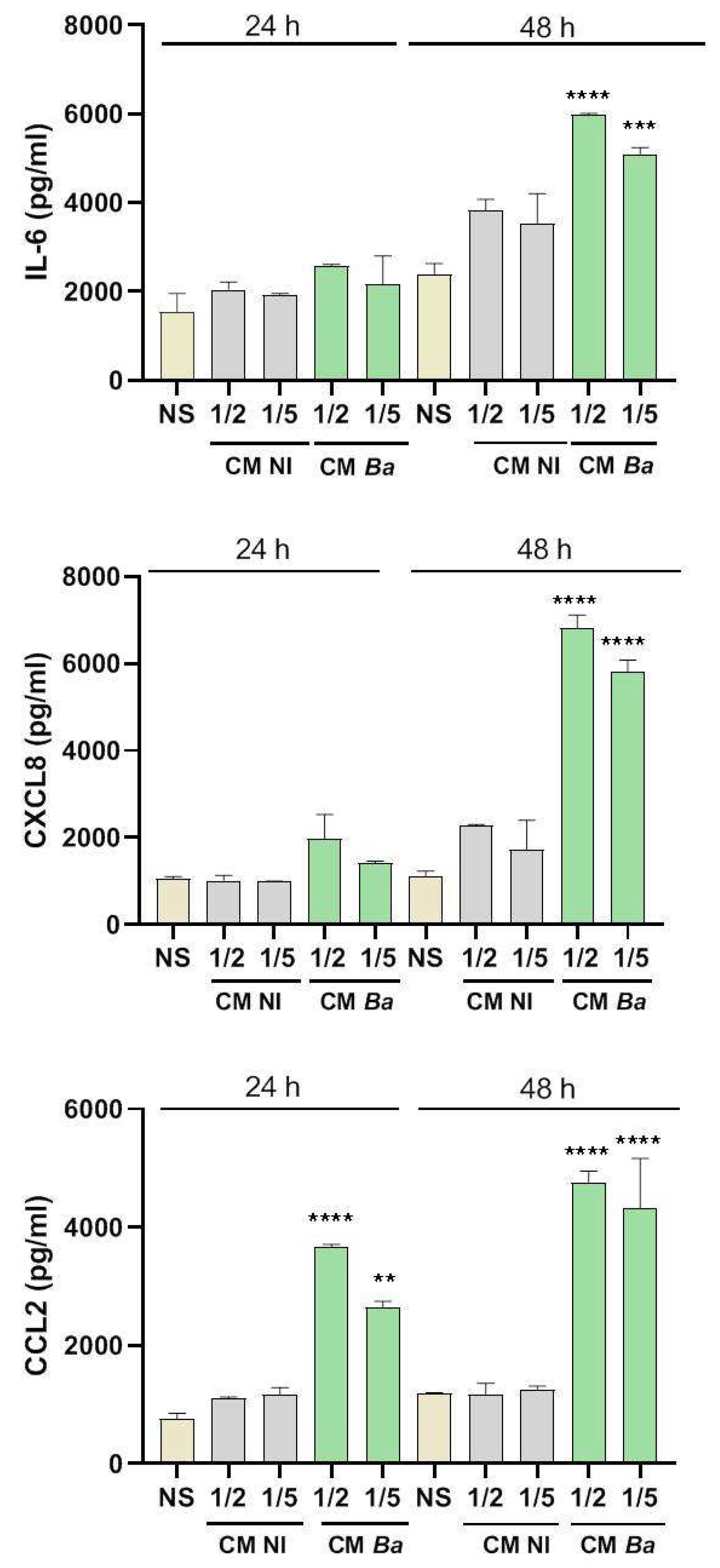
Figure 4.
Brucella infection reduces the ability of decidualized T-HESC cells to stimulate progesterone production in trophoblasts. Swan-71 trophoblasts were stimulated with CM from B. abortus-, B suis-, or B. melitensis–infected and later decidualized THESC cells (CM Ba, CM Bs and CM Bm, respectively) or from non-infected but decidualized cells (CM NI), or were kept non-stimulated (NS). At 48 h post-stimulation culture supernatants were harvested to measure progesterone with a commercial ELISA. The results are expressed as mean ± SD of three independent experiments performed in duplicate. **** p<0.0001; *** p= 0.0004; ** p= 0.002 versus CM NI.
Figure 4.
Brucella infection reduces the ability of decidualized T-HESC cells to stimulate progesterone production in trophoblasts. Swan-71 trophoblasts were stimulated with CM from B. abortus-, B suis-, or B. melitensis–infected and later decidualized THESC cells (CM Ba, CM Bs and CM Bm, respectively) or from non-infected but decidualized cells (CM NI), or were kept non-stimulated (NS). At 48 h post-stimulation culture supernatants were harvested to measure progesterone with a commercial ELISA. The results are expressed as mean ± SD of three independent experiments performed in duplicate. **** p<0.0001; *** p= 0.0004; ** p= 0.002 versus CM NI.
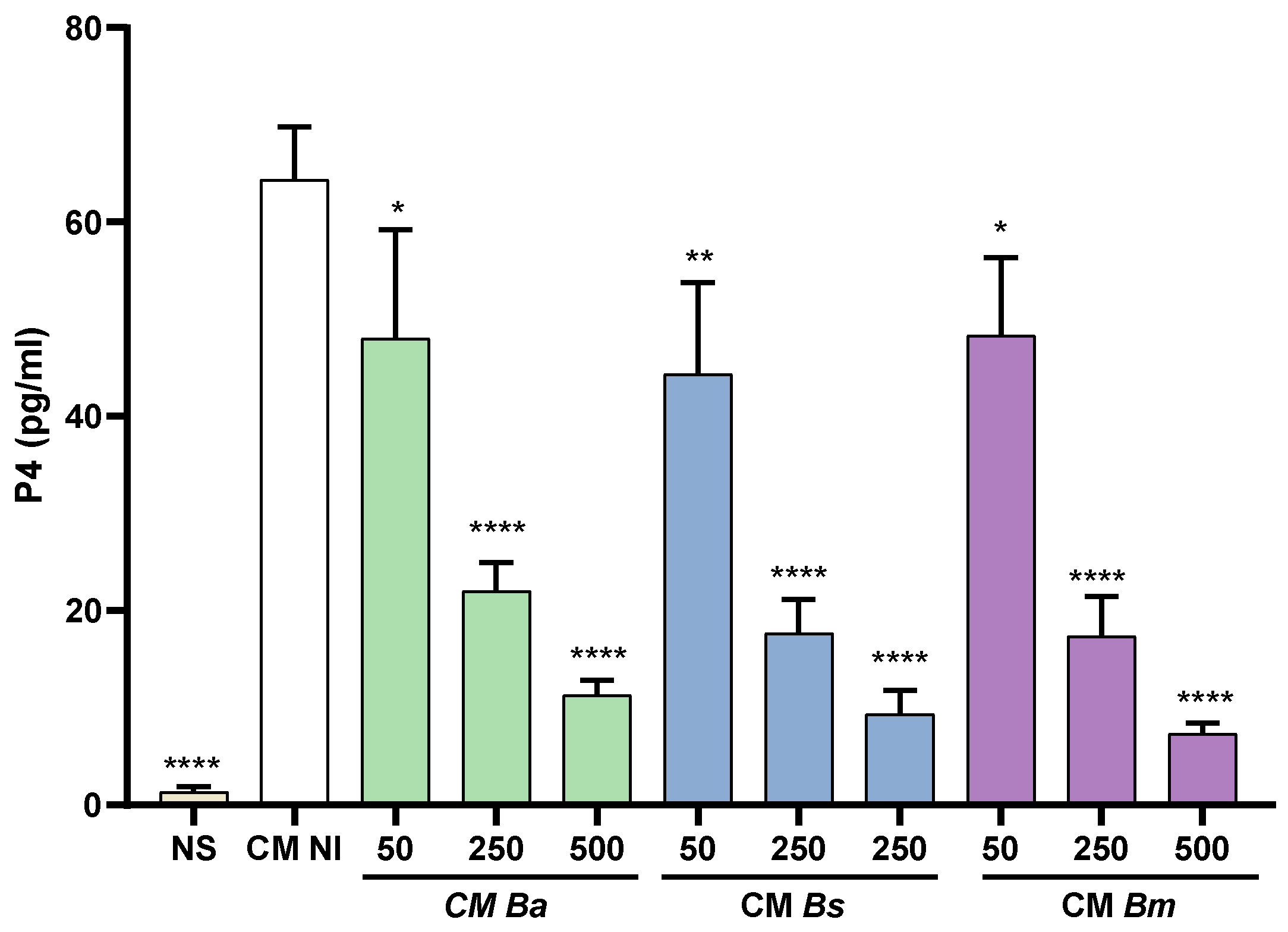
Figure 5.
Brucella infection of decidual stromal cells impairs the adhesion of trophoblast spheroids. THESC cells were infected with Brucella species at different multiplicities of infection (MI) or were left uninfected (NI), and were then decidualized. Blastocyst-like spheroids formed by non-infected trophoblasts were then laid on decidualized cells. Pictures were taken at 0 y a las 24 h to measure the area of spheroid outgrowth. Results are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over bars indicate differences versus the NI condition (** p<0.01; *** p<0.001; **** p<0.0001).
Figure 5.
Brucella infection of decidual stromal cells impairs the adhesion of trophoblast spheroids. THESC cells were infected with Brucella species at different multiplicities of infection (MI) or were left uninfected (NI), and were then decidualized. Blastocyst-like spheroids formed by non-infected trophoblasts were then laid on decidualized cells. Pictures were taken at 0 y a las 24 h to measure the area of spheroid outgrowth. Results are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over bars indicate differences versus the NI condition (** p<0.01; *** p<0.001; **** p<0.0001).
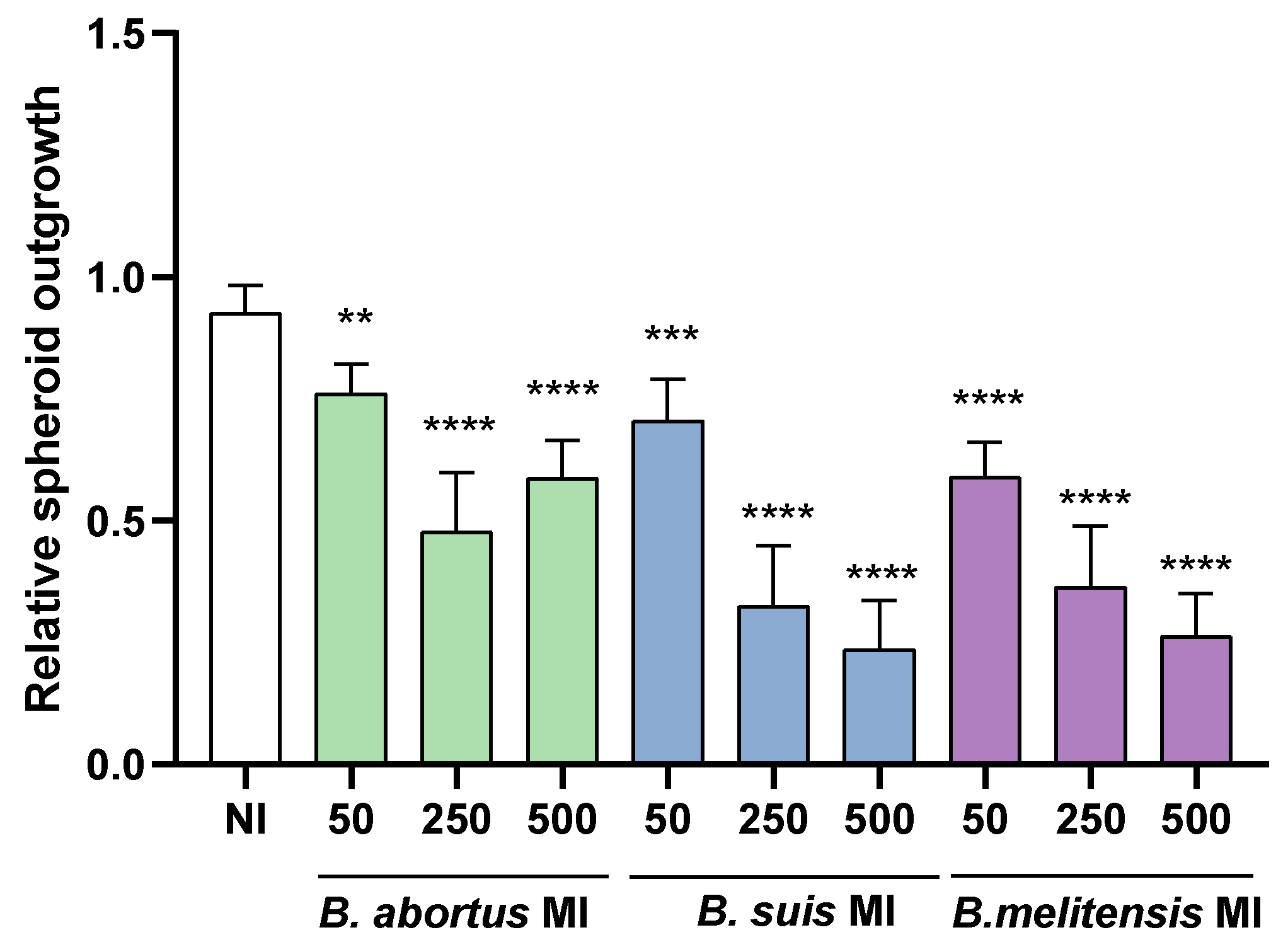
Figure 6.
Conditioned medium from Brucella abortus-infected decidual cells impairs trophoblast migration. Swan-71 trophoblasts were dispensed at 5 x 104 cells/well and were grown until confluence. A scratch was performed in the culture with a pipette tip, and then cells were stimulated with conditioned medium from B. abortus–infected and later decidualized THESC cells (CM Ba) or from non-infected but decidualized cells (CM NI). Stimulation with the corresponding CM was maintained during the whole assay. Swan-71 cells cultured in DMEM/F-12 supplemented with 10% FBS were used as a positive control of migration (+C). To evaluate wound closure, pictures were taken at time 0 and at 18 h post-stimulation on the same microscopic field. Pictures were processed using ImageJ software. The percentage of wound healing was calculated as: [(area time 0 h - area time = 18h) / area time = 0 h] x 100. In parallel experiments, CM Ba and CM NI were preincubated (or not) for 1 hour with two concentrations (0.5 and 1 µg/ml) of neutralizing antibodies against CXCL8 (A and B) or CCL2 (C and D) or a mixture of both (E and F) before performing the wound healing assay. Results are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over bars indicate differences versus the +C condition, whereas asterisks over lines indicate differences between antibody-treated and untreated conditions (*p<0.05; ** p<0.01; *** p<0.001; **** p<0.0001).
Figure 6.
Conditioned medium from Brucella abortus-infected decidual cells impairs trophoblast migration. Swan-71 trophoblasts were dispensed at 5 x 104 cells/well and were grown until confluence. A scratch was performed in the culture with a pipette tip, and then cells were stimulated with conditioned medium from B. abortus–infected and later decidualized THESC cells (CM Ba) or from non-infected but decidualized cells (CM NI). Stimulation with the corresponding CM was maintained during the whole assay. Swan-71 cells cultured in DMEM/F-12 supplemented with 10% FBS were used as a positive control of migration (+C). To evaluate wound closure, pictures were taken at time 0 and at 18 h post-stimulation on the same microscopic field. Pictures were processed using ImageJ software. The percentage of wound healing was calculated as: [(area time 0 h - area time = 18h) / area time = 0 h] x 100. In parallel experiments, CM Ba and CM NI were preincubated (or not) for 1 hour with two concentrations (0.5 and 1 µg/ml) of neutralizing antibodies against CXCL8 (A and B) or CCL2 (C and D) or a mixture of both (E and F) before performing the wound healing assay. Results are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over bars indicate differences versus the +C condition, whereas asterisks over lines indicate differences between antibody-treated and untreated conditions (*p<0.05; ** p<0.01; *** p<0.001; **** p<0.0001).
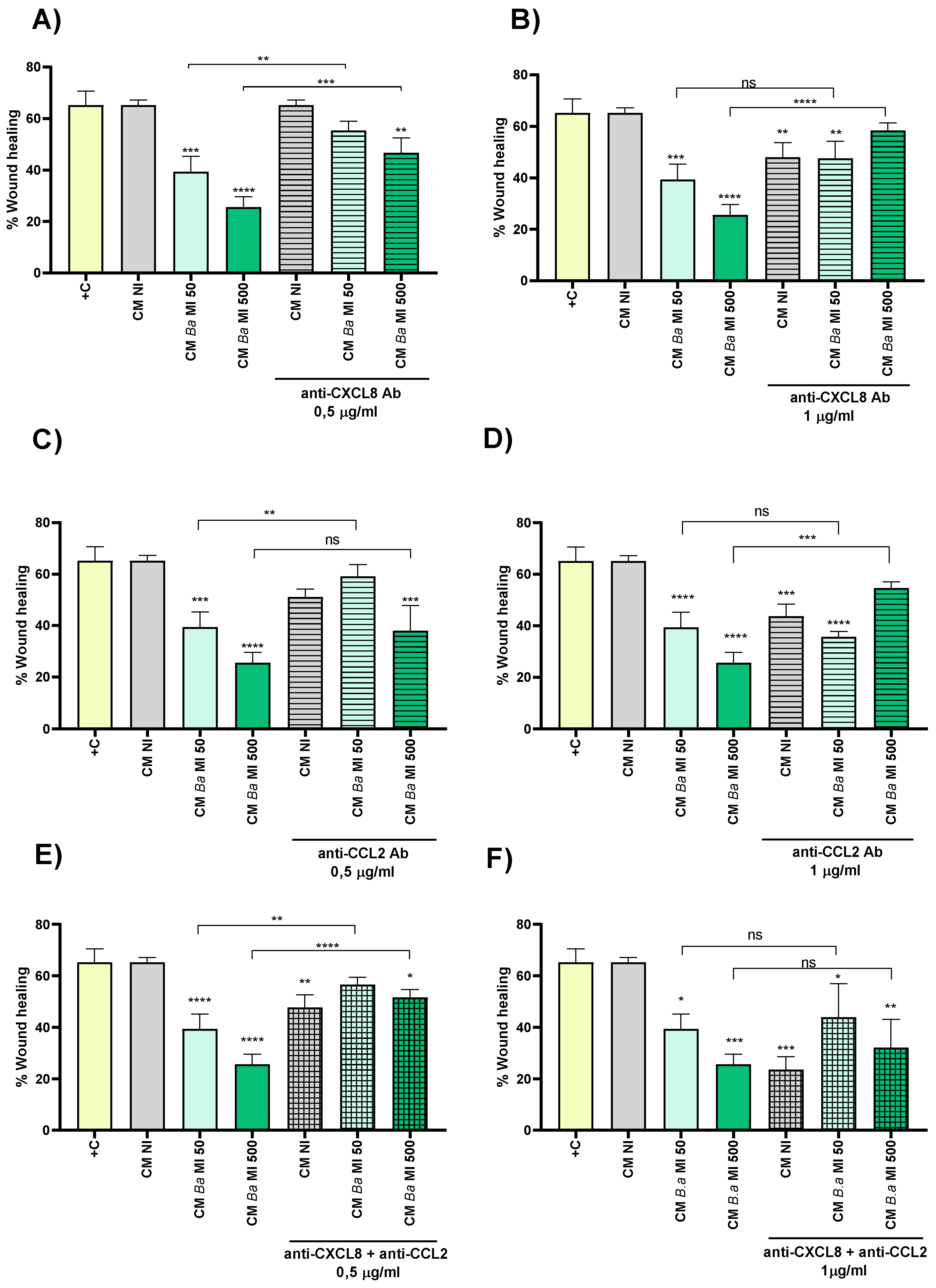
Figure 7.
Conditioned medium from infected decidualized stromal cells impairs trophoblast invasion capacity. Swan-71 trophoblasts were incubated for 1 hour with conditioned medium (CM) from decidual cells infected with B. abortus (Ba), B. suis (Bs) or B. melitensis (Bm), or from non-infected cells (NI). Then, trophoblasts were seeded onto a protein matrix-coated membrane placed in the upper compartment of a Transwell system and were maintained with continuous exposure to CM. The lower compartment was supplemented with DMEM/F12 containing 10% FBS to promote invasion. At 18 h of culture, the cells retained on the membrane were stained and quantified using microscopy-based image analysis. The same experiment was performed with pre-treatment of CM from both infected and non-infected cells with neutralizing antibodies against CXCL8 (A) or CCL2 (B). The results represent the number of cells in the membrane (migrating cells) and are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over bars indicate differences versus the CM NI condition whereas asterisks over lines indicate differences between antibody-treated and untreated conditions (** p<0.01; *** p<0.001; **** p<0.0001).
Figure 7.
Conditioned medium from infected decidualized stromal cells impairs trophoblast invasion capacity. Swan-71 trophoblasts were incubated for 1 hour with conditioned medium (CM) from decidual cells infected with B. abortus (Ba), B. suis (Bs) or B. melitensis (Bm), or from non-infected cells (NI). Then, trophoblasts were seeded onto a protein matrix-coated membrane placed in the upper compartment of a Transwell system and were maintained with continuous exposure to CM. The lower compartment was supplemented with DMEM/F12 containing 10% FBS to promote invasion. At 18 h of culture, the cells retained on the membrane were stained and quantified using microscopy-based image analysis. The same experiment was performed with pre-treatment of CM from both infected and non-infected cells with neutralizing antibodies against CXCL8 (A) or CCL2 (B). The results represent the number of cells in the membrane (migrating cells) and are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over bars indicate differences versus the CM NI condition whereas asterisks over lines indicate differences between antibody-treated and untreated conditions (** p<0.01; *** p<0.001; **** p<0.0001).
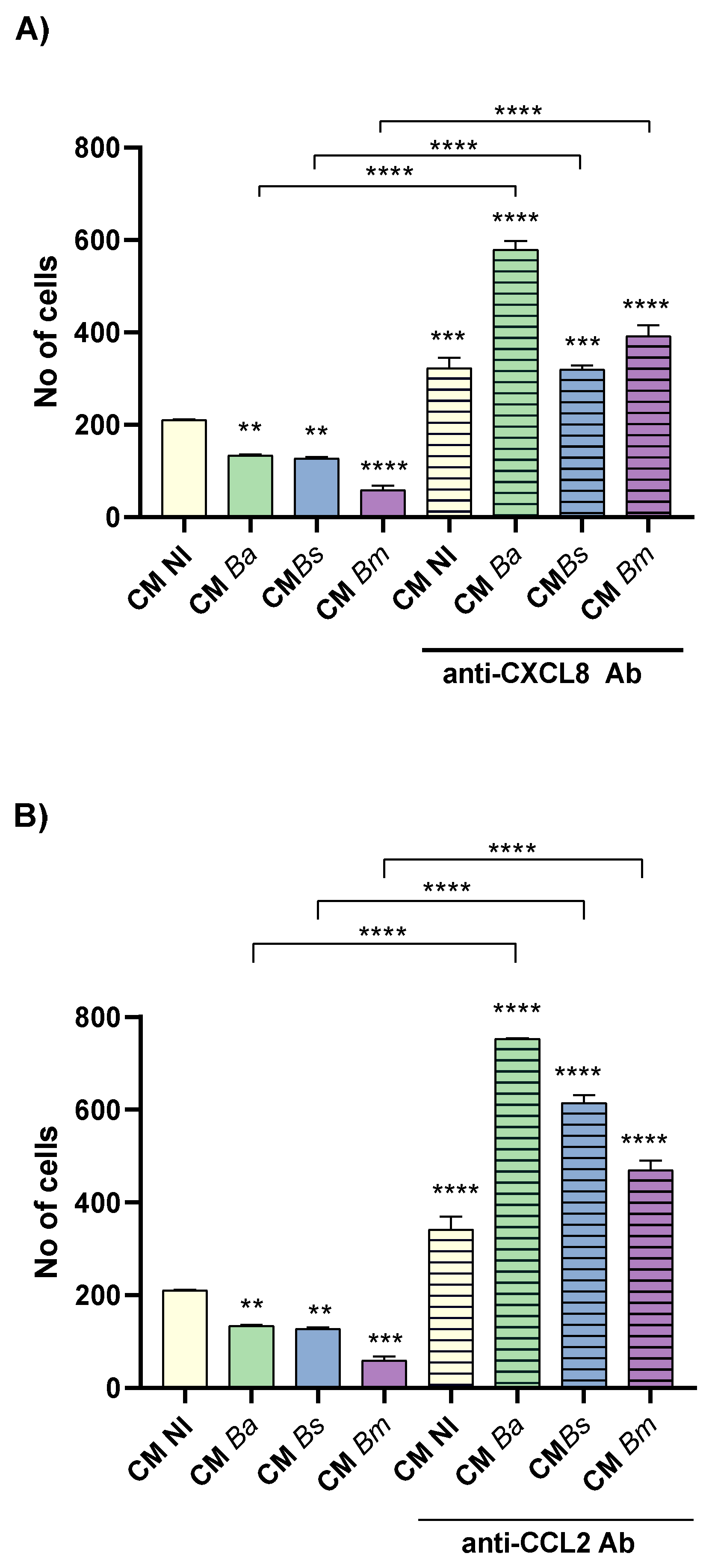
Figure 8.
Brucella infection of decidualized stromal cells impairs their chemotactic capacity for trophoblasts. Swan-71 cells suspended in medium with 2% FBS were seeded onto a protein matrix-coated membrane placed in the upper compartment of a Transwell system. Conditioned media (CM) from decidual stromal cells infected with B. abortus (Ba), B. suis (Bs) or B. melitensis (Bm) or from non-infected cells (NI) were placed in the lower compartment to evaluate chemotactic activity. After 6 hours of incubation, cells retained on the membrane were stained and quantified using microscopy-based image analysis. The same experiment was performed with pre-treatment of CM from both infected and non-infected cells with neutralizing antibodies against CXCL8 (A) or CCL2 (B). The results represent the number of cells in the membrane (migrating cells) and are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over groups indicate differences versus the CM NI condition, while asterisks over lines indicate differences between antibody-treated and untreated conditions (***p<0.001; ****p<0.0001; ns: non-significant).
Figure 8.
Brucella infection of decidualized stromal cells impairs their chemotactic capacity for trophoblasts. Swan-71 cells suspended in medium with 2% FBS were seeded onto a protein matrix-coated membrane placed in the upper compartment of a Transwell system. Conditioned media (CM) from decidual stromal cells infected with B. abortus (Ba), B. suis (Bs) or B. melitensis (Bm) or from non-infected cells (NI) were placed in the lower compartment to evaluate chemotactic activity. After 6 hours of incubation, cells retained on the membrane were stained and quantified using microscopy-based image analysis. The same experiment was performed with pre-treatment of CM from both infected and non-infected cells with neutralizing antibodies against CXCL8 (A) or CCL2 (B). The results represent the number of cells in the membrane (migrating cells) and are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over groups indicate differences versus the CM NI condition, while asterisks over lines indicate differences between antibody-treated and untreated conditions (***p<0.001; ****p<0.0001; ns: non-significant).
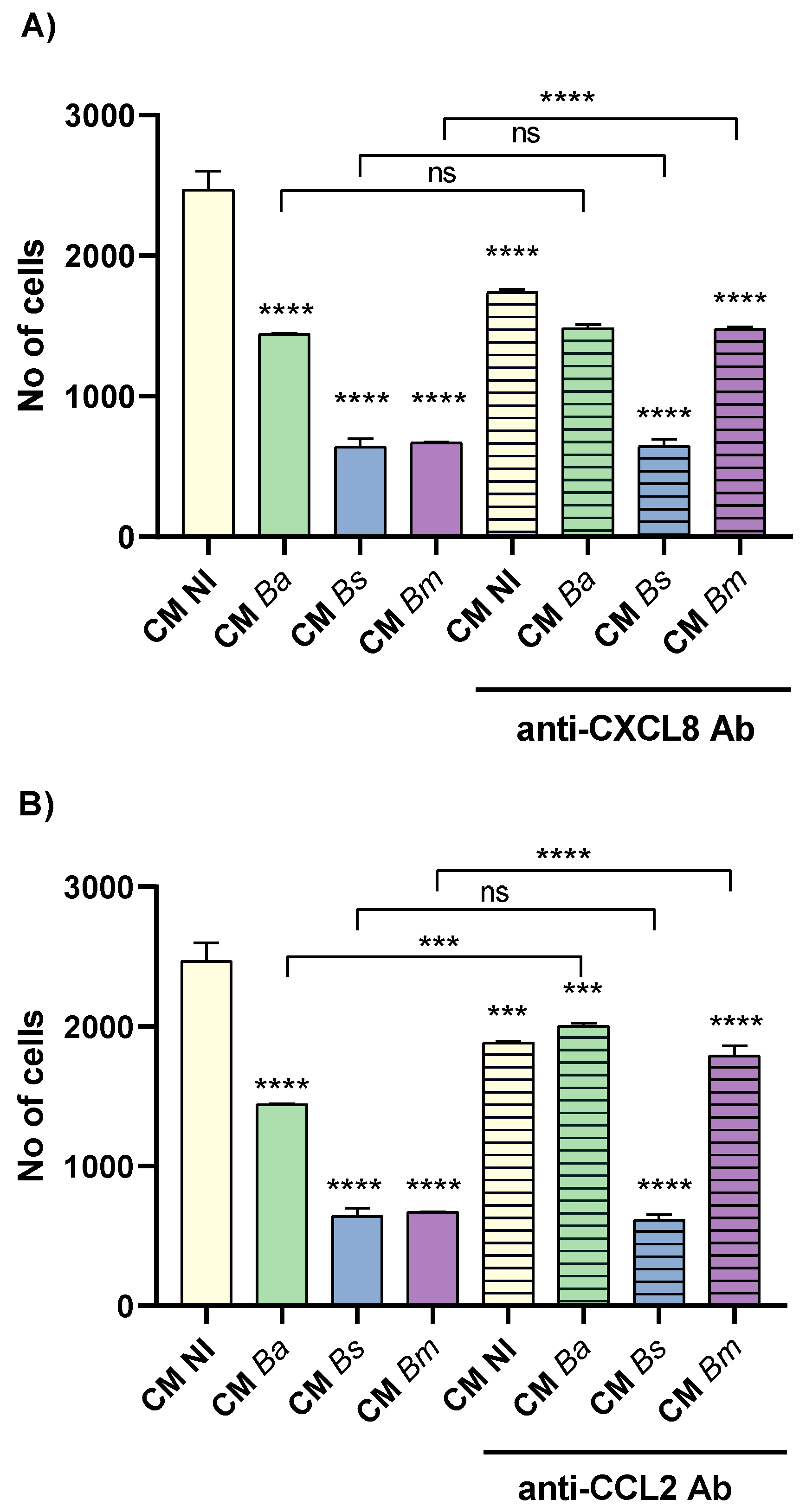
Figure 9.
Factors produced by Brucella-infected decidualized stromal cells impair trophoblast tubulogenesis. Swan-71 cells were seeded on a protein matrix (Geltrex) and incubated for 6 h with conditioned media (CM) from B. abortus, B. suis or B. melitensis-infected decidual cells (MI 50, 250 or 500) or from non-infected cells (NI), or were left untreated (Basal). After incubation, images were captured using a microscope to assess the number of master segments (A), meshes (B) and master junctions (C). The results are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over the bars indicate differences versus the Basal condition whereas asterisks over lines indicate differences versus CM NI (** p<0.01; *** p<0.001; **** p<0.0001; ns: non-significant). Representative images of the assay are shown in panel D.
Figure 9.
Factors produced by Brucella-infected decidualized stromal cells impair trophoblast tubulogenesis. Swan-71 cells were seeded on a protein matrix (Geltrex) and incubated for 6 h with conditioned media (CM) from B. abortus, B. suis or B. melitensis-infected decidual cells (MI 50, 250 or 500) or from non-infected cells (NI), or were left untreated (Basal). After incubation, images were captured using a microscope to assess the number of master segments (A), meshes (B) and master junctions (C). The results are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over the bars indicate differences versus the Basal condition whereas asterisks over lines indicate differences versus CM NI (** p<0.01; *** p<0.001; **** p<0.0001; ns: non-significant). Representative images of the assay are shown in panel D.
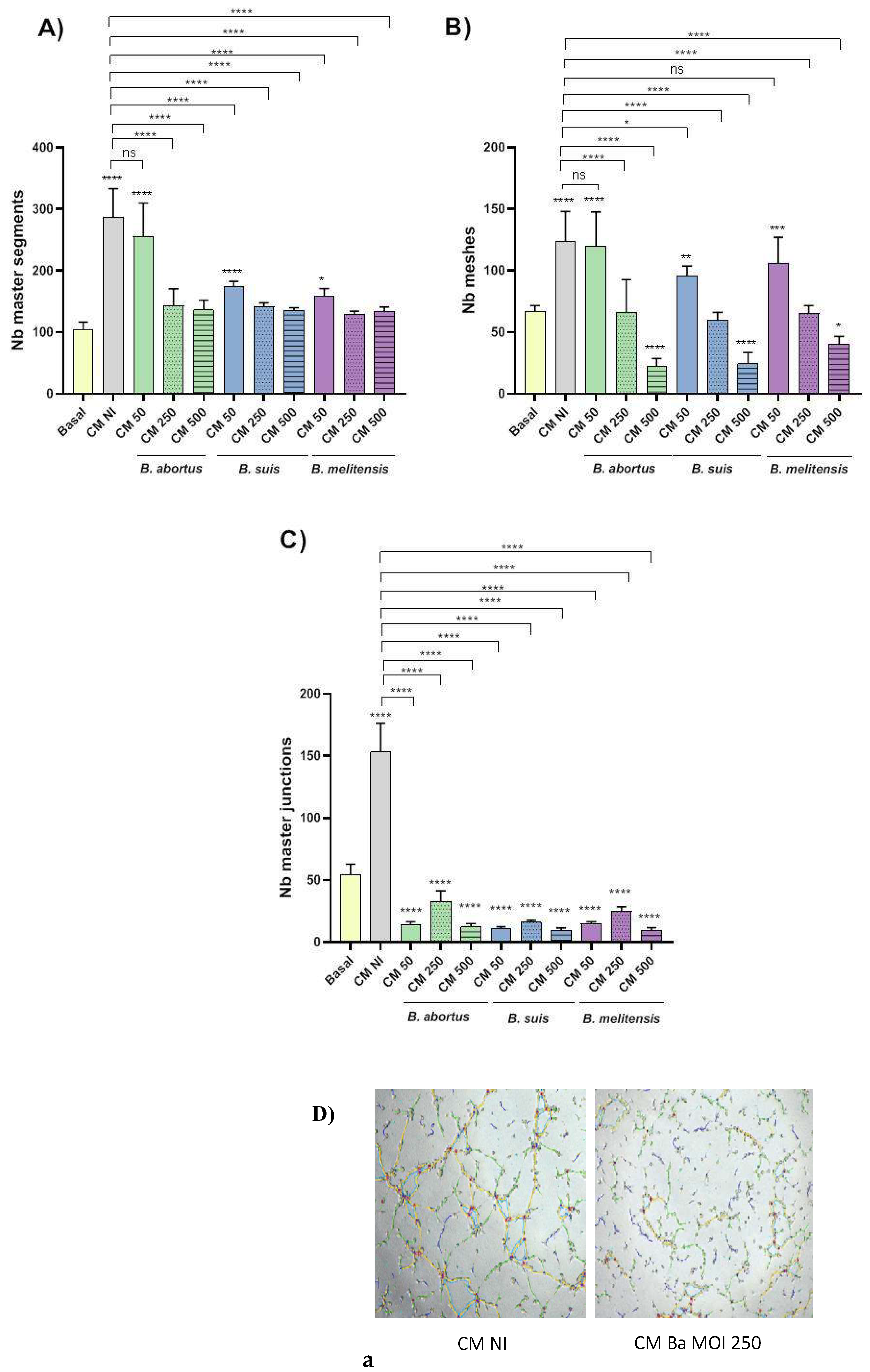
Figure 10.
Proinflammatory chemokines induced by Brucella infection in decidual cells contribute to the reduced ability of trophoblasts to form tubular structures. A tubulogenesis assay was performed as indicated in Figure 9, but conditioned media (CM) from Brucella-infected decidual cells were preincubated with neutralizing antibodies (Ab) against CXCL8 or CCL2 before addition to Swan-71 trophoblasts. After incubation, images were captured using a microscope to assess the number of master segments (A), meshes (B) and master junctions (C). The results are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over bars indicate differences versus the untreated CM NI (white bar), whereas asterisks over lines indicate differences between each Ab-treated and the untreated CM for each Brucella species (*p <0.05; ** p<0.01; **** p<0.0001; ns: non-significant).
Figure 10.
Proinflammatory chemokines induced by Brucella infection in decidual cells contribute to the reduced ability of trophoblasts to form tubular structures. A tubulogenesis assay was performed as indicated in Figure 9, but conditioned media (CM) from Brucella-infected decidual cells were preincubated with neutralizing antibodies (Ab) against CXCL8 or CCL2 before addition to Swan-71 trophoblasts. After incubation, images were captured using a microscope to assess the number of master segments (A), meshes (B) and master junctions (C). The results are expressed as mean ± SD of three independent experiments performed in duplicate. Asterisks over bars indicate differences versus the untreated CM NI (white bar), whereas asterisks over lines indicate differences between each Ab-treated and the untreated CM for each Brucella species (*p <0.05; ** p<0.01; **** p<0.0001; ns: non-significant).
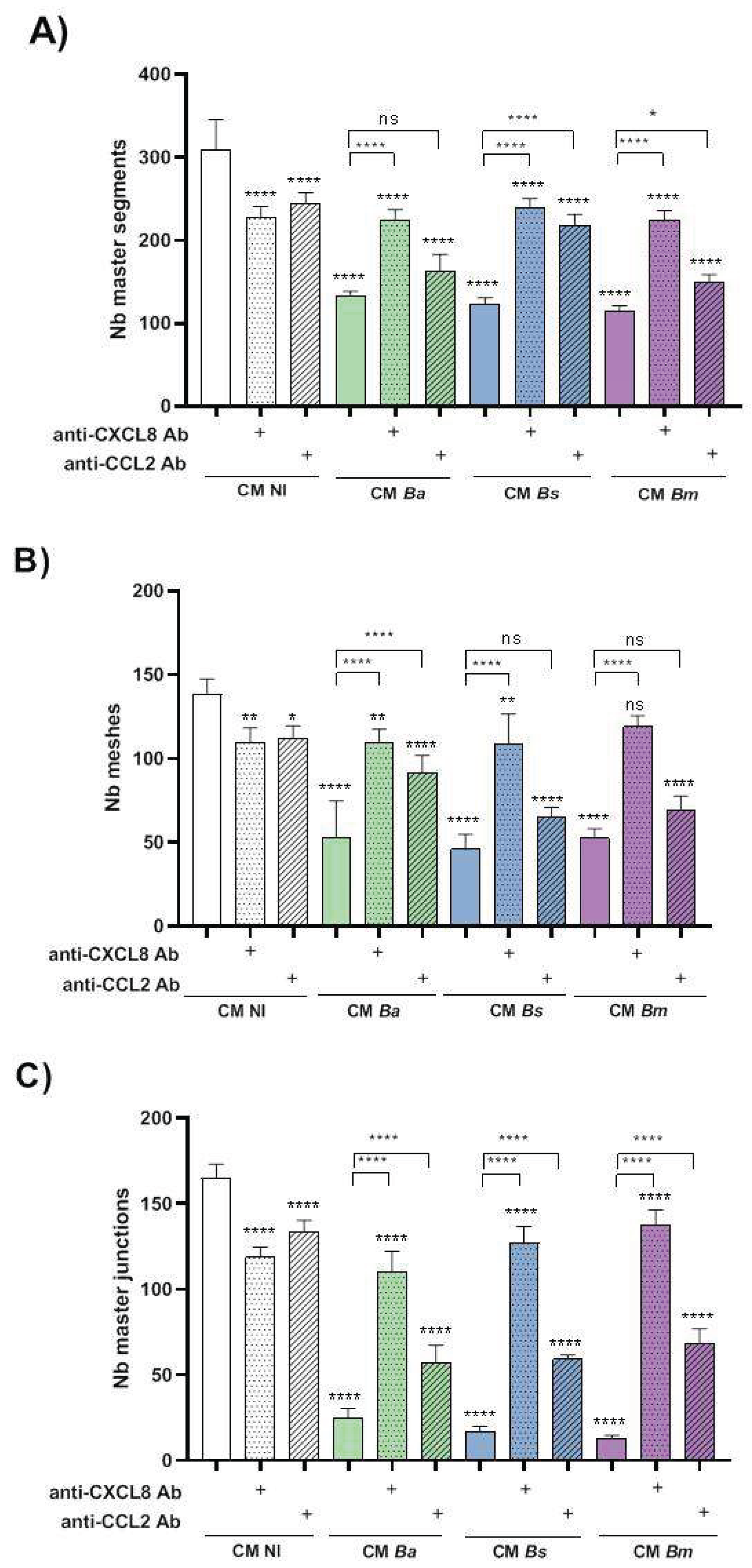
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



