
Submitted:
23 June 2025
Posted:
26 June 2025
You are already at the latest version
Abstract
Background: Sickle cell disease (SCD) is an inherited blood disorder characterized by chronic hemolysis, inflammation, and vaso-occlusive crises (VOC), leading to several complications and reduced life expectancy of affected individuals. Limited effective treatment options are currently available; however, pharmacogenomic (PGx) insights derived from recent genomic findings, particularly in underrepresented populations such as Saudi Arabians, may offer new hope for predicting molecular-guided treatments. This study aimed to identify approved drugs suitable for repurposing based on their interaction with SCD-associated genetic variants and discover novel druggable targets within genetic pathways linked to disease severity by utilizing genome-wide association study (GWAS) data from Saudi SCD patients.
Methods: Our PGx analysis used bioinformatic pipelines to evaluate drug-gene interactions and identify potential therapeutic targets to treat SCD based on GWAS data derived from the Saudi population. Selected approved drugs were suggested for repurposing based on their interactions with genes of interest known to impact SCD pathophysiology, using the Drug-Gene Interaction Database (DGIdb 5.0). New drug targets were also proposed based on assessing the simulated pockets of gene products according to the 3D protein structures provided by the protein data bank (PDB) and the AlphaFold database. New molecules with higher druggability scores, estimated based on the DoGSiteScorer database, were predicted to have a higher success rate in developing new SCD treatments.
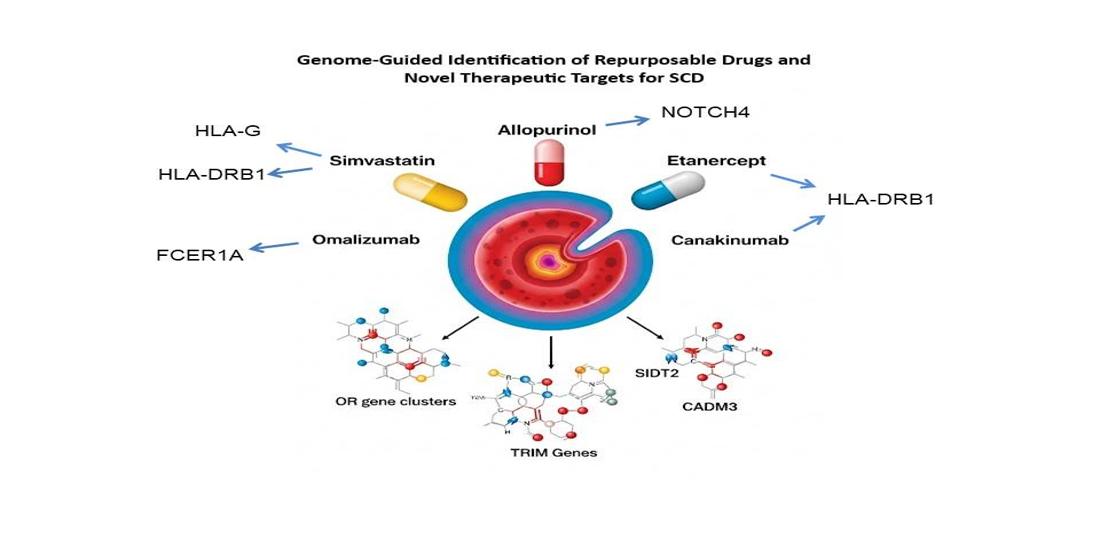
Results: Our analysis identified 78 approved medications that showed potential for repurposing in SCD; this list was narrowed down to 21 candidates based on their safety profiles and interactions with key genetic pathways in SCD. Among them, simvastatin, allopurinol, omalizumab, canakinumab, and etanercept were suggested as the most promising agents. Furthermore, novel drug targets encoded by olfactory receptor (OR) gene clusters (OR51V1, OR52A1, OR52A5, OR51B5, and OR51S1), TRIM genes, SIDT2, and CADM3, had high druggability scores.
Conclusion: This study provides a robust framework for drug repurposing and novel drug discovery in SCD, particularly tailored to the Saudi population. The findings underscore the potential of leveraging genomic data to identify targeted therapies, offering a pathway to more personalized and effective treatments for SCD patients. Future clinical trials are essential to validate these findings and translate them into clinical practice.
Keywords:
Sickle cell disease
; Drug repurposing
; Saudi population
; Precision medicine
; Pharmacogenomics
1. Introduction
Sickle cell disease (SCD) is a distressing inherited hematological disorder caused by a pathological hemoglobin variant (rs334 c.20 A>T) in the HBB gene, which induces rigid deformation or “sickling” of red blood cells (erythrocytes) [1]. This sickling leads to a range of downstream vascular complications, including vaso-occlusive episodes, thrombosis, and multiorgan infarctions, contributing to the clinical severity of SCD from early childhood onward [1,2]. The severe nature of these complications is further highlighted by the reduced life expectancy among patients compared to unaffected individuals in the general population [3].
From a public health perspective, SCD imposes a substantial global burden, with recent estimates indicating that over 7 million individuals live with the disease worldwide, with the highest prevalence found in sub-Saharan Africa, where the sickle cell trait has been historically promoted by the region's malaria endemicity [4]. Significant burdens also exist in India, the Americas, the Mediterranean, and the Middle East, though accurate epidemiological data is often limited by inadequate surveillance and historically high childhood mortality rates [5].
Reflecting this global burden, Saudi Arabia faces a notably high prevalence of SCD, with carrier rates nearing 27% and disease prevalence reaching 2.6% [6,7]. This substantial burden is likely exacerbated by high rates of consanguineous marriages, which concentrate inherited disease variants [6,8].
Despite the significant unmet medical need, the range of approved therapies for SCD remains remarkably limited. Initially, hydroxyurea was the only treatment approved by the US FDA for both adults and children with SCD. Since 2017, the therapeutic landscape has evolved, with the US FDA approving three new agents targeting distinct aspects of SCD pathophysiology, including: L-glutamine, crizanlizumab, and voxelotor [9,10]. However, recent developments have highlighted the challenges in this field. Notably, as of September 2024, voxelotor was voluntarily withdrawn globally due to safety concerns, and in May 2023, crizanlizumab was withdrawn from the European market [11,12]. These events underscore the complexities of SCD treatment and emphasize the critical importance of ongoing research, rigorous long-term safety monitoring, and continuous re-evaluation of approved therapies.
While bone marrow transplantation (BMT) offers a potential cure, its application is often hindered by significant challenges, including high costs, donor availability, and associated risks. The recent approval of the first cell and gene therapies, Casgevy and Lyfgenia, on December 8, 2023, marked a substantial advancement in SCD treatment. However, these cutting-edge therapies remain inaccessible to many patients due to their extremely high costs [13]. This situation highlights the urgent need for further research into innovative therapies that offer safer, more effective, and affordable treatment options for SCD.
Furthermore, the complex genetic nature of SCD, coupled with recently identified disease-associated genetic variants from genome-wide association studies (GWAS), presents an ideal opportunity for pharmacogenomic (PGx) approaches to guide both drug repurposing efforts and novel therapeutic development. [14,15] Drug repurposing represents a systematic methodology to propose new indications for currently marketed drugs based on their interactions with key genes that modulate cellular pathways and potentially ameliorate disease outcomes [16]. This computationally driven approach, which relies on in silico gene target analyses, has been used effectively to repurpose multiple drugs for various disease conditions [17].
Our research focuses on repurposing existing drugs and discovering new molecules for SCD, which would then undergo clinical trials. Such advancements could revolutionize SCD management, enhancing scientific knowledge, technical capabilities, and clinical practice through the identification of novel genetic targets. These efforts have the potential to lead to groundbreaking treatments, setting new standards in the field and driving further innovation in personalized medicine.
In addition to the HbS variant, the severity spectrum of SCD manifests extensive clinical heterogeneity influenced by multiple genes, [18] making it particularly suitable for PGx investigation. Previous genetic studies have uncovered polymorphisms that modulate different disease molecular pathways, including hemolysis, inflammation, oxidative stress, and vascular dysfunction [18,19,20]. Examples include fetal hemoglobin (HbF)-regulating variants in BCL11A and HBS1L-MYB, as well as pro-inflammatory cytokine polymorphisms in TNF-α and VCAM-1, which are associated with an increased risk of stroke [19]. These genetic insights not only provide critical frameworks for repurposing existing drugs to target emergent SCD mechanisms but also pave the way for developing innovative, tailored therapies that could significantly improve patient outcomes [21].
The shortage of viable treatments for SCD has prompted the exploration of alternative strategies, such as pathway-based, phenotypic, or in silico drug repurposing, leveraging existing compound libraries or approved agents to target novel SCD disease mechanisms [22,23,24]. Integrating genetic data from recent high-throughput genomic studies across diverse SCD patient populations empowers the rational identification of putative druggable targets and prioritizes candidates for screening [18,25]. However, most investigations have predominantly focused on African or African-American ancestries, with limited examination of other susceptible populations such as those in the Mediterranean and Middle Eastern regions [26].
Recently, our group uncovered Saudi-enriched genetic variants that drive thromboembolic complications in SCD patients, contrasting with markers identified in other cohorts [25]. Additionally, findings from our current data have identified several loci within known genes linked to molecular pathophysiological pathways potentially influencing SCD severity, underscoring the need for tailored interventions [15]. Expanding the characterization of population-specific molecular underpinnings through comprehensive genomics offers two key advantages: it uncovers novel therapeutic targets specific to Saudis and clarifies the heterogeneity in pathogenic mechanisms across the global SCD patient spectrum.
Therefore, in this study, we aimed to leverage findings from our GWAS analysis in conjunction with druggable target enrichment analysis, specifically focusing on Saudi SCD patients, to propose new SCD treatment options. Our overarching goal is to develop an informed, genetically guided roadmap that accelerates the development of innovative, population-tailored therapeutics for this historically overlooked SCD patient group. This holistic, molecular knowledge-based approach also seeks to evaluate and select the most promising druggable, non-targeted genes, thereby refining and expediting future drug discovery efforts.
2. Method
2.1. The Pipeline for Drug Repurposing and Novel Drug Development
We utilized data derived from our previous Saudi SCD GWAS analysis [15] to assess the druggability of genes identified in Saudi SCD patients with potential impact on disease outcomes, aiming to establish their feasibility as therapeutic targets. Our analysis focused on identifying potential gene-drug interactions to pinpoint candidates for drug repurposing and suggesting new molecules to be developed as future treatment options. The analysis in this study followed a previously published pipeline (Figure 1) [16].
2.2. Selection of SCD Severity-Associated Genes
We focused on the reported 31 genes associated with SCD severity [15]. These genes were selected based on their involvement in key SCD pathological pathways, including inflammation regulation, vascular function, endothelial modulation, and other disease-relevant biological processes. The selection criteria incorporated both pathway analysis of the identified genes and a comprehensive literature review, which demonstrated the functional relevance of these genes on SCD clinical phenotypes.
2.3. Drug-Gene Interaction Analysis for Drug Repurposing
Gene products were filtered based on their potential to interact with medicinal drugs using the Drug-Gene Interaction Database (DGIdb) version 5.0, which consolidates data from multiple sources, including DrugBank and the Pharmacogenomics Knowledge Base (PharmGKB), to identify potential drug repurposing opportunities [27]. The database provides detailed annotation of drug-gene interactions and gene druggability, covering approved drugs and experimental compounds.
Since gene products can interact with multiple drugs, we first excluded agents not clinically available (e.g., terminated or investigational drugs) due to their lack of immediate clinical applicability. Approved medications were then mapped to the genes of interest known impact disease pathways, excluding non-drug products.
During the initial filtration, we excluded terminated or withdrawn drugs. We prioritized medications targeting common SCD complications (e.g., inflammation, vascular dysfunction, immune dyregulation), as these pathways directly influence SCD severity. Diagnostic agents, vitamins, supplements, and superseded medications (replaced by more effective alternatives) were also excluded.
Candidate medications were further prioritized using predicted safety profiles. Using VigiBase (the largest global adverse event repository, https://www.vigiaccess.org/) and the Side Effect Resource (SIDER 4.1) database (http://sideeffects.embl.de/), we excluded drugs linked to severe adverse reactions [28,29]. Chemotherapy agents and drugs with serious toxicities (e.g., hepatotoxicity, cardiotoxicity) were excluded due to high-risk safety profiles. Agents posing a risk of hemolytic anemia were also removed to prevent exacerbating hemolysis in SCD patients. For comprehensive identification, we used the Search Tool for Interactions of Chemicals (STITCH) and the World Health Organization (WHO) databases to standardize drug nomenclature to match the International Nonproprietary Names (INN) and United States Adopted Names (USAN) and assign Anatomical Therapeutic Chemical (ATC) codes [30,31]. We prioritized avoiding medications that could worsen SCD severity, especially those with risks of infection, cognitive impairment, or blood pressure instability, given SCD patients’ heightened vulnerability to these complications.
2.4. Novel Drug Development Pipeline
Genes identified in our GWAS study [15], with no known interaction with approved drugs, were labeled as non-targeted genes. Selection criteria included: 1) variant consequences, 2) reported associations with SCD phenotypes, 3) gene function, and 4) expression in SCD-relevant tissues. Druggability scores were calculated to assess the likelihood of a compound interacting with a these gene products, based on the structural compatibility of their binding pockets with drug molecules.
Protein druggability was ranked using DoGSiteScorer [32], which analyzes Protein Data Bank (PDB) structures to evaluate physicochemical interactions and identify ligand-binding pockets ((https://www.rcsb.org/) [33]. DoGSiteScorer employs a difference Gaussian filter to detect potential pockets based solely on the 3D protein structures. Score ranges between 0 and 1, with values >0.5 indicating potential druggability. This threshold was validated using 12 drug-targeted protein structures (scores: 0.68 to 0.88; Supplementary Table), enabling prioritization of candidate genes for drug development.
To address the absence of experimentally determined structures of some proteins in PDB, we used the AlphaFold database [34], which provides highly accurate artificial intelligence (AI)-predicted structures. AlphaFold employs deep learning to predict protein structures with near-experimentally accuracy. Notably, its developers were awarded the 2024 Nobel Prize in chemistry for this breakthrough [35]. Combined with DoGSiteScorer, these tools enable identification of theoretical binding sites even without experimental structural data.
Predicted binding sites were visualized using UCSF ChimeraX V1.8 to generate 3D protein structure figures. Gene product expression was validated via the Human Protein Atlas (https://www.proteinatlas.org/) [36], the Gene Expression Omnibus (GEO) of the National Center for Biotechnology Information (NCBI, https://www.ncbi.nlm.nih.gov/geo/) [37], or the Genotype-Tissue Expression (GTEX) portal (https://www.gtexportal.org/) [38]. Genes with predominant expression in tissues unaffected by SCD were excluded (Supplementary Table).
3. Results
3.1. Promising Candidates for Drug Repurposing
Out of the 31 genes suggested as modifiers for SCD phenotypes, 12 genes (ACKR1, AGER, FCER1A, HBG2, HLA-A, HLA-DQB1, HLA-G, HLA-DRB1, TRIM5, NOTCH4, RRM1, and STIM1) showed the ability to interact with a total of 114 distinct drug molecules. Some of these drugs (n = 36) are not yet approved for clinical use and therefore excluded. The HBG2 gene did not show any interaction with approved drugs. Among the remaining 78 drugs, chemotherapies (n = 21), vitamins and diagnostic agents (n = 3), vaccines (n = 3), and antimicrobials (n = 12) were excluded. The remaining medications (n = 39) were further filtered based on their safety profiles.
Consequently, we excluded anticonvulsants (n = 5), antidepressants (n = 5), antithyroid (n = 3), glucocorticoids, ticlopidine, and hydralazine due to their undesirable side effects. Hydroxyurea and the analgesic agents (e.g., acetaminophen and lumiracoxib) were identified by our pipeline but were not included in the final suggested list, as they are routinely prescribed for managing SCD in clinical practice. Detection of the genes targeted by hydroxyurea (RRM1) and the analgesics (HLA-DQB1) served as benchmarks to validate the pipeline's performance (Supplementary Excel Sheet).
As a result, we narrowed down the list to the interactions between 21 medications and 6 genes, with 19 of these medications interacting with four HLA genes (HLA-DQB1, DRB1, G, and A). The remaining two medications (omalizumab and allopurinol) interact with two distinct genes (FCER1A and NOTCH4, respectively) (Table 1). The most frequently detected therapeutic class was the immunosuppressants (n = 12), including adalimumab, canakinumab, infliximab, omalizumab, tocilizumab, etanercept, rilonacept, peginterferon alfa-2a and 2b, azathioprine, glatiramer, and anakinra. The second most common class was lipid-modifying agents (n = 7), which included fenofibrate and statins (pitavastatin, pravastatin, atorvastatin, simvastatin, rosuvastatin, and fluvastatin).
Ultimately, we proposed immunomodulators, particularly monoclonal antibodies, and statins as potentially suitable candidates for repurposing to manage SCD severity based on their interactions with key genes that have known impact on SCD pathological pathways, as supported by literature evidence[15,39]. Three immunomodulators working on different gene targets (omalizumab targeting FCER1A and canakinumab and etanercept targeting HLA-DRB1) and a statin agent (simvastatin interacting with HLA-G and HLA-DRB1) demonstrated a high drug-gene interaction score. The antihyperuricemic agent (allopurinol), which targets NOTCH4, also represents a potential repurposing option.
3.2. Druggability Analysis of Non-Targeted Loci
Among the 31 genes suggested by our GWAS study [15], 12 genes were shortlisted as candidates for drug repurposing, while the remaining 19 genes were assessed to identify their chemical structures and evaluate their druggability. No protein structures were detected for three gene products (OR10J8P, OR10J9P, and HBBP1), resulting in their exclusion. Additionally, three more genes were excluded as well: POC5 due to an unclear role in SCD pathological mechanisms, SCAND3 and MMP26 due to predominant expression in irrelevant tissues (e.g., testis and endometrium).
Thirteen suitable gene targets were prioritized for drug development, with druggability scores ranging from 0.7 to 0.87. Detailed descriptions of all molecules that passed the pipeline filters are presented in Table 2. The olfactory receptor (OR) gene cluster (OR51V1, OR52A1, OR52A5, OR51B5, and OR51S1) and the Tripartite Motif gene family (TRIM6, TRIM22, and TRIM34) emerged as prominent candidates. More details about all screened molecules are available in Supplementary Table. The OR gene cluster, particularly OR51V1, OR51A1, and OR51B5, has been extensively linked to SCD phenotypes in the literature. Additionally, TRIM6, SIDT2, and CADM3 were suggested due to their high expression in SCD-related tissues and their potential roles in the related complications. These promising protein candidates, along with their structural visualizations and druggability scores, are shown in Figure 2.
4. Discussion
Applying genetics knowledge to enable the repurposing of drugs has emerged as a promising and innovative strategy in the field of pharmacology. In 2021, the U.S. Food and Drug Administration (FDA) approved approximately two-thirds of new medications based on genetic insights [40,41]. The development and application of advanced genetic analysis approaches such as GWAS and next-generation sequencing (NGS) [e.g., whole-genome sequencing (WGS) and whole-exome sequencing (WES)] have significantly deepened our understanding of the genetic underpinnings of complex traits [42,43]. This progress facilitated the identification of promising drug targets through the discovery of causative genetic variants [44]. However, effectively translating these genetic discoveries into clinical practice remains challenging, particularly for conditions like SCD with diverse phenotypes influenced by multifactorial genetic associations. The use of already authorized medications for new applications at more affordable costs is increasingly recognized as a valuable and expanding approach [16,40,45]. Several previous studies have utilized in silico pipeline to suggest potential drug candidates and identify target molecules for treating various medical conditions by leveraging findings from publicly available GWAS analyses [16,46,47]. Different GWAS databases are available and can be used to understand the genetic basis of selected diseases. Therefore, it is possible to translate identified signals into therapeutic targets using genomic-based approaches and a well-established pipeline [16]. In our project, we adopted a similar methodology to propose novel drug treatment options that could potentially target specific genes known to impact outcomes in SCD patients.
In a previous GWAS study, we identified multiple genes that play a significant role in SCD and its complications [15]. Furthermore, the current study aimed to maximize the utility of existing data to translate genetic knowledge into clinical care of SCD. Thus, 21 approved agents were identified as having strong affinity to interact with the reported genes modulating various SCD phenotypes. The chosen drug candidates are classified into lipid-modifying and immunomodulatory agents, with the potential to modulate multiple pathways in SCD, including sickling, hemolysis, and inflammation [48]. Based on unique criteria, the filtering process highlighted five highly promising candidates for repurposing: simvastatin, allopurinol, canakinumab, omalizumab, and etanercept.
In addition to simvastatin, other statins were also identified—such as atorvastatin, pravastatin, fluvastatin, rosuvastatin, and pitavastatin—have also shown promise for SCD management due to their anti-inflammatory properties. These agents downregulate the transcription factor NF-KB, inhibit the expression of key pro-inflammatory cytokines such as TNF and IL1B, and enhance endothelial function by restoring nitric oxide (NO) bioavailability and reducing the expression of adhesion molecule, collectively contributing to the vascular protective effects [49,50]. Notably, the JUPITER trial, a key intervention study evaluating rosuvastatin, demonstrated the immunomodulatory potential of statins, with rosuvastatin significantly reducing C-reactive protein (CRP) levels by approximately 37%, further supporting the role of statins in mitigating inflammation-related complications [51].
Simvastatin was suggested as the optimal candidate among all statins due to its superior interaction score and clinical relevance. While other statins interact with the HLA-DRB1 only, which encodes a major histocompatibility complex protein, simvastatin uniquely interacts with both HLA-DRB1 and HLA-G. These proteins are integral to immune responses that exacerbate SCD complications, such as vaso-occlusive crises and organ damage [52,53,54,55]. Furthermore, simvastatin has shown a greater capacity to reduce von Willebrand factor (vWF) levels, underscoring its crucial role in managing SCD-associated hypercoagulibility [56,57].
A clinical study in a small SCD cohort (n = 26) demonstrated that short-term simvastatin administration significantly improved nitric oxide bioavailability by approximately 52% (p = 0.01) and suppressed systemic inflammatory biomarkers such as CRP and IL-6, both of which are associated with increased vascular dysfunction risks in SCD [58]. Moreover, a pilot study on 19 SCD patients reported that simvastatin reduced the occurrence of vaso-occlusive pain by 85% (p = 0.0003) and decreased the use of analgesics by 67% (p = 0.003). It also showed a significant reduction in circulating CRP, soluble (s) E-selectin, intercellular adhesion molecule 1 (sICAM-1), sICAM-3, and vascular endothelial growth factor, with the most pronounced effects observed in patients concurrently receiving hydroxyurea [59].
In vitro studies further underscore simvastatin's potential, showing a 1.9-fold increase in fetal hemoglobin expression and a [30,31,32,33,34,35]% reduction in irreversibly sickled cells under hypoxic conditions [60]. An ongoing clinical trial is currently assessing simvastatin's effects on central nervous system vasculature in SCD patients (ClinicalTrials.gov Identifier: NCT03599609) [10]. Based on this robust body of evidence and specific criteria, we recommend simvastatin as the ideal drug for repurposing in SCD, although other statins may also offer benefits depending on individual patient factors. Further research, including large randomized controlled trials, is warranted to fully elucidate simvastatin’s therapeutic potential in this context.
Another suggested repurposing candidate is allopurinol, which targets products of the NOTCH4 gene. This gene is a member of the NOTCH family, well known for its role in regulating endothelial function and exerting anti-inflammatory effects, as well as influencing hematopoiesis, all of which may contribute to alleviating disease severity [61,62]. In the Saudi SCD dataset, we identified the marker rs3132946 in NOTCH4, which has been previously reported as a marker linked to interstitial lung diseases (ILD) [63]. This condition is notably common in SCD patients, with 74% of adults in a large prospective cohort study exhibiting a restrictive pattern on pulmonary function tests [64]. While not all adults with SCD show the common features of sickle cell lung disease (SCLD), this restrictive pattern remains the most consistent clinical manifestation, underscoring its significant impact on this population.
Allopurinol, a well-established xanthine oxidase inhibitor long used to treat hyperuricemia in patients with gout or tumor lysis syndrome [65], has demonstrated broader vascular benefits. It can protect vascular tissue from oxidative stress and repetitive reperfusion injury, both of which are critical components of SCD pathology [66]. By inhibiting the production of reactive oxygen species (ROS), allopurinol preserves nitric oxide availability, thereby supporting vascular relaxation and function [67]. Preclinical studies in sickle cell models show mixed results; while one study reports no significant effect on cell adhesion, another showed improved blood flow and reduced leukocyte recruitment upon exposure to allpurinol [68,69]. Given the oxidative stress and endothelial dysfunction observed in SCD, further exploration of allopurinol's therapeutic potential, particularly in combination with other agents, is warranted. However, careful consideration of its side effects, including hypersensitivity and rare severe reactions, is crucial. Even in populations where the HLA-B*5801 allele is not highly prevalent, such as the Saudi population [70], it is still important to genotype for HLA-B*5801 to mitigate the risk of hypersensitivity reactions when prescribing allopurinol [71].
Omalizumab was initially approved in 2003 by the US FDA for the treatment of moderate to severe persistent asthma, with expanded indications in subsequent years [72]. This medication, which targets the FCER1A gene product, has emerged as one of the top candidate repurposing drugs. This recombinant immunoglobulin G (IgG) is a monoclonal antibody that selectively binds to free IgE, thereby attenuating allergic asthma. By preventing IgE from interacting with high-affinity Fcε receptors on effector cells such as mast cells, omalizumab modulates downstream pro-inflammatory signaling that leads to airway inflammation and may also potentially reduce coagulation abnormalities associated with disease activity [73].
In the Saudi SCD cohort, the marker rs2494250 in the FCER1A was detected. This variant was previously reported as significantly associated with elevated levels of inflammatory biomarkers, including CCL2/MCP-1 [74]. Furthermore, studies have reported a higher prevalence of IgE among Saudis with SCD, potentially increasing the risk of acute chest syndrome (ACS). Notably, ACS accounts for 25% of ICU admissions and 28.5% of deaths among SCD patients, with particularly high rates observed in individuals from Al-Hasa region [75,76,77]. Thus, further clinical studies in SCD patients are warranted to confirm omalizumab benefits in mitigating this disease complication.
Our pipeline has also identified other biological disease-modifying drugs targeting the HLA-DRB1 gene. Canakinumab and etanercept were suggested as favorable candidates for repurposing in SCD management. Canakinumab can selectively target IL-1β, a cytokine with a central role in the inflammatory process, and may contribute to modulating disease pathways in SCD [10,78].
Moreover, canakinumab has been studied on 74 children and young adults with SCD in a recent double-blind, randomized study [10,79]. It demonstrated a tolerable safety profile, in contrast to other monoclonal antibodies that are often associated with a high risk of infection [80,81]. Although this clinical study did not achieve its primary goal of reducing daily pain compared to baseline, it showed several beneficial effects, including lower levels of inflammatory biomarkers, reduced hospitalization rates, and improvements in patient-reported outcomes such as fatigue and absenteeism from school or work [79,82]. Additionally, there is an ongoing clinical study evaluating its safety in pediatric SCD patients (ClinicalTrials.gov Identifier: NCT02961218) [10]. Consequently, further clinical studies involving a larger number of patients are warranted to confirm canakinumab's role in reducing inflammation and improving disease outcomes in Saudi SCD patients.
On the other hand, etanercept, a TNF-α receptor blocking agent, may halt inflammatory signaling initiated by TNF-α. Episodes of ischemia-reperfusion injury that commonly lead to vascular dysfunction and subsequent vaso-occlusion in SCD, may be minimized with etanercept therapy [24]. Long-term exposure of mice with SCD to etanercept resulted in a reduction of acute disease manifestations, including vaso-occlusion, responses to pain stimuli, leukocytes, and inflammatory biomarkers [83]. In addition, two case reports of patients with rheumatoid arthritis and SCD were treated successfully and safely with etanercept [84]. This encouraging evidence supports the need for conducting clinical trials to assess etanercept’s outcomes in SCD patients.
These suggested agents show promising potential in managing SCD severity and offer diverse features, including different routes of administration and cost advantages, which could facilitate their clinical adoption. For instance, simvastatin and allopurinol are widely used, available as oral formulations, and offered as affordable generics, which enhances their accessibility for long-term use [85,86]. In contrast, monoclonal antibodies also impact disease severity pathways; however, they require subcutaneous injections, often administered during hospital visits every 4–8 weeks, making them less convenient. Additionally, their higher costs compared to oral medications may limit accessibility [87,88]. Thus, oral agents offer greater convenience as they can be taken at home daily, potentially improving adherence, especially in chronic conditions like SCD. While injectable biologics may be more potent, they present challenges related to frequent healthcare interactions, administration, and higher costs. Strategically selecting these medications based on patient compliance and healthcare setting considerations may provide valuable therapeutic options for managing SCD complications.
The complex pathophysiology of SCD necessitates targeting multiple possible disease mechanisms. Thus, a multi-drug approach is suggested to address factors such as fetal hemoglobin modulation, cell adhesions and sickling, inflammation, ischemia/reperfusion, oxidative stress, coagulation, and free heme toxicity. To achieve this, a combination of drug therapies with different mechanisms is warranted [24].
In exploring new potential drug targets, we successfully identified the druggability pocket in several previously non-targeted gene products within this pipeline. We strongly recommend OR gene clusters as novel targets, particularly OR51V1, OR52A1, and OR51B5. Important mutations in these genes—such as rs7933549 (missense), rs112098990 (frameshift), and rs147062602 (frameshift), respectively—are commonly carried by SCD individuals and are associated with defects in protein function and subsequent degradation. As shown in the GTEx portal, this gene cluster is notably expressed in whole blood and erythroid cells near the β-globin cluster, which plays a regulatory role in hemoglobinopathies [89,90,91]. Although these are pseudogenes—non-functional DNA segments that do not directly contribute to phenotypic traits—they may regulate gene expression and function. Numerous genetic variants mapped to these gene products have been reported in the literature with multiple SCD phenotypes from previous GWAS and large meta-analyses, providing strong evidence of OR genes involvement in hematological parameters and SCD complications [18,25]. The reported traits associated with various mutations in the OR genes include level measurements of hemoglobin, hematocrit, erythrocyte count, mean corpuscular volume, red blood cell distribution width, mean corpuscular hemoglobin concentration, mean reticulocyte volume, fetal hemoglobin level, leukocyte count, monocytes, neutrophils, CRP, and platelet levels. Clinical complications such as hemolysis, thromboembolism, ischemia, and cardiomyopathy in SCD have also been previously linked to these genes [90,91,92].
The pipeline also suggests TRIM 6, SIDT2, and CADMD3 as novel, promising therapeutic targets. TRIM family proteins, including TRIM6, TRIM22, and TRIM34, are involved in innate immune modulation, cell cycle progression, and transcriptional regulation, suggesting they may influence inflammation, oxidative stress, hematopoietic stem cell differentiation, and immune responses [93]. These processes are critical to SCD pathophysiology and its complications. Furthermore, the SIDT2 gene product may contribute to SCD-related outcomes, as its dysregulation is linked to several metabolic traits associated with hepatic disorders, cardiovascular dysfunctions, and SCD-nephropathy [94,95]. Meanwhile, CADM3, which is involved in cell cycle regulation and epithelial proliferation [96], may impact hematological traits, thereby affecting blood cell formation and function in SCD. Each of these targets has a high druggable score, ranging from 0.7 to 0.86, highlighting them as strong candidates for future drug development.
3.5. Limitation
While the current study presents a promising framework to repurpose approved drugs as well as identify new drug targets for managing SCD severity, several limitations should be considered. First, our approach relies in part on in silico predictions and bioinformatic analyses, which, while powerful, cannot fully capture the complexities of drug-target interactions in vivo. This was partially addressed by the availability of various animal and small clinical trials that support our findings, yet larger, prospective clinical trials remain necessary. Additionally, the druggability scores that were used to prioritize the candidate targets for discovering new drug molecules were derived from computational models, which may not accurately reflect true biological conditions. Furthermore, the absence of direct clinical evidence concerning some of the proposed targets, such as SIDT2 and CADM3, in relation to SCD outcomes underscores the need for further experimental validation.
3.6. Conclusions
Utilizing an established pipeline enabled us to identify several promising candidates for drug repurposing, notably statins, immunomodulatory agents, and allopurinol. Additionally, novel targets—specifically OR and TRIM gene families—were identified for potential drug development to mitigate SCD severity and complications, with a focus on the Saudi population. The selection of these gene targets was supported by high pocket druggability scores and their involvement in key molecular pathways underlying the disease. These findings not only enhance our understanding of the genetic basis of SCD but also pave the way for more targeted and effective therapeutic approaches. Future clinical trials are essential to validate these computational predictions and translate them into actionable therapies that can improve outcomes for SCD patients.
Institutional Review Board Statement
The study received ethical approvals from the Institutional Review Boards (IRBs) of King Abdullah International Medical Research Center (KAIMRC), Saudi Arabia (Ref: IRBC/1414/19), and Virginia Commonwealth University (VCU), the United States of America (HM20030698). No informed consent was required for this study, as only publicly available data were used and no individual-level or identifiable information was accessed.
Authors’ contribution
AA: Conceptualization, Methodology, Data curation, Software, Formal analysis, Writing – original draft; MAA: Supervision, Project administration, Conceptualization, Methodology, Data curation, Formal analysis, Writing – review & editing; DW: Validation, Writing – review & editing; MA: Data curation, Resources, Writing – review & editing; SA: Investigation, Resources, Writing – review & editing; KA: Data curation, Software, Writing – review & editing; MK: Resources, Validation, Writing – review & editing; ASA: Supervision, Funding acquisition, Writing – review & editing; II: Conceptualization, Supervision, Writing – review & editing.
Funding
The financial support for this research project was obtained from KAIMRC, Riyadh, Saudi Arabia, with award number RC19-083-R.
Conflict of Interest
The authors declare no conflict of interest.
Acknowledgments
The authors extend their sincere gratitude to Elvin Price, Joseph McClay, Mary Peace McRae, and Patricia Slattum, at Virginia Commonwealth University, Virginia, USA, for their thorough review of the study and their constructive and insightful feedback, which significantly enhanced the quality of this work.
References
- Inusa BPD, Hsu LL, Kohli N, et al. Sickle Cell Disease-Genetics, Pathophysiology, Clinical Presentation and Treatment. Int J Neonatal Screen. 2019, 5, 20. [Google Scholar] [CrossRef]
- Belisário AR, Silva CM, Velloso-Rodrigues C, Viana MB. Genetic, laboratory and clinical risk factors in the development of overt ischemic stroke in children with sickle cell disease. Hematol Transfus Cell Ther. 2018, 40, 166–181. [Google Scholar] [CrossRef] [PubMed]
- Nnodu OE, Oron AP, Sopekan A, Akaba GO, Piel FB, Chao DL. Child mortality from sickle cell disease in Nigeria: a model-estimated, population-level analysis of data from the 2018 Demographic and Health Survey. Lancet Haematol. 2021, 8, e723–e731. [Google Scholar] [CrossRef]
- GBD 2021 Sickle Cell Disease Collaborators. Global, regional, and national prevalence and mortality burden of sickle cell disease, 2000-2021: a systematic analysis from the Global Burden of Disease Study 2021. Lancet Haematol. 2023, 10, e585–e599. [Google Scholar] [CrossRef]
- Ware RE, de Montalembert M, Tshilolo L, Abboud MR. Sickle cell disease. Lancet. 2017, 390, 311–323. [Google Scholar] [CrossRef]
- Jastaniah, W. Epidemiology of sickle cell disease in Saudi Arabia. Annals of Saudi Medicine. 2011, 31, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Bin Zuair A, Aldossari S, Alhumaidi R, Alrabiah M, Alshabanat A. The Burden of Sickle Cell Disease in Saudi Arabia: A Single-Institution Large Retrospective Study. Int J Gen Med. 2023, 16, 161–171. [Google Scholar] [CrossRef]
- el-Hazmi MA, al-Swailem AR, Warsy AS, al-Swailem AM, Sulaimani R, al-Meshari AA. Consanguinity among the Saudi Arabian population. J Med Genet. 1995, 32, 623–626. [Google Scholar] [CrossRef]
- Kavanagh PL, Fasipe TA, Wun T. Sickle Cell Disease: A Review. JAMA. 2022, 328, 57–68. [Google Scholar] [CrossRef]
- Salinas Cisneros G, Thein SL. Recent Advances in the Treatment of Sickle Cell Disease. Front Physiol. 2020, 11, 435. [Google Scholar] [CrossRef]
- Pfizer Voluntarily Withdraws All Lots of Sickle Cell Disease Treatment OXBRYTA® (voxelotor) From Worldwide Markets | Pfizer. Available online: https://www.pfizer.com/news/press-release/press-release-detail/pfizer-voluntarily-withdraws-all-lots-sickle-cell-disease (accessed on 27 September 2024).
- Revocation of authorisation for sickle cell disease medicine Adakveo | European Medicines Agency (EMA). May 26, 2023. Available online: https://www.ema.europa.eu/en/news/revocation-authorisation-sickle-cell-disease-medicine-adakveo (accessed on 30 August 2024).
- Commissioner O of the. FDA Approves First Gene Therapies to Treat Patients with Sickle Cell Disease. FDA. August 9, 2024. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-gene-therapies-treat-patients-sickle-cell-disease (accessed on 30 August 2024).
- Burt T, Dhillon S. Pharmacogenomics in early-phase clinical development. Pharmacogenomics. 2013, 14, 1085–1097. [Google Scholar] [CrossRef]
- Alghubayshi A, Wijesinghe D, Alwadaani D, et al. Unraveling the Complex Genomic Interplay of Sickle Cell Disease Among the Saudi Population: A Case-Control GWAS Analysis. Int J Mol Sci. 2025, 26, 2817. [Google Scholar] [CrossRef]
- Tragante V, Hemerich D, Alshabeeb M, et al. Druggability of Coronary Artery Disease Risk Loci. Circ Genom Precis Med. 2018, 11, e001977. [Google Scholar] [CrossRef]
- Pushpakom, S. Introduction and Historical Overview of Drug Repurposing Opportunities. Available online: https://books.rsc.org/books/edited-volume/1885/chapter/2472263/Introduction-and-Historical-Overview-of-Drug (accessed on 4 September 2024).
- Kirkham JK, Estepp JH, Weiss MJ, Rashkin SR. Genetic Variation and Sickle Cell Disease Severity: A Systematic Review and Meta-Analysis. JAMA Netw Open. 2023, 6, e2337484. [Google Scholar] [CrossRef]
- Pincez T, Ashley-Koch AE, Lettre G, Telen MJ. Genetic Modifiers of Sickle Cell Disease. Hematol Oncol Clin North Am. 2022, 36, 1097–1124. [Google Scholar] [CrossRef]
- Page GP, Kanias T, Guo YJ, et al. Multiple-ancestry genome-wide association study identifies 27 loci associated with measures of hemolysis following blood storage. J Clin Invest. 2021, 131, e146077. [Google Scholar] [CrossRef] [PubMed]
- Pritchard JLE, O’Mara TA, Glubb DM. Enhancing the Promise of Drug Repositioning through Genetics. Front Pharmacol. 2017, 8, 896. [Google Scholar] [CrossRef] [PubMed]
- Metaferia B, Cellmer T, Dunkelberger EB, et al. Phenotypic screening of the ReFRAME drug repurposing library to discover new drugs for treating sickle cell disease. Proc Natl Acad Sci U S A. 2022, 119, e2210779119. [Google Scholar] [CrossRef]
- Agu PC, Afiukwa CA, Orji OU, et al. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management. Sci Rep. 2023, 13, 13398. [Google Scholar] [CrossRef]
- Telen MJ, Malik P, Vercellotti GM. Therapeutic strategies for sickle cell disease: towards a multi-agent approach. Nat Rev Drug Discov. 2019, 18, 139–158. [Google Scholar] [CrossRef]
- Alshabeeb MA, Alwadaani D, Al Qahtani FH, et al. Impact of Genetic Variations on Thromboembolic Risk in Saudis with Sickle Cell Disease. Genes (Basel). 2023, 14, 1919. [Google Scholar] [CrossRef]
- Driss A, Asare KO, Hibbert JM, Gee BE, Adamkiewicz TV, Stiles JK. Sickle Cell Disease in the Post Genomic Era: A Monogenic Disease with a Polygenic Phenotype. Genomics Insights. 2009, 2009, 23–48. [Google Scholar]
- Cannon M, Stevenson J, Stahl K, et al. DGIdb 5.0: rebuilding the drug-gene interaction database for precision medicine and drug discovery platforms. Nucleic Acids Res. 2024, 52, D1227–D1235. [Google Scholar] [CrossRef]
- VigiAccess. Available online: https://www.vigiaccess.org/ (accessed on 4 September 2024).
- Kuhn M, Letunic I, Jensen LJ, Bork P. The SIDER database of drugs and side effects. Nucleic Acids Res. 2016, 44, D1075–1079. [Google Scholar] [CrossRef]
- Kuhn M, Szklarczyk D, Franceschini A, von Mering C, Jensen LJ, Bork P. STITCH 3: zooming in on protein-chemical interactions. Nucleic Acids Res. 2012, 40, D876–880. [Google Scholar] [CrossRef] [PubMed]
- Van Bever E, Wirtz VJ, Azermai M, et al. Operational rules for the implementation of INN prescribing. Int J Med Inform. 2014, 83, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Volkamer A, Kuhn D, Rippmann F, Rarey M. DoGSiteScorer: a web server for automatic binding site prediction, analysis and druggability assessment. Bioinformatics. 2012, 28, 2074–2075. [Google Scholar] [CrossRef]
- Berman H, Henrick K, Nakamura H, Markley JL. The worldwide Protein Data Bank (wwPDB): ensuring a single, uniform archive of PDB data. Nucleic Acids Res. 2007, 35, D301–303. [Google Scholar] [CrossRef]
- Jumper J, Evans R, Pritzel A, et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Callaway, E. Chemistry Nobel goes to developers of AlphaFold AI that predicts protein structures. Nature. 2024, 634, 525–526. [Google Scholar] [CrossRef]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 4 September 2024).
- Home - GEO - NCBI. Available online: https://www.ncbi.nlm.nih.gov/geo/ (accessed on 4 September 2024).
- GTEx Portal. Available online: https://www.gtexportal.org/home/ (accessed on 4 September 2024).
- Kirkham JK, Estepp JH, Weiss MJ, Rashkin SR. Genetic Variation and Sickle Cell Disease Severity: A Systematic Review and Meta-Analysis. JAMA Netw Open. 2023, 6, e2337484. [Google Scholar] [CrossRef] [PubMed]
- Kang H, Pan S, Lin S, Wang YY, Yuan N, Jia P. PharmGWAS: a GWAS-based knowledgebase for drug repurposing. Nucleic Acids Res. 2024, 52, D972–D979. [Google Scholar] [CrossRef] [PubMed]
- Ochoa D, Karim M, Ghoussaini M, Hulcoop DG, McDonagh EM, Dunham I. Human genetics evidence supports two-thirds of the 2021 FDA-approved drugs. Nat Rev Drug Discov. 2022, 21, 551. [Google Scholar] [CrossRef]
- Ullah, A. Perspective Chapter: Technological Advances in Population Genetics. IntechOpen; 2025. [CrossRef]
- Ku CS, Loy EY, Pawitan Y, Chia KS. The pursuit of genome-wide association studies: where are we now? J Hum Genet. 2010, 55, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Reay WR, Cairns MJ. Advancing the use of genome-wide association studies for drug repurposing. Nat Rev Genet. 2021, 22, 658–671. [Google Scholar] [CrossRef]
- Drug Repurposing (DR): An Emerging Approach in Drug Discovery | IntechOpen. Available online: https://www.intechopen.com/chapters/72744 (accessed on 4 September 2024).
- Nanda H, Ponnusamy N, Odumpatta R, Jeyakanthan J, Mohanapriya A. Exploring genetic targets of psoriasis using genome wide association studies (GWAS) for drug repurposing. 3 Biotech. 2020, 10, 43. [Google Scholar] [CrossRef]
- Xu J, Mao C, Hou Y, et al. Interpretable deep learning translation of GWAS and multi-omics findings to identify pathobiology and drug repurposing in Alzheimer’s disease. Cell Rep. 2022, 41, 111717. [Google Scholar] [CrossRef]
- Chowdhury FA, Colussi N, Sharma M, et al. Fatty acid nitroalkenes - Multi-target agents for the treatment of sickle cell disease. Redox Biol. 2023, 68, 102941. [Google Scholar] [CrossRef]
- Adam SS, Hoppe C. Potential role for statins in sickle cell disease. Pediatr Blood Cancer. 2013, 60, 550–557. [Google Scholar] [CrossRef]
- Bereal-Williams C, Machado RF, McGowan V, et al. Atorvastatin reduces serum cholesterol and triglycerides with limited improvement in vascular function in adults with sickle cell anemia. Haematologica. 2012, 97, 1768–1770. [Google Scholar] [CrossRef]
- Ridker PM, Danielson E, Fonseca FAH, et al. Rosuvastatin to Prevent Vascular Events in Men and Women with Elevated C-Reactive Protein. N Engl J Med. 2008, 359, 2195–2207. [Google Scholar] [CrossRef] [PubMed]
- Wong K, Lai WK, Jackson DE. HLA Class II regulation of immune response in sickle cell disease patients: Susceptibility to red blood cell alloimmunization (systematic review and meta-analysis). Vox Sang. 2022, 117, 1251–1261. [Google Scholar] [CrossRef]
- Tamouza R, Neonato MG, Busson M, et al. Infectious complications in sickle cell disease are influenced by HLA class II alleles. Hum Immunol. 2002, 63, 194–199. [Google Scholar] [CrossRef]
- Mahdi N, Al-Ola K, Al-Subaie AM, et al. HLA class II haplotypes distinctly associated with vaso-occlusion in children with sickle cell disease. Clin Vaccine Immunol. 2008, 15, 729–731. [Google Scholar] [CrossRef] [PubMed]
- Martins JO, Pagani F, Dezan MR, et al. Impact of HLA-G +3142C>G on the development of antibodies to blood group systems other than the Rh and Kell among sensitized patients with sickle cell disease. Transfus Apher Sci. 2022, 61, 103447. [Google Scholar] [CrossRef] [PubMed]
- Akaba K, Nwogoh B, Oshatuyi O. Determination of von Willebrand factor level in patient with sickle cell diseasein vaso-occlusive crisis. Res Pract Thromb Haemost. 2020, 4, 902–905. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar A, Serban C, Ursoniu S, et al. The impact of statin therapy on plasma levels of von Willebrand factor antigen. Systematic review and meta-analysis of randomised placebo-controlled trials. Thromb Haemost. 2016, 115, 520–532. [Google Scholar] [CrossRef]
- Hoppe C, Kuypers F, Larkin S, Hagar W, Vichinsky E, Styles L. A pilot study of the short-term use of simvastatin in sickle cell disease: effects on markers of vascular dysfunction. Br J Haematol. 2011, 153, 655–663. [Google Scholar] [CrossRef]
- Hoppe C, Jacob E, Styles L, Kuypers F, Larkin S, Vichinsky E. Simvastatin reduces vaso-occlusive pain in sickle cell anaemia: a pilot efficacy trial. Br J Haematol. 2017, 177, 620–629. [Google Scholar] [CrossRef]
- Xi C, Palani C, Takezaki M, et al. Simvastatin-Mediated Nrf2 Activation Induces Fetal Hemoglobin and Antioxidant Enzyme Expression to Ameliorate the Phenotype of Sickle Cell Disease. Antioxidants (Basel). 2024, 13, 337. [Google Scholar] [CrossRef]
- López-López S, Romero de Ávila MJ, Hernández de León NC, et al. NOTCH4 Exhibits Anti-Inflammatory Activity in Activated Macrophages by Interfering With Interferon-γ and TLR4 Signaling. Front Immunol. 2021, 12, 734966. [Google Scholar] [CrossRef]
- Zhou B, Lin W, Long Y, et al. Notch signaling pathway: architecture, disease, and therapeutics. Signal Transduct Target Ther. 2022, 7, 95. [Google Scholar] [CrossRef] [PubMed]
- Fingerlin TE, Murphy E, Zhang W, et al. Genome-wide association study identifies multiple susceptibility loci for pulmonary fibrosis. Nat Genet. 2013, 45, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Field JJ, Burdick MD, DeBaun MR, et al. The role of fibrocytes in sickle cell lung disease. PLoS One. 2012, 7, e33702. [Google Scholar] [CrossRef]
- Adeyinka A, Bashir K. Tumor Lysis Syndrome. In: StatPearls. StatPearls Publishing; 2024. Available online: http://www.ncbi.nlm.nih.gov/books/NBK518985/ (accessed on 4 September 2024).
- Pritchard KA Jr, Besch TL, Wang J, et al. Differential Effects of Lovastatin and Allopurinol Therapy in a Murine Model of Sickle Cell Disease. Blood. 2005, 106, 2325. [Google Scholar] [CrossRef]
- Kelkar A, Kuo A, Frishman WH. Allopurinol as a cardiovascular drug. Cardiol Rev. 2011, 19, 265–271. [Google Scholar] [CrossRef]
- Wood KC, Hebbel RP, Granger DN. Endothelial cell NADPH oxidase mediates the cerebral microvascular dysfunction in sickle cell transgenic mice. FASEB J. 2005, 19, 989–991. [Google Scholar] [CrossRef]
- Kaul DK, Liu XD, Choong S, Belcher JD, Vercellotti GM, Hebbel RP. Anti-inflammatory therapy ameliorates leukocyte adhesion and microvascular flow abnormalities in transgenic sickle mice. Am J Physiol Heart Circ Physiol. 2004, 287, H293–301. [Google Scholar] [CrossRef]
- Dashti M, Al-Matrouk A, Channanath A, Hebbar P, Al-Mulla F, Thanaraj TA. Distribution of HLA-B Alleles and Haplotypes in Qatari: Recommendation for Establishing Pharmacogenomic Markers Screening for Drug Hypersensitivity. Front Pharmacol. 2022, 13, 891838. [Google Scholar] [CrossRef]
- Dean L, Kane M. Allopurinol Therapy and HLA-B*58:01 Genotype. In: Pratt VM, Scott SA, Pirmohamed M, Esquivel B, Kattman BL, Malheiro AJ, eds. Medical Genetics Summaries. National Center for Biotechnology Information (US); 2012. Available online: http://www.ncbi.nlm.nih.gov/books/NBK127547/ (accessed on 4 September 2024).
- Xolair (omalizumab) FDA Approval History. Drugs.com. Available online: https://www.drugs.com/history/xolair.html (accessed on 4 September 2024).
- Kaplan AP, Giménez-Arnau AM, Saini SS. Mechanisms of action that contribute to efficacy of omalizumab in chronic spontaneous urticaria. Allergy. 2017, 72, 519–533. [Google Scholar] [CrossRef]
- Benjamin EJ, Dupuis J, Larson MG, et al. Genome-wide association with select biomarker traits in the Framingham Heart Study. BMC Med Genet. 2007, 8 (Suppl 1). [Google Scholar] [CrossRef]
- Al-Suliman A, Elsarraf NA, Baqishi M, Homrany H, Bousbiah J, Farouk E. Patterns of mortality in adult sickle cell disease in the Al-Hasa region of Saudi Arabia. Ann Saudi Med. 2006, 26, 487–488. [Google Scholar] [CrossRef]
- Alhaj Zeen M, Mohamed NE, Mady AF, et al. Predictors of Mortality in Adults With Sickle Cell Disease Admitted to the Intensive Care Unit in King Saud Medical City, Saudi Arabia. Cureus. 2023, 15, e38817. [Google Scholar] [CrossRef]
- An P, Barron-Casella EA, Strunk RC, Hamilton RG, Casella JF, DeBaun MR. Elevation of IgE in children with sickle cell disease is associated with doctor diagnosis of asthma and increased morbidity. J Allergy Clin Immunol. 2011, 127, 1440–1446. [Google Scholar] [CrossRef] [PubMed]
- Awojoodu AO, Keegan PM, Lane AR, et al. Acid sphingomyelinase is activated in sickle cell erythrocytes and contributes to inflammatory microparticle generation in SCD. Blood. 2014, 124, 1941–1950. [Google Scholar] [CrossRef]
- Rees DC, Kilinc Y, Unal S, et al. A randomized, placebo-controlled, double-blind trial of canakinumab in children and young adults with sickle cell anemia. Blood. 2022, 139, 2642–2652. [Google Scholar] [CrossRef]
- Fernández-Ruiz M, Aguado JM. Risk of infection associated with anti-TNF-α therapy. Expert Rev Anti Infect Ther. 2018, 16, 939–956. [Google Scholar] [CrossRef]
- Contejean A, Janssen C, Orsini-Piocelle F, Zecchini C, Charlier C, Chouchana L. Increased risk of infection reporting with anti-BCMA bispecific monoclonal antibodies in multiple myeloma: A worldwide pharmacovigilance study. Am J Hematol. 2023, 98, E349–E353. [Google Scholar] [CrossRef]
- Novelli, EM. Extinguishing the fire in sickle cell anemia. Blood. 2022, 139, 2578–2580. [Google Scholar] [CrossRef]
- Solovey A, Somani A, Belcher JD, et al. A monocyte-TNF-endothelial activation axis in sickle transgenic mice: Therapeutic benefit from TNF blockade. Am J Hematol. 2017, 92, 1119–1130. [Google Scholar] [CrossRef]
- Adelowo O, Edunjobi AS. Juvenile idiopathic arthritis coexisting with sickle cell disease: two case reports. BMJ Case Rep. 2011, 2011, bcr1020114889. [Google Scholar] [CrossRef]
- Simvastatin Prices, Coupons, Copay Cards & Patient Assistance - Drugs.com. Available online: https://www.drugs.com/price-guide/simvastatin (accessed on 4 September 2024).
- Allopurinol Prices, Coupons, Copay Cards & Patient Assistance - Drugs.com. Available online: https://www.drugs.com/price-guide/allopurinol (accessed on 4 September 2024).
- Ilaris Prices, Coupons, Copay Cards; Patient Assistance. Drugs.com. Available online: https://www.drugs.com/price-guide/ilaris (accessed on 4 September 2024).
- Xolair Prices, Coupons, Copay Cards & Patient Assistance - Drugs.com. Available online: https://www.drugs.com/price-guide/xolair (accessed on 4 September 2024).
- Bulger M, Bender MA, van Doorninck JH, et al. Comparative structural and functional analysis of the olfactory receptor genes flanking the human and mouse beta-globin gene clusters. Proc Natl Acad Sci U S A. 2000, 97, 14560–14565. [Google Scholar] [CrossRef] [PubMed]
- Yang Q, Kathiresan S, Lin JP, Tofler GH, O’Donnell CJ. Genome-wide association and linkage analyses of hemostatic factors and hematological phenotypes in the Framingham Heart Study. BMC Med Genet. 2007, 8 (Suppl. 1). [Google Scholar] [CrossRef]
- Feingold EA, Penny LA, Nienhuis AW, Forget BG. An olfactory receptor gene is located in the extended human beta-globin gene cluster and is expressed in erythroid cells. Genomics. 1999, 61, 15–23. [Google Scholar] [CrossRef]
- Chen Z, Zhao H, Fu N, Chen L. The diversified function and potential therapy of ectopic olfactory receptors in non-olfactory tissues. J Cell Physiol. 2018, 233, 2104–2115. [Google Scholar] [CrossRef]
- Uchil PD, Hinz A, Siegel S, et al. TRIM protein-mediated regulation of inflammatory and innate immune signaling and its association with antiretroviral activity. J Virol. 2013, 87, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Qian D, Cong Y, Wang R, Chen Q, Yan C, Gong D. Structural insight into the human SID1 transmembrane family member 2 reveals its lipid hydrolytic activity. Nat Commun. 2023, 14, 3568. [Google Scholar] [CrossRef]
- Geng MY, Wang L, Song YY, et al. Sidt2 is a key protein in the autophagy-lysosomal degradation pathway and is essential for the maintenance of kidney structure and filtration function. Cell Death Dis. 2021, 13, 7. [Google Scholar] [CrossRef]
- Estelius J, Lengqvist J, Ossipova E, et al. Mass spectrometry-based analysis of cerebrospinal fluid from arthritis patients—immune-related candidate proteins affected by TNF blocking treatment. Arthritis Res Ther. 2019, 21, 60. [Google Scholar] [CrossRef]
Figure 1.
The Pipeline for Drug Repurposing and Novel Drug Target Identification.
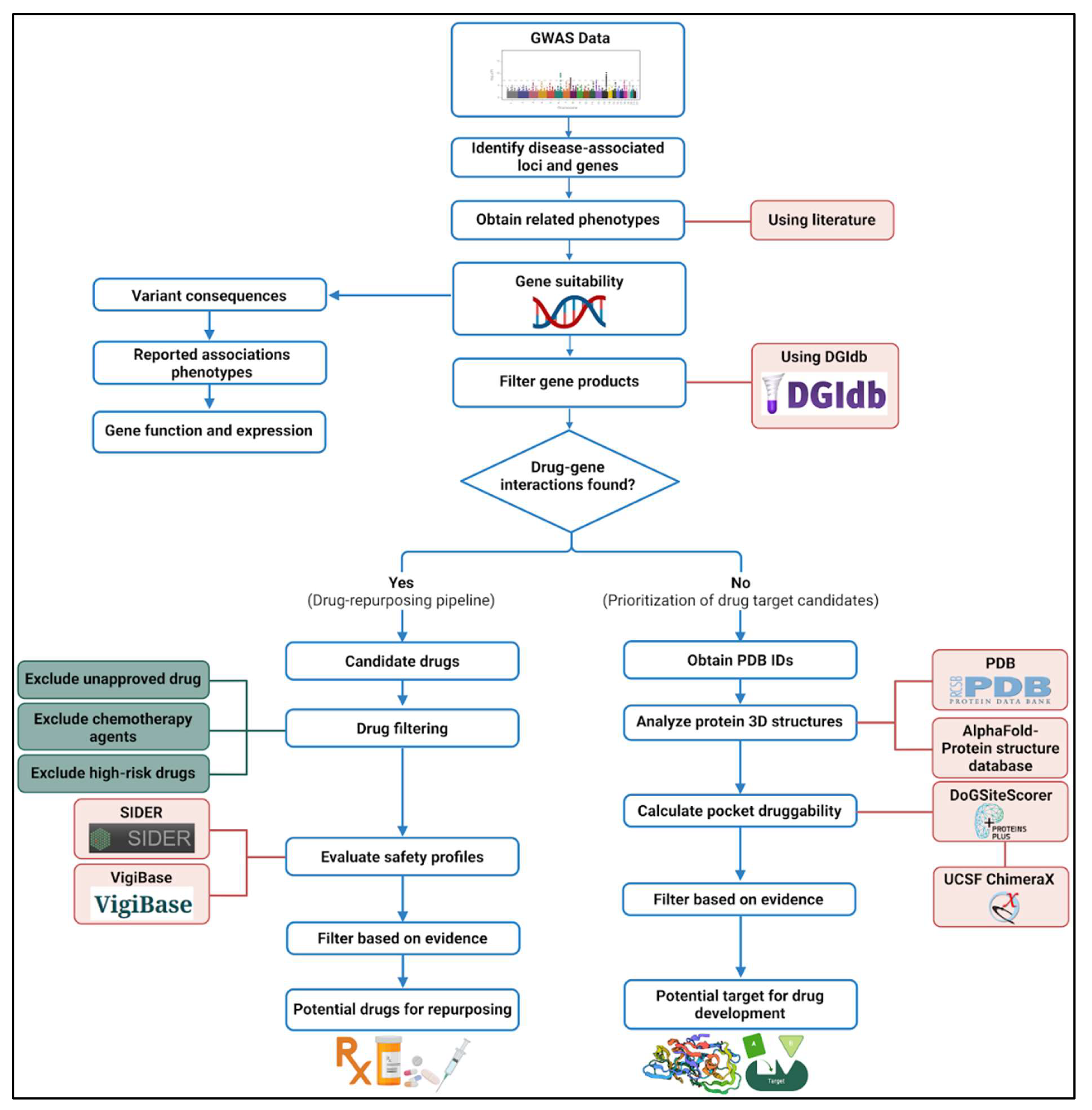
Figure 2.
: Three-dimensional structural representations of selected proposed proteins. Legend: A) OR51V1, B) OR52A1, C) OR51B5, D) TRIM6, E) SIDT2, and F) CADM3. The predicted optimal druggable binding site for each protein is highlighted in green.
Figure 2.
: Three-dimensional structural representations of selected proposed proteins. Legend: A) OR51V1, B) OR52A1, C) OR51B5, D) TRIM6, E) SIDT2, and F) CADM3. The predicted optimal druggable binding site for each protein is highlighted in green.
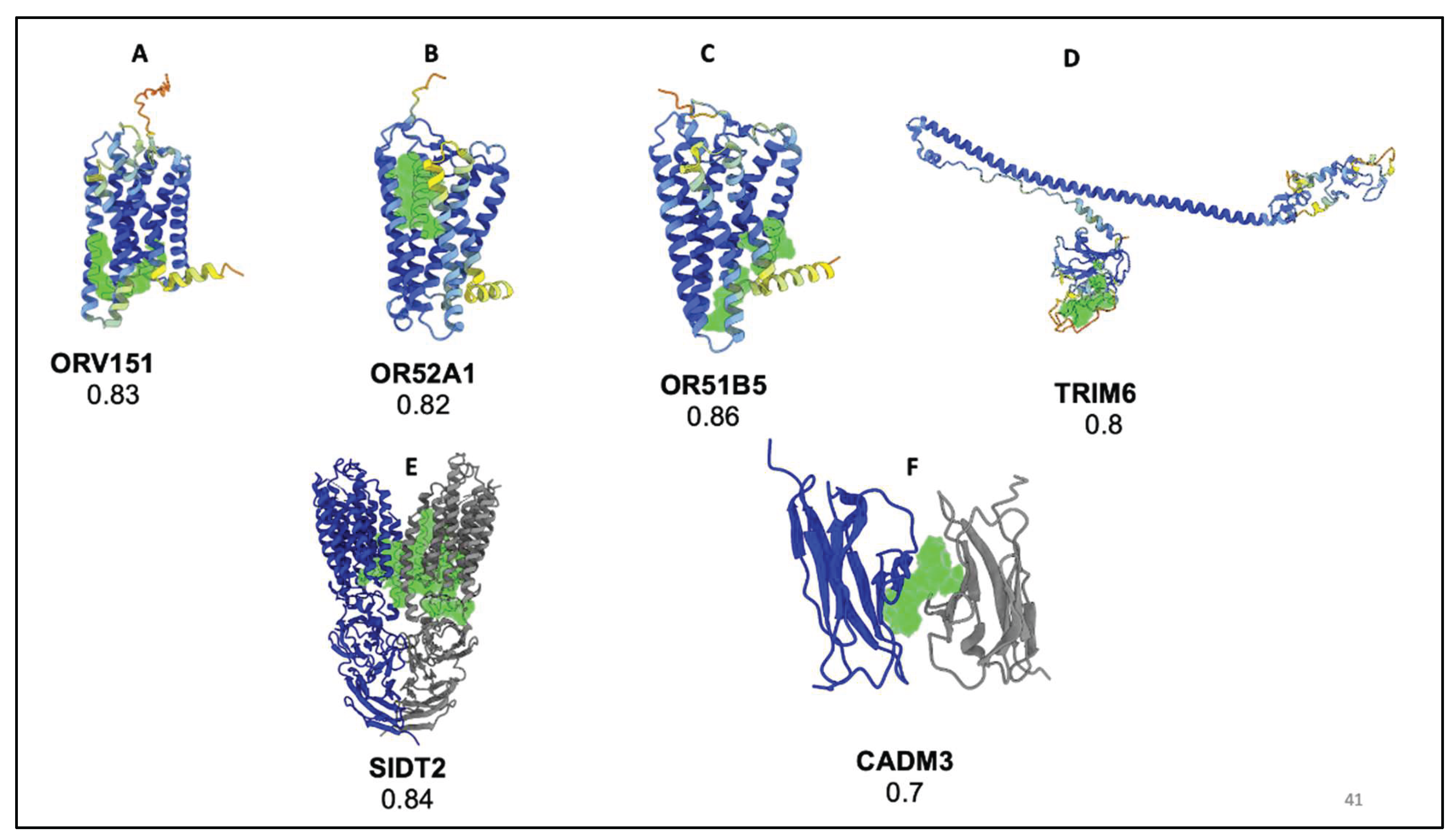

Table 1.
Predicted Drug-Gene Interactions Identified in DGI Database (V 5.0).
| SNP * | Gene * | Interacting drug | ATC code (s) | Interaction Score |
|---|---|---|---|---|
| rs3135006 | HLA-DQB1 | Infliximab | L04AB02 | 0.23 |
| Aspirin | B01AC06/ N02BA01 | 0.10 | ||
| rs2395522 | HLA-DRB1 | Pitavastatin | C10AA08 | 0.48 |
| Azathioprine | L04AX01 | 0.06 | ||
| Pravastatin | C10AA03 | 0.09 | ||
| Atorvastatin | C10AA05 | 0.04 | ||
| Simvastatin | C10AA01 | 0.05 | ||
| Rosuvastatin | C10AA07 | 0.10 | ||
| Fluvastatin | C10AA04 | 0.20 | ||
| Tocilizumab | L04AC07 | 0.48 | ||
| Adalimumab | L04AB04 | 0.07 | ||
| Rilonacept | L04AC04 | 0.80 | ||
| Canakinumab | L04AC08 | 1.19 | ||
| Anakinra | L04AC03 | 0.27 | ||
| Etanercept | L04AB01 | 0.09 | ||
| Infliximab | L04AB02 | 0.23 | ||
| Glatiramer | L03AX13 | 0.51 | ||
| Aspirin | B01AC06/ N02BA01 | 0.03 | ||
| rs2844806 | HLA-A | Fenofibrate | C10AB05 | 0.15 |
| Peginterferon alfa-2a | L03AB11 | 0.13 | ||
| Peginterferon alfa-2b | L03AB10 | 0.12 | ||
| rs2524035 | HLA-G | Simvastatin | C10AA01 | 1.05 |
| rs2494250 | FCER1A | Omalizumab | R03DX05 | 0.94 |
| rs3132946 rs3132940 rs9267898 rs3096702 |
NOTCH4 | Allopurinol | M04AA01 | 0.58 |
| * Genetic variants and mapped genes associated with sickle cell disease severity identified in our genome wide association study involving Saudi patients [15]. | ||||
Table 2.
Most Suitable Drug Targets Based on Predicted Pocket Interactions Utilizing DogSiteScorer Database.
Table 2.
Most Suitable Drug Targets Based on Predicted Pocket Interactions Utilizing DogSiteScorer Database.
| SNP | Gene | Pocket Score | PDB Code | AlphaFold Code |
|---|---|---|---|---|
| rs7933549 | OR51V1 | 0.83 | ---- | AF-Q9H2C8-F1 |
| rs112098990 | OR52A1 | 0.82 | ---- | AF-Q9UKL2-F1 |
| rs2472530 | OR52A5 | 0.82 | ---- | AF-Q9H2C5-F1 |
| rs147062602 rs10838058 rs10837853 rs78253695 rs180750244 |
OR51B5 | 0.86 | ---- | AF-Q9H339-F1 |
| rs12361955 | OR51S1 | 0.84 | ---- | AF-Q8NGJ8-F1 |
| rs3740999 rs11038294 rs12272467 |
TRIM6 | 0.8 | ---- | AF-Q9C030-F1 |
| rs67573252 | TRIM22 | 0.87 | ---- | AF-Q8IYM9-F1 |
| rs2342380 | TRIM34 | 0.82 | 2EGP | ---- |
| rs10535646 | SIDT2 | 0.84 | 7Y68 | ---- |
| rs3845624 | CADM3 | 0.7 | 1Z9M | ---- |
| MPTX1 | 0.81 | ---- | AF-A8MV57-F1 | |
| * Genetic variants and mapped genes associated with sickle cell disease severity identified in our genome wide association study involving Saudi patients [15]. | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



