
Submitted:
30 June 2025
Posted:
01 July 2025
You are already at the latest version
Abstract
Background/Objectives: Messenger RNA (mRNA) modifications regulate key steps in gene expression, including splicing, translation, and stability. Despite over 170 known RNA modifications, the subset occurring in mRNA remains understudied compared to tRNA and rRNA. This review aims to systematically evaluate 14 known naturally occurring mRNA-specific modifica-tions, rank them by publication frequency, and highlight emerging frontiers in epi-transcriptomics, including discovering new naturally occurring mRNA modifications, environmental RNA (eRNA) epitranscriptomics, and mirror-RNA generation and de-tection. Methods: We conducted a structured literature review of PubMed-indexed publications to rank mRNA modifications by citation prevalence. Key modifications such as m⁶A, m⁵C, Ψ, and m¹A were analyzed in terms of enzymatic machinery (“writers,” “erasers,” and “read-ers”), molecular functions, and physiological relevance. We also reviewed technological advances, with a focus on nanopore sequencing for detection of RNA modifications in native and environmental contexts. Results: m⁶A was identified as the most studied mRNA modification, followed by Ψ, m⁵C, and A-to-I editing (inosine). These modifications influence diverse mRNA processes, in-cluding translation efficiency, localization, and immune evasion. Cap-specific modifica-tions such as Cap0, Cap1, and Cap2 were also described, highlighting their role in transcript stability and innate immune regulation. Advances in nanopore sequencing have enabled direct detection of RNA modifications and offer promise for eRNA (envi-ronmental RNA) surveys. The potential for nanopore sequencing of many other of the 335 known RNA modifications in the MODOMICS database using existing nanopore tech-nologies and mirror-RNAs and mirror RNA modifications using mirror helicases is also discussed. Conclusions: mRNA modifications represent a critical, yet incompletely mapped, layer of gene reg-ulation. Continued research—especially using nanopore and machine learning tech-nologies—will help uncover their full biological significance. Exploration of eRNA and mirror-RNA modification space may redefine our understanding of RNA biology and origins of life.
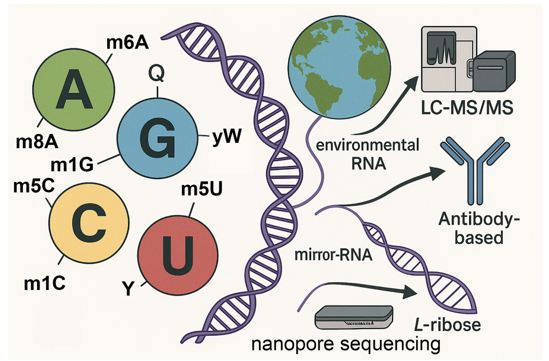
Keywords:
mRNA modification
; epitranscriptomics
; nanopore sequencing
; pseudouridine
; methyladenosine
; environmental RNA
; mirror RNA
; RNA caps
1. Introduction
The regulation of gene expression extends far beyond the DNA sequence, encompassing a complex landscape of chemical modifications that affect RNA molecules. Among these, modifications of messenger RNA (mRNA)—the direct templates for protein synthesis—represent a rapidly emerging frontier in molecular biology. These chemical marks, collectively referred to as the epitranscriptome, play vital roles in mRNA processing, stability, translation, and localization [1,2,3].
Over 150 types of RNA modifications have been cataloged to date, with the majority studied in transfer RNA (tRNA) and ribosomal RNA (rRNA) (see MODOMICS database, [4]). However, only a subset of these modifications occurs on mRNA, where they are less understood but potentially more dynamic and responsive to cellular context. Prominent among these is N6-methyladenosine (m⁶A), which has become a paradigm for reversible, regulatory RNA modifications [5], which can now be done at the single-cell level [6,7]. Other notable mRNA modifications include pseudouridine (Ψ or Y) [8,9], 5-methylcytidine (m⁵C) [8,10,11], N1-methyladenosine (m¹A) [12,13], inosine (I) [14,15], and various mRNA cap modifications such as m⁷G [16,17] and m⁶Am [18,19] at the 5′ end [20].
The functional consequences of these modifications are profound: they can influence transcript lifespan, translation efficiency, splice site selection, and cellular localization. Moreover, several modifications help mRNAs evade innate immune sensors such as RIG-I, highlighting their role in immune regulation and therapeutic mRNA design [21,22]. RIG-I (Retinoic acid-inducible gene I) is a crucial cytoplasmic RNA sensor that plays a vital role in the innate immune system’s response to viral infections. It functions as a pattern recognition receptor (PRR) that detects viral RNA and initiates a signaling cascade leading to the production of type I interferons, which are critical for antiviral defense. RIG-I is particularly important for recognizing RNA viruses, such as influenza and hepatitis C, by binding to specific viral RNA structures like 5’-triphosphorylated RNA [21,22].
Despite rapid advances, key controversies persist. For example, while m⁶A is known to be dynamically regulated, there is debate over how reversible and site-specific its deposition truly is [13,14]. Similarly, the functional roles of modifications such as m⁵C and Ψ in mRNA remain incompletely defined and sometimes contradictory depending on cell type and experimental model [15]. The contradictory results might be explained by mRNA modifications like m⁵C and Ψ being involved in a wide range of cellular processes, including:
- RNA export: m⁵C can play a role in transporting mRNA from the nucleus to the cytoplasm.
- Translation: Both m⁵C and Ψ can influence the rate and fidelity of protein synthesis, potentially even leading to alternative protein products.
- mRNA stability: Ψ, for example, can enhance mRNA stability by affecting its structure and protecting it from degradation.
- Development and disease: Alterations in these modifications are linked to various physiological and pathological processes, including embryonic development and tumor formation.
The development of new sequencing technologies has revolutionized the field. Mass spectrometry-based methods and antibody-enrichment sequencing techniques such as MeRIP-seq (Methyl RNA Immunoprecipitation) [23] and miCLIP (mRNA individual nucleotide resolution CrossLinking Immunoprecipitation) [24] have mapped many modifications, albeit with limitations in resolution and specificity [16]. More recently, nanopore sequencing has enabled direct RNA sequencing and detection of modifications in native RNA molecules without prior conversion to cDNA, opening new avenues for studying both endogenous and environmental RNA (eRNA) [17,18].
The aim of this review is to systematically summarize the major known modifications of mRNA, ranking them by their prevalence in the scientific literature as a proxy for research emphasis. We describe their biochemical mechanisms, molecular functions, and emerging detection technologies. A final section highlights future directions, including nanopore-based discovery of novel modifications in environmental and mirror-RNA contexts, as proposed in our POLET hypothesis [19].
2. Materials and Methods
To rank mRNA modifications by their prevalence in the literature, we systematically queried the PubMed database (https://pubmed.ncbi.nlm.nih.gov) on June 20, 2025 using both the full chemical name and the abbreviated code for each RNA modification (e.g., “N1-methyladenosine” and “m1A”). Each modification listed in the MODOMICS database (https://iimcb.genesilico.pl/modomics/) was included if it was classified as naturally occurring in cellular RNA. These included modifications found in messenger RNA (mRNA), transfer RNA (tRNA), ribosomal RNA (rRNA), and non-coding RNAs (ncRNAs).
To ensure the specificity of our results, search terms were restricted to exact phrase matches when possible, and abstracts were manually screened to confirm that the citation referenced RNA modifications rather than unrelated chemical analogs or non-RNA-related methylation. Both nucleoside (base + sugar) and nucleotide (base + sugar + phosphate) forms were included in the initial screening; however, final analysis focused exclusively on naturally occurring nucleosides.
The number of PubMed-indexed publications for each RNA modification was tallied and used as a proxy for research attention and biological relevance. These citation counts were used to rank the most studied mRNA modifications, and the results are presented in Table 1. For broader context, a more extensive list of RNA modifications—including those found primarily in tRNAs and rRNAs—is provided in Supplementary Table S1.
3. Results
3.1. Ranking of mRNA Modifications by PubMed Prevalence
We systematically ranked naturally occurring mRNA modifications by their prevalence in the scientific literature, using PubMed-indexed citations as a proxy for research emphasis. Queries included both chemical names and common abbreviations, and results were manually filtered for relevance to mRNA biology (excluding tRNA, rRNA, or DNA-specific contexts). The results reflect not only the biochemical importance of these modifications but also trends in technological accessibility and biomedical interest (Table 1 and Figure 1).
- N6-methyladenosine (m6A) ranked first with over 7,000 citations. This modification is extensively studied due to its widespread presence in mRNAs and its central roles in splicing, export, translation efficiency, and decay. Its regulation by “writers” (METTL3/14) (m6A methyltransferase 3/14), “erasers” (FTO, ALKBH5) (Fatso alpha-ketoglutarate-dependent dioxygenase, AlkB homolog 5), and “readers” has defined the emerging field of epitranscriptomics (reviewed in [25,26]).
- Pseudouridine (Ψ) was second, with ~1,000 citations. Once thought to be restricted to non-coding RNAs, Ψ is now recognized as a key player in mRNA stability, stress response, and synthetic mRNA vaccine design. Enzymes like PUS1 and PUS7 catalyze site-specific isomerization [27].
- 5-methylcytidine (m5C), with ~800 citations, modulates mRNA stability and nuclear export. It is written by NSUN2 (NOP2/Sun RNA methyltransferase 2) and DNMT2 (DNA/RNA Nucleotide Methyltransferase 2) and may act in conjunction with binding proteins such as ALYREF (Aly/REF Export Factor) to regulate cytoplasmic localization [28,29,30].
- N1-methyladenosine (m1A), with ~400 publications, affects translation initiation and mRNA secondary structure. Though less abundant, its functional impact can be significant in mitochondrial and stress-induced contexts [33].
- 5’ Cap modifications (Cap0, Cap1, Cap2) had ~300 citations combined. These modifications, installed by RNGTT (RNA Guanylyltransferase And 5’-Phosphatase), RNMT (RNA guanine-7 methyltransferase), CMTR1/2 (Cap MethylTRansferase), help evade innate immune detection and regulate cap-dependent translation [36,37].
- 5-methyluridine (m5U) and 2’-O-methyladenosine (Am) each had <100 citations, reflecting their recent or understudied roles in mRNA, although both are common in tRNA. TRMT2A/B (TRNA Methyltransferase 2 Homolog A) and FTSJ1 (FtsJ RNA 2’-O-Methyltransferase 1) are the main associated enzymes, respectively [38,39].
- N4-acetylcytidine (ac4C) (<50 citations) is a stress-responsive modification installed by NAT10 (N-acetyltransferase 10), linked to increased translation and mRNA stability [40].
- N7-methylguanosine (m7G) (<50 citations) forms part of the 5’ cap structure but has also been detected internally in some mRNAs. It plays roles in nuclear export and translation. Recent studies have suggested that mRNA internal m7G and its writer protein METTL1 (Methyltransferase 1, tRNA Methylguanosine) are closely related to cell metabolism and cancer regulation. The IGF2BP (Insulin Growth Factor 2 Binding Protein) family proteins IGF2BP1-3 can preferentially bind internal mRNA m7G and regulate mRNA stability [41].
- 2’-O-methylguanosine (Gm) and 2’-O-methylcytidine (Cm) each had <30 citations. These modifications occur both in cap-adjacent and internal positions, potentially contributing to mRNA longevity and translation efficiency [42].
- 5-hydroxymethylcytidine (hm5C) (<20 citations) has a poorly defined role in RNA, though its presence suggests possible epigenetic-like regulation analogous to its role in DNA (reviewed in [28]).
- Finally, co-modified m6A/Ψ sites had <10 citations, indicating a nascent field exploring combinatorial regulation of RNA structure and function at dual-modified loci. Long read nanopore sequencing is especially adept at discovering co-modified mRNAs [8].
3.2. Interpretation of Modification Ranking
The prevalence of publications reflects not only biological abundance but also technical detectability and perceived functional importance. m6A dominates the field due to the early availability of high-affinity antibodies and the development of m6A-seq, which catalyzed mechanistic discoveries across diverse biological systems [5,43]. Pseudouridine and m5C followed as sequencing and chemical mapping methods improved [27,44].
Inosine ranks highly due to its role in transcriptome diversification through RNA editing—a uniquely dynamic modification that alters coding potential. The high rank of cap modifications highlights their long-known essential role in translation and immune modulation, particularly relevant to viral and vaccine RNA biology. ICE-seq (Inosine Chemical Erasing) was developed in 2015 which helps explain the high ranking of inosine in mRNA publications [45].
Lower-ranked modifications like ac4C, Gm, Cm, and hm5C likely suffer from limited detection methods and ambiguous biological roles rather than true scarcity. The low ranking of m6A/Ψ co-modifications underscores how technical limitations may obscure complex regulatory interplay, which future single-molecule and multi-modification sequencing technologies are poised to reveal [8].
3.3. Disease Relevance of Top RNA Modifications
RNA modifications play emerging roles in development, disease, and therapy. The m6A mRNA modification is implicated in cancer progression [46], stem cell differentiation [47], and neurodevelopmental disorders [48]. Its dysregulation via altered METTL3 or FTO expression is linked to glioblastoma [49], leukemia [50], and metabolic diseases [51].
Pseudouridine is foundational in the design of synthetic mRNA therapeutics—particularly COVID-19 vaccines—which use Ψ to evade innate immune detection and enhance translation [52]. The m5C RNA modifications or NSUN2 deletions are associated with intellectual disability [53,54] and cancer [55].
Inosine levels are altered in neurodegenerative diseases like ALS [56] and in immune dysregulation syndromes [14,57]. m1A and m6Am modulate translation efficiency and stress responses [58], potentially contributing to cancer cell plasticity and adaptation to hypoxia [59].
Emerging modifications (e.g., ac4C, hm5C) may become important biomarkers or therapeutic targets as detection tools improve [60]. Understanding how these marks influence gene expression in disease-relevant tissues remains a major unmet need.
The growing relevance of epitranscriptomic alterations in human disease underscores the importance of developing high-throughput, base-specific detection tools to enable functional and diagnostic studies across a broad spectrum of RNA modifications.

Table 1.
All known mRNA modifications ranked in order of PubMed citations.
| Rank | Modification | Abbreviation | Enzyme(s) | Role | Estimated PubMed Citations | Reference(s) |
| 1 | N6-methyladenosine | m6A | METTL3, METTL14, FTO, ALKBH5 | Splicing, translation, decay, export | >7000 | [61,62,63] |
| 2 | Pseudouridine | Ψ | PUS1, PUS7 | Stability, decoding, stress response | ~1000 | [64,65] |
| 3 | 5-methylcytidine | m5C | NSUN2, DNMT2 | Export, stability | ~800 | [66,67] |
| 4 | Inosine | I | ADAR1, ADAR2 | A-to-I editing, recoding | ~750 | [68] |
| 5 | N1-methyladenosine | m1A | TRMT6/TRMT61A | Translation initiation, structure | ~400 | [69,70] |
| 6 | N6,2’-O-dimethyladenosine | m6Am | PCIF1 | Cap-proximal stability | ~200 | [71] |
| 7 | 5’ cap modifications | Cap0, Cap1, Cap2 | RNGTT, RNMT, CMTR1, CMTR2 | Immune evasion, translation | ~150–300 | [72,73] |
| 8 | 5-methyluridine | m5U | TRMT2A/B | tRNA-like stability role in mRNA | <100 | [74] |
| 9 | 2’-O-methyladenosine | Am | FTSJ1, CMTR1 | Cap stability and processing | <100 | [73,75] |
| 10 | N4-acetylcytidine | ac4C | NAT10 | Translation, stress response | <50 | [76] |
| 11 | N7-methylguanosine | m7G | RNGTT, RNMT, METTL1 | 5’ cap structure, nuclear export | <50 | [72,77] |
| 12 | 2’-O-methylguanosine | Gm | CMTR2 | Cap and internal stability | <30 | [72,73] |
| 13 | 2’-O-methylcytidine | Cm | FTSJ1 | Stability, cap modification | <30 | [39] |
| 14 | 5-hydroxymethylcytidine | hm5C | TET2 | Epigenetic-like regulation | <20 | [78] |
| 15 | m6A:Ψ co-modified sites | m6A/Ψ | Multiple | Dynamic regulation, RNA structure | <10 | [78] |
1 Rank is based on the number of PubMed citations (see Materials and Methods). References are the first or most relevant PubMed citation of the enzymes involved in the mRNA modification.
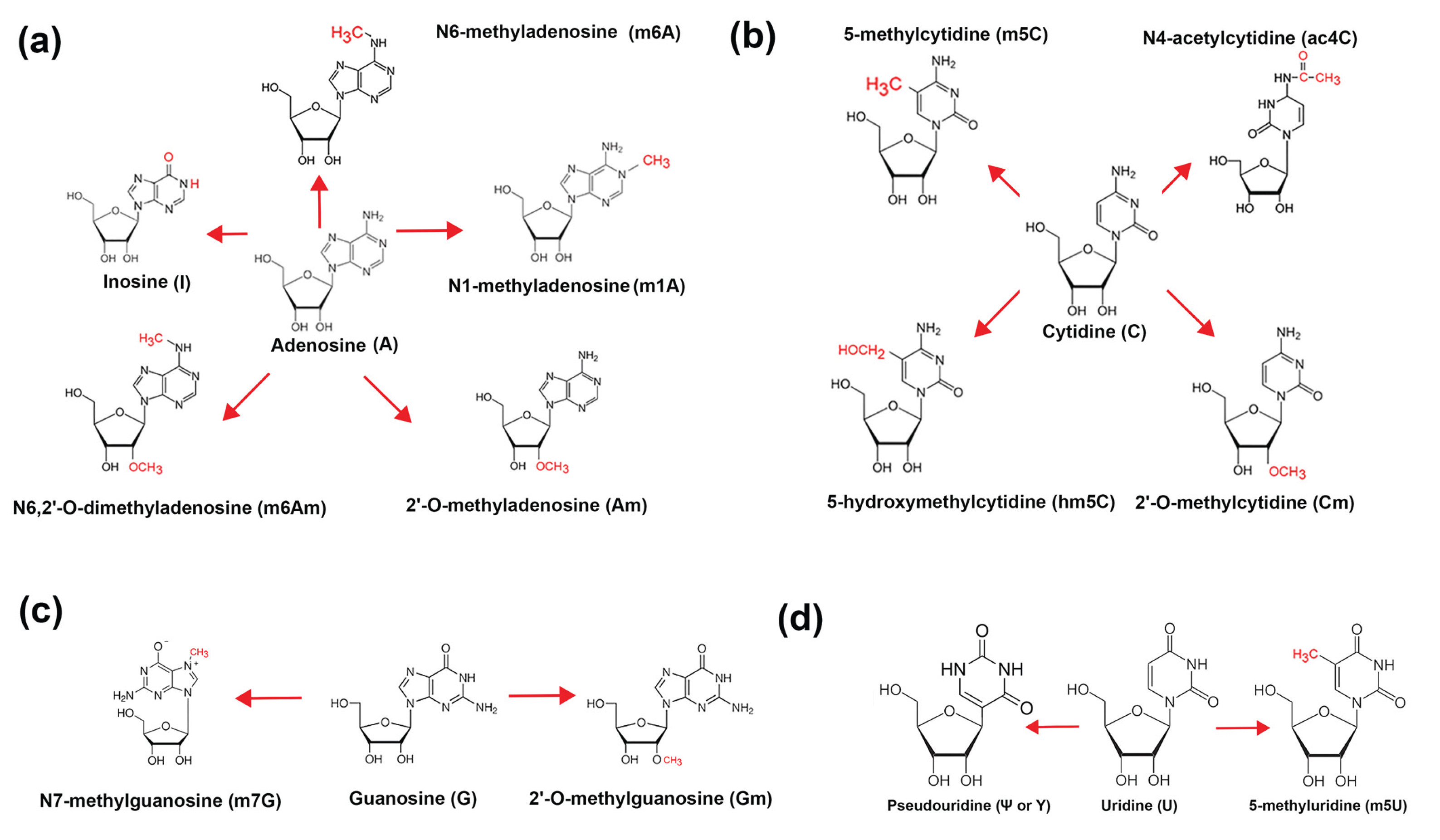
Figure 1.
Modifications of bases on mRNA. (a) Adenosine modifications; (b) Cytidine modifications; (c) Guanosine modifications; and (d) Uridine modifications.
Figure 1.
Modifications of bases on mRNA. (a) Adenosine modifications; (b) Cytidine modifications; (c) Guanosine modifications; and (d) Uridine modifications.
4. Discussion
The findings presented in this review emphasize the central role of m6A in mRNA regulation, reflecting both its biological relevance and the technical accessibility of its detection. The strong representation of m6A in the literature—surpassing 7000 PubMed citations—demonstrates its position as a dominant epitranscriptomic regulator involved in nearly every stage of mRNA life, from splicing and nuclear export to translation and degradation (reviewed in [25]). This prominence has been further amplified by the development of antibody-based enrichment techniques and transcriptome-wide mapping methods such as MeRIP-seq, which have made m6A one of the most tractable RNA modifications for large-scale studies [79].
In contrast, other modifications such as pseudouridine (Ψ), m5C, and inosine also show significant presence in the literature, but their detection and interpretation have historically required more specialized tools. Ψ, for instance, has been associated with enhanced transcript stability and stress response, yet lacks reliable transcriptome-wide detection without specialized chemical treatment [27]. Similarly, A-to-I editing by ADARs, while essential for neural and immune development, is sometimes overlooked in the context of dynamic mRNA regulation (reviewed in [80]).
RNA capping is a critical post-transcriptional modification that governs RNA stability, processing, and translational efficiency. The canonical eukaryotic cap structures—Cap0 (m⁷GpppN), Cap1 (m⁷GpppNm), and Cap2 (m⁷GpppNmNm)—play essential roles in promoting translation initiation and protecting transcripts from innate immune surveillance. Despite their fundamental importance, these cap modifications remain underrepresented in the literature, likely due to longstanding technical challenges in isolating and sequencing intact 5′ caps, especially within full-length mRNAs. Recent advances, however, have begun to reveal the broader biological relevance of cap methylation patterns. Differential cap modifications are now recognized as key determinants in viral mimicry, host-pathogen interactions, and the optimization of synthetic mRNA therapeutics. The field has also undergone a paradigm shift with the discovery of non-canonical RNA caps, initially in bacteria and now recognized across all domains of life. The repertoire of RNA caps has expanded well beyond the classic m⁷G structure to include metabolite-derived caps such as NAD⁺, FAD, Coenzyme A (CoA), UDP-glucose, and ADP-ribose. In addition, cells produce dinucleoside polyphosphate “alarmone” caps and methylated phosphate-containing cap-like structures. These non-canonical caps open new avenues for studying RNA regulation, signaling, and host-pathogen dynamics, while also posing intriguing questions about cap recognition and processing machinery across species. (reviewed in [81]).
The relatively low citation counts for certain mRNA modifications—such as N¹-methyladenosine (m¹A), 5-methyluridine (m⁵U), and 2’-O-methyladenosine (Am)—should not be interpreted as evidence of limited biological significance. Instead, they reflect persistent technological and methodological barriers that have hindered our ability to detect, map, and quantify these modifications at transcriptome-wide scale in messenger RNAs. In contrast, these marks have long been studied in the context of tRNA and rRNA, where they are more abundant and their structural or functional roles more clearly defined. For example, m¹A is known to alter base pairing and RNA secondary structure, which can influence translation initiation or pause sites (reviewed in [82]), yet its detection in mRNA is complicated by its lability and the need for specialized chemical treatment or ultra-sensitive sequencing approaches. Similarly, m⁵U and Am, though functionally well-established in non-coding RNAs and tRNA fragments [83], lack high-throughput mapping tools specific to their low-abundance and dispersed presence in mRNAs. Many of these modifications require enrichment strategies or site-specific antibodies that are currently unavailable or unreliable.
These technical limitations result in underrepresentation of such modifications in both experimental studies and the literature. As novel chemical labeling methods, direct RNA sequencing platforms (e.g., nanopore), and improved base calling algorithms evolve, the field is poised to uncover the broader regulatory roles these modifications may play in mRNA metabolism, localization, and translation—particularly under stress or in specialized cell types. Thus, low citation prevalence is more a reflection of our current technological blind spots than an accurate measure of functional importance.
These findings underscore the urgent need for comprehensive and unbiased platforms capable of detecting the full spectrum of RNA modifications. Nanopore-based direct RNA sequencing—elaborated in the Future Directions section—offers a particularly promising solution. Unlike traditional methods that rely on reverse transcription, nanopore sequencing reads native RNA molecules directly, enabling the detection of base-specific modifications through characteristic shifts in ionic current. A major advantage of this approach is its ability to simultaneously detect multiple modifications along a single full-length mRNA transcript, preserving the contextual relationships between marks [83]. As the accuracy, resolution, and computational interpretation of nanopore data continue to improve, this technology has the potential to transform the field—broadening our understanding of RNA modifications across diverse transcript classes, cell types, and physiological conditions.
5. Conclusions and Future Directions
The field of RNA modification research is rapidly evolving, yet significant challenges remain in the comprehensive identification and functional annotation of the full diversity of RNA modifications. The MODOMICS database currently catalogs over 335 distinct chemical modifications of RNA, most of which remain poorly characterized in terms of their distribution, dynamics, and biological function (Figure 2) [4]. A central future goal is the development of both biochemical and sequencing technologies capable of detecting these modifications with high sensitivity and specificity. Among the most promising approaches are antibody-based enrichment techniques and direct RNA sequencing using nanopore technologies. However, the current suite of available antibodies targets only a limited set of well-studied modifications such as m⁶A, m⁵C, and pseudouridine [84,85]. There is a pressing need to develop a broader repertoire of high-affinity, modification-specific antibodies, ideally with minimal cross-reactivity, to facilitate immunoprecipitation-based enrichment and mapping of additional modification types across the transcriptome [85].
Parallel to biochemical methods, advances in nanopore sequencing hold transformative potential for detecting RNA modifications directly, without prior conversion or enrichment. The Oxford Nanopore platform has already demonstrated the ability to detect some common modifications by characteristic shifts in ionic current (reviewed in [86]), but most of the 335 known modifications in the MODOMICS database remain indistinguishable due to limitations in both experimental resolution and current basecalling algorithms [6]. Future work must therefore focus on enhancing both the hardware sensitivity of nanopores and the machine learning-based bioinformatics tools that interpret the raw electrical signal [4]. These improvements will require training datasets that include synthetic RNAs bearing single, known modifications in defined sequence contexts [87]. Ideally, comprehensive modification “barcodes” could be integrated into public databases to facilitate community-wide benchmarking [6].
The MODOMICS database assigns a unique one-character code to each of the more than 500 known natural and synthetic RNA modifications, enabling streamlined annotation and computational analysis [6]. To accommodate the growing diversity of chemical modifications beyond the canonical nucleotides (A, U, G, C), MODOMICS utilizes an extended character set that includes not only Latin letters, but also symbols from Greek, Cyrillic, and Chinese scripts, as well as other Unicode characters. This expansive coding system allows for the efficient representation of structurally diverse nucleotide variants, including rare, heavily modified, or synthetic analogs used in RNA therapeutics and research [6]. While powerful, this multilingual symbolic system poses challenges for data standardization, software compatibility, and cross-platform interoperability—highlighting the need for harmonized bioinformatics tools capable of parsing and visualizing these expanded alphabets within RNA sequences.
An additional frontier involves the detection and sequencing of mirror RNAs—hypothetical nucleic acid structures composed of D-ribose rather than the naturally occurring L-ribose. The POLET hypothesis (Preprokaryotic Organismal Lifeforms Existing Today) proposes that such mirror RNAs may have existed alongside standard RNAs during early evolution and could persist in protected or engineered environments (doi: 10.20944/preprints202506.1269.v1). Detecting these stereoisomeric RNAs poses unique technical challenges, as current polymerases and helicases used in sequencing platforms are stereospecific for natural L-RNA. Thus, a critical innovation will be the development of mirror helicases capable of processing D-RNA through nanopore sequencers or modified platforms [88]. These synthetic motor proteins could unlock access to previously undetectable nucleic acid species, including mirror RNAs that may be relics of ancient biochemistry or products of deliberate design [89].
The growing interest in mirror RNA extends well beyond theoretical or academic curiosity. One of its most promising applications lies in the development of antisense oligonucleotide (ASO) drugs [90]. Because mirror RNAs are composed of D-ribose—opposite in chirality to natural L-RNA—they are completely resistant to degradation by endogenous nucleases. This remarkable stability offers a major advantage for therapeutic applications, enabling longer half-lives and reduced dosing frequency. Mirror-RNA-based ASOs could therefore represent a next-generation platform for treating genetic and infectious diseases with enhanced durability and specificity [90].
At the same time, the resilience of mirror RNA has also raised legitimate concerns about its potential misuse. Theoretical models suggest that mirror viruses—constructed entirely from mirror nucleotides—could evade immune detection and remain invisible to conventional RNA-based diagnostics, posing a unique challenge to biosecurity [91]. As a result, the development of analytical tools capable of detecting and characterizing mirror RNA is not only a scientific imperative but also a strategic priority. Emerging solutions may include stereochemistry-specific probes, chiral aptamers, or customized nanopore sensors engineered to distinguish D- from L-RNA at the single-molecule level [91]. Together, these efforts will not only mitigate risk but also unlock the vast, untapped potential of mirror RNA in both medicine and synthetic biology.
In conclusion, future directions in RNA modification research must encompass expansion of antibody and nanopore-based technologies to detect the full spectrum of natural modifications, as well as pioneering efforts to sequence synthetic or mirror RNAs in the context of both evolutionary biology and national security.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: Short description and number of citations for many of the 335 RNA modifications from MODOMICS database [4].
Author Contributions
DMR has conducted all research and writing of this manuscript.
Funding
This research was funded by the National Institutes of Health, grant numbers 5UG3OD023285, 5P42ES030991, and 1P30ES036084.
Acknowledgments
The author has reviewed and edited the output and takes full responsibility for the content of this publication.
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDPI | Multidisciplinary Digital Publishing Institute |
| POLET | Preprokaryotic Organismal Lifeforms Existing Today |
| m6A | N6-methyladenosine |
| Ψ | Pseudouridine |
| m5C | 5-methylcytidine |
| I | Inosine |
| m1A | N1-methyladenosine |
| m6Am | N6,2’-O-dimethyladenosine |
| Cap0, Cap1, Cap2 | 5’ cap modifications using m7G |
| m5U | 5-methyluridine |
| Am | 2’-O-methyladenosine |
| ac4C | N4-acetylcytidine |
| m7G | N7-methylguanosine |
| Gm | 2’-O-methylguanosine |
| Cm | 2’-O-methylcytidine |
| hm5C | 5-hydroxymethylcytidine |
| m6A/Ψ | m6A:Ψ co-modified sites |
References
- Alasar, A.A., et al., Genomewide m(6)A Mapping Uncovers Dynamic Changes in the m(6)A Epitranscriptome of Cisplatin-Treated Apoptotic HeLa Cells. Cells, 2022. 11(23). [CrossRef]
- Meyer, K.D. and S.R. Jaffrey, The dynamic epitranscriptome: N6-methyladenosine and gene expression control. Nat Rev Mol Cell Biol, 2014. 15(5): p. 313-26. [CrossRef]
- Pilala, K.M., et al., Exploring the methyl-verse: Dynamic interplay of epigenome and m6A epitranscriptome. Mol Ther, 2025. 33(2): p. 447-464. [CrossRef]
- Cappannini, A., et al., MODOMICS: a database of RNA modifications and related information. 2023 update. Nucleic Acids Res, 2024. 52(D1): p. D239-D244. [CrossRef]
- Dominissini, D., et al., Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature, 2012. 485(7397): p. 201-6. [CrossRef]
- Tegowski, M., et al., Single-cell m(6)A profiling in the mouse brain uncovers cell type-specific RNA methylomes and age-dependent differential methylation. Nat Neurosci, 2024. 27(12): p. 2512-2520. [CrossRef]
- Zhang, B., et al., Defining context-dependent m(6)A RNA methylomes in Arabidopsis. Dev Cell, 2024. 59(20): p. 2772-2786 e3. [CrossRef]
- Huang, S., A.C. Wylder, and T. Pan, Simultaneous nanopore profiling of mRNA m(6)A and pseudouridine reveals translation coordination. Nat Biotechnol, 2024. 42(12): p. 1831-1835. [CrossRef]
- Li, H., et al., Quantitative RNA pseudouridine maps reveal multilayered translation control through plant rRNA, tRNA and mRNA pseudouridylation. Nat Plants, 2025. 11(2): p. 234-247. [CrossRef]
- Guo, G., et al., Advances in mRNA 5-methylcytosine modifications: Detection, effectors, biological functions, and clinical relevance. Mol Ther Nucleic Acids, 2021. 26: p. 575-593. [CrossRef]
- Wang, R., et al., The Quiet Giant: Identification, Effectors, Molecular Mechanism, Physiological and Pathological Function in mRNA 5-methylcytosine Modification. Int J Biol Sci, 2024. 20(15): p. 6241-6254. [CrossRef]
- Chen, S., J. Meng, and Y. Zhang, Quantitative profiling N1-methyladenosine (m1A) RNA methylation from Oxford nanopore direct RNA sequencing data. Methods, 2024. 228: p. 30-37. [CrossRef]
- Liu, Y., et al., Research progress of N1-methyladenosine RNA modification in cancer. Cell Commun Signal, 2024. 22(1): p. 79. [CrossRef]
- Mendoza, H.G. and P.A. Beal, Structural and functional effects of inosine modification in mRNA. RNA, 2024. 30(5): p. 512-520. [CrossRef]
- Poyau, A., et al., Identification and relative quantification of adenosine to inosine editing in serotonin 2c receptor mRNA by CE. Electrophoresis, 2007. 28(16): p. 2843-52. [CrossRef]
- Xia, L. and P. Yin, Upregulated m7G methyltransferase METTL1 is a potential biomarker and tumor promoter in skin cutaneous melanoma. Front Immunol, 2025. 16: p. 1575219. [CrossRef]
- Xu, C., et al., The m(7)G methylation modification: An emerging player of cardiovascular diseases. Int J Biol Macromol, 2025. 309(Pt 3): p. 142940. [CrossRef]
- Akichika, S. and T. Suzuki, Cap-specific m(6)Am modification: A transcriptional anti-terminator by sequestering PCF11 with implications for neuroblastoma therapy. Mol Cell, 2024. 84(21): p. 4051-4052. [CrossRef]
- Liu, J.F., CROWN-seq reveals m(6)Am landscapes and transcription start site diversity. Nat Rev Genet, 2025. [CrossRef]
- Dohnalkova, M., et al., Essential roles of RNA cap-proximal ribose methylation in mammalian embryonic development and fertility. Cell Rep, 2023. 42(7): p. 112786. [CrossRef]
- Kishore, U. and T.A. Kufer, Editorial: Updates on RIG-I-like receptor-mediated innate immune responses. Front Immunol, 2023. 14: p. 1153410. [CrossRef]
- Kouwaki, T., et al., RIG-I-Like Receptor-Mediated Recognition of Viral Genomic RNA of Severe Acute Respiratory Syndrome Coronavirus-2 and Viral Escape From the Host Innate Immune Responses. Front Immunol, 2021. 12: p. 700926. [CrossRef]
- Li, Y., et al., MeRIP-PF: an easy-to-use pipeline for high-resolution peak-finding in MeRIP-Seq data. Genomics Proteomics Bioinformatics, 2013. 11(1): p. 72-5. [CrossRef]
- Hawley, B.R. and S.R. Jaffrey, Transcriptome-Wide Mapping of m(6) A and m(6) Am at Single-Nucleotide Resolution Using miCLIP. Curr Protoc Mol Biol, 2019. 126(1): p. e88.
- Dong, Q., et al., N6-Methyladenosine Modification of the Three Components “Writers”, “Erasers”, and “Readers” in Relation to Osteogenesis. Int J Mol Sci, 2025. 26(12).
- Wang, S., et al., New Targets for Immune Inflammatory Response in Rheumatoid Arthritis: Focus on the Potential Significance of N6-Methyladenosine, Ferroptosis and Cuproptosis. J Inflamm Res, 2025. 18: p. 8085-8106. [CrossRef]
- Zhang, L.S., et al., BID-seq for transcriptome-wide quantitative sequencing of mRNA pseudouridine at base resolution. Nat Protoc, 2024. 19(2): p. 517-538. [CrossRef]
- Li, M., et al., 5-methylcytosine RNA methyltransferases and their potential roles in cancer. J Transl Med, 2022. 20(1): p. 214. [CrossRef]
- Ye, X., et al., Pan-cancer analysis of RNA 5-methylcytosine reader (ALYREF). Oncol Res, 2024. 32(3): p. 503-515. [CrossRef]
- Yang, X., et al., 5-methylcytosine promotes mRNA export - NSUN2 as the methyltransferase and ALYREF as an m(5)C reader. Cell Res, 2017. 27(5): p. 606-625. [CrossRef]
- Roth, S.H., E.Y. Levanon, and E. Eisenberg, Genome-wide quantification of ADAR adenosine-to-inosine RNA editing activity. Nat Methods, 2019. 16(11): p. 1131-1138. [CrossRef]
- Yuting, K., D. Ding, and H. Iizasa, Adenosine-to-Inosine RNA Editing Enzyme ADAR and microRNAs. Methods Mol Biol, 2021. 2181: p. 83-95.
- Deng, Y., et al., N1-methyladenosine RNA methylation patterns are associated with an increased risk to biochemical recurrence in prostate cancer and serve as a potential novel biomarker for patient stratification. Int Immunopharmacol, 2024. 143(Pt 2): p. 113404. [CrossRef]
- Mauer, J., et al., Reversible methylation of m(6)A(m) in the 5’ cap controls mRNA stability. Nature, 2017. 541(7637): p. 371-375. [CrossRef]
- Sugita, A., et al., Cap-Specific m(6)Am Methyltransferase PCIF1/CAPAM Regulates mRNA Stability of RAB23 and CNOT6 through the m(6)A Methyltransferase Activity. Cells, 2024. 13(20).
- Chen, P., et al., Capping Enzyme mRNA-cap/RNGTT Regulates Hedgehog Pathway Activity by Antagonizing Protein Kinase A. Sci Rep, 2017. 7(1): p. 2891. [CrossRef]
- Pearson, L.A., et al., Characterisation of RNA guanine-7 methyltransferase (RNMT) using a small molecule approach. Biochem J, 2025. 482(4). [CrossRef]
- Witzenberger, M., et al., Human TRMT2A methylates tRNA and contributes to translation fidelity. Nucleic Acids Res, 2023. 51(16): p. 8691-8710. [CrossRef]
- Freude, K., et al., Mutations in the FTSJ1 gene coding for a novel S-adenosylmethionine-binding protein cause nonsyndromic X-linked mental retardation. Am J Hum Genet, 2004. 75(2): p. 305-9. [CrossRef]
- Liu, W.C., et al., Inhibition of tumor-intrinsic NAT10 enhances antitumor immunity by triggering type I interferon response via MYC/CDK2/DNMT1 pathway. Nat Commun, 2025. 16(1): p. 5154.
- Liu, C., et al., IGF2BP3 promotes mRNA degradation through internal m(7)G modification. Nat Commun, 2024. 15(1): p. 7421. [CrossRef]
- Tardu, M., et al., Identification and Quantification of Modified Nucleosides in Saccharomyces cerevisiae mRNAs. ACS Chem Biol, 2019. 14(7): p. 1403-1409. [CrossRef]
- Yang, J., et al., Machine learning-augmented m6A-Seq analysis without a reference genome. Brief Bioinform, 2025. 26(3). [CrossRef]
- Carlile, T.M., M.F. Rojas-Duran, and W.V. Gilbert, Pseudo-Seq: Genome-Wide Detection of Pseudouridine Modifications in RNA. Methods Enzymol, 2015. 560: p. 219-45.
- Suzuki, T., et al., Transcriptome-wide identification of adenosine-to-inosine editing using the ICE-seq method. Nat Protoc, 2015. 10(5): p. 715-32. [CrossRef]
- Tao, Q. and Q. Zhang, Translating the m(6)A epitranscriptome for prostate cancer. Nat Rev Urol, 2025. [CrossRef]
- Feng, B., et al., Transcriptomic Analysis of the m6A Reader YTHDF2 in the Maintenance and Differentiation of Human Embryonic Stem Cells. Stem Cells, 2025. [CrossRef]
- Widagdo, J. and V. Anggono, The m6A-epitranscriptomic signature in neurobiology: from neurodevelopment to brain plasticity. J Neurochem, 2018. 147(2): p. 137-152. [CrossRef]
- Liu, H., et al., METTL3/YTDHF1 Stabilizes MTCH2 mRNA to Regulate Ferroptosis in Glioma Cells. Front Biosci (Landmark Ed), 2025. 30(2): p. 25718.
- Liu, X., et al., Novel Associations Between METTL3 Gene Polymorphisms and Pediatric Acute Lymphoblastic Leukemia: A Five-Center Case-Control Study. Front Oncol, 2021. 11: p. 635251.
- Wang, S., et al., M6A methyltransferase METTL3 promotes glucose metabolism hub gene expression and induces metabolic dysfunction-associated steatotic liver disease (MASLD). BMC Genomics, 2025. 26(1): p. 188. [CrossRef]
- Morais, P., H. Adachi, and Y.T. Yu, The Critical Contribution of Pseudouridine to mRNA COVID-19 Vaccines. Front Cell Dev Biol, 2021. 9: p. 789427. [CrossRef]
- Abbasi-Moheb, L., et al., Mutations in NSUN2 cause autosomal-recessive intellectual disability. Am J Hum Genet, 2012. 90(5): p. 847-55. [CrossRef]
- Yang, Q., et al., A novel variant in NSUN2 causes intellectual disability in a Chinese family. BMC Med Genomics, 2024. 17(1): p. 95. [CrossRef]
- Wang, Z., et al., FOXA1-dependent NSUN2 facilitates the advancement of prostate cancer by preserving TRIM28 mRNA stability in a m5C-dependent manner. NPJ Precis Oncol, 2025. 9(1): p. 127. [CrossRef]
- Yang, Y., S. Okada, and M. Sakurai, Adenosine-to-inosine RNA editing in neurological development and disease. RNA Biol, 2021. 18(7): p. 999-1013. [CrossRef]
- Cui, N., et al., Extracellular Inosine Induces Anergy in B Cells to Alleviate Autoimmune Hepatitis. Cell Mol Gastroenterol Hepatol, 2025: p. 101539. [CrossRef]
- Alriquet, M., et al., The protective role of m1A during stress-induced granulation. J Mol Cell Biol, 2021. 12(11): p. 870-880. [CrossRef]
- Ren, M., et al., Exploration and validation of a combined Hypoxia and m6A/m5C/m1A regulated gene signature for prognosis prediction of liver cancer. BMC Genomics, 2023. 24(1): p. 776. [CrossRef]
- Yang, L., et al., NAT10 inhibition alleviates astrocyte autophagy by impeding ac4C acetylation of Timp1 mRNA in ischemic stroke. Acta Pharm Sin B, 2025. 15(5): p. 2575-2592. [CrossRef]
- Liu, J., et al., A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat Chem Biol, 2014. 10(2): p. 93-5. [CrossRef]
- Jia, G., et al., N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol, 2011. 7(12): p. 885-7. [CrossRef]
- Zheng, G., et al., ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell, 2013. 49(1): p. 18-29. [CrossRef]
- Patton, J.R., et al., Mitochondrial myopathy and sideroblastic anemia (MLASA): missense mutation in the pseudouridine synthase 1 (PUS1) gene is associated with the loss of tRNA pseudouridylation. J Biol Chem, 2005. 280(20): p. 19823-8.
- de Brouwer, A.P.M., et al., Variants in PUS7 Cause Intellectual Disability with Speech Delay, Microcephaly, Short Stature, and Aggressive Behavior. Am J Hum Genet, 2018. 103(6): p. 1045-1052.
- Moon, H.J. and K.L. Redman, Trm4 and Nsun2 RNA:m5C methyltransferases form metabolite-dependent, covalent adducts with previously methylated RNA. Biochemistry, 2014. 53(45): p. 7132-44.
- Schaefer, M., et al., Azacytidine inhibits RNA methylation at DNMT2 target sites in human cancer cell lines. Cancer Res, 2009. 69(20): p. 8127-32. [CrossRef]
- Gerber, A., et al., Tad1p, a yeast tRNA-specific adenosine deaminase, is related to the mammalian pre-mRNA editing enzymes ADAR1 and ADAR2. EMBO J, 1998. 17(16): p. 4780-9. [CrossRef]
- Wang, B., et al., RNA m1A Methyltransferase TRMT6 Predicts Poorer Prognosis and Promotes Malignant Behavior in Glioma. Front Mol Biosci, 2021. 8: p. 692130. [CrossRef]
- Chujo, T. and T. Suzuki, Trmt61B is a methyltransferase responsible for 1-methyladenosine at position 58 of human mitochondrial tRNAs. RNA, 2012. 18(12): p. 2269-76. [CrossRef]
- Akichika, S., et al., Cap-specific terminal N (6)-methylation of RNA by an RNA polymerase II-associated methyltransferase. Science, 2019. 363(6423).
- Pillutla, R.C., et al., Human mRNA capping enzyme (RNGTT) and cap methyltransferase (RNMT) map to 6q16 and 18p11.22-p11.23, respectively. Genomics, 1998. 54(2): p. 351-3. [CrossRef]
- Belanger, F., et al., Characterization of hMTr1, a human Cap1 2’-O-ribose methyltransferase. J Biol Chem, 2010. 285(43): p. 33037-33044.
- Carter, J.M., et al., FICC-Seq: a method for enzyme-specified profiling of methyl-5-uridine in cellular RNA. Nucleic Acids Res, 2019. 47(19): p. e113. [CrossRef]
- Guy, M.P., et al., Yeast Trm7 interacts with distinct proteins for critical modifications of the tRNAPhe anticodon loop. RNA, 2012. 18(10): p. 1921-33. [CrossRef]
- Liu, H., et al., DNA damage induces N-acetyltransferase NAT10 gene expression through transcriptional activation. Mol Cell Biochem, 2007. 300(1-2): p. 249-58. [CrossRef]
- Bahr, A., et al., Molecular analysis of METTL1, a novel human methyltransferase-like gene with a high degree of phylogenetic conservation. Genomics, 1999. 57(3): p. 424-8. [CrossRef]
- Fan, W., et al., Tet2 modulates M2 macrophage polarization via mRNA 5-methylcytosine in allergic rhinitis. Int Immunopharmacol, 2024. 143(Pt 3): p. 113495. [CrossRef]
- Meyer, K.D., et al., Comprehensive analysis of mRNA methylation reveals enrichment in 3’ UTRs and near stop codons. Cell, 2012. 149(7): p. 1635-46. [CrossRef]
- Du, Y., et al., Evolutionary Origins and Adaptive Significance of A-to-I RNA Editing in Animals and Fungi. Bioessays, 2025. 47(5): p. e202400220. [CrossRef]
- Potuznik, J.F. and H. Cahova, If the 5’ cap fits (wear it) - Non-canonical RNA capping. RNA Biol, 2024. 21(1): p. 1-13.
- Pajdzik, K., et al., Chemical manipulation of m(1)A mediates its detection in human tRNA. RNA, 2024. 30(5): p. 548-559. [CrossRef]
- Guo, G., et al., Disease Activity-Associated Alteration of mRNA m(5) C Methylation in CD4(+) T Cells of Systemic Lupus Erythematosus. Front Cell Dev Biol, 2020. 8: p. 430. [CrossRef]
- Itoh, K., M. Mizugaki, and N. Ishida, Detection of elevated amounts of urinary pseudouridine in cancer patients by use of a monoclonal antibody. Clin Chim Acta, 1989. 181(3): p. 305-15. [CrossRef]
- Kumar, S. and M. Sinha, Targeting intracellular mRNA m(6)A-modifiers in advancing immunotherapeutics. J Adv Res, 2025.
- Ranga, S., et al., Modifications of RNA in cancer: a comprehensive review. Mol Biol Rep, 2025. 52(1): p. 321. [CrossRef]
- Spangenberg, J., et al., The RMaP challenge of predicting RNA modifications by nanopore sequencing. Commun Chem, 2025. 8(1): p. 115.
- Xu, Y. and T.F. Zhu, Mirror-image T7 transcription of chirally inverted ribosomal and functional RNAs. Science, 2022. 378(6618): p. 405-412. [CrossRef]
- Service, R.F., A big step toward mirror-image ribosomes. Science, 2022. 378(6618): p. 345-346. [CrossRef]
- Zhang, Y., Y. Dantsu, and W. Zhang, Construction of a Mirror-Image RNA Nanostructure for Enhanced Biostability and Drug Delivery Efficiency. ACS Biomater Sci Eng, 2025. 11(4): p. 2408-2421. [CrossRef]
- Adamala, K.P., et al., Confronting risks of mirror life. Science, 2024. 386(6728): p. 1351-1353. [CrossRef]
Figure 2.
The 335 naturally occurring nucleosides and nucleotides in the most recent MODOMICS database showing their distribution on U, A, C, and G.
Figure 2.
The 335 naturally occurring nucleosides and nucleotides in the most recent MODOMICS database showing their distribution on U, A, C, and G.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



