Submitted:
19 June 2025
Posted:
20 June 2025
You are already at the latest version
Abstract
Recent data from the CDC indicate that the incidence of Autism Spectrum Disorder (ASD), a neurodevelopmental disorder characterized by deficits in social communication and the presence of restricted interests and repetitive behaviors , has increased to 1 in 31 children. Individuals with ASD have a constellation of neurological, behavioral, sensory, feeding, gastrointestinal, and immunological issues. This manuscript discusses how these can be explained by disruption of the gut-blood and blood-brain barriers due to microbiome dysbiosis which then results in chronic endotoxemia leading to chronic cycle of gut-brain inflammation via mast cell and microglial activation. We present how various environmental, pathogenic and stress factors can disrupt the gut-brain homeostasis to create susceptibility and epigenetic effects contributing to the development of ASD. We also offer a safe, simple, and effective treatment approach to addressing some of the key pathogenetic underpinnings of ASD.
Keywords:
brain
; flavonoids
; folinic acid
; gut
; inflammation
; luteolin
; mast cells
; microbiome
; microglia
; toxins
1. Introduction
Autism Spectrum Disorder (ASD) is increasing at an alarming rate and now impacts 1 in 31 children in the United States [2], with a projected total cost of $461 billion by 2025 [3]. While genetic vulnerabilities are known to be a significant contributing factor to ASD, environmental factors, particularly environmental toxicants, may be another reason for the increased prevalence of ASD [4,5]. The microbiome and total gut physiology can also influence the onset of ASD and can be highly predictive of ASD [6].
Environmental toxins, including glyphosates, heavy metals, synthetic compounds and plastics, mold toxins, air pollution, COVID, and excessive use of antibiotics are just some examples of factors that can compromise gut permeability, induce gut-mediated inflammation, and disrupt the microbiomes [3,7,8,9,10,11,12,13]. Neonatal events such as C-section delivery and early use of antibiotics are also known to disrupt the microbiome [14] and may contribute to an increased risk of ASD [15,16,17]. What is not yet well understood is how transient, yet significant, exposures to environmental toxins and stressors contribute to ASD.
This paper discusses how such exposures disrupt the gut-brain axis leading to a chronic state of neuroinflammation primarily via activation of mast cells and microglial that can explain the physical, cognitive, psychological, and social findings in at least a subset of individuals with ASD. It further offers a simple, safe, and inexpensive set of recommendations to mitigate the inflammatory response and improve subjects with ASD.
2. Relevant Clinical Findings
Individuals with ASD have abnormal neuronal apoptosis [18], myelination [19] and neuroplasticity [20]. They also have dysfunction in multiple areas of cognition including: attention, executive functioning, working memory, praxis and motor planning [21]. Moreover, individuals with ASD experience significantly higher rates of anxiety [22], obsessive compulsive disorder (OCD) [23], social anxiety [24], abnormal perception of fear [25], anger and aggression [26]. This chronic state of psychological stress can trigger inflammation within the nervous system [27] and gastrointestinal tract [28]. These individuals also struggle with abnormalities in the processing of complex sensory information [29], with sensory processing disorders affecting up to 80-95% of individuals with ASD. The severity of these sensory processing disorders is a significant predictor of ASD severity and everyday functioning [30,31,32].
Children with ASD are also at a 500% higher risk of developing feeding problems [33], such as food selectivity, food refusal, and poor oral intake, as compared to neurodevelopmentally normal children [34,35]. Children with ASD have texture aversion and strong preferences for foods like carbohydrates and processed foods [36,37], and a higher risk of healthy food avoidance: vegetables (56% refusal), eggs (43%), fruits (42%), chicken (35%), and meat (24%) [35].
Individuals with ASD have a significantly higher prevalence of intestinal inflammation and inflammatory bowel disease [38] and are four times more likely to have gastrointestinal symptoms vs controls [36,39], including constipation (odds ratio 3.86), diarrhea (OR 3.63), abdominal pain (OR 2.45) [39], and these symptoms can begin as early as 6-18 months of age [40]. Individuals with ASD also have abnormal intestinal permeability [41], abnormal microbiomes [42] with a possible [43] higher prevalence of candida versus controls [44]. These individuals also have complex patterns of systemic immune dysregulation [45,46], with brain inflammation including microglia activation [47] and elevations of various cytokines in the cerebrospinal fluid [46,48], including the neuropilin disruptor matrix metalloproteinase-9 (MMP-9) [49]. The immune dysregulation also presents as higher rates of food allergies [50] and atopic diseases including asthma, allergies, and eczema, which are also strongly correlated with the risk of ASD50.
3. How Environmental Exposures Create Chronic Gastrointestinal Inflammation and Dysfunction
ASD is likely due to genetic vulnerabilities activated by to environmental toxins and stressors [51], with a significant contribution from the environmental factors themselves [52,53]. Some of these vulnerabilities occur at the gut level [54], including how host genetics interact with the gut microbes to shape the immune and metabolic state of ASD [55]. One such example is the role of the human leukocyte antigen (HLA) haplotypes, of which HLA haplotypes A2, DR4, and DR11 have been found to create major susceptibility for ASD45. These haplotypes can determine the specificity of T lymphocyte and natural killer (NK) cell responses, which then influence the makeup of the commensal bacteria [56].
As such, each individual has a genetically determined threshold of gut resilience and tolerance to various stressors or toxins. Thus, if an environmental factor or combination of factors supersede this threshold, the gastrointestinal tract and microbiome can become compromised setting off a complex cascade of gut-immune-brain dysregulation. At the center of this dysregulated axis are the gastrointestinal mast cells (Figure 1).
4. The Role of Mast Cells in Chronic Gastrointestinal Inflammation
Mast cells are the sentinels of our immune system [57] and can be activated by many of the same triggers that disrupt the gut, including: heavy metals, herbicides (including glyphosates), polychlorinated biphenyl (PCB), lipopolysaccharides (LPS), mycotoxins and molds, as well as pathogens [58] including Borrelia [59,60], and Helicobacter pylori [61] (Figure 2). Once activated, they orchestrate complex arrays of immune activation including mediating the allergy response [62], a finding common in individuals with ASD.
The largest population of mast cells is in the gastrointestinal tract, where they play a fundamental role in maintaining the intestinal barrier by regulating epithelial function and integrity, managing the defensive and immuno-regulatory function, modulating both innate and adaptive mucosal immunity, and maintaining neuro-immune interactions. These functions all play a fundamental role in the health or disease of the gut [63,64].
The gastrointestinal tract harbors trillions of bacteria, fungi (the mycobiome), viruses, LPS, and dietary antigens. The microbiota are well known to play an important role in neuropsychiatric disorders [65]. Separating this complex and potentially deadly “internal” gut content from our circulation and organ systems, including the brain, is a tightly regulated interface consisting of a mucus layer, immunoglobulin A, defensins, a single epithelial layer held together by tight junction proteins, and a carefully orchestrated immune surveillance in the lamina propria of the gut [66]. As long as this gut barrier is healthy and intact, and the bacterial and fungal communities are healthy, the likelihood of systemic and chronic inflammation is low. If there is a disruption in the microbial communities or gut barriers, the likelihood of a chronic systemic and nervous system inflammatory responses rise dramatically [67,68,69,70].
Zonulin and occludin are two important tight junction proteins that ensure the integrity of the intestinal barrier. Aberrant mast cell activation can cause profound disruption to the gut through damage to these tight junction proteins via mediators such as histamine, MMP-9, TNF-α and tryptase, resulting in increased intestinal permeability and inflammation, a critical step that allows the translocation of the commensal bacteria [63,66,71]. Through the release of numerous mediators mast cells can further diminish the integrity of the gut barrier, sensitize dendritic cells to microbial signals, including LPS, and influence the behavior of the innate and adaptive immune response [72,73,74]. Small intestinal mucosal damage may decrease the activity of diamine oxidase (DAO) [75], a key enzyme that degrades histamine, and thus exacerbate the detrimental effects of histamine that is released by activated mast cells. Mast cells can directly and indirectly suppress the function of T regulatory cells, while simultaneously promoting the activation, recruitment, proliferation and cytokine secretion in multiple T-cell subsets [62].
These combined changes to the gastrointestinal tract barrier and microbiome are sufficient for the pathogenesis of food allergies [76]. Especially in young children, even minor changes to the barrier function early in life can lead to exposure to luminal antigens which can result in allergies in later stages of life [77], as found in individuals with ASD. These gut abnormalities can also explain the gastrointestinal symptoms found in individuals with ASD, including abdominal pain, constipation, and diarrhea [78,79,80].
Perhaps, the most significant consequence of this toxin-mediated gut dysfunction is how it influences the makeup of the microbiome. Abnormalities in T-cell activity directly impact microbial diversity [81]. Inflammation-mediated dysbiosis, an emerging concept where gut inflammation alters the intestine’s oxidative and metabolic environment, can also profoundly influence the survival and growth of the gut bacteria and lead to a deviation of the commensal population from a healthy, diverse symbiotic profile to microbial communities with reduced complexity and over-representation of particular taxa of microbes [66,82].
Furthermore, certain toxins, like mycotoxins can alter the production of intestinal specific immunoglobulins [83], which then significantly influence the composition and behavior of the bacterial and fungal makeup of the microbiome [84,85,86,87,88]. Mast cell activation and the concurrent gut and microbiome disruptions can also induce candida species to switch from a harmless commensal to a virulent pathobiont, which is then able to invade tissues and disseminate in the body [71,89,90]. In turn, the invasive form of candida, and the toxin its hyphae form secretes, candidalysin [91], can trigger significant mast cell activation [92,93], thus creating a vicious cycle of chronic gut and immune dysfunction. Further complicating this picture, candida can alter the makeup of the microbiome through multiple mechanisms [94] and prevent the regrowth of lactobacilli after antibiotic treatment, while promoting the colonization of enterococcus [95]. All of these factors in turn perpetuate candida’s virulent pathobiont state. These mycobiome abnormalities also influence the maturation and priming of the immune system [96] and create an additional factor to induce or exacerbate the allergic state [97] that, as noted, is more prevalent in ASD [98].
5. Gut Disruption and Bacterial Translocation
The innate immune system within the gut has pattern recognition receptors (PRRs) whose primary purpose is to detect pathogens, including the commensal bacteria, by recognizing molecules and structures on the pathogens called pathogen-associated molecular patterns (PAMPs) [101]. The disruption of the gut barrier allows pathogenic commensal bacteria to breach the gut, at which point their PAMPs are recognized by the PRRs, which then activate the intestinal epithelial cells (IECs) to secrete cytokines and chemokines. This inflammatory reaction triggers a larger immune response to clean up the foreign and damaging antigens and threats, thus resulting in further inflammation and cytokine release [66]. These inflammatory cytokines further disrupt the epithelial barrier by downregulating tight junctions (claudin-1, occludin, zonula occludens protein-1) and adherens junctions (E-cadherin) in IECs [66]. One consequence of this pathogenic breach of the gut barrier and the ensuing inflammatory response becomes the key step in the induction and persistence of gut inflammation inflammatory bowel diseases [102], which is more prevalent in children with ASD versus healthy controls [103].
Another serious consequence of the compromised gut barrier is the entry of dangerous bacterial components into the systemic circulation. The gram-negative bacterial surface glycolipid lipopolysaccharide ( LPS), is among the most potent pro-inflammatory neurotoxins [104,105]. Gut dysbiosis and a disrupted gut barrier allow for the translocation of LPS through the gut lining into the systemic circulation to allow the development of a low-grade, chronic generalized toxin-associated effects [106], as is found in individuals with ASD [107]. The toxicity of the LPS increases with the degree of dysbiosis [105]. Numerous animal models suggest a strong link between generalized toxin effects and the features of ASD [108,109,110,111]. One study in rats reported autistic features after a single prenatal exposure to LPS [112]. Propionic acid is also known to cause similar effects [113,114].
Once absorbed, LPS can trigger systemic inflammation, with a reduction in T regulatory (Tregs) lymphocytes, increase in Th17 and Th1 lymphocytes, along with increased TNF-α, NFkB, IL-6, IL-8, IL- 10, and IL-12 [105,115]. Children with ASD also have activation of inflammasome complexes, including the NLRP3 inflammasome [116], which are complex systems that play a critical role in the regulation and activation of the body’s inflammatory response. LPS can rapidly prime and activate this inflammasome [117], which could be further primed by candidalysin [117,118]. Furthermore, within the intestinal lining, LPS binds to PRRs (TLR4) on intestinal cells [106] and mast cells [119,120], thus triggering additional inflammation within the gut and further disrupting the tight junction proteins and gut barrier.
The most concerning effect of LPS occurs within the nervous system. The microglia are the innate immune cells that modulate the inflammatory response within the central nervous system [46]. Peripheral gut inflammation and damage can activate the microglia [121,122,123]. In healthy adult volunteers, 1 ng/kg of LPS (similar levels found in some healthy adults) can trigger robust microglial activation in most areas of the brain as measured by PET within three hours of injection [115]. In rodents, a single intraperitoneal injection of 5 mg LPS/kg causes microglial activation that persists for at least 12 months [115]. Several other animal studies have also shown systemic LPS can activate the microglia [124,125,126,127]. If animals are given multiple doses of 1 mg LPS/kg (over several days), a model for chronic generalized toxin effects, they experience neuroinflammation, BBB permeability, and rapid neurodegeneration [115], findings that are common in individuals with ASD [128,129].
Even low levels of LPS can induce sickness behavior through elevation of inflammatory cytokines [115,130,131,132]. Sickness behavior, an adaptive change in behavior as a result of inflammation, has been described in individuals with ASD and can present as anxiety, appetite loss, depression, headache, impaired alertness and focus, lethargy, muscle pain, and social withdrawal [115,133].
The impact of LPS on the CNS is likely through multiple mechanisms, including the vagus nerve (Figure 2) [123]. LPS cannot pass through or directly disrupt the blood-brain barrier (BBB) [115]. On the other hand, central histamine, various toxins, a high fat, high sugar diet are just a few of the factors that can compromise the BBB [134,135,136,137,138], thus allowing LPS to directly enter with CNS. Stress via the release of corticotropin-releasing hormone CRH) can also disrupt the BBB through activation of mast cells [139,140,141].
Microglia shape neuronal plasticity and connectivity, and synaptic function and wiring through neuronal pruning [142,143,144]. They regulate myelin growth and integrity [145] and when activated may cause severe demyelination [146]. They are also involved in abnormal neuronal apoptosis [147]. By disrupting neuronal circuitry, microglial activation impairs the processing and integration of various sensory and emotional responses as part of the presentation of ASD [46,148,149]. Individuals with ASD have been found to have microglial activation [150], also confirmed through postmortem findings [151] and on functional positron emission tomography [152].
Microglial activation also alter the neuronal pathways of the amygdala, which has been shown to disrupt the fear threshold within human beings and may present as ASD [153]. Disruptions in the amygdala can also play a significant role in the pathogenesis of aggressive behavior [154], addictive behaviors [155,156], anxiety disorders [157], impulse control disorder [158], attention deficit disorder [159], depression, and a host of other neuropsychiatric findings [160]. These behavioral and psychiatric findings are all noted in individuals with ASD [161,162,163,164,165].
Furthermore, activated microglia may induce post-synaptic calcium elevation causing increased neuronal reactivity and disrupt glutamate signaling [144]. Microglia can also alter the levels of quinolinic acid [166], a potent neurotoxin implicated in ASD [167]. Microglial activation can also disrupt the behavior of the astrocytes [168], which in turn can further disrupt glutamate homeostasis, upset GABA regulation, and neuronal pruning [144].
Beyond the effects of the microglia, human and animal studies have shown that endotoxin-induced inflammation can also increase the neural responses in the anterior cingulate cortex and prefrontal regions further impacting the processing of social and emotional information [169]. Beyond the microglia are the mast cells, which serve as an “immune gate to the brain” and communicate with microglia in a two-way activation [170,171,172,173].
6. Mast Cells and ASD
Peripheral gut inflammation activates mast cells within the nervous system [174]. This is further impacted by psychological stress [175,176]. Mast cell activity is intimately tied to microglial activity, and the activation of one cell line can lead to the activation of other immune cells through multiple pathways (Figure 3) [168].
The role of mast cell and microglial activity has been well described by Theoharides, et al. in individuals with ASD [46,177,178,179]. Mast cell activity can disrupt the blood-brain barrier, and release multiple chemokines, cytokines, tissue disruptors and neurotoxic molecules, like IL-6 that create localized inflammation in the area of the basic ganglia that disrupts neuronal connectivity and contributes to ASD-related behaviors [180]. Mast cells also trigger a neuroinflammatory response through multiple mechanisms [46,47,181]. They can induce mitochondrial translocation to the cell surface with the secretion of extracellular mitochondrial adenosine triphosphate (ATP) and DNA that is then detected by the immune system as ‘innate pathogens’ triggering a significant inflammatory response, potentially contributing to ASD [182]. This mitochondrial DNA may also induce a neuro-inflammatory response, which has been found to alter behavior in mouse models [183]. Theoharides has suggested that this profound sequence of neuro-inflammatory response is sufficient to cause ASD in some [180].
Gut-mediated mast cell activation (and gut histamine) can increase histamine levels within the CNS, likely through multiple pathways, although the exact mechanisms are not yet clear [184]. It is clear that activated microglia and mast cells within the brain produce 50% of all brain histamine [185], and in animal models, intraperitoneal LPS injection can activate brain mast cells and cause a rapid elevation of central histamine within 6 hours [186].
Histamine plays a critical role in modulating the nervous system [187]. It regulates alertness and is also a key wake-promoting neurotransmitter that influences the circadian rhythm and sleep-wake behavior [185]. Sleep disruption is a common feature of ASD [188]. Elevated levels of central histamine can also disrupt the vestibular system, which is critical for balance, motor planning, and sensory perception [187]. Histamine has also been shown in animal models to directly or indirectly influence various sensory pathways: sound processing [189], tactile sensation [190], and olfactory perception [187,191]. The disruption of these sensory pathways may explain the host of sensory findings found in individuals with autism, including eating problems and food texture avoidance [192]. The intensity of sensory issues has been associated with more significant social difficulties, lower adaptive functioning, and lower or divergent visual exploration of social environments in children with ASD [193]. These sensory abnormalities are also strongly associated with social motivation and difficulties with initiating or maintaining social interactions, or feeling tense in social situations [193].
Furthermore, because of these sensory abnormalities and eating problems, individuals with ASD avoid fiber-rich vegetables and foods that are often rich in simple carbohydrates, fats, and sometimes highly processed foods [35,194]. These dietary preferences can exacerbate any microbiome disruption [195] and increase the abundance of candida [196]. The high amounts of carbohydrates and fats, and few dietary fibers can also dramatically enhance the absorption of LPS and induce an inflammatory response [197]. However, certain fruits and vegetable contain histamine (e.g. avocado, pineapple, spinach, tomatoes) that should best be avoided, especially in those with polymorphisms in the histamine degrading enzymes diamine oxidase (DAO) and histamine N-methyl transferase (HNMT). [198]
7. How to Address Gut-Brain Inflammation Effectively
While it is beyond the scope of this paper, numerous options are available to address many parts of this proposed cycle of physiological dysfunction and endogenous toxicity (Table 1).
Theoharides, et al. have demonstrated significant clinical improvements of symptoms in children with ASD from a dietary supplement which contains liposomal (formulated in olive pomace oil) luteolin (100 mg/softgel) quercetin (70 mg/softgel), and the quercetin glycoside rutin (30 mg/softgel)(NeuroProtek) [199,200,201]. The noted studies all suggest that these supplements are safe and well tolerated.
The terahydroxyflavone luteolin is a well-studied bioflavonoid with a host of anti-inflammatory properties [202,203,204]. Luteolin has been found to inhibit mast cell and T cell activation [205], and decrease levels of histamine and TNF [206,207]. In fact, luteolin was recently shown to be a more potent inhibitor that the “mast cell stabilizer” drug cromolyn [208]. Luteolin can also enter the brain and reduce microglial activation [209], particularly as a result of LPS [210], as well as have antibacterial properties [211]. Pentaxydroxyflavonol quercetin can inhibit mast cells and is more effective than cromolyn at blocking mast cell cytokine release [212]. It can also balance the Th1/Th2 immune response [213], reduces gut permeability while improving microbial diversity [214], and protects against LPS-induced gut damage through multiple mechanisms [215]. Rutin is another important compound because of its ability to liberate quercetin in the gut. The delivery base is just as important, since olive pomace oil that not only increases absorption from the gut, but it offers the well-known cytoprotective properties of olive oil [216]. Unfortunately, many cheaper preparations of luteolin and quercetin in powder form have flooded the market but are either of low purity or the daily dose requires multiple capsules [217]. The common notion that if you take higher amounts of luteolin or quercetin in powder form will eventually allow some of the flavonoids to be absorbed is not only wrong but dangerous as the unabsorbed flavonoids accumulate in the gut and disrupt the microflora [218].
The structural luteolin analogue, tetramethoxyluteolin, is even more potent than luteolin in inhibiting both mast cells and microglia [219,220,221,222,223], and has been incorporated in the novel anti-allergic skin lotion (GentleDerm) [224], which is particularly useful in those individuals with both eczema and ASD.
Two additional compounds that may play an useful role in the treatment of ASD are palmitoylethanolamide (PEA) and diamine oxidase (DAO) enzymes. PEA is a naturally occurring fatty acid amine found in soybean lecithin, egg yolk, and peanut meal. PEA has noticeable anti-inflammatory properties and can regulate mast cell activation [225,226,227] by reducing release of TNF-a and histamine [228]. Furthermore, PEA displays neuroprotective properties and can inhibit microglial activation [229], particularly as a result of LPS exposure [230]. In isolated case reports, PEA has been shown to be beneficial in ASD [230].
DAO enzymes are naturally occurring enzymes within the gastrointestinal tract that are responsible for the degradation of histamine within the gut. Since intestinal mucosal damage may decrease the DAO activity [75], additional supplementation may play an important role in normalizing histamine levels within the gut and reducing the inflammatory and possibly neurological findings. Histamine intolerance has also been associated with anxiety disorders [231]. Exogenous supplementation with DAO enzymes can significantly reduce histamine levels within the gastrointestinal tract and the signs and symptoms of histamine intolerance [232], including extra-intestinal symptoms such as headaches [233]. To our knowledge, DAO enzymes are safe and well tolerated. However, DAO preparations vary considerably in their stated activity and most the enzyme will be degraded by the stomach acidity unless they are in acid resistant formulations.
Addition of berberine, lactoferrin and Bifidobacteria infantis/longum could provide additional benefits as they all have both antibacterial and anti-inflammatory properties [234,235,236,237,238,239]. The combination of these naturally occurring compounds may prove useful in the treatment of ASD.
Addition of folinic acid (Calcium folinate, Leucovorin) has been shown to significantly improve brain health, cognition and language by bypassing surface folate receptors and the enzyme MTHFR, especially in those with anti-folate receptor antibodies and MTHFR polymorphisms (Table 1) [240].
There are obvious limitations to the suggested treatment approached discussed above. The clinical efficacy of these compounds may be limited in the face of significant gastrointestinal disease or serious environmental exposures, such as living in a home with severe mold contamination.
8. Beyond the Clinical Benefits
Unfortunately, at the time of this publication, there are few clinically available and reliable diagnostic tools that can help clinicians accurately assess these assessing gut-mediated pathophysiological events, including:
- (1)
- Assessing candida and the mycobiome – There are many commercially available stool kits to assess the bacterial component of the microbiome. Unfortunately, these kits lack the sensitivity to accurately detect disturbances of candida or other fungal components of the microbiome (mycobiome).
- (2)
- Assessing microglial activation – No commercially available diagnostic modalities are available to accurately assess microglial activation.
- (3)
- Assessing mast cell activity and histamine – Serum histamine has a half-life of less than two minutes and thus cannot be used to accurately detect histamine imbalances. Serum tryptase can be used to assess significant mast cell burden (e.g. systemic mastocytosis), which may limit its ability to detect more subtle forms of mast cell activation, including within the central nervous system. Urinary N-methylhistamine, Prostaglandin F2 alpha and leukotrience E4 must be collected cold in 24-hour urine and most clinical labs do not perform them.
- (4)
- Assessing endotoxemia – At this time there is no commercially available diagnostic tool available to directly assess endotoxemia.
- (5)
- Assessing total toxin load – Currently, only specialty tests are available to assess select categories of toxins. These tests are not FDA-approved and their results are at times called into question.
These points demonstrate the profound limitations in our current diagnostic tools to detect these critical physiological processes in individuals with ASD. They also highlight how important future well-designed research studies are in helping us further our understanding of ASD, and to allow this science to be effectively utilized in the clinical setting.
9. Conclusion
There is no singular trigger, event, genetic or physiological process that is solely responsible for the onset of ASD in the majority of cases. It is the total load of pathogenic and environmental toxins, which varies from individual to individual, compounded by other infectious [241], microbiome/gut, physiological and psychological stressors that may be responsible for the onset of ASD in some individuals [242]. We believe that in these individuals, there is a moment during gestation/delivery, in infancy or early childhood where the gut barriers, microbiome and gut-mediated immune responses surpass a threshold of homeostasis, and enter into a perpetual cycle of neuroinflammation.
While there is sufficient data to identify the individual factors contributing to ASD risk, additional research is needed to bring this vast array of findings into one cohesive model that has the power to assess the unique exposome for each individual and appreciate the total physiological impact it can have, including on the mitochondria and other organ system dysfunctions that have been associated with ASD. Additional research is also needed to assess the role that bacterial endotoxins may play in the manifestation of ASD.
In the meantime, the proposed interventions can inhibit some of the pathogenetic pathways and allow the gut-brain axis to recover as has been the case in numerous individuals with ASD.
Author Contributions
Conceptualization and writing-original draft, P.K., writing contribution – review and editing, T.H. and R.F., visualization, T.H. and P.K..
Disclosures
TCT is the Scientific Director of Algonot LLC(Sarasota, FL) that formulates and markets unique dietary supplements.
References
- American Psychiatric Association 1994 Diagnostic and Statistical Manual of Mental Disorders: DSM-IV. 4th ed. American Psychiatric Association, Washington, D.C.
- Shaw, K.A.; Williams, S.; Patrick, M.E.; Valencia-Prado, M.; Durkin, M.S.; Howerton, E.M.; Ladd-Acosta, C.M.; Pas, E.T.; Bakian, A.V.; Bartholomew, P.; Nieves-Muñoz, N.; Sidwell, K.; Alford, A.; Bilder, D.A.; DiRienzo, M.; Fitzgerald, R.T.; Furnier, S.M.; Hudson, A.E.; Pokoski, O.M.; Shea, L.; Tinker, S.C.; Warren, Z.; Zahorodny, W.; Agosto-Rosa, H.; Anbar, J.; Chavez, K.Y.; Esler, A.; Forkner, A.; Grzybowski, A.; Agib, A.H.; Hallas, L.; Lopez, M.; Magaña, S.; Nguyen, R.H.N.; Parker, J.; Pierce, K.; Protho, T.; Torres, H.; Vanegas, S.B.; Vehorn, A.; Zhang, M.; Andrews, J.; Greer, F.; Hall-Lande, J.; McArthur, D.; Mitamura, M.; Montes, A.J.; Pettygrove, S.; Shenouda, J.; Skowyra, C.; Washington, A.; Maenner, M.J. Prevalence and Early Identification of Autism Spectrum Disorder Among Children Aged 4 and 8 Years - Autism and developmental disabilities monitoring network, 16 sites, United States, 2022. MMWR Surveill Summ. 2025, 74, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Chiu, K.; Warner, G.; Nowak, R.A.; Flaws, J.A.; Mei, W. The Impact of environmental chemicals on the gut microbiome. Toxicol Sci. 2020, 176, 253–284. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, D.A.; Genuis, S.J.; Frye, R.E. Environmental toxicants and autism spectrum disorders: a systematic review. Transl Psychiatry. 2014, 4, e360. [Google Scholar] [CrossRef] [PubMed]
- Chaste, P.; Leboyer, M. Autism risk factors: genes, environment, and gene-environment interactions. Dialogues Clin Neurosci. 2012, 14, 281–292. [Google Scholar] [CrossRef]
- Su, Q.; Wong, O.W.H.; Lu, W.; Wan, Y.; Zhang, L.; Xu, W.; Li, M.K.T.; Liu, C.; Cheung, C.P.; Ching, J.Y.L.; Cheong, P.K.; Leung, T.F.; Chan, S.; Leung, P.; Chan, F.K.L.; Ng, S.C. Multikingdom and functional gut microbiota markers for autism spectrum disorder. Nat Microbiol. 2024, 9, 2344–2355, Erratum in: Nat Microbiol. 2025, 10, 600. doi: 10.1038/s41564-024-01900-w. [Google Scholar] [CrossRef] [PubMed]
- Tu, P.; Chi, L.; Bodnar, W.; Zhang, Z.; Gao, B.; Bian, X.; Stewart, J.; Fry, R.; Lu, K. Gut microbiome toxicity: connecting the environment and gut microbiome-associated diseases. Toxics. 2020, 8, 19. [Google Scholar] [CrossRef]
- Sondergaard, T.E.; Fredborg, M.; Oppenhagen Christensen, A.M.; Damsgaard, S.K.; Kramer, N.F.; Giese, H.; Sørensen, J.L. Fast screening of antibacterial compounds from fusaria. Toxins (Basel). 2016, 8, 355. [Google Scholar] [CrossRef]
- Liew, W.P.; Mohd-Redzwan, S. Mycotoxin: Its impact on gut health and microbiota. Front Cell Infect Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef]
- Guerre, P. Mycotoxin and gut microbiota interactions. Toxins (Basel). 2020, 12, 769. [Google Scholar] [CrossRef]
- Bernard-Raichon L, Venzon M, Klein J, Axelrad JE, Zhang C, Sullivan AP, Hussey GA, Casanovas-Massana A, Noval MG, Valero-Jimenez AM, Gago J, Putzel G, Pironti A, Wilder, E.; Yale IMPACTResearch, T.e.a.m.; Thorpe LE, Littman DR, Dittmann M, Stapleford KA, Shopsin B, Torres VJ, Ko AI, Iwasaki A, Cadwell K, Schluter, J. Gut microbiome dysbiosis in antibiotic-treated COVID-19 patients is associated with microbial translocation and bacteremia. Nat Commun. 2022, 13, 5926. [CrossRef]
- Fouladi, F.; Bailey, M.J.; Patterson, W.B.; Sioda, M.; Blakley, I.C.; Fodor, A.A.; Jones, R.B.; Chen, Z.; Kim, J.S.; Lurmann, F.; Martino, C.; Knight, R.; Gilliland, F.D.; Alderete, T.L. Air pollution exposure is associated with the gut microbiome as revealed by shotgun metagenomic sequencing. Environ Int. 2020, 138, 105604. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, E.A.; Comba, I.Y.; Cho, T.; Engen, P.A.; Yazıcı, C.; Soberanes, S.; Hamanaka, R.B.; Niğdelioğlu, R.; Meliton, A.Y.; Ghio, A.J.; Budinger, G.R.S.; Mutlu, G.M. Inhalational exposure to particulate matter air pollution alters the composition of the gut microbiome. Environ Pollut. 2018, 240, 817–830. [Google Scholar] [CrossRef]
- Korpela, K. Impact of delivery mode on infant gut microbiota. Ann Nutr Metab. 2021, 1–9. [Google Scholar] [CrossRef]
- Nitschke, A.S.; do Valle, H.A.; Vallance, B.A.; Bickford, C.; Ip, A.; Lanphear, N.; Lanphear, B.; Weikum, W.; Oberlander, T.F.; Hanley, G.E. Association between prenatal antibiotic exposure and autism spectrum disorder among term births: A population-based cohort study. Paediatr Perinat Epidemiol. 2023, 37, 516–526. [Google Scholar] [CrossRef]
- Al-Zalabani, A.H.; Al-Jabree, A.H.; Zeidan, Z.A. Is cesarean section delivery associated with autism spectrum disorder? Neurosciences (Riyadh). 2019, 24, 11–15. [Google Scholar] [CrossRef]
- Hamad, A.F.; Alessi-Severini, S.; Mahmud, S.M.; Brownell, M.; Kuo, I.F. Early childhood antibiotics use and autism spectrum disorders: a population-based cohort study. Int J Epidemiol. 2018, 47, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Alberts, I.; Li, X. The apoptotic perspective of autism. Int J Dev Neurosci. 2014, 36, 13–8. [Google Scholar] [CrossRef] [PubMed]
- Galvez-Contreras, A.Y.; Zarate-Lopez, D.; Torres-Chavez, A.L.; Gonzalez-Perez, O. Role of oligodendrocytes and myelin in the pathophysiology of autism spectrum disorder. Brain Sci. 2020, 10, 951. [Google Scholar] [CrossRef]
- Anashkina, A.A.; Erlykina, E.I. Molecular mechanisms of aberrant neuroplasticity in autism spectrum disorders (Review). Sovrem Tekhnologii Med. 2021, 13, 78–91. [Google Scholar] [CrossRef]
- Zwick, G.P. Neuropsychological assessment in autism spectrum disorder and related conditions. Dialogues Clin Neurosci. 19, 373–379. [CrossRef]
- White, S.W.; Oswald, D.; Ollendick, T.; Scahill, L. Anxiety in children and adolescents with autism spectrum disorders. Clin Psychol Rev. 2009, 29, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.M.; Petersen, L.; Schendel, D.E.; Mattheisen, M.; Mortensen, P.B.; Mors, O. Obsessive-compulsive disorder and autism spectrum disorders: longitudinal and offspring risk. PLoS One. 2015, 10, e0141703. [Google Scholar] [CrossRef] [PubMed]
- Montaser, J.; Umeano, L.; Pujari, H.P.; Nasiri, S.M.Z.; Parisapogu, A.; Shah, A.; Khan, S. Correlations between the development of social anxiety and individuals with autism spectrum disorder: a systematic review. Cureus. 2023, 15, e44841. [Google Scholar] [CrossRef] [PubMed]
- Mayes, S.D.; Calhoun, S.L.; Aggarwal, R.; Baker, C.; Mathapati, S.; Molitoris, S.; Mayes, R.D. Unusual fears in children with autism. Research in Autism Spectrum Disorders. 2013, 7, 151–158. [Google Scholar] [CrossRef]
- Patel, S.; Day, T.N.; Jones, N.; Mazefsky, C.A. Association between anger rumination and autism symptom severity, depression symptoms, aggression, and general dysregulation in adolescents with autism spectrum disorder. Autism. 2017, 21, 181–189. [Google Scholar] [CrossRef]
- Theoharides, T.C. Effect of stress on neuroimmune processes. Clin Ther. 2020, 42, 1007–1014. [Google Scholar] [CrossRef]
- Katiraei, P.; Bultron, G. Need for a comprehensive medical approach to the neuro-immuno-gastroenterology of irritable bowel syndrome. World J Gastroenterol. 2011, 17, 2791–2800. [Google Scholar] [CrossRef]
- Leekam, S.R.; Nieto, C.; Libby, S.J.; Wing, L.; Gould, J. Describing the sensory abnormalities of children and adults with autism. J Autism Dev Disord. 2007, 37, 894–910. [Google Scholar] [CrossRef]
- Sanz-Cervera, P.; Pastor-Cerezuela, G.; Fernández-Andrés, M.I.; Tárraga-Mínguez, R. Sensory processing in children with autism spectrum disorder: relationship with non-verbal IQ, autism severity and attention deficit/hyperactivity disorder symptomatology. Res Dev Disabil. 2015, 45-46, 188–201. [Google Scholar] [CrossRef]
- Tomchek, S.D.; Dunn, W. Sensory processing in children with and without autism: a comparative study using the short sensory profile. Am J Occup Ther. 2007, 61, 190–200. [Google Scholar] [CrossRef]
- Suarez, M.A. Sensory processing in children with autism spectrum disorders and impact on functioning. Pediatr Clin North Am. 2012, 59, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Kanner, L. Austic disturbances of affective contact. Nervous Child. 1943, 2, 217–250. [Google Scholar]
- Nadon, G.; Feldman, D.E.; Dunn, W.; Gisel, E. Association of sensory processing and eating problems in children with autism spectrum disorders. Autism Res Treat. 2011, 541926. [Google Scholar] [CrossRef]
- Cherif, L.; Boudabous, J.; Khemekhem, K.; Mkawer, S.; Ayadi, H.; Moalla, Y. Feeding problems in children with autism spectrum disorders. Journal of Family Medicine. 2018, 1, 30–39. [Google Scholar] [CrossRef]
- Madra, M.; Ringel, R.; Margolis, K.G. Gastrointestinal issues and autism spectrum disorder. Child Adolesc Psychiatr Clin N Am. 2020, 29, 501–513. [Google Scholar] [CrossRef]
- Baraskewich, J.; von Ranson, K.M.; McCrimmon, A.; McMorris, C.A. Feeding and eating problems in children and adolescents with autism: A scoping review. Autism. 2021, 25, 1505–1519. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Choi, M.J.; Ha, S.; Hwang, J.; Koyanagi, A.; Dragioti, E.; Radua, J.; Smith, L.; Jacob, L.; Salazar de Pablo, G.; Lee, S.W.; Yon, D.K.; Thompson, T.; Cortese, S.; Lollo, G.; Liang, C.S.; Chu, C.S.; Fusar-Poli, P.; Cheon, K.A.; Shin, J.I.; Solmi, M. Association between autism spectrum disorder and inflammatory bowel disease: A systematic review and meta-analysis. Autism Res. 2022, 15, 340–352. [Google Scholar] [CrossRef]
- Wasilewska, J.; Klukowski, M. Gastrointestinal symptoms and autism spectrum disorder: links and risks - a possible new overlap syndrome. Pediatric Health Med Ther. 2015, 6, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Bresnahan, M.; Hornig, M.; Schultz, A.F.; Gunnes, N.; Hirtz, D.; Lie, K.K.; Magnus, P.; Reichborn-Kjennerud, T.; Roth, C.; Schjølberg, S.; Stoltenberg, C.; Surén, P.; Susser, E.; Lipkin, W.I. Association of maternal report of infant and toddler gastrointestinal symptoms with autism: evidence from a prospective birth cohort. JAMA Psychiatry. 2015, 72, 466–474. [Google Scholar] [CrossRef]
- de Magistris, L.; Familiari, V.; Pascotto, A.; Sapone, A.; Frolli, A.; Iardino, P.; Carteni, M.; De Rosa, M.; Francavilla, R.; Riegler, G.; Militerni, R.; Bravaccio, C. Alterations of the intestinal barrier in patients with autism spectrum disorders and in their first-degree relatives. J Pediatr Gastroenterol Nutr. 2010, 51, 418–424. [Google Scholar] [CrossRef]
- Morton, J.T.; Jin, D.M.; Mills, R.H.; Shao, Y.; Rahman, G.; McDonald, D.; Zhu, Q.; Balaban, M.; Jiang, Y.; Cantrell, K.; Gonzalez, A.; Carmel, J.; Frankiensztajn, L.M.; Martin-Brevet, S.; Berding, K.; Needham, B.D.; Zurita, M.F.; David, M.; Averina, O.V.; Kovtun, A.S.; Noto, A.; Mussap, M.; Wang, M.; Frank, D.N.; Li, E.; Zhou, W.; Fanos, V.; Danilenko, V.N.; Wall, D.P.; Cárdenas, P.; Baldeón, M.E.; Jacquemont, S.; Koren, O.; Elliott, E.; Xavier, R.J.; Mazmanian, S.K.; Knight, R.; Gilbert, J.A.; Donovan, S.M.; Lawley, T.D.; Carpenter, B.; Bonneau, R.; Taroncher-Oldenburg, G. Multi-level analysis of the gut-brain axis shows autism spectrum disorder-associated molecular and microbial profiles. Nat Neurosci. 2023, 26, 1208–1217. [Google Scholar] [CrossRef] [PubMed]
- Alookaran, J.; Liu, Y.; Auchtung, T.A.; Tahanan, A.; Hessabi, M.; Asgarisabet, P.; Rahbar, M.H.; Fatheree, N.Y.; Pearson, D.A.; Mansour, R.; Van Arsdall, M.R.; Navarro, F.; Rhoads, J.M. Fungi: friend or foe? a mycobiome evaluation in children with autism and gastrointestinal symptoms. J Pediatr Gastroenterol Nutr. 2022, 74, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Han, Y.; Dy, A.B.C.; Hagerman, R.J. The gut microbiota and autism spectrum disorders. Front Cell Neurosci. 2017, 11, 120. [Google Scholar] [CrossRef] [PubMed]
- Robinson-Agramonte, M.L.A.; Noris García, E.; Fraga Guerra, J.; Vega Hurtado, Y.; Antonucci, N.; Semprún-Hernández, N.; Schultz, S.; Siniscalco, D. Immune dysregulation in autism spectrum disorder: what do we know about It? Int J Mol Sci. 2022, 23, 3033. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Tsilioni, I.; Patel, A.B.; Doyle, R. Atopic diseases and inflammation of the brain in the pathogenesis of autism spectrum disorders. Transl Psychiatry. 2016, 6, e844. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Asadi, S.; Patel, A.B. Focal brain inflammation and autism. J Neuroinflammation. 2013, 10, 46. [Google Scholar] [CrossRef]
- Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Pessah, I.; Van de Water, J. Elevated plasma cytokines in autism spectrum disorders provide evidence of immune dysfunction and are associated with impaired behavioral outcome. Brain Behav Immun. 2011, 25, 40–45. [Google Scholar] [CrossRef]
- Rexrode, L.E.; Hartley, J.; Showmaker, K.C.; Challagundla, L.; Vandewege, M.W.; Martin, B.E.; Blair, E.; Bollavarapu, R.; Antonyraj, R.B.; Hilton, K.; Gardiner, A.; Valeri, J.; Gisabella, B.; Garrett, M.R.; Theoharides, T.C.; Pantazopoulos, H. Molecular profiling of the hippocampus of children with autism spectrum disorder. Mol Psychiatry. 2024, 29, 1968–1979. [Google Scholar] [CrossRef]
- Xu, G.; Snetselaar, L.G.; Jing, J.; Liu, B.; Strathearn, L.; Bao, W. Association of food allergy and other allergic conditions with autism spectrum disorder in children. JAMA Netw Open. 2018, 1, e180279. [Google Scholar] [CrossRef]
- Roe, K. Autism spectrum disorder initiation by inflammation-facilitated neurotoxin transport. Neurochem Res. 2022, 47, 1150–1165. [Google Scholar] [CrossRef]
- Masini, E.; Loi, E.; Vega-Benedetti, A.F.; Carta, M.; Doneddu, G.; Fadda, R.; Zavattari, P. An overview of the main genetic, epigenetic and environmental factors involved in autism spectrum disorder focusing on synaptic activity. Int J Mol Sci. 2020, 21, 8290. [Google Scholar] [CrossRef]
- Serkan, Y.; Beyazit, U.; Ayhan, A.B. Mycotoxin exposure and autism: a systematic review of the molecular mechanism. Curr Mol Pharmacol. 2021, 14, 853–859. [Google Scholar] [CrossRef]
- Niesler, B.; Rappold, G.A. Emerging evidence for gene mutations driving both brain and gut dysfunction in autism spectrum disorder. Mol Psychiatry. 2021, 26, 1442–1444. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Mao, X.; Dan, Z.; Pei, Y.; Xu, R.; Guo, M.; Liu, K.; Zhang, F.; Chen, J.; Su, C.; Zhuang, Y.; Tang, J.; Xia, Y.; Qin, L.; Hu, Z.; Liu, X. Gene variations in autism spectrum disorder are associated with alteration of gut microbiota, metabolites and cytokines. Gut Microbes. 2021, 13, 1–16. [Google Scholar] [CrossRef]
- Andeweg, S.P.; Keşmir, C.; Dutilh, B.E. Quantifying the impact of human leukocyte antigen on the human gut microbiota. mSphere. 2021, 6, e0047621. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Perlman, A.I.; Twahir, A.; Kempuraj, D. Mast cell activation: beyond histamine and tryptase. Expert Rev Clin Immunol. 2023, 19, 639–654. [Google Scholar] [CrossRef]
- Abraham, S.N.; St John, A.L. Mast cell-orchestrated immunity to pathogens. Nat Rev Immunol. 2010, 10, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Tsilioni, I.; Ren, H. Recent advances in our understanding of mast cell activation - or should it be mast cell mediator disorders? Expert Rev Clin Immunol. 2019, 15, 639–656. [Google Scholar] [CrossRef]
- Kovacheva, E.; Gevezova, M.; Maes, M.; Sarafian, V. Mast cells in autism spectrum disorder-the enigma to be solved? Int J Mol Sci. 2024, 25, 2651. [Google Scholar] [CrossRef]
- Boziki, M.; Theotokis, P.; Kesidou, E.; Nella, M.; Bakirtzis, C.; Karafoulidou, E.; Tzitiridou-Chatzopoulou, M.; Doulberis, M.; Kazakos, E.; Deretzi, G.; Grigoriadis, N.; Kountouras, J. Impact of mast cell activation on neurodegeneration: a potential role for gut-brain axis and helicobacter pylori infection. Neurol Int. 2024, 16, 1750–1778. [Google Scholar] [CrossRef]
- Amin, K. The role of mast cells in allergic inflammation. Respir Med. 2012, 106, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Albert-Bayo, M.; Paracuellos, I.; González-Castro, A.M.; Rodríguez-Urrutia, A.; Rodríguez-Lagunas, M.J.; Alonso-Cotoner, C.; Santos, J.; Vicario, M. Intestinal mucosal mast cells: key modulators of barrier function and homeostasis. Cells. 2019, 8, 135. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Asadi, S.; Chen, J.; Huizinga, J.D. Irritable bowel syndrome and the elusive mast cells. Am J Gastroenterol. 2012, 107, 727–729. [Google Scholar] [CrossRef] [PubMed]
- Petra, A.I.; Panagiotidou, S.; Hatziagelaki, E.; Stewart, J.M.; Conti, P.; Theoharides, T.C. Gut-microbiota-brain axis and its effect on neuropsychiatric disorders with suspected immune Dysregulation. Clin Ther. 2015, 37, 984–995. [Google Scholar] [CrossRef]
- Thoo, L.; Noti, M.; Krebs, P. Keep calm: the intestinal barrier at the interface of peace and war. Cell Death Dis. 2019, 10, 849. [Google Scholar] [CrossRef]
- Di Vincenzo, F.; Del Gaudio, A.; Petito, V.; Lopetuso, L.R.; Scaldaferri, F. Gut microbiota, intestinal permeability, and systemic inflammation: a narrative review. Intern Emerg Med. 2024, 19, 275–293. [Google Scholar] [CrossRef]
- Potrykus, M.; Czaja-Stolc, S.; Stankiewicz, M.; Kaska, Ł.; Małgorzewicz, S. Intestinal microbiota as a contributor to chronic inflammation and its potential modifications. Nutrients. 2021, 13, 3839. [Google Scholar] [CrossRef]
- Solanki, R.; Karande, A.; Ranganathan, P. Emerging role of gut microbiota dysbiosis in neuroinflammation and neurodegeneration. Front Neurol. 2023, 14, 1149618. [Google Scholar] [CrossRef] [PubMed]
- Boziki, M.; Theotokis, P.; Kesidou, E.; Nella, M.; Bakirtzis, C.; Karafoulidou, E.; Tzitiridou-Chatzopoulou, M.; Doulberis, M.; Kazakos, E.; Deretzi, G.; Grigoriadis, N.; Kountouras, J. Impact of mast cell activation on neurodegeneration: a potential role for gut-brain axis and helicobacter pylori infection. Neurol Int. 2024, 16, 1750–1778. [Google Scholar] [CrossRef]
- Renga, G.; Bellet, M.M.; Stincardini, C.; Pariano, M.; Oikonomou, V.; Villella, V.R.; Brancorsini, S.; Clerici, C.; Romani, L.; Costantini, C. To be or not to be a pathogen: candida albicans and celiac disease. Front Immunol. 2019, 10, 2844. [Google Scholar] [CrossRef]
- Smolinska, S.; Jutel, M.; Crameri, R.; O'Mahony, L. Histamine and gut mucosal immune regulation. Allergy. 2014, 69, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Potts, R.A.; Tiffany, C.M.; Pakpour, N.; Lokken, K.L.; Tiffany, C.R.; Cheung, K.; Tsolis, R.M.; Luckhart, S. Mast cells and histamine alter intestinal permeability during malaria parasite infection. Immunobiology. 2016, 221, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Tan, Y.; Huan, R.; Guo, J.; Yang, S.; Deng, M.; Xiong, Y.; Han, G.; Liu, L.; Liu, J.; Cheng, Y.; Zha, Y.; Zhang, J. Mast cell activation mediates blood-brain barrier impairment and cognitive dysfunction in septic mice in a histamine-dependent pathway. Front Immunol. 2023, 14, 1090288. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Akbari, P.; Garssen, J.; Fink-Gremmels, J.; Braber, S. Epithelial integrity, junctional complexes, and biomarkers associated with intestinal functions. Tissue Barriers. 2022, 10, 1996830. [Google Scholar] [CrossRef]
- Poto, R.; Fusco, W.; Rinninella, E.; Cintoni, M.; Kaitsas, F.; Raoul, P.; Caruso, C.; Mele, M.C.; Varricchi, G.; Gasbarrini, A.; Cammarota, G.; Ianiro, G. The role of gut microbiota and leaky gut in the pathogenesis of food allergy. Nutrients. 2023, 16, 92. [Google Scholar] [CrossRef]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch Toxicol. 2017, 91, 1007–1029. [Google Scholar] [CrossRef]
- Akiho, H.; Ihara, E.; Nakamura, K. Low-grade inflammation plays a pivotal role in gastrointestinal dysfunction in irritable bowel syndrome. World J Gastrointest Pathophysiol. 2010, 1, 97–105. [Google Scholar] [CrossRef]
- Ray, K. Bacterial histamine and abdominal pain in IBS. Nat Rev Gastroenterol Hepatol, 2022, 19, 623. [Google Scholar] [CrossRef]
- Rothenberg, M.E. An allergic basis for abdominal pain. N Engl J Med. 2021, 384, 2156–2158. [Google Scholar] [CrossRef]
- Kawamoto, S.; Maruya, M.; Kato, L.M.; Suda, W.; Atarashi, K.; Doi, Y.; Tsutsui, Y.; Qin, H.; Honda, K.; Okada, T.; Hattori, M.; Fagarasan, S. Foxp3(+) T cells regulate immunoglobulin a selection and facilitate diversification of bacterial species responsible for immune homeostasis. Immunity. 2014, 41, 152–165. [Google Scholar] [CrossRef]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017, 10, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: meta-analysis of published experiments in animals. Toxins (Basel). 2013, 5, 396–430. [Google Scholar] [CrossRef]
- Yang, Y.; Palm, N.W. Immunoglobulin A and the microbiome. Curr Opin Microbiol. 2020, 56, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Doron, I.; Kusakabe, T.; Iliev, I.D. Immunoglobulins at the interface of the gut mycobiota and anti-fungal immunity. Semin Immunol. 2023, 67, 101757. [Google Scholar] [CrossRef]
- Suzuki, K.; Ha, S.A.; Tsuji, M.; Fagarasan, S. Intestinal IgA synthesis: a primitive form of adaptive immunity that regulates microbial communities in the gut. Semin Immunol. 2007, 19, 127–135. [Google Scholar] [CrossRef]
- Kawamoto, S.; Maruya, M.; Kato, L.M.; Suda, W.; Atarashi, K.; Doi, Y.; Tsutsui, Y.; Qin, H.; Honda, K.; Okada, T.; Hattori, M.; Fagarasan, S. Foxp3(+) T cells regulate immunoglobulin a selection and facilitate diversification of bacterial species responsible for immune homeostasis. Immunity. 2014, 41, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Kato, L.M.; Kawamoto, S.; Maruya, M.; Fagarasan, S. The role of the adaptive immune system in regulation of gut microbiota. Immunol Rev. 2014, 260, 67–75. [Google Scholar] [CrossRef]
- Suhr, M.J.; Hallen-Adams, H.E. The human gut mycobiome: pitfalls and potentials--a mycologist's perspective. Mycologia. 2015, 107, 1057–1073. [Google Scholar] [CrossRef]
- Renga, G.; Moretti, S.; Oikonomou, V.; Borghi, M.; Zelante, T.; Paolicelli, G.; Costantini, C.; De Zuani, M.; Villella, V.R.; Raia, V.; Del Sordo, R.; Bartoli, A.; Baldoni, M.; Renauld, J.C.; Sidoni, A.; Garaci, E.; Maiuri, L.; Pucillo, C.; Romani, L. IL-9 and mast cells are key players of candida albicans commensalism and pathogenesis in the gut. Cell Rep. 2018, 23, 1767–1778. [Google Scholar] [CrossRef]
- Song, P.; Peng, G.; Yue, H.; Ogawa, T.; Ikeda, S.; Okumura, K.; Ogawa, H.; Niyonsaba, F. Candidalysin, a virulence factor of candida albicans, stimulates mast cells by mediating cross-talk between signaling pathways activated by the dectin-1 receptor and MAPKs. J Clin Immunol. 2022, 42, 1009–1025. [Google Scholar] [CrossRef]
- Jiao, Q.; Luo, Y.; Scheffel, J.; Zhao, Z.; Maurer, M. The complex role of mast cells in fungal infections. Exp Dermatol. 2019, 28, 749–755. [Google Scholar] [CrossRef] [PubMed]
- De Zuani, M.; Paolicelli, G.; Zelante, T.; Renga, G.; Romani, L.; Arzese, A.; Pucillo, C.E.M.; Frossi, B. Mast cells respond to candida albican infections and modulate macrophages phagocytosis of the fungus. Front Immunol. 2018, 9, 2829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Aschenbrenner, D.; Yoo, J.Y.; Zuo, T. The gut mycobiome in health, disease, and clinical applications in association with the gut bacterial microbiome assembly. Lancet Microbe. 2022, 3, e969–e983. [Google Scholar] [CrossRef] [PubMed]
- Suhr, M.J.; Hallen-Adams, H.E. The human gut mycobiome: pitfalls and potentials--a mycologist's perspective. Mycologia. 2015, 107, 1057–1073. [Google Scholar] [CrossRef]
- Li, X.V.; Leonardi, I.; Iliev, I.D. Gut Mycobiota in Immunity and Inflammatory Disease. Immunity. 2019, 50, 1365–1379. [Google Scholar] [CrossRef]
- Richard, M.L.; Sokol, H. The gut mycobiota: insights into analysis, environmental interactions and role in gastrointestinal diseases. Nat Rev Gastroenterol Hepatol. 2019, 16, 331–345. [Google Scholar] [CrossRef]
- Chua, R.X.Y.; Tay, M.J.Y.; Ooi, D.S.Q.; Siah, K.T.H.; Tham, E.H.; Shek, L.P.; Meaney, M.J.; Broekman, B.F.P.; Loo, E.X.L. Understanding the link between allergy and neurodevelopmental disorders: a current review of factors and mechanisms. Front Neurol. 2021, 11, 603571. [Google Scholar] [CrossRef]
- Su, Q.; Wong, O.W.H.; Lu, W.; Wan, Y.; Zhang, L.; Xu, W.; Li, M.K.T.; Liu, C.; Cheung, C.P.; Ching, J.Y.L.; Cheong, P.K.; Leung, T.F.; Chan, S.; Leung, P.; Chan, F.K.L.; Ng, S.C. Multikingdom and functional gut microbiota markers for autism spectrum disorder. Nat Microbiol. 2024, 9, 2344–2355, Erratum in: Nat Microbiol. 2025, 10, 600. doi: 10.1038/s41564-024-01900-w. [Google Scholar] [CrossRef]
- Li, Q.; Han, Y.; Dy, A.B.C.; Hagerman, R.J. The gut microbiota and autism spectrum disorders. Front Cell Neurosci. 2017, 11, 120. [Google Scholar] [CrossRef]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct Target Ther. 2021, 6, 291. [Google Scholar] [CrossRef]
- Liew, W.P.; Mohd-Redzwan, S. mycotoxin: its impact on gut health and microbiota. Front Cell Infect Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef]
- Kim, J.Y.; Choi, M.J.; Ha, S.; Hwang, J.; Koyanagi, A.; Dragioti, E.; Radua, J.; Smith, L.; Jacob, L.; Salazar de Pablo, G.; Lee, S.W.; Yon, D.K.; Thompson, T.; Cortese, S.; Lollo, G.; Liang, C.S.; Chu, C.S.; Fusar-Poli, P.; Cheon, K.A.; Shin, J.I.; Solmi, M. Association between autism spectrum disorder and inflammatory bowel disease: A systematic review and meta-analysis. Autism Res. 2022, 15, 340–352. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jaber, V.R.; Pogue, A.I.; Sharfman, N.M.; Taylor, C.; Lukiw, W.J. Lipopolysaccharides (LPSs) as potent neurotoxic glycolipids in alzheimer's disease (AD). Int J Mol Sci. 2022, 23, 12671. [Google Scholar] [CrossRef] [PubMed]
- Candelli, M.; Franza, L.; Pignataro, G.; Ojetti, V.; Covino, M.; Piccioni, A.; Gasbarrini, A.; Franceschi, F. Interaction between Lipopolysaccharide and Gut Microbiota in Inflammatory Bowel Diseases. Int J Mol Sci. 2021, 22, 6242. [Google Scholar] [CrossRef]
- Violi, F.; Cammisotto, V.; Bartimoccia, S.; Pignatelli, P.; Carnevale, R.; Nocella, C. Gut-derived low-grade endotoxaemia, atherothrombosis and cardiovascular disease. Nat Rev Cardiol. 2023, 20, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Emanuele, E.; Orsi, P.; Boso, M.; Broglia, D.; Brondino, N.; Barale, F.; di Nemi, S.U.; Politi, P. Low-grade endotoxemia in patients with severe autism. Neurosci Lett. 2010, 471, 162–165. [Google Scholar] [CrossRef]
- Li, F.; Ke, H.; Wang, S.; Mao, W.; Fu, C.; Chen, X.; Fu, Q.; Qin, X.; Huang, Y.; Li, B.; Li, S.; Xing, J.; Wang, M.; Deng, W. Leaky gut plays a critical role in the pathophysiology of autism in mice by activating the Llpopolysaccharide-mediated toll-like receptor 4-myeloid differentiation factor 88-nuclear factor kappa b signaling pathway. Neurosci Bull. 2023, 39, 911–928. [Google Scholar] [CrossRef]
- Xiao, L.; Yan, J.; Feng, D.; Ye, S.; Yang, T.; Wei, H.; Li, T.; Sun, W.; Chen, J. Critical Role of TLR4 on the microglia activation induced by maternal LPS exposure leading to ASD-like behavior of offspring. Front Cell Dev Biol. 2021, 9, 634837. [Google Scholar] [CrossRef]
- Kirsten, T.B.; Chaves-Kirsten, G.P.; Chaible, L.M.; Silva, A.C.; Martins, D.O.; Britto, L.R.; Dagli, M.L.; Torrão, A.S.; Palermo-Neto, J.; Bernardi, M.M. Hypoactivity of the central dopaminergic system and autistic-like behavior induced by a single early prenatal exposure to lipopolysaccharide. J Neurosci Res. 2012, 90, 1903–1912. [Google Scholar] [CrossRef]
- Kirsten, T.B.; Taricano, M.; Maiorka, P.C.; Palermo-Neto, J.; Bernardi, M.M. Prenatal lipopolysaccharide reduces social behavior in male offspring. Neuroimmunomodulation. 2010, 17, 240–251. [Google Scholar] [CrossRef]
- Kirsten, T.B.; Palermo-Neto, J.; Bernardi, M.M. A rat model of autism induced by a single early prenatal exposure to LPS. Brain, Behavior, and Immunity. 2012, 26, S4. [Google Scholar] [CrossRef]
- Tadas, M.; Wankhede, N.; Chandurkar, P.; Kotagale, N.; Umekar, M.; Katariya, R.; Waghade, A.; Kokare, D.; Taksande, B. Postnatal propionic acid exposure disrupts hippocampal agmatine homeostasis leading to social deficits and cognitive impairment in autism spectrum disorder-like phenotype in rats. Pharmacol Biochem Behav. 2025, 174030. [Google Scholar] [CrossRef]
- Benitah, K.C.; Kavaliers, M.; Ossenkopp, K.P. The enteric metabolite, propionic acid, impairs social behavior and increases anxiety in a rodent ASD model: Examining sex differences and the influence of the estrous cycle. Pharmacol Biochem Behav. 2023, 231, 173630. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C. The endotoxin hypothesis of neurodegeneration. J Neuroinflammation. 2019, 6, 180. [Google Scholar] [CrossRef] [PubMed]
- Saresella, M.; Piancone, F.; Marventano, I.; Zoppis, M.; Hernis, A.; Zanette, M.; Trabattoni, D.; Chiappedi, M.; Ghezzo, A.; Canevini, M.P.; la Rosa, F.; Esposito, S.; Clerici, M. Multiple inflammasome complexes are activated in autistic spectrum disorders. Brain Behav Immun. 2016, 57, 125–133. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An overview of mechanisms of activation and regulation. Int J Mol Sci. 2019, 20, 3328. [Google Scholar] [CrossRef]
- Rogiers, O.; Frising, U.C.; Kucharíková, S.; Jabra-Rizk, M.A.; van Loo, G.; Van Dijck, P.; Wullaert, A. Candidalysin crucially contributes to nlrp3 inflammasome activation by candida albicans hyphae. mBio. 2019, 10, e02221–e18. [Google Scholar] [CrossRef] [PubMed]
- McCurdy, J.D.; Lin, T.J.; Marshall, J.S. Toll-like receptor 4-mediated activation of murine mast cells. J Leukoc Biol. 2001, 70, 977–984. [Google Scholar] [CrossRef]
- Yu, M.; Song, X.T.; Liu, B.; Luan, T.T.; Liao, S.L.; Zhao, Z.T. The emerging role of mast cells in response to fungal infection. Front Immunol. 2021, 12, 688659. [Google Scholar] [CrossRef]
- Gonzalez, A.; Hammock, E.A.D. Oxytocin and microglia in the development of social behaviour. Philos Trans R Soc Lond B Biol Sci. 2022, 377, 20210059. [Google Scholar] [CrossRef]
- Caetano-Silva, M.E.; Rund, L.; Vailati-Riboni, M.; Matt, S.; Soto-Diaz, K.; Beever, J.; Allen, J.M.; Woods, J.A.; Steelman, A.J.; Johnson, R.W. The emergence of inflammatory microglia during gut inflammation is not affected by FFAR2 expression in intestinal epithelial cells or peripheral myeloid cells. Brain Behav Immun. 2024, 118, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Haq, R.; Schlachetzki, J.C.M.; Glass, C.K.; Mazmanian, S.K. Microbiome-microglia connections via the gut-brain axis. J Exp Med. 2019, 216, 41–59. [Google Scholar] [CrossRef]
- Qin, L.; Li, G.; Qian, X.; Liu, Y.; Wu, X.; Liu, B.; Hong, J.S.; Block, M.L. Interactive role of the toll-like receptor 4 and reactive oxygen species in LPS-induced microglia activation. Glia. 2005, 52, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Jalabi, W.; Shpargel, K.B.; Farabaugh, K.T.; Dutta, R.; Yin, X.; Kidd, G.J.; Bergmann, C.C.; Stohlman, S.A.; Trapp, B.D. Lipopolysaccharide-induced microglial activation and neuroprotection against experimental brain injury is independent of hematogenous TLR4. J Neurosci. 2012, 32, 11706–11715. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Zhu, M.; Che, X.; Wang, H.; Liang, X.J.; Wu, C.; Xue, X.; Yang, J. Lipopolysaccharide induces neuroinflammation in microglia by activating the MTOR pathway and downregulating Vps34 to inhibit autophagosome formation. J Neuroinflammation. 2020, 17, 18. [Google Scholar] [CrossRef]
- Jung, H.; Lee, D.; You, H.; Lee, M.; Kim, H.; Cheong, E.; Um, J.W. LPS induces microglial activation and GABAergic synaptic deficits in the hippocampus accompanied by prolonged cognitive impairment. Sci Rep. 2023, 13, 6547. [Google Scholar] [CrossRef]
- Gozal, E.; Jagadapillai, R.; Cai, J.; Barnes, G.N. Potential crosstalk between sonic hedgehog-WNT signaling and neurovascular molecules: Implications for blood-brain barrier integrity in autism spectrum disorder. J Neurochem. 2021, 159, 15–28. [Google Scholar] [CrossRef]
- Wei, H.; Alberts, I.; Li, X. The apoptotic perspective of autism. Int J Dev Neurosci. 2014, 36, 13–18. [Google Scholar] [CrossRef]
- Hines, D.J.; Choi, H.B.; Hines, R.M.; Phillips, A.G.; MacVicar, B.A. Prevention of LPS-induced microglia activation, cytokine production and sickness behavior with TLR4 receptor interfering peptides. PLoS One. 2013, 8, e60388. [Google Scholar] [CrossRef]
- Bassi, G.S.; Kanashiro, A.; Santin, F.M.; de Souza, G.E.; Nobre, M.J.; Coimbra, N.C. Lipopolysaccharide-induced sickness behaviour evaluated in different models of anxiety and innate fear in rats. Basic Clin Pharmacol Toxicol. 2012, 110, 359–369. [Google Scholar] [CrossRef]
- Biesmans, S.; Meert, T.F.; Bouwknecht, J.A.; Acton, P.D.; Davoodi, N.; De Haes, P.; Kuijlaars, J.; Langlois, X.; Matthews, L.J.; Ver Donck, L.; Hellings, N.; Nuydens, R. Systemic immune activation leads to neuroinflammation and sickness behavior in mice. Mediators Inflamm. 2013, 271359. [Google Scholar] [CrossRef] [PubMed]
- Onore, C.; Careaga, M.; Ashwood, P. The role of immune dysfunction in the pathophysiology of autism. Brain Behav Immun. 2012, 26, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Behrens, M.; Hüwel, S.; Galla, H.J.; Humpf, H.U. Blood-Brain Barrier Effects of the Fusarium Mycotoxins Deoxynivalenol, 3 Acetyldeoxynivalenol, and moniliformin and their transfer to the brain. PLoS One. 2015, 10, e0143640. [Google Scholar] [CrossRef]
- Patel, R.; Hossain, M.A.; German, N.; Al-Ahmad, A.J. Gliotoxin penetrates and impairs the integrity of the human blood-brain barrier in vitro. Mycotoxin Res. 2018, 34, 257–268. [Google Scholar] [CrossRef]
- Doi, K.; Uetsuka, K. Mechanisms of mycotoxin-induced neurotoxicity through oxidative stress-associated pathways. Int J Mol Sci. 2011, 12, 5213–5237. [Google Scholar] [CrossRef]
- Abbott, N.J. Inflammatory mediators and modulation of blood-brain barrier permeability. Cell Mol Neurobiol. 2000, 20, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Song, R.; Liu, Y.; Wu, Z.; Zhang, X. Effects of ultra-processed foods on the microbiota-gut-brain axis: The bread-and-butter issue. Food Res Int. 2023, 167, 112730. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Konstantinidou, A.D. Corticotropin-releasing hormone and the blood-brain-barrier. Front Biosci. 2007, 12, 1615–1628. [Google Scholar] [CrossRef]
- Esposito, P.; Chandler, N.; Kandere, K.; Basu, S.; Jacobson, S.; Connolly, R.; Tutor, D.; Theoharides, T.C. Corticotropin-releasing hormone and brain mast cells regulate blood-brain-barrier permeability induced by acute stress. J Pharmacol Exp Ther. 2002, 303, 1061–1066. [Google Scholar] [CrossRef]
- Theoharides, T.C. Mast cells: the immune gate to the brain. Life Sci. 1990, 46, 607–617. [Google Scholar] [CrossRef]
- Schafer, D.P.; Lehrman, E.K.; Kautzman, A.G.; Koyama, R.; Mardinly, A.R.; Yamasaki, R.; Ransohoff, R.M.; Greenberg, M.E.; Barres, B.A.; Stevens, B. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron. 2012, 74, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Hammock, E.A.D. Oxytocin and microglia in the development of social behaviour. Philos Trans R Soc Lond B Biol Sci. 2022, 377, 20210059. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Chen, J.; Li, Y. Microglia and astrocytes underlie neuroinflammation and synaptic susceptibility in autism spectrum disorder. Front Neurosci. 2023, 17, 1125428. [Google Scholar] [CrossRef]
- McNamara, N.B.; Munro, D.A.D.; Bestard-Cuche, N.; Uyeda, A.; Bogie, J.F.J.; Hoffmann, A.; Holloway, R.K.; Molina-Gonzalez, I.; Askew, K.E.; Mitchell, S.; Mungall, W.; Dodds, M.; Dittmayer, C.; Moss, J.; Rose, J.; Szymkowiak, S.; Amann, L.; McColl, B.W.; Prinz, M.; Spires-Jones, T.L.; Stenzel, W.; Horsburgh, K.; Hendriks, J.J.A.; Pridans, C.; Muramatsu, R.; Williams, A.; Priller, J.; Miron, V.E. Microglia regulate central nervous system myelin growth and integrity. Nature. 2023, 613, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Liu, C.; Deng, S.; Gan, L.; Zhang, Z.; Yang, G.Y.; Tian, H.; Tang, Y. The roles of microglia and astrocytes in myelin phagocytosis in the central nervous system. J Cereb Blood Flow Metab. 2023, 43, 325–340. [Google Scholar] [CrossRef]
- Marín-Teva, J.L.; Cuadros, M.A.; Martín-Oliva, D.; Navascués, J. Microglia and neuronal cell death. Neuron Glia Biol. 2011, 7, 25–40. [Google Scholar] [CrossRef]
- Tay, T.L.; Béchade, C.; D'Andrea, I.; St-Pierre, M.K.; Henry, M.S.; Roumier, A.; Tremblay, M.E. Microglia gone rogue: impacts on lsychiatric disorders across the lifespan. Front Mol Neurosci. 2018, 10, 421. [Google Scholar] [CrossRef]
- Rodriguez, J.I.; Kern, J.K. Evidence of microglial activation in autism and its possible role in brain underconnectivity. Neuron Glia Biol. 2011, 7, 205–213. [Google Scholar] [CrossRef]
- Gupta, S.; Ellis, S.E.; Ashar, F.N.; Moes, A.; Bader, J.S.; Zhan, J.; West, A.B.; Arking, D.E. Transcriptome analysis reveals dysregulation of innate immune response genes and neuronal activity-dependent genes in autism. Nat Commun. 2014, 5, 5748. [Google Scholar] [CrossRef]
- Frick, L.R.; Williams, K.; Pittenger, C. Microglial dysregulation in psychiatric disease. Clin Dev Immunol. 2013, 608654. [Google Scholar] [CrossRef] [PubMed]
- Petrelli, F.; Pucci, L.; Bezzi, P. Astrocytes and microglia and their potential link with autism spectrum disorders. Front Cell Neurosci. 2016, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kavalioti, M.; Tsilioni, I. Mast cells, stress, fear and autism spectrum disorder. Int J Mol Sci. 2019, 20, 3611. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, F.V.; Hamani, C.; Fonoff, E.T.; Brentani, H.; Alho, E.J.L.; de Morais, R.M.C.B.; de Souza, A.L.; Rigonatti, S.P.; Martinez, R.C.R. Amygdala and hypothalamus: historical overview with focus on aggression. Neurosurgery. 2019, 85, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Melbourne, J.K.; Chandler, C.M.; Van Doorn, C.E.; Bardo, M.T.; Pauly, J.R.; Peng, H.; Nixon, K. Primed for addiction: A critical review of the role of microglia in the neurodevelopmental consequences of adolescent alcohol drinking. Alcohol Clin Exp Res. 2021, 45, 1908–1926. [Google Scholar] [CrossRef]
- da Silva, M.C.M.; Iglesias, L.P.; Candelario-Jalil, E.; Khoshbouei, H.; Moreira, F.A.; de Oliveira, A.CP. Role of microglia in psychostimulant addiction. Curr Neuropharmacol. 2023, 21, 235–259. [Google Scholar] [CrossRef]
- Won, E.; Kim, Y.K. Neuroinflammation-associated alterations of the brain as potential neural biomarkers in anxiety disorders. Int J Mol Sci. 2020, 21, 6546. [Google Scholar] [CrossRef]
- Luo, Y. The crosstalk between the “inflamed” mind and the “impulsive” mind: activation of microglia and impulse control disorders. Second International Conference on Biological Engineering and Medical Science. 2023, 126112H. [Google Scholar] [CrossRef]
- Yokokura, M.; Takebasashi, K.; Takao, A.; Nakaizumi, K.; Yoshikawa, E.; Futatsubashi, M.; Suzuki, K.; Nakamura, K.; Yamasue, H.; Ouchi, Y. In vivo imaging of dopamine D1 receptor and activated microglia in attention-deficit/hyperactivity disorder: a positron emission tomography study. Mol Psychiatry. 2021, 26, 4958–4967. [Google Scholar] [CrossRef]
- Zhu, H.; Guan, A.; Liu, J.; Peng, L.; Zhang, Z.; Wang, S. Noteworthy perspectives on microglia in neuropsychiatric disorders. J Neuroinflammation. 2023, 20, 223. [Google Scholar] [CrossRef]
- Underwood, J.F.G.; DelPozo-Banos, M.; Frizzati, A.; Rai, D.; John, A.; Hall, J. Neurological and psychiatric disorders among autistic adults: a population healthcare record study. Psychol Med. 2023, 53, 5663–5673. [Google Scholar] [CrossRef]
- White, S.W.; Oswald, D.; Ollendick, T.; Scahill, L. Anxiety in children and adolescents with autism spectrum disorders. Clin Psychol Rev. 2009, 29, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Montaser, J.; Umeano, L.; Pujari, H.P.; Nasiri, S.M.Z.; Parisapogu, A.; Shah, A.; Khan, S. Correlations between the development of social anxiety and individuals with autism spectrum disorder: a systematic review. Cureus. 2023, 15, e44841. [Google Scholar] [CrossRef] [PubMed]
- Mayes SD, Calhoun SL, Aggarwal R, Baker C, Mathapati S, Molitoris S, Mayes RD. Unusual fears in children with autism. Research in Autism Spectrum Disorders. 2013, 7, 151–158. [CrossRef]
- Patel, S.; Day, T.N.; Jones, N.; Mazefsky, C.A. Association between anger rumination and autism symptom severity, depression symptoms, aggression, and general dysregulation in adolescents with autism spectrum disorder. Autism. 2017, 21, 181–189. [Google Scholar] [CrossRef]
- Lugo-Huitrón, R.; Ugalde Muñiz, P.; Pineda, B.; Pedraza-Chaverrí, J.; Ríos, C.; Pérez-de la Cruz, V. Quinolinic acid: an endogenous neurotoxin with multiple targets. Oxid Med Cell Longev. 2013, 104024. [Google Scholar] [CrossRef]
- Yildirim, V.; Simsek, S.; Cetin, I.; Dokuyucu, R. Kynurenine, kynurenic acid, quinolinic acid and interleukin-6 levels in the serum of patients with autism spectrum disorder. Medicina (Kaunas). 2023, 59, 1906. [Google Scholar] [CrossRef]
- Carthy, E.; Ellender, T. Histamine, Neuroinflammation and Neurodevelopment: A Review. Front Neurosci. 2021, 15, 680214. [Google Scholar] [CrossRef]
- Lasselin, J.; Lekander, M.; Benson, S.; Schedlowski, M.; Engler, H. Sick for science: experimental endotoxemia as a translational tool to develop and test new therapies for inflammation-associated depression. Mol Psychiatry. 2021, 26, 3672–3683. [Google Scholar] [CrossRef]
- Theoharides, T.C. Mast cells: the immune gate to the brain. Life Sci. 1990, 46, 607–617. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L.; Giusti, P. Mast cells, glia and neuroinflammation: partners in crime? Immunology. 2014, 141, 314–327. [Google Scholar] [CrossRef]
- Zhang, S.; Zeng, X.; Yang, H.; Hu, G.; He, S. Mast cell tryptase induces microglia activation via protease-activated receptor 2 signaling. Cell Physiol Biochem. 2012, 29, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Dong, H.; Xu, Y.; Zhang, S. Induction of microglial activation by mediators released from mast cells. Cell Physiol Biochem. 2016, 38, 1520–1531. [Google Scholar] [CrossRef]
- Conesa, M.P.B.; Blixt, F.W.; Peesh, P.; Khan, R.; Korf, J.; Lee, J.; Jagadeesan, G.; Andersohn, A.; Das, T.K.; Tan, C.; Di Gesu, C.; Colpo, G.D.; Moruno-Manchón, J.F.; McCullough, L.D.; Bryan, R.; Ganesh, B.P. Stabilizing histamine release in gut mast cells mitigates peripheral and central inflammation after stroke. J Neuroinflammation. 2023, 20, 230. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kavalioti, M.; Tsilioni, I. Mast cells, stress, fear and autism spectrum disorder. Int J Mol Sci. 2019, 20, 3611. [Google Scholar] [CrossRef]
- Theoharides, T.C. The impact of psychological stress on mast cells. Ann Allergy Asthma Immunol. 2020, 25, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Stewart, J.M.; Panagiotidou, S.; Melamed, I. Mast cells, brain inflammation and autism. Eur J Pharmacol. 2016, 778, 96–102. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Doyle, R. Autism, gut-blood-brain barrier, and mast cells. J Clin Psychopharmacol. 2008, 28, 479–483. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Zhang, B. Neuro-inflammation, blood-brain barrier, seizures and autism. J Neuroinflammation. 2011, 8, 168. [Google Scholar] [CrossRef]
- Theoharides, T.C. Is a subtype of autism an allergy of the brain? Clin Ther. 2013, 35, 584–591. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Stewart, J.M.; Panagiotidou, S.; Melamed, I. Mast cells, brain inflammation and autism. Eur J Pharmacol. 2016, 778, 96–102. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Asadi, S.; Panagiotidou, S.; Weng, Z. The "missing link" in autoimmunity and autism: extracellular mitochondrial components secreted from activated live mast cells. Autoimmun Rev. 2013, 12, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, K.H.; Moldestad, O.; Eide, L.; Carlsen, H.; Nesse, G.; Storm, J.F.; Mansuy, I.M.; Bergersen, L.H.; Klungland, A. Mitochondrial DNA toxicity in forebrain neurons causes apoptosis, neurodegeneration, and impaired behavior. Mol Cell Biol. 2010, 30, 1357–1367. [Google Scholar] [CrossRef]
- Duncan, J.G.; Waton, N.G. Absorption of histamine from the gastrointestinal tract of dogs in vivo. J Physiol. 1968, 198, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Scammell, T.E.; Jackson, A.C.; Franks, N.P.; Wisden, W.; Dauvilliers, Y. Histamine: neural circuits and new medications. Sleep. 2019, 42, zsy183. [Google Scholar] [CrossRef]
- Wang, Y.; Sha, H.; Zhou, L.; Chen, Y.; Zhou, Q.; Dong, H.; Qian, Y. The Mast Cell Is an Early Activator of Lipopolysaccharide-Induced Neuroinflammation and blood-brain barrier dysfunction in the jippocampus. Mediators Inflamm. 2020, 8098439. [Google Scholar] [CrossRef]
- Haas, H.L.; Sergeeva, O.A.; Selbach, O. Histamine in the nervous system. Physiol Rev. 2008, 88, 1183–1241. [Google Scholar] [CrossRef] [PubMed]
- Devnani, P.A.; Hegde, A.U. Autism and sleep disorders. J Pediatr Neurosci. 2015, 10, 304–307. [Google Scholar] [CrossRef]
- Ji, W.; Suga, N. Histaminergic modulation of nonspecific plasticity of the auditory system and differential gating. J Neurophysiol. 2013, 109, 792–802. [Google Scholar] [CrossRef]
- Fabian, R.; Seyfarth, E.A. Acetylcholine and histamine are transmitter candidates in identifiable mechanosensitive neurons of the spider Cupiennius salei: an immunocytochemical study. Cell Tissue Res. 1997, 287, 413–423. [Google Scholar] [CrossRef]
- Orona, E.; Ache, B.W. Physiological and pharmacological evidence for histamine as a neurotransmitter in the olfactory CNS of the spiny lobster. Brain Res. 1992, 590, (1–2), 136. [Google Scholar] [CrossRef]
- Saure, E.; Lepistö-Paisley, T.; Raevuori, A.; Laasonen, M. Atypical sensory processing is associated with lower body mass index and increased aating disturbance in individuals with anorexia nervosa. Front Psychiatry. 2022, 13, 850594. [Google Scholar] [CrossRef] [PubMed]
- Kojovic, N.; Ben Hadid, L.; Franchini, M.; Schaer, M. Sensory processing issues and their association with social difficulties in children with autism spectrum disorders. J Clin Med. 2019, 8, 1508. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Choi, M.J.; Ha, S.; Hwang, J.; Koyanagi, A.; Dragioti, E.; Radua, J.; Smith, L.; Jacob, L.; Salazar de Pablo, G.; Lee, S.W.; Yon, D.K.; Thompson, T.; Cortese, S.; Lollo, G.; Liang, C.S.; Chu, C.S.; Fusar-Poli, P.; Cheon, K.A.; Shin, J.I.; Solmi, M. Association between autism spectrum disorder and inflammatory bowel disease: A systematic review and meta-analysis. Autism Res. 2022, 15, 340–352. [Google Scholar] [CrossRef]
- Fu, J.; Zheng, Y.; Gao, Y.; Xu, W. Dietary Fiber Intake and Gut Microbiota in Human Health. Microorganisms. 2022, 10, 2507. [Google Scholar] [CrossRef]
- Hoffmann, C.; Dollive, S.; Grunberg, S.; Chen, J.; Li, H.; Wu, G.D.; Lewis, J.D.; Bushman, F.D. Archaea and fungi of the human gut microbiome: correlations with diet and bacterial residents. PLoS One. 2013, 8, e66019. [Google Scholar] [CrossRef]
- Erlanson-Albertsson, C.; Stenkula, K.G. The importance of food for endotoxemia and an inflammatory response. Int J Mol Sci. 2021, 22, 9562. [Google Scholar] [CrossRef]
- Jochum, C. Histamine intolerance: symptoms, diagnosis, and beyond. Nutrients. 2024, 16, 1219. [Google Scholar] [CrossRef] [PubMed]
- Tsilioni, I.; Taliou, A.; Francis, K.; Theoharides, T.C. Children with autism spectrum disorders, who improved with a luteolin-containing dietary formulation, show reduced serum levels of TNF and IL-6. Transl Psychiatry. 2015, 5, e647. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Asadi, S.; Panagiotidou, S. A case series of a luteolin formulation (NeuroProtek®) in children with autism spectrum disorders. Int J Immunopathol Pharmacol. 2012, 25, 317–323. [Google Scholar] [CrossRef]
- Taliou, A.; Zintzaras, E.; Lykouras, L.; Francis, K. An open-label pilot study of a formulation containing the anti-inflammatory flavonoid luteolin and its effects on behavior in children with autism spectrum disorders. Clin Ther. 2013, 35, 592–602. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.M. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr Cancer Drug Targets. 2008, 8, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C. Luteolin: The wonder flavonoid. Biofactors. 2021, 47, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Kim, M.Y.; Cho, J.Y. Immunopharmacological activities of luteolin in chronic diseases. Int J Mol Sci. 2023, 24, 2136. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kempuraj, D.; Iliopoulou, B.P. Mast cells, T cells, and inhibition by luteolin: implications for the pathogenesis and treatment of multiple sclerosis. Adv Exp Med Biol. 2007, 601, 423–430. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Tsilioni, I.; Ren, H. Recent advances in our understanding of mast cell activation - or should it be mast cell mediator disorders? Expert Rev Clin Immunol. 2019, 15, 639–656. [Google Scholar] [CrossRef]
- Tsilioni, I.; Taliou, A.; Francis, K.; Theoharides, T.C. Children with autism spectrum disorders, who improved with a luteolin-containing dietary formulation, show reduced serum levels of TNF and IL-6. Transl Psychiatry. 2015, 5, e647. [Google Scholar] [CrossRef]
- Tsilioni, I.; Theoharides, T. Luteolin is more potent than cromolyn in their ability to inhibit mediator release from cultured human mast cells. Int Arch Allergy Immunol. 2024, 185, 803–809. [Google Scholar] [CrossRef]
- Jang, S.; Dilger, R.N.; Johnson, R.W. Luteolin inhibits microglia and alters hippocampal-dependent spatial working memory in aged mice. J Nutr. 2010, 140, 1892–1898. [Google Scholar] [CrossRef]
- Burton, M.D.; Rytych, J.L.; Amin, R.; Johnson, R.W. Dietary luteolin reduces proinflammatory microglia in the brain of senescent mice. Rejuvenation Res. 2016, 19, 286–292. [Google Scholar] [CrossRef]
- Chagas, M.D.S.D.S.; Moragas Tellis, C.J.; Silva, A.R.; Brito, M.A.D.S.M.; Teodoro, A.J.; de Barros Elias, M.; Ferrarini, S.R.; Behrens, M.D.; Gonçalves-de-Albuquerque, C.F. Luteolin: A novel approach to fight bacterial infection. Microb Pathog. 2025, 204, 107519. [Google Scholar] [CrossRef]
- Weng, Z.; Zhang, B.; Asadi, S.; Sismanopoulos, N.; Butcher, A.; Fu, X.; Katsarou-Katsari, A.; Antoniou, C.; Theoharides, T.C. Quercetin is more effective than cromolyn in blocking human mast cell cytokine release and inhibits contact dermatitis and photosensitivity in humans. PLoS One. 2012, 7, e33805. [Google Scholar] [CrossRef]
- Jafarinia, M.; Sadat Hosseini, M.; Kasiri, N.; Fazel, N.; Fathi, F.; Ganjalikhani Hakemi, M.; Eskandari, N. Quercetin with the potential effect on allergic diseases. Allergy Asthma Clin Immunol. 2020, 16, 36. [Google Scholar] [CrossRef]
- Shi, T.; Bian, X.; Yao, Z.; Wang, Y.; Gao, W.; Guo, C. Quercetin improves gut dysbiosis in antibiotic-treated mice. Food Funct. 2020, 11, 8003–8013. [Google Scholar] [CrossRef]
- Feng, J.; Li, Z.; Ma, H.; Yue, Y.; Hao, K.; Li, J.; Xiang, Y.; Min, Y. Quercetin alleviates intestinal inflammation and improves intestinal functions via modulating gut microbiota composition in LPS-challenged laying hens. Poult Sci. 2023, 102, 102433. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, P.; Polissidis, A.; Kythreoti, G.; Sagnou, M.; Stefanatou, A.; Theoharides, T.C. Anti-inflammatory and neuroprotective polyphenols derived from the european olive tree, Olea europaea L., in long COVID and other conditions involving cognitive impairment. Int J Mol Sci. 2024, 25, 11040. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C. Luteolin supplements: All that glitters is not gold. Biofactors. 2021, 47, 242–244. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J Physiol Pharmacol. 2012, 63, 497–503. [Google Scholar]
- Patel, A.B.; Tsilioni, I.; Leeman, S.E.; Theoharides, T.C. Neurotensin stimulates sortilin and mTOR in human microglia inhibitable by methoxyluteolin, a potential therapeutic target for autism. Proc Natl Acad Sci U S A. 2016, 113, E7049–E7058, Erratum in: Proc Natl Acad Sci U S A. 2016, 113, E7138. doi: 10.1073/pnas.1616587113. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Patel, A.B.; Theoharides, T.C. Methoxyluteolin inhibits neuropeptide-stimulated proinflammatory mediator release via mTOR activation from human mast cells. J Pharmacol Exp Ther. 2017, 361, 462–471. [Google Scholar] [CrossRef]
- Taracanova, A.; Tsilioni, I.; Conti, P.; Norwitz, E.R.; Leeman, S.E.; Theoharides, T.C. Substance P and IL-33 administered together stimulate a marked secretion of IL-1β from human mast cells, inhibited by methoxyluteolin. Proc Natl Acad Sci U S A. 2018, 115, E9381–E9390. [Google Scholar] [CrossRef]
- Bawazeer, M.A.; Theoharides, T.C. IL-33 stimulates human mast cell release of CCL5 and CCL2 via MAPK and NF-κB, inhibited by methoxyluteolin. Eur J Pharmacol. 2019, 865, 172760. [Google Scholar] [CrossRef]
- Weng, Z.; Patel, A.B.; Panagiotidou, S.; Theoharides, T.C. The novel flavone tetramethoxyluteolin is a potent inhibitor of human mast cells. J Allergy Clin Immunol. 2015, 135, 1044–1052.e5. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Stewart, J.M.; Tsilioni, I. Tolerability and benefit of a tetramethoxyluteolin-containing skin lotion. Int J Immunopathol Pharmacol. 2017, 30, 146–151. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, D.; Negro, L.; Vaia, M.; Cinelli, M.P.; Iuvone, T. New insights in mast cell modulation by palmitoylethanolamide. CNS Neurol Disord Drug Targets. 2013, 12, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Schiano Moriello, A. Palmitoylethanolamide: a nutritional approach to keep neuroinflammation within physiological boundaries-a systematic review. Int J Mol Sci. 2020, 21, 9526. [Google Scholar] [CrossRef]
- Landolfo, E.; Cutuli, D.; Petrosini, L.; Caltagirone, C. Effects of palmitoylethanolamide on neurodegenerative diseases: a review from rodents to humans. Biomolecules. 2022, 12, 667. [Google Scholar] [CrossRef]
- Cerrato, S.; Brazis, P.; della Valle, M.F.; Miolo, A.; Puigdemont, A. Effects of palmitoylethanolamide on immunologically induced histamine, PGD2 and TNFalpha release from canine skin mast cells. Vet Immunol Immunopathol. 133, 9–15. [CrossRef]
- D'Aloia, A.; Molteni, L.; Gullo, F.; Bresciani, E.; Artusa, V.; Rizzi, L.; Ceriani, M.; Meanti, R.; Lecchi, M.; Coco, S.; Costa, B.; Torsello, A. Palmitoylethanolamide modulation of microglia activation: characterization of mechanisms of action and implication for Its neuroprotective effects. Int J Mol Sci. 2021, 22, 3054. [Google Scholar] [CrossRef]
- Antonucci, N.; Cirillo, A.; Siniscalco, D. Beneficial effects of palmitoylethanolamide on expressive language, cognition, and behaviors in autism: a report of two cases. Case Rep Psychiatry. 2015, 325061. [Google Scholar] [CrossRef]
- Nosková, E.; Vochosková, K.; Knop, V.; Stopková, P.; Kopeček, M. Histamine intolerance and anxiety disorders: pilot cross-sectional study of histamine intolerance prevalence in cohort of patients with anxiety disorders. Eur Psychiatry. 2022, 65 (Suppl 1), S387–8. [Google Scholar] [CrossRef]
- Schnedl, W.J.; Schenk, M.; Lackner, S.; Enko, D.; Mangge, H.; Forster, F. Diamine oxidase supplementation improves symptoms in patients with histamine intolerance. Food Sci Biotechnol. 2019, 28, 1779–1784. [Google Scholar] [CrossRef]
- Izquierdo-Casas, J.; Comas-Basté, O.; Latorre-Moratalla, M.L.; Lorente-Gascón, M.; Duelo, A.; Soler-Singla, L.; Vidal-Carou, M.C. Diamine oxidase (DAO) supplement reduces headache in episodic migraine patients with DAO deficiency: A randomized double-blind trial. Clin Nutr. 2019, 38, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhang, J.; You, H.; Ye, F.; Yang, Y.; Zhu, C.; Jiang, Y.C.; Tang, Z.X. Berberine ameliorates inflammation by inhibiting MrgprB2 receptor-mediated activation of mast cell in mice. Eur J Pharmacol. 2024, 985, 177109. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Cui, Y.; Zhang, Y.; Fu, W.; Liu, Y.; Liu, F. Berberine hydrochloride enhances innate immunity to protect against pathogen infection via p38 MAPK pathway. Front Immunol. 2025, 16, 1536143. [Google Scholar] [CrossRef] [PubMed]
- Gori A, Brindisi G, Daglia M, Giudice MMD, Dinardo G, Di Minno A, Drago L, Indolfi C, Naso M, Trincianti C, Tondina E, Brunese FP, Ullah H, Varricchio A, Ciprandi G, Zicari AM; Nutraceutical and Medical Device Task Force of the Italian Society of Pediatric Allergy, Immunology (SIAIP). Exploring the role of lactoferrin in managing allergic airway diseases among children: unrevealing a potential breakthrough. Nutrients. 2024, 16, 1906. [CrossRef]
- Gunning, L.; O'Sullivan, M.; Boutonnet, C.; Pedrós-Garrido, S.; Jacquier, J.C. Effect of in vitro simulated gastrointestinal digestion on the antibacterial properties of bovine lactoferrin. J Dairy Res. 2024, 1–8. [Google Scholar] [CrossRef]
- Dev, S.; Mizuguchi, H.; Das, A.K.; Matsushita, C.; Maeyama, K.; Umehara, H.; Ohtoshi, T.; Kojima, J.; Nishida, K.; Takahashi, K.; Fukui, H. Suppression of histamine signaling by probiotic Lac-B: a possible mechanism of its anti-allergic effect. J Pharmacol Sci. 2008, 107, 159–166. [Google Scholar] [CrossRef]
- Dong, J.; Ping, L.; Cao, T.; Sun, L.; Liu, D.; Wang, S.; Huo, G.; Li, B. Immunomodulatory effects of the Bifidobacterium longum BL-10 on lipopolysaccharide-induced intestinal mucosal immune injury. Front Immunol. 2022, 13, 947755. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Frye, R.E. Cerebral folate deficiency, folate receptor alpha autoantibodies and leucovorin (folinic acid) treatment in autism spectrum disorders: a systematic review and meta-analysis. J Pers Med. 2021, 11, 1141, Erratum in: J Pers Med. 2022, 12, 721. DOI: 10.3390/jpm12050721. [Google Scholar] [CrossRef]
- Shuid, A.N.; Jayusman, P.A.; Shuid, N.; Ismail, J.; Kamal Nor, N.; Mohamed, I.N. Association between viral infections and risk of autistic disorder: an overview. Int J Environ Res Public Health. 2021, 18, 2817. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Genuis, S.J.; Frye, R.E. Environmental toxicants and autism spectrum disorders: a systematic review. Transl Psychiatry. 2014, 4, e360. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
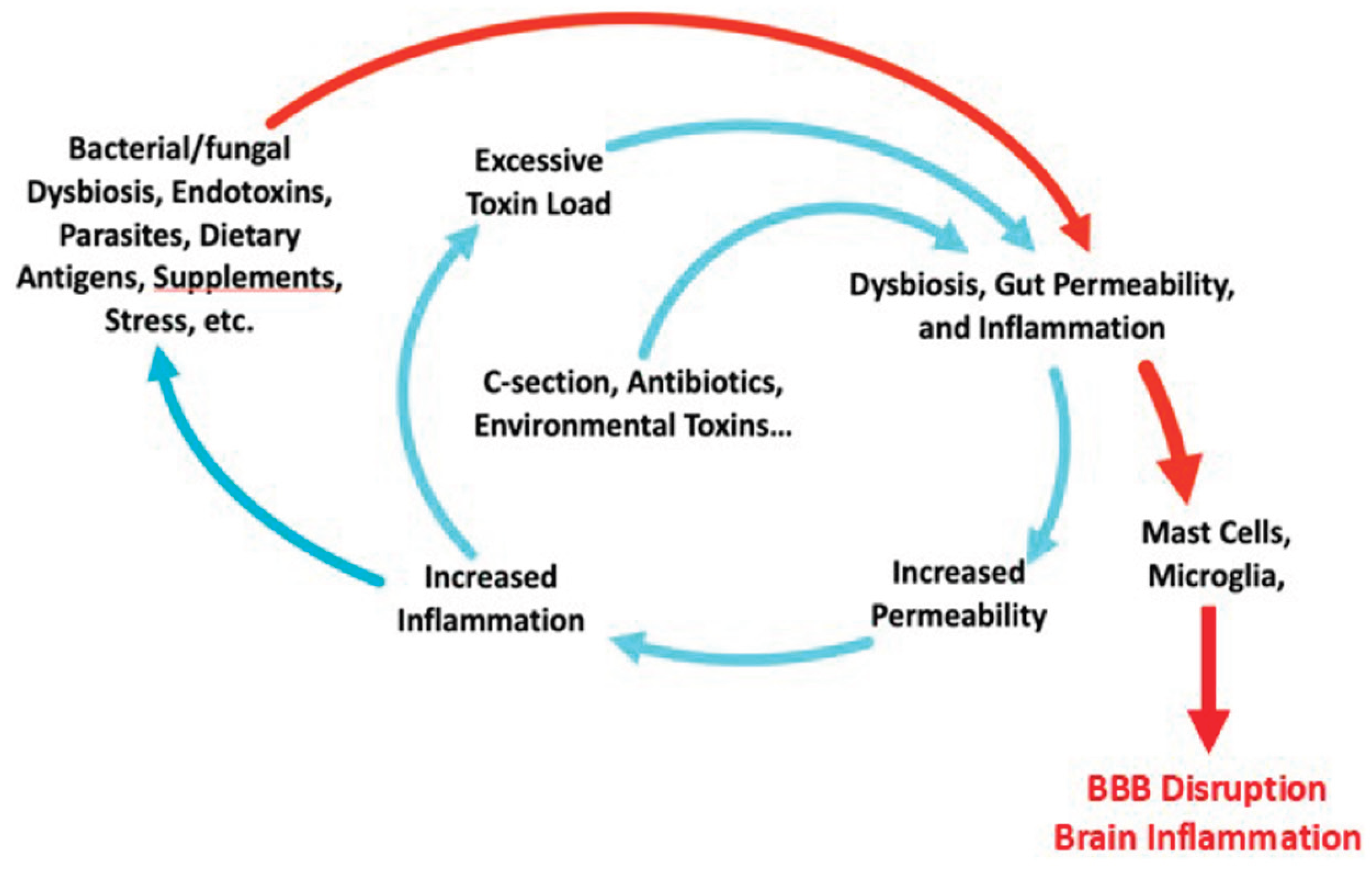
Figure 2.
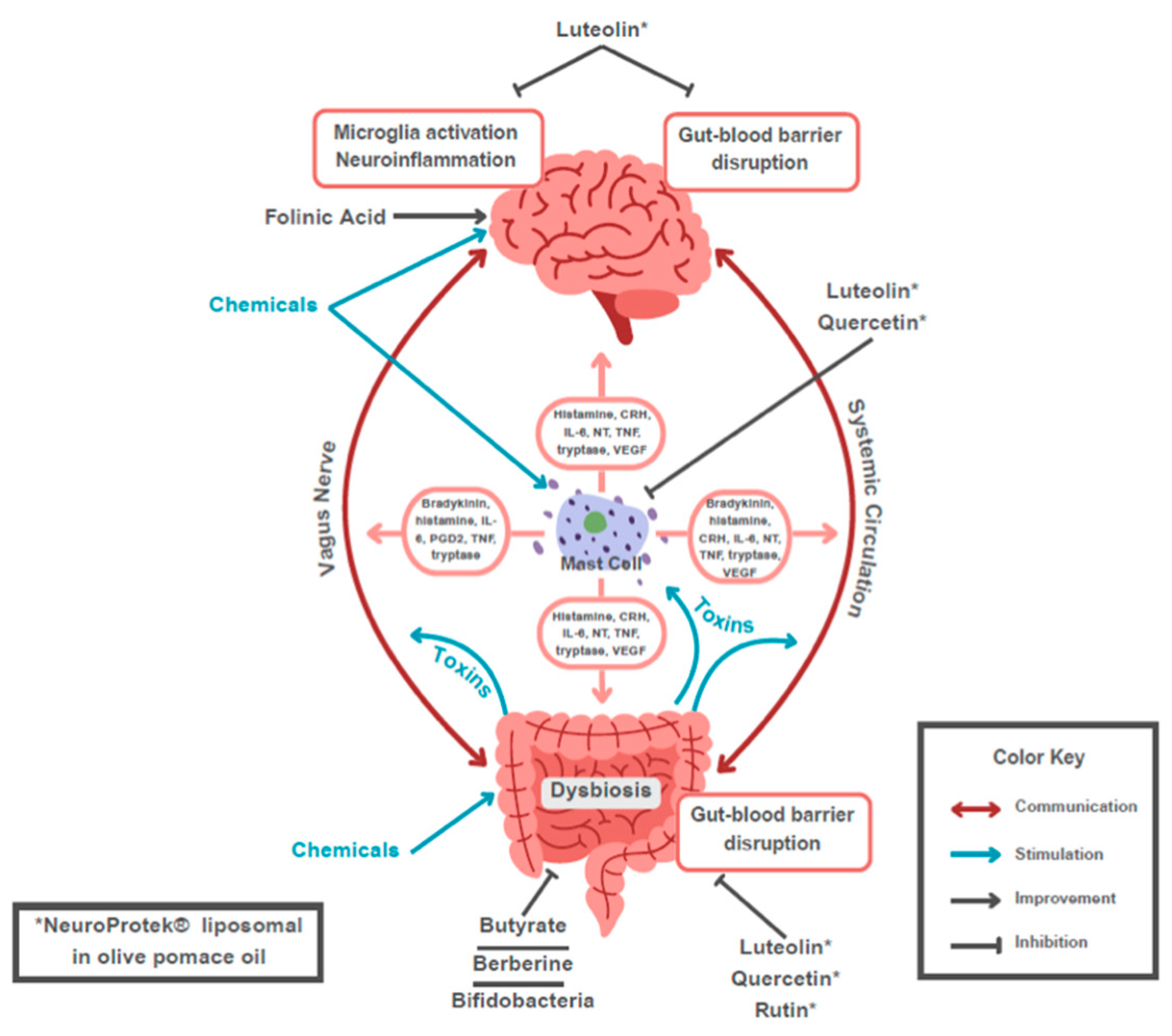
Figure 3.

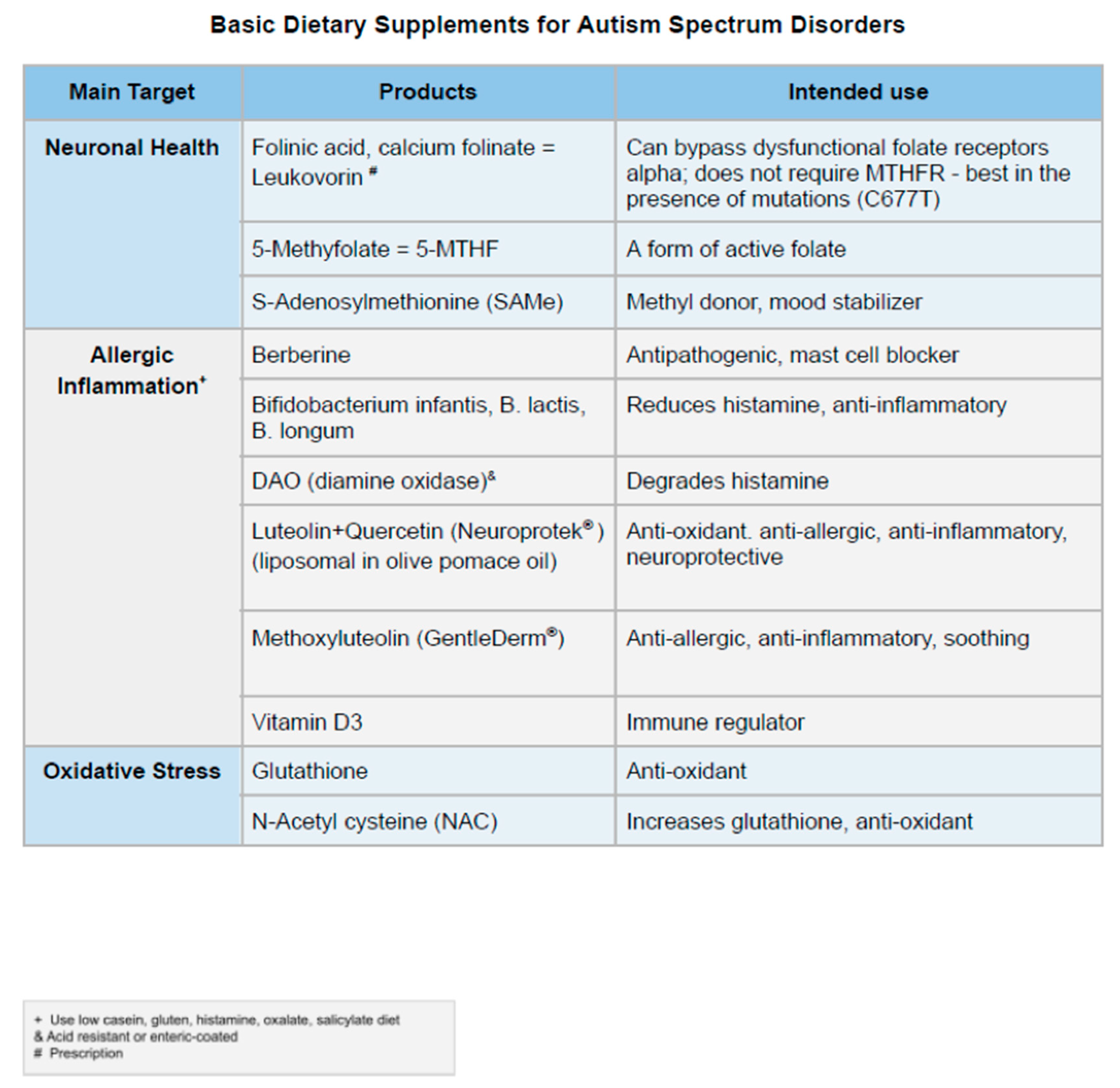
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



