Submitted:
25 June 2025
Posted:
26 June 2025
You are already at the latest version
Abstract
The domestic dog has been intensively selectively bred since the Victorian era for morphological traits. Questions about the implications on canine welfare of some of those traits have been asked since that time. More recently, legislation has been proposed and enacted in several countries to guide selection for specific morphologies. We describe common canine phenotypes with associated disorders and the biological mechanisms driving those disorders. We additionally describe opportunities for further study identifying which ranges of morphologies support robust canine health while still preserving the uniqueness of different breeds. Focusing on morphologies with a range of expression, rather than focusing on harmful characteristics prevalent in particular breeds, is a new perspective on canine welfare which we expect to support evidence-based efforts to set future breeding goals, and to increase awareness of these issues by the people who purchase (and love) dogs.
Keywords:
dogs
; animal welfare
; canine welfare
; canine health
; animal breeding
1. Introduction
The domestic dog is the most morphologically diverse mammalian species [1], in part because of the species’ unique evolutionary history [2]. Dogs arose 11,000 to 40,000 years ago [3,4,5] from a population of gray wolves that is now extinct [3,6,7,8]. Like free-living village dogs today, early dogs were human commensals subject to natural selection [9]. Around the Early Bronze period, several cultures began to manage dog populations to form loosely defined types [10,11,12,13]. While it is debated when the first dog breeds were formed [14], the oldest dog breeds in existence today are based on landraces which developed around 500 years ago for functions such as hunting or pulling sleds [15,16], although some have argued for potential earlier emergence [17].
By contrast, most extant dog breeds have much more recent origins [18]. British dog enthusiasts of the late Victorian period were interested in improving the appearances of dogs, which led them to establish breed standards and narrow gene pools [19,20,21]. In many dog breeds, this facilitated selection for novel morphologies that did not exist prior to this period, such as severely shortened snouts [22]. While many Victorians admired dogs with these traits, some called them “monstrosities” [20]. More recently, in several European countries legislative attempts have addressed concerns about extreme morphologies in light of growing scientific evidence that they are associated with compromised welfare [21,23,24,25,26] and the first American municipality has banned the breeding of specified morphologies [27]. Many of the concerning traits may cause pain, which has secondary effects on behavior [28].
Figure 1.
George Du Maurier, “Dog Fashions for 1889.” Punch, 25 Jan. 1889.

This review provides a broad overview of common canine phenotypes with associated disorders. We detail biological mechanisms where they are understood, as well as resulting welfare implications. We argue that clearly defined guidelines are a necessary part of any attempt to modify dog breeding to be more favorable to canine welfare. We hope this review will serve as a base for future work developing those guidelines, which may be of use when extreme morphology becomes a welfare issue, to support evidence-based efforts to set new breeding goals, and to increase awareness of these issues by the people who purchase (and love) dogs.
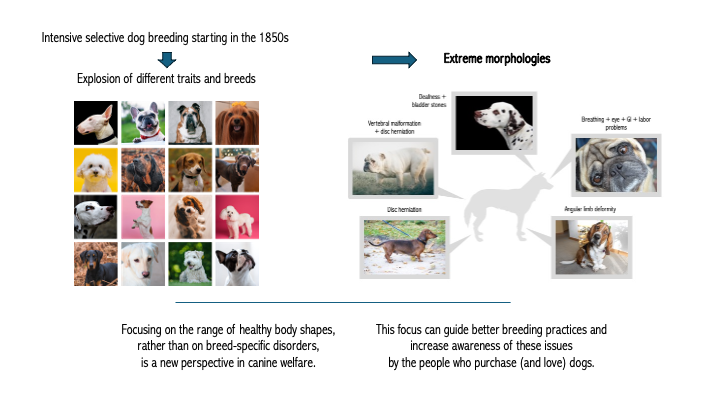
Figure 2.
Extreme morphologies selected for in dog breeds. A. “Screw tail,” associated with congenital vertebral malformations, which in turn are associated with kyphosis and intervertebral disc herniation. B. Extreme white coat color, associated with deafness; spots, associated with urinary tract stones. C. Brachycephaly, associated with brachycephalic obstructive airway syndrome, ocular disorders, skin fold infections, ear infections, and GI disorders. D. “Wrap-around front” legs, associated with angular limb deformity. E. Chondrodystrophy, associated with intervertebral disc herniation.
Figure 2.
Extreme morphologies selected for in dog breeds. A. “Screw tail,” associated with congenital vertebral malformations, which in turn are associated with kyphosis and intervertebral disc herniation. B. Extreme white coat color, associated with deafness; spots, associated with urinary tract stones. C. Brachycephaly, associated with brachycephalic obstructive airway syndrome, ocular disorders, skin fold infections, ear infections, and GI disorders. D. “Wrap-around front” legs, associated with angular limb deformity. E. Chondrodystrophy, associated with intervertebral disc herniation.

2. Size
Dogs have a wide range of sizes, ranging from the Chihuahua (which can stand under 15 cm tall and weigh just 1.5 kg] to the Irish Wolfhound (up to about 94 cm tall) and Mastiff [up to about 105 kg]. An IGF-1 polymorphism is the largest single contributor to the size variation in selectively-bred dogs [29,30], but variations in other genes also play a role in determining canine size [31,32,33,34]. Comparisons between breeds of dramatically different sizes indicate that IGF-1 genetic differences influence circulating hormone levels, which decline throughout life from an early postnatal high, suggesting that IGF-1 variation influences body size by influencing growth rate [35]. Canine allometric scaling differs from scaling of similarly sized mammals of other species, and this difference may be associated with pathologies; for example, the brain, liver, lungs, and kidneys are larger in 2 kg dogs than other mammals of similar size, and smaller in 70 kg dogs; gestation length is fixed in dogs across all sizes, despite being closely correlated to size in other mammalian species [36]. Variation in body size has far-ranging impacts on other traits as well, which we outline in the following sections.
2.1. Lifespan
Small dogs are well known to live longer than larger dogs [37], with a predicted lifespan difference of 4.5 years between dogs of the smallest and largest breeds [38]. This difference is relatively smoothly graded; a 1 kg body mass increase between breeds is associated with about 26 days’ decrease in expected lifespan [38]. The expected lifespans of giant breed dogs are notably short; their age of senescence [i.e., the point of lowest risk of mortality, after which mortality risk begins to increase] can fall before they have reached their full adult growth [39]. Larger dogs have also been found to receive more veterinary care throughout their lives [40].
Larger dogs are more likely to die at every time point once maturity is reached, and the speed of increase of this mortality hazard is positively correlated to body size [37,39]. Postnatal growth rates are negatively correlated with lifespan in mammals [37,41] and large breed dogs grow faster and longer to reach their larger size [35,42] - early growth rates increase by an average of 0.22 kg/week per 10 kg of adult size [37]. This difference in growth rate is most likely primarily driven by differences in circulating levels of IGF-1. Differences in levels of this hormone are associated with longevity differences across species, from nematodes to mammals; in mammals, reduced IGF-1 signaling is associated with increased longevity [43].
While the correlation between size and lifespan has been well documented in dogs, existing analysis has focused on describing correlation across the complete size range. Does an inflection point exist in the relationship between size and lifespan? In other words, how big is too big? Answering this question may help guide future breeding programs.
2.2. Cancer
As dogs increase in size, so too increases their risk of cancer [37,38,43,44,45,46]. The correlation between weight increase and risk of mortality due to cancer is linear from toy to small and medium sized dogs; the line flattens for large and giant sized dogs, likely because cancer is typically a disease of old age, and larger breeds have such reduced lifespans that they do not live into the age range of high cancer risk [46,47].
Why are larger dogs at higher cancer risk? Within a species, species-specific protective mechanisms are likely to be ineffective for individuals of unusually large size. This predicts that larger dogs will have greater cancer risk, which [while allowing for some cancers specific to specific breeds] has been demonstrably true [37,48]. The size-cancer risk relationship is well illustrated by canine osteosarcoma. This disease primarily affects the long bones of the limbs in dogs [48,49]. The mortality related to this disease is high, with a one year survival rate of around 45% [50]. Risk of osteosarcoma in dogs is associated with size: dogs weighing over 60 kg have more than 36 times the risk of developing osteosarcoma compared to dogs of 10-20 kg [51], though height may have a stronger influence on risk than weight alone [52].
It is likely that the increased growth rates in large and giant breed dogs, required to generate such long bones, is the strongest driver of osteosarcoma risk, rather than breed-specific mutations or environmental risk factors [48]. Indeed, genome wide association studies of canine osteosarcoma have found associations between the disease and genes associated with growth: IGF-1 [53], described previously; FGF9 [53], a gene associated with long bone development [54]; and CDKN2A/B [54,55], tumor suppressor genes which operate through interactions with the cell cycle [54,55,56]. Therefore, the most effective preventive approach for this disease may be to breed dogs of less extreme size [51], though the ideal size range for reduction of cancer risk is unknown.
2.3. Orthopedics
The welfare impacts of different orthopedic disorders vary. However, all of them predispose dogs to development of secondary osteoarthritis [57], a disease typically associated with old age which is very commonly diagnosed and can lead to pain and associated decrease in quality of life [58]. Increasing body weight is also associated with risk of osteoarthritis, though frame size has not been well separated from body condition in the literature [58,59].
While large dogs have been found to be at higher risk of orthopedic disease overall [44,46], small dogs are more susceptible to some specific orthopedic disorders, most commonly patellar luxation, with dogs under 10kg having 2.8 greater odds of diagnosis compared to dogs in the 10-19.9 kg range [60]. Canine hip dysplasia (CHD) is a common developmental disorder in which the hip joint is malformed, potentially causing lameness in young dogs, but more commonly resulting in osteoarthritis in older dogs [61]. This disease is more common [or more commonly clinically symptomatic] in medium sized dogs and larger, particularly large and giant sized dogs [62,63,64]. Another common developmental orthopedic disorder of large dogs is elbow dysplasia, an umbrella term for a suite of disorders which may, together, significantly contribute to the later development of osteoarthritis [65,66]. In one study, the risk of elbow dysplasia increased steadily from a baseline of dogs under 10 kg to 15.03 times the odds in dogs over 40 kg [66]. One of the most common orthopedic injuries in dogs is to the cranial cruciate ligament (CCL) [67]. While the etiology of CCL disease is complex, large dogs are diagnosed with it at younger ages [67,68,69,70] and greater frequencies overall [67,68]. The preponderance of studies about risk of orthopedic disease in dogs are breed-specific, and direct analyses of the connection between frame size and risk are needed for many disorders.
2.4. Heart Disease
Small dogs are more likely to be diagnosed with cardiac disease [46]: 75% of breeds averaging under 9 kg suffer cardiac disease as a major cause of death, versus only 25% of breeds averaging over 9 kg [71,72]. The type of cardiac diseases common in small dogs differ from large dogs as well; small dogs are more likely to be diagnosed with mitral valve disease (MVD) [71,72]. The cause is unknown, but hypotheses include a small chest crowding the heart, genes for small size which are known to affect heart development, or simply shared ancestry between many small breeds [71]. Large dogs, conversely, more commonly suffer from dilated cardiomyopathy (DCM), with increased risk reported in dogs over 30 lbs [73] and median weight of DCM cases around 37 kg [74]. Because of lack of coordinated studies of risk, there is no clear guidance around the range of frame size or bodyweight to most greatly reduce the risk of cardiac disease. Does risk spike above and below particular weight ranges? This question has not been addressed, and more studies of risk are called for [72], although medium sized dogs appear to be at lower risk.
2.5. Dental Disease
Risk of periodontal disease (PD - disease of the tissues surrounding the teeth and holding them in place) increases as adult body weight decreases [75]. Dog breeds averaging under 6.5 kg in adult body weight have been found to have up to five times the risk of PD compared to giant breeds, and the majority of dogs diagnosed with PD were from breeds of medium-small size or smaller [76]. Risk of retained deciduous teeth increases as adult size decreases, with a prevalence of 15% in extra small dogs compared to less than 1% in medium-large and larger dogs [77]. As for crowded teeth, smaller dogs may be at increased risk due to having teeth of a larger size relative to their skulls [78], and the consequences of disease in dogs of this size may be greater due to their relatively less alveolar bone [79].
2.6. Energy Regulation
Body mass affects energy regulation, with challenges at both extremes. Very small dogs have an elevated risk of hypoglycemia as a consequence of their low body mass index [80] and may need to allocate more of their metabolism to thermoregulation than larger dogs [81]. Conversely, larger dogs have more difficulty dissipating heat due to reduced surface area to volume ratio, and are at higher risk of heatstroke (dogs weighing over 50 kg have 3.42 times the risk compared to dogs under 10 kg) - an increasing problem in many parts of the world as global temperatures rise [82].
2.7. Labor Difficulties
Bitches at both extremes of size are at increased risk for dystocia (difficulty in labor) [83,84]. The risk for dystocia in small breed dogs is widely recognized, with an increased risk of 3.3 in toy breeds [83,84]. The mechanism for this is hypothesized to be the decreased litter size seen in smaller dogs, leading to larger individual puppy size and a mismatch between puppy size and the birth canal [36,83,84,85]. Dystocia in large breed dogs appears to have been less commonly studied; potentially, the larger litter size in large dogs [85] could lead to exhaustion of the uterine muscles during prolonged labor, which is a risk factor for dystocia [86,87]. Labor difficulties can also result from other extreme morphologies; see section 3.6.
2.8. Behavior
Behavioral differences have been described in smaller breeds, including increased risk of fearful and/or aggressive behavior [88,89,90,91,92] and decreased ability to follow human pointing gestures [93]. Meanwhile, trainability is positively associated with breed body size [88], and behavior scores can cluster by breed body size [94]. However, a study which included mixed-breed dogs and assessed individual rather than breed-average behavior scores suggested that individual (as opposed to breed) body size may not correlate with behavioral differences [34], suggesting breed differences may have more influence on behavior than size alone. Alternatively, small dogs may show reductions in neocortex, a component of the brain linked to trainability and behavioral flexibility, and enlargement in subcortical regions, which are linked to fear, aggression, and behavioral reactivity [95]. This recapitulates an allometric scaling pattern that has long been observed in a broadly cross-species context [96,97,98]. Alternately, behavior differences in small dogs could be due to differences in training and management, as owners of small dogs may be less likely to play with, train, and socialize them [91,92,99]. Size-based behavioral differences could be due to a variety of potential mechanisms, which could be distinguished through further research. These could include genetic influences, which are possibly breed-specific; neuroanatomical differences; visual perceptual differences; and/or differential training regimens by owners who are more likely to tolerate behavior issues in smaller dogs. Alternatively, simply living as a small dog in a big world may lower the threshold for threat perception and increase reliance on behaviors which mitigate perceived threat, such as reactive aggression.
3. Skull Shape
Skull shape varies across dog breeds from the very long and narrow (dolichocephalic) to the canid-average (mesocephalic) to the shortened muzzle and wide head (brachycephalic). Brachycephaly has shown dramatic increases in popularity over recent decades [100,101] despite many associated health disorders resulting in increased veterinary costs and decreased lifespans [40,100,101]. The attraction of the brachycephalic phenotype and cognitive dissonance in relationship to its welfare implications is an active area of study [101,102,103,104]. Future work may focus on the question of how to redirect public interest towards a healthier phenotype.
3.1. Brachycephalic Obstructive Airway Syndrome (BOAS)
The syndrome best associated with brachycephalic dogs is BOAS. Brachycephalic breeds have undergone artificial selection to reduce the bones of the muzzle; however, other structures of the head have not been reduced in concert. The resulting size mismatches in the brachycephalic head cause upper airway obstruction, the primary cause of the syndrome, which results in secondary pathologies due to chronically increased negative airway pressure. Sequelae as the disorder progresses include dyspnea (difficulty breathing), exercise and heat intolerance, increased airway sounds (snoring while awake), sleep apnea, gastrointestinal signs (such as reflux, regurgitation, or vomiting), collapse, and death [105,106,107].
Congenital soft tissue changes associated with the shortening of the muzzle begin at the nostrils, which may be narrowed to slits allowing little space for air passage. The soft palate is thickened, a change which may be primary [105,106,107] but may also be secondary due to chronic struggle to move air [108,109]. The brachycephalic tongue may be thickened, again either a primary or secondary change [110], which can block the upper airway. The trachea may be narrowed, further increasing airway resistance [105,106,107,110,111]. The nasal turbinates - a delicate bony structure in the sinus space, designed to support thermoregulation - may protrude into and impede the airway [112,113]. These various deformations may result in single to multiple points of mucosal-mucosal contact throughout the upper airway, i.e., blockages decreasing the space for air passage and causing increased negative airway pressure [112,113].
The disorder is often progressive due to this chronic resistance to airflow [114]. Secondary changes may include thickening of soft tissues such as the soft palate [108] and tongue [110], potentially followed by inflammation and degradation of the laryngeal tissue [105,106,107,111]. Laryngeal collapse may follow, i.e., temporary complete blockage of the airway [105,106,107]. The increased negative airway pressure may also affect the gastrointestinal tract, and BOAS is commonly associated with reflux, regurgitation, vomiting, and temporary herniation of parts of the stomach into the thoracic cavity [115,116]. Thermoregulation can also be markedly affected; an English Bulldog has been documented as hyperthermic while standing at rest in a temperature controlled room [117]. The majority of the canine airflow passes through the nostrils rather than the mouth, and thermoregulation through panting is most efficient when the air passes in through the nostrils and out through the mouth [118], meaning that decreases in upper airway airflow such as seen in brachycephalic dogs are particularly consequential for thermoregulation.
Figure 3.
Computer tomography of mesocephalic skull (A,C,E) and brachycephalic skull (B,D,F). Tongue is highlighted with green. (Aqua represents the oropharynx, blue the nasopharynx, and pink the palatal tissues.) Note the difference in tongue thickness between the two skull types and its potential to block the airway (marked with red lines in A, B) [110].
Figure 3.
Computer tomography of mesocephalic skull (A,C,E) and brachycephalic skull (B,D,F). Tongue is highlighted with green. (Aqua represents the oropharynx, blue the nasopharynx, and pink the palatal tissues.) Note the difference in tongue thickness between the two skull types and its potential to block the airway (marked with red lines in A, B) [110].
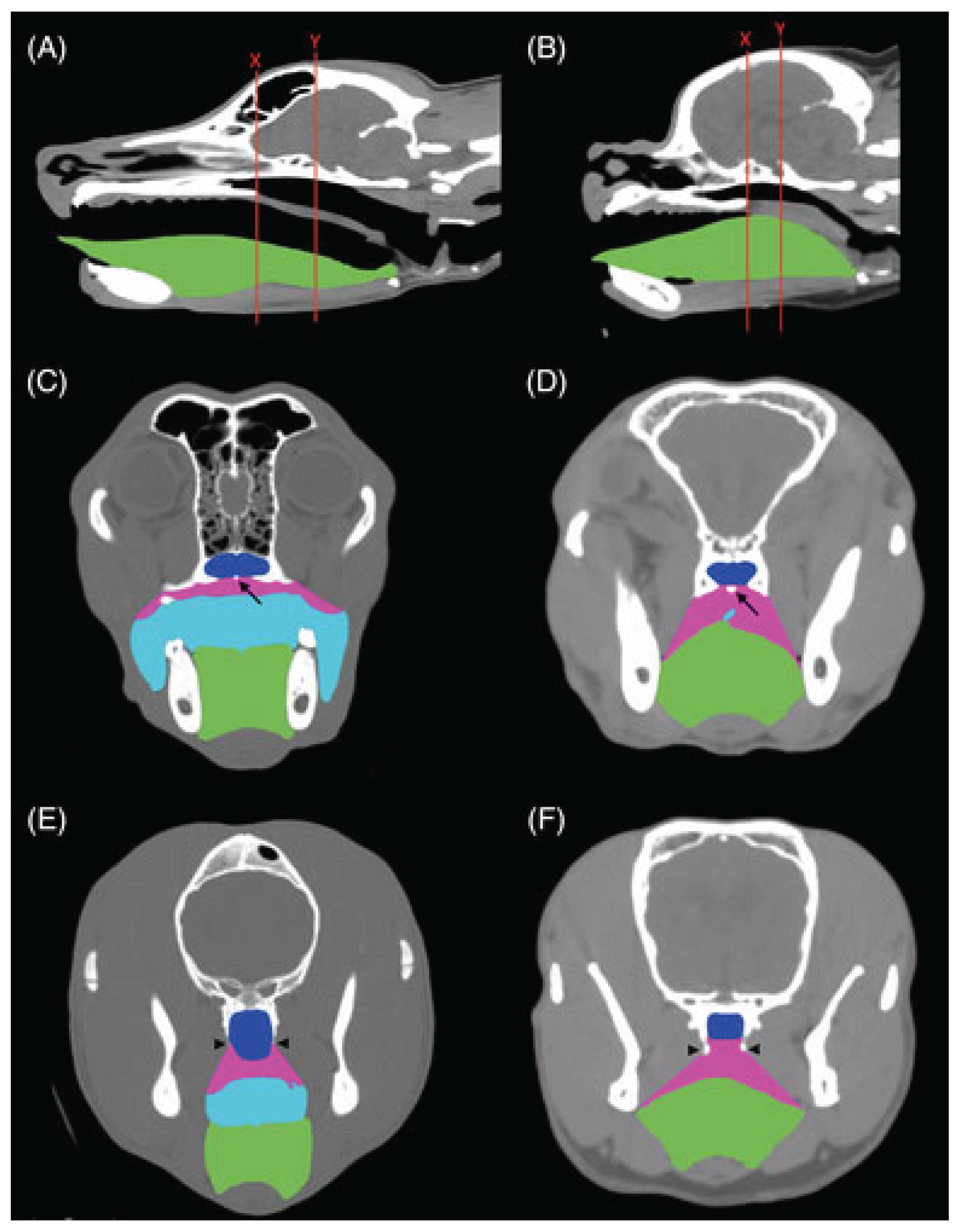
BOAS is a complex set of changes and an individual dog may be diagnosed with one or more of them. Breed differences have been recognized, and appear to support the theory that reduced airway space further towards the nostrils (rostral) affects function more than reduced space further down the airway (caudal) [119].
3.2. Brain and Nervous System Abnormalities
The changes in skull shape associated with brachycephaly predispose many toy and brachycephalic breeds to disorders of cerebrospinal fluid [CSF] flow, Chiari-type malformation [CM], and syringomyelia (SM) [120,121,122,123]. CM is characterized by crowding of the brain and cervical spinal cord and is often associated with behavior indicating pain [121,122,124]. SM is closely associated with CM and, in many breeds, rarely seen in its absence [120]. It is defined as the presence of a cavitation (syrinx) in the spinal cord [121,122,123]. Dogs with a syrinx may be asymptomatic, but commonly present with signs including pain, reduced activity, phantom scratching, scoliosis, ataxia, and weakness [123,125,126]. In Chihuahuas, open cranial sutures (persistent fontanelles), commonly considered of no clinical consequence, are associated with SM [127]. SM has significant welfare consequences, as affected dogs commonly are treated with analgesics, and may be euthanized due to quality of life issues [123,126,128,129]. SM is often progressive due to compensation for blocked CSF flow [122,126,130,131]. Both CM and SM are strongly associated with a shortened, domed skull shape and tighter angle between the nasal and frontal bone [124,132]; CM is almost fixed in some breeds with distinctively domed heads [Griffon Brussels 80% [121], Cavalier King Charles Spaniel (CKCS) 91-100% [120,131,133,134]]. SM is a more complex disorder but also at high prevalence in breeds with the risk head shape (e.g., CKCS 70-74% [133,135]). True prevalence of SM may be higher than reported as diagnosis requires MRI, an expensive and invasive test, and due to its progressive nature and potentially late onset [129,131,136]. While symptomatic SM is highly heritable [130,137], attempts to breed away from it have resulted in only minimal reduction of prevalence [138], perhaps due to the challenges of MRI-based screening, the disorder’s causal association with a head shape that is considered desirable, and because of low variation of that head shape in the affected breeds [124,132,137,138].
Additionally, the brachycephalic phenotype may predispose to an increased risk of structural brain lesions associated with epilepsy [139].
3.3. Eye Disease
A short muzzle and compressed skull shape predispose brachycephalic dogs to eye disease, loss of the globe, and blindness, as reviewed in [140]. Corneal irritation and increased risk of ulceration arise from shallow orbits resulting in protrusion of the globe; nasal folds lead to caruncular trichiasis (hair rubbing on the surface of the eye); macroblepharon (abnormally large opening between the eyelids, resulting in decreased protection of the eye); decreased corneal innervation leading to decreased sensitivity; and lagophthalmos (incomplete blinking due to inability of the eyelid to cover the eye). The recurrent irritation and ulceration of the cornea increases risk for ulcerative keratitis and corneal pigmentation, resulting in potential loss of vision and of the globe itself. Shallow eye orbits also increase the risk of traumatic proptosis (the eye being displaced out of the orbit). Other increased risks include keratoconjunctivitis sicca, which also carries risk of blindness and loss of the eye, and prolapse of the nictitating gland of the third eyelid [140]. Exact risk differs by breed [141]. Increased risk for corneal ulcerative disease in brachycephalic compared to mesocephalic dogs has been calculated at 11.18 [142], but this may be an underrepresentation of the true prevalence of the disease in flat-faced dogs, as some forms of ulcerative disease may be challenging for primary care veterinarians to diagnose [141]. Constant irritation of the eye is painful, as are corneal ulcers, and ulcerative disease can contribute to euthanasia decisions [142].
Selective breeding to more moderate conformation can reduce these risks. Dogs with nasal folds may be at 5 times the risk of corneal ulcers compared to those without, and dogs with a craniofacial ratio < 0.5 are at 20 times the risk [143]. A 10% increase in the angle of the eyelid aperture triples the risk of ulcers, as does exposed eye white [144].
3.4. Ear Infections
Ear infections (otitis externa and otitis media) are a common health concern in brachycephalic breeds - the most common diagnosis in primary care for bulldogs [145] and third most common for pugs [146]. The brachycephalic head shape is associated with narrowing of the canal in the outer ear [147,148] which may result in drainage issues predisposing to infection [147]. Middle ear effusion is seen in brachycephalic dogs even when asymptomatic, again suggestive of drainage issues predisposing to infection [148,149].
3.5. Developmental Abnormalities of the Skull
Brachycephalic dogs are overrepresented in the population of dogs presenting with cleft lip or palate (CL/CP) [150,151,152]. While environmental influences and inbreeding likely contribute to this predisposition in addition to conformation [150,151], the developmental origins of CL/CP are suggestive of a link to brachycephalic conformation. Specifically, the hard and soft palate develop from a complex series of fusions of tissues from different developmental origins; this series is initiated by fusion of nasal and maxillary processes [153]. Moreover, CL/CP dogs frequently have other craniomaxillofacial abnormalities visible on CT [154], indicating that these abnormalities may be associated with other developmental abnormalities that are more difficult to recognize. Notably, while brachycephalics are overrepresented among CL/CP diagnoses, dolichocephalics appear to rarely if ever suffer from this condition [151,152]. CL/CP is a welfare issue, as it may be commonly associated with neonatal euthanasia [154], surgery is the only definitive treatment [152], CL/CP surgery is complex and often requires multiple attempts for resolution [155], and surgery will not be curative in 15% of dogs [152].
3.6. Labor Difficulties
A mismatch between maternal pelvic opening and fetal head size is a common cause of labor difficulty (dystocia) [156,157,158]. Breeds with typically large heads, such as the English bulldog, Boston terrier, and French bulldog, are overrepresented in dystocia cases [83,156,157]. The true prevalence of dystocia among these breeds is difficult to assess, as most owners elect for Caesarean section. C-section rates in these breeds may be over 80% [156]; one report estimated 95% of English bulldog births were via C-section, of which 90% were elective [157]. Rates of dystocia in these three breeds are so high that some insurance companies will not insure against it [159].
4. Dwarfism
4.1. Chondrodysplasia
Canine “dwarfism” is caused by marked shortening of the limbs [160,161]. An fgf4 retrogene in chromosome 18 results in chondrodysplasia, the most significant shortening of leg length [160,162], due to a disorder of cartilage cells in the growth plates [163]. While the limb shortening itself does not appear to be associated with welfare issues, in some breeds it is closely related to forelimb curvature, or angular limb deformity (ALD) [164], and the mutation has been linked to increased risk of rotation of the ulna (ulnar valgus) [160]. Chondrodysplasia increases the risk of complex ALD [165]. The retardation of ulnar growth due to the mutation can result in compensatory bowing of the radius to compensate, as well as a turned out carpus [valgus], both of which issues increase the risk of elbow joint disease at a much younger age than is expected in dogs with straight legs [166,167,168,169,170]. Shorter and more deformed the limbs are associated with greater joint space between the humerus and ulna and a greater risk of lameness [171,172].
Prevalence of this issue is challenging to assess, as it is considered normal for many breeds and therefore underreported [171]. For example, an AKC breed description notes that "Dachshunds aren't built for distance running, leaping, or strenuous swimming" [173]. This issue was recently introduced as one with significant welfare repercussions which is deserving of more study [164]. While the connection between chondrodysplasia and leg bowing appears not yet fully understood, bowed legs [or “crook’d legs”, or “wrap-around front”] and splayed carpi are explicitly called for in the conformation standards for the Skye terrier, dachshund, and Cardigan Welsh corgi, and in addenda to the Basset hound standard [174,175,176], and a guide to judging Bassets notes that “Bassets with straight front legs are called ‘wide’ in front. A wide front is incorrect” [177]. Therefore, it is possible that the fgf4 insertion on chromosome 18 works in concert with another mutation to predispose to ALD, which has been selected for in some breeds but not others (for example, compare the bowed limbs of the Cardigan Welsh corgi with the straight limbs of the Pembroke Welsh corgi). More research into this relationship would be valuable.
4.2. Chondrodystrophy
An fgf4 retrogene in chromosome 12 results in chondrodystrophy, another phenotype characterized by shortened leg length [178]. While this mutation contributes significantly less to leg length reduction than does chondrodysplasia, it does provide additive reduction with that mutation, and is therefore at high frequency in many short-legged breeds [160]. It has additionally been found in many breeds not considered short-legged - as many as 40 of 75 common breeds [179] - potentially due to some other phenotypic effects. Like its relative on chromosome 12, this is a gain of function mutation and therefore dominant [178].
Dogs with this mutation, in addition to mild decrease in leg length, have increased risk of intervertebral disc herniation [178,179]. Disc herniation, commonly referred to as intervertebral disc disease, is classified into type I [seen in young, chondrodystrophic dogs] and type II [seen in older, non-chondrodystrophic dogs] [180,181,182]. Disc degeneration occurs with age, but chondrodystrophic dogs have degeneration in nearly 100% of their discs [183]. Degeneration is seen in newborn chondrodystrophic dogs and progresses quickly, with herniation observed in chondrodystrophic dogs as young as two years of age [184]. Herniation risk is polygenic and affected by environment [185,186] as well as breed-specific genetic architecture [187]; however, it is almost always higher in dogs with the mutation [160].
Disc herniation is a significant welfare issue. It is so often acutely painful that back pain causing the dog to “scream” is strongly suggestive of this disorder; the majority of dogs are unable to walk at time of diagnosis, and 60% may be treated surgically [186] (Packer 2016). Some animals are euthanized after diagnosis or failed treatment, and even those who do well are reported to be reluctant to perform everyday activities like getting on furniture [188,189]. Chondrodystrophic dogs may have new herniations at different sites subsequent to their initial diagnosis [189,190]. At the time of diagnosis, quality of life scores for the dog average 43-47%, and owner coping scores 3/10 [191].
Disc herniation is at highest prevalence in dachshunds but varies by breed type (14-33%) [179,180,185,186,188,191,192]. Two other chondrodystrophic breeds with elevated risk are the beagle (6%) and cocker spaniel [5-6%] [180]. Other, unidentified alleles appear to contribute to variations in breed risk [179,186,187], but without breeding away from the fgf4 chromosome 12 mutation, selective breeding can have only limited effect. The risk of disc herniation is normalized in dachshunds, which are frequently presented as dogs who should simply not be allowed to do things like jump on furniture or run down stairs [173]. The risk allele could be selectively bred out of populations with moderate to low allele frequencies. However, the question of how to remove it from breeds with high allele frequency (dachshund 94%, beagle 99%, cocker 91%) [187,193] is thorny, despite the fact that the mutation’s contribution to extreme short legs is mild to moderate.
5. Chest Depth
The ratio of thoracic width to depth is associated with risk of gastric dilatation and volvulus (GDV) [194,195,196,197,198]. GDV, commonly referred to as “bloat,” is the dilation and twist of the stomach, associated with significant morbidity and mortality [199,200]. While it can be seen in any dog, it is more common in large dogs (6% incidence [201]). A “deep chest” may result in a large abdominal cavity, allowing a very full stomach to stretch the hepatogastric ligament, which in turn allows the stomach to twist and cut off blood flow [198]. The risk of GDV is a complex trait, associated with many genes and environmental influences [196,198]. However, the risk is clearly higher in dogs with deeper chests; from the deepest thorax to the shallowest, the odds ratio for GDV may be 8.45 [197]. The most commonly affected breeds are German Shepherd Dogs and Great Danes [198,202]. GDV is an extremely serious diagnosis. Some affected dogs are dead by the time they reach veterinary care; many are euthanized rather than treated due to poor prognosis or financial concerns; 80-90% will survive surgery [202,203]. To mitigate this innate risk, owners may opt for preventative gastropexy, in which the stomach is sutured to the abdominal wall. Selective breeding away from the deep chest phenotype may be effective, but the German Shepherd Dog breed standard mandates a chest that “is well filled and carried well down between the legs. It is deep and capacious, never shallow” [204] and the Great Dane standard likewise mandates a “deep” chest [205].
6. Skin Folds
6.1. Facial Skin Folds in Brachycephalic Dogs
Many dogs of brachycephalic breeds have excessive skin on the face, resulting in skin folds. The skin between these folds is predisposed to skin fold dermatitis [intertrigo], or skin infections, due to the close proximity of areas of skin to each other [206,207]. For example, English bulldogs seen in primary care have 49 times the risk of developing intertrigo compared to crossbred dogs, French bulldogs 26 times the risk, and pugs 16 times the risk; these numbers may under-represent true risk factors due to underreporting of the issue, as abnormal skin may be difficult to visualize deep in folds [207]. Intertrigo may require surgical intervention to reduce the depth of folding [207]. Both owners and veterinarians may normalize excessive skin folding and struggle to recognize welfare issues associated with this morphology [145,208]. However, intertrigo causes itching, behavior change, and potentially pain, and has been named a welfare concern [206,209].
6.2. Entropion and Ectropion
Loose skin can affect ocular health, either when folds bring fur in contact with the globe (entropion) or when the lower lid everts away from the eye and fails to protect it (ectropion) [210]. Skin folds alone may obscure vision in entropion, but the contact of the fur with the globe can lead to more serious issues such as conjunctivitis, dry eye, and ulceration of the cornea, potentially eventually blindness, and may require surgical correction [211,212]. Entropion is the most common reported disorder in Shar-peis, with 18% of them affected, and is likely the root cause of the common eye problems in the breed [208], though it is seen in other breeds as well [208,211]. Eye tacking is a surgical procedure commonly used in puppies to prevent entropion and may be normalized among fanciers of this breed [213].
Inversely, when skin below the eye is too loose (ectropion), the lower eyelid cannot assist in maintenance of lubrication. Eventually the mucous tissue around the eye becomes inflamed, and chronic ocular disease may develop. Again, surgery is the treatment for severe ectropion [210]. Loose skin around the eyes is written into the Basset hound breed standard: “tight skin” is a fault, and the eyes should show a “prominent haw,” or third eyelid [176,210]. Neapolitan Mastiffs suffer from both ectropion (24% of dogs) and entropion (another 24% of dogs); 92% of dogs in this breed suffer from ophthalmologic disorders, the majority of which are related to their skin folds [214].
6.3. Shar pei Skin Folds
Two genetic mutations have been identified in association with the deep skin folds of the Shar pei. The folding and mutations are linked to Shar pei fever [a disease specific to the breed characterized by periodic bouts of fever and inflammation] and ongoing chronic inflammation [215], as well as intertrigo and ear infections [208]. This folding is considered a welfare concern in the breed [208], but the Shar Pei breed standard specifies “profuse wrinkles on the forehead continuing into side wrinkles framing the face” [216].
7. Tail Length and Curliness
7.1. Neurological Disorders
The association between tightly curled tails and neurological disorders is complex. Dogs with tightly curled tails have a high incidence of congenital vertebral malformations or CVMs [217,218,219,220]. The specific type and location of CVM, as well as type of associated neurological disorder risk, is highly breed specific [218,220]. While all tightly curled tails have historically been referred to as “screw tails,” the 2018 discovery of the DISHEVELLED 2 (DVL2) mutation clarified that the “screw tail” terminology is most appropriate for breeds such as English and French bulldogs, whose tails are both tightly curled and extremely short; the pug tail phenotype, tightly curled but normal length, depends on a different and not yet identified mutation [219,221,222]. Additional genetic architecture may be associated with the screw-tail phenotype in bulldogs, as DVL2 does not have as strong an effect on tail phenotype in other breeds; DVL2 may also affect the brachycephaly phenotype [222], but more research is called for to assess this relationship.
In the case of both pugs and dogs of true screw-tailed breeds, CVM prevalence is extremely high, with percentages in neurologically normal French bulldogs, English bulldogs, and/or Boston terriers reported from around 50% [219,223] to around 80% [218,219,224]. CVM prevalence in pugs may be close to 100% [219]. As many dogs with CVMs are neurologically normal, veterinary opinion has historically been that the malformations are benign [218]. While pathology directly caused by CVMs appears rare, CVMs are associated with and may in fact cause spinal curvature (kyphosis) [217,224]. More extreme angles of kyphosis [over 35 degrees] are associated with neurologic deficits [225], and these deficits may be severe enough to result in non-ambulatory dogs from birth or as young as a year of age; treatment is spinal surgery which is not always successful [226,227]. Milder kyphosis is associated with spinal instability, which may increase the risk of disc herniation above or below the region of curvature [228,229] and predispose to discospondylitis, or infection of the intervertebral discs and surrounding structures [230]. French Bulldogs are particularly affected by this process; while their rates of disc herniation are somewhat lower than that of dachshunds, the average French Bulldogs herniation is on average two years earlier than the average dachshund herniation [228,231]. French Bulldogs may be at a disadvantage due to the combined effects of high prevalence of CVMs due to their screw tails, and high prevalence [91%] of the chromosome 12 FGF4 retrogene allele (associated with chondrodystrophy) [193]. Because of these associations, authors have suggested that selective breeding away from the screw-tail phenotype would be in the dogs’ best interest [232].
7.2. Skin Folds Around the Tail in Screw-Tailed Dogs
7.3. Communication
Dogs use their tails to communicate both with other dogs and with humans; emotional states and intentions conveyed by tail position and motion include threat vs appeasement or friendliness, fear or anxiety vs confidence, and excitability [235]. A comparison of canine responses to robot dogs with long versus short tails showed that dogs responded differently to long tails when they were wagging versus when they were still, but did not differentiate wagging versus still short tails - suggesting that dogs do get more information from the movements of longer tails [236]. Dogs with short tails may therefore have greater difficulty communicating than dogs with normal length tails, and additional research would be beneficial to better understand this relationship.
8. Coat
8.1. Ridge
The Rhodesian Ridgeback breed standard specifies a “ridge” along the dog’s spine, which is a line of hair growing in the opposite direction to the main coat. A genome-wide association study identified the causative mutation for the ridge, a 133-kb duplication including three fibroblast growth factor genes, and also linked increasing copy number of the duplication with increasing risk of dermoid sinus [237,238]. Dermoid sinus (DS) is a developmental disorder resulting in a sinus opening running from the skin along the backbone, potentially as deep as the dura mater around the spinal cord [239]. DS can introduce infection past the skin barrier; surgical excision is the treatment of choice but may not always be curative [240,241]. DS is prevalent in Rhodesian Ridgebacks, occuring in around 5% of dogs, and breeders actively select away from it [242,243]. However, breeding away from DS is complicated by its association with the preferred trait of having a ridge. While the mutation causing the ridge phenotype may not entirely explain the risk for DS, the risk clearly increases with increasing copy numbers of the duplicated locus [237]. Therefore, producing heterozygous dogs (who express the ridge phenotype and have decreased DS risk) may be the ideal solution, but as this involves using ridgeless dogs for breeding, there has been resistance in the breeding community [242,243].
8.2. Hairlessness
Hairlessness in the Chinese Crested (CC) is due to a mutation in FOXI3 and results in the canine ectodermal dysplasia (CED) phenotype [244]. CED is characterized by dysplastic hair follicles and numerous comedones (acne) [245]. Additionally, it is associated with a variable number of malformed, malpositioned, undersized, or entirely missing teeth [246].
Figure 4.
3D renderings of skulls from (a) coated and (b) hairless dog [246]. Note the significant lack of teeth in the hairless dog.
Figure 4.
3D renderings of skulls from (a) coated and (b) hairless dog [246]. Note the significant lack of teeth in the hairless dog.
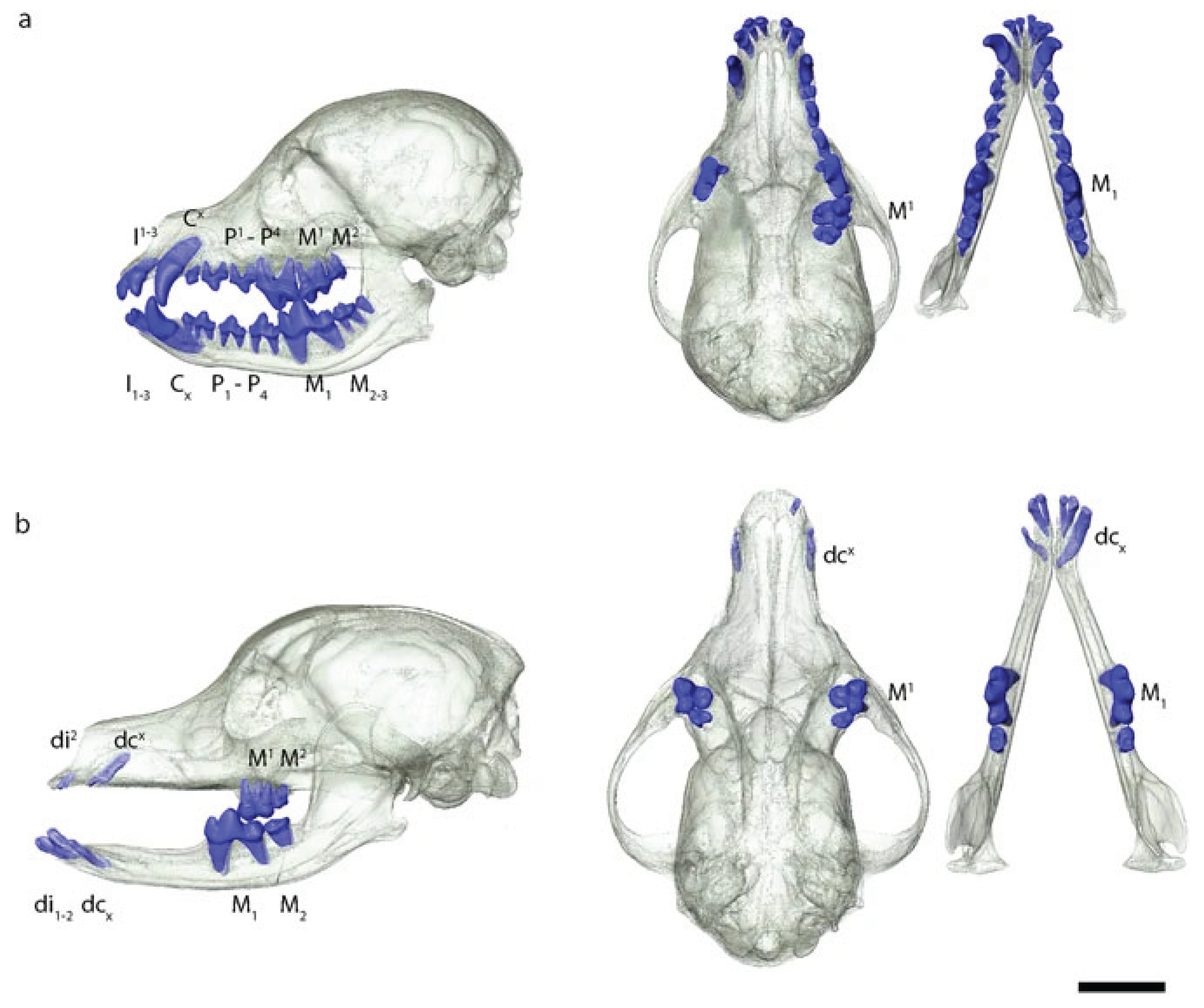
8.3. Color and Pattern
A variety of coat colors and patterns are associated with disorders in the dog. Extensive white markings are associated with congenital deafness in multiple species, including in over 90 breeds of dogs [247], and are likely a problem in any breed with white coat color due to melanocyte suppression [248]. This association has been studied the most extensively in the Dalmatian, in which extreme white markings covering the entire body give the appearance of an all-white dog [the spots are controlled by a separate mutation]. A mutation in the MITF gene is associated with white markings, blue eyes, and deafness [249,250] due to a lack of cochlear melanocytes and resulting degeneration of cochlear hair cells and nerves [251]. In Dalmatians, black patches on the ears are correlated with reduced risk of deafness [251]. Nevertheless, the AKC Dalmatian breed standard notes that patches are disqualifying [252]. About 22% of Dalmatians are unilaterally deaf, and about 8% bilaterally [and therefore clinically] deaf [251].
The Dalmatian coat is distinctive not just for its extreme white, but also for its spots. These appear to be controlled by the gene for roaning in conjunction with another, not yet identified gene to decrease the “frosted” look of a roan pattern [247,253,254]. This second, unidentified mutation is located close to (in linkage with) a mutation in SLC2A9, or may be the same mutation. The SLC2A9 mutation disrupts uric acid transport; as a result the Dalmatian breed, prior to an outcross to Pointer, was fixed for this mutation, resulting in a high risk of urinary stones [255]. The Pointer outcross produced a population of Dalmatians heterozygous or homozygous for the wild-type allele, with normal uric acid transport. However, use of these dogs is resisted by some breeders as their spots are perceived as often “frosted” [256], potentially a sign that the unidentified mutation that works epistatically with roaning to produce spots is not present, or present in altered form, in this population. Better characterization of the mutation [s] controlling Dalmatian spots may provide valuable tools to the breeding community.
Merle is another dramatic coat pattern selected for in many breeds. Heterozygous merle coats display patchy areas of dilute color; homozygous merle coats are mostly white with patches of dilute. The merle allele is a variable-length SINE insertion in the SILV gene [257,258,259]. Homozygous merles are at high risk of deafness [unilateral or bilateral], blindness, microphthalmia, and sterility; heterozygous merles are also at risk, though it is reduced compared to homozygotes [251,260]. Nevertheless, breeding recommendations focus on avoiding homozygous merles rather than moving away from the pattern entirely [247,259]. Meanwhile, merle is being introduced to new breeds, such as the Poodle and French Bulldog.
Several variants in the MLPH gene cause coat color to be “diluted,” for example from black to “blue” or brown to “isabella” or “fawn” [244,261,262,263]. Dilution has long been associated with color dilution alopecia (CDA) [264]. One of the MLPH variants (d1) is clearly associated with CDA in various breeds [265], while potential association of the other two variants (d2, d3) has yet to be investigated [247]. CDA is a dysfunction of the hair follicle, characterized by clumping of melanin and progressive hair loss [265,266]; the skin may become itchy and infected [264]. The association between dilute color and CDA varies by breed, suggesting additional genetic architecture modifying the phenotype [265]. For example, it is less common and the phenotype less extreme in Weimaraners [267], a breed in which the d1 variant is fixed. In other breeds, the association may be quite strong, for example, as high as 60% prevalence in dilute Dobermans [264]. Nevertheless, dilution variants have been recently introduced into some breeds, such as Labrador Retrievers [268], and are becoming increasingly popular in others, such as French bulldogs [269].
9. Discussion
Many common morphologies in artificially selected dogs have well-documented negative impacts on their health and welfare [270]. In this Perspective, we have summarized major health outcomes of common morphological traits under artificial selection. Additional traits were not included here due to space limitations, for example the association of pendulous ears with ear infections [271,272]; meanwhile, other traits are anecdotally linked to negative health impacts but it appears that the supporting research has not yet been done, for example, the black tongue of the Chow Chow and increased risk of oral melanoma. These negative impacts raise an important question – what does a physically sound dog look like? Free-living village dogs may offer some insight. Unlike selectively bred dogs, village dogs evolve under natural selection, which favors phenotypes that promote survival and reproduction. Consequently, village dogs may demonstrate a range of physical characteristics that are associated with robust innate health. For example, most village dogs are medium-sized (11-20 kg) irrespective of their geographic origin [273,274,275,276,277], suggesting that this might be an optimal body size for dogs, at least in the geo-climatic regions in which most sampling has occurred to date. However, purpose-bred dogs and village dogs generally live in different environments, so the same physical traits may not necessarily promote health in all populations. This is an important topic for further study.
The exaggerated morphologies observed in some purpose-bred dogs, but not village dogs, are the product of modern selective breeding practices based on aesthetic preference [278]. This is exemplified by the French Bulldog, the most popular breed in the United States in 2024 [279], which exhibits several extreme traits associated with adverse health outcomes, such as airway obstruction, gastrointestinal issues, labor difficulties, skin fold infections, and vertebral disc herniation [207,228,280]. The popularity of the French Bulldog has not only increased the prevalence of extreme phenotypes, but has also helped normalize them [103]. These morphologies are frequently associated with physical discomfort and reduced functionality, and their impacts can extend beyond the specific morphology itself. For example, physical discomfort from conditions like chronic pain or severe itching has been linked to behavioral changes, including increased aggression and decreased trainability [28,281].
The accumulated evidence presented in this Perspective strongly supports the conclusion that artificial selection based on physical appearance can have substantial and often detrimental impacts on the health and welfare of purebred dogs. Many current breeding practices permit the continued propagation of harmful traits rather than actively altering breeding regimes to lessen them. To mitigate these issues, there is a pressing need to reconsider how we approach breeding practices and breed standards - as well as how to mitigate consumer demand for “exotic” dogs with extreme phenotypes. This consumer demand drives the introduction of extreme phenotypes into new breeds (such as merle coloring or hairlessness into French Bulldogs), or the further exaggeration of existing phenotypes (such as “big rope” French Bulldogs with even more extreme wrinkling).
Future directions in this field could include welfare epidemiology research to better understand the relationship between morphology, health, and behavior across different dog populations. Efforts to prioritize innate health, such as those outlined by organizations like the Brachycephalic Working Group [282], offer a promising path forward. Defining core health attributes for dogs, such as functional respiratory anatomy and sound musculoskeletal structure, could guide breeding toward healthier populations - work which depends on evidence-based assessments of when morphologies become “extreme” and thereby detrimental. Alternatives to traditional breeding practices should also be considered. These might include adjusting breed standards to prioritize health and functionality, promoting the value of mixed-breed dogs, and supporting alternative registries that emphasize both physical and behavioral wellbeing [21]. Change is possible, as demonstrated by recent adjustments to breed standards in the United Kingdom aimed at reducing harmful traits [283]. By shifting the focus from appearance to health, and by evaluating which morphologies negatively affect health, we can take meaningful steps to improve the lives of dogs [284].
10. Conclusions
In this review, we have summarized the impacts of artificially selected morphological traits on dogs’ wellbeing. It is clear that artificial selection to suit arbitrary human aesthetic preferences can result in poor outcomes for dogs’ physical and behavioral health. Notably, many of these breeding practices do not include selection for the niche in which dogs now live - inside human households - which is very different from their niche even 100 years ago.
While lineages of dogs have differed in morphological phenotype for many hundreds of years, strong, intensive, intentional selection for traits related to appearance began in earnest during the Victorian era. At that time, the health consequences of selection on these traits were not well understood. Now they are, and we have a responsibility to the animals we breed to take their welfare into consideration.
In this Perspective, we have only skimmed the surface of each of the covered traits. Our aim is to create a basis for future work that can delve more deeply. Two fundamental needs are apparent: first, to better research and understand the mechanisms and etiologies of health outcomes of morphological selection; and second, to enact changes in breeding practices. If we can define clear guidelines for when extreme morphology becomes a welfare issue, this can support evidence-based efforts to set new breeding goals. How should we determine the boundary between morphologies that do and do not have substantial impairment on innate health? How should physiological and health data be gathered and interpreted to guide breeding practices? How can members of the general public be informed about the ethical consequences of their financial support for particular selection regimes?
The researchers listed below have added their names to those of the article’s authors in calling for the scientific and veterinary communities - and the general public - to consider these issues.
Carlos E. Alvarez, PhD
Departments of Pediatrics and Veterinary Clinical Sciences, The Ohio State University Colleges of Medicine and Veterinary Medicine
Marc Bekoff, PhD
Professor Emeritus of Ecology and Evolutionary Biology, University of Colorado, Boulder
Mia Cobb, PhD
Chaser Innovation Research Fellow, Animal Welfare Science Centre, The University of Melbourne, Australia
Craig Daniell
Chief Executive Officer, British Columbia Society for the Prevention of Cruelty to Animals
Nicholas H. Dodman, BVMS, DACVA, DACVB
Professor Emeritus, Cummings School of Veterinary Medicine, Tufts University
Hille Fieten, DVM, PhD, Diplomate ECVIM-CA internal medicine, MSc genetic epidemiology
Expertise Centre of Veterinary Genetics, Faculty of Veterinary Medicine, Utrecht University, the Netherlands
Simon Gadbois, PhD
Ethology and Canine Olfaction Lab, Dalhousie University
Temple Grandin, PhD
Distinguished Professor, Department of Animal Science, Colorado State University
Åke Hedhammar, DVM, PhD
Hereditary Disease Committee Member, World Small Animal Veterinary Association
Professor Emeritus in Internal Medicine, Swedish University of Agricultural Sciences
Alexandra Horowitz, PhD
Senior Research Fellow, Psychology, Barnard College
Katherine Houpt VMD PhD DACVB
Emeritus Professor of Veterinary Behavior, Cornell University College of Veterinary Medicine
Ana G. Jiménez, PhD
Vice-Chair of the Canine Science Society
Associate Professor in Biology, Colgate University
Juliane Kaminski, PhD
Associate Professor, Director Dog Cognition Centre, Centre for Comparative and Evolutionary Psychology, University of Portsmouth
Susan Penelope Knowler, BSc, PGDE, PhD
Visiting Researcher, School of Veterinary Medicine, University of Surrey, UK
Anna Kukekova, PhD
Associate Professor, Department of Animal Sciences, University of Illinois Urbana-Champaign
Evan MacLean, PhD
Associate Professor, College of Veterinary Medicine, University of Arizona
Monique Megens, DVM
Chair, International Collaborative on Extreme Conformations In Dogs [ICECDogs]
Ádám Miklósi, PhD
Department of Ethology, Eötvös Loránd University, Budapest, Hungary
Daniel S. Mills BVSc PhD CBiol FRSB FHEA CCAB Dip ECAWBM[BM] FRCVS
RCVS and EBVS European Veterinary Specialist in Behavioural Medicine
Emma Milne, BVSc, FRCVS
Founder of Vets Against Brachycephalism
Siobhan Mullan, DVM, PhD
UCD, Chair of the Brachycephalic sub-group of the Advisory Council on Companion Animal Welfare, Ireland
Dan O'Neill, MVB, BSc (hons), GPCert (SAP), GPCert (FelP), GPCert (Derm), GPCert (B&PS), PGCertVetEd, FHEA, MSc (VetEpi), PhD, FRCVS
Associate Professor in Companion Animal Epidemiology, Royal Veterinary College, London, UK
Alexandra Protopopova, PhD, CAAB
Associate Professor, Animal Welfare Program, Faculty of Land and Food Systems, The University of British Columbia
Rodrigo Gutierrez Quintana, MVZ, MVM, DipECVN, MRCVS
Professor of Comparative Neurology, School of Biodiversity, One Health and Veterinary Medicine, University of Glasgow
Rick F. Sanchez DVM, DipECVO, CertVetEd / FHEA, Diplomate of the European College of Veterinary Ophthalmology
Specialistische Dierenkliniek Utrecht (SDU-AniCUra)
Peter Sandøe, DPhil
Professor of Bioethics at the University of Copenhagen and Director of the Danish Centre for Companion Animal Welfare
Lionel Sebbag, DVM, PhD, DACVO
Veterinary Ophthalmologist, Expert in Brachycephalic Ocular Syndrome
James Serpell, PhD
Emeritus Professor of Animal Welfare, School of Veterinary Medicine, Dept of Clinical Sciences & Advanced Medicine, University of Pennsylvania
Rosemary Strasser, PhD
Director of the Neuroscience & Behavior Graduate Program, University of Nebraska Omaha
Silvan R. Urfer, Dr. Med. Vet.
Dog Aging Project, Seattle, WA
Carolyn Walsh, PhD
Associate Professor, Canine Research Unit, Psychology Department, Memorial University of Newfoundland and Labrador, Canada
C. D. L. Wynne, PhD
Professor, Department of Psychology, Arizona State University
Isain Zapata, PhD
Associate Professor, Department of Biomedical Sciences, Rocky Vista University
Conflicts of Interest
The authors declare no conflicts of interest.
References
- A. L. Shearin, E. A. Ostrander, Canine morphology: hunting for genes and tracking mutations. PLoS Biol 8, e1000310 (2010). [CrossRef]
- P. W. Hedrick, L. Andersson, Are dogs genetically special? Heredity (Edinb) 106, 712–713 (2011). [CrossRef]
- A. H. Freedman, et al., Genome sequencing highlights the dynamic early history of dogs. PLoS Genet 10, e1004016 (2014). [CrossRef]
- L. R. Botigué, et al., Ancient European dog genomes reveal continuity since the Early Neolithic. Nat Commun 8, 16082 (2017). [CrossRef]
- A. R. Perri, et al., Dog domestication and the dual dispersal of people and dogs into the Americas. Proc Natl Acad Sci U S A 118 (2021). [CrossRef]
- P. Skoglund, E. Ersmark, E. Palkopoulou, L. Dalén, Ancient wolf genome reveals an early divergence of domestic dog ancestors and admixture into high-latitude breeds. Curr Biol 25, 1515–1519 (2015). [CrossRef]
- Z. Fan, et al., Worldwide patterns of genomic variation and admixture in gray wolves. Genome Res 26, 163–173 (2016). [CrossRef]
- A. Bergström, et al., Origins and genetic legacy of prehistoric dogs. Science 370, 557–564 (2020). [CrossRef]
- A. R. Boyko, et al., Complex population structure in African village dogs and its implications for inferring dog domestication history. Proc Natl Acad Sci U S A 106, 13903–13908 (2009). [CrossRef]
- D. Brewer, “Hunting, animal husbandry and diet in ancient Egypt” in A History of the Animal World in the Ancient Near East, (Brill, 2002), pp. 425–456.
- F. Hole, C. Wyllie, The Oldest Depictions of Canines and a Possible Early Breed of Dog in Iran. Paléorient 33, 175–185 (2007). [CrossRef]
- S. H. Lonsdale, Attitudes Towards Animals in Ancient Greece1. Greece Rome 26, 146–159 (1979). [CrossRef]
- A. T. Lin, et al., The history of Coast Salish “woolly dogs” revealed by ancient genomics and Indigenous Knowledge. Science 382, 1303–1308 (2023). [CrossRef]
- C. Jung, D. Pörtl, How old are (Pet) Dog Breeds? Pet Behav. Sci. 29–37 (2019). [CrossRef]
- B. M. vonHoldt, et al., Genome-wide SNP and haplotype analyses reveal a rich history underlying dog domestication. Nature 464, 898–902 (2010). [CrossRef]
- A. Tonoike, et al., Comparison of owner-reported behavioral characteristics among genetically clustered breeds of dog (Canis familiaris). Sci. Rep. 5, 17710 (2015). [CrossRef]
- C. Ameen, et al., Specialized sledge dogs accompanied Inuit dispersal across the North American Arctic. Proc. Biol. Sci. 286, 20191929 (2019). [CrossRef]
- G. Larson, et al., Rethinking dog domestication by integrating genetics, archeology, and biogeography. Proc. Natl. Acad. Sci. U. S. A. 109, 8878–8883 (2012). [CrossRef]
- M. E. Derry, Made to Order: The Designing of Animals (University of Toronto Press, 2022).
- M. Worboys, J.-M. Strange, N. Pemberton, The invention of the modern dog: breed and blood in Victorian Britain (Johns Hopkins University Press, 2018).
- H. F. Proschowsky, et al., A new future for dog breeding. Anim. Welf. 34, e1 (2025). [CrossRef]
- C. Brassard, et al., Unexpected morphological diversity in ancient dogs compared to modern relatives. Proc. Biol. Sci. 289, 20220147 (2022). [CrossRef]
- A. Herzog, Torture breeding, definitions, judgement, pathogenesis. (1997).
- H. Howe, T. Katamine, Using the law to address harmful conformation in dogs-is a breed-specific breed ban the answer?
- Luxembourg, Law of June 27, 2018 on the protection of animals. (2018). Available at: https://legilux.public.lu/eli/etat/leg/loi/2018/06/27/a537/jo [Accessed 13 January 2025].
- Eurogroup for Animals, Landmark ruling against unethical dog breeding in Norway. (2023). Available at: https://www.eurogroupforanimals.org/news/landmark-ruling-against-unethical-dog-breeding-norway [Accessed 13 January 2025].
- W. Woods II, Ojai becomes first U.S. city to ban torture breeding. (2024). Available at: https://www.vcstar.com/story/news/local/california/2024/10/24/ojai-bans-torture-breeding-of-animals/75814787007/ [Accessed 13 January 2025].
- D. S. Mills, et al., Pain and problem behavior in cats and dogs. Animals (Basel) 10, 318 (2020). [CrossRef]
- N. B. Sutter, et al., A single IGF1 allele is a major determinant of small size in dogs. Science 316, 112–115 (2007). [CrossRef]
- B. C. Hoopes, M. Rimbault, D. Liebers, E. A. Ostrander, N. B. Sutter, The insulin-like growth factor 1 receptor (IGF1R) contributes to reduced size in dogs. Mamm. Genome 23, 780–790 (2012). [CrossRef]
- A. R. Boyko, et al., A simple genetic architecture underlies morphological variation in dogs. PLoS Biol. 8, e1000451 (2010). [CrossRef]
- M. Rimbault, et al., Derived variants at six genes explain nearly half of size reduction in dog breeds. Genome Res. 23, 1985–1995 (2013). [CrossRef]
- J. Plassais, et al., Whole genome sequencing of canids reveals genomic regions under selection and variants influencing morphology. Nat. Commun. 10, 1489 (2019). [CrossRef]
- K. Morrill, et al., Ancestry-inclusive dog genomics challenges popular breed stereotypes. Science 376, eabk0639 (2022). [CrossRef]
- M. A. Tryfonidou, et al., Hormonal regulation of calcium homeostasis in two breeds of dogs during growth at different rates. J. Anim. Sci. 81, 1568–1580 (2003). [CrossRef]
- J. K. Kirkwood, The influence of size on the biology of the dog. J. Small Anim. Pract. 26, 97–110 (1985). [CrossRef]
- J. da Silva, B. J. Cross, Dog life spans and the evolution of aging. Am. Nat. 201, E140–E152 (2023). [CrossRef]
- C. Kraus, N. Snyder-Mackler, D. E. L. Promislow, How size and genetic diversity shape lifespan across breeds of purebred dogs. GeroScience 45, 627–643 (2023). [CrossRef]
- C. Kraus, S. Pavard, D. E. L. Promislow, The size–life span trade-off decomposed: why large dogs die young. Am. Nat. 181, 492–505 (2013). [CrossRef]
- D. Bannasch, et al., The effect of inbreeding, body size and morphology on health in dog breeds. Canine Med. Genet. 8, 1–9 (2021). [CrossRef]
- J. P. de Magalhães, J. Costa, G. M. Church, An analysis of the relationship between metabolism, developmental schedules, and longevity using phylogenetic independent contrasts. J. Gerontol. A Biol. Sci. Med. Sci. 62, 149–160 (2007). [CrossRef]
- K. A. Greer, L. M. Hughes, M. M. Masternak, Connecting serum IGF-1, body size, and age in the domestic dog. Age (Dordr.) 33, 475–483 (2011). [CrossRef]
- D. E. Berryman, J. S. Christiansen, G. Johannsson, M. O. Thorner, J. J. Kopchick, Role of the GH/IGF-1 axis in lifespan and healthspan: lessons from animal models. Growth Horm. IGF Res. 18, 455–471 (2008). [CrossRef]
- J. M. Fleming, K. E. Creevy, D. E. L. Promislow, Mortality in North American dogs from 1984 to 2004: an investigation into age-, size-, and breed-related causes of death. J. Vet. Intern. Med. 25, 187–198 (2011). [CrossRef]
- S. M. Schwartz, et al., Lifetime prevalence of malignant and benign tumours in companion dogs: Cross-sectional analysis of Dog Aging Project baseline survey. Vet. Comp. Oncol. 20, 797–804 (2022). [CrossRef]
- Y. Nam, et al., Dog size and patterns of disease history across the canine age spectrum: Results from the dog aging project. PLoS One 19, e0295840 (2024). [CrossRef]
- L. Nunney, The effect of body size and inbreeding on cancer mortality in breeds of the domestic dog: a test of the multi-stage model of carcinogenesis. R. Soc. Open Sci. 11, 231356 (2024). [CrossRef]
- K. M. Makielski, et al., Risk factors for development of canine and human osteosarcoma: a comparative review. Vet. Sci. 6, 48 (2019). [CrossRef]
- T. T. Samaras, H. Elrick, L. H. Storms, Birthweight, rapid growth, cancer, and longevity: a review. J. Natl. Med. Assoc. 95, 1170 (2003).
- S. Simpson, et al., Comparative review of human and canine osteosarcoma: morphology, epidemiology, prognosis, treatment and genetics. Acta Vet. Scand. 59, 1–11 (2017). [CrossRef]
- D. G. O’Neill, et al., Dog breeds and conformations predisposed to osteosarcoma in the UK: a VetCompass study. Canine Med. Genet. 10, 8 (2023). [CrossRef]
- G. Ru, B. Terracini, L. T. Glickman, Host related risk factors for canine osteosarcoma. Vet. J. 156, 31–39 (1998). [CrossRef]
- I. Zapata, et al., Risk-modeling of dog osteosarcoma genome scans shows individuals with Mendelian-level polygenic risk are common. BMC Genomics 20, 1–14 (2019). [CrossRef]
- M. Harada, K. Akita, Mouse fibroblast growth factor 9 N143T mutation leads to wide chondrogenic condensation of long bones. Histochem. Cell Biol. 153, 215–223 (2020). [CrossRef]
- E. K. Karlsson, et al., Genome-wide analyses implicate 33 loci in heritable dog osteosarcoma, including regulatory variants near CDKN2A/B. Genome Biol. 14, 1–16 (2013). [CrossRef]
- Y. Jiao, Y. Feng, X. Wang, Regulation of tumor suppressor gene CDKN2A and encoded p16-INK4a protein by covalent modifications. Biochemistry (Mosc.) 83, 1289–1298 (2018). [CrossRef]
- R. A. Pettitt, A. J. German, Investigation and management of canine osteoarthritis. In Pract. 37, 1–8 (2015). [CrossRef]
- K. L. Anderson, et al., Prevalence, duration and risk factors for appendicular osteoarthritis in a UK dog population under primary veterinary care. Sci. Rep. 8, 5641 (2018). [CrossRef]
- K. L. Anderson, H. Zulch, D. G. O’Neill, R. L. Meeson, L. M. Collins, Risk factors for canine osteoarthritis and its predisposing arthropathies: a systematic review. Front. Vet. Sci. 7, 220 (2020). [CrossRef]
- D. G. O’Neill, R. L. Meeson, A. Sheridan, D. B. Church, D. C. Brodbelt, The epidemiology of patellar luxation in dogs attending primary-care veterinary practices in England. Canine Genet. Epidemiol. 3, 1–12 (2016). [CrossRef]
- M. D. King, Etiopathogenesis of canine hip dysplasia, prevalence, and genetics. Vet. Clin. North Am. Small Anim. Pract. 47, 753–767 (2017). [CrossRef]
- F. H. Comhaire, F. Snaps, Comparison of two canine registry databases on the prevalence of hip dysplasia by breed and the relationship of dysplasia with body weight and height. Am. J. Vet. Res. 69, 330–333 (2008). [CrossRef]
- M. Ginja, A. M. Silvestre, J. M. Gonzalo-Orden, A. J. A. Ferreira, Diagnosis, genetic control and preventive management of canine hip dysplasia: a review. Vet. J. 184, 269–276 (2010). [CrossRef]
- T. H. Witsberger, J. A. Villamil, L. G. Schultz, A. W. Hahn, J. L. Cook, Prevalence of and risk factors for hip dysplasia and cranial cruciate ligament deficiency in dogs. J. Am. Vet. Med. Assoc. 232, 1818–1824 (2008). [CrossRef]
- J. Michelsen, Canine elbow dysplasia: aetiopathogenesis and current treatment recommendations. Vet. J. 196, 12–19 (2013). [CrossRef]
- D. G. O’Neill, D. C. Brodbelt, R. Hodge, D. B. Church, R. L. Meeson, Epidemiology and clinical management of elbow joint disease in dogs under primary veterinary care in the UK. Canine Med. Genet. 7, 1–15 (2020). [CrossRef]
- G. Harasen, Canine cranial cruciate ligament rupture in profile: 2002–2007. Can. Vet. J. 49, 193 (2008). [CrossRef]
- J. M. Duval, S. C. Budsberg, G. L. Flo, J. L. Sammarco, Breed, sex, and body weight as risk factors for rupture of the cranial cruciate ligament in young dogs. J. Am. Vet. Med. Assoc. 215, 811–814 (1999). [CrossRef]
- E. J. Comerford, K. Smith, K. Hayashi, Update on the aetiopathogenesis of canine cranial cruciate ligament disease. Vet. Comp. Orthop. Traumatol. 24, 91–98 (2011). [CrossRef]
- G. W. Niebauer, B. Restucci, Etiopathogenesis of canine cruciate ligament disease: a scoping review. Animals (Basel) 13, 187 (2023). [CrossRef]
- H. G. Parker, P. Kilroy-Glynn, Myxomatous mitral valve disease in dogs: does size matter? J. Vet. Cardiol. 14, 19–29 (2012). [CrossRef]
- M. J. Mattin, et al., Degenerative mitral valve disease: Survival of dogs attending primary-care practice in England. Prev. Vet. Med. 122, 436–442 (2015). [CrossRef]
- A. Tidholm, J. Häggström, M. Borgarelli, A. Tarducci, Canine idiopathic dilated cardiomyopathy. Part I: aetiology, clinical characteristics, epidemiology and pathology. Vet. J. 162, 92–107 (2001). [CrossRef]
- M. W. S. Martin, M. J. Stafford Johnson, B. Celona, Canine dilated cardiomyopathy: a retrospective study of signalment, presentation and clinical findings in 369 cases. J. Small Anim. Pract. 50, 23–29 (2009). [CrossRef]
- D. G. O’Neill, et al., Epidemiology of periodontal disease in dogs in the UK primary-care veterinary setting. J. Small Anim. Pract. 62, 1051–1061 (2021). [CrossRef]
- C. Wallis, E. K. Saito, C. Salt, L. J. Holcombe, N. G. Desforges, Association of periodontal disease with breed size, breed, weight, and age in pure-bred client-owned dogs in the United States. Vet. J. 275, 105717 (2021). [CrossRef]
- C. Wallis, A. Ivanova, L. J. Holcombe, Persistent deciduous teeth: Association of prevalence with breed, breed size and body weight in pure-bred client-owned dogs in the United States. Res. Vet. Sci. 169, 105161 (2024). [CrossRef]
- M. A. Gioso, F. Shofer, P. S. M. Barros, C. E. Harvey, Mandible and mandibular first molar tooth measurements in dogs: relationship of radiographic height to body weight. J. Vet. Dent. 18, 65–68 (2001). [CrossRef]
- M. Kyllar, B. Doskarova, V. Paral, Morphometric assessment of periodontal tissues in relation to periodontal disease in dogs. J. Vet. Dent. 30, 146–149 (2013). [CrossRef]
- O. Idowu, K. Heading, Hypoglycemia in dogs: Causes, management, and diagnosis. Can. Vet. J. 59, 642 (2018).
- A. G. Jimenez, K. Paul, A. Zafar, A. Ay, Effect of different masses, ages, and coats on the thermoregulation of dogs before and after exercise across different seasons. Vet. Res. Commun. 47, 833–847 (2023). [CrossRef]
- E. J. Hall, A. J. Carter, D. G. O’Neill, Dogs don’t die just in hot cars—exertional heat-related illness (Heatstroke) is a greater threat to UK dogs. Animals (Basel) 10, 1324 (2020). [CrossRef]
- D. G. O’Neill, et al., Canine dystocia in 50 UK first-opinion emergency-care veterinary practices: prevalence and risk factors. Vet. Rec. 181, 88–88 (2017). [CrossRef]
- A. Münnich, U. Küchenmeister, Dystocia in numbers–evidence-based parameters for intervention in the dog: causes for dystocia and treatment recommendations. Reprod. Domest. Anim. 44, 141–147 (2009). [CrossRef]
- K. S. Borge, R. Tønnessen, A. Nødtvedt, A. Indrebø, Litter size at birth in purebred dogs—A retrospective study of 224 breeds. Theriogenology 75, 911–919 (2011). [CrossRef]
- A. J. Cornelius, R. Moxon, J. Russenberger, B. Havlena, S. H. Cheong, Identifying risk factors for canine dystocia and stillbirths. Theriogenology 128, 201–206 (2019). [CrossRef]
- D. Bennett, Canine dystocia—a review of the literature. J. Small Anim. Pract. 15, 101–117 (1974).
- P. D. McGreevy, et al., Dog behavior co-varies with height, bodyweight and skull shape. PLoS One 8, e80529 (2013). [CrossRef]
- I. Zapata, J. A. Serpell, C. E. Alvarez, Genetic mapping of canine fear and aggression. BMC Genomics 17, 1–20 (2016). [CrossRef]
- I. Zapata, A. W. Eyre, C. E. Alvarez, J. A. Serpell, Latent class analysis of behavior across dog breeds reveal underlying temperament profiles. Sci. Rep. 12, 15627 (2022). [CrossRef]
- C. Arhant, H. Bubna-Littitz, A. Bartels, A. Futschik, J. Troxler, Behaviour of smaller and larger dogs: Effects of training methods, inconsistency of owner behaviour and level of engagement in activities with the dog. Appl. Anim. Behav. Sci. 123, 131–142 (2010). [CrossRef]
- A. Bassi, L. Pierantoni, S. Cannas, C. Mariti, Dog’s size affects owners’ behaviour and attitude during dog walking. Dog behavior 2, 1–8 (2016).
- W. S. Helton, N. D. Helton, Physical size matters in the domestic dog’s (Canis lupus familiaris) ability to use human pointing cues. Behav. Processes 85, 77–79 (2010). [CrossRef]
- B. Wilson, J. Serpell, H. Herzog, P. McGreevy, Prevailing clusters of canine behavioural traits in historical US demand for dog breeds (1926–2005). Animals (Basel) 8, 197 (2018). [CrossRef]
- E. E. Hecht, et al., Neurodevelopmental scaling is a major driver of brain–behavior differences in temperament across dog breeds. Brain Struct. Funct. 226, 2725–2739 (2021). [CrossRef]
- B. L. Finlay, R. B. Darlington, Linked regularities in the development and evolution of mammalian brains. Science 268, 1578–1584 (1995). [CrossRef]
- B. L. Finlay, R. B. Darlington, N. Nicastro, Developmental structure in brain evolution. Behav. Brain Sci. 24, 263–278 (2001). [CrossRef]
- B. L. Finlay, K. Huang, Developmental duration as an organizer of the evolving mammalian brain: scaling, adaptations, and exceptions. Evol. Dev. 22, 181–195 (2020). [CrossRef]
- F. Pirrone, et al., Correlation between the size of companion dogs and the profile of the owner: A cross-sectional study in ItalyMarian. Dog behavior 1, 32–43 (2015). [CrossRef]
- D. G. O’Neill, et al., Unravelling the health status of brachycephalic dogs in the UK using multivariable analysis. Sci. Rep. 10, 17251 (2020). [CrossRef]
- R. M. A. Packer, D. G. O’Neill, F. Fletcher, M. J. Farnworth, Great expectations, inconvenient truths, and the paradoxes of the dog-owner relationship for owners of brachycephalic dogs. PLoS One 14, e0219918 (2019). [CrossRef]
- R. M. A. Packer, A. Hendricks, C. C. Burn, Do dog owners perceive the clinical signs related to conformational inherited disorders as “normal”for the breed? A potential constraint to improving canine welfare. Anim. Welf. 21, 81–93 (2012). [CrossRef]
- R. M. A. Packer, A. Wade, J. Neufuss, Nothing Could Put Me Off: Assessing the Prevalence and Risk Factors for Perceptual Barriers to Improving the Welfare of Brachycephalic Dogs. Pets 1, 458–484 (2024). [CrossRef]
- E. S. Paul, et al., That brachycephalic look: Infant-like facial appearance in short-muzzled dog breeds. Anim. Welf. 32, e5 (2023). [CrossRef]
- D. Krainer, G. Dupré, Brachycephalic obstructive airway syndrome. Vet. Clin. North Am. Small Anim. Pract. 52, 749–780 (2022). [CrossRef]
- S. Mitze, V. R. Barrs, J. A. Beatty, S. Hobi, P. M. Bęczkowski, Brachycephalic obstructive airway syndrome: much more than a surgical problem. Vet. Q. 42, 213–223 (2022). [CrossRef]
- K. J. Ekenstedt, K. R. Crosse, M. Risselada, Canine brachycephaly: anatomy, pathology, genetics and welfare. J. Comp. Pathol. 176, 109–115 (2020). [CrossRef]
- J. Grand, S. Bureau, Structural characteristics of the soft palate and meatus nasopharyngeus in brachycephalic and non-brachycephalic dogs analysed by CT. J. Small Anim. Pract. 52, 232–239 (2011). [CrossRef]
- D. A. Barker, C. Rubiños, O. Taeymans, J. L. Demetriou, Magnetic resonance imaging evaluation of olfactory bulb angle and soft palate dimensions in brachycephalic and nonbrachycephalic dogs. Am. J. Vet. Res. 79, 170–176 (2018). [CrossRef]
- B. A. Jones, B. J. Stanley, N. C. Nelson, The impact of tongue dimension on air volume in brachycephalic dogs. Vet. Surg. 49, 512–520 (2020). [CrossRef]
- J. A. Ginn, M. S. A. Kumar, B. C. McKiernan, B. E. Powers, Nasopharyngeal turbinates in brachycephalic dogs and cats. J. Am. Anim. Hosp. Assoc. 44, 243–249 (2008). [CrossRef]
- R. Schuenemann, G. U. Oechtering, Inside the brachycephalic nose: intranasal mucosal contact points. J. Am. Anim. Hosp. Assoc. 50, 149–158 (2014). [CrossRef]
- M. Auger, K. Alexander, G. Beauchamp, M. Dunn, Use of CT to evaluate and compare intranasal features in brachycephalic and normocephalic dogs. J. Small Anim. Pract. 57, 529–536 (2016). [CrossRef]
- F. J. Fasanella, J. M. Shivley, J. L. Wardlaw, S. Givaruangsawat, Brachycephalic airway obstructive syndrome in dogs: 90 cases (1991–2008). J. Am. Vet. Med. Assoc. 237, 1048–1051 (2010). [CrossRef]
- A. Conte, S. Morabito, R. Dennis, D. Murgia, Computed tomographic comparison of esophageal hiatal size in brachycephalic and non-brachycephalic breed dogs. Vet. Surg. 49, 1509–1516 (2020). [CrossRef]
- E. J. Reeve, D. Sutton, E. J. Friend, C. M. R. Warren-Smith, Documenting the prevalence of hiatal hernia and oesophageal abnormalities in brachycephalic dogs using fluoroscopy. J. Small Anim. Pract. 58, 703–708 (2017). [CrossRef]
- L. Lilja-Maula, et al., Comparison of submaximal exercise test results and severity of brachycephalic obstructive airway syndrome in English bulldogs. Vet. J. 219, 22–26 (2017). [CrossRef]
- K. Schmidt-Nielsen, W. L. Bretz, C. R. Taylor, Panting in dogs: unidirectional air flow over evaporative surfaces. Science 169, 1102–1104 (1970). [CrossRef]
- R. Oshita, S. Katayose, E. Kanai, S. Takagi, Assessment of nasal structure using CT imaging of brachycephalic dog breeds. Animals (Basel) 12, 1636 (2022). [CrossRef]
- I. Carrera, R. Dennis, D. J. Mellor, J. Penderis, M. Sullivan, Use of magnetic resonance imaging for morphometric analysis of the caudal cranial fossa in Cavalier King Charles Spaniels. Am. J. Vet. Res. 70, 340–345 (2009). [CrossRef]
- S. P. Knowler, et al., Quantitative analysis of Chiari-like malformation and syringomyelia in the Griffon Bruxellois dog. PLoS One 9, e88120 (2014). [CrossRef]
- C. Rusbridge, New considerations about Chiari-like malformation, syringomyelia and their management. In Pract. 42, 252–267 (2020). [CrossRef]
- C. Rusbridge, P. Knowler, The need for head space: brachycephaly and cerebrospinal fluid disorders. Life (Basel) 11, 139 (2021). [CrossRef]
- S. P. Knowler, et al., Facial changes related to brachycephaly in Cavalier King Charles Spaniels with Chiari-like malformation associated pain and secondary syringomyelia. J. Vet. Intern. Med. 34, 237–246 (2020). [CrossRef]
- C. Rusbridge, H. Carruthers, M. Dubé, M. Holmes, N. D. Jeffery, Syringomyelia in cavalier King Charles spaniels: the relationship between syrinx dimensions and pain. J. Small Anim. Pract. 48, 432–436 (2007). [CrossRef]
- A. C. Freeman, et al., Chiari-like malformation and syringomyelia in American Brussels griffon dogs. J. Vet. Intern. Med. 28, 1551–1559 (2014). [CrossRef]
- A. Kiviranta, C. Rusbridge, A. K. Lappalainen, J. J. T. Junnila, T. S. Jokinen, Persistent fontanelles in Chihuahuas. Part I. Distribution and clinical relevance. J. Vet. Intern. Med. 35, 1834–1847 (2021). [CrossRef]
- I. N. Plessas, et al., Long-term outcome of Cavalier King Charles spaniel dogs with clinical signs associated with Chiari-like malformation and syringomyelia. Vet. Rec. 171, 501–501 (2012). [CrossRef]
- S. Sanchis-Mora, et al., Dogs attending primary-care practice in England with clinical signs suggestive of Chiari-like malformation/syringomyelia. Vet. Rec. 179, 436–436 (2016). [CrossRef]
- M. S. Thøfner, et al., Prevalence and heritability of symptomatic syringomyelia in Cavalier King Charles Spaniels and long-term outcome in symptomatic and asymptomatic littermates. J. Vet. Intern. Med. 29, 243–250 (2015). [CrossRef]
- K. Wijnrocx, et al., Twelve years of chiari-like malformation and syringomyelia scanning in Cavalier King Charles Spaniels in the Netherlands: Towards a more precise phenotype. PLoS One 12, e0184893 (2017). [CrossRef]
- T. J. Mitchell, S. P. Knowler, H. van den Berg, J. Sykes, C. Rusbridge, Syringomyelia: determining risk and protective factors in the conformation of the Cavalier King Charles Spaniel dog. Canine Genet. Epidemiol. 1, 1–12 (2014). [CrossRef]
- S. Cerda-Gonzalez, N. J. Olby, E. H. Griffith, Medullary position at the craniocervical junction in mature cavalier king charles spaniels: relationship with neurologic signs and syringomyelia. J. Vet. Intern. Med. 29, 882–886 (2015). [CrossRef]
- J. Couturier, D. Rault, L. Cauzinille, Chiari-like malformation and syringomyelia in normal cavalier King Charles spaniels: a multiple diagnostic imaging approach. J. Small Anim. Pract. 49, 438–443 (2008). [CrossRef]
- J. E. Parker, S. P. Knowler, C. Rusbridge, E. Noorman, N. D. Jeffery, Prevalence of asymptomatic syringomyelia in Cavalier King Charles spaniels. Vet. Rec. 168, 667–667 (2011). [CrossRef]
- T. R. Pedersen, et al., Clinical predictors of syringomyelia in Cavalier King Charles Spaniels with chiari-like malformation based on owners’ observations. Acta Vet. Scand. 66, 5 (2024). [CrossRef]
- T. Lewis, C. Rusbridge, P. Knowler, S. Blott, J. A. Woolliams, Heritability of syringomyelia in Cavalier King Charles spaniels. Vet. J. 183, 345–347 (2010). [CrossRef]
- C. Limpens, V. T. M. Smits, H. Fieten, P. J. J. Mandigers, The effect of MRI-based screening and selection on the prevalence of syringomyelia in the Dutch and Danish Cavalier King Charles Spaniels. Front. Vet. Sci. 11, 1326621 (2024). [CrossRef]
- A. Prodger, S. Khan, G. Harris, Prevalence of structural and idiopathic epilepsy in brachycephalic and non-brachycephalic dogs in the context of the International Veterinary Epilepsy Task Force guidelines. J. Small Anim. Pract. (2025). [CrossRef]
- L. Sebbag, R. F. Sanchez, The pandemic of ocular surface disease in brachycephalic dogs: The brachycephalic ocular syndrome. Vet. Ophthalmol. 26, 31–46 (2023). [CrossRef]
- S. V. Palmer, F. E. Gomes, J. A. A. McArt, Ophthalmic disorders in a referral population of seven breeds of brachycephalic dogs: 970 cases (2008–2017). J. Am. Vet. Med. Assoc. 259, 1318–1324 (2021). [CrossRef]
- D. G. O’Neill, M. M. Lee, D. C. Brodbelt, D. B. Church, R. F. Sanchez, Corneal ulcerative disease in dogs under primary veterinary care in England: epidemiology and clinical management. Canine Genet. Epidemiol. 4, 1–12 (2017). [CrossRef]
- R. M. A. Packer, A. Hendricks, C. C. Burn, Impact of facial conformation on canine health: corneal ulceration. PLoS One 10, e0123827 (2015). [CrossRef]
- R. M. A. Packer, A. Hendricks, M. S. Tivers, C. C. Burn, Impact of facial conformation on canine health: brachycephalic obstructive airway syndrome. PLoS One 10, e0137496 (2015). [CrossRef]
- D. G. O’Neill, et al., Disorders of Bulldogs under primary veterinary care in the UK in 2013. PLoS One 14, e0217928 (2019). [CrossRef]
- D. G. O’Neill, E. C. Darwent, D. B. Church, D. C. Brodbelt, Demography and health of Pugs under primary veterinary care in England. Canine Genet. Epidemiol. 3, 1–12 (2016). [CrossRef]
- T. Töpfer, C. Köhler, S. Rösch, G. Oechtering, Brachycephaly in French bulldogs and pugs is associated with narrow ear canals. Vet. Dermatol. 33, 214–e60 (2022). [CrossRef]
- R. Salgüero, M. Herrtage, M. Holmes, P. Mannion, J. Ladlow, Comparison between computed tomographic characteristics of the middle ear in nonbrachycephalic and brachycephalic dogs with obstructive airway syndrome. Vet. Radiol. Ultrasound 57, 137–143 (2016). [CrossRef]
- W. Stern-Bertholtz, L. Sjöström, N. W. Håkanson, Primary secretory otitis media in the Cavalier King Charles spaniel: a review of 61 cases. J. Small Anim. Pract. 44, 253–256 (2003). [CrossRef]
- M. V. Estevam, et al., Congenital malformations in brachycephalic dogs: A retrospective study. Front. Vet. Sci. 9, 981923 (2022). [CrossRef]
- N. Roman, P. C. Carney, N. Fiani, S. Peralta, Incidence patterns of orofacial clefts in purebred dogs. PLoS One 14, e0224574 (2019). [CrossRef]
- S. Peralta, R. D. Campbell, N. Fiani, K. H. Kan-Rohrer, F. J. M. Verstraete, Outcomes of surgical repair of congenital palatal defects in dogs. J. Am. Vet. Med. Assoc. 253, 1445–1451 (2018). [CrossRef]
- K. M. Kelly, J. Bardach, “Biologic basis of cleft palate and palatal surgery” in Oral and Maxillofacial Surgery in Dogs and Cats, (Elsevier, 2012), pp. 343–350.
- A. Nemec, L. Daniaux, E. Johnson, S. Peralta, F. J. M. Verstraete, Craniomaxillofacial abnormalities in dogs with congenital palatal defects: computed tomographic findings. Vet. Surg. 44, 417–422 (2015). [CrossRef]
- S. Peralta, A. Nemec, N. Fiani, F. J. M. Verstraete, Staged double-layer closure of palatal defects in 6 dogs. Vet. Surg. 44, 423–431 (2015). [CrossRef]
- K. M. Evans, V. J. Adams, Proportion of litters of purebred dogs born by caesarean section. J. Small Anim. Pract. 51, 113–118 (2010). [CrossRef]
- E. Wydooghe, E. Berghmans, T. Rijsselaere, A. Van Soom, International breeder inquiry into the reproduction of the English bulldog. Vlaams Diergeneeskd. Tijdschr. 82 (2013).
- A. Eneroth, C. Linde-Forsberg, M. Uhlhorn, M. Hall, Radiographic pelvimetry for assessment of dystocia in bitches: a clinical study in two terrier breeds. J. Small Anim. Pract. 40, 257–264 (1999). [CrossRef]
- A. Bergström, A. N. E. Nødtvedt, A. Lagerstedt, A. Egenvall, Incidence and breed predilection for dystocia and risk factors for cesarean section in a Swedish population of insured dogs. Vet. Surg. 35, 786–791 (2006). [CrossRef]
- D. Bannasch, et al., The effects of FGF4 retrogenes on canine morphology. Genes (Basel) 13, 325 (2022). [CrossRef]
- H. G. Parker, et al., An expressed fgf4 retrogene is associated with breed-defining chondrodysplasia in domestic dogs. Science 325, 995–998 (2009). [CrossRef]
- H. G. Parker, A. Harris, D. L. Dreger, B. W. Davis, E. A. Ostrander, The bald and the beautiful: hairlessness in domestic dog breeds. Philos. Trans. R. Soc. Lond. B Biol. Sci. 372, 20150488 (2017). [CrossRef]
- S. Martínez, R. Fajardo, J. Valdés, R. Ulloa-Arvizu, R. Alonso, Histopathologic study of long-bone growth plates confirms the basset hound as an osteochondrodysplastic breed. Can. J. Vet. Res. 71, 66 (2007).
- A. K. Lappalainen, et al., Breed-typical front limb angular deformity is associated with clinical findings in three chondrodysplastic dog breeds. Front. Vet. Sci. 9, 1099903 (2023). [CrossRef]
- T. W. Kwan, D. J. Marcellin-Little, O. L. A. Harrysson, Correction of biapical radial deformities by use of bi-level hinged circular external fixation and distraction osteogenesis in 13 dogs. Vet. Surg. 43, 316–329 (2014). [CrossRef]
- R. E. Lau, Inherited premature closure of the distal ulnar physis. (1977).
- C. B. Carrig, Growth abnormalities of the canine radius and ulna. (1983). [CrossRef]
- D. B. Fox, J. L. Tomlinson, J. L. Cook, L. M. Breshears, Principles of uniapical and biapical radial deformity correction using dome osteotomies and the center of rotation of angulation methodology in dogs. Vet. Surg. 35, 67–77 (2006). [CrossRef]
- J. Temwichitr, et al., Evaluation of radiographic and genetic aspects of hereditary subluxation of the radial head in Bouviers des Flandres. Am. J. Vet. Res. 71, 884–890 (2010). [CrossRef]
- J. L. Knapp, J. L. Tomlinson, D. B. Fox, Classification of angular limb deformities affecting the canine radius and ulna using the center of rotation of angulation method. Vet. Surg. 45, 295–302 (2016). [CrossRef]
- A. K. Lappalainen, T. Hyvärinen, J. Junnila, O. Laitinen-Vapaavuori, Radiographic evaluation of elbow incongruity in Skye terriers. J. Small Anim. Pract. 57, 96–99 (2016). [CrossRef]
- L. F. H. Theyse, G. Voorhout, H. A. W. Hazewinkel, Prognostic factors in treating antebrachial growth deformities with a lengthening procedure using a circular external skeletal fixation system in dogs. Vet. Surg. 34, 424–435 (2005). [CrossRef]
- American Kennel Club, Dachshund Dog Breed Information. Available at: https://www.akc.org/dog-breeds/dachshund/ [Accessed 2024].
- Skye Terrier Club of America, Skye Terrier Breed Standard. Available at: https://stca.us/about-skye-terriers/skye-terrier-breed-standard/.
- Cardigan Welsh Corgi Club of America, Official Standard of the Cardigan Welsh Corgi. Available at: https://cardigancorgis.com/cwcca/breed/standard [Accessed 2024].
- American Kennel Club, Official standard for the Basset Hound. Available at: https://images.akc.org/pdf/breeds/standards/BassetHound.pdf [Accessed 2024].
- The Basset Hound Club of America, Inc, Pocket guide to the Basset Hound: an aid for judges. Available at: https://basset-bhca.org/wp-content/uploads/2018/12/Pocket-Guide-3112015.pdf [Accessed 2024].
- E. A. Brown, et al., FGF4 retrogene on CFA12 is responsible for chondrodystrophy and intervertebral disc disease in dogs. Proc. Natl. Acad. Sci. U. S. A. 114, 11476–11481 (2017). [CrossRef]
- K. Batcher, et al., Phenotypic effects of FGF4 retrogenes on intervertebral disc disease in dogs. Genes (Basel) 10, 435 (2019). [CrossRef]
- N. Bergknut, et al., Intervertebral disc degeneration in the dog. Part 1: Anatomy and physiology of the intervertebral disc and characteristics of intervertebral disc degeneration. Vet. J. 195, 282–291 (2013). [CrossRef]
- N. D. Jeffery, J. M. Levine, N. J. Olby, V. M. Stein, Intervertebral disk degeneration in dogs: consequences, diagnosis, treatment, and future directions. J. Vet. Intern. Med. 27, 1318–1333 (2013). [CrossRef]
- H.-J. Hansen, A pathologic-anatomical study on disc degeneration in dog: With special reference to the so-called enchondrosis intervertebralis. Acta Orthop. Scand. 23, 1–130 (1952). [CrossRef]
- V. L. J. Reunanen, T. S. Jokinen, M. K. Hytönen, J. J. T. Junnila, A. K. Lappalainen, Evaluation of intervertebral disc degeneration in young adult asymptomatic Dachshunds with magnetic resonance imaging and radiography. Acta Vet. Scand. 65, 42 (2023). [CrossRef]
- L. A. Smolders, et al., Intervertebral disc degeneration in the dog. Part 2: chondrodystrophic and non-chondrodystrophic breeds. Vet. J. 195, 292–299 (2013). [CrossRef]
- R. M. A. Packer, A. Hendricks, H. A. Volk, N. K. Shihab, C. C. Burn, How long and low can you go? Effect of conformation on the risk of thoracolumbar intervertebral disc extrusion in domestic dogs. PLoS One 8, e69650 (2013). [CrossRef]
- R. M. A. Packer, I. J. Seath, D. G. O’Neill, S. De Decker, H. A. Volk, DachsLife 2015: an investigation of lifestyle associations with the risk of intervertebral disc disease in Dachshunds. Canine Genet. Epidemiol. 3, 1–15 (2016). [CrossRef]
- P. J. Dickinson, D. L. Bannasch, Current understanding of the genetics of intervertebral disc degeneration. Front. Vet. Sci. 7, 431 (2020). [CrossRef]
- A. K. Lappalainen, E. Vaittinen, J. Junnila, O. Laitinen-Vapaavuori, Intervertebral disc disease in Dachshunds radiographically screened for intervertebral disc calcifications. Acta Vet. Scand. 56, 1–7 (2014). [CrossRef]
- B. A. Brisson, S. L. Moffatt, S. L. Swayne, J. M. Parent, Recurrence of thoracolumbar intervertebral disk extrusion in chondrodystrophic dogs after surgical decompression with or without prophylactic enestration: 265 cases (1995–1999). J. Am. Vet. Med. Assoc. 224, 1808–1814 (2004). [CrossRef]
- P. D. Mayhew, et al., Risk factors for recurrence of clinical signs associated with thoracolumbar intervertebral disk herniation in dogs: 229 cases (1994–2000). J. Am. Vet. Med. Assoc. 225, 1231–1236 (2004). [CrossRef]
- J. M. Levine, et al., Owner-perceived, weighted quality-of-life assessments in dogs with spinal cord injuries. J. Am. Vet. Med. Assoc. 233, 931–935 (2008). [CrossRef]
- M. U. Ball, J. A. McGuire, S. F. Swaim, B. F. Hoerlein, Patterns of occurrence of disk disease among registered dachshunds. J. Am. Vet. Med. Assoc. 180, 519–522 (1982). [CrossRef]
- UC Davis Veterinary Medicine Veterinary Genetics Laboratory, Chondrodystrophy (CDDY and IVDD) and Chondrodysplasia (CDPA). Available at: https://vgl.ucdavis.edu/test/cddy-cdpa [Accessed 2024].
- L. T. Glickman, N. W. Glickman, C. M. Pérez, D. B. Schellenberg, G. C. Lantz, Analysis of risk factors for gastric dilatation and dilatation-volvulus in dogs. J. Am. Vet. Med. Assoc. 204, 1465–1471 (1994). [CrossRef]
- L. Glickman, et al., Radiological assessment of the relationship between thoracic conformation and the risk of gastric dilatation-volvulus in dogs. Vet. Radiol. Ultrasound 37, 174–180 (1996). [CrossRef]
- R. H. Schaible, et al., Predisposition to gastric dilatation-volvulus in relation to genetics of thoracic conformation in Irish setters. J. Am. Anim. Hosp. Assoc. 33, 379–383 (1997). [CrossRef]
- D. Schellenberg, Q. Yi, N. W. Glickman, L. T. Glickman, Influence of thoracic conformation and genetics on the risk of gastric dilatation-volvulus in Irish setters. J. Am. Anim. Hosp. Assoc. 34, 64–73 (1998). [CrossRef]
- J. S. Bell, Inherited and predisposing factors in the development of gastric dilatation volvulus in dogs. Top. Companion Anim. Med. 29, 60–63 (2014). [CrossRef]
- C. R. Sharp, E. A. Rozanski, Cardiovascular and systemic effects of gastric dilatation and volvulus in dogs. Top. Companion Anim. Med. 29, 67–70 (2014). [CrossRef]
- A. Pike, T. M. Smalle, Gastric dilatation-volvulus in dogs: A review of the literature from 2000-2020. Aust. Vet. Pr. 52 (2022).
- L. T. Glickman, N. W. Glickman, D. B. Schellenberg, M. Raghavan, T. Lee, Non-dietary risk factors for gastric dilatation-volvulus in large and giant breed dogs. J. Am. Vet. Med. Assoc. 217, 1492–1499 (2000). [CrossRef]
- D. G. O’neill, et al., Gastric dilation-volvulus in dogs attending UK emergency-care veterinary practices: prevalence, risk factors and survival. J. Small Anim. Pract. 58, 629–638 (2017). [CrossRef]
- K. K. Song, S. E. Goldsmid, J. Lee, D. J. Simpson, Retrospective analysis of 736 cases of canine gastric dilatation volvulus. Aust. Vet. J. 98, 232–238 (2020). [CrossRef]
- The American Kennel Club, Official Standard of the German Shepherd Dog. Available at: https://images.akc.org/pdf/breeds/standards/GermanShepherdDog.pdf [Accessed 23 January 2025].
- The American Kennel Club, The Official Standard of the Great Dane. Available at: https://images.akc.org/pdf/breeds/standards/GreatDane.pdf [Accessed 23 January 2025].
- S. Hobi, V. R. Barrs, P. M. Bęczkowski, Dermatological problems of brachycephalic dogs. Animals (Basel) 13, 2016 (2023). [CrossRef]
- D. G. O’NeillI, D. Rowe, D. C. Brodbelt, C. Pegram, A. Hendricks, Ironing out the wrinkles and folds in the epidemiology of skin fold dermatitis in dog breeds in the UK. Sci. Rep. 12, 10553 (2022). [CrossRef]
- D. G. O’Neill, et al., Is it now time to iron out the wrinkles? Health of Shar Pei dogs under primary veterinary care in the UK. Canine Med. Genet. 10, 11 (2023). [CrossRef]
- Universities Federation for Animal Welfare, Genetic Welfare Problems of Companion Animals - English Bulldog. Available at: https://www.ufaw.org.uk/dogs/english-bulldog [Accessed 2024].
- F. C. Stades, K. N. Gelatt, Diseases and surgery of the canine eyelid. Essentials of Veterinary Ophthalmology 22, 53 (2008).
- A. Van Der Woerdt, Adnexal surgery in dogs and cats. Vet. Ophthalmol. 7, 284–290 (2004).
- E. A. Giuliano, Don’t worry; he’ll grow out of his entropion… (BSAVA Library, 2015).
- J. Vidt, Eye Tacking. Available at: https://cspca.com/eye-tacking/ [Accessed 2024].
- A. Guandalini, et al., Epidemiology of ocular disorders presumed to be inherited in three large Italian dog breeds in Italy. Vet. Ophthalmol. 20, 420–426 (2017). [CrossRef]
- M. Olsson, et al., A novel unstable duplication upstream of HAS2 predisposes to a breed-defining skin phenotype and a periodic fever syndrome in Chinese Shar-Pei dogs. PLoS Genet. 7, e1001332 (2011). [CrossRef]
- American Kennel Club, Official standard of the Chinese Shar-Pei. Available at: https://images.akc.org/pdf/breeds/standards/ChineseSharPei.pdf [Accessed 2024].
- J. L. Bouma, Kyphosis and kyphoscoliosis associated with congenital malformations of the thoracic vertebral bodies in dogs. Congenital Abnormalities of the Skull, Vertebral Column, and Central Nervous System, An Issue of Veterinary Clinics of North America: Small Animal Practice, E-Book: Congenital Abnormalities of the Skull, Vertebral Column, and Central Nervous System, An Issue of Veterinary Clinics of North America: Small Animal Practice, E-Book 46, 295 (2016). [CrossRef]
- R. Ryan, R. Gutierrez-Quintana, G. Ter Haar, S. De Decker, Prevalence of thoracic vertebral malformations in French bulldogs, Pugs and English bulldogs with and without associated neurological deficits. Vet. J. 221, 25–29 (2017). [CrossRef]
- S. Bertram, G. Ter Haar, S. De Decker, Congenital malformations of the lumbosacral vertebral column are common in neurologically normal French bulldogs, English bulldogs, and pugs, with breed-specific differences. Vet. Radiol. Ultrasound 60, 400–408 (2019). [CrossRef]
- S. De Decker, C. Rohdin, R. Gutierrez-Quintana, Vertebral and spinal malformations in small brachycephalic dog breeds: Current knowledge and remaining questions. Vet. J. 106095 (2024). [CrossRef]
- T. A. Mansour, et al., Whole genome variant association across 100 dogs identifies a frame shift mutation in DISHEVELLED 2 which contributes to Robinow-like syndrome in Bulldogs and related screw tail dog breeds. PLoS Genet. 14, e1007850 (2018). [CrossRef]
- J. E. Niskanen, et al., Canine DVL2 variant contributes to brachycephalic phenotype and caudal vertebral anomalies. Hum. Genet. 140, 1535–1545 (2021). [CrossRef]
- C. Rohdin, et al., Presence of thoracic and lumbar vertebral malformations in pugs with and without chronic neurological deficits. Vet. J. 241, 24–30 (2018). [CrossRef]
- P. Moissonnier, P. Gossot, S. Scotti, Thoracic kyphosis associated with hemivertebra. Vet. Surg. 40, 1029–1032 (2011). [CrossRef]
- J. Guevar, et al., Computer-assisted radiographic calculation of spinal curvature in brachycephalic “screw-tailed” dog breeds with congenital thoracic vertebral malformations: reliability and clinical evaluation. PLoS One 9, e106957 (2014). [CrossRef]
- T. Aikawa, S. Kanazono, Y. Yoshigae, N. J. H. Sharp, K. R. Muñana, Vertebral stabilization using positively threaded profile pins and polymethylmethacrylate, with or without laminectomy, for spinal canal stenosis and vertebral instability caused by congenital thoracic vertebral anomalies. Vet. Surg. 36, 432–441 (2007). [CrossRef]
- M. Charalambous, et al., Surgical treatment of dorsal hemivertebrae associated with kyphosis by spinal segmental stabilisation, with or without decompression. Vet. J. 202, 267–273 (2014). [CrossRef]
- T. Aikawa, et al., A comparison of thoracolumbar intervertebral disc extrusion in French bulldogs and dachshunds and association with congenital vertebral anomalies. Vet. Surg. 43, 301–307 (2014). [CrossRef]
- M. C. C. M. Inglez de Souza, et al., Evaluation of the influence of kyphosis and scoliosis on intervertebral disc extrusion in French bulldogs. BMC Vet. Res. 14, 1–8 (2018). [CrossRef]
- N. Grapes, S. Bertram, R. Gonçalves, S. De Decker, Prevalence of discospondylitis and association with congenital vertebral body malformations in English and French bulldogs. J. Vet. Intern. Med. (2024). [CrossRef]
- K. Faller, et al., The effect of kyphoscoliosis on intervertebral disc degeneration in dogs. Vet. J. 200, 449–451 (2014). [CrossRef]
- E. Schlensker, O. Distl, Heritability of hemivertebrae in the French bulldog using an animal threshold model. Vet. J. 207, 188–189 (2016). [CrossRef]
- L. Roses, F. Yap, E. Welsh, Surgical management of screw-tail in dogs. Companion Anim. 23, 287–292 (2018). [CrossRef]
- P. Abrescia, et al., Caudectomy for resolution of tail folds intertrigo in brachycephalic dogs: about eleven cases. (2017). [CrossRef]
- M. Siniscalchi, S. d’Ingeo, M. Minunno, A. Quaranta, Communication in dogs. Animals (Basel) 8, 131 (2018). [CrossRef]
- Leaver, Reimchen, Behavioural responses of Canis familiaris to different tail lengths of a remotely-controlled life-size dog replica. Behaviour 145, 377–390 (2008). [CrossRef]
- N. H. C. Salmon Hillbertz, et al., Duplication of FGF3, FGF4, FGF19 and ORAOV1 causes hair ridge and predisposition to dermoid sinus in Ridgeback dogs. Nat. Genet. 39, 1318–1320 (2007). [CrossRef]
- X. Zhang, O. Distl, A study of Rhodesian Ridgeback dogs indicates that the duplication responsible for hair ridge is not identical with the hypothesized locus for dermoid sinus. Anim. Genet. 53 (2022). [CrossRef]
- C. H. Vite, In: Braund’s Clinical Neurology in Small Animals: Localization, Diagnosis and Treatment, CH Vite (Ed.).
- J. S. Gammie, Dermoid sinus removal in a Rhodesian Ridgeback dog. Can. Vet. J. 27, 250 (1986).
- N. J. Volstad, D. Tyrrell, J. Beck, Sacrocaudal dermoid sinus in three Rhodesian Ridgeback dogs. Aust. Vet. Pr. 46 (2016).
- V. Moritz, U. Ridgebacks, Rhodesian Ridgeback Health and Genetics A Breeders perspective.
- S. R. R. Sverige, Strategies for Health and Long-term Sustainable breeding. Available at: https://www.srrs.org/srrs/nyhetsarkiv/20231206-strategies-health-and-long-term-sustainable-breeding [Accessed 2024].
- C. Drögemüller, U. Philipp, B. Haase, A.-R. Günzel-Apel, T. Leeb, A noncoding melanophilin gene (MLPH) SNP at the splice donor of exon 1 represents a candidate causal mutation for coat color dilution in dogs. J. Hered. 98, 468–473 (2007). [CrossRef]
- D. J. Wiener, et al., Clinical and histological characterization of hair coat and glandular tissue of Chinese crested dogs. Vet. Dermatol. 24, 274–e62 (2013). [CrossRef]
- K. Kupczik, A. Cagan, S. Brauer, M. S. Fischer, The dental phenotype of hairless dogs with FOXI3 haploinsufficiency. Sci. Rep. 7, 5459 (2017). [CrossRef]
- L. Brancalion, B. Haase, C. M. Wade, Canine coat pigmentation genetics: a review. Anim. Genet. 53, 3–34 (2022). [CrossRef]
- G. M. Strain, Canine deafness. Vet. Clin. North Am. Small Anim. Pract. 42, 1209–1224 (2012). [CrossRef]
- M. F. Rothschild, P. S. Van Cleave, K. L. Glenn, L. P. Carlstrom, N. M. Ellinwood, Association of MITF with white spotting in Beagle crosses and Newfoundland dogs. Anim. Genet. 37 (2006). [CrossRef]
- S. Stritzel, A. Wöhlke, O. Distl, A role of the microphthalmia-associated transcription factor in congenital sensorineural deafness and eye pigmentation in Dalmatian dogs. J. Anim. Breed. Genet. 126, 59–62 (2009). [CrossRef]
- G. M. Strain, Deafness prevalence and pigmentation and gender associations in dog breeds at risk. Vet. J. 167, 23–32 (2004). [CrossRef]
- American Kennel Club, Official standard of the Dalmatian. Available at: https://images.akc.org/pdf/breeds/standards/Dalmatian.pdf [Accessed 2024].
- M. Reissmann, A. Ludwig, Pleiotropic effects of coat colour-associated mutations in humans, mice and other mammals (Elsevier, 2013). [CrossRef]
- T. Kawakami, et al., R-locus for roaned coat is associated with a tandem duplication in an intronic region of USH2A in dogs and also contributes to Dalmatian spotting. PLoS One 16, e0248233 (2021). [CrossRef]
- D. Bannasch, Mutations in the SLC2A9 gene cause hyperuricosuria and hyperuricemia in the dog. PLoS Genet. 4, 1–8 (2008). [CrossRef]
- Dalmatian Club of America, Dalmatians and Uric Acid HUA and LUA. Available at: https://dcaf.org/dalmatian-health/urinary-stones/lua-dalmatians/ [Accessed 2024].
- L. A. Clark, J. M. Wahl, C. A. Rees, K. E. Murphy, Retrotransposon insertion in SILV is responsible for merle patterning of the domestic dog. Proc. Natl. Acad. Sci. U. S. A. 103, 1376–1381 (2006). [CrossRef]
- B. C. Ballif, et al., The PMEL gene and merle in the domestic dog: A continuum of insertion lengths leads to a spectrum of coat color variations in Australian shepherds and related breeds. Cytogenet. Genome Res. 156, 22–34 (2018). [CrossRef]
- M. Langevin, H. Synkova, T. Jancuskova, S. Pekova, Merle phenotypes in dogs–SILV SINE insertions from Mc to Mh. PLoS One 13, e0198536 (2018). [CrossRef]
- I. Reetz, M. Stecker, W. Wegner, Audiometric findings in dachshunds (merle gene carriers). (1977).
- U. Philipp, et al., Polymorphisms within the canine MLPH gene are associated with dilute coat color in dogs. BMC Genet. 6, 1–15 (2005). [CrossRef]
- A. Bauer, A. Kehl, V. Jagannathan, T. Leeb, A novel MLPH variant in dogs with coat colour dilution. Anim. Genet. 49, 94–97 (2018). [CrossRef]
- S. L. Van Buren, et al., A third MLPH variant causing coat color dilution in dogs. Genes (Basel) 11, 639 (2020). [CrossRef]
- W. H. Miller Jr, Colour dilution alopecia in Doberman Pinschers with blue or fawn coat colours: a study on the incidence and histopathology of this disorder. Vet. Dermatol. 1, 113–122 (1990). [CrossRef]
- M. Welle, et al., MLPH genotype—melanin phenotype correlation in dilute dogs. J. Hered. 100, S75–S79 (2009). [CrossRef]
- S. M. Schmutz, T. G. Berryere, Genes affecting coat colour and pattern in domestic dogs: a review. Anim. Genet. 38, 539–549 (2007). [CrossRef]
- C. Laffort-Dassot, L. Beco, D. N. Carlotti, Follicular dysplasia in five Weimaraners. Vet. Dermatol. 13, 253–260 (2002). [CrossRef]
- F. O. Smith, The Issue of the Silver Labrador. Available at: https://thelabradorclub.com/the-issue-of-the-silver-labrador/.
- French Bull Dog Club of America, Interpretation of the French Bulldog Standard on Color. Available at: https://frenchbulldogclub.org/color/ [Accessed 2024].
- L. Asher, G. Diesel, J. F. Summers, P. D. McGreevy, L. M. Collins, Inherited defects in pedigree dogs. Part 1: Disorders related to breed standards. The Veterinary Journal 182, 402–411 (2009). [CrossRef]
- G. Lehner, C. S. Louis, R. S. Mueller, Reproducibility of ear cytology in dogs with otitis externa. Vet. Rec. 167, 23–26 (2010). [CrossRef]
- D. G. O’Neill, et al., Frequency and predisposing factors for canine otitis externa in the UK–a primary veterinary care epidemiological view. Canine Med. Genet. 8, 1–16 (2021). [CrossRef]
- M.-D. Crapon De Caprona, P. Savolainen, Extensive Phenotypic Diversity among South Chinese Dogs. ISRN Evol. Biol. 2013, 1–8 (2013). [CrossRef]
- A. T. Vanak, M. E. Gompper, Interference competition at the landscape level: the effect of free-ranging dogs on a native mesocarnivore. J. Appl. Ecol. 47, 1225–1232 (2010). [CrossRef]
- E. Ruiz-Izaguirre, et al., Roaming characteristics and feeding practices of village dogs scavenging sea-turtle nests. Anim. Conserv. 18, 146–156 (2015). [CrossRef]
- D. Krauze-Gryz, J. Gryz, Free-Ranging Domestic Dogs ( Canis familiaris ) in Central Poland: Density, Penetration Range and Diet Composition. Pol. J. Ecol. 62, 183–193 (2014). [CrossRef]
- L. Vychodilova, et al., Genetic diversity and population structure of African village dogs based on microsatellite and immunity-related molecular markers. PLoS One 13, e0199506 (2018). [CrossRef]
- G. Leroy, S.-Z. Wang, T. Lewis, S. Licari, Ancient and Recent Changes in Breeding Practices for Dogs. Dogs, Past and Present 24 (2023).
- American Kennel Club, Most Popular Dog Breeds. (2024). Available at: https://www.akc.org/most-popular-breeds/ [Accessed 20 January 2025].
- D. G. O’Neill, et al., French Bulldogs differ to other dogs in the UK in propensity for many common disorders: a VetCompass study. Canine Med. Genet. 8, 1–14 (2021). [CrossRef]
- L. R. McAuliffe, C. S. Koch, J. Serpell, K. L. Campbell, Associations between atopic dermatitis and anxiety, aggression, and fear-based behaviors in dogs. J. Am. Anim. Hosp. Assoc. 58, 161–167 (2022). [CrossRef]
- Brachycephalic Working Group, Brachycephalic Working Group. Available at: https://www.ukbwg.org.uk/ [Accessed 20 January 2025].
- F. W. Nicholas, Response to the documentary pedigree dogs exposed: three reports and their recommendations. [Preprint] (2011).
- E. Morel, L. Malineau, C. Venet, V. Gaillard, F. Péron, Prioritization of Appearance over Health and Temperament Is Detrimental to the Welfare of Purebred Dogs and Cats. Animals (Basel) 14, 1003 (2024). [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author[s] and contributor[s] and not of MDPI and/or the editor[s]. MDPI and/or the editor[s] disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



