Submitted:
04 July 2025
Posted:
08 July 2025
You are already at the latest version
Abstract
Alcohols are biostimulants and growth enhancers in agricultural crops. The aim of the present study was to improve the growth of freesia refracta via photosynthesis enhance-ment following foliar application of 0, 10, 20, and 30% methanol and ethanol. The highest net photosynthesis (10.54 µmol CO2 m-2 s-1) was obtained by spraying of 30% methanol + 30% ethanol which was significantly higher than the lowest mean value (3.18 µmol CO2 m-2 s) obtained by spraying of 10% ethanol. Similarly, plants treated by 30% methanol + 30% ethanol showed the highest transpiration rate (3.68 mol H2O m-2 s-1), instantaneous water-use efficiency (3.67 µmol CO2. mmol-1 H2O) and mesophyll conductance (0.057 mmol CO2 m-2 s-1). The heaviest corms (12.43 g) were produced by spraying of 10% meth-anol which was approximately 2.5 times higher than the corm weight of non-treated plants (5.13 g). Spraying of 10% methanol + 30% ethanol doubled dry weight of florets compared to non-treated plants (0.92 g vs. 0.47 g). Statistical analyses confirmed that methanol was more effective than ethanol for improving plant growth of studied genotype of freesia. Findings of this work help greenhouse growers and open field farmers to im-prove the growth of their crops by alcohol application.
Keywords:
alcohol spraying
; foliar carbon nutrient
; photosynthesis
; underground organs
1. Introduction
Freesia is an ornamental cormous plant from the Iridaceae family placed among top ten cut flowers sold in the world. Flowers are fragrant and come in a wide range of colors. Tetraploid cultivars of modern freesia (Freesia ×hybrida Bailey) are widely sold in the international flower market [1]. These cultivars possess long floral scape and larger corms than diploids. However, cultivars of F. refracta, a diploid species, produce more pleasant fragrant flowers, makes them very popular. Seeds, corms and tissue culture techniques are used to propagate freesias [1,2]. Propagation by corms is a usual method used by many growers. Thus, increase in the weight of daughter corms produced in the last season improves the quality of next year flowers. Moreover, the number of cormlets produced by freesia is low (2-3 cormlets per plant) and increase in the number and weight of cormlets is one of the main aims of freesia propagators. Enhancing photosynthesis is one of the main horticultural practices that provides more assimilates to increase the size and number of underground organs of geophytes. CO2 enrichments is a way to enhance photosynthesis and promote plant growth [3,4]. However, this technique has some difficulties and its operation and handling is not easy for all growers and may be not feasible in all greenhouses. Interestingly, literature reviews show that foliar application of some alcohols can be a beneficial cultural practice to enhance photosynthesis and growth of some crops [5,6,7].
Methanol (CH3OH) is endogenously produced within plant tissues. Todays, methanol is considered more than as a plant by-product. Beside its impact on the physiological processes, methanol emitted by plants triggers signaling pathways to improve plant immunity. In fact, gaseous methanol plays a key role in plant-plant and plant-animal communications. Emission of methanol by damaged plants triggers defense reactions in other healthy plants and thus activates methanol-inducible genes (MIGs) that regulates plant resistance [8]. Methanol is mainly generated by pectin demethylesterification, especially following cell wall damage. Thus, wounding or other stresses increase the expression of pectin methylesterases [9,10]. Methanol is also biosynthesized by demethylation of macromolecules (nucleic acids and proteins) [10]. There are some evidences indicate that foliar application of methanol could improves plant growth. Exogenously applied methanol enhances carbon fixation by plant tissues and thus improves plant growth and yield. In bell pepper (Capsicum annuum L.), foliar application of methanol significantly increased carbon conversion efficiency. Three times treatment of tomato (Lycopersicon esculentum) plants by 10% methanol under direct sunlight increased the number of internodes (12-16) compared to non-treated plants (9-10). Stem diameter was increased by 25-50%. Fruits contained 10-12% greater sugar and were harvested 5-10 days earlier than non-treated plants [5]. Moreover, methanol spraying increases cell turgidity and reduces water requirement in some crops [11].
Ethanol (CH3CH2OH) is an organic compound produced in response to environmental stresses and plays critical physiological roles in plants. Ethanol is primarily generated by alcohol dehydrogenase (ADH) [12,13]. As ADH enzymes are highly active under stress conditions, exogenous application of ethanol is suggested to mitigate stress impacts on the plants [14,15]. Not only ethanol increases the plant tolerance to abiotic stress, but also possesses an impact on the plant growth and development. Wu et al. (2019) reported that application of low concentrations of ethanol (0.0125-0.05 ml/L) increased the biomass of oilseed rape by 16.62-44.08%. Nutrients accumulation was also affected by exogenous ethanol. They found that total nitrogen, phosphorus, and potassium of the studied plants was raised. However, higher concentrations of ethanol inhibited the growth [16]. In soybean, spraying of 20 mM ethanol improved growth performance of stressed plants by increase in biomass, leaf area, photosynthetic pigment contents, gas exchange parameters, water-use-efficiency, and leaf relative water content [17].
The aims of the present study were to 1) reveal weather methanol and ethanol spraying enhances net photosynthesis rate and subsequent growth and flowering of freesia; 2) evaluate the effects of methanol and ethanol on the production and enlargement of freesia corm/cormlet which improves their propagation rate.
2. Materials and Methods
2.1. Plant Materials and Experimental Conditions
Corms of a local genotype of diploid freesia (Freesia refracta Klatt) were obtained from gene bank of Iranian Center for Flower and Ornamental Plants Research. After disinfection by fungicide (1.5% mancozeb), mother corms of similar size and weight were planted at 3 cm depth of 10 × 25 cm plastic pots filled with cocopeat (50%) + perlite (50%). Pots were kept in a plastic greenhouse located in Rafsanjan city (latitude 30°24′24″ N, longitude 55°59′38″ E, 1514 m above mean sea level).
The greenhouse equipment for climatic control were set to provide day and night temperatures of 24 ± 2 °C and 15 ± 2 °C, respectively. Relative humidity was maintained at approximately 40-50%. The natural light intensity received by plants was about 10000 – 12000 lux.
2.2. Alcohol Treatments
Fresh aqueous solutions of ethanol (0, 10, 20, and 30%) and methanol (0, 10, 20, and 30%) were sprayed on freesia plants. Alcohol sprays (20 ml per pot) were applied two times before flower harvest (on plants with at least 7 fully expanded leaves) and two times after flower harvest. The intervals between two applications was 7 days. A hand sprayer was used to apply alcohols on leaves.
2.3. Measured Traits
2.3.1. Determination of Leaf Chlorophyll Index (LCI) and Fluorescence
To non-destructively measure the chlorophyll content of leaves, a chlorophyll meter (SPAD, Konica, P502, Japan) was used to record the leaf chlorophyll index (LCI) of young fully expanded leaves. The average of three measurements per leaf was considered as LCI.
Measurement of chlorophyll fluorescence shows any probable stress caused by alcohol application. a portable Handy Plant Efficiency Analyzer (PEA, Hansatech, UK) was used to determine chlorophyll fluorescence (Bulgari et al., 2015). Leaf clips were attached to leaves, and after 15 min, the maximal quantum yield of PSII efficiency (Fv/Fm) and Performance Index (PI) were recorded on dark-adapted leaf tissues. PI shows the performance index for energy conservation from photons absorbed by the PSII antenna to the reduction of QB during photosynthesis.
2.3.2. Gas Exchange Measurements
Seven days after the last spray, gas exchange was recorded on fully expanded healthy leaves between 9:30 and 11:30 am. using a portable LCi photosynthesis system (ADC BioScientific Ltd., UK). Net photosynthetic rate (Pn, μmol CO2 m-2 s-1), stomatal conductance of H2O (gs, mol H2O m-2 s-1), mesophyll conductance of CO2 (gm, mmol m-2 s-1), internal CO2 (ci, vpm), and transpiration rate (E, mol H2O m-2 s-1) were determined on each leaf. The ratio of Pn to E was calculated and reported as instantaneous water use efficiency (WUEins, μmol CO2 mol-1 H2O).
2.3.3. Morphological Traits
Days to harvest, height of floral scape, length of flower axis, number of florets, florets dry weight, and floral scape dry weight were measured after flower harvest. At the end of the plant life, underground organs were lifted up and corm weight, number of cormlets, largest cormlet weight, and mean weight of all cormlets were measured/recorded. To determine dry weights, flora parts were placed overnight in a 70ºC oven.
2.4. Statistical Analyses
The experiment was carried out as factorial based on a completely randomized design (CRD) with four replicates. Factors were sprays of methanol (0, 10, 20, and 30%) and sprays of ethanol (0, 10, 20, and 30%). Data were subjected to a two-way analysis of variance (ANOVA) and means were compared using Duncan’s new multiple range test (DNMRT) at 5% level of significance (P ≤ 0.05) using IBM SPSS 27.
3. Results
3.1. Gas Exchange Measurements
Leaf gas exchange parameters were significantly affected by spraying of methanol and ethanol. Net photosynthesis (Pn) showed an increase of 54% by alcohol application. The maximum Pn (10.54 µmol CO2 m-2 s-1) was obtained by spraying of 30% methanol + 30% ethanol. In contrast, the minimum mean value (3.18 µmol CO2 m-2 s-1) was recorded for plants that received 10% ethanol (Figure 1). The quantum efficiency of photosynthesis (Pn/Qleaf) showed a relatively similar trend and was increased by 55%. The highest mean value of Pn/Qleaf (0.045) was recorded for plants sprayed with 10% methanol + 10% ethanol. However, plants that received 10% ethanol showed the lowest mean value (0.015) (Figure 2).
The transpiration rate (Tr) and instantaneous water-use efficiency (WUEins) were significantly changed by alcohol application. Plants sprayed by 30% methanol + 30% ethanol possess the highest Tr mean value (3.68 mol H2O m-2 s-1) which was approximately 35% higher than the mean value of non-treated plants (2.74 mol H2O m-2 s-1). The highest mean value of WUEins (3.67 µmol CO2. mmol-1 H2O) was observed in plants treated by 10% methanol + 10% ethanol. Spraying of 10% ethanol resulted in the lowest mean value of Tr (1.48 mol H2O m-2 s-1) and WUEins (2.16 µmol CO2. mmol-1 H2O), respectively (Figure 3 and Figure 4).
The mean values of sub-stomatal CO2 (Ci) were contrary to the results obtained for Pn and Tr. The highest Ci concentration (292.7 vpm) was observed by application of 10% ethanol. In contrast, plants that were sprayed by 30% methanol + 30% ethanol showed the lowest ci mean value (184.0 vpm) (Figure 5).
Stomatal conductance (gs) and mesophyll conductance (gm) of alcohol treated plants showed significant differences compared to non-treated plants. The lowest mean values of gs (53.33 nmol H2O m-2 s-1) and gm (0.013 mmol CO2 m-2 s-1) were obtained by foliar application of 10% ethanol. However, when ethanol was constituted by 20% methanol, the mean value of gs almost doubled (107.50 nmol H2O m-2 s-1). Mesophyll conductance showed the highest mean value (0.057 mmol CO2 m-2 s-1) in plants received 30% methanol + 30% ethanol (Figure 6 and Figure 7).
3.2. Morphological Traits
Daughter corms weight showed a significant increase by alcohol application. The heaviest corms (12.43 g) were produced by spraying 10% methanol which was approximately 2.5 times higher than the corm weight of non-treated plants (5.13 g). The lowest mean value of daughter corm weight (3.15 g) was recorded for plants treated by 10% methanol + 10% ethanol (Figure 8).
Number of cormlets changed depends on the concentration and combination of methanol and ethanol. Foliar application of 30% ethanol reduced the number of cormlets to 1.5, while the average number of cormlets produced by non-treated plants was 4. However, the highest mean value was obtained by spraying 10 % methanol + 10% ethanol or 30 % methanol + 30% ethanol (Figure 9).
The highest (1.92 g) and the lowest (0.33 g) weight of the largest cormlet were obtained using 20% methanol + 20% ethanol and by spraying 30% ethanol, respectively. The maximum (1.65 g) and the minimum (0.29 g) mean values recorded for the average weight of all cormlets showed a similar trend (Figure 10 and Figure 11).
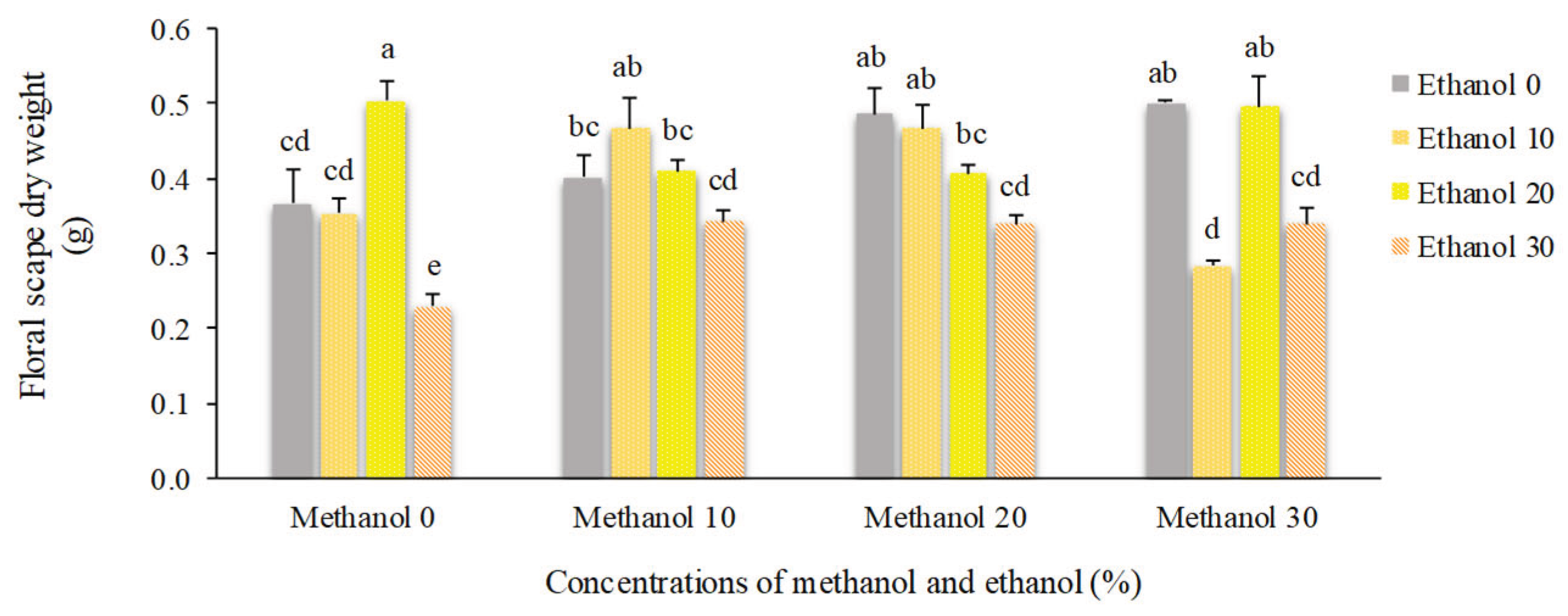
Application of 20% methanol resulted in the highest number of florets (13.25). Plants that received no alcohol possess 10 florets. However, higher concentrations of alcohol decreased the number of florets to 9. Spraying of 10% methanol + 30% ethanol doubled dry weight of florets compared to non-treated plants (0.92 g vs. 0.47 g). By foliar application of 20% ethanol, floral scape dry weight showed a 45% increase compared to non-treated plants. Higher concentrations of alcohol possess negative impacts on this trait (Figure 12, Figure 13 and Figure 14).
4. Discussion
The results indicated that photosynthetic parameters were dramatically changed by alcohol application. Although 10 or 30% ethanol negatively affected some of these parameters, higher concentrations of methanol + ethanol increased the efficacy of photosynthesis apparatus. Simultaneous application of methanol and ethanol increased net photosynthesis, transpiration rate, stomatal conductance, and mesophyll conductance compared to non-treated plants. In contrast, sub-stomatal conductance and water-use efficiency showed a significant decrease. It has been shown that under both greenhouse and field conditions alcohols are able to promote photosynthesis and subsequently increase the growth and yield of C3 plants [18,19,20]. However, there are contradictory reports in this regard. Rajala et al. (1998) concluded that comparing to the capacity of carbon fixation of plants canopy during growing season, applied concentrations of alcohols were not enough to increase photosynthesis and plant growth [21]. Similarly, alcohols did not improve photosynthesis of cotton (Gossypium hirsutum L.) [22]. Alcohols alters carbon fixation efficacy in plants via different pathways. Fructose-1,6-bisphosphatase (FBPase) is a key enzyme in photosynthesis. It has been reported that alcohols stabilize the binding between FBPase and thylakoid membrane and thus enhance its activity. The concentration needed to maximize this activity is decreased by increase in alcohol chain length [21]. In pea (Pisum sativum L.), FBPase activity was increased by application of methanol, ethanol, propanol, and butanol, probably by enhancing hydrophobic binding between FBPase and membrane-bound proteins [6]. Moreover, alcohols delay ethylene-induced senescence in plant tissues and thus prolong photosynthesis duration and increase CO2 fixation [23,24]. On the other hand, conversion of methanol to CO2, provides more carbon for photosynthesis [25]. It means that methanol is finally used to synthesis of sugars. Hemming et al. (1995) reported that the positive effects of methanol on the photosynthesis of bell pepper lasted several weeks [7]. It means that plant has enough time to assimilate the applied alcohols and unlike CO2 enrichment, application of alcohols may be time independent. Thus, we have much more freedom in using methanol than CO2 injection, as CO2 application should be in sunny days. However, the presence of light to encourage photosynthesis in next days is crucial. It has been shown that turgidity of guard cells is noticeably higher after methanol application, leads to more open stomata [5]. Turgid guard cells and open stomata permit more carbon dioxide fixation and plant growth. A concomitant increase is happened in transpiration rate which indicate good water conditions for plant growth [26].
The present study showed that foliar application of alcohols improved the growth of freesia plants. A sharp increase in the weight of corms and cormlets was recorded following foliar application of alcohols. Corm weight was increased at lower concentrations of methanol. By the treatments that the number of cormlets was higher, corm weight slightly decreased. It could be due to growth competition between cormlets and new daughter corms. As mentioned above, alcohols could enhance plant photosynthesis and thus provide more carbohydrates for new growth points. It could be concluded that improved growth of underground organs and increase in dry weight of floral parts is a result of enhanced photosynthesis and subsequently more availability of carbohydrates for growth. Interestingly, enhancing photosynthesis resulted in the higher number of cormlets than larger corms. Thus, treatments that improved photosynthesis can be useful to increase propagation rate of the studied freesia plants.
There are not a lot of reports on the growth promoting effects of alcohols in agricultural crops. However, among alcohols applied to promote plant growth, methanol has been in the heart of studies. In regard to the effects of alcohols on the plant growth there are contradictory reports. Some studies report positive effects and the other works indicate no changes in plant growth. Many researchers reported that alcohols have no effect on plant growth [22,27,28,29]. Rajala et al. (1998) studied the effects of foliar application of methanol and ethanol on the growth of some crops (barely, oat, wheat, pea, spring turnip rape) under field and greenhouse conditions. At both conditions, no positive effect was observed on the plant growth [21]. Contrary, foliar application of alcohols improved growth of tomato, pea, radish and wheat seedlings grown in greenhouse [19,20]. Nonomura and Benson (1992a) reported that methanol (10-50%) foliar application significantly increased biomass and water use efficiency of some C3 crops under warm and arid conditions. After treatment of savoy cabbages (Brassica oleracea capitata) leaves with 20% methanol, plant growth was improved in fall [5].
Different reports on the alcohols impact on the plant growth could be explained from various aspects. Rajala et al., (1998) believed that the reproducibility of beneficial effects of alcohols on the plant growth is questionable [21]. McGiffen and Manthey (1996) believed that methanol is not a reliable stimulant for improving plant growth [27]. In fact, even by duplicating the procedures defined by Nonomura and Benson’s (1992a, 1992b) [5,30], researchers did not achieve growth improvement by methanol application [31,32,33]. One of the main reasons is that alcohols are not useful in all crops and their impact is completely genotype dependent. For example, Hemming et al. (1995) reported that maize cultivars (C4 plants) did not respond to exogenous methanol application [7]. Nonomura and Benson (1992a) did not report growth improvement of C4 crops (corn, sorghum, bermuda grass, or johnson grass) by methanol application. They also stated that winter crops did not show growth improvement by methanol application [5]. Beside genotype, reaction of different plant tissues to alcohols is not the same. Cossins (1964) showed that over 90% of applied methanol to carrot root tissue was utilized, while only 10% was metabolized by pea cotyledons [25]. It is clear that alcohol concentration also plays a key role in the obtained results. Optimal concentrations of alcohols to promote plant growth varies with target tissue and plant species and varieties. Roots of tomato were damaged by 10% ethanol or methanol, whereas above ground organs withstood higher concentrations [20]. Similar results were reported by Hemming et al. (1995) on tomato [7]. Nonomura and Benson (1992) reported that trunk sections of some trees resist up to 100% methanol, while 20% methanol is optimal for enhancing cereals growth. They also showed that foliar application of 50% methanol was optimal for palm and eucalyptus, while eggplant required 10% methanol [5]. However, Rajala et al. (1998) found that up to 50% of methanol or ethanol was not deleterious for oat seedlings [21]. Similarly, pea and winter wheat tolerated up to 80–90% methanol with no symptoms of sever phytotoxicity [34]. On the other hand, environmental conditions and time of application highly affect the efficacy of alcohols application. Rajala et al. (1998) indicated that methanol and ethanol promoted the plant growth only under greenhouse conditions. In fact, they showed that field grown crops did not respond to alcohol treatments. They indicated that for field application of alcohols, penetration to leaf tissues is greater during night with lower temperatures and no wind result in lower evaporation [21]. Nonomura and Benson (1992a) showed that under shade, not only no growth improvement was obtained by methanol application, but also repeated applications resulted in phytotoxicity [5]. Contrarily, Cossins (1964) reported that darkness promotes methanol utilization and its conversion to sugars and amino acids [25].
In some commercial cultivars of greenhouse rose, methanol accelerated flowering and increased the number and weight of leaves and flowers. Plants treated with methanol bloomed within 62-70 days after treatment. Control plants that received no methanol treatment, required 75-80 days for flowering. In 'Miss All-American Beauty', flowers weighed 26 g while the flower weight of control plants was 18 g. In 'Paul Harris', the number of fully opened flowers was doubled compared to control (8 vs 4 flowers) at the first harvest. Moreover, methanol treated plants were healthier and pest-free [5]. Ten daily application of 20% methanol (contained glycine + glycerophosphate) on Coleus blumei increased plant weight (43 g) compared to the weight (36 g) of each water sprayed plant [5]. Foliar application of methanol and ethanol on greenhouse-grown tomato plants increased stem and leaf weight up to 19%. Methanol showed a somewhat greater effect than ethanol [20].
On the other hand, alcohols may provide more nutrients required for plant growth. For example, methanol can promote plant growth by altering the nutrients content in plant tissues. Methanol-soluble nutrients have been used to correct nutrients deficiency in some crops. In fact, it has been hypothesized that methanol maybe acts as a carrier of low water-soluble nutrients in plant tissues [33,35]. In rose, 0.08 g Fe-EDTA in methanol was used to promote plant growth. Repeated applications of iron and nitrogen methanolic solutions corrected nutrient-deficiency symptoms in citrus and rice [35].
Besides providing carbon source and nutrients, alcohols may act as osmoprotectants to protect plant physiological processes especially under stress conditions. Moreover, alcohols blocks ethylene-induced senescence in plant tissues and enhance plant recovery after stress. On the other hand, increase in leaf sugars content following methanol application changes osmotic potential of cells that leads to higher turgidity and stomata conductance. Open stomata allow to higher photosynthesis, plant growth, and maturation [27]. In rose, plants treated by 20% methanol (supplemented by glycine and glycerophosphate) produced turgid leaves and flowers. Foliar application of 10% methanol (contained glycine + glycerophosphate) improved turgidity and growth of Chrysanthemum indicum, Dieffenbachia sequine, Syngonium podophyllum, Scindapsus aureus and Ficus elastica [5]. Moreover, foliar application of methanol has increased turgidity and reduced irrigation requirement in some crops under field conditions. Under high temperatures and direct noon sunlight in cotton fields, foliar application of 30% methanol increased leaf turgidity and plants were 15% taller than non-treated plants. Leaf surface area and thickness were increased by 20% and 20-50%, respectively. Treated plants required fewer irrigation cycles that reduced water orders by half, from 122 × 105 liters/ha to 60 × 105 liters/ha [5]. Similar results in savoy cabbage were reported by these authors. They showed that under high temperatures plants were wilted unless those treated by 20% methanol. Moreover, they showed that three applications of 20% methanol contained soluble nutrients maintained the turgidity of durum wheat (Triticum durum) plants under water stress and direct sunlight. Treated plants showed larger leaves and more seeds. However, it seems that under hydroponic conditions that plants receive enough water and nutrients via fertigation, alcohol application may not result in significant improvement in plant turgidity. Under field conditions plants are exposed to sunlight and wind. Thus, under these conditions, application of alcohols significantly improves plants turgidity.
5. Conclusions
The present study showed that foliar application of alcohols, especially methanol, encourages the production of photoassimilates and growth of plants. Statistical analyses confirmed that low concentrations of methanol are beneficial for corm growth and weighing. However, higher concentrations of methanol + ethanol improves photosynthesis and the number of cormlets. Totally, methanol was more effective than ethanol for improving plant growth of studied genotype of freesia. Findings of this work suggest that methanol can be as a foliar carbon nutrient in plants. However, it may be highly genotype and concentration dependent. Moreover, assessing the substitution of CO2 enrichment with methanol application, as a carbon source, in greenhouse production of plants could be suggested.
Funding
This research was funded by Vali-e-Asr University of Rafsanjan.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections.
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Pn | Net photosynthesis |
| Pn/Qleaf | The quantum efficiency of photosynthesis |
| Tr | Transpiration rate |
| WUEins | Instantaneous water-use efficiency |
| Ci | Sub-stomatal CO2 |
| gs | Stomatal conductance |
| gm | Mesophyll conductance |
References
- Anderson, N. O. Flower breeding and genetics issues, challenges and opportunities for the 21st century; Springer: The Netherlands, 2007; pp. 665–693. [Google Scholar]
- De Hertogh, A.; Le Nard, M. The physiology of flower bulbs; Elsevier: The Netherlands, 1993; pp. 285–296. [Google Scholar]
- Nelson, P.V. Greenhouse operation and management – Seventh Edition; Pearson: USA, 2014; pp. 336–342. [Google Scholar]
- Wang, A.; Lv, J.; Wang, J.; Shi, K. CO2 enrichment in greenhouse production: Towards a sustainable approach. Front. Plant Sci. 2022, 13, 1029901. [Google Scholar] [CrossRef]
- Nonomura, A.M.; Benson, A.A. The path of carbon in photosynthesis: Improved crop yields with methanol. Proc. Nati. Acad. Sci. 1992, 89, 9794–9798. [Google Scholar] [CrossRef] [PubMed]
- Andre´s, R.; La`zaro, J.; Chueca, A.; Hermoso, R.; Gorge´, L. Effect of alcohols on the association of photosynthetic fructose-1,6-bisphosphatase to thylakoid membranes. Physiol. Plant. 1990, 78, 409–413. [Google Scholar] [CrossRef]
- Hemming, D.; Criddle, R.; Hansen, L. Effects of methanol on plant respiration. J. Plant Physiol. 1995, 146, 193–198. [Google Scholar] [CrossRef]
- Dorokhov, Y.L.; Sheshukova, E.V; Komarova, T.V. Methanol in plant life. Front. Plant Sci. 2018, 9, 1623. [Google Scholar] [CrossRef] [PubMed]
- Komarova, T.V.; Sheshukova, E.V.; Dorokhov, Y.L. Cell wall methanol as a signal in plant immunity. Front. Plant Sci. 2014, 5, 101. [Google Scholar] [CrossRef]
- Dorokhov, Y.L.; Shindyapina, A.V.; Sheshukova, E.V.; Komarova, T.V. Metabolic methanol: molecular pathways and physiological roles. Physiol. Rev. 2015, 95, 603–644. [Google Scholar] [CrossRef]
- Nishio, J.N.; Huang, S.; Winder, T.L.; Brownson, M.P.; Ngo, L. Physiological aspects of methanol feeding to higher plants. In Proceedings of the 20th Annual Meeting of Plant Growth Regulator Society of America, USA; 1994; pp. 8–13. [Google Scholar]
- Speight, J.G. Biomass Processes and Chemicals; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Tesniere, C.; Torregrosa, L.; Pradal, M.; Souquet, J.-M.; Gilles, C.; Dos Santos, K.; Chatelet, P.; Gunata, Z. Effects of genetic manipulation of alcohol dehydrogenase levels on the response to stress and the synthesis of secondary metabolites in grapevine leaves. J. Exp. Bot. 2006, 57, 91–99. [Google Scholar] [CrossRef]
- Das, A.K.; Anik, T.R.; Rahman, M.M.; Keya, S.S.; Islam, M.R.; Rahman, M.A.; Sultana, S.; Ghosh, P.K.; Khan, S.; Ahamed, T. Ethanol treatment enhances physiological and biochemical responses to mitigate saline toxicity in soybean. Plants 2022, 11, 272. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Sako, K.; Matsui, A.; Suzuki, Y.; Mostofa, M.G.; Ha, C.V.; Tanaka, M.; Tran, L.-S.P.; Habu, Y.; Seki, M. Ethanol enhances high-salinity stress tolerance by detoxifying reactive oxygen species in Arabidopsis thaliana and rice. Front. Plant Sci. 2017, 8, 1001. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; Jiang, L.; Zhang, Z.; Song, H.; Rong, X.; Han, Y. Low concentration of exogenous ethanol promoted biomass and nutrient accumulation in oilseed rape (Brassica napus L.). Plant Signal Behav. 2019, 14, 1681114. [Google Scholar] [CrossRef]
- Rahman, M.M.; Mostofa, M.G.; Das, A.K.; Anik, T.R.; Keya, S.S.; Ahsan, S.M.; Khan, M.A.R.; Ahmed, M.; Rahman, M.A.; Hossain, M.M.; Tran, L.S.P. Ethanol positively modulates photosynthetic traits, antioxidant defense and osmoprotectant levels to enhance drought acclimatization in soybean. Antioxidants 2022, 11, 516. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.; Prasad, R. Effect of plant growth regulators based on long chain aliphatic alcohols on seed and straw yield of lentil. Lens Newsl. 1990, 17, 19–20. [Google Scholar]
- Devlin, R.; Bhowmik, P.; Karczmarczyck, S. Influence of methanol on plant growth. PGRSA Q. 1994, 22, 102–108. [Google Scholar]
- Rowe, R.; Farr, D.; Richards, B. Effects of foliar and root applications of methanol or ethanol on the growth of tomato plants (Lycopersicon esculentum Mill. ). N.Z. J. Crop. Hort. Sci. 1994, 22, 335–337. [Google Scholar] [CrossRef]
- Rajala, A.; Karkkainen, J.; Peltonen, J.; Peltonen-Sainio, P. Foliar applications of alcohols failed to enhance growth and yield of C3 crops. Ind. Crops Prod. 1998, 7, 129–137. [Google Scholar] [CrossRef]
- Iersel, M.; Heitholt, J.; Wells, R.; Oosterhuis, M. Foliar methanol applications to cotton in the southeastern United States: leaf physiology, growth, and yield components. Agron. J. 1995, 87, 1157–1160. [Google Scholar] [CrossRef]
- Satler, S.; Thimann, K. The influence of aliphatic alcohols on leaf senescence. Plant Physiol. 1980, 66, 395–399. [Google Scholar] [CrossRef]
- Saltveit, M. Effect of alcohols and their interaction with ethylene on the ripening of epidermal pericarp discs of tomato fruit. Plant Physiol. 1989, 90, 167–174. [Google Scholar] [CrossRef]
- Cossins, E.A. The utilization of carbon-1-compounds by plants. Can. J. Bot. 1964, 42, 1793–1802. [Google Scholar]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- McGiffen, M.E.; Manthey, J.A. The role of methanol in promoting plant growth: A current evaluation. HortScience. 1996, 31, 1092–1096. [Google Scholar] [CrossRef]
- Esensee, V.; Leskovar, D.; Boales, A. Inefficacy of methanol as a growth promoter in selected vegetable crops. HortTechnology. 1995, 5, 253–256. [Google Scholar] [CrossRef]
- Feibert, E.; James, S.; Rykbost, K.; Mitchell, A.; Shock, A. Potato yield and quality not changed by foliar-applied methanol. HortScience. 1995, 30, 494–495. [Google Scholar] [CrossRef]
- Nonomura, A.M.; Benson, A.A. The path of carbon in photosynthesis: Methanol and light. In Research in photosynthesis; Murata, N., Ed.; Kluwer Academic Publisher: Dordrecht, Netherlands, 1992b; pp. 911–914. [Google Scholar]
- Hartz, T.K.; Mayberry, K.S.; McGiffen, M.E., Jr.; LeStrange, M.; Miyao, G.; Baameur, A. Foliar methanol application ineffective in tomato and melon. HortScience, 1994, 22, 1087. [Google Scholar] [CrossRef]
- Mauney, J.R.; Gerik, T.J. Evaluating methanol usage in cotton. In: Proc. Beltwide Cotton Conf., San Diego, Calif., vol. 1. Natl. Cotton Council of Amer., Memphis, Tenn, 1994; pp. 39–40.
- McGiffen, M.E.; Green, R.L.; Manthey, J.A.; Faber, B.A.; Downer, A.J.; Sakovich, N.J.; Aguiar, J. Field tests of methanol as a crop yield enhancer. HortScience 1995, 30, 1225–1228. [Google Scholar] [CrossRef]
- Albrecht, S.L.; Douglas, C.L., Jr.; Klepper, E.L.; Rasmussen, P.E.; Rickman, R.W.; Smiley, R.W.; Wilkins, D.E.; Wysocki, D.J. Effects of foliar methanol applications on crop yield. Crop Sci. 1995, 35, 1642–1646. [Google Scholar] [CrossRef]
- Nonomura, A.M.; Nishio, J.N.; Benson, A.A. Stimulated growth and correction of Fe deficiency with trunk- and foliar-applied methanol- soluble nutrient amendments. In: Iron nutrition in soils and plants. J. Abadia (ed.). Kluwer Academic Publisher, Dordrecht, Netherlands, 1995; pp. 329–333.
Figure 1.
The effect of methanol and ethanol spraying on the net photosynthesis (Pn) of freesia leaves.
Figure 1.
The effect of methanol and ethanol spraying on the net photosynthesis (Pn) of freesia leaves.
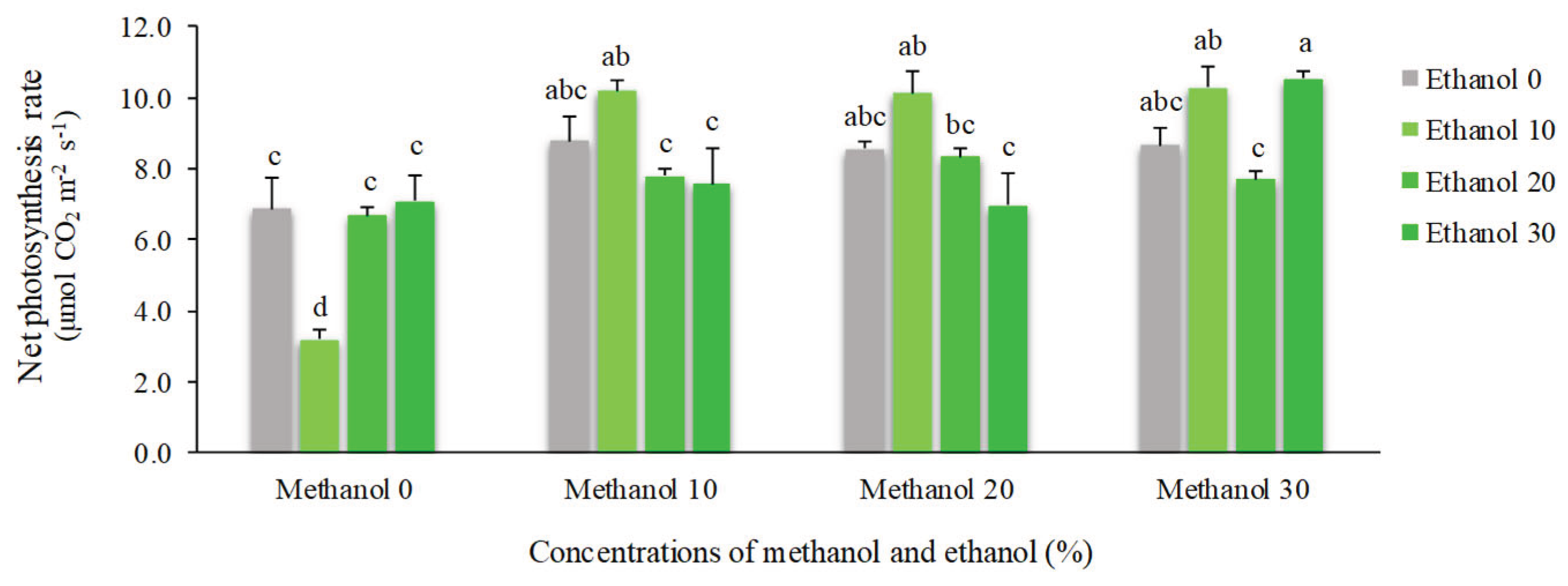
Figure 2.
The effect of methanol and ethanol spraying on the quantum efficiency of photosynthesis (Pn/Qleaf) of freesia leaves.
Figure 2.
The effect of methanol and ethanol spraying on the quantum efficiency of photosynthesis (Pn/Qleaf) of freesia leaves.
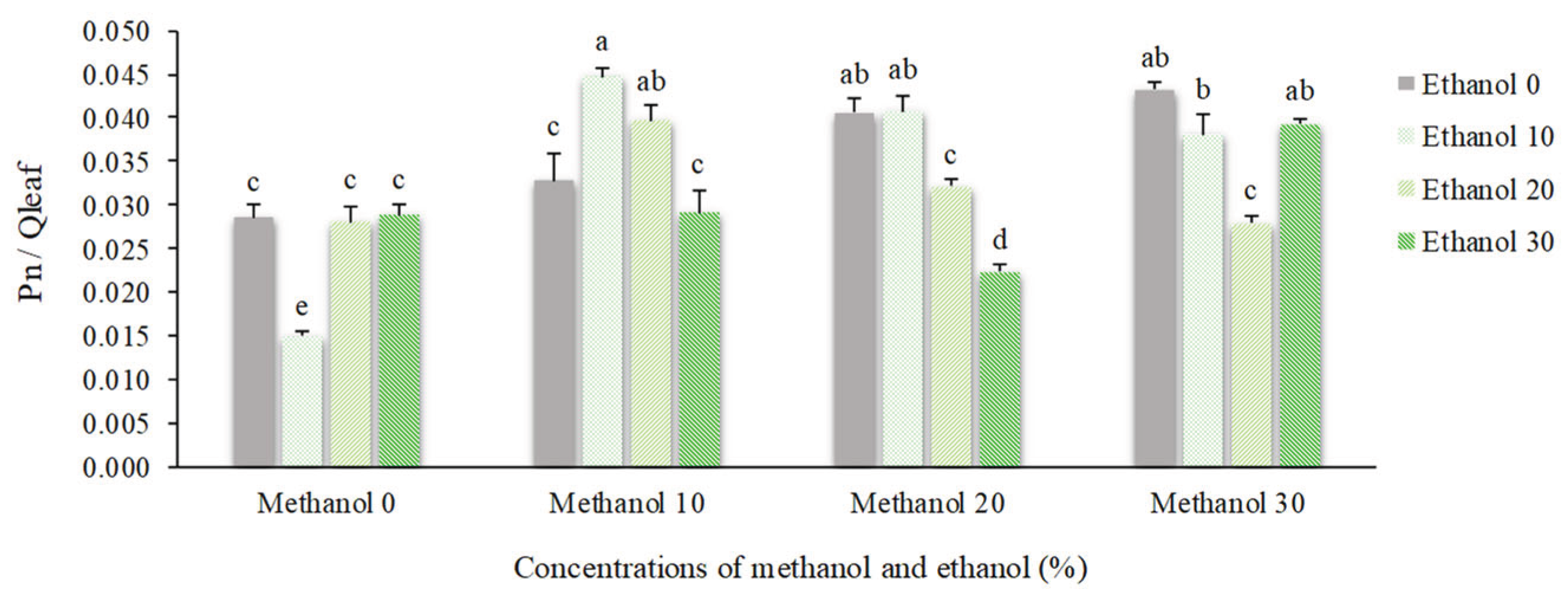
Figure 3.
The effect of methanol and ethanol spraying on the transpiration rate (Tr) of freesia leaves.
Figure 3.
The effect of methanol and ethanol spraying on the transpiration rate (Tr) of freesia leaves.
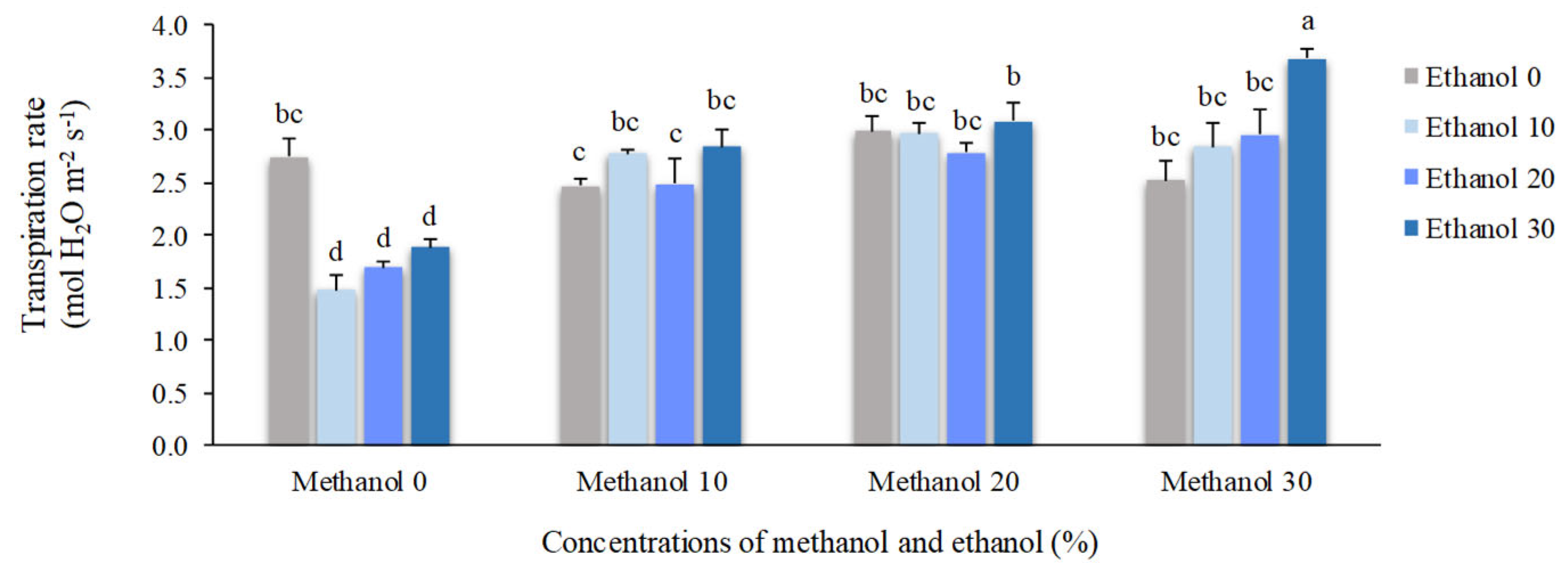
Figure 4.
The effect of methanol and ethanol spraying on the water-use efficiency (WUEins) of freesia leaves.
Figure 4.
The effect of methanol and ethanol spraying on the water-use efficiency (WUEins) of freesia leaves.
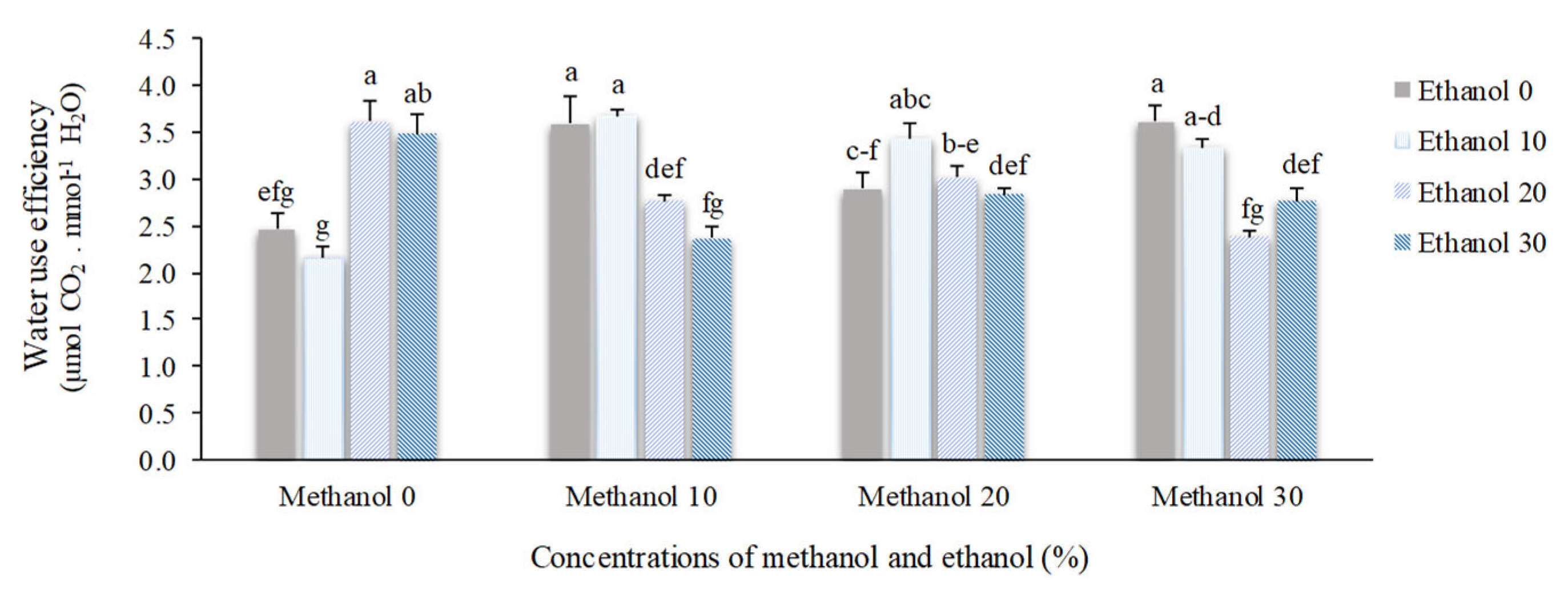
Figure 5.
The effect of methanol and ethanol spraying on the sub-stomatal CO2 (Ci) of freesia leaves.
Figure 5.
The effect of methanol and ethanol spraying on the sub-stomatal CO2 (Ci) of freesia leaves.

Figure 6.
The effect of methanol and ethanol spraying on the stomatal conductance (gs) of freesia leaves.
Figure 6.
The effect of methanol and ethanol spraying on the stomatal conductance (gs) of freesia leaves.
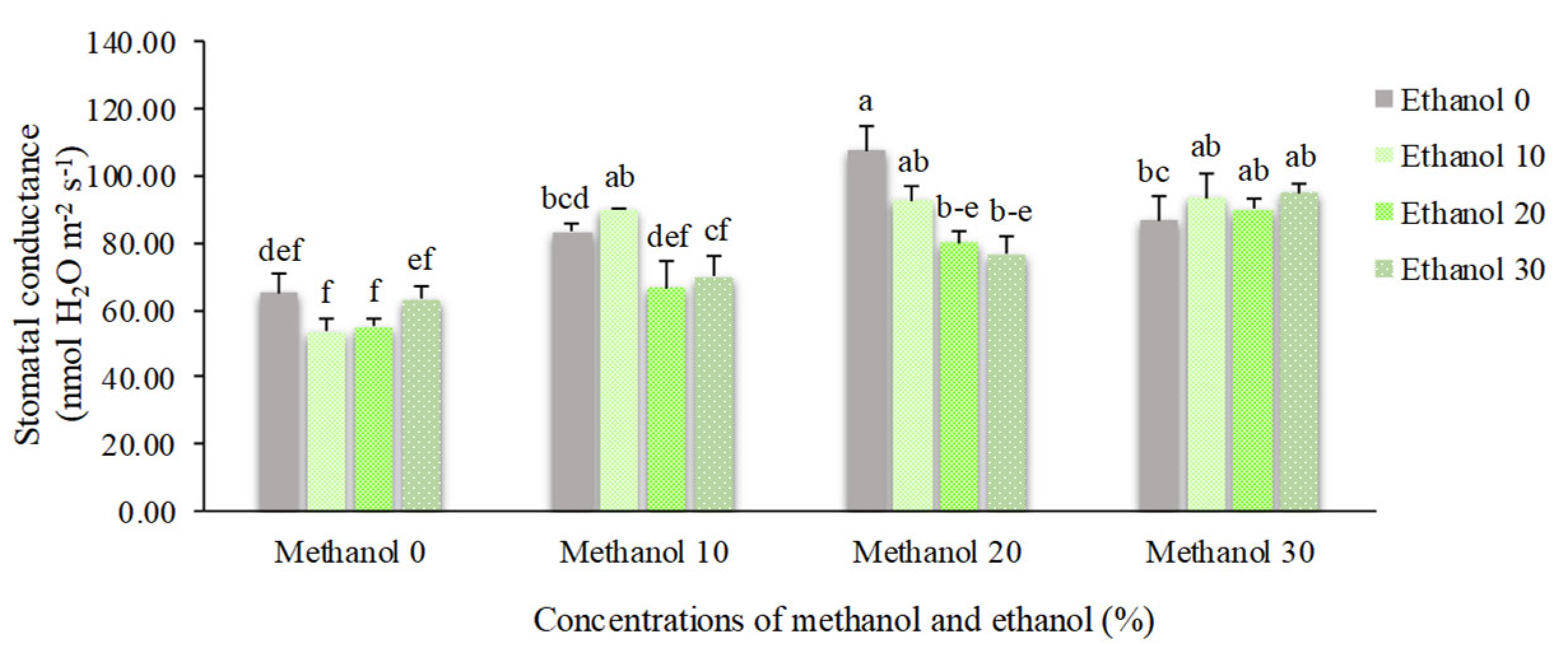
Figure 7.
The effect of methanol and ethanol spraying on the mesophyll conductance (gm) of freesia leaves.
Figure 7.
The effect of methanol and ethanol spraying on the mesophyll conductance (gm) of freesia leaves.
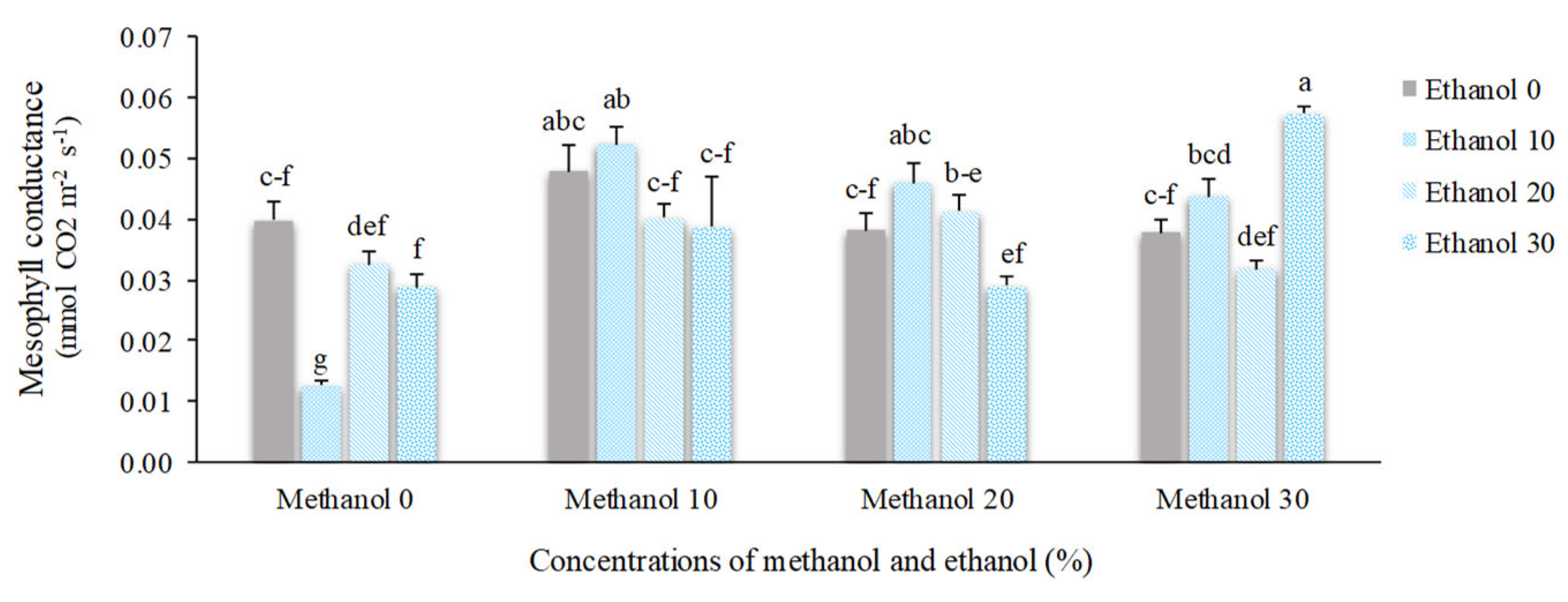
Figure 8.
The effect of methanol and ethanol spraying on the daughter corm weight of freesia plants.
Figure 8.
The effect of methanol and ethanol spraying on the daughter corm weight of freesia plants.
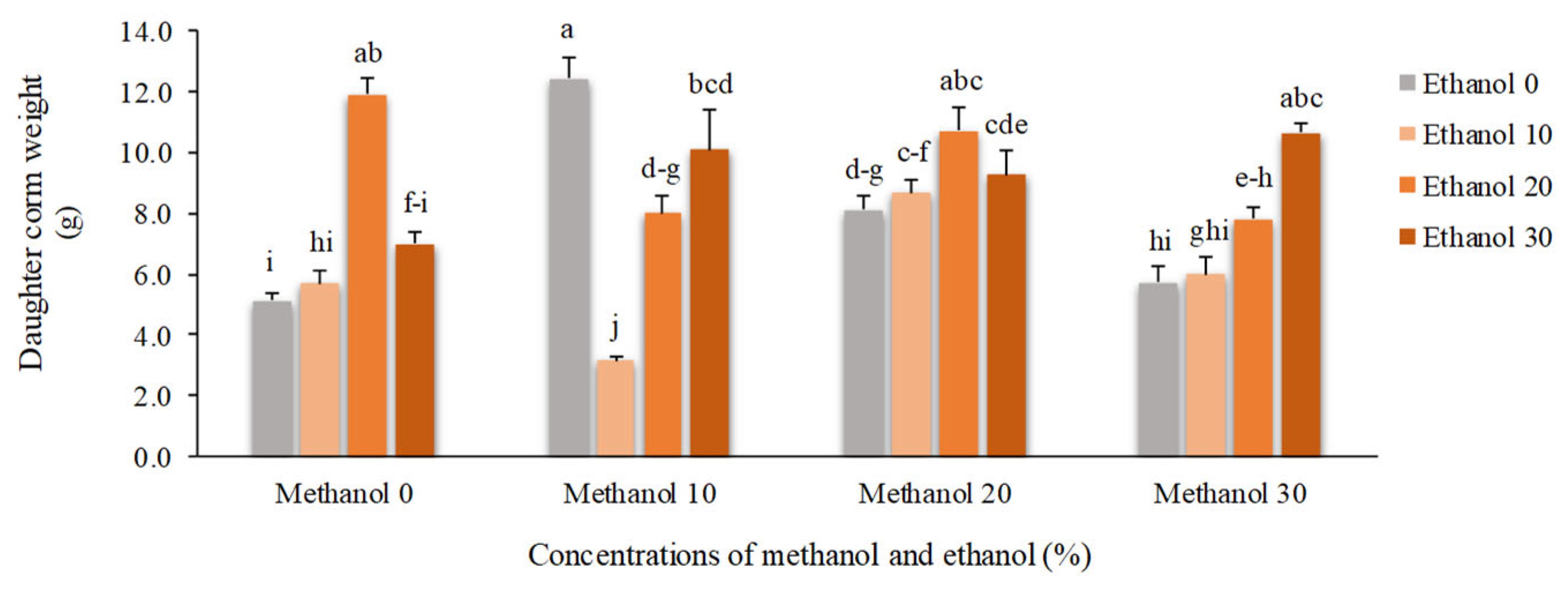
Figure 9.
The effect of methanol and ethanol spraying on the number of cormlets produced by freesia plants.
Figure 9.
The effect of methanol and ethanol spraying on the number of cormlets produced by freesia plants.
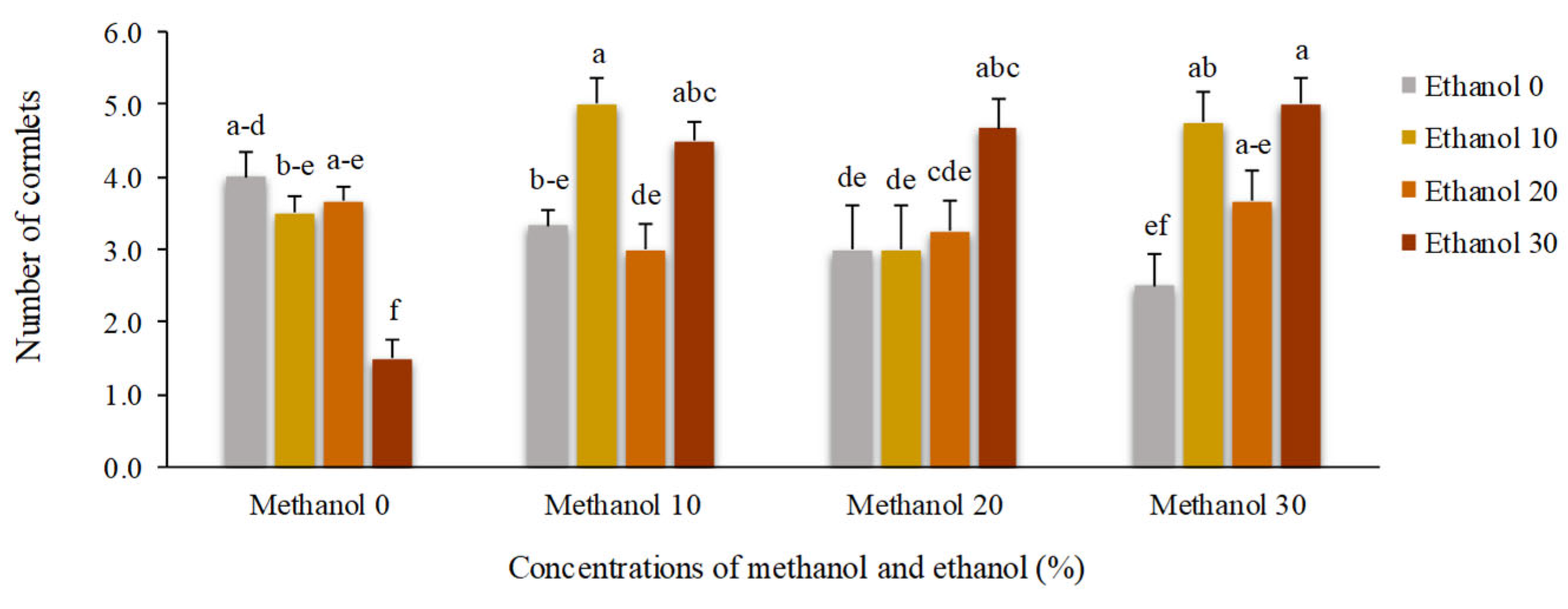
Figure 10.
The effect of methanol and ethanol spraying on the largest cormlet weight produced by freesia plants.
Figure 10.
The effect of methanol and ethanol spraying on the largest cormlet weight produced by freesia plants.
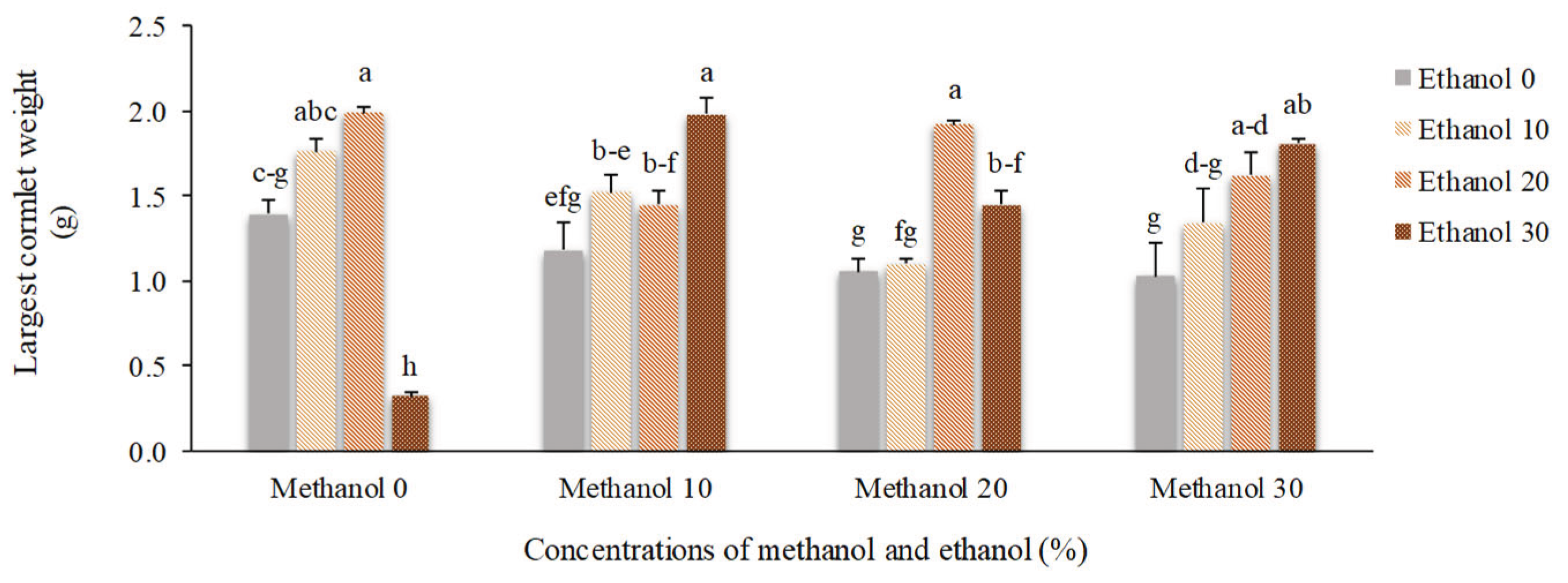
Figure 11.
The effect of methanol and ethanol spraying on the mean weight of all cormlets produced by freesia plants.
Figure 11.
The effect of methanol and ethanol spraying on the mean weight of all cormlets produced by freesia plants.
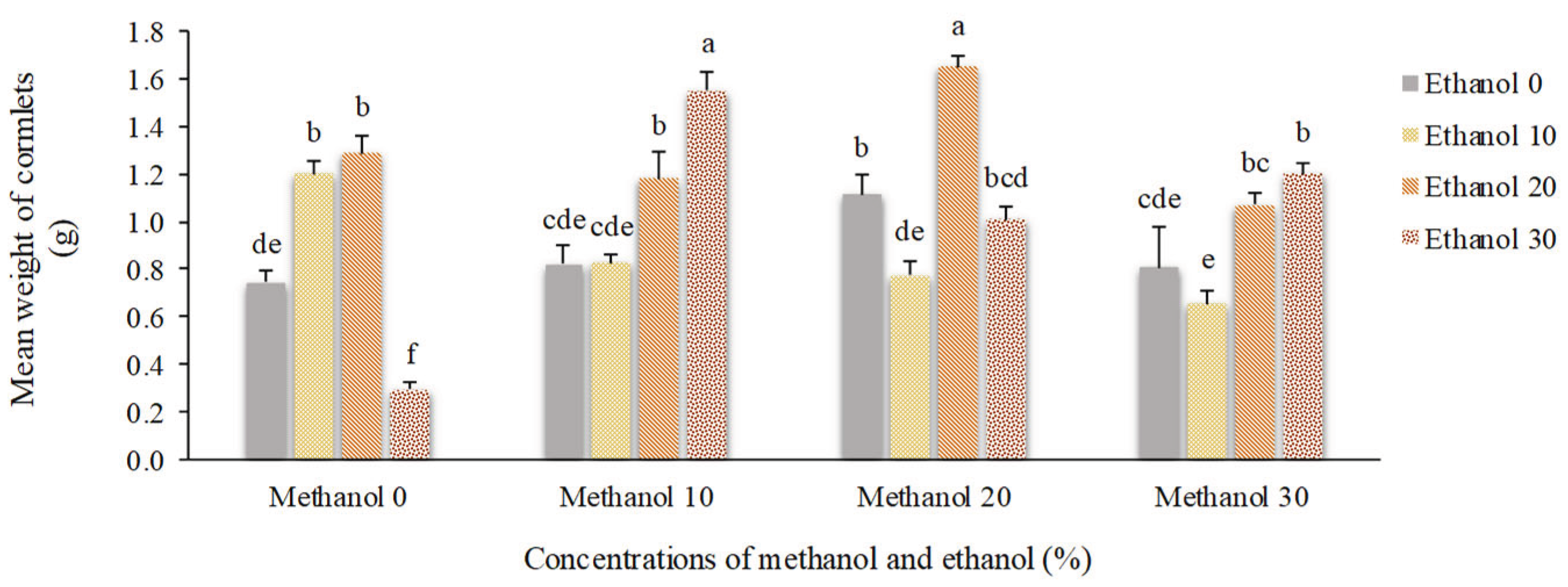
Figure 12.
The effect of methanol and ethanol spraying on the number of florets of freesia plants.
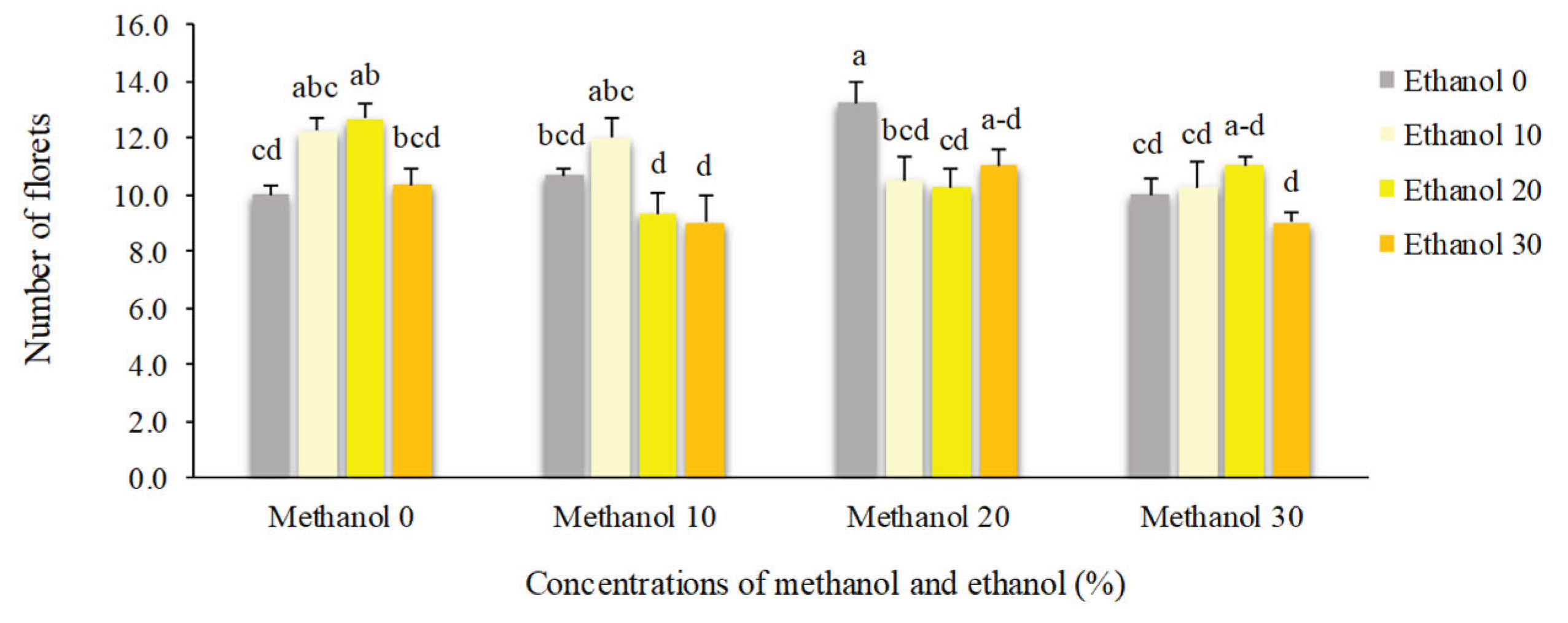
Figure 13.
The effect of methanol and ethanol spraying on the floret dry weight of freesia plants.
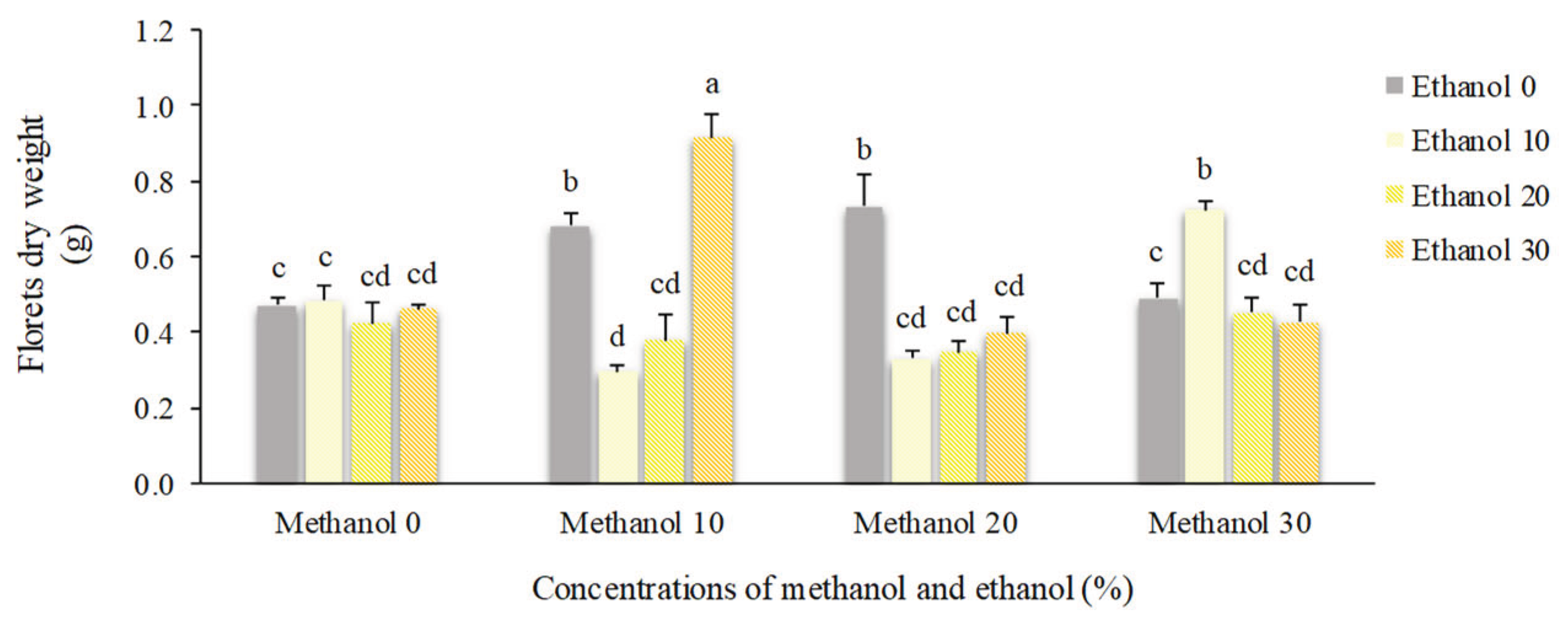
Figure 14.
The effect of methanol and ethanol spraying on the floret scape dry weight of freesia plants.
Figure 14.
The effect of methanol and ethanol spraying on the floret scape dry weight of freesia plants.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



