Submitted:
20 June 2025
Posted:
23 June 2025
You are already at the latest version
Abstract
Porcine circovirus type 2 (PCV2) infection causes porcine circovirus disease (PCVD), a global immunosuppressive disease in pigs. Its clinical manifestations include post-weaning multisystemic wasting syndrome (PMWS) and porcine dermatitis and nephropathy syndrome (PDNS), which cause significant economic losses to the swine industry. The Cap protein, which is the major protective antigen of PCV2, can self-assemble to form virus-like particles (VLPs) when expression in the insect baculovirus expression system. Few studies have compared the expression of Cap proteins in different baculovirus expression systems. In this study, we compared two commonly commercialized baculovirus construction systems to expressing the Cap protein in various insect cells. The results demonstrated that the flashBAC system expressing Cap protein at higher levels than the Bac-to-Bac system. Notably, when expressing four copies of the Cap protein, the flashBAC system achieved the highest protein yield in High Five cells, reaching 432 μg/mL at 5 days post-infection (dpi) at 27°C cultivation. Animal experiments confirmed that the purified Cap protein effectively induced specific antibody production in mice and swine. This study provides critical data for optimizing the production of PCV2 Cap protein, which is of great significance for reducing the production cost of PCV2 vaccines and improving industrial production efficiency.
Keywords:
Porcine circovirus type 2 (PCV2)
; Baculovirus
; Insect expression system
; flashBAC
; Bac-to-Bac
1. Introduction
Circoviruses are recognized as the smallest viruses known to infect animals. Four types of porcine circoviruses (PCV1–PCV4) have been sequentially identified in swine [1]. Porcine circovirus type 1 (PCV1), initially identified as the first member of the porcine circovirus family, has been characterized as a nonpathogenic virus [2]. PCV2 remains the main pathogen of related diseases. The pathogenicity of PCV3 remains controversial. PCV4 is a recently discovered pathogen, and its pathogenicity in pigs requires further investigation [3,4]. PCV2 infection causes porcine circovirus disease (PCVD), a global immunosuppressive disease in pigs. Its clinical manifestations include post-weaning multisystemic wasting syndrome (PMWS) and porcine dermatitis and nephropathy syndrome (PDNS), which cause significant economic losses to the swine industry [5,6]. PCV2 is classified into 8 genotypes (PCV2a–PCV2h) [7,8]. Recently, a novel genotype, PCV2i, was identified [9]. PCV2d is the predominant circulating subtype of PCV2 [5]. The PCV2 genome is a covalently closed circular single-stranded DNA (ssDNA), approximately 1.7 kilobases (kb) [10]. The software analysis identified 11 open reading frames (ORFs) in the PCV2 genome, but only 4 are utilized for protein expression [11]. ORF2 encodes the sole structural protein - the capsid (Cap) protein. The Cap protein represents the primary antigenic determinant of the virus, which serves as a critical determinant of the host immune response and vaccine efficacy [12].
For the prevention of PCVD, vaccination remains the primary strategy [6]. Virus-like particles (VLPs), as a type of subunit vaccine, have emerged as the preferred candidate due to their lack of nucleic acids, inability to replicate autonomously, and enhanced safety profile. The Cap protein can self-assemble into VLPs across various expression systems [13]. Although bacterial, yeast, and insect cell platforms are capable of expressing the Cap protein, commercially available vaccines (e.g., Ingelvac CircoFLEX® and Circumvent®) exclusively utilize the baculovirus-insect cell system [14,15,16]. This preference is likely attributed to its advantages in high-yield production, appropriate post-translational modifications, and efficient VLP assembly. While alternative expression systems such as E. coli and yeast are being explored for cost-effectiveness, they currently face challenges related to immunogenicity and scalability limitations [17,18].
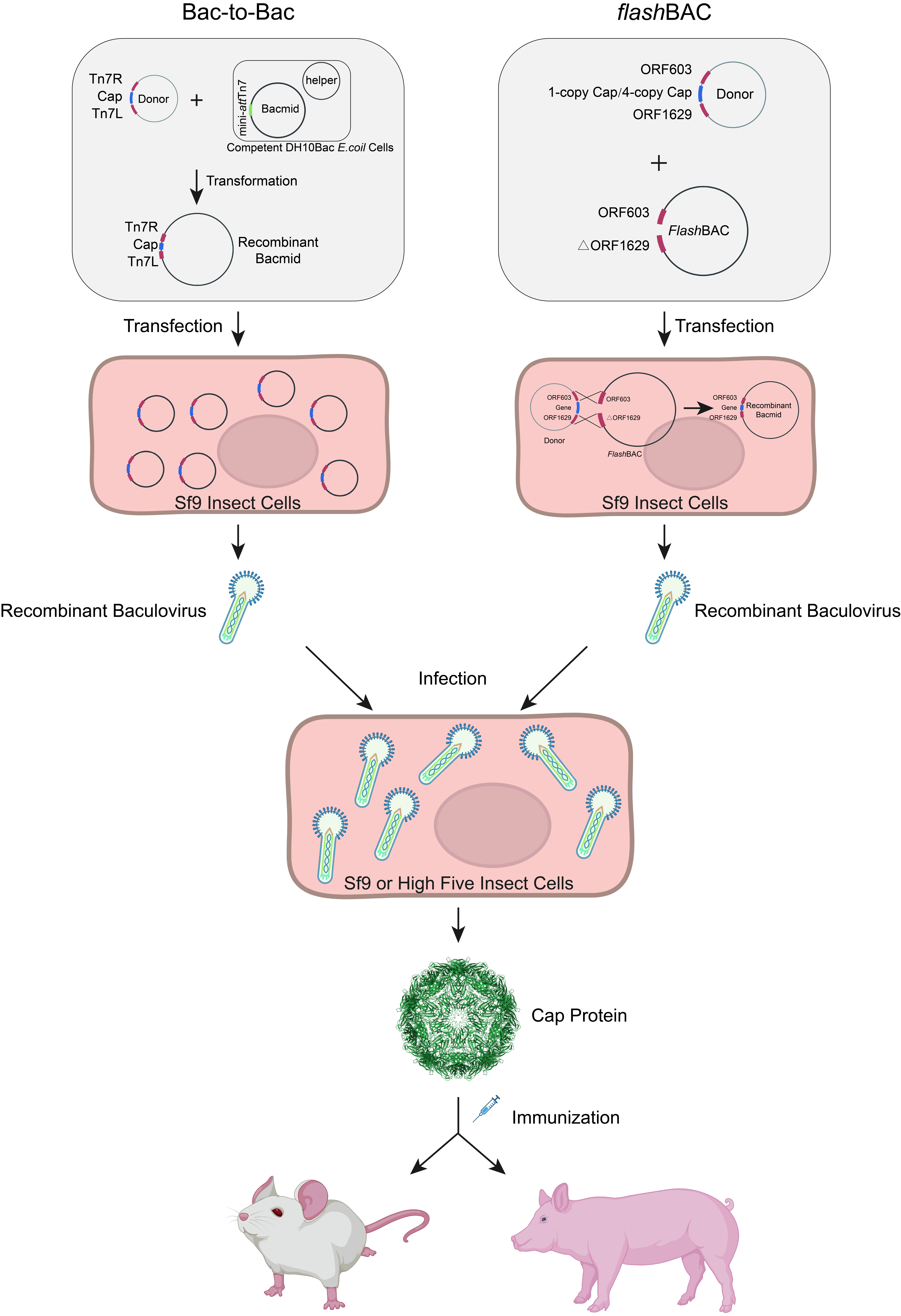
The Bac-to-Bac and flashBAC systems are two commonly commercialized baculovirus construction systems. Each system has its own advantages and limitations in terms of the operation procedures and application scope [19,20]. In the Bac-to-Bac system, the target gene is first cloned into a pFast series vector. The recombinant plasmid is then transformed into E. coli DH10Bac competent cells to generate the bacmid through site-specific transposition. Positive clones are selected via blue-white screening, followed by bacmid extraction and transfection into insect cells for viral packaging (Figure 1. A) [21]. In contrast, the flashBAC system employs compatible transfer vectors (e.g., pBAC or pOET series) to construct the expression cassette. The recombinant transfer vector is co-transfected with a linearized bacmid into insect cells, where homologous recombination occurs in vivo to generate the recombinant baculovirus directly (Figure 1. B) [19,22]. Although both systems are commercial, laboratories choose one system based on their existing platform and research costs, seldom comparing the two systems side by side [20]. Despite numerous studies have documented the application of both systems for Cap protein expression, applications of both systems in Cap protein expression, simultaneous expression comparisons remain unexplored. For certain proteins, increasing the gene copy number can enhance the expression level of recombinant proteins [23,24]. It remains unknown whether increasing the copy number of cap gene enhances protein expression in the insect baculovirus system.
In this study, we compared two commonly commercialized baculovirus construction systems for expressing the Cap protein in various insect cells. Our findings demonstrated that the flashBAC system achieved significantly higher Cap protein expression levels compared to the Bac-to-Bac system, with increased gene copy number of the Cap protein further enhancing expression efficiency. High Five insect cells demonstrated superior performance for Cap protein expression compared to Sf9 cells. Notably, when expressing four copies of the Cap protein, the flashBAC system achieved the highest protein yield of 432 μg/mL in High Five cells. Animal immunization experiments confirmed that the expressed Cap protein effectively induced specific antibody production in both mice and swine. This study provides critical data for optimizing the production of PCV2 Cap protein, which is of great significance for cost reduction in porcine circovirus vaccine manufacturing.
2. Material and Method
2.1. Construction of Vectors
The target gene Cap (GenBank accession no. MK751862.1) was codon-optimized for expression in Spodoptera frugiperda cells and synthesized by Zhejiang Sunya Biotechnology Co., Ltd. Two expression systems were employed: the pFast-Dual vector (Bac-to-Bac system) and the pBAC-1 vector (flashBAC system). Construction of the pFast-Cap Vector: The synthesized Cap gene was used as a template for polymerase chain reaction (PCR)amplification using PrimeSTAR® Max DNA Polymerase (Takara, R045Q, Shiga, Japan) with specific primers (pFast-Cap-F/R). The target fragment was purified by 1% agarose gel electrophoresis. The pFast-Dual vector was double-digested with BamHI (Takara, 1605, Shiga, Japan) and HindIII (Takara, 1615, Shiga, Japan), and the linearized vector was gel-purified. The insert and vector were ligated at a 3:1 molar ratio (insert:vector) using the ClonExpress Ultra One-Step Cloning Kit (Vazyme, C115-01, Nanjing, China) and transformed into DH5α competent cells (TransGen Biotech, CD201, Beijing, China); Positive clones were selected on Luria-Bertani (LB) agar plates containing 100 μg/mL ampicillin (Sangon Biotech, A610028, Shanghai, China) after overnight incubation at 37°C; Colony PCR and sequencing were performed for validation, and the positive recombinant plasmid was extracted using a Plasmid Extraction Kit (Tiangen, DP106, Beijing, China). Construction of the pBAC-1-Cap Vector: The same strategy was applied; The Cap gene was amplified using primers pBAC-1-Cap-F/R, and the pBAC-1 vector was digested with BamHI/HindIII for subsequent cloning. Construction of the pBAC-4×Cap Vector: A pUC19-4×Cap recombinant vector was synthesized by Shanghai Shangya Biotech; The multi-copy Cap gene fragment was obtained by double digestion with BglII (Takara, 1606, Shiga, Japan) and HindIII, then ligated into the similarly digested pBAC-1 vector using T4 DNA Ligase (NEB, M0202V) at a 5:1 insert-to-vector ratio; The ligation product was transformed into Sure competent cells (WEIDI, DL1065S, Shanghai, China). Positive clones were verified by sequencing, and the plasmid was extracted for downstream applications.
2.2. Construction of Bacmid
The Bac-to-Bac method requires the construction of bacmid by transfecting DH10Bac chemically competent cells (Weidi, DL1071, Shanghai, China) with an expression plasmid. The pFast-Cap plasmid was transformed into DH10Bac chemically competent cells, and the heat-shocked cells were plated on Luria-Bertani (LB) agar plates containing 50 μg/mL kanamycin (Sangon Biotech, A506636, Shanghai, China), 7 μg/mL gentamicin (Sangon Biotech, A428430, Shanghai, China), 7 μg/mL tetracycline (Sangon Biotech, A430165, Shanghai, China), 40 μg/mL X-gal (Sangon Biotech, A600083, Shanghai, China), and 40 μg/mL IPTG (Sangon Biotech, A600168, Shanghai, China). Positive white colonies were selected via blue-white screening and further verified by PCR using pFast-Cap-F/R, M13(40)/Tn7R and Tn7L/M13R primers. The bacmid DNA was extracted using the BAC/PAC DNA Kit (Omega Bio-Tek, D2156, Norcross, Georgia, USA) for subsequent baculovirus packaging.
2.3. Baculovirus Packaging and Amplification
Culture Sf9 cells in SF-SMF cell culture medium in a 27°C incubator. Ensure the cells are in the logarithmic growth phase (density approximately 1-2×106 cells/mL). Seed 2.5×106 cells in a 6-well plate with a culture volume of 2 mL. After 1 hour, replace the medium with 2.5 mL of Transfection Media (Oxford Expression Technologies, 500312, Oxford OX3 0BP, UK). For the Bac-to-Bac system, add 8 μg of Bacmid to 100 μL of Transfection Media, followed by 1.2 μL of BaculoFECTIN II (Oxford Expression Technologies, 300105, Oxford OX3 0BP, UK). For the flashBAC system, add 100 ng of flashBAC GOLD baculovirus DNA (Oxford Expression Technologies, 100202, Oxford OX3 0BP, UK) and 500 ng of pBAC-1-Cap/pBAC4×-1-Cap to 100 μL of Transfection Media, followed by 1.2 μL of BaculoFECTIN II (Oxford Expression Technologies, 300105, Oxford OX3 0BP, UK). Incubate the mixture at room temperature for 15 minutes before adding it to the 6-well plate. After 5 days, collect the cell supernatant to obtain the P1 generation of virus and store it at -80°C. To obtain the P2 generation of virus, inoculate Sf9 cells at a density of 2×106 cells/mL with multiplicity of infection (MOI)=0.05 of the P1 baculovirus stock. Collect the cell supernatant when the cell viability reaches 50%. The P3 generation of virus is obtained using the same method as for the P2 generation.
2.4. Identification and Titer Test of Baculoviruses
The second-generation (P2) baculovirus DNA was extracted using a viral genome DNA/RNA extraction kit (Tiangen, DP315, Beijing, China). PCR amplification was performed using baculovirus VP80-specific primers, and the target band was detected by agarose gel electrophoresis. Viral titer was determined using a GP64-specific antibody against baculovirus. Sf9 cells were seeded in a 96-well plate at a density of 7 × 106 cells diluted in 10 mL of medium, with 100 μL of the cell suspension added to each well. After incubation at 27°C for 30 minutes, the viral stock was serially diluted in 10-fold increments across 10 gradients. Each dilution was added to a column of 8 wells in the 96-well plate, with the last two columns left as blanks. The plate was incubated at 27°C for 4 days. Subsequently, the cells were fixed with 4% paraformaldehyde, permeabilized with 0.3% Triton X-100 in Phosphate-Buffered Saline (PBS), and blocked with 3% BSA in PBS. The cells were then incubated with a 1:1000 dilution of GP64 antibody (Santa Cruz, sc-65499, CA, USA) at 37°C for 2 hours, followed by three washes. A 1:2000 dilution of anti-mouse IgG (H+L) cross-adsorbed secondary antibody conjugated with Alexa Fluor™ 488 (Thermo, A-11001, MA, USA) was added and incubated at 37°C for 2 hours, followed by another three washes. Positive wells were observed under a fluorescence microscope, and the viral titer (TCID50/mL) was calculated using the Reed & Muench method.
2.5. The Expression of Cap Protein Was Detected by SDS-PAGE and ELISA
The protein sample was treated with protein loading buffer (TransGen Biotech, DL101-02, Beijing, China) at 95°C for 5 minutes. The treated sample was then loaded onto a 12% SDS-PAGE gel (Yamei, PG113, Shanghai, China), and electrophoresis was performed at 80V for 30 minutes, followed by 120V for 1 hour and 20 minutes. Protein bands were visualized by staining with Coomassie Brilliant Blue R-250. The concentration of the protein sample was measured using the Quantitative Determination for the Total Concentration of PCV2-Cap Protein ELISA Kit (Womei, FGPC292002, Suzhou, Jiangsu, China), with the specific methodology detailed in the manufacturer’s instructions.
2.6. Purification and Identification of Cap Protein
Purification of Cap protein, based on its properties, primarily employs cation exchange chromatography. A chromatography column was packed with SP Sepharose Fast Flow cation exchange resin (Bestchrom, AI0011, Jiaxing, Zhejiang, China) and equilibrated with 5 column volumes (CV) of equilibration buffer (20 mM sodium phosphate, pH 6.0) until the pH and conductivity of the effluent stabilized. The insect cell culture supernatant was centrifuged (12,000 × g, 30 min, 4°C), and the supernatant was filtered through a 0.45 μm membrane (Millipore, SLHP033N, Burlington, MA, USA). The sample was then diluted with equilibration buffer to achieve a conductivity below 5 mS/cm, and the pH was adjusted to 6.0. The processed sample was loaded onto the equilibrated cation exchange column at a flow rate of 1 mL/min to ensure efficient binding of the target protein. The column was washed with 10 CV of equilibration buffer to remove unbound impurities until the UV absorbance (280 nm) returned to baseline. A linear gradient elution was performed using equilibration buffer containing 1 M NaCl (Sangon Biotech, A610476, Shanghai, China) at a flow rate of 1 mL/min. The eluted peaks were collected while monitoring UV absorbance (280 nm). The fractions corresponding to the elution peak were pooled and analyzed for target protein purity using SDS-PAGE and Western blot. For Western blotting, proteins from an unstained PAGE gel were transferred to a PVDF membrane (Biorad, 1620177, CA, USA) using a wet transfer system. The membrane was blocked with 5% skim milk (Oxoid, LP0031B, MA, USA) in PBS for 2 hours, incubated overnight at 4°C with a 1:1000 dilution of primary antibody against PCV2 (Bioss, bs-10057R, Beijing, China) in PBS, washed five times with PBS-Tween, incubated with a 1:5000 dilution of anti-rabbit secondary antibody (Thermo, 31460, MA, USA) at 37°C for 2 hours, washed five times with PBS-Tween, and finally developed with ECL substrate (Vazyme, E423-01/02, Shanghai, China) before imaging with a gel imaging system.
2.7. Animal Immunity and Efficacy Evaluation
Evaluate the immunogenicity of the Cap protein in mice and pigs. Six-week-old BALB/c mice (Sipeifu, Beijing, China) were randomly divided into two groups, with 5 mice in each group. The vaccine group was immunized with 50 μg of Cap protein mixed 1:1 with a water-based adjuvant, while the control group was immunized with PBS mixed 1:1 with the same adjuvant. Blood was collected from the mice 5 weeks post-immunization, and the antibody titers in the serum were determined. Healthy weaned piglets aged 2 weeks were randomly divided into two groups, with 10 piglets in each group. The vaccine group was immunized with 100 μg of Cap protein mixed 1:1 with a water-based adjuvant, while the control group was immunized with PBS mixed 1:1 with the same volume of adjuvant. Blood was collected from the piglets 5 weeks post-immunization, and the antibody titers in the serum were determined. Serum samples were collected at weeks 0 (pre-immunization), 1, 4, 7, 10, 13, 16, and 19 post-immunization, and the antibody titers were measured. The antibody titers in piglet serum were determined by indirect ELISA (INGENASA, 11.PCV.K.1/5, Madrid, Spain) to detect Cap protein-specific antibodies. A sample was considered positive if the OD450 nm value was ≥0.5 and at least 2.1 times higher than the negative control. The antibody titers in mouse serum were measured by indirect ELISA (INGENASA, 11.PCV.K.1/5, Madrid, Spain), with the secondary antibody replaced by an HRP-labeled anti-mouse secondary antibody (Thermo, G-21040, MA, USA).
3. Result
3.1. Construction of Expression Vector and Bacmid
This experiment required the construction of three vectors: pFast-Cap, pBAC-1-Cap, and pBAC-4×Cap. Among them, pFast-Cap needed to be transfected into DH10Bac to generate bacmid, and the schematic diagram of the construction is shown in Figure 1. A. Construction of the pFast-Cap vector: The synthesized Cap gene was used as a template, and PCR amplification was performed with specific primers (pFast-Cap-F/R). The target fragment size was 746 bp, as confirmed by 1% agarose gel electrophoresis (Figure 2. B). The pFast-Dual vector was double-digested with BamHI/HindIII, yielding a target fragment of 5166 bp (Figure 2. C). Construction of the pBAC-1-Cap vector: Using the synthesized Cap gene as a template, the target gene was amplified with primers pBAC-1-Cap-F/R, resulting in a product of 749 bp (Figure 2. D). The pBAC-1 vector was double-digested with BamHI/HindIII, producing a target fragment of 5234 bp (Figure 2. E). Construction of the pBAC-4×Cap vector: The recombinant vector was synthesized by Shangya Biotechnology; The multi-copy Cap gene fragment (3753 bp, Figure 2. F) was obtained by BglII/HindIII double digestion of pUC19-4×Cap and ligated into the similarly digested pBAC-1 vector (5132 bp, Figure 2. G). The pFast-Cap plasmid was transformed into DH10Bac competent cells, yielding distinct blue and white colonies (Figure 2. H). The white colonies were positive clones, and PCR screening was performed using three primer pairs: M13 (40)/TnR-R, Cap-F/R, and TnL-F/M13R, which successfully amplified the target bands (Figure 2. I).
3.2. Identification and Titration of Baculovirus
To determine whether the baculovirus was successfully packaged, the DNA of the packaged viral cells was extracted and detected using VP80-specific primers. PCR results showed that baculovirus-specific bands, with a size of 193 bp, were successfully detected using the baculovirus VP80-specific primers (Figure 3. A). The results indicate that the baculovirus was successfully packaged. The 50% tissue culture infectious dose (TCID50) of the baculovirus was measured by immunofluorescence staining using a baculovirus GP64-specific antibody. The results showed that the viral titers of pFast-Cap, pBAC-1, and pBAC4×-1 were 7.35 log10TCID50/mL, 8.41 log10TCID50/mL, and 8.01 log10TCID50/mL, respectively (Figure 3. B and C).
3.3. Growth Parameters of Sf9 and High Five Cells after Baculovirus Infection
To determine the optimal time for harvesting the protein, the viable cell count, total cell count, viability, and cell diameter of Sf9 and High Five insect cells infected with baculovirus were measured every 24 hours until the cell viability dropped below 20%. Sf9 Cells total cell count initially increased and then decreased. On the first day post-infection, the cells proliferated normally, but after 24 hours, the proliferation rate of the infected group was significantly lower than that of the blank control group (Figure 4. A). Sf9 viable cell count also showed an initial increase followed by a decrease, rising during days 1–2 but declining after day 3 (Figure 4. B). Sf9 cell viability remained above 90% for the first three days but dropped by the fourth day. By the fifth day, the viability of some infected groups fell to 20%, whereas the control group also exhibited a decline but at a significantly slower rate compared to the infected group (Figure 4. C). Sf9 cell diameter first increased and then decreased with infection time. From days 1–3 post-infection, the cell diameter gradually increased, with the infected group showing a faster rate of increase than the control group. However, the diameter decreased on days 4–5 (Figure 4. D). High Five total cell count increased with infection time, peaking on the fifth day, though the proliferation rate of the infected group was notably lower than that of the control group (Figure 4. E). High Five viable cell count exhibited an initial rise followed by a decline, increasing during days 1–2 but decreasing after day 3, similar to the trend observed in Sf9 cells (Figure 4. F). High Five cell viability remained above 90% for the first three days but decreased by the fourth day, with some infected groups dropping to 20% viability by the fifth day (Figure 4. G). High Five cell diameter showed a trend of first decreasing, then increasing, and finally decreasing again. On day 1 post-infection, the diameter decreased, but from days 2–3, it gradually increased, with the infected group displaying a faster rate of enlargement than the control. By days 4–5, the cell diameter decreased again. (Figure 4. H).
3.4. Analysis of Cap Protein Expression in Baculovirus-Infected Sf9 and High Five Cell Lines
The constructed baculovirus pFastBac-Cap infected Sf9 cells, and the expression level of Cap protein increased with the number of infection days, reaching 164 μg/mL on 5 dpi. When pFastBac-Cap-infected High Five cells, the expression of Cap protein also increased with infection time, peaking at 129 μg/mL on 5 dpi (Figure 5. A and B). For the baculovirus pBAC-1-Cap infecting Sf9 cells, the expression of Cap protein rose with prolonged infection, reaching its highest level of 298 μg/mL on 4 dpi. Similarly, in High Five cells infected with pBAC-1-Cap, Cap protein expression increased over time, with the highest level of 392 μg/mL observed on 5 dpi. In the case of pBAC-4×Cap baculovirus infecting Sf9 cells, Cap protein expression initially increased and then decreased, peaking at 319 μg/mL on 4 dpi before declining to 287 μg/mL on 5 dpi (Figure 5. A and B). When pBAC-4×Cap infected High Five cells, Cap protein expression showed a similar trend, rising initially and then decreasing, with the highest expression level of 431 μg/mL on 4 dpi, followed by a drop to 395 μg/mL on 5 dpi (Figure 5. A and B).
3.5. Cap Protein Purification
To improve the purity of Cap, cell debris was removed by centrifugation, the culture supernatant was clarified using a 0.45 μm filter, and the Cap protein was finally purified by cation-exchange chromatography. The purified Cap was detected by Western blot using a specific antibody. We found that centrifugation effectively removed cell debris, and the cation-exchange purification yielded a higher concentration of Cap protein with a one-step elution showing better results (Figure 6. A). Western blot analysis of the purified protein using a Cap-specific antibody confirmed specific binding (Figure 6. B). The results demonstrate that cation-exchange chromatography can successfully purify the protein, and the purified product is the Cap protein.
3.6. Evaluation of Immune Effect of Cap Protein
The purified Cap protein was emulsified with adjuvant and used to immunize mice and pigs. Serum samples were collected at designated time points, and anti-Cap antibody titers were measured by ELISA (Figure 7.A). In mice, Cap-specific antibodies were detectable at 5 weeks post-immunization, with a peak titer of 1:12,800 (Figure 7.B). In pigs, antibody titers increased progressively from weeks 4 to 7 but declined thereafter, reaching levels comparable to the control group by week 10 and remaining undetectable through week 19 (Figure 7.C). These findings demonstrate that the Cap protein effectively induces an immune response, eliciting robust but transient antibody production in immunized animals.
4. Discussion
PCV2 infection causes PCVD, a global immunosuppressive disease in pigs. Its clinical manifestations include PMWS and PDNS, which cause significant economic losses to the swine industry [5,6]. Vaccination remains the primary strategy for preventing PCVD [6]. Cap, as the protective antigen of porcine circovirus, has been expressed in various protein expression systems, such as E. coli, yeast and the insect baculovirus system [25,26,27]. However, currently marketed products are exclusively expressed using the insect baculovirus system, indicating its superior suitability for protein expression. Common baculovirus construction systems include Bac-to-Bac and the homologous recombination method flashBAC [19,20], yet few laboratories have compared the impact of different baculovirus construction systems on protein expression. Our results demonstrate that the baculovirus constructed using flashBAC is more suitable for Cap protein expression than that constructed using the Bac-to-Bac system, regardless of whether Sf9 or High Five cells are employed. Furthermore, the insect cell line High Five cells are more conducive to Cap protein expression compared to Sf9 cells. Previous literature and patents report protein expression levels ranging from 50 to 150 μg/mL, with the highest patent-reported level reaching 200 μg/mL [28]. In this study, the maximum expression level achieved was 432 μg/mL, significantly surpassing the values reported in the literature and patents.
In addition to codon optimization, increasing the gene copy number can enhance protein expression [23,29]. In this study, the flashBAC system was used to construct baculoviruses carrying 1-copy and 4-copy Cap protein genes. We found that elevating the copy number could improve the expression level of the Cap protein, but not to the extent of a 4-fold increase—only a 12% enhancement compared to the 1-copy construct. This limited improvement may be due to the restricted expression capacity of insect cells or the presence of negative feedback mechanisms. Meanwhile, the baculovirus expressing 4 copies of the Cap protein reached its peak expression on the fourth day post-infection, followed by a decline on the fifth day. This reduction might be attributed to cellular negative feedback mechanisms or the onset of cell lysis, releasing intracellular proteases that degrade the protein. Although the Cap protein in this experiment was designed for intracellular expression and its levels decreased by the fifth day, harvesting at this time point was deemed more suitable for industrial-scale downstream purification. This is because a higher proportion of cell death at this stage facilitates the subsequent purification process.
In addition to codon optimization, increasing the gene copy number can enhance protein expression [30]. In this study, the flashBAC system was used to construct baculoviruses carrying 1-copy and 4-copy genes of the Cap protein. We found that elevating the copy number could improve the expression level of the Cap protein, but not to the extent of a 4-fold increase—only a 12% enhancement compared to the 1-copy construct. This limited improvement might be attributed to the constrained expression capacity of insect cells or the presence of negative feedback mechanisms [31]. Furthermore, the baculovirus with 4 copies of the Cap protein reached peak expression on the 4 dpi, followed by a decline on the fifth day. This reduction could be due to cellular negative feedback mechanisms or the onset of cell lysis, releasing intracellular proteases that degrade the protein. Although the Cap protein in this study was designed for intracellular expression and its levels decreased by the fifth day, harvesting at this time point was deemed more suitable for industrial-scale downstream purification. This is because a higher proportion of cell death at this stage facilitates the subsequent purification process.
Baculoviruses also exhibit passage instability. With successive viral passages, an increasing number of defective viruses incapable of expressing the target protein are generated. These non-functional viral particles proliferate at a higher rate compared to viruses carrying the target gene. Consequently, the expression levels of the target protein progressively decline with extended passaging [32]. Both industrial production guidelines and scientific literature therefore recommend limiting baculovirus passages to a maximum of five generations to maintain optimal protein expression. While increasing the copy number can enhance expression, it also raises the risk of homologous recombination in the viral genome. Thus, the probability of viral genome recombination increases with more passages [33]. To ensure stable protein expression, it is advisable to restrict baculovirus passages to 4–5 generations. This limited number of passages effectively mitigates the risk of viral genome instability caused by increased copy numbers.
The optimization of the MOI in baculovirus infection is also an important factor in improving protein expression [34]. In this experiment, the MOI was not optimized, and the recommended MOI of 0.05 from the kit was used. Future studies will consider MOI optimization to further enhance the expression level of the Cap protein. In addition to enhancing protein yield by optimizing the codon usage of the target protein to match that of insect cells [29], promoter engineering is another frequently employed strategy to improve protein expression. Studies have shown that cis-linking the polyhedrin promoter to the p10 promoter enhances protein production, while co-expressing molecular chaperones and transcription-associated proteins such as baculovirus transactivation factors IE1 and IE0 can improve baculovirus protein expression levels [35]. This study did not explore the optimization of promoter activity or the expression of molecular chaperones. Whether the use of tandem promoter combinations or the co-expression of molecular chaperones could further enhance protein expression warrants further investigation.
5. Conclusion
This study utilized different baculovirus construction systems, Bac-to-Bac and flashBAC, to generate recombinant baculoviruses expressing the Cap protein. Additionally, the flashBAC system was employed to construct baculoviruses with varying copy numbers of the Cap protein. The expression levels of the Cap protein were evaluated by infecting different insect cell lines, High Five and Sf9. The results demonstrated that the recombinant baculovirus constructed using the flashBAC system yielded the highest Cap protein expression when infecting High Five cells. Furthermore, our findings revealed that increasing the copy number of the cap gene in the baculovirus enhanced Cap protein expression levels. This study provides critical data for optimizing the production process of the Cap protein, which holds significant importance for reducing the production cost of porcine circovirus vaccines and improving vaccine manufacturing efficiency.
Abbreviations
Porcine circovirus type 2 (PCV2); porcine circovirus disease (PCVD); post-weaning multisystemic wasting syndrome (PMWS); porcine dermatitis and nephropathy syndrome (PDNS); virus-like particles; dpi: days post-infection (VLPs); post-infection (dpi); kilobases (kb); open reading frames (ORFs); capsid (Cap); polymerase chain reaction (PCR); Luria-Bertani (LB); multiplicity of infection (MOI); Phosphate-Buffered Saline (PBS); 50% tissue culture infectious dose (TCID50); column volumes (CV).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. Primers used in this article. Table S2. DNA sequence of PCV2 Cap protein optimized for insect cell codon preference.
Authors’ contributions
Designed the experiments: L.C., D.X., Y.Z. and S.Z.; writing—original draft: L.C.; Writing - review & editing: W.J., Y.Z. and S.Z.; performed the experiments: X.Y., F.Y., X.Y. and N.G.; analyzed the data: Y.Z. and M.Y. All authors read and approved the final manuscript.
Funding
This work was supported in part by the Zhejiang Hisun Animal Healthcare Products Co., Ltd. Fund Project, Jiangsu Province Agricultural Science and Technology Independent Innovation Fund Program (Grant No. CX(24)1010).
Institutional Review Board Statement
Animal experiments were approved by the Animal Ethics Committee of the Zhejiang Hisun Animal Healthcare Products Co., Ltd. The Animal Ethics Committee approval number was YS2024-05. During the entire experimental process, all protocols used were consistent with the rules for animal ethics.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets supporting the conclusions in this article are included within the article.
Acknowledgments
We thank all of the authors of the primary studies included in this article.
Declaration of Generative AI in Scientific Writing
The textual content of this article was written by the author, with partial content translated using DeepSeek AI-Assisted Translation to improve readability and language quality. All figures, images, and artwork were created without utilizing AI technology.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Lv, W.; Cao, L.; Yang, L.; Wang, N.; Li, Z.; Huang, S.; Wen, F.; Guo, J. The Prevalence and Genetic Diversity of Porcine Circoviruses (PCVs) during 2017–2023 in Guangdong Province, China. Animals 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Tischer, I.; Gelderblom, H.; Vettermann, W.; Koch, M.A. A very small porcine virus with circular single-stranded DNA. Nature 1982, 295, 64–66. [Google Scholar] [CrossRef]
- Opriessnig, T.; Karuppannan, A.K.; Castro, A.M.M.G.; Xiao, C.-T. Porcine circoviruses: current status, knowledge gaps and challenges. Virus Research 2020, 286. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.B.; Zhao, Y.; Cui, J.T.; Zheng, H.H.; Xu, T.; Hou, C.Y.; Wang, Z.Y.; Li, X.S.; Zheng, L.L.; Chen, H.Y. Molecular detection and phylogenetic analysis of Porcine circovirus 4 in Henan and Shanxi Provinces of China. Transboundary and Emerging Diseases 2020, 68, 276–282. [Google Scholar] [CrossRef]
- Meng, X.-J. Porcine Circovirus Type 2 (PCV2): Pathogenesis and Interaction with the Immune System. Annual Review of Animal Biosciences 2013, 1, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Hou, L.; Zhou, J.; Wang, D.; Cui, Y.; Feng, X.; Liu, J. Porcine Circovirus Type 2 Vaccines: Commercial Application and Research Advances. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Wang, F.; Guo, X.; Ge, X.; Wang, Z.; Chen, Y.; Cha, Z.; Yang, H. Genetic variation analysis of Chinese strains of porcine circovirus type 2. Virus Res 2009, 145, 151–156. [Google Scholar] [CrossRef]
- Sola, C.; Franzo, G.; Segalés, J. Porcine circovirus 2 (PCV-2) genotype update and proposal of a new genotyping methodology. Plos One 2018, 13. [Google Scholar] [CrossRef]
- Wang, Y.; Noll, L.; Lu, N.; Porter, E.; Stoy, C.; Zheng, W.; Liu, X.; Peddireddi, L.; Niederwerder, M.; Bai, J. Genetic diversity and prevalence of porcine circovirus type 3 (PCV3) and type 2 (PCV2) in the Midwest of the USA during 2016–2018. Transboundary and Emerging Diseases 2020, 67, 1284–1294. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, Z.; Tao, Q.; Xu, L.; Gu, S.; Huang, Y.; Liu, Z.; Zhang, Y.; Wen, J.; Lai, S.; et al. Construction of recombinant pseudorabies virus expressing PCV2 Cap, PCV3 Cap, and IL-4: investigation of their biological characteristics and immunogenicity. Frontiers in Immunology 2024, 15. [Google Scholar] [CrossRef]
- Hamel, A.L.; Lin, L.L.; Nayar, G.P. Nucleotide sequence of porcine circovirus associated with postweaning multisystemic wasting syndrome in pigs. Journal of virology 1998, 72, 5262–5267. [Google Scholar] [CrossRef]
- Karuppannan, A.; Opriessnig, T. Porcine Circovirus Type 2 (PCV2) Vaccines in the Context of Current Molecular Epidemiology. Viruses 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Sun, M.; Wang, N.; Zhang, S.; Deng, Z.; Xu, H.; Yang, H.; Gu, H.; Fang, W.; He, F. Efficacy comparison in cap VLPs of PCV2 and PCV3 as swine vaccine vehicle. International journal of biological macromolecules 2024, 278, 134955. [Google Scholar] [CrossRef]
- Weibel, H.; Sydler, T.; Brugnera, E.; Voets, H.; Grosse Liesner, B.; Sidler, X. Efficacy of simultaneous vaccination with Enterisol® Ileitis and Ingelvac® CircoFLEXTM in a Swiss breeding farm. Schweizer Archiv fur Tierheilkunde 2012, 154, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yu, P.; Bao, Y.; Ren, Y.; Zhao, S.; Zhang, X. Production of virus-like particles of porcine circovirus 2 in baculovirus expression system and its application for antibody detection. BMC Veterinary Research 2023, 19. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Zhan, Y.; Gong, Q.; Yu, W.; Deng, Z.; Wang, A.; Yang, Y.; Wang, N. Generation of E. coli-derived virus-like particles of porcine circovirus type 2 and their use in an indirect IgG enzyme-linked immunosorbent assay. Archives of virology 2016, 161, 1485–1491. [Google Scholar] [CrossRef]
- Tripathi, N.K.; Shrivastava, A. Recent Developments in Bioprocessing of Recombinant Proteins: Expression Hosts and Process Development. Frontiers in bioengineering and biotechnology 2019, 7, 420. [Google Scholar] [CrossRef]
- Hayat, S.M.G.; Farahani, N.; Golichenari, B.; Sahebkar, A. Recombinant Protein Expression in Escherichia coli (E.coli): What We Need to Know. Current pharmaceutical design 2018, 24, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Hitchman, R.B.; Possee, R.D.; King, L.A. High-Throughput Baculovirus Expression in Insect Cells. In Recombinant Gene Expression; Methods in Molecular Biology; 2012; pp. 609-627.
- Stolt-Bergner, P.; Benda, C.; Bergbrede, T.; Besir, H.; Celie, P.H.N.; Chang, C.; Drechsel, D.; Fischer, A.; Geerlof, A.; Giabbai, B.; et al. Baculovirus-driven protein expression in insect cells: A benchmarking study. Journal of Structural Biology 2018, 203, 71–80. [Google Scholar] [CrossRef]
- Lu, B.; Tang, Q.; Wang, Q.; Liu, X.; Peng, H.; Zhu, B.; Xie, L.; Li, Z.; Wang, H.; Zheng, Z.; et al. Recovery Infectious Enterovirus 71 by Bac-to-Bac Expression System in vitro and in vivo. Front Microbiol 2022, 13, 825111. [Google Scholar] [CrossRef]
- Vilca-Machaca, L.S.; Calvay-Sanchez, K.D.; Zarate-Sulca, Y.; Jimenez-Vasquez, V.; Ramirez, P.; Mendoza-Mujica, G. Baculovirus-Assisted Production of Bartonella bacilliformis Proteins: A Potential Strategy for Improving Serological Diagnosis of Carrion’s Disease. Pathogens 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Sauer, C.; Syvertsson, S.; Bohorquez, L.C.; Cruz, R.; Harwood, C.R.; van Rij, T.; Hamoen, L.W. Effect of Genome Position on Heterologous Gene Expression in Bacillus subtilis: An Unbiased Analysis. ACS synthetic biology 2016, 5, 942–947. [Google Scholar] [CrossRef]
- Gulyás, D.; Kocsis, B.; Szabó, D. Plasmid copy number and qnr gene expression in selection of fluoroquinolone-resistant Escherichia coli. Acta microbiologica et immunologica Hungarica 2019, 66, 169–178. [Google Scholar] [CrossRef]
- Li, P.-c.; Qiao, X.-w.; Zheng, Q.-s.; Hou, J.-b. Immunogenicity and immunoprotection of porcine circovirus type 2 (PCV2) Cap protein displayed by Lactococcus lactis. Vaccine 2016, 34, 696–702. [Google Scholar] [CrossRef]
- Bucarey, S.A.; Noriega, J.; Reyes, P.; Tapia, C.; Sáenz, L.; Zuñiga, A.; Tobar, J.A. The optimized capsid gene of porcine circovirus type 2 expressed in yeast forms virus-like particles and elicits antibody responses in mice fed with recombinant yeast extracts. Vaccine 2009, 27, 5781–5790. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, F.; Yuan, C.; Zhang, Y.; Ren, J.; Yue, H.; Ma, T.; Song, Q. Comparison of immune effects of porcine circovirus type 2d (PCV2d) capsid protein expressed by Escherichia coli and baculovirus-insect cells. Vaccine 2024, 42, 2848–2857. [Google Scholar] [CrossRef]
- ROOF, M.B.; HAYES, P.W.; EICHMEYER, M.; NITZEL, G.; SCHAEFFER, M. Multivalent PCV2 immunogenic compositions and methods of producing such compositions. EP Patant 3320919B1, 05 July 2007.
- Wu, X.; Xu, M.; Yang, J.-R.; Lu, J. Genome-wide impact of codon usage bias on translation optimization in Drosophila melanogaster. Nature Communications 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Vassileva, A.; Chugh, D.A.; Swaminathan, S.; Khanna, N. Effect of copy number on the expression levels of hepatitis B surface antigen in the methylotrophic yeast Pichia pastoris. Protein Expr Purif 2001, 21, 71–80. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Z. Engineering strategies for enhanced production of protein and bio-products in Pichia pastoris: A review. Biotechnol Adv 2018, 36, 182–195. [Google Scholar] [CrossRef]
- Ramoska, W.A.; Hink, W.F. Electron microscope examination of two plaque variants from a nuclear polyhedrosis virus of the alfalfa looper, Autographa californica. Journal of invertebrate pathology 1974, 23, 197–201. [Google Scholar] [CrossRef]
- Triplett, M.K.; Johnson, M.J.; Symington, L.S. Induction of homologous recombination by site-specific replication stress. DNA repair 2024, 142, 103753. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Guardalini, L.G.; Moura Dias, F.; Omae Camalhonte, S.; Leme, J.; Consoni Bernardino, T.; Soares Sposito, F.; Dias, E.; Mancini Astray, R.; Tonso, A.; Attie Calil Jorge, S.; et al. Multiplicity of infection and culture medium on the SARS-CoV-2 virus like-particles production by baculovirus/insect system. Biotechnology letters 2025, 47, 32. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gómez-Sebastián, S.; López-Vidal, J.; Escribano, J.M. Significant Productivity Improvement of the Baculovirus Expression Vector System by Engineering a Novel Expression Cassette. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of Bac-to-Bac and flashBAC construction of baculovirus.
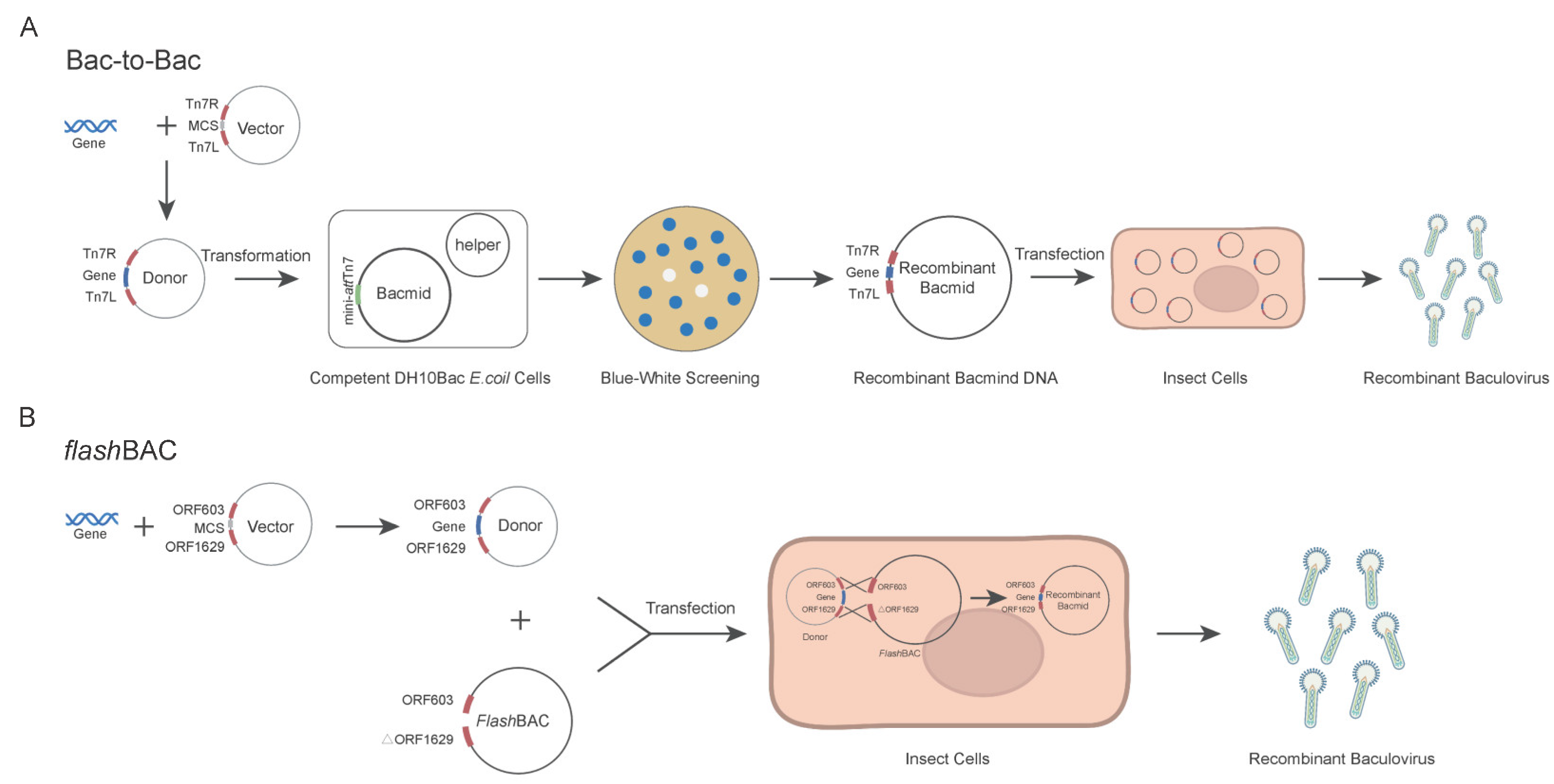
Figure 2.
Construction of pFast-Cap, pBAC-1-Cap and pBAC-4×Cap vectors. (A) Gene expression cassette schematic diagram. (B) pFast-Cap PCR. (C) Digestion of pFast-dual with BamHI and HindIII. (D) pBAC-Cap PCR. (E) Digestion of pBAC-1 with BamHI and HindIII. (F) Digestion of pUC19-4×Cap with BglII and HindIII. (G) Digestion of pBAC-1 with BglII and HindIII. (H) Transforming DH10Bac E. coli competent cells with pFast-Cap. (I) PCR-based identification of bacmid.
Figure 2.
Construction of pFast-Cap, pBAC-1-Cap and pBAC-4×Cap vectors. (A) Gene expression cassette schematic diagram. (B) pFast-Cap PCR. (C) Digestion of pFast-dual with BamHI and HindIII. (D) pBAC-Cap PCR. (E) Digestion of pBAC-1 with BamHI and HindIII. (F) Digestion of pUC19-4×Cap with BglII and HindIII. (G) Digestion of pBAC-1 with BglII and HindIII. (H) Transforming DH10Bac E. coli competent cells with pFast-Cap. (I) PCR-based identification of bacmid.
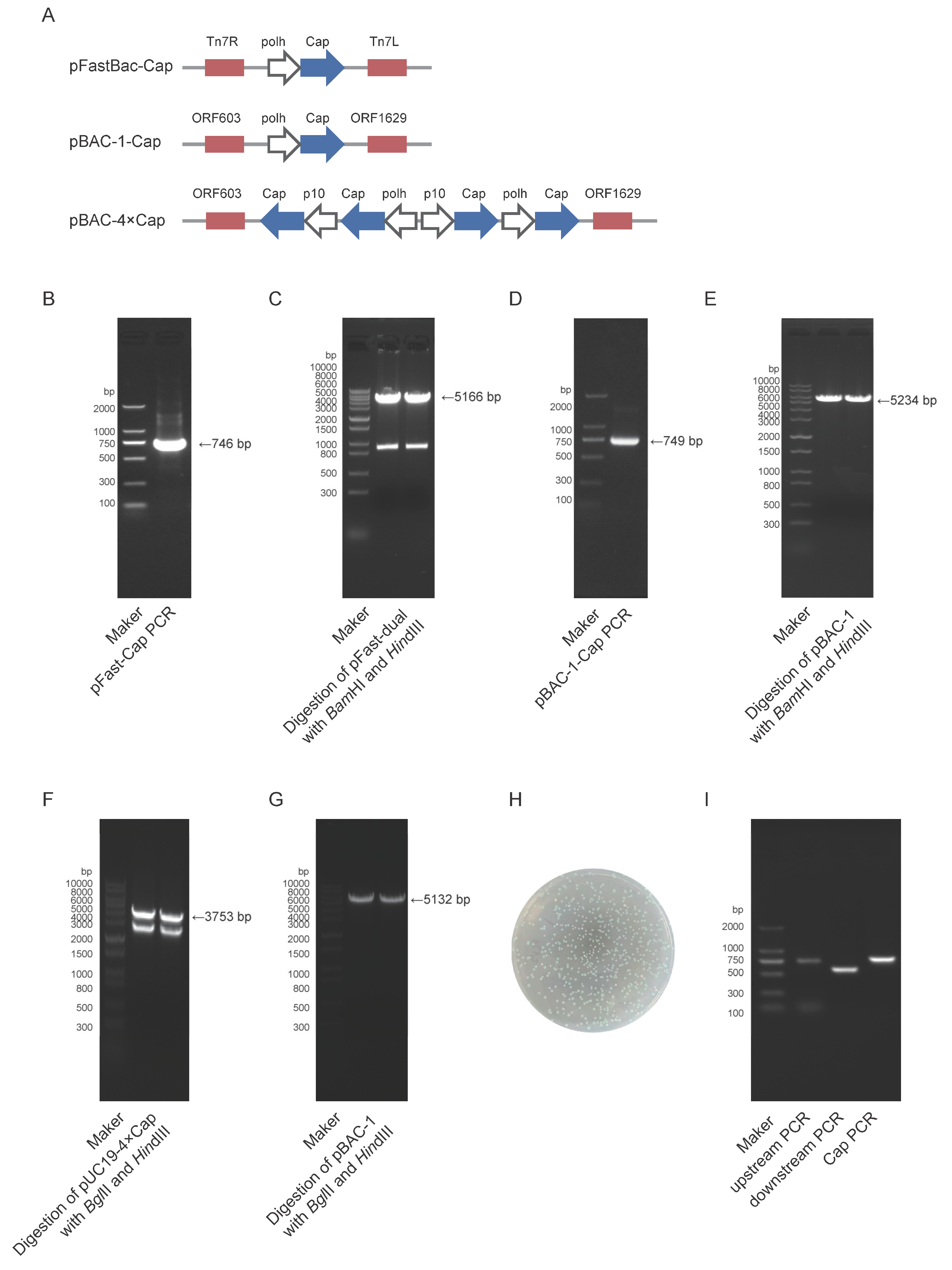
Figure 3.
Baculovirus identification by PCR and titer determination by immunofluorescent staining. (A) Identification of baculovirus using VP80 primers. (B) TCID50 of baculovirus. (C) Immunofluorescence staining of baculovirus.
Figure 3.
Baculovirus identification by PCR and titer determination by immunofluorescent staining. (A) Identification of baculovirus using VP80 primers. (B) TCID50 of baculovirus. (C) Immunofluorescence staining of baculovirus.
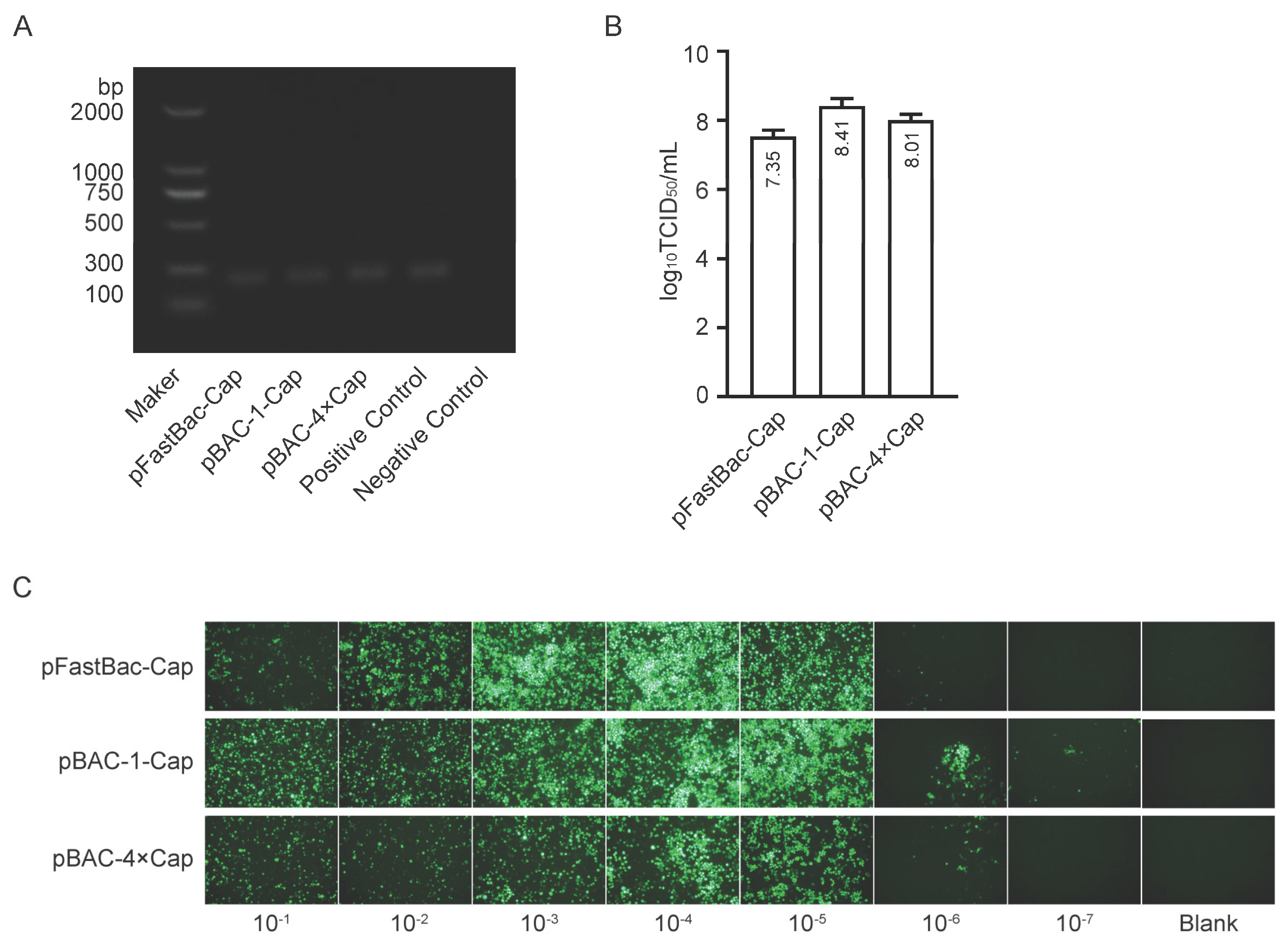
Figure 4.
Growth parameters of Sf9 and High Five cells after baculovirus infection. (A) Total cell number of Sf9 cells post-infection. (B) Viable cell number of Sf9 cells post-infection. (C) Viability of Sf9 cells post-infection. (D) Diameter of Sf9 cells post-infection. (E) Total cell number of High Five cells post-infection. (F) Viable cell number of High Five cells post-infection. (G) Viability of High Five cells post-infection. (H) Diameter of High Five cells post-infection.
Figure 4.
Growth parameters of Sf9 and High Five cells after baculovirus infection. (A) Total cell number of Sf9 cells post-infection. (B) Viable cell number of Sf9 cells post-infection. (C) Viability of Sf9 cells post-infection. (D) Diameter of Sf9 cells post-infection. (E) Total cell number of High Five cells post-infection. (F) Viable cell number of High Five cells post-infection. (G) Viability of High Five cells post-infection. (H) Diameter of High Five cells post-infection.
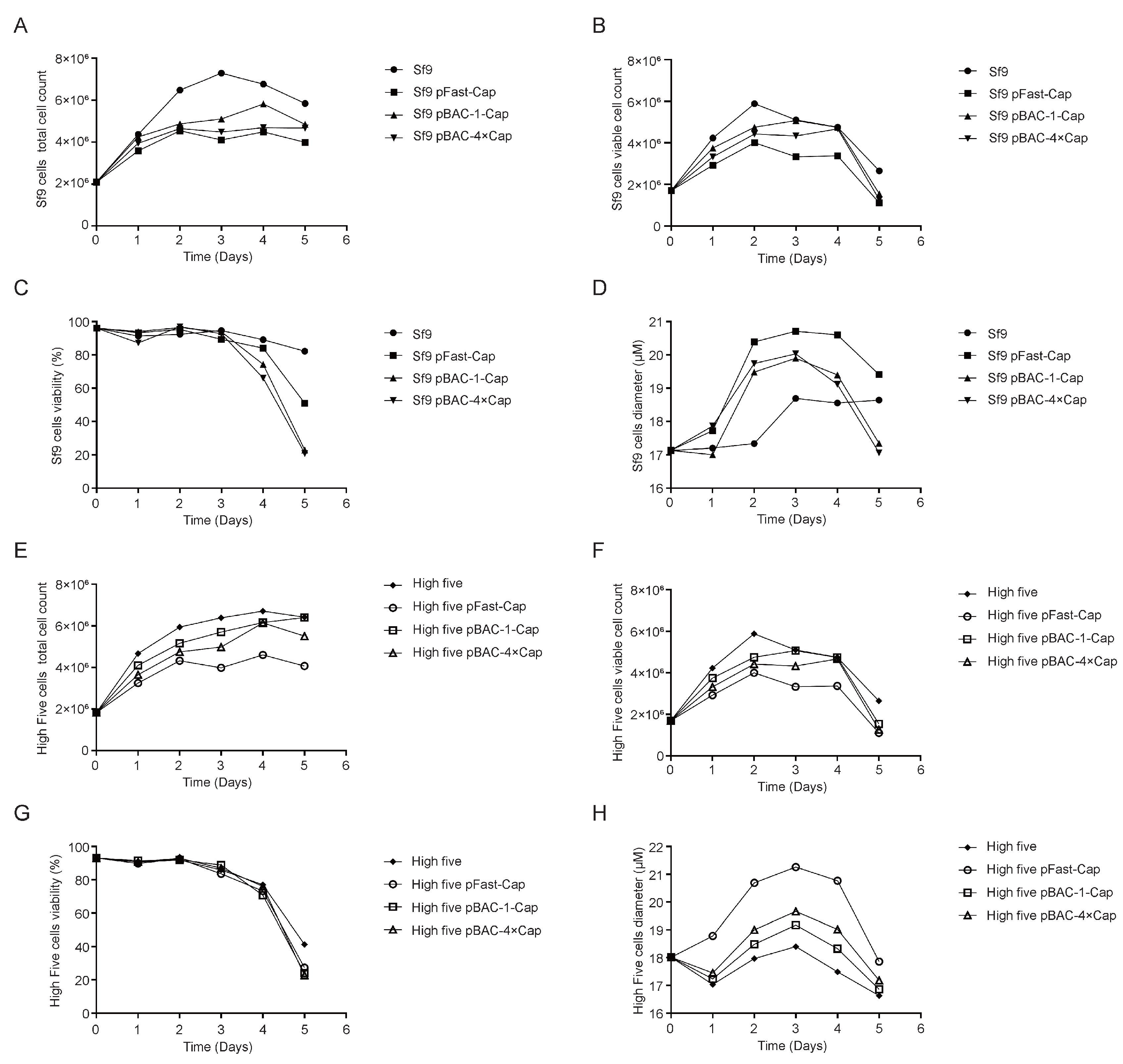
Figure 5.
Cap protein expression in baculovirus-infected Sf9 and High Five cell Lines. (A) Analysis of the expressed Cap protein by SDS-PAGE. (B) Determination of Cap protein concentration by ELISA.
Figure 5.
Cap protein expression in baculovirus-infected Sf9 and High Five cell Lines. (A) Analysis of the expressed Cap protein by SDS-PAGE. (B) Determination of Cap protein concentration by ELISA.
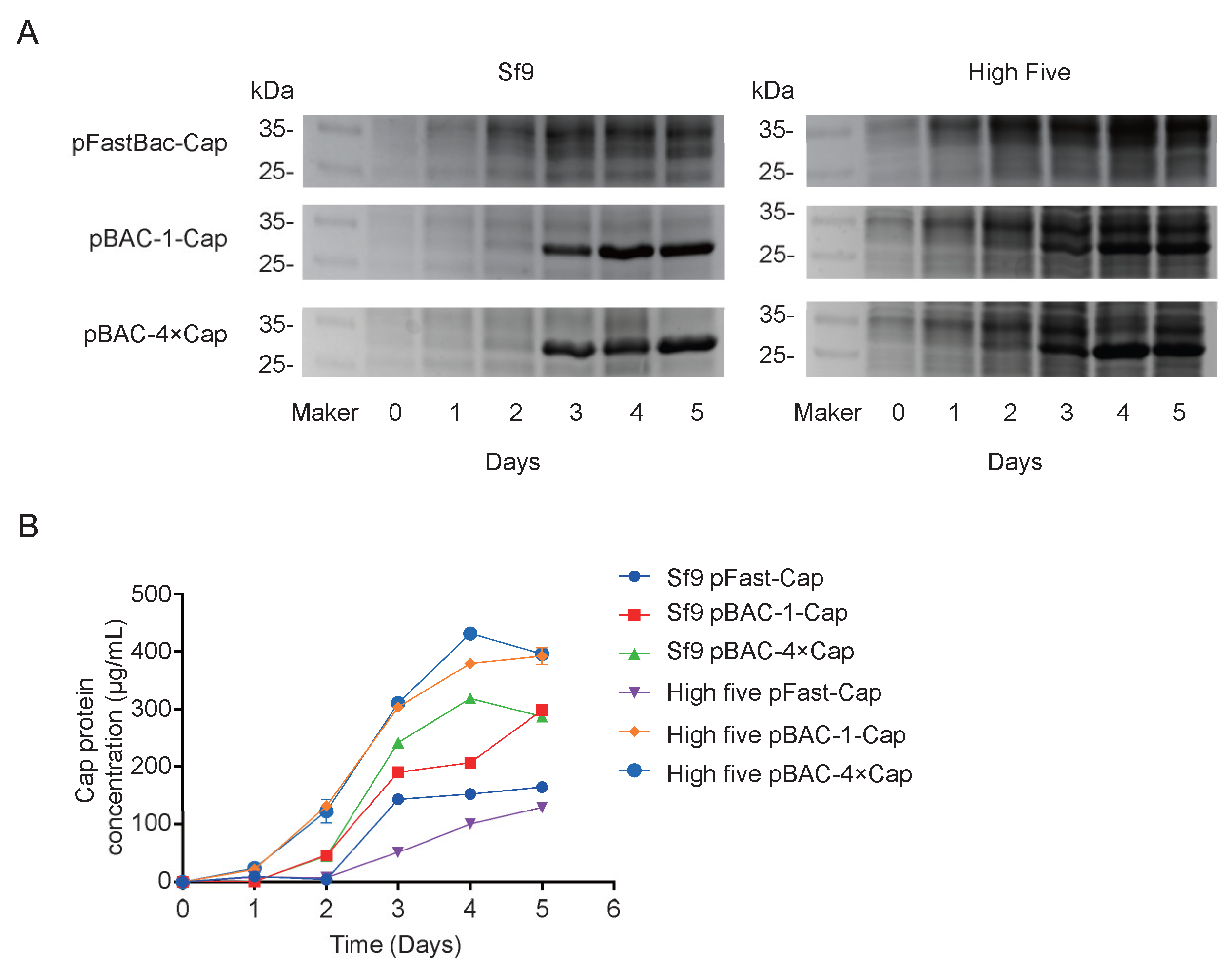
Figure 6.
Purification of Cap Protein by Cation Exchange Chromatography. (A) Cation exchange chromatography profile. (B) Analysis of the Cap protein by SDS-PAGE. (C) Analysis of the Cap protein by western blotting.
Figure 6.
Purification of Cap Protein by Cation Exchange Chromatography. (A) Cation exchange chromatography profile. (B) Analysis of the Cap protein by SDS-PAGE. (C) Analysis of the Cap protein by western blotting.
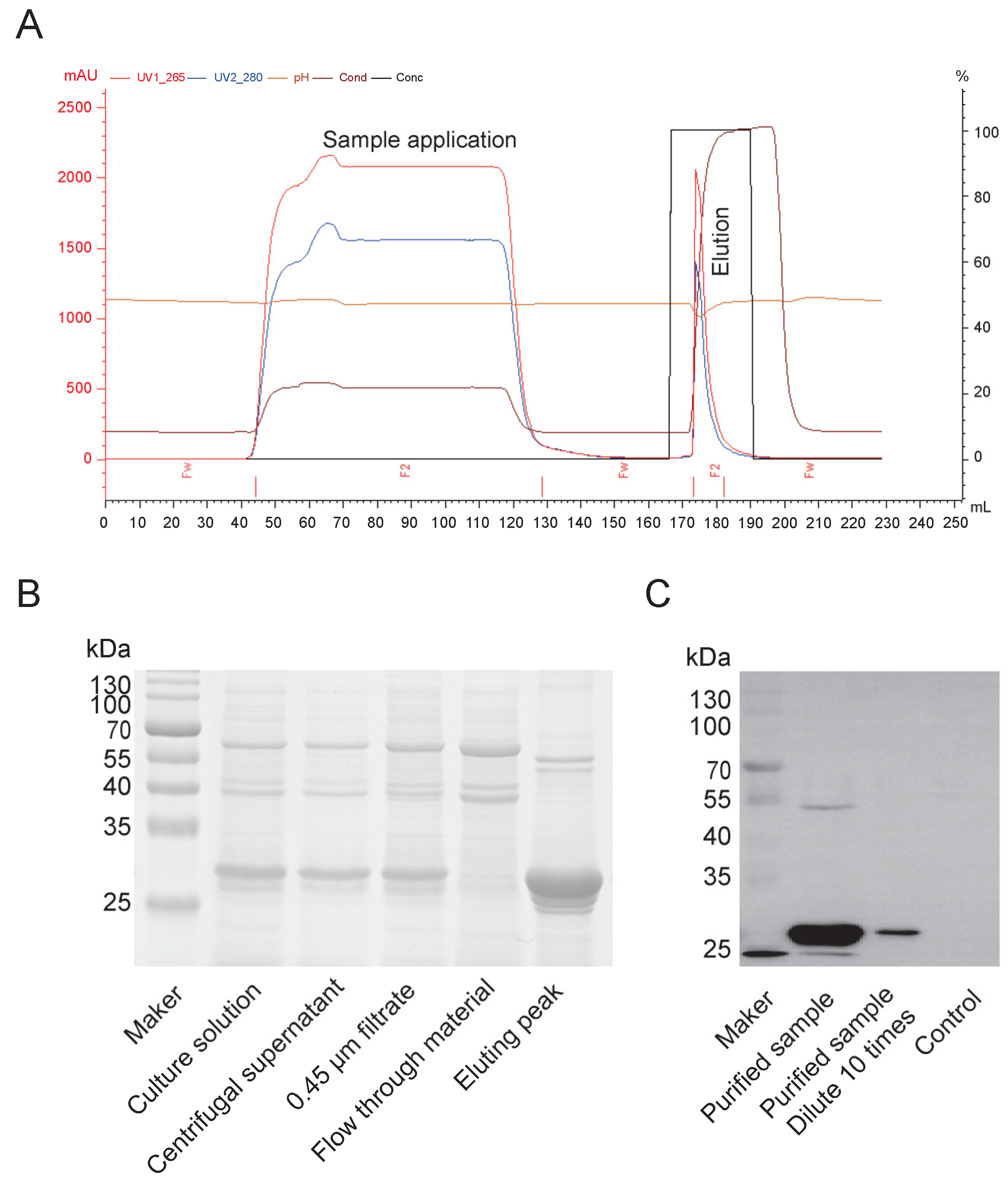
Figure 7.
Serum antibody content in mice and swine after immunization. (A) Timeline of animal immunization and serum collection. (B) Post-immunization IgG antibody in mice. (C) Post-immunization IgG antibody dynamics in swine.
Figure 7.
Serum antibody content in mice and swine after immunization. (A) Timeline of animal immunization and serum collection. (B) Post-immunization IgG antibody in mice. (C) Post-immunization IgG antibody dynamics in swine.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



