Submitted:
12 June 2025
Posted:
13 June 2025
You are already at the latest version
Abstract
Geminiviruses infect many crops and weeds globally, especially in the tropical and subtropical regions, where there are waves of epidemics mainly of the begomovirus group of these viruses. These begomovirus epidemics are frequently associated with three DNA satellites, betasatellites, alphasatellites, and deltasatellites. Except for the origin of replication, these satellites show no sequence identity with the helper begomovirus. Alphasatellites and betasatellites encode -Rep and βC1 proteins, respectively, while deltasatellites encode no proteins. The -Rep, which functions like the Rep of the helper begomoviruses, ensures alphasatellite replication autonomy while betasatellites and deltasatellites depend wholly on the helper virus for replication. The betasatellite βC1 protein is a pathogenicity determinant and suppressor of RNA silencing. The associations between satellites and helper viruses vary, depending on the virus and the host, and the roles of these satellites on disease development are an active area of investigation. This review highlights current information on the role of DNA satellites on begomovirus diseases and examines commonalities and differences between, and within these satellites under prevailing conditions. Furthermore, two episomes, SEGS-1 and SEGS-2, associated with cassava mosaic geminiviruses, and their possible status as DNA satellites are discussed. Taken together, DNA satellites are a major factor in begomovirus infections, which are a major constraint to crop production especially in tropical and subtropical regions. Thus, areas for future research efforts, as well as implications in the biotechnological management of these viruses, are discussed in this review.
Keywords:
n/a
; Alphasatellites
; Betasatellites
; Deltasatellites
; Begomoviruses
1. Introduction
Geminiviruses (family Geminiviridae) are the largest known group of plant viruses and are of global economic concern as they cause devastating diseases in many important crops. The family Geminiviridae is comprised of fourteen genera—Begomovirus, Capulavirus, Citlodavirus, Curtovirus, Grablovirus, Maldovirus, Mastrevirus, Mulcrilevirus, Opunvirus, Topilevirus, Topocuvirus, Turncurtovirus, Becurtovirus, Eragrovirus —based on genome organization, host range, and insect vector [1]. Geminiviruses have circular, single-stranded DNA (ssDNA) genomes, and except for the genus Begomovirus that has some members with two component (bipartite) genomes (DNA-A and DNA-B), have single component (monopartite) genomes. Viral DNA replication follows a combination of rolling circle and recombination-dependent modes of replication, which produce double-stranded (ds) DNA replication form intermediates [2].
Gene transcription in geminiviruses occurs bidirectionally from the dsDNA template from divergent promoters located in a DNA region containing cis-acting signals required for gene transcription and DNA replication [3]. DNA-A components of bipartite begomoviruses contain canonical genes, two sense genes—AV1, which encodes the coat protein (CP), and AV2. The AV2 ORF is found in Old World bipartite begomoviruses, but not in New World viruses. Four genes are encoded in the complementary sense strand: AC1, whose gene product is the replication associated protein Rep), AC2 and AC3, which code for the transcription activation protein (TrAP) and the replication enhancer protein (REn), and AC4 that is located within AC1 at the 5’ end but in a different reading frame [4] (Figure 1). DNA-B has two genes, BV1 and BC1, which code for the movement protein (MP) and nuclear shuttle protein (NSP), respectively. DNA-A and DNA-B show no sequence similarities, except for an approximately 200 nucleotide segment of the 5′ intergenic region (IR), designated the common region (CR). The CR contains an origin of replication organized modularly, including a stem–loop structure containing the invariant nonanucleotide TAATATTAC or TAAGATTCC sequence, where cleavage, synthesis and joining of the DNA virion strand occur during replication [5,6]. All monopartite geminivirus proteins are encoded in one genomic component. The ORFs include the virion-sense V1 (CP) and V2 (MP) genes, and the complementary-sense C1 (Rep), C2 (TrAP), C3 (REn) and C4 (C4) genes. Analogous to bipartite geminiviruses, the IR harbors sequence elements found in the CR of bipartite begomoviruses, including the invariant nonanucleotide TAATATTAC sequence that harbors the origin of replication (Figure 1b).
Although RNA satellites have long been found in association with RNA viruses [7,8], the first report of a DNA satellite was in 1997 when a DNA satellite was found in northern Australia in association with tomato leaf curl virus (ToLCV), a monopartite begomovirus [9]. Since then, many begomoviruses have been found in association with three classes of satellite molecules, namely betasatellites, alphasatellites, and deltasatellites. Like the helper virus, these satellites have a circular ssDNA genome and replicate using a combination of rolling circle and recombination-dependent replication (RDR) from the dsDNA replication intermediate. RDR mode of geminivirus replication is similar to the replication mechanism of the bacteriophage φ29. Thus, the rolling circle amplification (RCA) approach, which is based on the φ29 DNA polymerase was developed and has been used extensively to identify previously undetected viruses and DNA satellites. The key advantage of this detection method is that amplification does not require previous knowledge of nucleotide sequences [10,11]. The increase in the number of DNA satellites associated with begomoviruses is therefore likely due mainly to the availability of RCA than to an emergence of these molecules.
1.1. Structure and Function of Begomovirus Satellite Molecules
Alphasatellites and betasatellites are half the size (~1.3 kb) of their helper begomovirus genome (for monopartite begomoviruses) or genome component (for bipartite begomoviruses). Like the helper viruses, these DNA satellites have circular, ssDNA genomes and contain a putative origin of replication marked by a canonical nonanucleotide motif (TAA/GTATTAC) in the loop region of the stem-loop structure. Both DNA satellites encode at least one protein. In contrast, deltasatellites, which are about ~700 nucleotides in size, encode no proteins. Specific structural features of each DNA satellite detailed below.
1.2. Betasatellites (family Tolecusatellitidae)
Betasatellites encode in the complementary strand, a 13.5k Da protein, known as βC1. βC1 proteins from several betasatellites are associated with increased levels of viral DNA and pronounced viral disease symptoms, due to suppression of host antiviral RNA silencing properties [12,13,14,15,16,17]. Some betasatellites have been found to encode a second protein in the virus sense of the molecule designated βVl, which is also a pathogenicity determinant [18,19]. Betasatellite genomes also contain a conserved sequence known as satellite conserved region (SCR) with an approximate size of ~125 nt (Figure 2A), and a 160- to 280-nt adenine-rich (A-rich) (or AT-rich) region. A search of inverted repeats with potential to form stem–loop structures with a stem of at least 6 nucleotides and a loop of at least 4 nucleotides identified only one such repeat, which harbors the TAATATTAC nonanucleotide sequence containing the origin of replication [20]. That the betasatellites have the same origin of virion-strand replication as the helper viruses is consistent with the fact that unlike alphasatellites, they depend wholly on the helper virus for replication. We must note that although the SCR was suggested to be the site where the helper begomovirus Rep binds to initiate replication [21,22,23], more recent studies suggest that Rep trans-replication of betasatellites is mediated by sequences between the A-rich region and the SCR [24]. It should be noted that unlike some Old World monopartite begomoviruses, begomoviruses found in the New World, where betasatellites are absent [25,26,27], have highly specific Rep binding sites in DNA-A and DNA-B [20].
A biological function has not yet been assigned for the betasatellite A-rich region. As discussed below, the alphasatellite A-rich region was suggested to have been used to increase the size of nanovirus DNA component from which alphasatellites are believed to have evolved [21,28]. Thus, by analogy, the betasatellite A-rich region might have the same origins and role.
1.3. Alphasatellites (family Alphasatellitidae)
Alphasatellites are of similar size to betasatellites, approximately 1.3 kb. The alphasatellites encode the ∼37kDa α-Rep, which functions like the geminivirus Rep protein and ensures autonomous replication [26]. Similar to betasatellites, alphasatellites have an A-rich region and a predicted hairpin structure (Figure 2B). A-rich sequences are thought to increase sizes of alphasatellite molecules to half the size of the begomovirus components [29], and might have been acquired from a nanoviruses [30,31]. A similar search for potential stem–loop structures as described for betasatellites identified a stem–loop structure harboring the nonanucleotide TAGTATTAC common to nanoviruses, and analogous to the TAATATTAC nonanucleotide sequence in helper virus stem–loop (Figure 2B). The slight difference between the nonanucleotide loop sequences of alphasatellites and helper geminiviruses can, at least partly, be explained by the fact that alphasatellites self-replicate using α-Rep that is more specific for the alphasatelilite loop sequence. Alphasatellites have at least three other predicted stem–loop structures (Figure 2B), the biological relevance of which will need to be examined.
1.4. Deltasatellites (family Tolecusatellitidae)
The least studied of the three begomovirus DNA satellites identified so far are deltasatellites, which unlike betasatellites and alphasatellites, do not code for a protein and the molecules are half the size of betasatellites and alphasatellites. There are three groups of deltasatellites, deltasatellites associated with the monopartite ToLCV in Australia [9], those associated with sweet potato viruses in Spain and Venezuela [28,32], and deltasatellites associated with New World bipartite begomoviruses, including sida golden yellow vein virus (SiGYVV) [28,33,34]. Structurally, deltasatellites associated with ToLCV are similar to the one associated with the New World bipartite begomovirus, SiGYVV, and both are characterized by the SCR, conserved stem–loop, A-rich region (Figure 2C). The SCR is absent in SPLCV-SPLCD1, the deltasatellite associated with sweet potato leaf curl virus (SPLCV). Half of the SPLCV-SPLCD1 genome has sequence identity to a truncated C1 and IR of SPLCV and was likely acquired from a recombination event (Figure 2C).
Structurally, stem–loop structures are found to be the most common features in these satellite molecules. In addition to harboring the origin of replication, stem–loop structures provide specificity during virus genome assembly and packaging. In ssRNA viruses,for example, packaging proteins specifically recognize and bind to the stem–loop with high affinity, followed by encapsidation of only its genetic material [35,36,37,38,39,40].
2. Role of DNA Satellites on Virus Disease Development
Begomoviruses cause important crop diseases and are a threat to global agriculture. Symptoms displayed by plants infected by these viruses include leaf curling, distortion, yellowing, mosaic patterns, stunting, and vein swelling; this may lead to substantial yield losses. Begomoviruses are geographically separated into New World and Old World viruses. Most New World begomoviruses have a bipartite genome, while Old World viruses tend to have monopartite genomes, which are frequently associated with DNA satellites. Alphasatellites are found in the Old World and in the New World. In the Old World they are associated with monopartite begomoviruses, and in many cases coinfect the same plant with betasatellites, while in the New World, they are associated with bipartite begomoviruses [27]. Like alphasatellites, deltasatellites are associated with begomoviruses in the Old World and New World, where betasatellites are absent.
2.1. Interaction between betasatellites and associated helper begomoviruses is synergistic
Begomovirus-betasatellite complexes have been reported in several crops, including cotton (Gossypium hirsutum), peppers (Capsicum spp.), okra (Abelmoschus esculentus), radish (Raphanus sativus), papaya (Carica papaya L.) and tomato (Solanum lycopersicum). In the Old World betasatellites have been shown to act synergistically in mixed infections with helper viruses, particularly in Malvaceae and Solanaceae hosts, but interestingly not in some Cucurbitaceae hosts. In Pakistan and India, synergistic interactions were reported between radish leaf curl virus (RaLCV) and its betasatellite, RaLCB, in infected radish plants [41,42]. Furthermore, studies in China reported a synergistic interaction between tomato yellow leaf curl China Virus (TYLCCNV) and tomato yellow leaf curl China betasatellite (TYLCCNB) in tomato [43]. Also, pepper chili leaf curl betasatellite (ChLCB) and pepper leaf curl Lahore virus (PepLCLV), which likely resulted from a recombination event between Papaya leaf curl virus (PaLCuV) and chili leaf curl virus (ChiLCV), caused more severe symptoms in pepper plants compared with single infection by PepLCLV [44]. The study further showed that ChiLCV and ChLCB do coinfect potato (Solanum tuberosum) in nature, thus indicating a broad host range for these betasatellites.
Although the whitefly is the vector of begomoviruses and associated DNA satellites, and therefore and important component in begomovirus disease epidemics, most studies on the incidence, distribution, and diversity of these viruses and DNA satellites have disproportionately been on plant hosts. In one of the few studies carried out so far, four cryptic whitefly species were sampled in pumpkin, cucumber, tomato, sponge gourd and ridge gourd fields in India. Analysis of these samples identified four types of betasatellites, tomato leaf curl Bundi betasatellite (ToLCBDB), okra enation leaf curl betasatellite (OLCuB), Bhendi yellow vein mosaic betasatellite (BYVB), and papaya leaf curl virus betasatellite (PaLCB), as well as the alphasatellites ageratum enation alphasatellite (AEV) and cotton leaf curl Multan alphasatellite (CLCuMuA) [45]. This stresses the importance of monitoring the incidence, distribution and diversity of these viruses and associated satellites in whitefly vectors.
Much of sub-Saharan Africa, especially West Africa, has a rich vegetation due to uniformly high temperatures and heavy rainfall year-round. This provides a rich environment for plant viruses, which infect both cultivated crops and weeds. Unfortunately, until recently, there has been a dearth of information on the occurrence of begomoviruses and associated DNA satellites in much of this region. A study carried out in the tropical rainforest part of Cameroon identified cotton leaf curl Gezira virus (CLCuGeV) and okra yellow crinkle virus (OYCrV), respectively in coinfection of okra cotton leaf curl Gezira betasatellite (CLCuGeB) [46]. This study followed a previous study by the same group that reported coinfection of Ageratum conyzoides (Family Asteraceae) by a previously unreported ageratum leaf curl Cameroon virus (ALCCMV) and its associated ageratum leaf curl Cameroon betasatellite (ALCCMB) [47]. A. conyzoides is an invasive weed that grows extensively in much of West Africa and therefore likely acts as a reservoir of these viruses and their DNA satellites. Also in the rainforest zone of West Africa, a previously unreported betasatellite associated with ToLCD, designated tomato leaf curl Togo betasatellite (ToLCTGB) was found to associate with tomato leaf curl Kumasi virus (ToLCKuV) and tomato leaf curl Nigeria virus (ToLCNGV) [48]. Similar to the previously reported tobacco curly shoot disease in China [49], ToLCTGB was found not to enhance ToLCD symptoms in tomato. In contrast, it caused an increase of symptoms of both viruses in Nicotiana spp. and Datura stramonium [48]. The same group found in Mali, in the Sudano-Sahelian climate zone, that tomato yellow leaf curl Mali virus (TYLCMLV), a recombinant virus, and CLCuGB caused severe symptoms in tomato and several other solanaceous hosts, but not in common bean [50]. Another study in Burkina Faso, which like Mali, is in the Sudano-Sahelian zone, reported coinfection of okra by cotton leaf curl Gezira betasatellite (CLCuGB) and its helper virus, CLCuGeV [51]. Furthermore, a study carried in Uganda, in eastern Africa, detected vernonia crinkle virus (VeCrV) and its associated and previously unidentified vernonia crinkle betasatellite (VeCrB) in african bitter leaf (Vernonia amygdalina), a non-solanaceous vegetable that grows in tropical Africa [52].
2.1.1. The βC1 RNA silencing suppression function contributes to betasatellite enhancement of helper virus symptoms
βC1 suppression of antiviral RNA silencing deployed by the host is partly due to its interaction with several proteins involved in the RNA silencing pathway, including S-adenosyl homocysteine hydrolase (SAHH) [43], SnRK1 (sucrose non-fermenting1-related protein kinase 1), Calmodulin-like protein (rgs-CaM) [53], as well as Argonaute 1 Argonaute 1 (AGO1) [54]. These interactions are suggested to inhibit methylation, antiviral phosphorylation, formation of the RNA-induced silencing complex (RISC), and thereby suppressing RNA silencing. This action stabilizes the begomovirus-betasatellite complexes and increases viral DNA levels, as well as symptom severity.
The ability of βC1 to antagonize host defenses is not limited to suppression of RNA silencing, in fact, it was found to be involved in virus movement. Specifically, it was shown that βC1 interacts with the CP of bhendi yellow vein mosaic virus (BYVMV), the helper virus, as well as with the host karyopherin α, a nuclear importin like protein, to facilitate shuttling between the nucleus and the cytoplasm [55]. Furthermore, βC1 forms a complex with Asymmetric leaves 1 (AS1) , a gene that facilitates the specification of leaf adaxial identity [56], and attenuates expression of selective jasmonic acid (JA)-responsive gene, leading to suppression of JA host defense responses [17]. Additionally, βC1 of CLCuMuB associated with cotton leaf curl disease, interacts with the ubiquitin-conjugating enzyme (UBC), thereby interfering in the ubiquitin proteasome pathway, which results in the development of leaf curling symptoms [57]. Sequence analysis shows that except for a few conserved residues, including the PFDFN motif found in the middle of the protein, βC1 exhibits considerable sequence variation, especially at the N- and C-termini of the proteins (Figure 3A). Taken together, these studies show the important role played by βC1 in betasatellite induction of severe symptoms during coinfection with helper viruses.
2.2. Alphasatellites tend to attenuate symptoms in coinfections with helper viruses.
Alphasatellites have been found not to cause an increase in disease symptoms or in helper viral DNA accumulation. In fact, many alphasatellites tend to modulate virulence and symptom development. For example, a cassava mosaic virus associated alphasatellite, closely related (80% sequence identity) to cotton leaf curl Gezira (CLCuGeA) was found not to influence cassava mosaic disease (CMD) symptoms in coinfected cassava in Madagascar [58]. The same is the interaction between CLCuGeA and okra leaf curl Burkina Faso alphasatellite (OLCuBFA) and a complex of CLCuGeV and OYCrV in okra plants in Cameroon [46]. Furthermore, two alphasatellites associated with the cotton leaf curl Multan disease in India were reported to attenuate and delay symptom development in N. benthamiana [59].
In the Americas, melon chlorotic mosaic virus-associated alphasatellite (MeCMA) was detected in watermelon samples collected from Venezuela in association with its help virus, melon chlorotic mosaic virus (MeCMV), a bipartite begomovirus [60]. Subsequent studies identified MeCMA in association with several bipartite begomoviruses infecting non-cultivated plants and watermelon crops [61,62]. In an in-depth survey of begomovirus-associated satellite DNAs carried out in the Old World and New World (USA, Guatemala, Israel, Puerto Rico, and Spain), whiteflies were collected in weeds, as well as in bean, eggplant, pumpkin, squash, and tomato fields, then analyzed using vector-enabled metagenomic (VEM) and RCA detection [63]. Results of the study confirmed the limited geographic distribution of alphasatellites in the New World compared with the Old World with only two new species detected in whiteflies collected from a single tomato field in Guatemala and Puerto Rico.
2.2.1. The ability of alphasatellites to attenuate begomovirus disease symptoms may be due to α
-Rep interaction with the Rep of the parent virus
The ability of two alphasatellites associated with cotton leaf curl Rajasthan virus (CLCuRaV) to attenuate helper virus levels and symptoms was attributed to interaction between the α-Rep protein and CLCuRaV Rep, which inhibited helper virus proper replication [26]. Additionally, asystasia yellow mosaic alphasatellite (AYMA) has been observed to cause yellowing symptoms in Asystasia gangetica, a weed common in western and central Africa, during mixed infection with West African Asystasia virus (WAAV) but with no apparent reduction in leaf size [64] (Figure 4). However, the yellowing symptoms are observed to be more intense and systemic in plants containing AYMA, compared with plants singly infected by WAAV. Interestingly, a survey in China showed that plants infected by alphasatellites tended to also contain betasatellites, yet only a few betasatellite-infected plants contained alphasatellites [65]. A case can be made that alphasatellite modulation of virus levels and disease severity ensures survival of the host, which ultimately helps the virus successfully complete its infection cycle. It is also possible that alphasatellites are molecular parasites that reduce levels of the helper begomovirus, thereby lessening virus disease symptoms.
It must be stated that there have been reports of some Old World alphasatellites encoded α-Rep increasing accumulation of the helper virus DNA by suppressing RNA silencing. This is the case with α-Reps of tomato yellow leaf curl China virus (TYLCCNA), the alphasatellites associated with TYLCCNV [66]. Similar results were recorded for the α-Rep of cotton leaf curl Multan alphasatellites (CLCuMuA) associated with CLCuMuV [67,68]. Strikingly, α-Reps of gossypium darwinii symptomless alphasatellite (GDarSLA) and gossypium mustelinium symptomless alphasatellite (GMusSLA) coinfecting Gossypium spp. with cotton leaf curl Rajasthan virus (CLCuRaV) were found to be suppressors of RNA silencing, but not βC1 of CLCuMuB, or viral TrAP, C4, and MP, which are known suppressors of RNA silencing [68]. Unsurprisingly, the α-Reps of TYLCCNA, GDarSLA and GMusSLA so far shown to suppress RNA silencing, group together in a phylogenetic tree (Figure 5). It is important to note that the New World euphorbia yellow mosaic alphabetasatellite (EuYMA) encoded α-Rep was found not to suppress RNA silencing [69]. Yet, EuYMA, as well as tomato severe rugose alphasatellite (ToSRA) were reported to differentially interact with EuYMV and ToYSV in different hosts. Thus, EuYMA increased symptoms of EuYMV and ToYSV in tomato and N. benthamiana, while ToYSA caused more severe symptoms of ToYSV in N. benthamiana and L. sibiricus but negatively affected the accumulation of ToYSV in both hosts [69,70]. This stresses the role of helper virus and host factors in these interactions.
Alignment of these α-Reps showed substantial sequence variation between New World and Old World α-Reps, especially at the N- and C-termini (Figure 3B), which likely explains the dichotomy in these biological functions. Future studies will elucidate the mechanism by which these α-Reps suppress RNA silencing and the basis of differential behavior between New World and Old World α-Reps. Furthermore, it is not yet clear whether α-Rep can trans-replicate the helper virus in the absence of the encoded Rep.
2.3. Deltasatellites do not enhance helper virus DNA levels or symptom severity
The first identified deltasatellite was found to be associated with ToLCV, designated ToLCV-sat, was reported not to influence helper virus accumulation [9]. Replication of this deltasatellite was supported by several monopartite and bipartite begomoviruses, including tomato yellow leaf curl virus (TYLCV), African cassava mosaic virus (ACMV), and the curtovirus beet curly top virus (BCTV) [71]. Like Old World deltasatellites, New World deltasatellites do not affect the symptoms induced by the helper begomoviruses, and tend to reduce virus accumulation [33]. In fact, viral DNA levels and symptom severity of SPLCV were reported to be lower in the presence of SPLCV- SPLCD1, its associated deltasatellite [72]. However, like for alphasatellites, there are differential effects of deltasatellites on the helper viruses. Indeed, recent research indicates that deltasatellites influence the replication dynamics of their helper begomoviruses, either by enhancing or by reducing their accumulation [32].
2.4. DNA satellites complementation of begomoviruses from different regions may lead to new disease epidemics
Since begomovirus acquisition or exchange of satellite DNAs may lead to adaptation to new plant hosts, it is important to investigate the diversity and distribution of these molecules. New DNA satellites continue to be identified in association with begomoviruses in different regions, especially in the Indian subcontinent and in Africa. It is clear that virus and satellite coevolve together resulting in a somewhat stable disease dynamic in the host under prevailing environmental conditions. This dynamic may however change if new viruses and satellites are introduced from other regions. Thus, there is a need to investigate complementation between begomoviruses and non-cognate satellites. In a recent study, therefore, CLCuKoV and CLCuMuV from Pakistan were found to each interact with CLCuGeB from Cameroon synergistically in N. benthamiana, which developed more severe symptoms with higher viral DNA levels compared with plants containing the virus alone [73]. The same study showed that CLCuKoV and CLCuMuV complemented infection by ALCCMB, also from Cameroon, while OYCrV from Cameroon and TYLCV from Australia functionally interacted with CLCuMuB, ALCCMB and CLCuGeB, respectively. This complementations appear not to be limited to betasatellites, given that the study further showed that all begomoviruses from different geographical origins that were tested supported ALCCMA from Africa [73]. Although these studies were carried out on N. benthamiana, an experimental host, there is reason to believe that similar interactions occur in natural hosts. For example, another study showed that CLCuGeB, which is associated with its cognate CLCuGeV in okra, was transmitted and assisted by TYLCV in tomato plant [74]. In the absence of available data there is the likelihood of high compatibility between alphasatellites and non-cognate helper begomoviruses, given that alphasatellites exhibit autonomous replication. Taken together, the ability of functional interactions between satellite and helper virus shows the evolutionary potential of these complexes, which may disturb the established equilibrium and lead to the emergence of new begomovirus epidemics.
3. Recombination Between Helper Viruses and DNA Satellites Contributes to Virus and Satellite Evolution
Although the evolutionary relation between begomoviruses and their DNA satellites, as well as amongst the DNA satellites, is still being examined, early evidence suggests occurrence of recombination events in the evolution of these molecules [75]. This is to be expected given that helper viruses and their satellites replicate using recombination-dependent replication. Thus, it is not surprising that recombination was reported between ageratum yellow vein virus (AYVV) and it associated betasatellite (AYVB), and with the alphasatellite (AYVA) in coinfected Ageratum conyzoides plants [23,31,76]. The recent identification of a chimeric DNA molecule with a similar size to alphasatellites and betasatellites and consisting of sequences derived from CLCuKoV, CLCuMuA, and croton yellow vein mosaic alphasatellite [77] further supports ongoing recombination between helper virus and associated DNA satellites. Additionally, recombination events are reported between CLCuKoV and CLCuMuV with their respective associated satellites CLCuMuB and CLCuMuA [78,79,80]. Moreover, in Pakistan, a recombinant DNA betasatellite with documented epidemiological impact on cotton plantings was found to contain a sequence derived from CLCuMuB, as well as from tomato leaf curl betasatellite [81], causing the breakdown of resistance to CLCuKoV [57,82,83,84].
Recombination events are not limited to protein-coding alphasatellites and betasatellites. In fact, half of the genome of SPLCV-SPLCD1, the deltasatellite associated with SPLCV, is thought to have been acquired from the latter as a result of a recombination event during coinfection of Convolvulaceae and/or sweet potato [77] (Figure 2C). Together, the role of recombination in interactions between helper virus and DNA satellite is evidenced in the fact that many new begomovirus epidemics involve recombinant molecules. This is evident, particularly in the Indian sub-continent, where there is ccurrently more documentation of begomovirus-DNA satellite complexes [44,50,57,75,77,78,79,80,82,83].
4. Cassava Geminivirus Associated SEGS-1 and SEGS-2 Episomes are Novel DNA Satellites
Two episomes designated as SEGS-1, and SEGS-2 “sequences enhancing geminivirus symptoms“, isolated in cassava plants exhibiting unusually severe CMD symptoms are reported in cassava fields in the coastal region of Tanzania and the tropical forest region of Cameroon [85,86]. One of the few more comprehensive surveys carried out in West Africa is from Ghana, where out of 110 cassava leaf samples affected by cassava mosaic begomoviruses, SEGS-2 and SEGS-1 were detected in 66 (60%) and 47 (43%), respectively [87]. It is yet unclear whether this high incidence is representative of the region. The two episomes (Figure 6) show no sequence identity to each other, nor to the associated cassava geminiviruses, or to the three DNA satellites discussed in this review. Interestingly, SEGS-1 shows 98% sequence identity to a satellite DNA molecule isolated in mentha (Mentha spicata) plants infected by tomato leaf curl Karnataka virus-Bangalore (ToLCKV-Ban) in Punjab, India [88]. This molecule was designated mentha leaf deformity associated DNA-II (MLDA-DNA-II) because of the deformation it caused in association with ToLCKV-Ban in coinfected mentha plants. Thus, SEGS-1, which like SEGS-2 induces deformed cassava leaves and enhanced CMD symptoms in cassava (Manihot esculenta Crantz) in coinfection with cassava mosaic geminiviruses (CMGs) [86], causes similar symptoms in mint plants in India.
Interestingly, sequences related to SEGS-1 and SEGS-2 occur in the cassava genome. A third of the SEGS-1 is 99% identical to the a segment of the eukaryotic ORF encoding the protein prefoldin subunit 6 (PFDN6), which is a subunit of the heteromeric prefoldin complex involved in the proper folding of cytoskeletal proteins [89,90]. Correspondingly, over a third of SEGS-2 shows 96% sequence identity with the cassava pentatricopeptide repeat (PPR) family of proteins, which regulate gene expression in chloroplasts and mitochondria, and are involved in many aspects of plant growth and development, including responses to stress and reproduction [91]. Additionally, the 3’ half of PPR in SEGS-2 contains a start codon of a potential 68-amino acid peptide. Curiously, this sequence maps to the first 68 amino acids of pre-mRNA-splicing factor ATP-dependent RNA helicase DEAH1 isoform X2 (DHX16 isoform X2). This DHX16 isoform X2 is part of the spliceosome complex involved in the processing of pre-mRNA into mRNA [92]. Unsurprisingly, DHX16 isoform X2 and PPR are found in the same region of the cassava genome (loci XM_043949572 and XM_043949652) and their ORFs overlap but are in different frames. Alignment of SEGS-2 and these two sequences shows the specific DNA fragment that became part of the SEGS-2 molecule (Figure 7). Furthermore, another segment of the SEGS-2 genome shows 83% sequence identity with a noncoding region of the cassava genome but outside the DHX16 isoform X2 and PPR loci. This adds complexity regarding the origins and evolutionary path of SEGS-2.
Other structural features of these episomes include a GC-rich region that covers a third of each molecule, with CpG/GpC ratios of 0.84, and therefore qualifying as CpG islands [93]. These CpG islands are positionally located in the recombinant PFDN6 and PPR regions, respectively (Figure 6). CpG islands are substrates for DNA methylation, especially in promoter regions, leading to gene silencing [94,95,96,97]. What role a heavily methylated region plays in the biology of these episomes is yet to be elucidated. It has been established that SEGS-1 episomes are only found in CMG-infected plants, consistent with infection promoting episome formation [98]. Given that begomovirus infection induces the host cell cycle S phase and DNA replication [99,100] and interferes with DNA methylation [101], it is plausible to think that the cell replicative state and/or the decrease in DNA methylation could lead to activation and generation of SEGS-1 episomes [98]. This supports the view that unlike SEGS-2, SEGS-1 is not a DNA satellite. Furthermore, SEGS-1 is ubiquitous in the cassava genome, thus, it was suggested to be perceived as a member of a repetitive sequence family that is inactivated by DNA methylation and/or sequestration into heterochromatin [102].
Both episomes display characteristic stem–loop structures; with a confidence level of >90%, this feature is real and likely has an important biological role, and were suggested to harbor the replication origin of the episomes [86]. Future investigations will determine the possibility that these hairpins are packaging signals for SEGS-2, which has been shown to be packaged by the helper CMGs [86].
Epidemiologically, SEGS-1 and SEGS-2 enhance CMG replication and symptom severity, and were reported to cause a break in resistance to CMD in cassava in the coastal region of Tanzania [85,86,102]. Correspondingly, in Cameroon, elite CMD-resistant cassava breeding lines containing the polygenic recessive resistance locus and a dominant monogenic resistance locus, display severe symptoms when coinfected by these episomes and ACMV. In more severe cases, the leaf is reduced to the midrib with a total absence of the leaf blade (Figure 8). These episomes were also found to cause ACMV infection and enhancement of symptoms when applied exogenously to Arabidopsis leaves, as well as in Arabidopsis containing an integrated copy of the episome [85,102]. The SEGS-2 encoded putative 68-amino acid protein that corresponds to DHX16 isoform X2 plant protein likely plays a role in severe CMD symptoms observed in coinfected plants [85]. Because SEGS-2 is packaged into virions and transmitted by whiteflies along the helper CMG [85,86], this episome qualifies as a DNA satellite.
A recent in depth study has shown that unlike SEGS-2, SEGS-1 activity in Arabidopsis is not mediated by an episome, but rather by the transgene and therefore chromosomal [102]. This is supported by the fact that SEGS-1 molecules have only been found in plants and not in virions or whiteflies, suggesting that they may originate from the cassava genome. Thus SEGS-1 is likely not a DNA satellite. This is supported by the fact that in the absence of exogenous SEGS-1, infected plants of some cassava cultivars, but not all, form SEGS-1 episomes during CMD [102].
5. Outlook
To contain the global begomovirus epidemics, considerable efforts are required to identify all components underpinning the epidemics; this will enable us to design appropriate control strategies. Here, some immediate research efforts to understand the epidemics, as well as use of prime editing to generate potentially non-GMO virus-resistant plants are discussed.
5.1. Knowledge gaps that need addressing on the begomovirus diseases
Future research efforts will need to determine whether DNA satellites influence efficiency of whitefly transmission of the helper virus. Also, an in-depth comparison between viruliferous whitefly and virus-free whitefly transmission efficiency will provide additional information on the development begomovirus epidemics. Moreover, it is not clear whether higher levels of virus in infected hosts necessarily translates to higher transmission efficiency. The role of Rep binding in betasatellites in the interaction between the latter and helper begomoviruses needs to be elucidated, given that betasatellites lack the iteron sequences of their helper begomoviruses. Thus, different Reps will bind with different efficiencies, thereby influencing effectivity. For example, ToLCKuV from Togo and ToLCNGV, both of which coinfect different host species in association with ToLCTGB causing different symptoms have different iterons, which likely influences ToLCTGB binding, and thus infectivity [48]. Therefore, investigating the role of the characteristic high affinity of Rep binding sites in New World begomoviruses, as opposed to betasatellite binding sites, would address a possible role of Rep binding sites on the absence of betasatellites in the New World. Furthermore, additional investigation would determine whether α-Reps can efficiently trans-replicate the parent virus, given that such a function would add to our understanding of virus infection.
Furthermore, tomato leaf curl New Delhi virus (ToLCNDV), causal agent of ToLCD, and begomoviruses responsible for TYLCD) are extending host range and spreading to new geographical regions, in the Middle East and the western Mediterranean Basin [103,104,105]. The role of DNA satellites in this expantion will need to be determined in future research efforts. Correspondingly, more comprehensive studies on the diversity, distribution, and impact of DNA satellites are needed in West Africa where continuing studies are documenting occurrence of new begomoviruses [46,47,48,51,64,87,106,107,108].
Also to be addressed further is the status of SEGS-1. Although it was recently shown not to be transmitted by the whitefly, and therefore does not qualify as a DNA satellite [98], this episome will need to be monitored in the future to see whether it becomes a DNA satellite. This would provide us important information on the origins and evolution of these DNA satellites.
5.2. DNA satellites and biotechnological management of begomovirus diseases
RNA satellites that attenuate symptoms of the helper RNA viruses have been shown to modulate symptoms and/or levels of the viral RNA, including RNA satellites associated with cucumber mosaic virus [109,110,111], tobacco ringspot virus [112], and bamboo mosaic virus [113]. Reduced virus levels are attributed mainly to competition between satellite RNA and viral genomic RNA for replication [110,114,115]. Indeed, bamboo mosaic virus RNA satellites have been used to generate virus resistant plants [113]. This approach has not yet been used in geminivirus control. The fact that alphasatellites and deltasatellites modulate helper virus symptoms suggests a natural defense mechanism and therefore can be harnessed to produce begomovirus-resistant plants. With recent advances in biotechnological strategies in plant virus management [116,117,118,119], the use of satellite DNA molecules in controlling geminiviruses needs to be explored. For example, prime editing (PE), the novel CRISPR-based genome editing method, can be used for targeted and precise modification of the genome [120]. A potential impactful strategy is to explore the phased secondary small interfering RNAs (phasiRNA) pathway. We and others have reported datasets of small RNAs of several crops impacted by begomoviruses and associated satellites [121,122,123,124,125]. These studies include phasiRNAs and their PHAS loci from which 21-24-nt phasiRNA are processed under natural conditions. This provides an excellent opportunity to employ PE to precisely insert 21-24-mers deigned from virus and/or satellite to generate virus-resistant plants [119].
Acknowledgments
This research was funded by NIFA Award #2020-67018-31180. I thank Linda Hanley-Bowdoin (North Carolina State University) and Roisin McGarry (University of North Texas) for critical reading of the manuscript. I equally thank Fidele Tiendrebeogo for providing the photo of Asystasia yellow mosaic alphasatellite coinfection of Asystasia gangetica with West African asystasia virus.
References
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A. ICTV Virus Taxonomy Profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Stenger, D.C.; Revington, G.N.; Stevenson, M.C.; Bisaro, D.M. Replicational Release of Geminivirus Genomes from Tandemly Repeated Copies: Evidence for Rolling-Circle Replication of a Plant Viral DNA. Proc. Natl. Acad. Sci. 1991, 88, 8029–8033. [Google Scholar] [CrossRef]
- Orozco, B.M.; Hanley-Bowdoin, L. A DNA Structure Is Required for Geminivirus Replication Origin Function. J. Virol. 1996, 70, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Fondong, V.N. Geminivirus Protein Structure and Function. Mol. Plant Pathol. 2013, 14, 635–649. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Lett, J.-M.; Martin, D.P.; Roumagnac, P.; Varsani, A.; Zerbini, F.M.; Navas-Castillo, J. ICTV Virus Taxonomy Profile: Geminiviridae 2021. J. Gen. Virol. 2021, 102, 001696. [Google Scholar] [CrossRef] [PubMed]
- Laufs, J.; Jupin, I.; David, C.; Schumacher, S.; Heyraud-Nitschke, F.; Gronenborn, B. Geminivirus Replication: Genetic and Biochemical Characterization of Rep Protein Function, a Review. Biochimie 1995, 77, 765–773. [Google Scholar] [CrossRef]
- Roossinck, M.J.; Sleat, D.; Palukaitis, P. Satellite RNAs of Plant Viruses: Structures and Biological Effects. Microbiol. Rev. 1992, 56, 265–279. [Google Scholar] [CrossRef]
- Simon, A.E. Satellite RNAs of Plant Viruses. Plant Mol. Biol. Report. 1988, 6, 240–252. [Google Scholar] [CrossRef]
- Dry, I.B.; Krake, L.R.; Rigden, J.E.; Rezaian, M.A. A Novel Subviral Agent Associated with a Geminivirus: The First Report of a DNA Satellite. Proc. Natl. Acad. Sci. 1997, 94, 7088–7093. [Google Scholar] [CrossRef]
- Haible, D.; Kober, S.; Jeske, H. Rolling Circle Amplification Revolutionizes Diagnosis and Genomics of Geminiviruses. J. Virol. Methods 2006, 135, 9–16. [Google Scholar] [CrossRef]
- Inoue-Nagata, A.K.; Albuquerque, L.C.; Rocha, W.B.; Nagata, T. A Simple Method for Cloning the Complete Begomovirus Genome Using the Bacteriophage Phi29 DNA Polymerase. J. Virol. Methods 2004, 116, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Li, G.; Wang, D.; Hu, D.; Zhou, X. A Begomovirus DNAbeta-Encoded Protein Binds DNA, Functions as a Suppressor of RNA Silencing, and Targets the Cell Nucleus. J. Virol. 2005, 79, 10764–10775. [Google Scholar] [CrossRef]
- Gnanasekaran, P.; Ponnusamy, K.; Chakraborty, S. A Geminivirus Betasatellite Encoded βC1 Protein Interacts with PsbP and Subverts PsbP-mediated Antiviral Defence in Plants. Mol. Plant Pathol. 2019, 20, 943–960. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tang, Y.; She, X.; Yu, L.; Lan, G.; Ding, S.; He, Z. Characterisation of a Betasatellite Associated With Tomato Yellow Leaf Curl Guangdong Virus and Discovery of an Unusual Modulation of Virus Infection Associated With C4 Protein. Mol. Plant Pathol. 2025, 26, e70051. [Google Scholar] [CrossRef]
- Mirzazadeh, M.; Heydarnejad, J.; Salari, A.; Massumi, H.; Esmaeili, M. Association of Tomato Leaf Curl Betasatellite (Betasatellite Solani) with the Turncurtovirus Sesame Curly Top Virus (Turncurtovirus Sesami) Increases the Severity of Symptoms in Watermelon Plants. Trop. Plant Pathol. 2025, 50, 5. [Google Scholar] [CrossRef]
- Saeed, M.; Zafar, Y.; Randles, J.W.; Rezaian, M.A. A Monopartite Begomovirus-Associated DNA Beta Satellite Substitutes for the DNA B of a Bipartite Begomovirus to Permit Systemic Infection. J. Gen. Virol. 2007, 88, 2881–2889. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Iwasaki, M.; Machida, C.; Machida, Y.; Zhou, X.; Chua, N.-H. betaC1, the Pathogenicity Factor of TYLCCNV, Interacts with AS1 to Alter Leaf Development and Suppress Selective Jasmonic Acid Responses. Genes Dev. 2008, 22, 2564–2577. [Google Scholar] [CrossRef]
- Hu, T.; Song, Y.; Wang, Y.; Zhou, X. Functional Analysis of a Novel βV1 Gene Identified in a Geminivirus Betasatellite. Sci. China Life Sci. 2020, 63, 688–696. [Google Scholar] [CrossRef]
- Gupta, N.; Reddy, K.; Gnanasekaran, P.; Zhai, Y.; Chakraborty, S.; Pappu, H.R. Functional Characterization of a New ORF βV1 Encoded by Radish Leaf Curl Betasatellite. Front. Plant Sci. 2022, 13, 972386. [Google Scholar] [CrossRef]
- Maliano, M.R.; Melgarejo, T.A.; Rojas, M.R.; Barboza, N.; Gilbertson, R.L. The Begomovirus Species Melon Chlorotic Leaf Curl Virus Is Composed of Two Highly Divergent Strains That Differ in Their Genetic and Biological Properties. Plant Dis. 2021, 105, 3162–3170. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.E.; Amin, I.; Idris, A.M.; Mansoor, S.; Bedford, I.D.; Dhawan, P.; Rishi, N.; Siwatch, S.S.; Abdel-Salam, A.M.; et al. Diversity of DNA Beta, a Satellite Molecule Associated with Some Monopartite Begomoviruses. Virology 2003, 312, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Stanley, J. Subviral Agents Associated with Plant Single-Stranded DNA Viruses. Virology 2006, 344, 198–210. [Google Scholar] [CrossRef]
- Saunders, K.; Norman, A.; Gucciardo, S.; Stanley, J. The DNA Beta Satellite Component Associated with Ageratum Yellow Vein Disease Encodes an Essential Pathogenicity Protein (betaC1). Virology 2004, 324, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Saunders, K.; Briddon, R.W.; Stanley, J. Replication Promiscuity of DNA-Beta Satellites Associated with Monopartite Begomoviruses; Deletion Mutagenesis of the Ageratum Yellow Vein Virus DNA-Beta Satellite Localizes Sequences Involved in Replication. J. Gen. Virol. 2008, 89, 3165–3172. [Google Scholar] [CrossRef]
- Sattar, M.N.; Kvarnheden, A.; Saeed, M.; Briddon, R.W. Cotton Leaf Curl Disease - an Emerging Threat to Cotton Production Worldwide. J. Gen. Virol. 2013, 94, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X. Advances in Understanding Begomovirus Satellites. Annu. Rev. Phytopathol. 2013, 51, 357–381. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Navas-Castillo, J. Begomoviruses: What Is the Secret(s) of Their Success? Trends Plant Sci. 2023, 28, 715–727. [Google Scholar] [CrossRef]
- Lozano, G.; Trenado, H.P.; Fiallo-Olivé, E.; Chirinos, D.; Geraud-Pouey, F.; Briddon, R.W.; Navas-Castillo, J. Characterization of Non-Coding DNA Satellites Associated with Sweepoviruses (Genus Begomovirus, Geminiviridae) – Definition of a Distinct Class of Begomovirus-Associated Satellites. Front. Microbiol. 2016, 7, 162. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.E.; Amin, I.; Mansoor, S.; Bedford, I.D.; Rishi, N.; Siwatch, S.S.; Zafar, Y.; Abdel-Salam, A.M.; Markham, P.G. Diversity of DNA 1: A Satellite-like Molecule Associated with Monopartite Begomovirus-DNA Beta Complexes. Virology 2004, 324, 462–474. [Google Scholar] [CrossRef]
- Mansoor, S.; Khan, S.H.; Bashir, A.; Saeed, M.; Zafar, Y.; Malik, K.A.; Briddon, R.; Stanley, J.; Markham, P.G. Identification of a Novel Circular Single-Stranded DNA Associated with Cotton Leaf Curl Disease in Pakistan. Virology 1999, 259, 190–199. [Google Scholar] [CrossRef]
- Saunders, K.; Stanley, J. A Nanovirus-like DNA Component Associated with Yellow Vein Disease of Ageratum Conyzoides: Evidence for Interfamilial Recombination between Plant DNA Viruses. Virology 1999, 264, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Ferro, C.G.; Zerbini, F.M.; Navas-Castillo, J.; Fiallo-Olivé, E. Revealing the Complexity of Sweepovirus-Deltasatellite-Plant Host Interactions: Expanded Natural and Experimental Helper Virus Range and Effect Dependence on Virus-Host Combination. Microorganisms 2021, 9, 1018. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Martínez-Zubiaur, Y.; Moriones, E.; Navas-Castillo, J. A Novel Class of DNA Satellites Associated with New World Begomoviruses. Virology 2012, 426, 1–6. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Navas-Castillo, J. Molecular and Biological Characterization of a New World Mono-/Bipartite Begomovirus/Deltasatellite Complex Infecting Corchorus Siliquosus. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Bennett, A.; Mietzsch, M.; Agbandje-McKenna, M. Understanding Capsid Assembly and Genome Packaging for Adeno-Associated Viruses. Future Virol. 2017, 12, 283–297. [Google Scholar] [CrossRef]
- Bikard, D.; Loot, C.; Baharoglu, Z.; Mazel, D. Folded DNA in Action: Hairpin Formation and Biological Functions in Prokaryotes. Microbiol. Mol. Biol. Rev. MMBR 2010, 74, 570–588. [Google Scholar] [CrossRef] [PubMed]
- Musatov, S.; Roberts, J.; Pfaff, D.; Kaplitt, M. A Cis-Acting Element That Directs Circular Adeno-Associated Virus Replication and Packaging. J. Virol. 2002, 76, 12792–12802. [Google Scholar] [CrossRef]
- Russel, M.; Linderoth, N.A.; Sali, A. Filamentous Phage Assembly: Variation on a Protein Export Theme. Gene 1997, 192, 23–32. [Google Scholar] [CrossRef]
- Russel, M.; Model, P. Genetic Analysis of the Filamentous Bacteriophage Packaging Signal and of the Proteins That Interact with It. J. Virol. 1989, 63, 3284–3295. [Google Scholar] [CrossRef]
- Sun, S.; Rao, V.B.; Rossmann, M.G. Genome Packaging in Viruses. Curr. Opin. Struct. Biol. 2010, 20, 114–120. [Google Scholar] [CrossRef]
- Singh, A.; Chattopadhyay, B.; Chakraborty, S. Biology and Interactions of Two Distinct Monopartite Begomoviruses and Betasatellites Associated with Radish Leaf Curl Disease in India. Virol. J. 2012, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, D.; Gnanasekaran, P.; Kumar, R.K.; Kushwaha, N.K.; Sharma, V.K.; Yusuf, M.A.; Chakraborty, S. A Geminivirus Betasatellite Damages the Structural and Functional Integrity of Chloroplasts Leading to Symptom Formation and Inhibition of Photosynthesis. J. Exp. Bot. 2015, 66, 5881–5895. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xie, Y.; Raja, P.; Li, S.; Wolf, J.N.; Shen, Q.; Bisaro, D.M.; Zhou, X. Suppression of Methylation-Mediated Transcriptional Gene Silencing by βC1-SAHH Protein Interaction during Geminivirus-Betasatellite Infection. PLOS Pathog. 2011, 7, e1002329. [Google Scholar] [CrossRef]
- Tahir, M.; Haider, M.S.; Briddon, R.W. Chili Leaf Curl Betasatellite Is Associated with a Distinct Recombinant Begomovirus, Pepper Leaf Curl Lahore Virus, in Capsicum in Pakistan. Virus Res. 2010, 149, 109–114. [Google Scholar] [CrossRef]
- Venkataravanappa, V.; Kodandaram, M.H.; Prasanna, H.C.; Reddy, M.K.; Reddy, C.N.L. Unraveling Different Begomoviruses, DNA Satellites and Cryptic Species of Bemisia Tabaci and Their Endosymbionts in Vegetable Ecosystem. Microb. Pathog. 2023, 174, 105892. [Google Scholar] [CrossRef]
- Leke, W.N.; Sattar, M.N.; Ngane, E.B.; Ngeve, J.M.; Kvarnheden, A.; Brown, J.K. Molecular Characterization of Begomoviruses and DNA Satellites Associated with Okra Leaf Curl Disease in Cameroon. Virus Res. 2013, 174, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Leke, W.N.; Brown, J.K.; Ligthart, M.E.; Sattar, N.; Njualem, D.K.; Kvarnheden, A. Ageratum Conyzoides: A Host to a Unique Begomovirus Disease Complex in Cameroon. Virus Res. 2012, 163, 229–237. [Google Scholar] [CrossRef]
- Kon, T.; Gilbertson, R.L. Two Genetically Related Begomoviruses Causing Tomato Leaf Curl Disease in Togo and Nigeria Differ in Virulence and Host Range but Do Not Require a Betasatellite for Induction of Disease Symptoms. Arch. Virol. 2012, 157, 107–120. [Google Scholar] [CrossRef]
- Li, Z.; Xie, Y.; Zhou, X. Tobacco Curly Shoot Virus DNAβ Is Not Necessary for Infection but Intensifies Symptoms in a Host-Dependent Manner. Phytopathology® 2005, 95, 902–908. [Google Scholar] [CrossRef]
- Chen, L.-F.; Rojas, M.; Kon, T.; Gamby, K.; Xoconostle-Cazares, B.; Gilbertson, R.L. A Severe Symptom Phenotype in Tomato in Mali Is Caused by a Reassortant between a Novel Recombinant Begomovirus (Tomato Yellow Leaf Curl Mali Virus) and a Betasatellite. Mol. Plant Pathol. 2009, 10, 415–430. [Google Scholar] [CrossRef]
- Tiendrébéogo, F.; Lefeuvre, P.; Hoareau, M.; Villemot, J.; Konaté, G.; Traoré, A.S.; Barro, N.; Traoré, V.S.; Reynaud, B.; Traoré, O.; et al. Molecular Diversity of Cotton Leaf Curl Gezira Virus Isolates and Their Satellite DNAs Associated with Okra Leaf Curl Disease in Burkina Faso. Virol. J. 2010, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Mollel, H.G.; Ndunguru, J.; Sseruwagi, P.; Alicai, T.; Colvin, J.; Navas-Castillo, J.; Fiallo-Olivé, E. A Novel East African Monopartite Begomovirus-Betasatellite Complex That Infects Vernonia Amygdalina. Arch. Virol. 2017, 162, 1079–1082. [Google Scholar] [CrossRef]
- Li, F.; Huang, C.; Li, Z.; Zhou, X. Suppression of RNA Silencing by a Plant DNA Virus Satellite Requires a Host Calmodulin-Like Protein to Repress RDR6 Expression. PLOS Pathog. 2014, 10, e1003921. [Google Scholar] [CrossRef] [PubMed]
- Eini, O. [A Betasatellite-Encoded Protein Regulates Key Components of Gene Silencing System in Plants]. Mol. Biol. (Mosk.) 2017, 51, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Kumar P, P.; Usha, R.; Zrachya, A.; Levy, Y.; Spanov, H.; Gafni, Y. Protein-Protein Interactions and Nuclear Trafficking of Coat Protein and betaC1 Protein Associated with Bhendi Yellow Vein Mosaic Disease. Virus Res. 2006, 122, 127–136. [Google Scholar] [CrossRef]
- Fu, Y.; Xu, L.; Xu, B.; Yang, L.; Ling, Q.; Wang, H.; Huang, H. Genetic Interactions between Leaf Polarity-Controlling Genes and ASYMMETRIC LEAVES1 and 2 in Arabidopsis Leaf Patterning. Plant Cell Physiol. 2007, 48, 724–735. [Google Scholar] [CrossRef]
- Zubair, M.; Zaidi, S.S.-A.; Shakir, S.; Amin, I.; Mansoor, S. An Insight into Cotton Leaf Curl Multan Betasatellite, the Most Important Component of Cotton Leaf Curl Disease Complex. Viruses 2017, 9, 280. [Google Scholar] [CrossRef]
- Harimalala, M.; De Bruyn, A.; Hoareau, M.; Andrianjaka, A.; Ranomenjanahary, S.; Reynaud, B.; Lefeuvre, P.; Lett, J.-M. Molecular Characterization of a New Alphasatellite Associated with a Cassava Mosaic Geminivirus in Madagascar. Arch. Virol. 2013, 158, 1829–1832. [Google Scholar] [CrossRef]
- Kumar, M.; Zarreen, F.; Chakraborty, S. Roles of Two Distinct Alphasatellites Modulating Geminivirus Pathogenesis. Virol. J. 2021, 18, 249. [Google Scholar] [CrossRef]
- Romay, G.; Chirinos, D.; Geraud-Pouey, F.; Desbiez, C. Association of an Atypical Alphasatellite with a Bipartite New World Begomovirus. Arch. Virol. 2010, 155, 1843–1847. [Google Scholar] [CrossRef]
- Jeske, H.; Kober, S.; Schäfer, B.; Strohmeier, S. Circomics of Cuban Geminiviruses Reveals the First Alpha-Satellite DNA in the Caribbean. Virus Genes 2014, 49, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Paprotka, T.; Metzler, V.; Jeske, H. The First DNA 1-like Alpha Satellites in Association with New World Begomoviruses in Natural Infections. Virology 2010, 404, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Rosario, K.; Marr, C.; Varsani, A.; Kraberger, S.; Stainton, D.; Moriones, E.; Polston, J.E.; Breitbart, M. Begomovirus-Associated Satellite DNA Diversity Captured Through Vector-Enabled Metagenomic (VEM) Surveys Using Whiteflies (Aleyrodidae). Viruses 2016, 8, 36. [Google Scholar] [CrossRef]
- Yoboué, A.A.N.; Kouakou, B.S.M.; Pita, J.S.; N’Zué, B.; Amoakon, W.J.-L.; Kouassi, K.M.; Vanié-Léabo, L.P.L.; Kouassi, N.K.; Sorho, F.; Zouzou, M. Emergence of Begomoviruses and DNA Satellites Associated with Weeds and Intercrops: A Potential Threat to Sustainable Production of Cassava in Côte d’Ivoire. Front. Plant Sci. 2025, 16. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wu, P.; Liu, P.; Gong, H.; Zhou, X. Characterization of Alphasatellites Associated with Monopartite Begomovirus/Betasatellite Complexes in Yunnan, China. Virol. J. 2010, 7, 178. [Google Scholar] [CrossRef]
- Zhao, L.; Che, X.; Wang, Z.; Zhou, X.; Xie, Y. Functional Characterization of Replication-Associated Proteins Encoded by Alphasatellites Identified in Yunnan Province, China. Viruses 2022, 14, 222. [Google Scholar] [CrossRef]
- Abbas, Q.; Amin, I.; Mansoor, S.; Shafiq, M.; Wassenegger, M.; Briddon, R.W. The Rep Proteins Encoded by Alphasatellites Restore Expression of a Transcriptionally Silenced Green Fluorescent Protein Transgene in Nicotiana Benthamiana. VirusDisease 2019, 30, 101–105. [Google Scholar] [CrossRef]
- Nawaz-Ul-Rehman, M.S.; Nahid, N.; Mansoor, S.; Briddon, R.W.; Fauquet, C.M. Post-Transcriptional Gene Silencing Suppressor Activity of Two Non-Pathogenic Alphasatellites Associated with a Begomovirus. Virology 2010, 405, 300–308. [Google Scholar] [CrossRef]
- Mar, T.B.; Mendes, I.R.; Lau, D.; Fiallo-Olivé, E.; Navas-Castillo, J.; Alves, M.S.; Murilo Zerbini, F. Interaction between the New World Begomovirus Euphorbia Yellow Mosaic Virus and Its Associated Alphasatellite: Effects on Infection and Transmission by the Whitefly Bemisia Tabaci. J. Gen. Virol. 2017, 98, 1552–1562. [Google Scholar] [CrossRef]
- Nogueira, A.M.; Nascimento, M.B.; Barbosa, T.M.C.; Quadros, A.F.F.; Gomes, J.P.A.; Orílio, A.F.; Barros, D.R.; Zerbini, F.M. The Association between New World Alphasatellites and Bipartite Begomoviruses: Effects on Infection and Vector Transmission. Pathogens 2021, 10, 1244. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Tovar, R.; Navas-Castillo, J. Deciphering the Biology of Deltasatellites from the New World: Maintenance by New World Begomoviruses and Whitefly Transmission. New Phytol. 2016, 212, 680–692. [Google Scholar] [CrossRef] [PubMed]
- Hassan, I.; Orílio, A.F.; Fiallo-Olivé, E.; Briddon, R.W.; Navas-Castillo, J. Infectivity, Effects on Helper Viruses and Whitefly Transmission of the Deltasatellites Associated with Sweepoviruses (Genus Begomovirus, Family Geminiviridae). Sci. Rep. 2016, 6, 30204. [Google Scholar] [CrossRef]
- Sattar, M.N.; Ligthart, M.; Kvarnheden, A. Compatibility and Interaction of Begomoviruses and DNA-Satellites Causing Leaf Curl Disease in Asia, Africa and Mediterranean Region. Eur. J. Plant Pathol. 2019, 155, 111–124. [Google Scholar] [CrossRef]
- Fouad, N.; Granier, M.; Blanc, S.; Thébaud, G.; Urbino, C. Demonstration of Insect Vector-Mediated Transfer of a Betasatellite between Two Helper Viruses. Viruses 2024, 16, 1420. [Google Scholar] [CrossRef]
- Varma, A.; Singh, M.K. The Role of Satellites in the Evolution of Begomoviruses. Viruses 2024, 16, 970. [Google Scholar] [CrossRef] [PubMed]
- Stanley, J.; Saunders, K.; Pinner, M.S.; Wong, S.M. Novel Defective Interfering DNAs Associated with Ageratum Yellow Vein Geminivirus Infection of Ageratum Conyzoides. Virology 1997, 239, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Navas-Castillo, J. The Role of Extensive Recombination in the Evolution of Geminiviruses. Curr. Top. Microbiol. Immunol. 2023, 439, 139–166. [Google Scholar] [CrossRef]
- Baig, M.S.; Akhtar, S.; Khan, J.A. Engineering Tolerance to CLCuD in Transgenic Gossypium Hirsutum Cv. HS6 Expressing Cotton Leaf Curl Multan Virus-C4 Intron Hairpin. Sci. Rep. 2021, 11, 14172. [Google Scholar] [CrossRef]
- Biswas, K.K.; Bhattacharyya, U.K.; Palchoudhury, S.; Balram, N.; Kumar, A.; Arora, R.; Sain, S.K.; Kumar, P.; Khetarpal, R.K.; Sanyal, A.; et al. Dominance of Recombinant Cotton Leaf Curl Multan-Rajasthan Virus Associated with Cotton Leaf Curl Disease Outbreak in Northwest India. PLoS ONE 2020, 15, e0231886. [Google Scholar] [CrossRef]
- Qadir, R.; Khan, Z.A.; Monga, D.; Khan, J.A. Diversity and Recombination Analysis of Cotton Leaf Curl Multan Virus: A Highly Emerging Begomovirus in Northern India. BMC Genomics 2019, 20, 274. [Google Scholar] [CrossRef]
- Amin, I.; Mansoor, S.; Amrao, L.; Hussain, M.; Irum, S.; Zafar, Y.; Bull, S.E.; Briddon, R.W. Mobilisation into Cotton and Spread of a Recombinant Cotton Leaf Curl Disease Satellite. Arch. Virol. 2006, 151, 2055–2065. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Amin, I.; Zaidi, S.S.-E.-A.; Rahman, S.U.; Farooq, M.; Fauquet, C.M.; Mansoor, S. Circular DNA Enrichment Sequencing Reveals the Viral/Satellites Genetic Diversity Associated with the Third Epidemic of Cotton Leaf Curl Disease. Biol. Methods Protoc. 2021, 6, bpab005. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Akbar, F.; Iqbal, Z.; Amrao, L.; Amin, I.; Saeed, M.; Mansoor, S. Effects of Genetic Changes to the Begomovirus/Betasatellite Complex Causing Cotton Leaf Curl Disease in South Asia Post-Resistance Breaking. Virus Res. 2014, 186, 114–119. [Google Scholar] [CrossRef]
- Mansoor, S.; Amin, I.; Iram, S.; Hussain, M.; Zafar, Y.; Malik, K.A.; Briddon, R.W. Breakdown of Resistance in Cotton to Cotton Leaf Curl Disease in Pakistan. Plant Pathol. 2003, 52, 784–784. [Google Scholar] [CrossRef]
- Aimone, C.D.; De León, L.; Dallas, M.M.; Ndunguru, J.; Ascencio-Ibáñez, J.T.; Hanley-Bowdoin, L. A New Type of Satellite Associated with Cassava Mosaic Begomoviruses. J. Virol. 2021, 95, e0043221. [Google Scholar] [CrossRef]
- Ndunguru, J.; De León, L.; Doyle, C.D.; Sseruwagi, P.; Plata, G.; Legg, J.P.; Thompson, G.; Tohme, J.; Aveling, T.; Ascencio-Ibáñez, J.T.; et al. Two Novel DNAs That Enhance Symptoms and Overcome CMD2 Resistance to Cassava Mosaic Disease. J. Virol. 2016, 90, 4160–4173. [Google Scholar] [CrossRef] [PubMed]
- Torkpo, S.K.; Amponsah, E.; Arhin, C.D.; Offei, S.K. Occurrence of Cassava Mosaic Begomovirus-Associated Satellites on Cassava in Ghana. Cogent Food Agric. 2021, 7, 1963929. [Google Scholar] [CrossRef]
- Borah, B.K.; Cheema, G.S.; Gill, C.K.; Dasgupta, I. A Geminivirus-Satellite Complex Is Associated with Leaf Deformity of Mentha (Mint) Plants in Punjab. Indian J. Virol. Off. Organ Indian Virol. Soc. 2010, 21, 103–109. [Google Scholar] [CrossRef]
- Geissler, S.; Siegers, K.; Schiebel, E. A Novel Protein Complex Promoting Formation of Functional Alpha- and Gamma-Tubulin. EMBO J. 1998, 17, 952–966. [Google Scholar] [CrossRef]
- Herranz-Montoya, I.; Park, S.; Djouder, N. A Comprehensive Analysis of Prefoldins and Their Implication in Cancer. iScience 2021, 24, 103273. [Google Scholar] [CrossRef]
- Li, X.; Sun, M.; Liu, S.; Teng, Q.; Li, S.; Jiang, Y. Functions of PPR Proteins in Plant Growth and Development. Int. J. Mol. Sci. 2021, 22, 11274. [Google Scholar] [CrossRef] [PubMed]
- Umate, P.; Tuteja, R.; Tuteja, N. Genome-Wide Analysis of Helicase Gene Family from Rice and Arabidopsis: A Comparison with Yeast and Human. Plant Mol. Biol. 2010, 73, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Ashikawa, I. Gene-Associated CpG Islands in Plants as Revealed by Analyses of Genomic Sequences. Plant J. Cell Mol. Biol. 2001, 26, 617–625. [Google Scholar] [CrossRef]
- Deaton, A.M.; Bird, A. CpG Islands and the Regulation of Transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [PubMed]
- Stephens, K.E.; Miaskowski, C.A.; Levine, J.D.; Pullinger, C.R.; Aouizerat, B.E. Epigenetic Regulation and Measurement of Epigenetic Changes. Biol. Res. Nurs. 2013, 15, 373–381. [Google Scholar] [CrossRef]
- Zhang, X.; Yazaki, J.; Sundaresan, A.; Cokus, S.; Chan, S.W.-L.; Chen, H.; Henderson, I.R.; Shinn, P.; Pellegrini, M.; Jacobsen, S.E.; et al. Genome-Wide High-Resolution Mapping and Functional Analysis of DNA Methylation in Arabidopsis. Cell 2006, 126, 1189–1201. [Google Scholar] [CrossRef]
- Zilberman, D.; Gehring, M.; Tran, R.K.; Ballinger, T.; Henikoff, S. Genome-Wide Analysis of Arabidopsis Thaliana DNA Methylation Uncovers an Interdependence between Methylation and Transcription. Nat. Genet. 2007, 39, 61–69. [Google Scholar] [CrossRef]
- Chiunga, E.; Aimone, C.D.; Rajabu, C.; Dallas, M.M.; Ndunguru, J.; Ascencio-Ibáñez, J.T.; Ateka, E.M.; Hanley-Bowdoin, L. SEGS-1 Episomes Generated during Cassava Mosaic Disease Enhance Disease Severity. Front. Plant Sci. 2024, 15, 1469045. [Google Scholar] [CrossRef]
- Nagar, S.; Hanley-Bowdoin, L.; Robertson, D. Host DNA Replication Is Induced by Geminivirus Infection of Differentiated Plant Cells. Plant Cell 2002, 14, 2995–3007. [Google Scholar] [CrossRef]
- Ascencio-Ibáñez, J.T.; Sozzani, R.; Lee, T.-J.; Chu, T.-M.; Wolfinger, R.D.; Cella, R.; Hanley-Bowdoin, L. Global Analysis of Arabidopsis Gene Expression Uncovers a Complex Array of Changes Impacting Pathogen Response and Cell Cycle during Geminivirus Infection. Plant Physiol. 2008, 148, 436–454. [Google Scholar] [CrossRef]
- Gui, X.; Liu, C.; Qi, Y.; Zhou, X. Geminiviruses Employ Host DNA Glycosylases to Subvert DNA Methylation-Mediated Defense. Nat. Commun. 2022, 13, 575. [Google Scholar] [CrossRef] [PubMed]
- Rajabu, C.A.; Dallas, M.M.; Chiunga, E.; De León, L.; Ateka, E.M.; Tairo, F.; Ndunguru, J.; Ascencio-Ibanez, J.T.; Hanley-Bowdoin, L. SEGS-1 a Cassava Genomic Sequence Increases the Severity of African Cassava Mosaic Virus Infection in Arabidopsis Thaliana. Front. Plant Sci. 2023, 14, 1250105. [Google Scholar] [CrossRef]
- Fortes, I.M.; Pérez-Padilla, V.; Romero-Rodríguez, B.; Fernández-Muñoz, R.; Moyano, C.; Castillo, A.G.; De León, L.; Moriones, E. Begomovirus Tomato Leaf Curl New Delhi Virus Is Seedborne but Not Seed Transmitted in Melon. Plant Dis. 2023, 107, 473–479. [Google Scholar] [CrossRef]
- Moriones, E.; Praveen, S.; Chakraborty, S. Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops. Viruses 2017, 9, 264. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Pendón, J.A.; Sánchez-Campos, S.; Fortes, I.M.; Moriones, E. Tomato Yellow Leaf Curl Sardinia Virus, a Begomovirus Species Evolving by Mutation and Recombination: A Challenge for Virus Control. Viruses 2019, 11, 45. [Google Scholar] [CrossRef]
- Leke, W.N.; Khatabi, B.; Mignouna, D.B.; Brown, J.K.; Fondong, V.N. Complete Genome Sequence of a New Bipartite Begomovirus Infecting Cotton in the Republic of Benin in West Africa. Arch. Virol. 2016, 161, 2329–2333. [Google Scholar] [CrossRef] [PubMed]
- Leke, W.N.; Mignouna, D.B.; Brown, J.K.; Fondong, V.N. First Report of Soybean Chlorotic Blotch Virus and West African Asystasia Virus 1 Infecting Cassava and a Wild Cassava Relative in Cameroon and Togo. New Dis. Rep. 2016, 33, 24–24. [Google Scholar] [CrossRef]
- Keita, G.K.; Kansaye, L.; Doumbia, L.; Keita, I.; Sangare, M.; Sinayoko, T.; Macalou, B.; Maiga, M.N.; Sanogo, N.P.; Koita, O. Molecular Identification and Characterization of a Begomovirus Associated with Okra Enation Leaf Curl Disease in Mali. Adv. Biosci. Biotechnol. 2023, 14, 429–438. [Google Scholar] [CrossRef]
- Baulcombe, D.C.; Saunders, G.R.; Bevan, M.W.; Mayo, M.A.; Harrison, B.D. Expression of Biologically Active Viral Satellite RNA from the Nuclear Genome of Transformed Plants. Nature 1986, 321, 446–449. [Google Scholar] [CrossRef]
- Cao, X.; Liu, S.; Yu, C.; Li, X.; Yuan, X. A New Strategy of Using Satellite RNA to Control Viral Plant Diseases: Post-inoculation with Satellite RNA Attenuates Symptoms Derived from Pre-infection with Its Helper Virus. Plant Biotechnol. J. 2019, 17, 1856–1858. [Google Scholar] [CrossRef]
- Cillo, F.; Finetti-Sialer, M.M.; Papanice, M.A.; Gallitelli, D. Analysis of Mechanisms Involved in the Cucumber Mosaic Virus Satellite RNA-Mediated Transgenic Resistance in Tomato Plants. Mol. Plant-Microbe Interactions® 2004, 17, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, W.L.; Llewellyn, D.; Haseloff, J. Construction of a Plant Disease Resistance Gene from the Satellite RNA of Tobacco Ringspot Virus. Nature 1987, 328, 802–805. [Google Scholar] [CrossRef]
- Lin, K.-Y.; Lin, N.-S. Interfering Satellite RNAs of Bamboo Mosaic Virus. Front. Microbiol. 2017, 8, 787. [Google Scholar] [CrossRef]
- Simon, A.E.; Roossinck, M.J.; Havelda, Z. PLANT VIRUS SATELLITE AND DEFECTIVE INTERFERING RNAS: New Paradigms for a New Century. Annu. Rev. Phytopathol. 2004, 42, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Tien, P.; Wu, G. Satellite RNA for the Biocontrol of Plant Disease. In Advances in Virus Research; Maramorosch, K., Murphy, F.A., Shatkin, A.J., Eds.; Academic Press, 1991; Vol. 39, pp. 321–339.
- Fondong, V.N. The Search for Resistance to Cassava Mosaic Geminiviruses: How Much We Have Accomplished, and What Lies Ahead. Front. Plant Sci. 2017, 8, 408. [Google Scholar] [CrossRef]
- Fondong, V.N.; Rey, C.; Fondong, V.N.; Rey, C. Recent Biotechnological Advances in the Improvement of Cassava; IntechOpen, 2018; ISBN 978-953-51-3741-2.
- Giudice, G.; Moffa, L.; Varotto, S.; Cardone, M.F.; Bergamini, C.; De Lorenzis, G.; Velasco, R.; Nerva, L.; Chitarra, W. Novel and Emerging Biotechnological Crop Protection Approaches. Plant Biotechnol. J. 2021, 19, 1495–1510. [Google Scholar] [CrossRef]
- Niraula, P.M.; Fondong, V.N. Development and Adoption of Genetically Engineered Plants for Virus Resistance: Advances, Opportunities and Challenges. Plants Basel Switz. 2021, 10, 2339. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-Replace Genome Editing without Double-Strand Breaks or Donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Baldrich, P.; Liu, A.; Meyers, B.C.; Fondong, V.N. An Atlas of Small RNAs from Potato. Plant Direct 2022, 6, e466. [Google Scholar] [CrossRef]
- Luan, W.; Dai, Y.; Li, X.-Y.; Wang, Y.; Tao, X.; Li, C.-X.; Mao, P.; Ma, X.-R. Identification of tRFs and phasiRNAs in Tomato (Solanum Lycopersicum) and Their Responses to Exogenous Abscisic Acid. BMC Plant Biol. 2020, 20, 320. [Google Scholar] [CrossRef]
- Khatabi, B.; Arikit, S.; Xia, R.; Winter, S.; Oumar, D.; Mongomake, K.; Meyers, B.C.; Fondong, V.N. High-Resolution Identification and Abundance Profiling of Cassava (Manihot Esculenta Crantz) microRNAs. BMC Genomics 2016, 17, 85. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Kim, T.; Park, J.H.; Yeom, S.-I.; Kim, S.; Seo, M.-K.; Shin, C.; Choi, D. Genome-Wide Comparative Analysis in Solanaceous Species Reveals Evolution of microRNAs Targeting Defense Genes in Capsicum Spp. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2018, 25, 561–575. [Google Scholar] [CrossRef]
- Zhao, T.; Tao, X.; Li, M.; Gao, M.; Chen, J.; Zhou, N.; Mei, G.; Fang, L.; Ding, L.; Zhou, B.; et al. Role of phasiRNAs from Two Distinct Phasing Frames of GhMYB2 Loci in Cis- Gene Regulation in the Cotton Genome. BMC Plant Biol. 2020, 20, 219. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Genome organization of some of bipartite and monopartite begomoviruses genome. (A) Bipartite begomovirus sida golden yellow vein virus (SiGYVV) DNA-A and DNA-B genome components; and monopartite begomoviruses (B) sweet potato leaf curl virus (SPLCV). Canonical genes encoding viral proteins are represented by curved arrows and are coat protein (CP), movement protein (MP), replication enhancer protein (Ren), replication-associated proteins (Rep and RepA), transcriptional activator protein (TrAP).
Figure 1.
Genome organization of some of bipartite and monopartite begomoviruses genome. (A) Bipartite begomovirus sida golden yellow vein virus (SiGYVV) DNA-A and DNA-B genome components; and monopartite begomoviruses (B) sweet potato leaf curl virus (SPLCV). Canonical genes encoding viral proteins are represented by curved arrows and are coat protein (CP), movement protein (MP), replication enhancer protein (Ren), replication-associated proteins (Rep and RepA), transcriptional activator protein (TrAP).
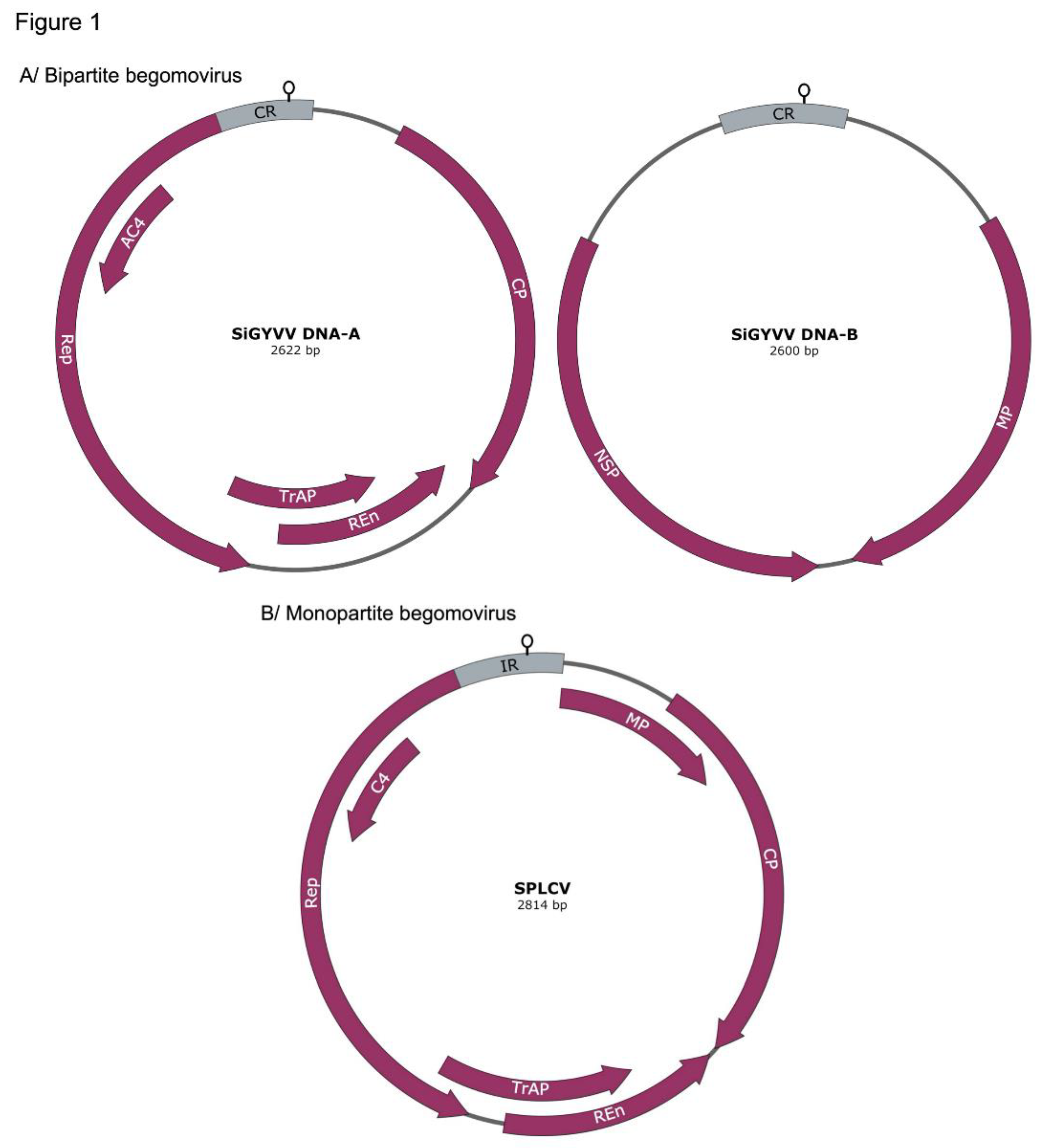
Figure 2.
Representative genomes of begomovirus DNA satellites discussed in this review. (A) Cotton leaf curl Multan betasatellite (CLCuMuB), (B) Cotton leaf curl Multan alphasatellite (CLCuMuA), (C) Sweet potato leaf curl deltasatellite (SPLCV-SPLCD1), (D) Tomato leaf curl deltasatellite (ToLCD), and (E) New World deltasatellite (NW Deltasatellite). Betasatellite encodes βC1, which is a pathogenicity determinant while alphasatellites encode the α-Rep protein for autonomous DNA replication. The hairpin structures shown harbor the DNA replication origin (a), CLCuMuA, ToLCD, and NW deltasatellites show other predicted hairpins identified as (b), (c), and (d). The confidence levels of these structures are predicted by SnapGene software (GLS Biotech).
Figure 2.
Representative genomes of begomovirus DNA satellites discussed in this review. (A) Cotton leaf curl Multan betasatellite (CLCuMuB), (B) Cotton leaf curl Multan alphasatellite (CLCuMuA), (C) Sweet potato leaf curl deltasatellite (SPLCV-SPLCD1), (D) Tomato leaf curl deltasatellite (ToLCD), and (E) New World deltasatellite (NW Deltasatellite). Betasatellite encodes βC1, which is a pathogenicity determinant while alphasatellites encode the α-Rep protein for autonomous DNA replication. The hairpin structures shown harbor the DNA replication origin (a), CLCuMuA, ToLCD, and NW deltasatellites show other predicted hairpins identified as (b), (c), and (d). The confidence levels of these structures are predicted by SnapGene software (GLS Biotech).
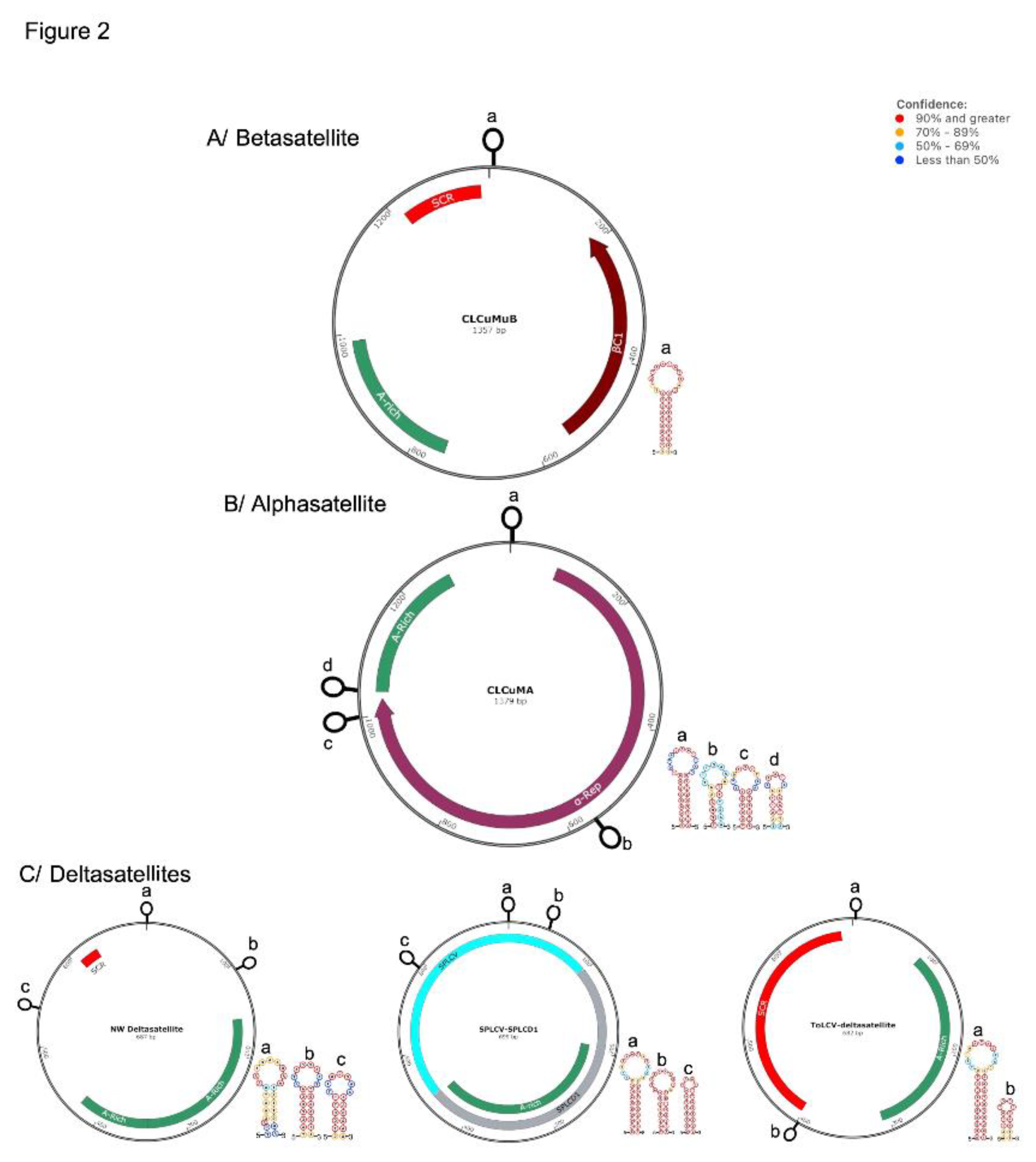
Figure 3.
Proteins encoded by betasatellites and alphasatellites show variation. (A) Except for the conserved PFDFN motif (boxed), βC1 sequence variation, especially at the N- and C-termini of the proteins; (B) α-Reps show sequence variation between New World and Old World α-Reps.
Figure 3.
Proteins encoded by betasatellites and alphasatellites show variation. (A) Except for the conserved PFDFN motif (boxed), βC1 sequence variation, especially at the N- and C-termini of the proteins; (B) α-Reps show sequence variation between New World and Old World α-Reps.
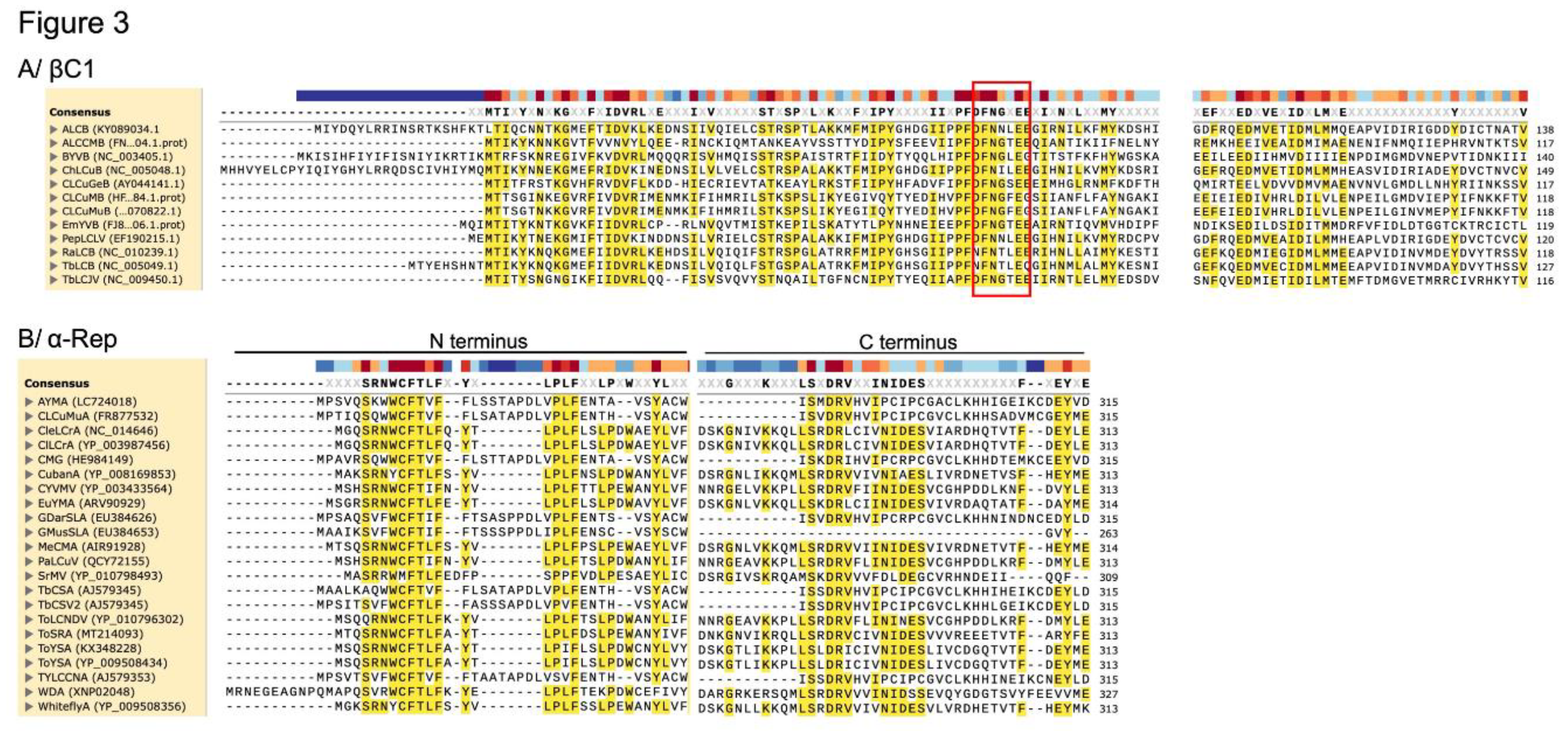
Figure 4.
Asystasia yellow mosaic alphasatellite (AYMA) causes yellowing symptoms in Asystasia gangetica in the presence of West African asystasia virus (WAAV).
Figure 4.
Asystasia yellow mosaic alphasatellite (AYMA) causes yellowing symptoms in Asystasia gangetica in the presence of West African asystasia virus (WAAV).

Figure 5.
Phylogenetic tree generated by the maximum-likelihood method from the alignment of α-Reps. α-Reps of GDarSLA and GMusSLA that have so far shown to suppress PTGS group together in a phylogenetic tree.
Figure 5.
Phylogenetic tree generated by the maximum-likelihood method from the alignment of α-Reps. α-Reps of GDarSLA and GMusSLA that have so far shown to suppress PTGS group together in a phylogenetic tree.
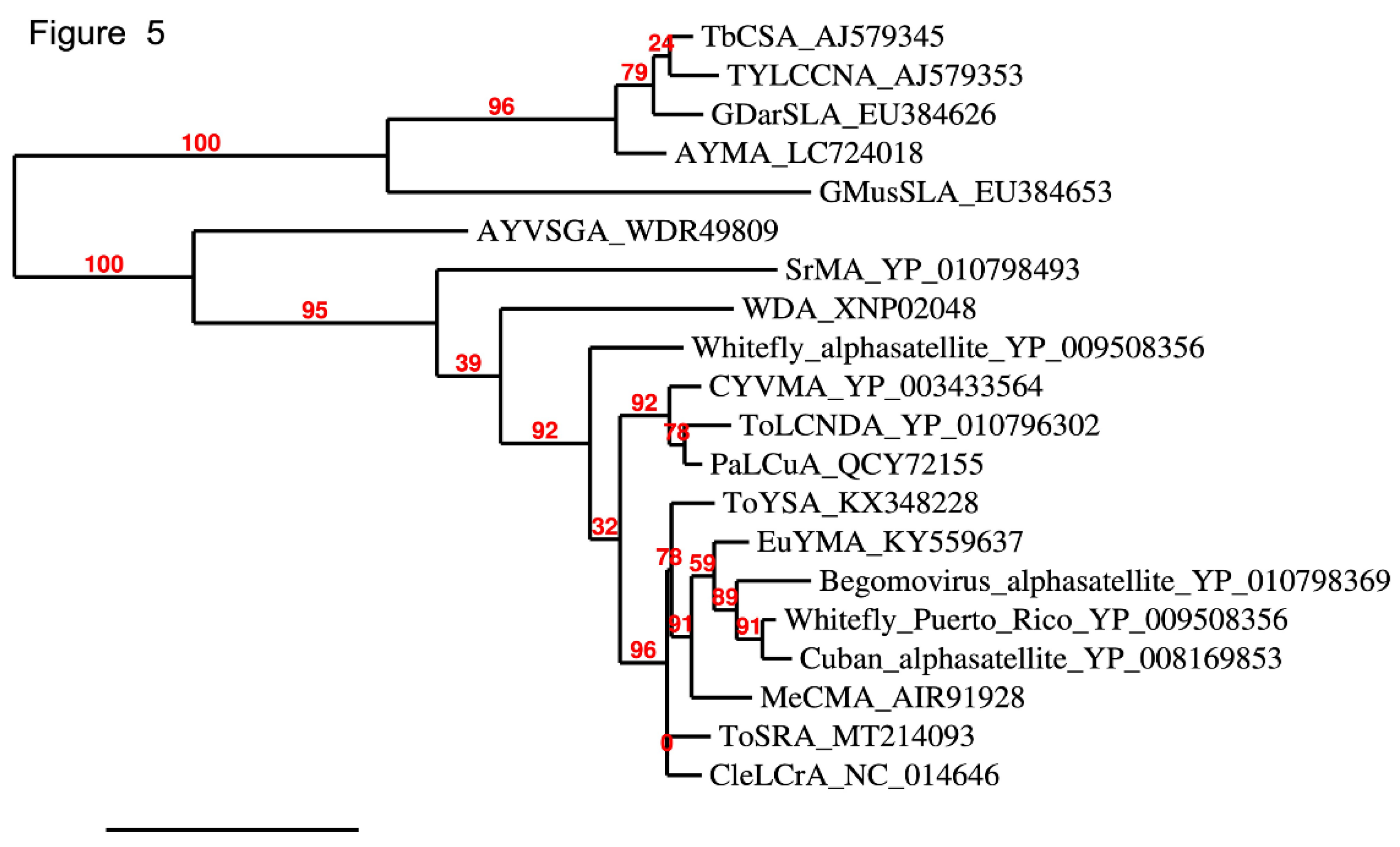
Figure 6.
Representative of geminivirus episomes SEGS. (A) SEGS-1 sequence has identity with the cassava protein prefoldin subunit 6 (PFDN6). (B) SEGS-2 shows sequence identity with the cassava pentatricopeptide repeat (PPR) family, as well as a potential protein of 68 amino acids that maps to the N-terminal first 68 amino acid segment of the pre-mRNA-splicing factor ATP-dependent RNA helicase DEAH1 isoform X2 (DHX16 isoform X2). Like SEGS-1, SEGS-2 has a CpG island and two predicted stem–loop structures. The confidence levels of these structures are predicted by SnapGene software (GLS Biotech).
Figure 6.
Representative of geminivirus episomes SEGS. (A) SEGS-1 sequence has identity with the cassava protein prefoldin subunit 6 (PFDN6). (B) SEGS-2 shows sequence identity with the cassava pentatricopeptide repeat (PPR) family, as well as a potential protein of 68 amino acids that maps to the N-terminal first 68 amino acid segment of the pre-mRNA-splicing factor ATP-dependent RNA helicase DEAH1 isoform X2 (DHX16 isoform X2). Like SEGS-1, SEGS-2 has a CpG island and two predicted stem–loop structures. The confidence levels of these structures are predicted by SnapGene software (GLS Biotech).
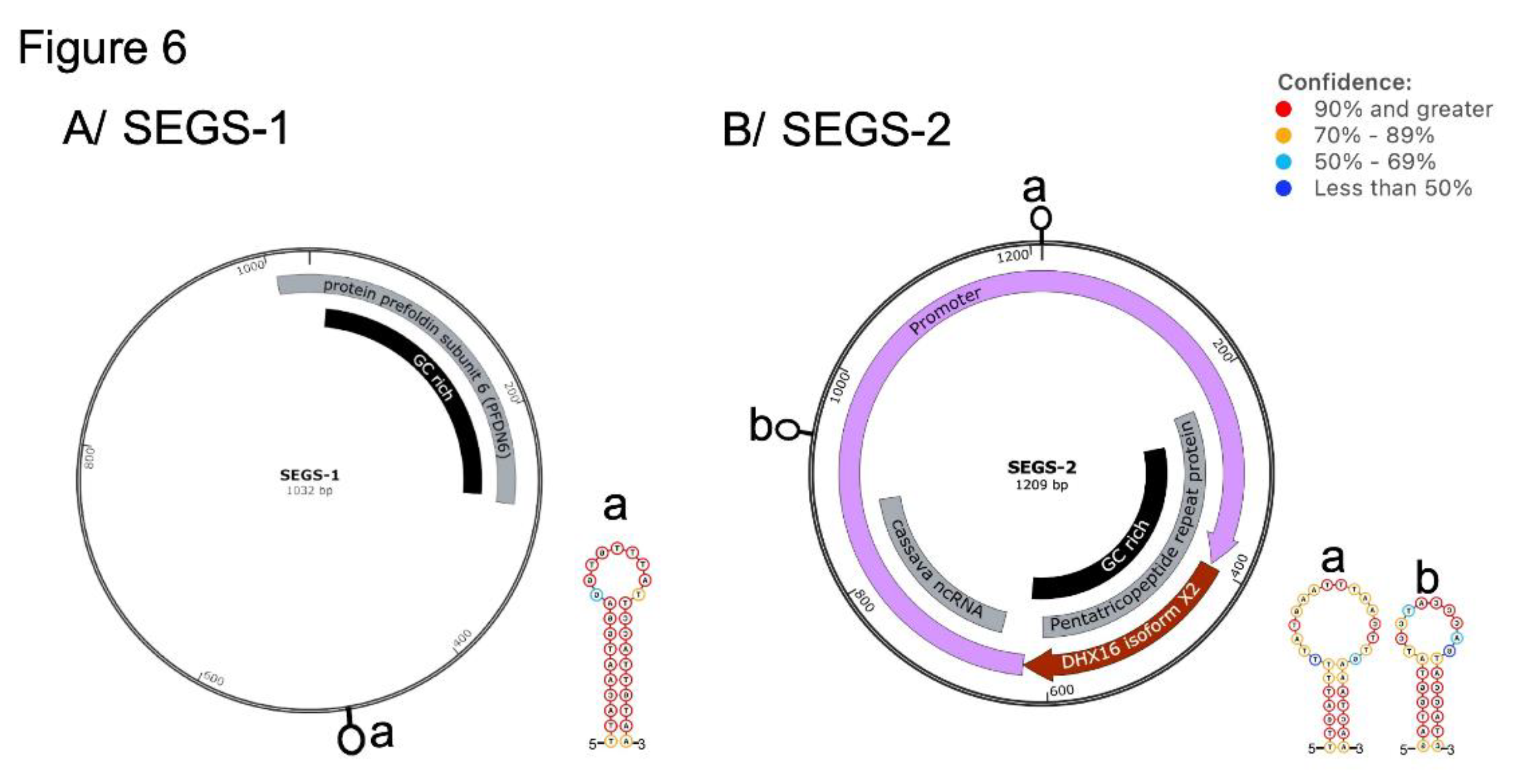
Figure 7.
Alignment of SEGS-2, PPR, and DHX16 isoform X2 shows the specific DNA fragment that became part of the SEGS-2 molecule.
Figure 7.
Alignment of SEGS-2, PPR, and DHX16 isoform X2 shows the specific DNA fragment that became part of the SEGS-2 molecule.
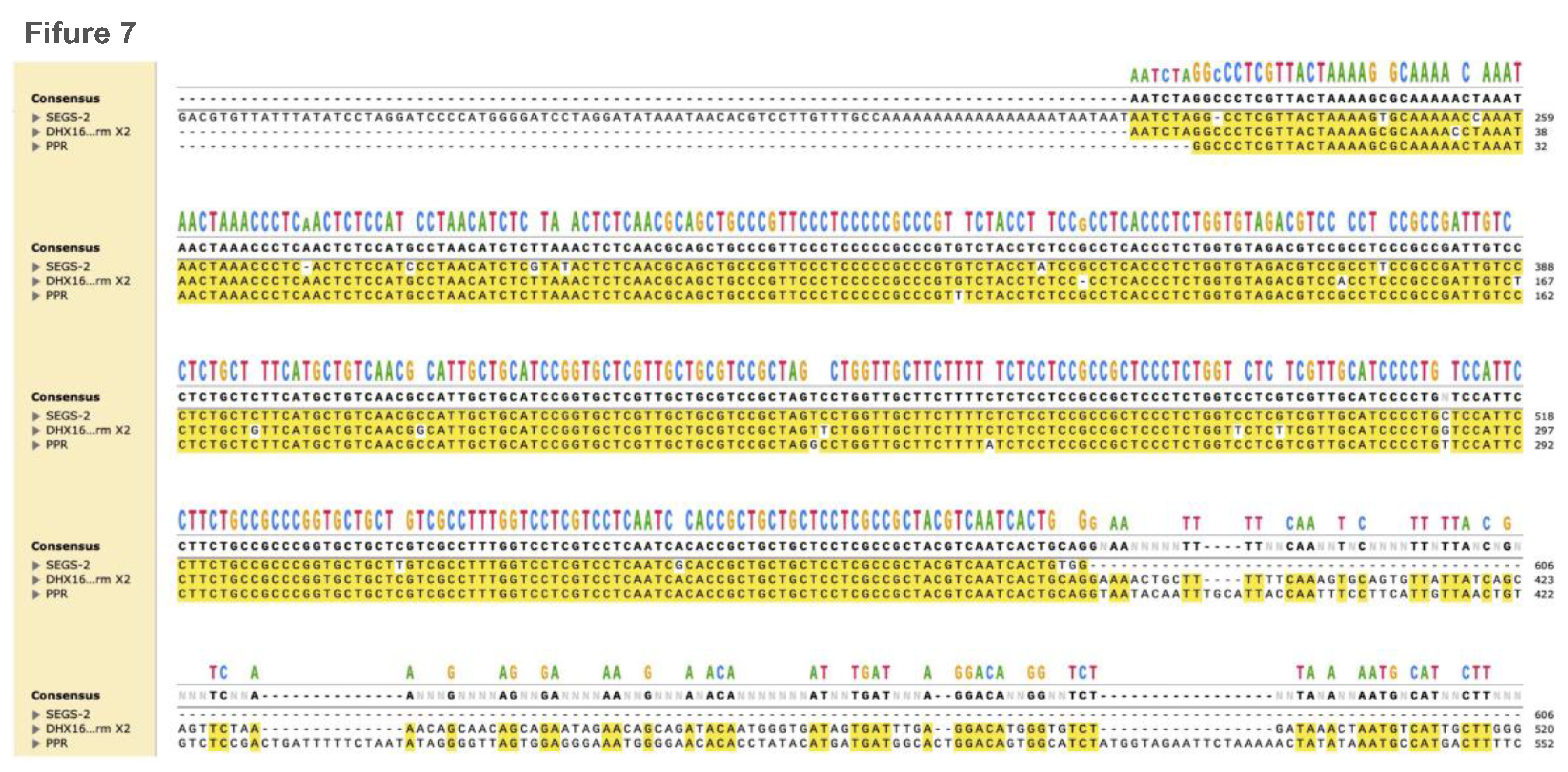
Figure 8.
SEGS episomes cause severe symptoms when mixed infected with cassava mosaic geminiviruses. CMG-resistant cassava showing (A) ACMV-free plant, (B) plant infected by ACMV alone, and (C) plant mixed infected by episomes and ACMV displaying leaves that are reduced to the midrib with a total absence of the leaf blade.
Figure 8.
SEGS episomes cause severe symptoms when mixed infected with cassava mosaic geminiviruses. CMG-resistant cassava showing (A) ACMV-free plant, (B) plant infected by ACMV alone, and (C) plant mixed infected by episomes and ACMV displaying leaves that are reduced to the midrib with a total absence of the leaf blade.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



