
Submitted:
30 June 2025
Posted:
30 June 2025
You are already at the latest version
Abstract
The application of artificial intelligence through the brain computer interface (BCI) is proving to be one of the great advances in neuroscience today. The development of surface electrodes over the cortex, and very fine electrodes that can be stereotactically implanted in the brain have moved the science forward to the extent that the paralyzed can play chess, and the blind can read letters. However, introduction of foreign bodies into deeper parts of the central nervous system results in foreign body reaction, scarring, apoptosis, and decreased signaling. Implanted electrodes activate microglia, causing release of inflammatory factors, recruitment of systemic inflammatory cells to the site of injury and ultimately glial scarring and encapsulation of the electrode. Recordings historically fail between 6 months to 1 year, the longest BCI in use has been 7years.
This article offers a biomolecular strategy provided by angiogenic cell precursors (ACP) and
nerve cell precursors (NCP), either intrathecally or intra-arterially. Together through downstream activation of neurotrophic factors, they exert a salutary immunomodulatory suppression of inflammation, anti-apoptosis, healing, homeostasis, angiogenesis, differentiation, synaptogenesis, neuritogenesis and learning-associated plasticity. This combination of cells is anticipated to sustain and promote learning across the BCI.
Keywords:
ACP
; NCP
; NF-κB
; NK cells
; Brain/AI interface
; M2 phenotype
; IL-8
; learning
; P75NTR
Research and Development of Brain-Computer Interfaces (BCI)s has flourished in the past several years, offering hope to individuals suffering from a myriad of conditions including paralysis, ALS, blindness and Parkinson’s. BCI implants from several companies have allowed paralyzed patients to control robotic arms, write emails, and play chess with their thoughts. A non-verbal man with ALS recovered his voice with a Neuralink implant, and a Prima implant gave blind individuals “artificial vision”, allowing them to perceive letters and shapes. BCI convert electrical signals from the brain into computer code which can be used for applications such as moving and clicking a mouse cursor. Non-invasive modalities are disadvantaged by weaker brain signals which require expensive amplification hardware and sophisticated signal processing techniques [1].
Invasive BCI have better signal quality. However, electrodes implanted into the brain cause inflammation, scarring and loss of neurons. There is an increased risk of infection and rejection of the implant as a foreign entity [1,2]. Perpetual inflammation following implantation can damage surrounding tissues, affect function and cause failure of the implant. Recordings historically fail between 6 months to 1 year; the longest BCI in use has been 7 years [3]. A more pervasive problem is that implanted electrodes cause glial scarring and encapsulation of the electrode. This is the result of activation of the microglia, which constitute 10% of brain cells- and the subsequent release of inflammatory factors and recruitment of systemic inflammatory cells to the site of injury [4]. The scarring is exacerbated by migration of meningeal fibroblasts along the electrodes [5]. These cells and activated glia eventually encapsulate the electrode with scar tissue and interfere or prevent signal transmission [6].
While the alacrity and skill of electrode placement, and the quality of electrode are significant positive variables affecting the quality of the electrode brain interface, the immune response and triggering of programmed cell death- apoptosis- are presently the limiting factors to long term success of BCIs. Robotic placement of the implants, and micro-scaling of the electrodes will minimize these risks. However, as deeper electrodes are contrived, surface area over the electrodes will necessarily increase, and ultimately result in some degree of inflammation, scarring, apoptosis and decreased signal transmission.
Adult neurons, moreover, have limited capacity for regeneration. Neurons do not undergo cell division, a feature that prohibits cell regeneration, and regeneration of the cellular processes is impeded by a central nervous system environment that generally impedes meaningful axonal and dendritic regenerative outgrowth [7].
Autologous Angiogenic Precursor Cells and Nerve Cell Precursors Offer a Biomolecular Solution to Inflammation, Scarring and Apoptosis
The biomolecular solution to the challenges and consequences of implanted foreign bodies would appear to be the creation of a cellular milieu from which the robust expression of cytokines and growth factors mitigate the scarring process, enhance migration of cells to the injured site, and promote cell survival. The particular combination of angiogenic cell precursors (ACP) and nerve cell precursors (NCP) serve to limit inflammation and apoptosis, and may additionally foster synaptogenesis, neuritogenesis, and ultimately enhance machine learning through long term potentiation (LTP) and memory consolidation.
ACP Attract NK Cells; NK Cells Modulate Scarring
The expression of interleukin-8 (IL-8 or CXCL8) by ACP attract natural killer (NK) cells [8,9,10] (Figure 1). As part of the innate lymphoid cell (ILC) population, NK cells suppress inflammation through receptor-ligand interactions and cytokine secretion. NK cytokine release alters Th subtypes, lysing auto-aggressive T cells, and accelerating maturation of monocytes and dendritic cells [11,12]. NK cells express IFNγ-a potent immunomodulating influencer - and other mediators which create an anti-inflammatory chemokine environment that inhibits the assembly of inflammatory cells, and limits both fibrosis and apoptosis [13,14]. NK cells serve in the first line of defense against intracellular pathogens through expression of type I interferons- IFN-α, IFN-β, and IFN-γ- which enhance the expression of major histocompatibility complex molecules of antigen -presenting cells, and initiate an anti-viral inflammatory cascade, preventing viral replication by detecting and destroying the infected resident cells [15,16]. NK cells are protective against pathogens - such as coxsackievirus B (CBV) and cytomegalovirus - causing meningitis, encephalitis, seizures and myelitis.
CXCR1/2 receptors are expressed on NK cells, myeloid-derived epithelial cells and other cell types [17]. The CXCR1 and CXCR2 receptors on NK cells are highly specific to the ligand chemokine CXCL8 [18,1][]. CXCL8 is the most potent human neutrophil-attracting chemokine, playing a dominant role in response to infection and tissue injury, the activity of which depends upon interaction with human CXC chemokine receptors CXCR1 and CXCR2 [20]. ACP induce high levels of expression of CXCL8 [8,9,10], which exhibits specific chemokine activity for NK cells. Moreover, CXCL8 exhibits a systemic effect that extends to remote populations of NK cells.
NF-κB Inhibits Apoptosis and Regulates Cell Survival
Nuclear Factor kappa B (NF-κB) is activated by the downstream effects of CXCL8 and may be proinflammatory or conversely exert an anti-apoptotic effect, depending on cell type and pathological state. Under the influence of NF-κB, microglia and glial cells in trauma produce excitotoxins, nitric oxide species(NOS), neurotoxic reactive oxygen species (ROS) and proinflammatory cytotoxins [21], and can induce apoptosis in response to certain death-inducing signals, depending upon the type and duration of stimulus. Alternatively, NF-κB proteins can promote the expression of anti-apoptotic genes.
In nerve cell populations the NF-κB influence is usually protective [22]. In neurons, NF-κB induce genetic coding for anti-apoptotic proteins, neurotrophic factors, antioxidant enzymes, and calcium-regulating proteins. NF-κB thus generally functions to inhibit the apoptotic cascade; conversely, blocking NF-κB results in apoptosis. For instance, the inhibition of NF-κB in macrophages results in release of Cytochrome C, an activator of the pro-apoptotic caspases [23,24,25,26,27]. These anti-apoptotic effects are especially demonstrated in sympathetic and sensory neurons, encompassing sequestration of ROS, and enhanced neurite growth [28,29,30,31,32,33,34,35,36,37,38,39,40]. Activation of the NF-κB–MnSOD pathway confers reduction of oxidative stress by mitochondrial sequestration of free radicals [41]. Neuronal target genes encoding for neuronal survival include neuronal apoptosis inhibitory protein (NAIP), calcium calmodulin kinase II δ, (CaMKII δ), brain derived neurotrophic factor (BDNF), μ-opioid receptors, neural cell adhesion molecule (NCAM), manganese superoxide dismutase (MnSOD), Bcl-2, and calbindin D28k [42]. Cultures of rat hippocampal cells and sympathetic neurons subjected to excitotoxic, or metabolic stress demonstrated increased survival, attributed to the neuroprotective effects of upregulated NF-κB and induction of MnSOD [43]. NF-κB also induces expression of cellular inhibitors of apoptosis (cIAPs), Bcl-2s, TRAF1/TRAF2, and may decrease expression of apoptosis-promoting cytokines such as TNFα and FAS ligand (FASL) [44,45,46,47,48].
Neural Progenitor Cells (NCP)
Neural progenitor cells or neural cell precursors (NCP) are generated from autologous peripheral blood by culturing the cells in medium supplemented with autologous serum, followed by activation in a defined medium containing specific differentiation-inducing factors. The cells acquire a neural phenotype characterized by development of neuroglial morphology with neurite-like extensions and expression of neural markers (Nestin, Beta-tubulin III, O4 and glial fibrillary acidic protein (GFAP), secrete neurotrophic factors including brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), glial cell-derived neurotrophic factor (GDNF), Macrophage Colony Stimulating Factor (M-CSF) and Stem Cell factor (SCF). Robust expression of neurotrophic factors: BDNF, GDNF, SCF and M-CSF is neuroprotective, promoting myelin formation, enhancing axon survival through interaction with the tyrosine kinases, p75 Neurotrophic factor, and NF-κB [49]. Preliminary results indicate that NCPs migrate to the place of injury and engraft in vivo in animal models of stroke and laser injury to the retina.
Nerve Growth Factor Induces NF-κB
NCP express high levels of NGF. NGF induction of NF-κB is essential to the normal development, function, and maintenance of cells in the nervous system. Activation of NF-κB by NGF is well characterized, mediating its effects through activation of p75 NTR or by tyrosine receptor kinase A (TrkA). NGF binding to TrkA generates signals leading to neuronal cell survival, differentiation and neurite process formation. Neurotrophic factors NGF, BDNF, NT-3, NT-4/5, and NT-6 promote neuron survival and differentiation by binding to TrkA, TrkB, and TrkC, which in turn upregulate the ERK/MAPKs. p75NTR expression in the presence of a neurotrophin may lead to programmed cell death through the caspase and a JNK pathways. However, when p75NTR is expressed together with the TrkA receptor, it facilitates neurotrophic function by increasing the affinity of NGF binding to the TrkA receptor [50,51], and the aggregate of NF-κB signal generated from both receptors exerts neuroprotective effects [35].
NSC May Mitigate the Pathophysiological Effects of CNS Injury Through Modulation of Expression of the N-Methyl D-Aspartate (NMDA) Receptors
N-Methyl-d-Aspartate (NMDA) –type receptors are ligand-gated ion channels which mediate excitatory transmission. Comprised of a diverse array of tetrameric receptor complexes that exhibit regional specificity, NMDA receptors exhibit distinct physiological roles across different neuronal cell types and brain regions. Sequence variants of NMDA receptors are associated with autistic spectrum disorder, developmental delay and intellectual disability, epilepsy, alterations of muscle, language problems, and other phenotypic manifestations [52]. NMDA receptors are pathologically upregulated in central nervous system injury, variably resulting in neural supersensitization, behavioral change, epilepsy or apoptosis. Glutamate binding to NMDA in the setting of CNS injury results in Ca2+ influx and direct excitotoxicity, release of apoptosis inducing factor (AIF) and cell death. NCP downregulate NMDA receptors, decreasing excitotoxicity and central sensitization [53]. The downregulation of NMDA receptors may be of importance in subjects with traumatic brain injury or post-traumatic stress disorder.
The CXCR4 /CXCL12 Axis Promotes ACP and NCP Migration Toward Areas of Hypoxic Injury and Stress
Chemokine CXCL12, also known as Stromal derived factor 1(SDF-1), is released in areas of CNS injury. CXCL12 is up-regulated in neurons and glial cells following injury, and induces migration and activation of hematopoietic progenitor cells (HPC)s and endothelial cells bearing a CXCR4 receptor [20]. ACP highly express the CXCR4 receptor, and migrate toward higher concentrations of CXCL12, repopulating injured CNS tissue [54]. Chemokine CXCL12 gene therapy has also been shown to promote angiogenesis, neurogenesis, and remyelination, attracting endothelial progenitor cells (EPCs), neural progenitor cells, and oligodendrocyte progenitor cells (OPCs) to the injured sites of the brain [55]. ACP secrete tissue regeneration factors such as CXCL8, vascular endothelial growth factor (VEGF) and angiogenin, promoting angiogenesis and tissue repair and preventing inflammation and apoptosis. CXCL8 recruits peripheral, perivascular CD34+ cells, which promote robust angiogenesis [10] (Figure 2). ACP-01 (Hemostemix Corp.) is a heterogenous population of cell types, including a large subpopulation of CD34+ cells.
NCP highly express the CXCR4 receptor, and migrate toward higher concentrations of CXCL12, repopulating injured CNS tissue [54]. NCP are specifically programmed to differentiate into neurons, astrocytes and oligodendrocytes, and have demonstrated meaningful repair of injured brain. Exogenous stem cell transplantation has been shown to accelerate immature neuronal development in damaged brain regions [56], and to engraft, differentiate into oligodendrocytes, and contribute to re-myelination of damaged axonal functions, facilitating recovery after spinal cord injury [57,58].
ACP Expression of IL-8 (CXCL8) Activates NF-κB – The “Learning Molecule”. NF-κB Is Essential to Synaptogenesis, Neuritogenesis, and Learning
CXCL8 is highly expressed by ACP [10] and causes downstream translocation of cytosolic NF-κB to the nucleus, a process of seminal importance in all processes of neural plasticity required for long-term memory (LTM) storage. NF-κB is ubiquitous in the cytosol of neurons in the cortex, amygdala and hippocampus. Constitutively present in neurite projections, and essential for synaptic plasticity, NF-κB proteins alone are not sufficient to induce neurite process formation [35], but nevertheless play a central role in neurite outgrowth, plasticity and neural cell longevity. In the process of memory formation, NF-κB is activated by synaptic activity, and acts as a messenger from synapse to nucleus [59,60].
Kandel laid the underpinnings of the molecular basis of the storage of learned information and memory consolidation, a process involving hippocampus-dependent molecular processes, which includes gene transcription [61]. Synaptic plasticity is the primary cellular mechanism for storage of LTM in the nervous system. Long-lasting neuronal plasticity and LTM require specific gene transcription and the de novo synthesis of m-RNA and protein [59]. Synaptic plasticity is regulated by the dynamic balance of excitatory and inhibitory neurotransmissions. Stimulus driven post-synaptic glutamate results in progressive postsynaptic depolarization. Opposing the excitability is the release of GABA by inhibitory GABAergic neurons, which affect hyperpolarization of the postsynaptic neuron. Stimulus-coupled changes in synaptic plasticity are required for the storage, retrieval, and removal of memory formation, the product of modification of synaptic substrate and new proteins, as regulated by excitatory and inhibitory neurotransmitters and specific transcription kinases [62].
Various families of transcription factors have been explored in the context of forming memory: Cyclic AMP response Element Binding Protein (CREB), Activating Protein 1 transcription factor (AP-1), CCAAT/Enhancing Binding Protein (C/EBP), and Early Growth Response transcription factor (EGR). However, NF-κB responsive genes play a critical role in learning and memory and are essential to all learning-associated neuronal plasticity, long term potentiation (LTP) and long-term depression (LTD), remodulation of dendritic spine and formation of new synapses (synaptogenesis), and outgrowth of axons and dendrites (neuritogenesis) [63,64,65,66,67,68,69,70,71,72,73,74,75,76,77]. Murine models of genetic disruption of NF-κB have demonstrated that NF-κB transcription factor is critical for synaptic transmission, LTP and induction of LTD. Pretreatment with NF-κB DNA element decoy interferes with the induction of both LTP and LTD [72].
Synaptic activity regulates NF-κB subcellular distribution, DNA binding activity, and transcription [66]. The list of signals activating NF-κB includes the excitatory neurotransmitter glutamate, depolarization, CXCL8, NGF, proinflammatory cytokines interleukin-1 and TNFα, kainate, amyloid β peptide, brain injury, and oxidative stress [31,78,79,80]. LTP induced by high frequency stimulation (HFS) or low-frequency stimulation (LFS) of hippocampal slices appears to increase mRNA of p50 and p65, and to decrease mRNA of the inhibitory subunit IkB. Increased active form of NF-κB is observed in hippocampal granular and pyramidal cells after LFS and HFS [72].
Transcription factor NF-κB, in its role of recruiting histone acetyltransferases (HAT)s and Histone deacetylase (HDAC) enzymes at the regulatory regions of target genes, plays a pivotal role in consolidation and persistence of object recognition memory [81]. The strength of a memory is related to the level of motivation and training, and the epigenetic mechanisms by which its gene expression is subjected. The NF-κB/Rel family helps to modulate the contested terrain of inhibitory interneuron function versus excitatory neuronal function through its effect on GAD65. Loss of neuronal NF-κB results in hyperexcitability and enhanced LTP [62]. Histone acetylation by HATs makes DNA accessible to the apparatus of transcription; and histone acetylation, methylation or phosphorylation are involved in long-term plasticity and memory consolidation [81,82]. Inhibition of NF-κB prevents memory consolidation [83].
The genes induced by NF-κB in the context of neuroplasticity and memory can be categorized into several fundamental groups, including cell signaling, cell adhesion, growth, cellular trafficking, neurotransmission, redox signaling, and transcription/ translation [59]. NF-κB epigenetically regulates memory consolidation by targeting the transcription factor early growth response-1 (EGR-1) (a.k.a. Zif268, also known as Ngfi-A, Krox 24, Tis 8, and Zenk 2d). NF-κB regulates the Zif268 promoter [84,85]. NF-κB upregulates EGR-1 at the promoter region in the hippocampus, resulting in increased acetylation of histone H3 [59].
CaMKII, an abundant synaptic signaling molecule essential for memory formation and metaplasticity, is only induced during consolidation of a persistent form of memory. CaMKII has four isoforms, of which the delta isoform is regulated by NF-κB after strong training [86]. In particular, the p65 monomer of NF-κB regulates histone acetylation during strong memory consolidation, achieving high levels of H3 acetylation in the NF-κB regulatory region of CaMKIIδ promoter region. Inhibition of NF-κB dependent histone acetylation impairs persistent memory, and HDAC inhibition renders memory more persistent. Moreover, weak memories due to weak training, which is unable to generate sufficient gene transcription, can be enhanced by HDAC inhibitors, and transformed into a stronger memory [81].
NF-κB family members are preformed proteins, the activation and activity of which are regulated to large extent by post-translational modification, as opposed to induction of their synthesis. Acetylation regulates the p65 subunits, p105, and p100 of NF-κB, but there is also regulation from S-nitrosylation, oxidation, nitration, and alkylation, and O-GlcNAcetylation [87,88]. NF-κB activation is site specific; acetylation at one site (Lys-310) enhances transcriptional activity of p65, where acetylation at Lys-122 and 123 inhibits P65 DNA binding and transcriptional activity [59].
Canonical and Non-Canonical Activation of NF-κB Factors
The NF-κB signaling pathway regulates transcription in the hippocampus during memory reconsolidation. NF-κB resides ubiquitously in the cytosol as an inactive form of three protein subunits, a transcription factor dimer and an inhibitory subunit, IκB. In neurons, the most common combination of subunit is p65 (also referred to as RelA), P50 and IκBα. However, other protein monomers (RelB, cRel, and p52) form homodimers or heterodimers which affect genetic transcription in response to specific stimuli [89].
Tightly regulated by either the canonical- NEMO (NF-κB Essential Modulator) dependent pathway - or the non-canonical, NEMO-independent pathway, the transcriptionally active NF-κB dimers are the levers of control for various programs of genetic expression, thus controlling broad gene expression programs. [90]. In the canonical mechanism of NF-κB activation, phosphorylation of the inhibitory IκB subunit by IκB Kinases (IKK) results in ubiquitination and proteasomal degradation of the IκB subunit, and release of the active form of NF-κB factor dimer to translocate to the nucleus (Figure 3). In the non-canonical control mechanism, IKKα directly modifies chromatin structure by the phosphorylation of histone H3, triggering acetylation of histone H3 by interacting with the CREB Binding Protein (CBP) [85,91].
NF-κB Signaling Pathway Regulates Transcription in Memory Re-Consolidation
The period of memory recall is one in which memories are susceptible to alteration and requires a process of reconsolidation after retrieval [94,95]. To enable memory reconsolidation, IKKα activity is necessary to regulate gene transcription and chromatin structure in hippocampus. Retrieval of contextual conditioned fear memories activates the NF-κB pathway via IKKα, and subsequent up-regulation in hippocampal NF-κB signaling activity increases histone H3 phosphorylation and acetylation. And as anticipated, IKK inhibition blocks regulation of both chromatin structure and NF-κB DNA binding during memory reconsolidation; this inhibition of the NF-κB signaling cascade at the level of the IKKα protein kinase blocks contextual conditioned fear memory reconsolidation and histone H3 posttranslational modifications in the hippocampus after memory retrieval. Conversely, elevating histone acetylation rescues this memory deficit in the face of IKK blockade [85,96,97,98].
The importance of NF-κB has been demonstrated in both invertebrate and vertebrate species. Inhibition of NF-κB in the hippocampus disrupts memory [91]. In the crab Chasmagnathus, habituation of the escape response elicited by a fear stimulus is paralleled by increased activated form of an NF-κB homolog in brain nuclei and in isolated synapses [63,99]. On the other hand, blocking NF-κB activation with IKK complex inhibitor prevents formation of escape memory, [100] and blocking NF-κB after memory recall results in amnesia [95]. In murine models, intrahippocampal injections of NF-κB decoy disrupts the reconsolidation of memory [83]. NF-κB is needed for radial maze mastery [101]. Intra-amygdala administration of NF-κB decoy disrupts the fear-potentiated startle response [102]. Inhibition of NF-κB function by overexpression of IkB in the mouse forebrain is associated with decreased mRNA of the catalytic subunit of protein kinase A (PKA), decreased CREB phosphorylation and decreased spatial memory [72].
Information Storage Is a Function of Synaptogenesis
The transcription factor NF-κB is centrally-involved in the upregulation of dendritic spines and synaptic density. NF-κB is necessary in the regulation of signals which induce neurite outgrowth during synaptic plasticity and memory. NF-κB influences the morphology and complexity of the dendritic arbors through regulation of NCAM, amyloid precursor protein (APP), Tenascin-C, and β1 integrin [59]. Moreover, the p65 subunit of NF-κB is enriched in dendritic spines, where it regulates density, size and function of dendritic spines and excitatory synapses, and provides feedback to control spine density and spine morphology. During the learning process, plasticity is manifest by a substantial increase in NF-κB over basal level in a number of dendritic spines and excitatory synapses [103] p65-mediated regulation of spine density absolutely depends on the ability of p65 to bind DNA, and to activate transcription of target genes. Altered strength and connectivity of synapses and dendritic spine morphology underlies neural plasticity [104,105,106,107]. Structural plasticity also involves change of dendrite head size. p65 increases dendrite head size. Larger spine heads and larger synapses correlate with increased numbers of AMPA receptors and increased AMPA receptor-mediated currents. Conversely, loss of NF-κB function leads to a decrease in dendritic spine density, and diminished spine head size [108,109].
NF-κB also regulates BDNF, which is important in energy metabolism, neuronal differentiation, survival, growth, neurogenesis, and synaptic plasticity and memory. In astrocytes and neurons, upregulated BDNF is profoundly manifested in terms of memory consolidation, cellular growth, differentiation, and neurogenesis [110].
Thus, the high levels of CXCR8 expressed by ACP activate NF-κB, and this effect is of profound importance to the potential for learning and memory storage.
NSC Favor the Non-Inflammatory M2 Phenotype, Resulting in Less Inflammation and Less Scarring
The grafting of human NSCs intracerebrally attenuates microglial/macrophage activation, favoring transition of the macrophage toward the M2 anti-inflammatory phenotype. Adoption of the M2 phenotype promotes axonal regrowth in the setting of traumatic injury [111]. Ubiquitously distributed throughout the CNS, microglia depending on their activation status can be pro-inflammatory and cytotoxic -the M1 “classical” phenotype- or conversely the neuroprotective, M2 “alternative” phenotype. Oxidative stress, low-grade inflammation, injury and neurotoxin exposure result in microglial priming to the inflammatory and harmful macrophage M1 phenotype. Pathogenic particles from damaged cells trigger release of pro-inflammatory cytokines IL-1β, TNF-α, IL-6, nitrous oxide, ROS, and reactive nitrogen species (RNS) reciprocally amplifying microglial activation, and resulting in neuron vulnerability and neuronal death [112,113,114]. This inflammatory response may be amplified through recruitment of additional cells by pro-inflammatory cytokines- TNF-α, IL-1β, IL-16 and IL-18 [115,116]. The M1 and M2 phenotypes represent a continuum, and microglia may move from one state to the other [117]. In contradistinction to the pro-inflammatory M1 phenotype that utilizes anaerobic glycolysis, the M2 macrophages have an intact tricarboxylic acid (TCA) cycle, expressing anti-inflammatory mediators, clearing apoptotic cells, promoting tissue repair, and driving angiogenesis. NSC signal preferential adoption of the M2 phenotype, thus promoting neuroprotection and tissue healing, and decreasing deleterious inflammatory changes and scarring in areas of central nervous system injury (Figure 4) [118,119]. Though neuroinflammation serves as a defense mechanism to protect the brain by removing or inhibiting diverse pathogens, sustained inflammation inhibits regeneration [120]. NSC thus dampen the inflammatory changes that occur in the first 24 hours after injury and lessen scarring by promoting the adoption of the M2 phenotype of microglia and macrophages. M2 microglia aid phagocytosis of cell debris and misfolded proteins, promote extracellular matrix reconstruction and tissue repair, and support neuron survival by release of neurotrophic factors [115]. The M2 phenotypical macrophages release anti-inflammatory cytokines- IL-4, IL-10, IL-13, transforming growth factor (TGF)-β), insulin-like growth factor-1 (IGF-1), fibroblast growth factor (FGF), colony stimulating factor (CSF)1, and neurotrophic growth factors (NGF)s- BDNF, neurotrophins and GDNF, inducing a condition of neuroprotectivity [121].
While microglia originate from infiltrated yolk sac progenitor cells during early embryonic development, and are maintained independently by self-proliferation, they are also maintained in number by circulating monocytes under disease conditions [122]. Within 24 hours of stroke, monocytes from the peripheral circulation adhere to damaged endothelium of the BBB and infiltrate the CNS to replenish the macrophage count [123].
Conclusions
The application of artificial intelligence through the brain computer interface is proving to be one of the great advances in neuroscience today. However, the introduction of foreign bodies into deeper parts of the central nervous system will ultimately result in foreign body reaction, scarring, apoptosis, and decreased signaling. One probable solution to these significant obstacles is the biomolecular strategy provided by the delivery of ACP and NCP, either intrathecally or intra-arterially, and the important consequences of which may be improved cell survival, angiogenesis, decreased inflammation and scarring, and improved learning and memory storage.
Author Contributions
Conceptualization: FH; Writing-original draft preparation FH; Writing-review & editing: FH, KT; Resources: FH, KT; Visualization: KT.
Funding
this research received no external funding.
Conflicts of Interest
Professor Henderson is a practicing, academic neurosurgeon who serves as Chief Medical Officer, and has stock in Hemostemix, Inc. Kelly Tuchman was paid for her time by Metropolitan Neurosurgery Group, and has no conflict of interest. Potential conflicts of interest have been transparently disclosed, and do not undermine the scientific validity of the work.
References
- Maiseli, B.; Abdalla, A.T.; Massawe, L.V.; Mbise, M.; Mkocha, K.; Nassor, N.A.; Ismail, M.; Michael, J.; Kimambo, S. Brain-computer interface: trend, challenges, and threats. Brain Inform. 2023, 10, 20. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kubben, P. Invasive Brain-Computer Interfaces: A Critical Assessment of Current Developments and Future Prospects. JMIR Neurotech 2024, 3, e60151. [Google Scholar] [CrossRef]
- Bedell, H.W.; Capadona, J.R. Anti-inflammatory Approaches to Mitigate the Neuroinflammatory Response to Brain-Dwelling Intracortical Microelectrodes. J Immunol Sci. 2018, 2, 15–21. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Raposo, C.; Graubardt, N.; Cohen, M.; Eitan, C.; London, A.; Berkutzki, T.; Schwartz, M. CNS repair requires both effector and regulatory T cells with distinct temporal and spatial profiles. J Neurosci. 2014, 34, 10141–10155. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Polikov, V.S.; Tresco, P.A.; Reichert, W.M. Response of brain tissue to chronically implanted neural electrodes. J Neurosci Methods. 2005, 148, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Griffith, R.W.; Humphrey, D.R. Long-term gliosis around chronically implanted platinum electrodes in the Rhesus macaque motor cortex. Neurosci Lett. 2006, 406, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Filbin, M.T. Myelin-associated inhibitors of axonal regeneration in the adult mammalian CNS. Nat Rev Neurosci. 2003, 4, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Schubart, J.R.; Zare, A.; Fernandez-de-Castro, R.M.; Figueroa, H.R.; Sarel, I.; Tuchman, K.; Esposito, K.; Henderson, F.C.; von Schwarz, E. Safety and outcomes analysis: transcatheter implantation of autologous angiogenic cell precursors for the treatment of cardiomyopathy. Stem Cell Res Ther. 2023, 14, 308. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Henderson, F.C.; Sarel, I.; Tuchman, K.; Lewis, S.; Hsiang, Y. Angiogenic Precursor Cell Treatment of Critical Limb Ischemia Decreases Ulcer Size, Amputation and Death Rate: Re-Examination of phase II ACP NO-CLI Trial Data. J Biomed Res Environ Sci. 2024, 5, 092–105. [Google Scholar] [CrossRef]
- Henderson, F.C.; Tuchman, K.; Sarel, I. Autologous Angiogenic Cell Precursors- A Molecular Strategy for the Treatment of Heart Failure: Response to Biocardia’s Cardiamp HF Trial. J Biomed Res Environ Sci. 12 May. [CrossRef]
- Knorr, M.; Münzel, T.; Wenzel, P. Interplay of NK cells and monocytes in vascular inflammation and myocardial infarction. Front Physiol. 2014, 5, 295. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ong, S.; Rose, N.R.; Čiháková, D. Natural killer cells in inflammatory heart disease. Clin Immunol. 2017, 175, 26–33. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ong, S.; Ligons, D.L.; Barin, J.G.; Wu, L.; Talor, M.V.; Diny, N.; Fontes, J.A.; Gebremariam, E.; Kass, D.A.; Rose, N.R.; Čiháková, D. Natural killer cells limit cardiac inflammation and fibrosis by halting eosinophil infiltration. Am J Pathol. 2015, 185, 847–861. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Boukouaci, W.; Lauden, L.; Siewiera, J.; Dam, N.; Hocine, H.R.; Khaznadar, Z.; Tamouza, R.; Borlado, L.R.; Charron, D.; Jabrane-Ferrat, N.; Al-Daccak, R. Natural killer cell crosstalk with allogeneic human cardiac-derived stem/progenitor cells controls persistence. Cardiovasc Res. 2014, 104, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Vosshenrich, C.A.; Di Santo, J.P. Developmental programming of natural killer and innate lymphoid cells. Curr Opin Immunol. 2013, 25, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Zhou, R.; Li, T.; Hua, Y.; Zhou, K.; Li, Y.; Luo, S.; An, Q. The Molecular Role of Immune Cells in Dilated Cardiomyopathy. Medicina (Kaunas). 2023, 59, 1246. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Russo, R.C.; Garcia, C.C.; Teixeira, M.M.; Amaral, F.A. The CXCL8/IL-8 chemokine family and its receptors in inflammatory diseases. Expert Rev Clin Immunol. 2014, 10, 593–619. [Google Scholar] [CrossRef] [PubMed]
- Vujanovic, L.; Ballard, W.; Thorne, S.H.; Vujanovic, N.L.; Butterfield, L.H. Adenovirus-engineered human dendritic cells induce natural killer cell chemotaxis via CXCL8/IL-8 and CXCL10/IP-10. Oncoimmunology. 2012, 1, 448–457. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Walle, T.; Kraske, J.A.; Liao, B.; Lenoir, B.; Timke, C.; von Bohlen Und Halbach, E.; Tran, F.; Griebel, P.; Albrecht, D.; Ahmed, A.; Suarez-Carmona, M.; Jiménez-Sánchez, A.; Beikert, T.; Tietz-Dahlfuß, A.; Menevse, A.N.; Schmidt, G.; Brom, M.; Pahl, J.H.W.; Antonopoulos, W.; Miller, M.; Perez, R.L.; Bestvater, F.; Giese, N.A.; Beckhove, P.; Rosenstiel, P.; Jäger, D.; Strobel, O.; Pe’er, D.; Halama, N.; Debus, J.; Cerwenka, A.; Huber, P.E. Radiotherapy orchestrates natural killer cell dependent antitumor immune responses through CXCL8. Sci Adv. 2022, 8, eabh4050. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cambier, S.; Gouwy, M.; Proost, P. The chemokines CXCL8 and CXCL12: molecular and functional properties, role in disease and efforts towards pharmacological intervention. Cell Mol Immunol. 2023, 20, 217–251. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nurmi, A.; Lindsberg, P.J.; Koistinaho, M.; Zhang, W.; Juettler, E.; Karjalainen-Lindsberg, M.L.; Weih, F.; Frank, N.; Schwaninger, M.; Koistinaho, J. Nuclear factor-kappaB contributes to infarction after permanent focal ischemia. Stroke. 2004, 35, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z. H. , Tao L. Y., Chen X. (2007). Dual roles of NF-kappaB in cell survival and implications of NF-kappaB inhibitors in neuroprotective therapy. Acta Pharmacol. Sin. 28 1859–1872. 10.1111/j.1745-7254.2007.00741.x.
- Liu, H.; Ma, Y.; Pagliari, L.J.; Perlman, H.; Yu, C.; Lin, A.; Pope, R.M. TNF-alpha-induced apoptosis of macrophages following inhibition of NF-kappa B: a central role for disruption of mitochondria. J Immunol. 2004, 172, 1907–1915. [Google Scholar] [CrossRef] [PubMed]
- Pazarentzos, E.; Mahul-Mellier, A.L.; Datler, C.; Chaisaklert, W.; Hwang, M.S.; Kroon, J.; Qize, D.; Osborne, F.; Al-Rubaish, A.; Al-Ali, A.; Mazarakis, N.D.; Aboagye, E.O.; Grimm, S. IκΒα inhibits apoptosis at the outer mitochondrial membrane independently of NF-κB retention. EMBO J. 2014, 33, 2814–2828. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Luo, J.L.; Kamata, H.; Karin, M. The anti-death machinery in IKK/NF-kappaB signaling. J Clin Immunol. 2005, 25, 541–550. [Google Scholar] [CrossRef] [PubMed]
- May, M.J.; Ghosh, S. IkappaB kinases: kinsmen with different crafts. Science. 1999, 284, 271–273. [Google Scholar] [CrossRef] [PubMed]
- Barkett, M.; Gilmore, T.D. Control of apoptosis by Rel/NF-kappaB transcription factors. Oncogene. 1999, 18, 6910–6924. [Google Scholar] [CrossRef] [PubMed]
- Maggirwar, S.B.; Sarmiere, P.D.; Dewhurst, S.; Freeman, R.S. Nerve growth factor-dependent activation of NF-kappaB contributes to survival of sympathetic neurons. J Neurosci. 1998, 18, 10356–10365. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hamanoue, M.; Middleton, G.; Wyatt, S.; Jaffray, E.; Hay, R.T.; Davies, A.M. p75-mediated NF-kappaB activation enhances the survival response of developing sensory neurons to nerve growth factor. Mol Cell Neurosci. 1999, 14, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Beg, A.A.; Sha, W.C.; Bronson, R.T.; Ghosh, S.; Baltimore, D. Embryonic lethality and liver degeneration in mice lacking the RelA component of NF-kappa B. Nature. 1995, 376, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Baldwin AS, Jr. The NF-kappa B and I kappa B proteins: new discoveries and insights. Annu Rev Immunol. 1996, 14, 649–683. [Google Scholar] [CrossRef] [PubMed]
- Beg, A.A.; Baltimore, D. An essential role for NF-kappaB in preventing TNF-alpha-induced cell death. Science. 1996, 274, 782–784. [Google Scholar] [CrossRef] [PubMed]
- Van Antwerp, D.J.; Martin, S.J.; Kafri, T.; Green, D.R.; Verma, I.M. Suppression of TNF-alpha-induced apoptosis by NF-kappaB. Science. 1996, 274, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Van Antwerp, D.; Mercurio, F.; Lee, K.F.; Verma, I.M. Severe liver degeneration in mice lacking the IkappaB kinase 2 gene. Science. 1999, 284, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Foehr, E.D.; Lin, X.; O’Mahony, A.; Geleziunas, R.; Bradshaw, R.A.; Greene, W.C. NF-kappa B signaling promotes both cell survival and neurite process formation in nerve growth factor-stimulated PC12 cells. J Neurosci. 2000, 20, 7556–7563. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sompol, P.; Xu, Y.; Ittarat, W.; Daosukho, C.; Clair, D.S. NF-κB-Associated MnSOD induction protects against β-amyloid-induced neuronal apoptosis. J Mol Neurosci. 2006, 29, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.F.; Guo, F.; Cao, Y.Z.; Shi, W.; Xia, Q. Neuroprotection by manganese superoxide dismutase (MnSOD) mimics: antioxidant effect and oxidative stress regulation in acute experimental stroke. CNS Neurosci Ther. 2012, 18, 811–818. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Massaad, C.A.; Klann, E. Reactive oxygen species in the regulation of synaptic plasticity and memory. Antioxid Redox Signal. 2011, 14, 2013–2054. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wu, M.; Lee, H.; Bellas, R.E.; Schauer, S.L.; Arsura, M.; Katz, D.; FitzGerald, M.J.; Rothstein, T.L.; Sherr, D.H.; Sonenshein, G.E. Inhibition of NF-kappaB/Rel induces apoptosis of murine B cells. EMBO J. 1996, 15, 4682–4690. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xia, Y.; Rao, J.; Yao, A.; Zhang, F.; Li, G.; Wang, X.; Lu, L. Lithium exacerbates hepatic ischemia/reperfusion injury by inhibiting GSK-3β/NF-κB-mediated protective signaling in mice. Eur J Pharmacol. 2012, 697, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Kiningham, K.K.; Xu, Y.; Daosukho, C.; Popova, B.; St Clair, D.K. Nuclear factor kappaB-dependent mechanisms coordinate the synergistic effect of PMA and cytokines on the induction of superoxide dismutase 2. Biochem J. 2001, 353, 147–156. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pizzi, M.; Goffi, F.; Boroni, F.; Benarese, M.; Perkins, S.E.; Liou, H.C.; Spano, P. Opposing roles for NF-κB/Rel factors p65 and c-Rel in the modulation of neuron survival elicited by glutamate and interleukin-1beta. J. Biol. Chem. 2002, 277, 20717–20723. [Google Scholar] [CrossRef]
- Mattson, M.P.; Goodman, Y.; Luo, H.; Fu, W.; Furukawa, K. Activation of NF-kappaB protects hippocampal neurons against oxidative stress-induced apoptosis: evidence for induction of manganese superoxide dismutase and suppression of peroxynitrite production and protein tyrosine nitration. J Neurosci Res. 1997, 49, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Christakos, S.; Mattson, M.P. Tumor necrosis factors protect neurons against metabolic-excitotoxic insults and promote maintenance of calcium homeostasis. Neuron. 1994, 12, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Tamatani, M.; Che, Y.H.; Matsuzaki, H.; Ogawa, S.; Okado, H.; Miyake, S.; Mizuno, T.; Tohyama, M. Tumor necrosis factor induces Bcl-2 and Bcl-x expression through NFkappaB activation in primary hippocampal neurons. J Biol Chem. 1999, 274, 8531–8538. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, L.; Klein, M.; Schlett, K.; Pfizenmaier, K.; Eisel, U.L. Tumor necrosis factor (TNF)-mediated neuroprotection against glutamate-induced excitotoxicity is enhanced by N-methyl-D-aspartate receptor activation. Essential role of a TNF receptor 2-mediated phosphatidylinositol 3-kinase-dependent NF-kappa B pathway. J Biol Chem. 2004, 279, 32869–32881. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Culmsee, C.; Klumpp, S.; Krieglstein, J. Neuroprotection by transforming growth factor-beta1 involves activation of nuclear factor-kappaB through phosphatidylinositol-3-OH kinase/Akt and mitogen-activated protein kinase-extracellular-signal regulated kinase1,2 signaling pathways. Neuroscience. 2004, 123, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Kovács, A.D.; Chakraborty-Sett, S.; Ramirez, S.H.; Sniderhan, L.F.; Williamson, A.L.; Maggirwar, S.B. Mechanism of NF-kappaB inactivation induced by survival signal withdrawal in cerebellar granule neurons. Eur J Neurosci. 2004, 20, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.R.; Miller, F.D. Signal transduction by the neurotrophin receptors. Curr Opin Cell Biol. 1997, 9, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, M.; Levi, A.; Chao, M.V. Differential expression of nerve growth factor receptors leads to altered binding affinity and neurotrophin responsiveness. Proc Natl Acad Sci U S A. 1993, 90, 7859–7863. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Barker, P.A.; Shooter, E.M. Disruption of NGF binding to the low affinity neurotrophin receptor p75LNTR reduces NGF binding to TrkA on PC12 cells. Neuron. 1994, 13, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, J.; Tang, W.; Mizu, R.K.; Kusumoto, H.; XiangWei, W.; Xu, Y.; Chen, W.; Amin, J.B.; Hu, C.; Kannan, V.; Keller, S.R.; Wilcox, W.R.; Lemke, J.R.; Myers, S.J.; Swanger, S.A.; Wollmuth, L.P.; Petrovski, S.; Traynelis, S.F.; Yuan, H. De novo GRIN variants in NMDA receptor M2 channel pore-forming loop are associated with neurological diseases. Hum Mutat. 2019, 40, 2393–2413. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, H.; Yu, S.W.; Koh, D.W.; Lew, J.; Coombs, C.; Bowers, W.; Federoff, H.J.; Poirier, G.G.; Dawson, T.M.; Dawson, V.L. Apoptosis-inducing factor substitutes for caspase executioners in NMDA-triggered excitotoxic neuronal death. J Neurosci. 2004, 24, 10963–10973. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lee, I.S.; Jung, K.; Kim, M.; Park, K.I. Neural stem cells: properties and therapeutic potentials for hypoxic-ischemic brain injury in newborn infants. Pediatr Int. 2010, 52, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chang, S.; Li, W.; Tang, G.; Ma, Y.; Liu, Y.; Yuan, F.; Zhang, Z.; Yang, G.Y.; Wang, Y. cxcl12-engineered endothelial progenitor cells enhance neurogenesis and angiogenesis after ischemic brain injury in mice. Stem Cell Res Ther. 2018, 9, 139. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tajiri, N.; Kaneko, Y.; Shinozuka, K.; Ishikawa, H.; Yankee, E.; McGrogan, M.; Case, C.; Borlongan, C.V. Stem cell recruitment of newly formed host cells via a successful seduction? Filling the gap between neurogenic niche and injured brain site. PLoS One. 2013, 8, e74857. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ben-Hur, T. Reconstructing neural circuits using transplanted neural stem cells in the injured spinal cord. J Clin Invest. 2010, 120, 3096–3098. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fandel, T.M.; Trivedi, A.; Nicholas, C.R.; Zhang, H.; Chen, J.; Martinez, A.F.; Noble-Haeusslein, L.J.; Kriegstein, A.R. Transplanted Human Stem Cell-Derived Interneuron Precursors Mitigate Mouse Bladder Dysfunction and Central Neuropathic Pain after Spinal Cord Injury. Cell Stem Cell. 2016, 19, 544–557. [Google Scholar] [CrossRef] [PubMed]
- Snow, W.M.; Stoesz, B.M.; Kelly, D.M.; Albensi, B.C. Roles for NF-κB and gene targets of NF-κB in synaptic plasticity, memory, and navigation. Mol Neurobiol. 2014, 49, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Lubin, F.D.; Johnston, L.D.; Sweatt, J.D.; Anderson, A.E. Kainate mediates nuclear factor-kappa B activation in hippocampus via phosphatidylinositol-3 kinase and extracellular signal-regulated protein kinase. Neuroscience. 2005, 133, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Kandel, E.R. The molecular biology of memory storage: a dialog between genes and synapses. Biosci Rep. 2004, 24, 475–522. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, A.; Raber, J.; Montano, M.; Foehr, E.; Han, V.; Lu, S.M.; Kwon, H.; LeFevour, A.; Chakraborty-Sett, S.; Greene, W.C. NF-kappaB/Rel regulates inhibitory and excitatory neuronal function and synaptic plasticity. Mol Cell Biol. 2006, 26, 7283–7298. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Freudenthal, R.; Romano, A. Participation of Rel/NF-kappaB transcription factors in long-term memory in the crab Chasmagnathus. Brain Res. 2000, 855, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Meberg, P.J.; Kinney, W.R.; Valcourt, E.G.; Routtenberg, A. Gene expression of the transcription factor NF-kappa B in hippocampus: regulation by synaptic activity. Brain Res Mol Brain Res. 1996, 38, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Kassed, C.A.; Willing, A.E.; Garbuzova-Davis, S.; Sanberg, P.R.; Pennypacker, K.R. Lack of NF-kappaB p50 exacerbates degeneration of hippocampal neurons after chemical exposure and impairs learning. Exp Neurol. 2002, 176, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Freudenthal, R.; Romano, A.; Routtenberg, A. Transcription factor NF-kappaB activation after in vivo perforant path LTP in mouse hippocampus. Hippocampus. 2004, 14, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Albensi, B.C.; Mattson, M.P. Evidence for the involvement of TNF and NF-kappaB in hippocampal synaptic plasticity. Synapse. 2000, 35, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Denis-Donini, S.; Dellarole, A.; Crociara, P.; Francese, M.T.; Bortolotto, V.; Quadrato, G.; Canonico, P.L.; Orsetti, M.; Ghi, P.; Memo, M.; Bonini, S.A.; Ferrari-Toninelli, G.; Grilli, M. Impaired adult neurogenesis associated with short-term memory defects in NF-kappaB p50-deficient mice. J Neurosci. 2008, 28, 3911–3919. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- O’Riordan, K.J.; Huang, I.C.; Pizzi, M.; Spano, P.; Boroni, F.; Egli, R.; Desai, P.; Fitch, O.; Malone, L.; Ahn, H.J.; Liou, H.C.; Sweatt, J.D.; Levenson, J.M. Regulation of nuclear factor kappaB in the hippocampus by group I metabotropic glutamate receptors. J Neurosci. 2006, 26, 4870–4879. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ahn, H.J.; Hernandez, C.M.; Levenson, J.M.; Lubin, F.D.; Liou, H.C.; Sweatt, J.D. c-Rel, an NF-kappaB family transcription factor, is required for hippocampal long-term synaptic plasticity and memory formation. Learn Mem. 2008, 15, 539–549. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- O’Sullivan, N.C.; Croydon, L.; McGettigan, P.A.; Pickering, M.; Murphy, K.J. Hippocampal region-specific regulation of NF-kappaB may contribute to learning-associated synaptic reorganisation. Brain Res Bull. 2010, 81, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Kaltschmidt, B.; Ndiaye, D.; Korte, M.; Pothion, S.; Arbibe, L.; Prüllage, M.; Pfeiffer, J.; Lindecke, A.; Staiger, V.; Israël, A.; Kaltschmidt, C.; Mémet, S. NF-kappaB regulates spatial memory formation and synaptic plasticity through protein kinase A/CREB signaling. Mol Cell Biol. 2006, 26, 2936–2946. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Oikawa, K.; Odero, G.L.; Platt, E.; Neuendorff, M.; Hatherell, A.; Bernstein, M.J.; Albensi, B.C. NF-κB p50 subunit knockout impairs late LTP and alters long term memory in the mouse hippocampus. BMC Neurosci. 2012, 13, 45. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- He, F.Q.; Qiu, B.Y.; Zhang, X.H.; Li, T.K.; Xie, Q.; Cui, D.J.; Huang, X.L.; Gan, H.T. Tetrandrine attenuates spatial memory impairment and hippocampal neuroinflammation via inhibiting NF-κB activation in a rat model of Alzheimer’s disease induced by amyloid-β(1-42). Brain Res. 2011, 1384, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Merlo, E.; Freudenthal, R.; Maldonado, H.; Romano, A. Activation of the transcription factor NF-kappaB by retrieval is required for long-term memory reconsolidation. Learn Mem. 2005, 12, 23–29. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Alberini, C.M. Transcription factors in long-term memory and synaptic plasticity. Physiol Rev. 2009, 89, 121–145. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Romano, A.; Freudenthal, R.; Merlo, E.; Routtenberg, A. Evolutionarily-conserved role of the NF-kappaB transcription factor in neural plasticity and memory. Eur J Neurosci. 2006, 24, 1507–1516. [Google Scholar] [CrossRef]
- Barger, S.W.; Hörster, D.; Furukawa, K.; Goodman, Y.; Krieglstein, J.; Mattson, M.P. Tumor necrosis factors alpha and beta protect neurons against amyloid beta-peptide toxicity: evidence for involvement of a kappa B-binding factor and attenuation of peroxide and Ca2+ accumulation. Proc Natl Acad Sci U S A. 1995, 92, 9328–9332. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Krushel, L.A.; Cunningham, B.A.; Edelman, G.M.; Crossin, K.L. NF-kappaB activity is induced by neural cell adhesion molecule binding to neurons and astrocytes. J Biol Chem. 1999, 274, 2432–2439. [Google Scholar] [CrossRef] [PubMed]
- Alberini, C.M.; Ledoux, J.E. Memory reconsolidation. Curr Biol. 2013, 23, R746–50. [Google Scholar] [CrossRef] [PubMed]
- Federman, N.; de la Fuente, V.; Zalcman, G.; Corbi, N.; Onori, A.; Passananti, C.; Romano, A. Nuclear factor κB-dependent histone acetylation is specifically involved in persistent forms of memory. J Neurosci. 2013, 33, 7603–7614. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gupta, S.; Kim, S.Y.; Artis, S.; Molfese, D.L.; Schumacher, A.; Sweatt, J.D.; Paylor, R.E.; Lubin, F.D. Histone methylation regulates memory formation. J Neurosci. 2010, 30, 3589–3599. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Boccia, M.; Freudenthal, R.; Blake, M.; de la Fuente, V.; Acosta, G.; Baratti, C.; Romano, A. Activation of hippocampal nuclear factor-kappa B by retrieval is required for memory reconsolidation. J Neurosci. 2007, 27, 13436–13445. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Carayol, N.; Chen, J.; Yang, F.; Jin, T.; Jin, L.; States, D.; Wang, C.Y. A dominant function of IKK/NF-kappaB signaling in global lipopolysaccharide-induced gene expression. J Biol Chem. 2006, 281, 31142–31151. [Google Scholar] [CrossRef] [PubMed]
- Lubin, F.D.; Sweatt, J.D. The IkappaB kinase regulates chromatin structure during reconsolidation of conditioned fear memories. Neuron. 2007, 55, 942–957. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lucchesi, W.; Mizuno, K.; Giese, K.P. Novel insights into CaMKII function and regulation during memory formation. Brain Res Bull. 2011, 85, 2–8. [Google Scholar] [CrossRef]
- Ramakrishnan, P.; Clark, P.M.; Mason, D.E.; Peters, E.C.; Hsieh-Wilson, L.C.; Baltimore, D. Activation of the transcriptional function of the NF-κB protein c-Rel by O-GlcNAc glycosylation. Sci Signal. 2013, 6, ra75. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hanover, J.A.; Krause, M.W.; Love, D.C. Bittersweet memories: linking metabolism to epigenetics through O-GlcNAcylation. Nat Rev Mol Cell Biol. 2012, 13, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Kaltschmidt, C.; Kaltschmidt, B.; Neumann, H.; Wekerle, H.; Baeuerle, P.A. Constitutive NF-kappa B activity in neurons. Mol Cell Biol. 1994, 14, 3981–3992. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shih, V.F.; Tsui, R.; Caldwell, A.; Hoffmann, A. A single NFκB system for both canonical and non-canonical signaling. Cell Res. 2011, 21, 86–102. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Freudenthal, R.; Boccia, M.M.; Acosta, G.B.; Blake, M.G.; Merlo, E.; Baratti, C.M.; Romano, A. NF-kappaB transcription factor is required for inhibitory avoidance long-term memory in mice. Eur J Neurosci. 2005, 21, 2845–2852. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Moerman, A.M.; Barger, S.W. Neuronal kappa B-binding factors consist of Sp1-related proteins. Functional implications for autoregulation of N-methyl-D-aspartate receptor-1 expression. J Biol Chem. 2002, 277, 44911–44919. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley Interdiscip Rev Syst Biol Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nader, K.; Schafe, G.E.; Le Doux, J.E. Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval. Nature. 7: 17;406(6797), 6797. [Google Scholar] [CrossRef] [PubMed]
- Merlo, E.; Freudenthal, R.; Maldonado, H.; Romano, A. Activation of the transcription factor NF-kappaB by retrieval is required for long-term memory reconsolidation. Learn Mem. 2005, 12, 23–29. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cheung, P.; Allis, C.D.; Sassone-Corsi, P. Signaling to chromatin through histone modifications. Cell. 2000, 103, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Viatour, P.; Legrand-Poels, S.; van Lint, C.; Warnier, M.; Merville, M.P.; Gielen, J.; Piette, J.; Bours, V.; Chariot, A. Cytoplasmic IkappaBalpha increases NF-kappaB-independent transcription through binding to histone deacetylase (HDAC) 1 and HDAC3. J Biol Chem. 2003, 278, 46541–46548. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Lin, Z.; SenBanerjee, S.; Jain, M.K. Tumor necrosis factor alpha-mediated reduction of KLF2 is due to inhibition of MEF2 by NF-kappaB and histone deacetylases. Mol Cell Biol. 2005, 25, 5893–5903. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Freudenthal, R.; Locatelli, F.; Hermitte, G.; Maldonado, H.; Lafourcade, C.; Delorenzi, A.; Romano, A. Kappa-B like DNA-binding activity is enhanced after spaced training that induces long-term memory in the crab Chasmagnathus. Neurosci Lett. 1998, 242, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Merlo, E.; Freudenthal, R.; Romano, A. The IkappaB kinase inhibitor sulfasalazine impairs long-term memory in the crab Chasmagnathus. Neuroscience. 2002, 112, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Meffert, M.K.; Chang, J.M.; Wiltgen, B.J.; Fanselow, M.S.; Baltimore, D. NF-kappa B functions in synaptic signaling and behavior. Nat Neurosci. 2003, 6, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.H.; Lin, C.H.; Lee, C.F.; Gean, P.W. A requirement of nuclear factor-kappaB activation in fear-potentiated startle. J Biol Chem. 2002, 277, 46720–46729. [Google Scholar] [CrossRef] [PubMed]
- Boersma, M.C.; Dresselhaus, E.C.; De Biase, L.M.; Mihalas, A.B.; Bergles, D.E.; Meffert, M.K. A requirement for nuclear factor-kappaB in developmental and plasticity-associated synaptogenesis. J Neurosci. 2011, 31, 5414–5425. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Matsuzaki, M.; Honkura, N.; Ellis-Davies, G.C.; Kasai, H. Structural basis of long-term potentiation in single dendritic spines. Nature. 2004, 429, 761–766. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nägerl, U.V.; Eberhorn, N.; Cambridge, S.B.; Bonhoeffer, T. Bidirectional activity-dependent morphological plasticity in hippocampal neurons. Neuron. 2004, 44, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Homma, K.J.; Poo, M.M. Shrinkage of dendritic spines associated with long-term depression of hippocampal synapses. Neuron. 2004, 44, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Kopec, C.D.; Real, E.; Kessels, H.W.; Malinow, R. GluR1 links structural and functional plasticity at excitatory synapses. J Neurosci. 2007, 27, 13706–13718. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nusser, Z.; Lujan, R.; Laube, G.; Roberts, J.D.; Molnar, E.; Somogyi, P. Cell type and pathway dependence of synaptic AMPA receptor number and variability in the hippocampus. Neuron. 1998, 21, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, M.; Ellis-Davies, G.C.; Nemoto, T.; Miyashita, Y.; Iino, M.; Kasai, H. Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nat Neurosci. 2001, 4, 1086–1092. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Saha, R.N.; Liu, X.; Pahan, K. Up-regulation of BDNF in astrocytes by TNF-alpha: a case for the neuroprotective role of cytokine. J Neuroimmune Pharmacol. 2006, 1, 212–222. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gao, J.; Grill, R.J.; Dunn, T.J.; Bedi, S.; Labastida, J.A.; Hetz, R.A.; Xue, H.; Thonhoff, J.R.; DeWitt, D.S.; Prough, D.S.; Cox CSJr Wu, P. Human Neural Stem Cell Transplantation-Mediated Alteration of Microglial/Macrophage Phenotypes after Traumatic Brain Injury. Cell Transplant. 2016, 25, 1863–1877. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, E.C.; Hunot, S. Neuroinflammation in Parkinson’s disease: a target for neuroprotection? Lancet Neurol. 2009, 8, 382–397. [Google Scholar] [CrossRef] [PubMed]
- Abou-Sleiman, P.M.; Muqit, M.M.; Wood, N.W. Expanding insights of mitochondrial dysfunction in Parkinson’s disease. Nat Rev Neurosci. 2006, 7, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity. 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in neurodegenerative disorders: the roles of microglia and astrocytes. Transl Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization From M1 to M2 in Neurodegenerative Diseases. Front Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- L’Episcopo, F.; Tirolo, C.; Serapide, M.F.; Caniglia, S.; Testa, N.; Leggio, L.; Vivarelli, S.; Iraci, N.; Pluchino, S.; Marchetti, B. Microglia Polarization, Gene-Environment Interactions and Wnt/β-Catenin Signaling: Emerging Roles of Glia-Neuron and Glia-Stem/Neuroprogenitor Crosstalk for Dopaminergic Neurorestoration in Aged Parkinsonian Brain. Front Aging Neurosci. 2018, 10, 12. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Peruzzotti-Jametti, L.; Bernstock, J.D.; Vicario, N.; Costa, A.S.H.; Kwok, C.K.; Leonardi, T.; Booty, L.M.; Bicci, I.; Balzarotti, B.; Volpe, G.; Mallucci, G.; Manferrari, G.; Donegà, M.; Iraci, N.; Braga, A.; Hallenbeck, J.M.; Murphy, M.P.; Edenhofer, F.; Frezza, C.; Pluchino, S. Macrophage-Derived Extracellular Succinate Licenses Neural Stem Cells to Suppress Chronic Neuroinflammation. Cell Stem Cell. 2018, 22, 355–368.e13. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kempuraj, D.; Thangavel, R.; Natteru, P.A.; Selvakumar, G.P.; Saeed, D.; Zahoor, H.; Zaheer, S.; Iyer, S.S.; Zaheer, A. Neuroinflammation Induces Neurodegeneration. J Neurol Neurosurg Spine. 2016, 1, 1003. [Google Scholar] [PubMed] [PubMed Central]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu Rev Immunol. 4: 26;35. [CrossRef] [PubMed]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; Samokhvalov, I.M.; Merad, M. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science. 2010, 330, 841–845. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rezai-Zadeh, K.; Gate, D.; Town, T. CNS infiltration of peripheral immune cells: D-Day for neurodegenerative disease? J Neuroimmune Pharmacol. 2009, 4, 462–475. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sedighzadeh, S.S.; Khoshbin, A.P.; Razi, S.; Keshavarz-Fathi, M.; Rezaei, N. A narrative review of tumor-associated macrophages in lung cancer: regulation of macrophage polarization and therapeutic implications. Transl Lung Cancer Res. 2021, 10, 1889–1916. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Omi, M.; Hata, M.; Nakamura, N.; Miyabe, M.; Ozawa, S.; Nukada, H.; Tsukamoto, M.; Sango, K.; Himeno, T.; Kamiya, H.; Nakamura, J.; Takebe, J.; Matsubara, T.; Naruse, K. Transplantation of dental pulp stem cells improves long-term diabetic polyneuropathy together with improvement of nerve morphometrical evaluation. Stem Cell Res Ther. 2017, 8, 279. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
Figure 1.
Created in BioRender. Tuchman, K. (2025) https://BioRender.com/6rfoaiv,Natural killer (NK) cells recruited by angiogenic cell precursors (ACP)s suppress inflammation through release of anti-inflammatory cytokines, dendritic cell and monocyte maturation and lysis of auto-aggressive T-cells.
Figure 1.
Created in BioRender. Tuchman, K. (2025) https://BioRender.com/6rfoaiv,Natural killer (NK) cells recruited by angiogenic cell precursors (ACP)s suppress inflammation through release of anti-inflammatory cytokines, dendritic cell and monocyte maturation and lysis of auto-aggressive T-cells.
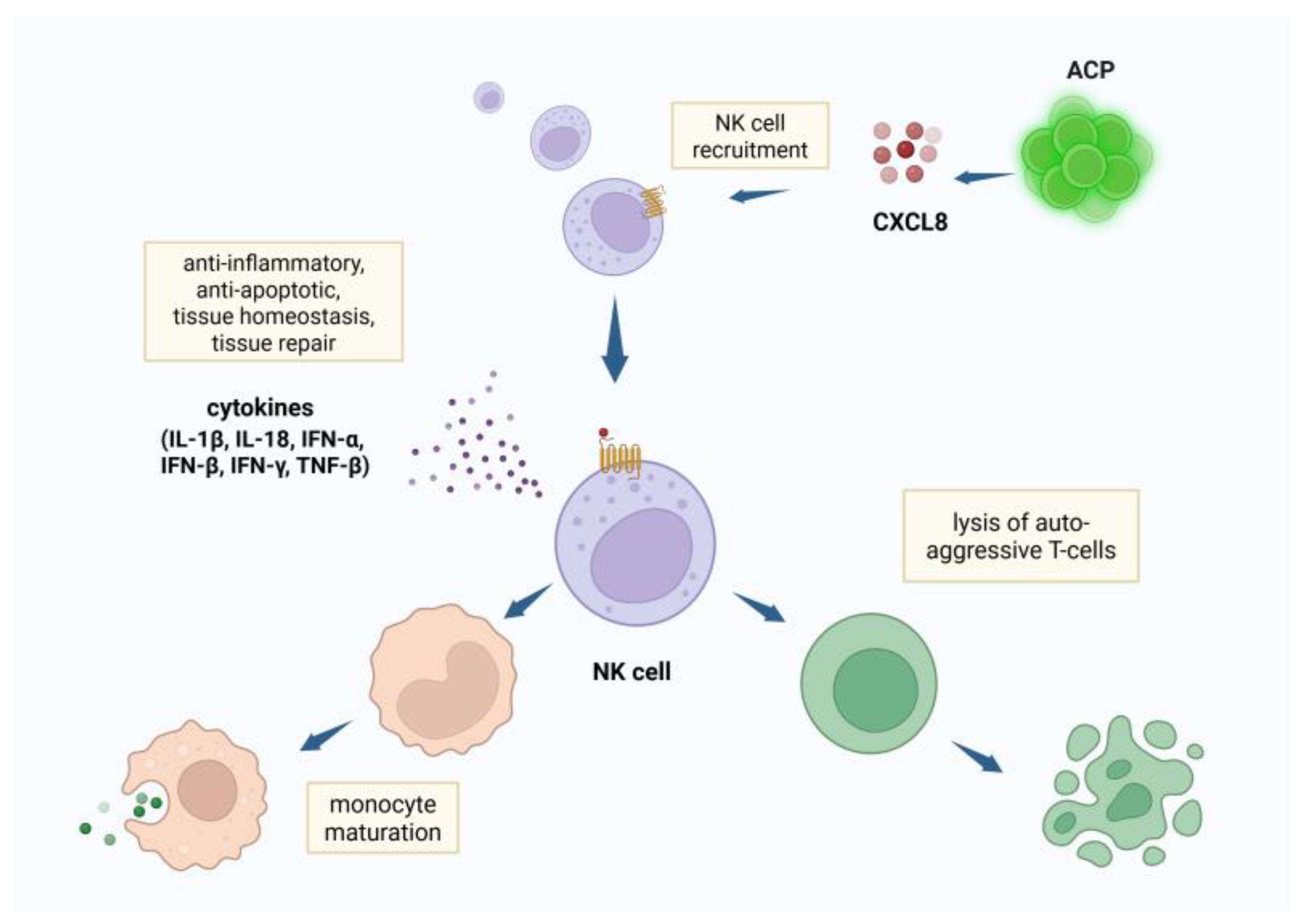
Figure 2.
Created in BioRender. Tuchman, K. (2025) https://BioRender.com/w2cthsg,ngiogenic cell precursors (ACP)s potentiate healing through secretion of tissue regeneration factors such as the chemokine interleukin-8 (CXCL8), vascular endothelial growth factor (VEGF) and angiogenin. CXCL8 recruits peripheral CD34+ precursor cells, further supporting angiogenesis.
Figure 2.
Created in BioRender. Tuchman, K. (2025) https://BioRender.com/w2cthsg,ngiogenic cell precursors (ACP)s potentiate healing through secretion of tissue regeneration factors such as the chemokine interleukin-8 (CXCL8), vascular endothelial growth factor (VEGF) and angiogenin. CXCL8 recruits peripheral CD34+ precursor cells, further supporting angiogenesis.
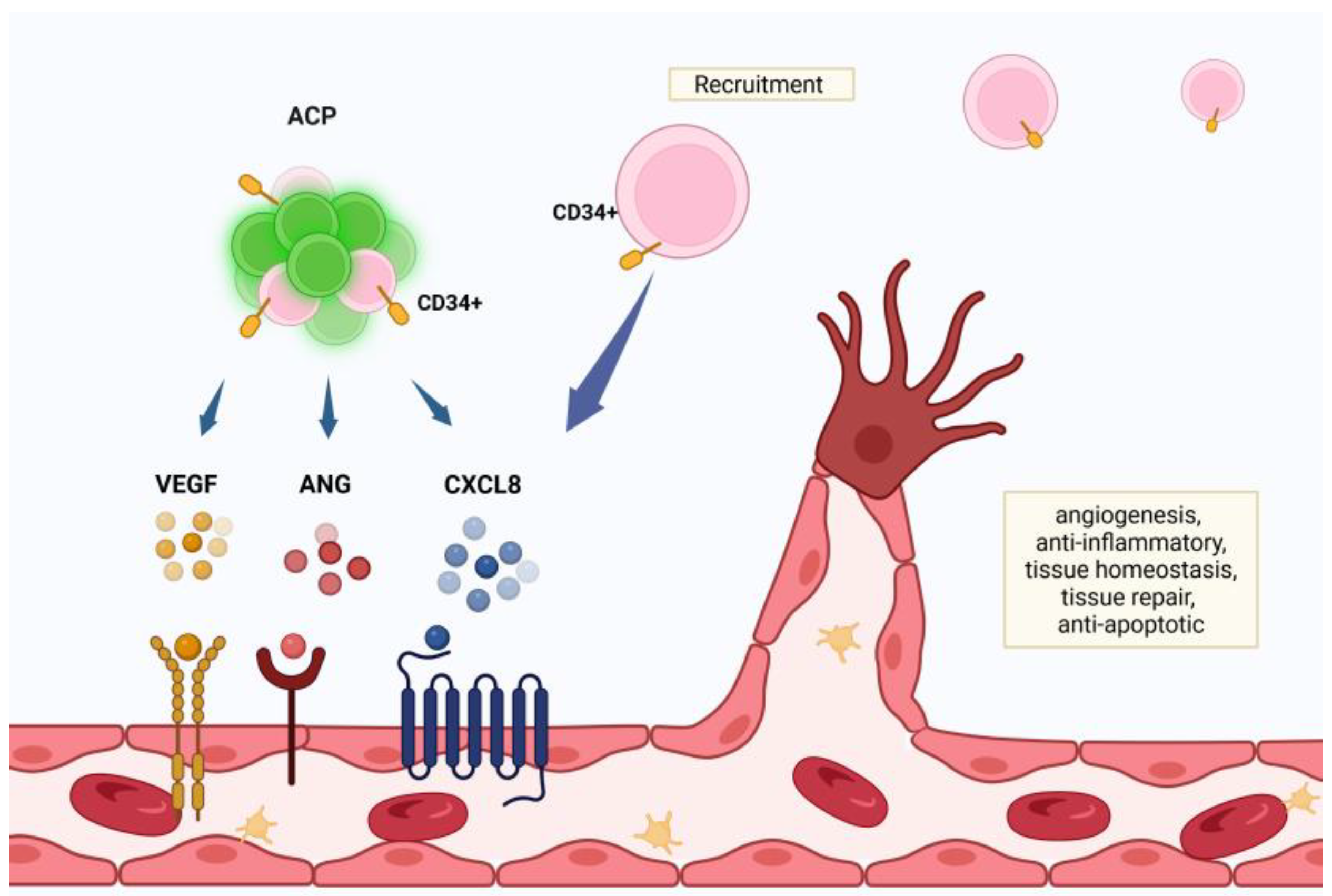
Figure 3.
Created in BioRender. Tuchman, K. (2025) https://BioRender.com/u2icrk5,Interleukin-8 (CXCL8) is expressed by angiogenic cell precursors (ACP)s and activates the canonical NF-κB pathway, resulting in gene transcription and protein synthesis necessary for memory formation and consolidation.
Figure 3.
Created in BioRender. Tuchman, K. (2025) https://BioRender.com/u2icrk5,Interleukin-8 (CXCL8) is expressed by angiogenic cell precursors (ACP)s and activates the canonical NF-κB pathway, resulting in gene transcription and protein synthesis necessary for memory formation and consolidation.
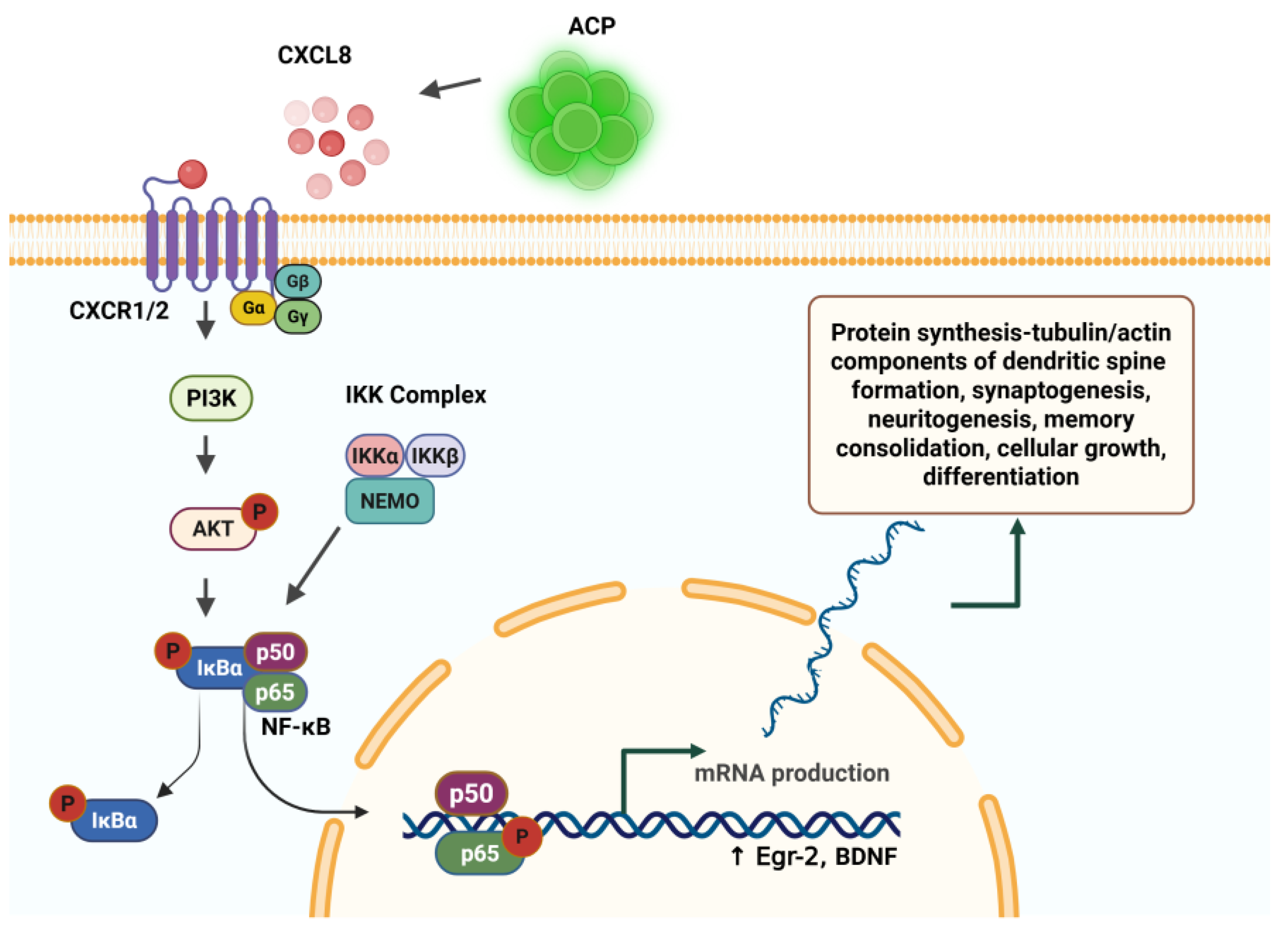
Figure 4.
Created in BioRender. Tuchman, K. (2025) https://BioRender.com/36j1vy7,Neural progenitor cells (NCP) have CXCR4 receptors, and migrate towards CXCL12 released by injured tissue. NCP promote the M2 (neuroprotective) macrophage phenotype, which release anti-inflammatory factors. The NCP differentiate into neuronal , glial or oligodendrocytic cells.
Figure 4.
Created in BioRender. Tuchman, K. (2025) https://BioRender.com/36j1vy7,Neural progenitor cells (NCP) have CXCR4 receptors, and migrate towards CXCL12 released by injured tissue. NCP promote the M2 (neuroprotective) macrophage phenotype, which release anti-inflammatory factors. The NCP differentiate into neuronal , glial or oligodendrocytic cells.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



