
Submitted:
02 July 2025
Posted:
02 July 2025
You are already at the latest version
Abstract
A new species of symbiotic copepod, Anchimolgus jejuicus n. sp., is described based on specimens collected from the external washings of the scleractinian coral Alveopora japonica Eguchi, 1965 in Jeju Island, Korea. The new species closely resembles A. multidentatus Kim, 2003 associated with Alveopora catalai Wells, 1968 from New Caledonia in having similar body lengths and similar shapes of the antennae, mouth organs, and swimming legs. However, the new species can be distinguished from the New Caledonian species by the shorter female caudal ramus, which is 1.70 times longer than wide, by the shorter third endopodal segment of the female antenna, which is 3.35 times longer than wide, by the minimized second spine of the distal lash of the maxilla, which is less than half as long as the first spine, by the relatively shorter inner distal spine of the second endopodal segment of female leg 4, which is 1.56 times longer than the outer spine, by the shorter exopodal segment of female leg 5, which is 3.39 times longer than wide, and by the exopod of male leg 5 armed with 1 spine and 1 seta. The present paper reports the northernmost distribution area of the genus Anchimolgus, near the latitude of 33°24'N in Jeju Island. In addition, the recent rapid increase in the population of the host coral Alveopora japonica around Jeju Island—likely linked to climate change—has prompted a discussion on the ecological implications for both the scleractinian coral and its newly discovered symbiotic copepod species.
Keywords:
Copepod symbiont
; taxonomy
; external association
; scleractinian coral
; Jeju Island
1. Introduction
Copepods are small aquatic crustaceans and are one of the most numerous metazoan groups in aquatic communities. Copepods may be free-living, symbiotic, or internal or external parasites on almost every major metazoan phylum [1]. The copepod family Anchimolgidae Humes and Boxshall, 1996 currently comprises 32 genera, and all members of this family are associates of scleractinian corals [2, 3]. Anchimolgus, the largest genus of the family, consists of 37 known species [4] distributed in the tropical seas, including Indonesia, New Caledonia, the east coast of Australia, and Madagascar [5, 6, 7, 8].
The corals (Cnidaria) are major hosts of the symbiotic copepods [9]. In Korea, the scleractinian coral Alveopora japonica Eguchi, 1965 recently thrives in shallow subtidal hard bottoms around Jeju Island [10, 11, 13, 14]. While examining the washings of the coral A. japonica collected from the west coast of Jeju Island, we unexpectedly found a new species of the genus Anchimolgus, which is to be described herein.
2. Materials and Methods
In October 2012, there were extensive A. japonica beds in the sampling site at Biyangdo, located to the northwest of Jeju Island, South Korea, where the beds formed dense and almost monospecific carpets at a depth of 10–17 m. These stands were punctuated by the regionally endemic kelp Ecklonia cava. A recent observation shows that this scleractinian coral is now the dominant taxon at 15 m at this site, covering up to 67% of the benthic substrate on an estimated area of >1 ha [10]. In 14 June 2023, the samples of the coral host, Alveopora japonica, were collected by Drs. Taewon Jung and Jong Guk Kim of the Honam National Institute of Biological Resources using SCUBA diving at a depth of 20 m near Biyangdo, Jeju Island (33°24'19.3"N, 126°12'17.0"E) (Figure 1). The collected coral samples were fixed with 80% ethanol, and copepods dislodged from the corals were sorted out. Before microscopic observation, selected copepod specimens were immersed in lactic acid for several minutes. Dissections and microscopic observations were done using the reversed slide method of Humes and Gooding [15]. Drawings were made with a drawing apparatus equipped on the microscope. The lengths of the appendage segments of copepod specimens are the average of the longest and shortest margins. The terminology adopted in this paper mainly follows Huys and Boxshall [16] and Humes and Boxshall [2]. The dissected paratype of Anchimolgus multidentatus Kim, 2003 was carefully re-examined and measured for a detailed comparison with the new species. Type specimens have been deposited in the Honam National Institute of Biological Resources (NNIBR), Mokpo, Korea.
3. Taxonomic Description
Class Copepoda Milne Edwards, 1840
Order Cyclopoida Burmeister, 1834
Family Anchimolgidae Humes and Boxshall, 1996
Genus Anchimolgus Humes and Stock, 1972
(Korean name: Dol-san-ho-no-beol-le)
Anchimolgus jejuicus n. sp.
(Korean name: Je-ju-dol-san-ho-no-beol-le)
Type material. Holotype (intact ♀; HNIBR IV7751), intact paratypes (15 ♀♀, 10 ♂♂; HNIBR IV7752), and dissected paratypes (2 ♀♀, 1 ♂) from washings of the scleractinian coral Alveopora japonica Eguchi, 1965, Biyangdo in Jeju Island (33°24'19.3"N, 126°12'17.0"E), depth 20 m, collected by Tae Won Jung and Jong Guk Kim, 14 June 2023. Intact type specimens have been deposited in the Honam National Institute of Biological Resources (HNIBR), Mokpo, Korea. Dissected paratypes are retained in the collection of I.-H. Kim.
Etymology. The name is derived from “Jeju Island”, where the type locality is located.
Female. Body (Figure 2A) rather stout. Mean body length 1.24 mm (1.16-1.36 mm), based on 10 specimens. Body length of figured and described specimen 1.33 mm. Prosome 867×594 μm. All prosomal somites with rounded lateral corners. Cephalothorax as long as wide, with dorsal suture line between cephalosome and first pedigerous somite. Urosome (Figure 2B) 5-segmented. Fifth pedigerous somite 180 μm wide, with several minute spinule at lateral apex. Genital double-somite 1.23 times longer than wide (193×157 μm), widest at anterior third; Genital apertures positioned laterally at about 60% region of double-somite length. Three free abdominal somites 45×91, 36×84, and 73×96 μm, respectively. Anal somite with minute spinules along posterior margin (Figure 2C). Caudal ramus (Figure 2C) 1.70 times longer than wide (68×40 μm), slightly shorter than anal somite, armed with 6 setae and ornamented with minute spinules on posteroventral margin; setae II and VII naked, other 4 setae pinnate. Egg sac 782×364 μm; each egg 150 μm in diameter.
Rostrum (Figure 2E) with indistinctly rounded apex. Antennule (Figure 2F) 444 μm long, 7-segmented; armature formula 4, 13, 6, 3, 4+aesthetasc, 2+aesthetasc, and 7+aesthetasc; third and last segments shortest; all setae naked; aesthetascs thin, setiform. Antenna (Figure 2G) 4-segmented, consisting of coxobasis and 3-segmented endopod; armature formula 1, 1, 3, 1+claw; setae small, rudimentary; third endopodal segment 3.35 times longer than wide (77×23 μm); terminal claw arched, 64 μm long, shorter than third endopodal segment.
Labrum (Figure 2H) broad, with broad posterior lobes; each lobe bearing broad membranous flange along its inner margin. Mandible (Figure 2I) with distinct inner notch; inner margin slightly bilobed, with thin spinules on proximal lobe and thicker spinules on distal lobe; outer side of gnathobase strongly protruded, bearing marginal row of minute spinules and row of about 7 small digitiform processes near margin of protruded region; distal lash elongated, spinulose along both margins. Maxillule (Figure 2J) lobate, bearing 1 inner and 3 distal setae; shortest innermost and outermost spinulose; middle one of 3 distal setae longest, naked. Maxilla (Figure 2K) 2-segmented; syncoxa (first segment) broad, unarmed; basis (distal segment) bearing elongated distal lash and 2 setae; distal lash with about 20 spines along its distal (outer) margin, first spine distinctly longer than other spines, second spine less than half as long as first spine, much smaller than other proximal spines; seta I (inner seta) short, broad, strongly tapering, with several spinules along margins; seta II curved, naked. Maxilliped (Figure 3A) 3-segmented, consisting of syncoxa, basis, and endopod; syncoxa unarmed; basis with 2 very unequal setae, distal one 3 times longer than proximal one; basis bearing 1 small spine, 1 seta, 2 subdistal rows of 3 to 5 small spinules, and terminated in slender, spiniform process.
Legs 1 (Figure 3B), 2 (Figure 3C), and 3 with 3-segmented rami. Leg 2 similar to leg 2, except for bearing 3 spines and 2 setae on third endopodal segment (Figure 3D). Leg 4 (Figure 3E) with 3-segmented exopod and 2-segmented endopod. Inner coxal seta well-developed in legs 1–3, but vestigial in leg 4. Outer seta of basis small, naked in legs 1–3, but longer and pinnate in leg 4. All spines on legs 1–4 with serrate margins. Leg 4 endopod with second segment of 51×29 μm; its 2 distal spines 32 (outer) and 50 μm (inner), respectively. Armature formula for legs 1–4 as follows:
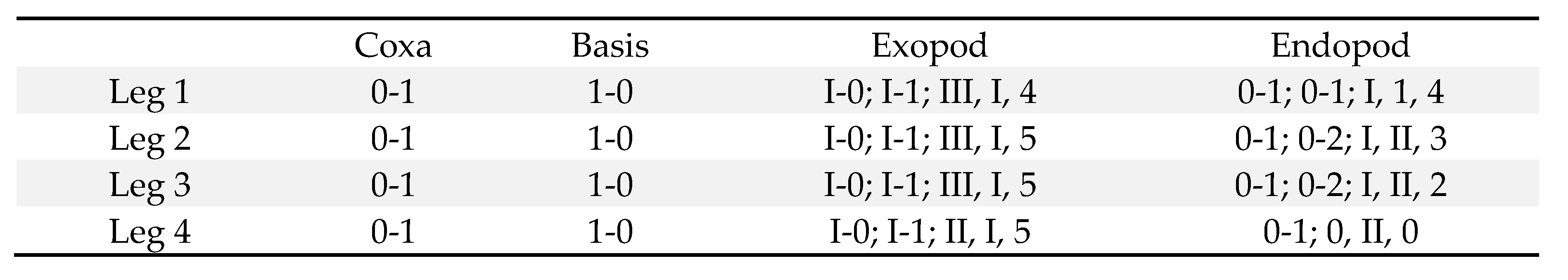
Leg 5 (Figure 2B) consisting of dorsolateral seta and free exopod; exopodal segment (Figure 3F) 3.39 times longer than wide (95×28 μm; width measured at proximal expanded region), 23 μm wide in midway, bearing prominent inner proximal inflation, distally 1 small serrate lamella and 2 naked setae (outer one 87 μm long and inner one 64 μm long), and ornamented with several minute spinules on outer margin. Leg 6 (Figure 3G) represented by 2 small setae (1 blunt and 1 attenuated) and 1 toothlike process on genital operculum.
Male. Body (Figure 4A) similar to that of female. Body length of figured and described specimen 1.02 mm. Prosome 630×395 μm. Cephalothorax 1.28 times longer than wide, with obscure dorsal suture line. Urosome (Figure 4B) 6-segmented. Fifth pedigerous somite 124 μm wide. Genital somite roughly quadrangular 179×180 μm, with rounded corners and well-developed genital operculum. Four abdominal somites 27×62, 31×64, 25×64, and 42×71 μm, respectively. Caudal ramus 1.69 times longer than wide (54×32 μm), armed as in female.
Rostrum as in female. Antennule with 3 additional aesthetascs, 2 on second segment and 1 on fourth at places indicated by dark dots in Figure 1F. Antenna (Figure 4C) armed as in female, but with additional spinules on inner margin of first and second endopodal segments; third endopodal segment 3.95 times longer than wide (75×19 μm).
Labrum, mandible, maxillule, and maxilla as in female. Maxilliped (Figure 4D) consisting of syncoxa, basis, endopod, and terminal claw; syncoxa and small endopod unarmed; basis with 2 naked, unequal setae, 1 row of spinules along inner margin, 2 small subdistal teeth; terminal claw arched, elongated, as long as remaining part of maxilliped, with 2 very unequal setae proximally, fringed with membrane along concave margin.
Leg 1 (Figure 4E) with 2 spines and 4 setae (formula I, I, 4) on third endopodal segment. Legs 2–4 as in female. Exopodal segment of leg 5 (Figure 4F) 2.53 times longer than wide (38×15 μm), lacking proximal expansion, bearing spinules on outer surface, armed distally with 1 seta (45 μm long) and 1 spinulose spine (15 μm long). Leg 6 (Figure 4B) represented by 2 thin setae on genital operculum.
4. Discussion
4.1. Species Characterization
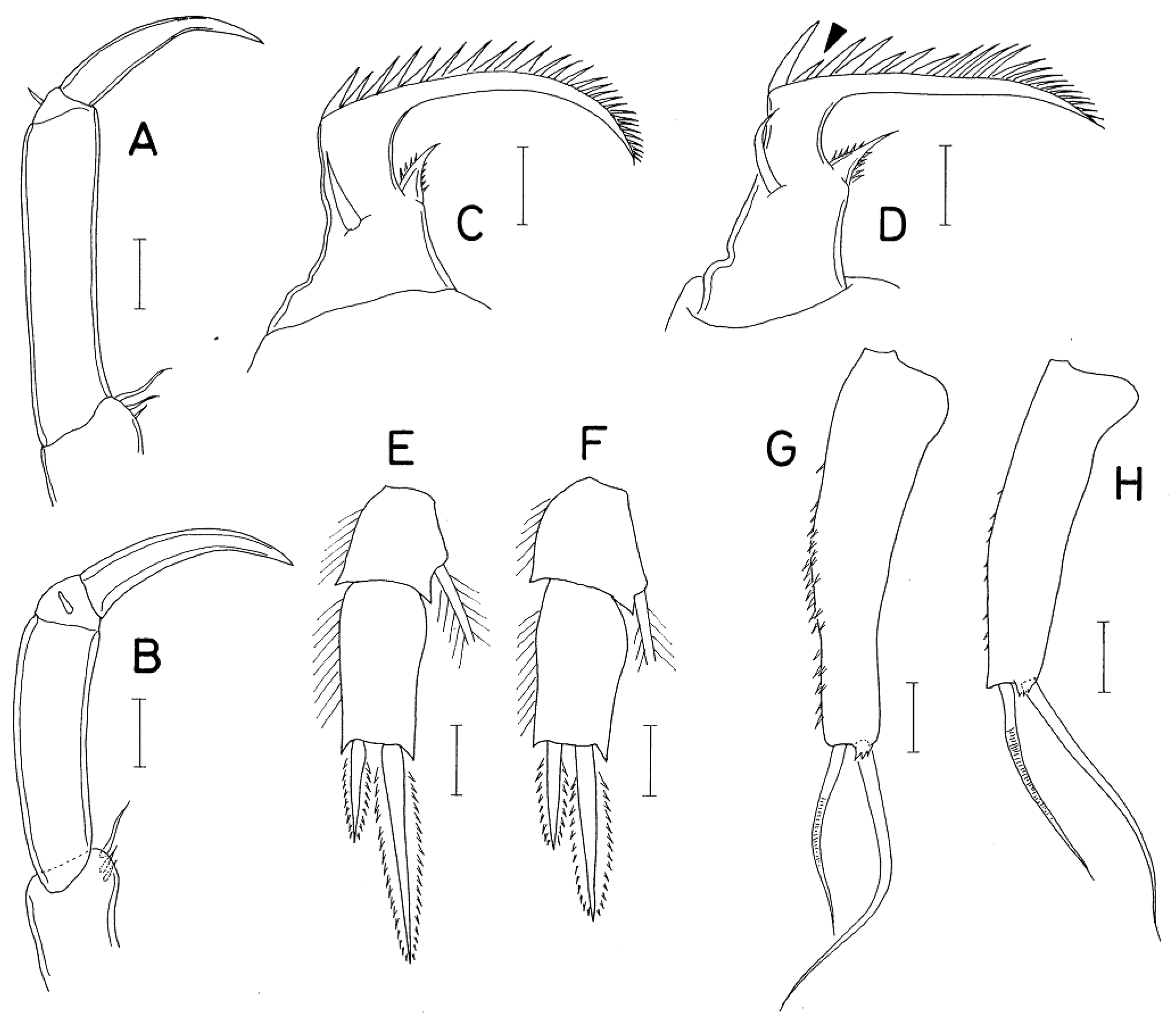
Species of the genus Anchimolgus can be divided into two groups: the digitatus-group and the contractus-group. In the digitatus-group, which consists of 25 known species, including the type species, A. digitatus (Humes and Ho, 1968), the third endopodal segment (terminal segment) of the antenna is shorter than or as long as the second endopodal segment (third segment). In the contractus-group, which consists of 12 known species, including A. contractus Humes, 1979, the third endopodal segment of the antenna is longer than the second segment. Anchimolgus jejuicus n. sp. belongs to the contractus-group, as the third endopodal segment of its antenna is much longer than the second endopodal segment. Within the contractus-group, the new species most closely resembles A. multidentatus Kim, 2003, which was described as an associate of Alveopora catalai Wells, 1969, from New Caledonia [6], as both species have similar body lengths and similar shapes of the antennae, mouth organs, and swimming legs. Moreover, since the two copepod species are the only species parasitizing coral hosts of the same genus, it was considered highly likely that they were the same species.
A careful comparison of specimens of A. jejuicus n. sp. and the original description and preserved specimen of A. multidentatus has led to the conclusion that they are not the same species. The morphological differences between the two species lies in the details, as follows: (1) the female caudal ramus of A. jejuicus n. sp. is 1.70 times longer than wide (68×40 μm), while it is 2.02 times longer than wide (85×42 μm) in A. multidentatus; (2) the third endopodal segment of the antenna of A. jejuicus n. sp. is 3.35 times longer than wide (77×23 μm) and 1.20 times longer than the terminal claw, while it is 3.96 times longer than wide (95×24 μm in measured dissected paratype) and 1.48 times longer than the terminal claw in A. multidentatus (compare Figure 4A and B); (3) the second spine (indicated by the arrowhead in Figure 5D) of the distal lash of the maxilla of A. jejuicus n. sp. is distinctly minimized, while it is only slightly shorter than the first spine in A. multidentatus (compare Figure 5C and D); (4) the inner distal spine (50 μm long) of the second endopodal segment of female leg 4 of A. jejuicus n. sp. is 1.56 times longer than the outer spine (32 μm long), while the inner spine (64 μm long) is 2.13 times longer than the outer spine (30 μm long) in A. multidentatus (compare Figure 5E and F); (5) the exopodal segment of female leg 5 of A. jejuicus n. sp. is 3.39 times longer than wide (95×28 μm), while it is more elongated, 3.83 times longer than wide (115×30 μm), and abruptly narrowed along the distal quarter, with more and larger spinules on the outer surface in A. multidentatus (compare Figure 5G and H); and (6) the exopod of male leg 5 of A. jejuicus n. sp. is armed with 1 spine and 1 spine, while it is armed with 2 setae in A. multidentatus.
4.2. Ecological Implications of the Discovery of the Symbiotic Copepod Anchimolgus jejuicus n. sp. on the Scleractinian Coral Alveopora japonica from Jeju Island
Seven zooxanthellate scleractinian coral species have been documented around Jeju Island, Korea [17]. Among them, Alveopora japonica Eguchi, 1968 is endemic to the Asia-Pacific region, occurring in subtropical to temperate waters of the northwestern Pacific, including Taiwan, Japan, and Jeju Island [18, 19]. Recent molecular analyses of A. japonica, using ITS and 23S rDNA sequences of its symbiotic dinoflagellates (family Symbiodiniaceae), suggest the presence of three cryptic species in the Northwest Pacific, with Taiwanese populations forming a distinct clade [20]. According to 75 georeferenced records from the Global Biodiversity Information Facility (GBIF), the species’ latitudinal range extends from 30°24'N in Tanegashima (Kagoshima Prefecture) to 36°06'N in Shimane Prefecture, Japan, and includes Jeju Island, Korea. This distribution appears to correlate with sea surface temperatures between 20°C and 24°C, as inferred from time-averaged SST data in the surrounding regional seas over the past two decades (August 2003–August 2023) (Figure 1).
Since its first report in Jeju Island in 1973 [21], A. japonica has shown significant population growth over the past three decades, likely driven by climate change and anthropogenic influences [10, 22, 11, 23, 14, 30]. Colonies are submassive to hemispherical, typically 2–3 cm in diameter, with occasional specimens exceeding 10 cm. Polyps, which extend during the day, bear 12 tentacles and exhibit pale brown to green coloration with white or cream tips [17, 20]. Observations in Tateyama Bay, Japan, indicate seasonal recruitment from August to November, potentially linked to planulation [24]. Colony density is positively associated with rising sea temperatures but negatively affected by sand substrates, which are themselves correlated with typhoon activity, suggesting that typhoons influence short-term population dynamics.
The high density of A. japonica colonies in the shallow rocky subtidal zone, particularly among stands of the endemic kelp Ecklonia cava Kjellman, 1885 in Jeju Island [10], has created a structurally distinct microhabitat resembling a “submarine tropical island”. While initially reducing local biodiversity during successional stages, this habitat has become a biodiversity hotspot. For instance, a recent survey reported 34 epibenthic mollusk species—20 bivalves and 14 gastropods—associated with A. japonica colonies in 1 m2 plots, many with tropical-subtropical affinities [13]. The discovery of a new symbiotic copepod species in this context is thus ecologically significant, particularly as A. japonica expands its range northward.
Coral-associated copepods are essential components of reef cryptofauna, with many species displaying host specificity [25, 26]. Globally, 148 scleractinian coral species host 363 copepod symbionts, including 141 species in the family Anchimolgidae [27]. This family encompasses 32 genera [2], all associated with scleractinian corals [3]. Anchimolgus, the largest genus in the family, contains 37 described species [4], which are distributed across tropical Indo-Pacific regions, including Indonesia, Thailand, New Caledonia, eastern Australia, and Madagascar [5, 6, 7, 8, 28]. Before this study, the northernmost record of Anchimolgus was Dongsha Atoll (20°40'N) in the South China Sea [29]. The discovery of A. jejuicus in Jeju Island (33°24'N) thus significantly extends the known geographic range of the genus northward.
Author Contributions
Conceptualization, Il-Hoi Kim and Jae-Sang Hong; Data curation, Il-Hoi Kim and Jae-Sang Hong; Investigation, Il-Hoi Kim and Jae-Sang Hong; Writing – original draft, Il-Hoi Kim; Writing – review & editing, Jae-Sang Hong.
Funding
This research received no funding.
Acknowledgments
We thank Drs. Taewon Jung and Jong-Guk Kim, Honam National Institute of Biological Resources (HNIBR), Mokpo, for providing us with the specimens of the new copepod species they collected. We also acknowledge the MODIS mission scientists and associated NASA personnel for producing the SST data used in this research effort. We are also indebted to Professor Chaolun Allen Chen for providing information on the distribution of Alveopora japonica in Taiwanese waters.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Walter, T.C.; Boxshall, G.A. World of Copepods Database 2025. Accessed at https://www.marinespecies.org/copepoda on 2025-04-16.
- Humes, A.G.; Boxshall, G.A. A revision of the lichomolgoid complex (Copepoda: Poecilostomatoida), with the recognition of six new families. J. Nat. Hist. 1996, 30, 175–227. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Halsey, A.H. An Introduction to Copepod Diversity. The Ray Society, London, 2004, 966 pp.
- WoRMS Editorial Board. World Register of Marine Species, 2025. Available from: http://www.marinespecies.org (accessed 14 April 2025).
- Humes, A.G. Cyclopoid copepods (Lichomolgidae) from fungiid corals in New Caledonia. Zool. Anz. 1973, 190, 312–333. [Google Scholar]
- Kim, I.-H. Copepods (Crustacea) associated with marine invertebrates from New Caledonia. Korean J. Syst. Zool. 2003, Special Issue 4. 1–167.
- Kim, I.-H. Copepods (Crustacea) associated with marine invertebrates from the Moluccas. Korean J. Syst. Zool. 2007, Special Issue 6, 1–126. [Google Scholar]
- Kim, I.-H. Siphonostomatoid Copepoda (Crustacea) associated with invertebrates from tropical waters. Korean J. Syst. Zool. 2010, Special Issue 8, 1–176. [Google Scholar]
- Humes, A.G. How many copepods? Hydrobiologia 1994, 292/293, 1–7. [Google Scholar] [CrossRef]
- Denis, V.; Chen, C.A.; Song, J.I.; Woo, S. Alveopora japonica beds thriving under kelp. Coral Reefs 2013, 32, 503. [Google Scholar] [CrossRef]
- Vieira, C.; Keshavmurthy, S.; Ju, S.-J.; Hyeong, K.; Seo, I.; Kang, C.-K.; Hong, H.-K.; Chen, C.A; Choi, K.S. Population dynamics of a high-latitude coral Alveopora japonica Eguchi from Jeju Island, off the southern coast of Korea. Mar. Freshwater Res. 2016, 67, 594–604. [Google Scholar] [CrossRef]
- Lee, K.-T.; Lee, H.-M.; Subramaniam,T. ; Yang, H.-S.; Patk, S.R.; Kang, C.-K.; Keshavmurthy, S.; Choi, K.-S. Dominance of the scleractinian coral Alveopora japonica in the barren subtidal hard bottom of high-latitude Jeju Island off the south coast of Korea assessed by high-resolution underwater images. PLoS One 2022, 17, e0275244. [Google Scholar] [CrossRef]
- Noseworthy, R.G.; Hong, H.-K.; Ju, S.-J.; Yang, H.-S.; Choi, K.-S. Mollusk species associated with the scleractinian coral Alveopora japonica Eguchi, 1968 forming a coral carpet in northwestern Jeju Island. Ocean Polar Res. 2022, 44, 331–338. [Google Scholar]
- Shin, S.; Ribas-Deulofeu, L.; Subramaniam, T.; Lee, K.T.; Kang, C-K. ; Denis, V.; Choi, K.-S. The vertical distribution of Alveopora japonica provides insight into the characteristics and factors controlling population expansion at Jeju Island off the south coast of Korea. Mar. Biodiversity 2024, 54, 20. [Google Scholar] [CrossRef]
- Humes, A.G.; Gooding, R.U. A method for studying the external anatomy of copepods, Crustaceana 1964, 6, 238–240. [CrossRef]
- Huys, R.; Boxshall, G.A. Copepod evolution. Ray Society, London, 1991, 1-468. ISBN 0-903-87421-0.
- Sugihara, K.; Yamano, H. ; Choi, K-S.; Hyeong, K. Zooxanthellate scleractinian corals of Jeju Island, Republic of Korea. In: Nakano SI, Yahara T, Nakashizuka T (eds) Integrative observations and assessments. Ecological Research Monographs, Springer, Tokyo, 2014; pp. 111–130. [Google Scholar]
- Veron, J.E.N. Corals of the World. Vols. 1–3. Aust. Inst. Mar. Sci. 2000, 1410 pp.
- Dai, C.F.; Horng, S. Scleractinia fauna of Taiwan. I. The complex group. National Taiwan University, Taipei 2009, 1–172.
- Kang. J.H.; Jang, J.E.; Kim, J.H.; Kim, S.; Keshavmurthy, S.; Agostini, S.; Rimer, J.D.; Chen, C.A.; Choi, K.-S.; Park, S.R.; Lee, H.J. The origin of the subtropical coral Alveopora japonica (Scleractinia: Acroporidae) in high-latitude environments. Front. Ecol. Evol. 2020, 8, 12. [Google Scholar] [CrossRef]
- Song, J.-I. A study on the classification of the Korean Anthozoa 7. Scleractinia (Hexacorallia). Korean J. Zool. 1982, 25, 131–148. [Google Scholar]
- Denis, V.; Ribas-Deulofeu, L.; Loubeyres, M.; De Palmas, S.; Hwang, S.-J.; Woo, S.; Song, J.-I.; Chen, C.A. Recruitment of the subtropical coral Alveopora japonica in the temperate waters of Jeju Island, South Korea. Bull. Mar. Sci. 2015, 91, 85–96. [Google Scholar] [CrossRef]
- Ribas-Deulofeu, L.; Loubeyres, M.; Denis, V.; de Palmas, S.; Hwang, S.-J.; Woo, S.; Song, J.-I.; Chen, C.A. Chen. Jeju Island: a sentinel for tracking ocean warming impacts on high-latitude benthic communities. Coral Reefs 2023, 42, 1097–1112. [Google Scholar] [CrossRef]
- Matsumoto, A. ; Hashimoto, S; Arakawa, H. Short-term population dynamics of high-latitude Alveopora japonica in Tateyama Bay, Japan. Galaxea, J. Coral Reef Stud 2015, 17, 33–39. [Google Scholar]
- Preston, N.P.; Doherty, P.J. Cross-shelf patterns in the community structure of coral-dwelling Crustacea in the central region of the Great Barrier Reef. II. Cryptofauna. Mar. Ecol. Prog. Ser. 1994, 104, 27–38. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; van der Meij. S.E.T.; Fransen, C.H.J.M. The mushroom coral as a habitat. J. Mar. Biol. Assoc. U. K. 2012, 92, 647–663. [Google Scholar] [CrossRef]
- Cheng, Y.R.; Mayfield, A.B.; Meng, P.-J; Dai, C.-F.; Huys, R. Copepods associated with scleractinian corals: a worldwide checklist and a case study of their impact on the reef-building coral Pocillopora damicornis (Linnaeus, 1758) (Pocilloporidae). Zootaxa 2016, 4174, 291–345. [Google Scholar] [CrossRef]
- Kim, I.-H.; Hong, J.-S. Copepods (Crustacea, Copepoda, Cyclopoida) associated with marine invertebrates from Thailand. Anim. Syst. Evol. Divers. 2014, 30, 274–318. [Google Scholar] [CrossRef]
- Cheng, Y.R. : Dai, C.F. Poecilostomatoid copepods associated with two species of widely distributed corals, Galaxea astreata (Lamarck, 1816) and Galaxea fascicularis (Linnaeus, 1767), in the South China Sea. Mar. Biodivers. 2016, 48, 1057–1072. [Google Scholar] [CrossRef]
- Kim, T.; Kim, T.; Yang, H.-S.; Choi, S K. ; Son, Y.B.; Kang, D.-H. Alveopora japonica conquering temperate reefs despite massive coral bleaching. Diversity, 2022, 14, 86. [Google Scholar] [CrossRef]
Figure 1.
Sampling locality of Anchimolgus jejuicus n. sp. on Jeju Island (★) and distribution map of its host coral Alveopora japonica in the surrounding seas of the Northwest Pacific (●) from the Global Biodiversity Information Facility (GBIF). https://www.gbif.org/species/2260435 based on 75 georeferenced records (accessed 12 June 2025). Additionally, the time averaged Sea Surface Temperature (See legend on the right, ℃) during the last two decades (August 2003–August 2023) in the regional seas was drawn using the satellite remote sensing data sets of the NASA Giovanni Data System.
Figure 1.
Sampling locality of Anchimolgus jejuicus n. sp. on Jeju Island (★) and distribution map of its host coral Alveopora japonica in the surrounding seas of the Northwest Pacific (●) from the Global Biodiversity Information Facility (GBIF). https://www.gbif.org/species/2260435 based on 75 georeferenced records (accessed 12 June 2025). Additionally, the time averaged Sea Surface Temperature (See legend on the right, ℃) during the last two decades (August 2003–August 2023) in the regional seas was drawn using the satellite remote sensing data sets of the NASA Giovanni Data System.
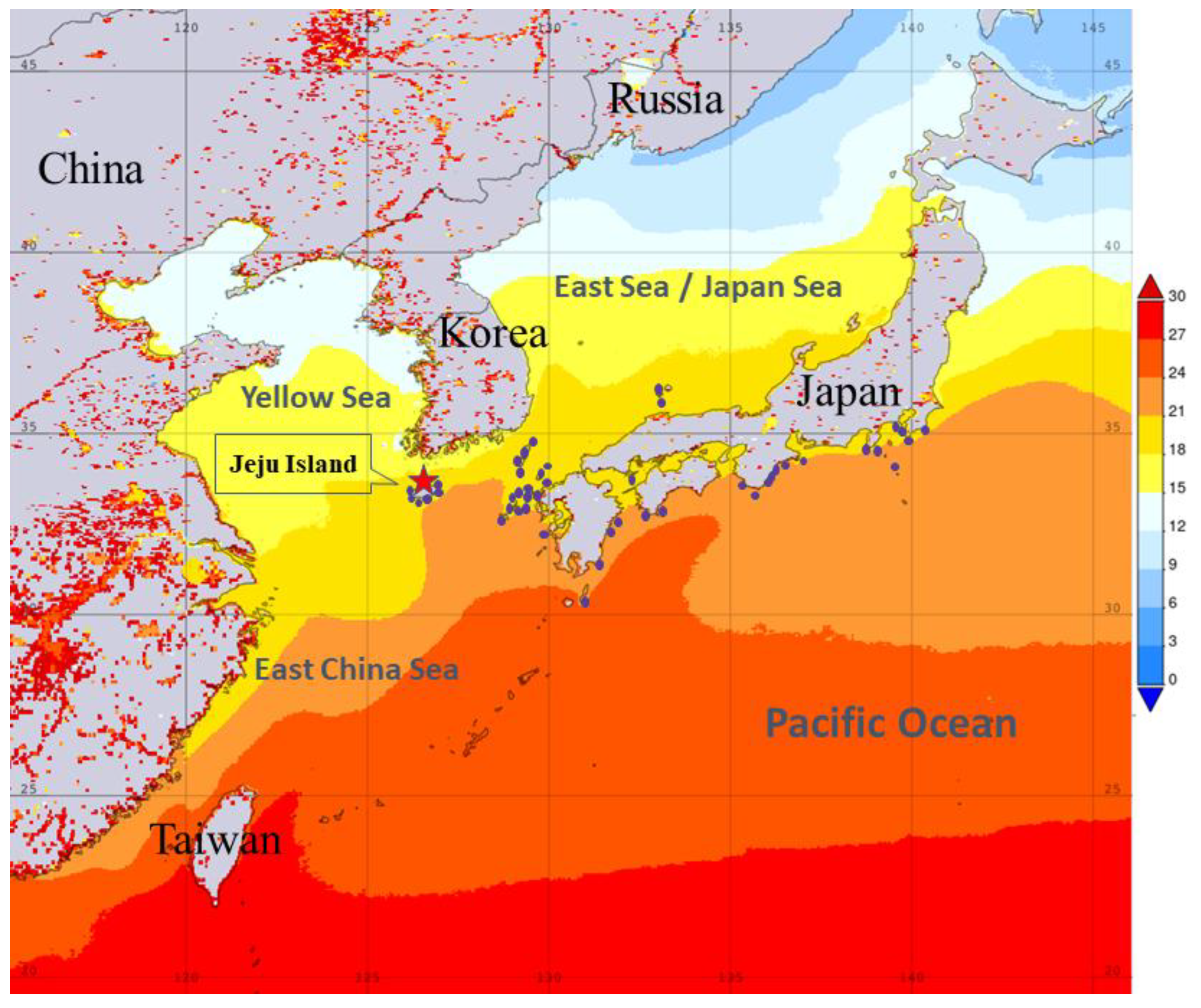
Figure 2.
Anchimolgus jejuicus n. sp., female. A, habitus, dorsal; B, urosome, dorsal; C, right caudal ramus, dorsal; D, egg sac; E, rostral area; F, antennule; G, antenna; H, labrum; I, mandible; J, maxillule; K, maxilla. Scale bars: A, D, 0.2 mm; B, E, 0.1 mm; C, H–K, 0.02 mm; F, G, 0.05 mm.
Figure 2.
Anchimolgus jejuicus n. sp., female. A, habitus, dorsal; B, urosome, dorsal; C, right caudal ramus, dorsal; D, egg sac; E, rostral area; F, antennule; G, antenna; H, labrum; I, mandible; J, maxillule; K, maxilla. Scale bars: A, D, 0.2 mm; B, E, 0.1 mm; C, H–K, 0.02 mm; F, G, 0.05 mm.
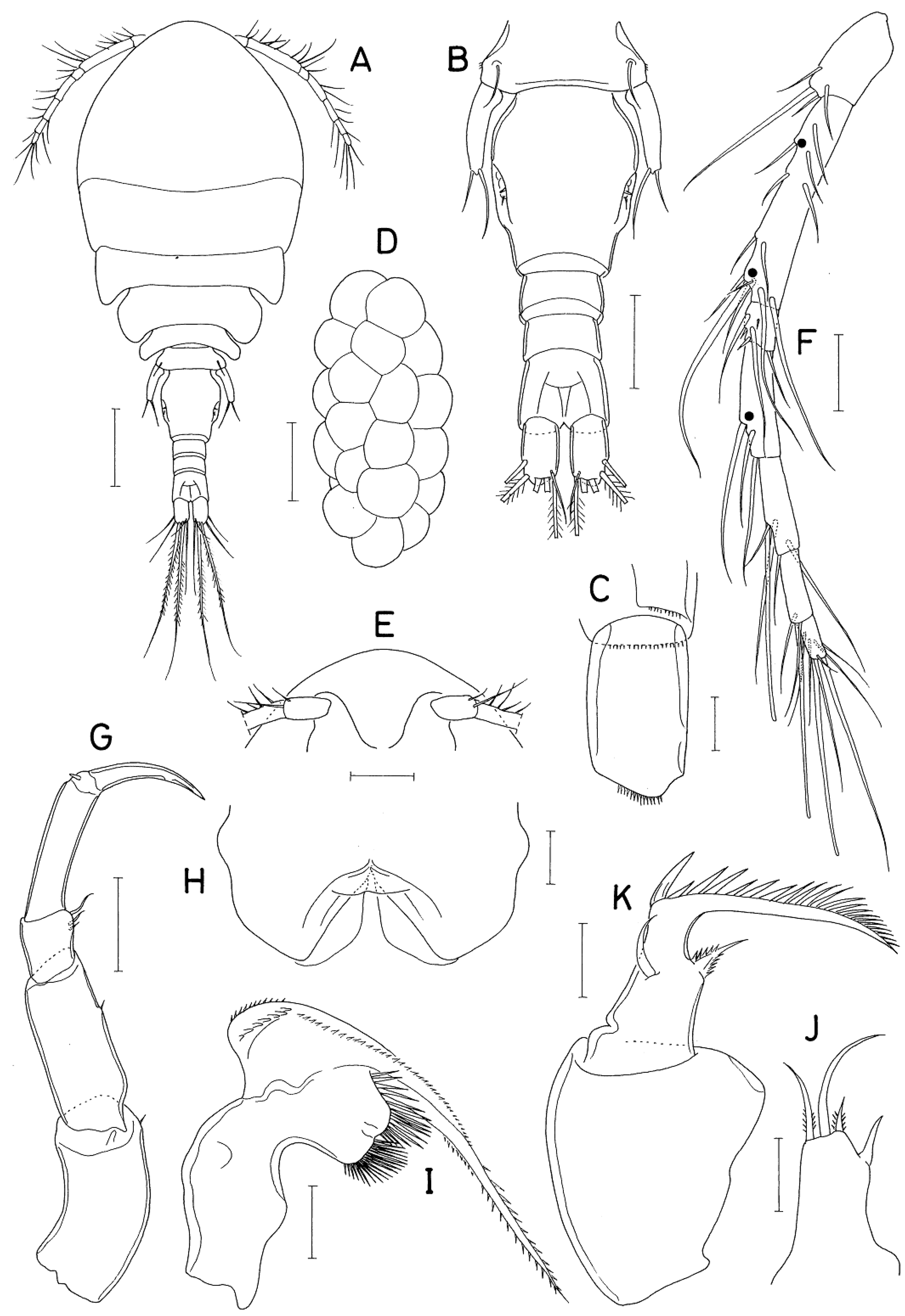
Figure 3.
Anchimolgus jejuicus n. sp., female. A, maxilliped; B, leg 1; C, leg 2; D, endopod of leg 3; E, leg 4; F, exopod of leg 5; G, left genital aperture, dorsal. Scale bars: A, G, 0.02 mm; B–F, 0.05 mm.
Figure 3.
Anchimolgus jejuicus n. sp., female. A, maxilliped; B, leg 1; C, leg 2; D, endopod of leg 3; E, leg 4; F, exopod of leg 5; G, left genital aperture, dorsal. Scale bars: A, G, 0.02 mm; B–F, 0.05 mm.
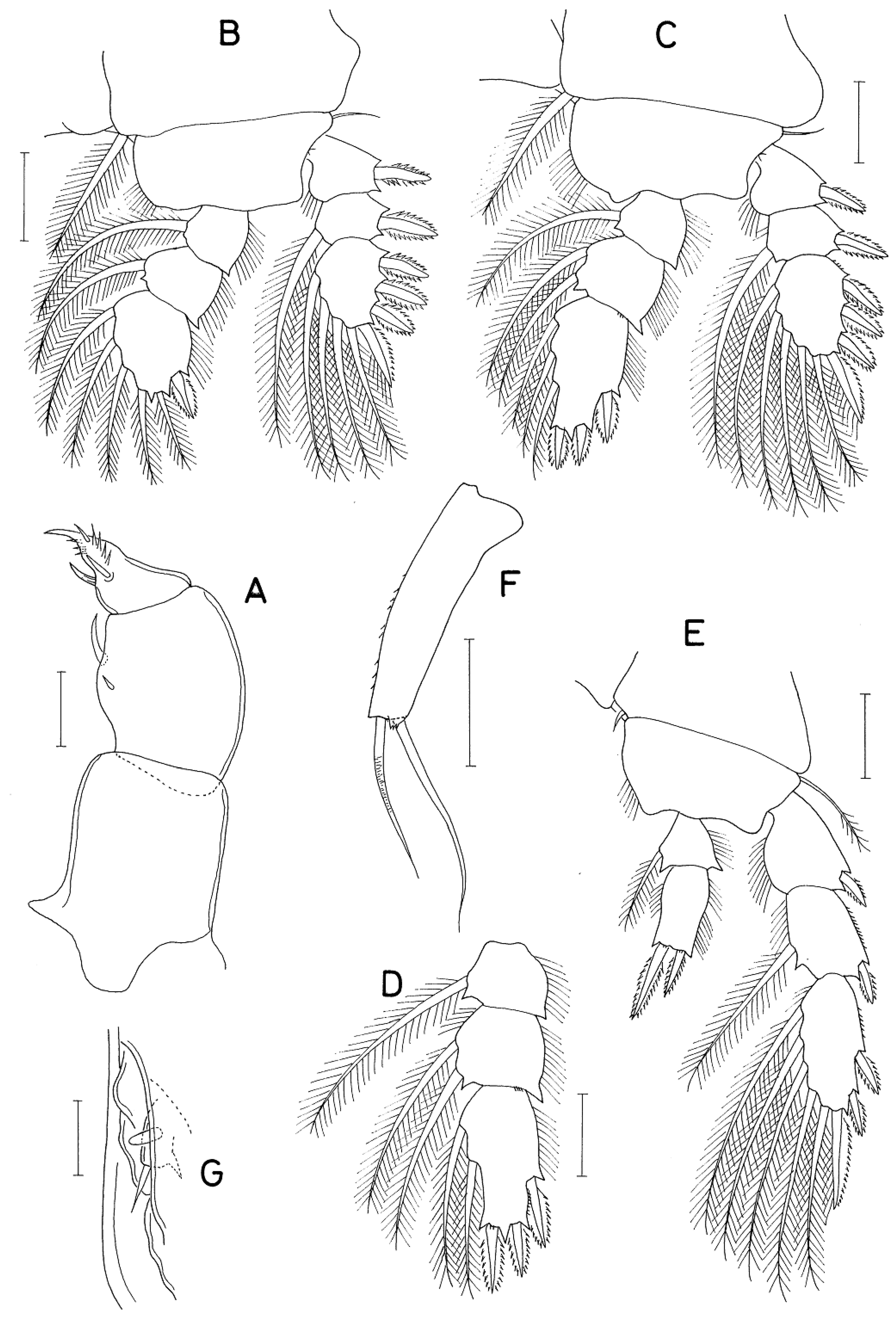
Figure 4.
Anchimolgus jejuicus n. sp., male. A, habitus, dorsal; B, urosome, ventral; C, antenna; D, maxilliped; E, leg 1; F, exopod of leg 5. Scale bars: A, 0.2 mm; B, 0.1 mm; C–E, 0.05 mm; F, 0.02 mm.
Figure 4.
Anchimolgus jejuicus n. sp., male. A, habitus, dorsal; B, urosome, ventral; C, antenna; D, maxilliped; E, leg 1; F, exopod of leg 5. Scale bars: A, 0.2 mm; B, 0.1 mm; C–E, 0.05 mm; F, 0.02 mm.
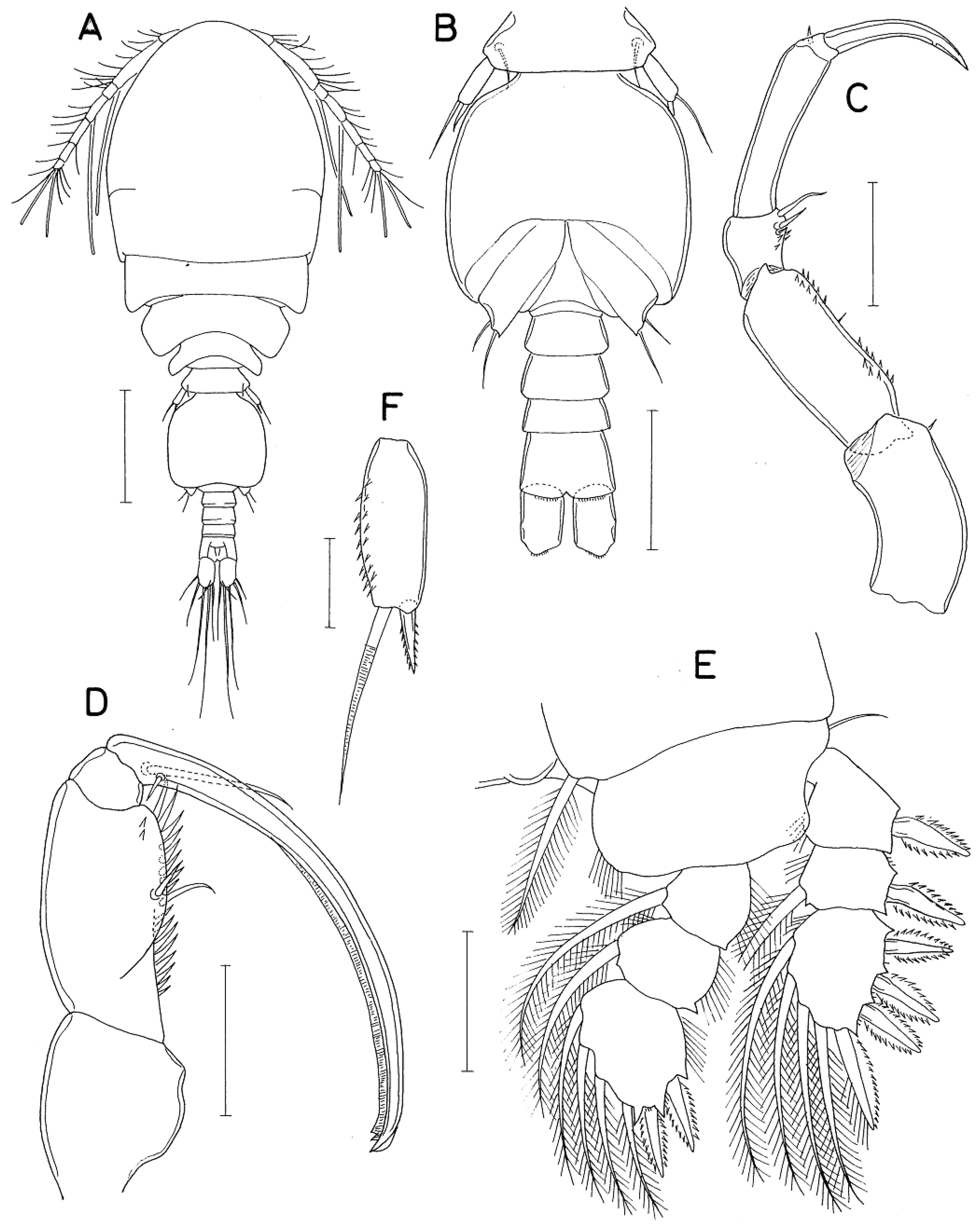
Figure 5.
Anchimolgus multidentatus Kim, 2003, female: A, distal part of antenna; C, basis of maxilla; E, endopod of leg 4; G, exopod of leg 5. Anchimolgus jejuicus n. sp., female: B, distal part of antenna; D, basis of maxilla; F, endopod of leg 4; H, exopod of leg 5. Scale bars: 0.02 mm.
Figure 5.
Anchimolgus multidentatus Kim, 2003, female: A, distal part of antenna; C, basis of maxilla; E, endopod of leg 4; G, exopod of leg 5. Anchimolgus jejuicus n. sp., female: B, distal part of antenna; D, basis of maxilla; F, endopod of leg 4; H, exopod of leg 5. Scale bars: 0.02 mm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



