
Submitted:
11 June 2025
Posted:
11 June 2025
You are already at the latest version
Abstract
Bread is consumed daily throughout the world as an important source of nutrients. How-ever, bakery products are highly susceptible to spoilage, especially fungal, which is a source of bread losses and a threat to food security and consumer health. The use of sour-doughs is the best alternative to chemical preservatives, while providing a number of ad-vantages to baked bread. This review highlights the main areas in the field of bread pro-tection, covers the principal representatives of sourdough microbiota and their contribu-tion to protecting bread from spoilage. The review is mainly based on publications in the field of research over the last five years, identifying new directions and strategies for bread protection related to the use of sourdoughs. A list of the main compounds produced by lactic acid bacteria of the sourdough, which contribute to the protection of bread from fungal spoilage, is presented. The contribution of other microorganisms to the antifungal effect is also considered. Finally, some prospects for the development of research in the field of sourdoughs are determined.
Keywords:
sourdough
; antifungal
; organic acids
; antimicrobial peptides
; preservation
1. Introduction
Bread is an important source of nutrients and a staple food consumed daily worldwide. The average consumption of bread across European countries is 50 kg per person per year, reaching up to 80 kg per person per year in some countries [1,2]. However, the susceptibility of bread to spoilage threatens global food security. The main cause of the spoilage of bakery products is mold damage. Due to high humidity and an optimal pH range, bread and bakery products are highly susceptible to fungal spoilage, which can occur during the shelf life. Bread losses due to mold spoilage are estimated at 20% [2]. In addition to fungal growth on the surface of bread and repelling consumers with the appearance, molds cause changes in taste and produce mycotoxins, which make bread potentially unsafe. Calcium propionate has traditionally been used to inhibit mold growth in bread and bakery products. However, this additive has been shown to cause hypersensitivity, visual irritability, attention, and sleep disturbances [3].
Recently, the number of health-conscious consumers and the demand for natural and healthy products have been growing, and much attention has been paid to the safety of consumed products and the absence of chemical additives in them. Which, in turn, also contributes to an increase in product spoilage. So, fungal spoilage remains an unsolved problem in bread making.
The most significant agents of bread spoilage are fungi of the genera Penicillium, Aspergillus, Paecilomyces, and Rhizopus [1,4,5,6,7,8], which are characterized by a higher growth rate and competitiveness on the bread surface compared to other fungi. Well-known species of the genus Penicillium include P. roqueforti, P. paneum, P. corylophilum, P. chrysogenum, P. brevicompactum, and others. The genus Aspergillus include A. chevalieri, A. niger, and A. pseudoglaucus, while Paecilomyces variotii is characteristic for the genus Paecilomyces. Bread spoilage has also been reported from the genera Mucor, Endomyces, Cladosporium, Fusarium, Wallemia, and Neurospora [5,7,9], whereas the ‘Chalk mold’ is usually caused by the yeast-like fungi Hyphopichia burtonii and Saccharomycopsis fibuligera [5].
Fungal spores present in raw materials are inactivated during baking, but can be released into the air during mixing of ingredients as aerosols and settle on finished products [5]. Fungi can also be transferred to products during the slicing and packaging stages from the air and via equipment surfaces. Fungal spores are ubiquitous in the atmosphere at up to 1000 spores per m3, but in bakery air, the number of spores can be much higher.
Although the fungal spores are destroyed during baking, mycotoxins may remain in bread as they are relatively heat-stable. Levels of mycotoxins such as aflatoxin and ochratoxin, as well as deoxynivalenol and zearalenone, are regulated. However, maximum levels for many other toxins have not yet been established, and the potential synergistic effects of different mycotoxins are also of great concern [2]. The particular problem is the intensified growth of mould in countries with higher humidity and temperatures, which requires greater attention to fungal spoilage in Asia and Africa.
Bacterial ropy spoilage of bread, characterized by sticky viscosity, color, and odor changes, is caused by the presence in flour of endophytic commensal wheat microbiota of the genus Bacillus or close genera that form heat-stable endospores [10]. The Bacillus species causing ropiness mainly include B. subtilis, B. licheniformis, B. amyloliquefaciens, and B. pumilus. However, spoilage of bread by other species of this and related genera of spore-forming bacteria has also been reported. It is believed that the increasing demand for preservative-free whole grain products, along with global warming, may contribute to the spread of bread ropy spoilage [10].
The high demand for natural products with a clean label is driving the desire to replace chemical preservatives with natural alternatives. Microbial fermentation is the most environmentally friendly way to preserve food and a significant substitute for chemical preservatives. Lactic acid bacteria (LAB) have long been used and studied as an alternative to chemical preservatives. LAB not only guarantees a clean label for products but also provides many benefits, such as improved sensory indicators, texture, and nutritional value. LAB have the status of GRAS (Generally Regarded as Safe) in the USA and are included in the list of Qualified Presumption of Safety (QPS) of the European Union [11].
Review papers in the field of improving the preservation and safety of bread by using sourdoughs summarize data on the dominant microorganisms of spontaneous sourdoughs, their mechanisms of spoilage control, and the effect on extending the shelf life of various types of bread. The presented review combines information on bread sourdough microorganisms, both traditional and previously unstudied in detail, but promising for use, including taxonomic affiliation, production of antifungal and antibacterial compounds, focusing on works published in the last five years, clearly outlines the directions that have emerged in recent years and prospects for the development of research in the field of bread preservation and safety.
To summarize the data on the most common microorganisms in bread sourdoughs, term mapping was performed from OpenAlex text data. Terms were extracted from the title and abstract fields of 1967 documents, containing the term “sourdough” in the abstract. Binary counting was used to determine the frequency of publications mentioning certain species of microorganisms. The term co-occurrence was at least five documents. Direct counting was used to determine the depth of the study. Among the extracted terms, the species names of microorganisms were selected, and the mention number for each species was summed up, taking into account abbreviations. A direct search for studies in the field of bread sourdoughs was carried out in the Scopus, Web of Science, and Pub Med databases from 2020 to 2025 using combinations of the following keywords in search queries: sourdough AND (inhibitory OR antimicrobial OR antifungal OR antagonistic OR antibacterial OR bacteriostatic), as well as antimould/antimold, microbiota, “lactic acid bacteria”, “bread spoilage”, rope, and others. The main research areas were identified for relevant articles where necessary, and a more specific search was conducted when needed, including in the Google Scholar database and in extended time intervals.
This review provides a detailed and thorough summary of the latest research results on the effects of bread starters on improving bread shelf life and safety, highlighting the main directions and new strategies.
The genera of LAB are presented in the review in a modern classification [12].
2. Sourdough Microbiota
2.1. Mapping Terms Among Sourdough-Related Documents
46,852 terms were extracted from 1,967 documents with the word “sourdough” in the abstract. From extracted terms, 2,843 were repeated at least five times (binary count relied only on the presence of the term in a publication) (Figure 1a). Terms were automatically placed into several groups by the application. The names of microorganisms were mostly included in the red group (Figure 1b), which contained the largest number of terms and occupied a central position among other groups on the map. In this group, the dominance of Lactiplantibacillus plantarum is striking.
Among 1706 most relevant terms, the occurrence of LAB species related to the sourdough microbiota was calculated. Certain species and genera of LAB were most frequently mentioned in sourdough literature. Thus, the diagram in Figure 2 represents LAB species repeating in at least five publications from 1971 related to sourdough.
L. plantarum occupies a central position in sourdough bread research, accounting for 26.9% of occurrences among species names of microorganisms (Figure 2). This is explained by the fact that this species is abundantly represented in flour of various origins and can dominate its fermentation [13].
LAB of the genus Limosilactobacillus are the second most common (15.1% total) after the genus Lactiplantibacillus. However, among the species, Levilactobacillus brevis (9.2%) and Fructilactobacillus sanfranciscensis (7.7%) are slightly more represented in studies compared to Limosilactobacillus reuteri (6.8%) and Limosilactobacillus fermentum (5.0%). Pediococcus pentosaceus also reaches a significant number of mentions (5.2%). The genera of LAB are arranged in the following descending order of frequency of their species occurrence in publications: Lactiplantibacillus, Limosilactobacillus, Levilactobacillus, Fructilactobacillus, Pediococcus, Lacticaseibacillus, Lactobacillus, Weissella, Lactococcus, Leuconostoc, Companilactobacillus, Latilactobacillus, and Enterococcus. Studies of the dynamics of microbial successions during sourdough maturation [13] show that microorganisms such as Enterococcus, Lactococcus, and Leuconostoc are characteristic of the first phase, and Lactobacillus, Pediococcus, and Weissella are characteristic of the second. In the third phase, the microbiota of bread sourdough is dominated by individual species of Lactobacillus sensu lato, previously grouped into the genus Lactobacillus but transferred to the present time into several new genera [12].
According to the term mapping, the genus Limosilactobacillus included the largest number of species, mentioned in at least five publications. In turn, only four genera were represented by a single species. These species were F. sanfranciscensis, Lactococcus lactis, Latilactobacillus curvatus, and Furfurilactobacillus rossiae.
At the same time, some microorganisms were mentioned and studied in more detail, as determined by the total term count. Among them are Limosilactobacillus reuteri (the occurrence is 12.2% compared to 6.8% in the binary count), acetic acid bacteria (6.8% and 3.1%, respectively), and Lacticaseibacillus paracasei (3.0% and 1.6%, respectively). This may indicate some special functions of these microorganisms in bread sourdoughs.
In the sourdough yeast microbiota, the most frequently reported genera were Kazachstania (30.8%), Candida (24.4%), Saccharomyces (14.9%), Pichia (12.7%), Wickerhamomyces (9.0%), and Kluyveromyces (8.1%) (Figure 3). The yeast species Kazachstania humilis (formerly Candida humilis) and Candida milleri were the most frequent in sourdough publications.
2.2. Types of Sourdoughs
Various researchers have described sourdough microbiota according to the type of production method and fermented flour. There are three main types of sourdoughs, differing in the production method [13,14]. Type I sourdough is an artisanal spontaneous sourdough obtained by back-slopping – renewal using a fresh mixture of flour and water repeated 5-13 or more times. According to De Bondt et al. scoping review [15], about 18% of articles on sourdough are devoted to type I sourdoughs. Type II sourdoughs refer to industrial liquid sourdoughs and are based on pure cultures of LAB added to yeast in a ratio of 100:1 [14]. This type is most often described in the literature because it includes known strains of microorganisms with easily standardized technology for obtaining sourdough and dough. Type III sourdough is an industrial dry sourdough. Only about 8% of publications are devoted to dry sourdoughs [15]. Sometimes, type IV sourdough is distinguished, which is a combination of type I and type II.
2.2.1. Type I Sourdough
Type I sourdough is usually a mixture of yeasts and mesophilic LAB, typical for a particular flour type. The formation of microbiota in spontaneous sourdoughs occurs gradually. In the first stage, cereal microbiota microorganisms such as Enterobacter, Acinetobacter, Pseudomonas, and Sphingomonas are found in large quantities [13]. Klebsiella, Serratia, Erwinia, Clostridium, and Staphylococcus were also identified as dominant genera in the first stage [16]. From day 2 to day 5, Enterococcus, Lactococcus, and Pediococcus cocci predominate, and Weissella spp. and Lactobacillus (in particular, homofermentative Lactobacillus delbrueckii subsp. lactis) are also detected [13]. However, the transition to the second stage can be observed much earlier. Thus, De Angelis et al. [16] showed a high number of Lactococcus, Enterococcus, and Weissella already after 8 hours. A more rapid change in microbial communities is more characteristic of whole-grain dough. Thus, Boreczek et al. [17], studying the microbiota of spontaneous sourdoughs from whole grain flour (wheat, rye, and spelt) during the period of community formation (24–72 hours), showed that the total number of Enterococcus, Lactococcus, and Leuconostoc did not exceed 2% after 24 hours. The reduction in the number of acid-forming cocci is probably due to a rapid increase in acidity, which contributes to sourdough stabilization and dominance of resistant lacticobacilli species. In the studies of Boreczek et al. [17], Weissella was the most abundant genus after 24 hours of rye sourdough fermentation, and its abundance decreased thereafter in all sourdoughs. In the Oshiro et al. [18] studies, Weissella spp. dominated in environments with a more neutral pH, while L. brevis was attached to lower pH values.
The relative abundance of Enterococcus, Lactococcus, and Leuconostocaceae in the sourdough varied across geographical zones and wheat varieties; in contrast, rod-shaped LAB did not dominate the microbial communities of the untreated raw materials [19]. Maintaining the sourdough for several days and increasing its acidity leads to the dominance of certain species of lactobacilli that are well adapted to the sourdough ecosystem, such as L. plantarum and L. fermentum [13]. Interestingly, Latilactobacillus curvatus, L. brevis, and Lactiplantibacillus sp., which dominate in rye sourdoughs during the first three days, were replaced after one month in the dense sourdough with F. sanfranciscensis and Companilactobacillus sp., and in the liquid sourdough with Limosilactobacillus pontis [20].
A shift in LAB from the genera Weissella and Lactococcus to Pediococcus and Leuconostoc, and at later stages to Lactobacillus, is characteristic of spontaneous fermentation of plant materials [21,22,23]. The reduction in cocci and active proliferation of lactobacilli coincide with a decrease in substrate pH, since they are usually acid-resistant.
Dissimilar results were obtained by Baev et al. [24]. They showed high Weissella contents in whole grain wheat sourdoughs (16.98–42.84%). In addition, mature sourdoughs included 3.24–20.05% Pseudomonadota (former Proteobacteria), as well as significant amounts of Cyanobacteria in some sourdoughs. Also, in one of the mature industrial sourdoughs studied by Debonne et al. [25], Weissella was the dominant genus (73.8%, of which 62.2% was Weissella confusa). Leuconostoc citreum in this sourdough was 3.6%, and 14.5% were other bacteria. Most likely, these features are related to the sourdough production technology, but the studies mentioned do not provide information on the methods for obtaining mature sourdoughs.
The time required for sourdough maturation varies depending on the substrate used. For example, Weckx et al. [26] showed that reverse mowing of rye sourdough resulted in a complete change in the main genera and species of LAB from Lactococcus, Leuconostoc, and Weissella via L. curvatus and P. pentosaceus to L. fermentum and L. plantarum in just four days.
The production technology has a significant impact on the sourdough microbiota. For example, there is evidence of a relative abundance of Leuconostoc after 10 days of back-slopping [16]. The studies of De Angelis et al. [16] also demonstrated the dominance of P. pentosaceus and a high relative abundance of Weissella species in both soft wheat (42–72%) and durum wheat (19–35%) sourdoughs. The reason for these discrepancies lies in the different sourdough production technologies. The dough in the studies was fermented for only 6–8 hours per day, followed by a resting period (16–18 hours at 4 °C), preventing acid-forming cocci's subsequent replacement by acid-resistant rods.
Specific technological parameters such as temperature, dough yield, and the addition of salt affect the biodiversity and the increase in the number of certain LAB species in the sourdough [27]. Sourdough-associated LAB originate from flour, production environment, and other ingredients, if available [19]. Minervini et al. showed the influence of the propagation environment on the composition of sourdough yeast and LAB microbiotas [28]. At the same time, technological features, home characteristics, and climatic factors explained only 9% and 5% of the variations in the composition of bacterial and fungal communities, respectively [29]. In the studies of Minervini et al. [28], major differences between the microbiota of artisan bakery- or laboratory-propagated sourdoughs were shown for yeasts, which are spore-forming microorganisms that survive well on various work surfaces, and for LAB of the genus Leuconostoc, which are characterized by biofilm formation due to increased production of extracellular polysaccharides [30,31,32]. Therefore, the environmental microorganisms of spontaneous sourdoughs can be attributed to the flour microorganisms that survive during the production process on utensils and work surfaces, rather than to the externally introduced microbiota. Later, Minervini et al. [33] demonstrated that endophytic LAB of the cereals used for flour production form the basis of sourdough microbiota. The authors showed that Lactobacillus, Streptococcus, Enterococcus, and Lactococcus were the main LAB genera in durum wheat epiphytic and endophytic microbiota. The relative abundance of genera depends on plant varieties, phenological stages, environmental factors, and agronomic techniques. The dominance of L. plantarum in various sourdoughs (Figure 1) is explained by its association with wheat plant organs throughout the life cycle.
It has been shown that type I wheat and rye sourdoughs are usually colonized by insect-adapted lactobacilli such as F. sanfranciscensis, while type II sourdoughs usually include vertebrate-adapted lactobacilli of the L. delbrueckii and L. reuteri groups [34]. In contrast, F. sanfranciscensis and Pediococcus parvulus were reported to be more abundant in older sourdoughs, while L. plantarum was more numerous in younger sourdoughs [20,29]. F. sanfranciscensis was also more often identified in private sourdoughs originating from a commercial source, while L. brevis prevailed in de novo sourdoughs [29]. The latter showed good growth performance in a separate culture and was well preserved in a pair with competing LAB and yeast, whereas the former species was characterized by low competitive ability and could survive only with the yeast K. humilis. It is known that F. sanfranciscensis is characterized by the smallest genome size among lactobacilli and a low GC content, which indicates adaptation to a narrow ecological niche [34,35]. The reductive evolution of this species is confirmed by the highest density of ribosomal RNA operons, indicating a high frequency of gene inactivation and elimination [35]. F. sanfranciscensis is the most abundant and predominant lactobacilli species in faecal samples of confused flour beetle (Tribolium confusum) and, consequently, insect excreta from stored grain products may be a natural reservoir of these bacteria [36]. At the same time, the ability of F. sanfranciscensis to synthesize de novo deficient in wheat amino acids facilitates its good growth in a sourdough environment [35].
Landis et al. [29] studied 500 sourdoughs mostly from the United States (429) and found strong patterns of species dominance or co-occurrence. Most sourdoughs were dominated by a single yeast and/or bacterial species, with a median of three LAB and/or acetic acid bacteria (AAB) and one yeast species per sourdough. Saccharomyces cerevisiae was the most abundant yeast in the studied sourdoughs, followed by F. sanfranciscensis among the bacteria. L. sanfranciscensis was also shown to be negatively correlated with L. plantarum and L. brevis. L. brevis, in turn, was the most frequently co-occurring species. Despite the widespread occurrence of S. cerevisiae and F. sanfranciscensis, these microorganisms tended to be mutually exclusive, while K. humilis was more often found together with F. sanfranciscensis [14,25,29]. However, the dominance of S. cerevisiae among yeasts was also documented for F. sanfranciscensis-dominated sourdoughs [25,37]. At the same time, AAB and other LAB were negatively correlated with F. sanfranciscensis [37].
Interesting data were obtained by Xing et al. [38], indicating the influence of terrain conditions on the composition of sourdoughs. Acetobacter was widespread only in the mountain samples, while P. pentosaceus was the dominant strain in the basin sourdough samples.
AAB are now known as constant companions of natural fermentation of sugar-containing substrates by LAB and yeasts [39]. Although AAB rarely dominate due to strictly aerobic metabolism, they influence metabolic pathways and final product properties. They are also a common and characteristic component of spontaneous sourdoughs [29,37,40,41,42]. Landis et al. [29] detected AAB in 147 sourdough samples of 500 with a relative abundance of more than 1%. In another study, AABs were detected in 29.4% of 500 sourdough samples from different continents, with an average relative abundance of 23.3% [41]. Interestingly, all Belgian sourdoughs investigated by Comasio et al. [37] contained AABs. Nevertheless, the absence of a sourdough-specific genomic cluster indicates that AAB are nomadic microorganisms [41].
The presence of AABs in bread sourdoughs has not been systematically investigated, so data on their abundance are incomplete. Several AAB species belonging to Acetobacter and Komagataeibacter were detected in sourdoughs [37]. Isolated AAB species included Acetobacter cerevisiae, Acetobacter oryzifermentans, Acetobacter senegalensis, Acetobacter fabarum, Acetobacter pasteurianus subsp. pasteurianus, Acetobacter sicerae, Komagataeibacter xylinus, and Komagataeibacter sucrofermentans. In turn, shotgun metagenomic sequencing by Landis et al. [29] showed the presence of Acetobacter malorum, A. pasteurianus, and Acetobacter lovaniensis in sourdoughs. The most common AAB were Acetobacter malorum/cerevisiae and Acetobacter oryzoeni/oryzifermentans/pasteurianus. The genera Gluconobacter and Komagataeibacter were also noted.
The functions of AAB and their importance in sourdoughs remain to be fully understood.
2.2.2. Type II Sourdough
In type II sourdoughs, the dominant microorganisms are LAB, and S. cerevisiae is added for leavening [7]. The choice of starter cultures for the most common type II sourdoughs traditionally used in industry depends on the metabolic traits of technological and functional interest. The use of sourdough makes it possible to influence the sensory and rheological properties of bread, its shelf life and safety. Sourdough can also improve the nutritional and functional properties of bakery products, enriching them with vitamins, essential amino acids, fatty acids and bioactive peptides, reducing the glycemic index and gluten content, and increasing the bioavailability of minerals [19,43,44].
Important parameters for starter microorganisms are their acidifying capacity and growth characteristics. Other important properties of starter microorganisms include the production of exopolysaccharides and various volatile compounds, as well as proteolytic and antagonistic activities [43,45,46].
Acid-tolerant strains of LAB belonging to the genera Limosilactobacillus and Lactobacillus [7] are commonly used for sourdoughs, such as Lactobacillus amylovorus, Limosilactobacillus panis, L. pontis, and Limosilactobacillus reuteri [14]. F. sanfranciscensis is often used as the sole leavening agent [7].
LAB proteolytic activity is crucial for the degradation of flour proteins for affecting bread texture and the production of bioactive peptides. De Vero et al. [21] revealed significant proteolytic activity of L. citreum PRO17 and P. pentosaceus, strains OA1 and S3N3, in whole wheat flour dough. Angiotensin I-converting enzyme, for controlling hypertension, and cancer-preventive peptide lunasin are some bioactive compounds of sourdough LAB. As De Vero et al. showed, L. curvatus SAL33 and L. brevis AM7 significantly increased the concentration of lunasin [21]. In turn, L. amylovorus DSM19280, L. brevis R2Δ, and L. reuteri R29 revealed good results as microorganisms with high antifungal activity, producing a total of 171–589 mg/kg antifungal compounds [47]. The antimicrobial activity of LAB from sourdoughs will be discussed in more detail in the following sections.
Microorganisms originating from sources other than the spontaneously produced microbiota of dough and cereals are often considered promising for Type II sourdoughs. Thus, Comasio et al. [48] studied the adaptation of LAB isolates from other sources to a cereal matrix and showed the promise of using L. fermentum from cocoa mass and Latilactobacillus sakei from fermented sausage in wheat sourdoughs. Zhang et al. [49] found high cell counts of Lentilactobacillus buchneri from African cereal products and Lentilactobacillus diolivorans from maize silage in wheat, rye, and buckwheat sourdoughs. The microorganisms also produced acetate and propionate, inhibiting the growth of Aspergillus clavatus, Cladosporium spp., and Mortierella spp. Another study [50] demonstrated the effectiveness of L. plantarum from pickles against the bakery spoilage fungi Penicillium citrinum, Aspergillus flavus, Aspergillus fumigatus, Fusarium graminearum, Aspergillus niger, and Aspergillus ochraceus. While Muhialdin et al. [51] showed shelf life extension of bread inoculated with A. niger and Aspergillus oryzae using LAB (L. fermentum, P. pentosaceus, L. pentosus, and L. paracasei) isolated from Malaysian fruits and fermented products. L. plantarum ZZUA493 derived from alfalfa inhibited the growth of A. niger, A. oryzae, Trichoderma longibrachiatum, A. flavus, and F. graminearum in Chinese steamed buns through the production of lactic, acetic, and phenyllactic acids [52]. At the same time, dairy (kefir grain-derived) P. pentosaceus SP2 and L. paracasei SP5 enhanced the resistance of baked bread to both fungal spoilage and rope spoilage [53,54].
AAB have largely escaped research attention due to the difficulties of maintaining them in pure culture. However, the detection of significant amounts of AAB in spontaneous fermentations of various substrates with LAB and yeast indicates the need for a more thorough investigation of the contribution of AAB to co-fermentation and the properties of the resulting products. To date, few studies have been conducted to investigate the contribution of AAB to sourdough fermentation. However, contributions of AAB to dough acidification (by 18.5% compared to yeast and LAB), volatile aroma profile, amino acid biosynthesis, and dough properties (rise, viscosity, and elasticity) have been reported [29,37,41,55,56].
Research on the contribution of AAB to bread sourdoughs has only begun in recent years, but several studies have already shown promise for their use. Considering the production of acetic acid, the antimicrobial properties of different types of vinegar [57,58], the information on the novel antimicrobial properties of AAB in combination with MAB [59,60,61], and the involvement of AAB in the biodetoxification of mycotoxins [62,63,64], the influence of AAB on the shelf life and safety of bread should be expected.
2.2.3. Type III Sourdough
Desiccation-resistant LAB, such as P. pentosaceus, L. plantarum, and L. brevis, usually predominate in type III sourdoughs [7,66,86]. However, the survival of microorganisms in such starters depends on the drying process. Therefore, type III sourdoughs are often added to doughs only to improve the taste of bread.
Nevertheless, the convenience of dry sourdoughs encourages researchers to develop efficient technologies for their preparation and use. The research on dry sourdoughs is still in the developmental stage. However, the studies by Lafuente et al. [66] showed the preservation of bread prepared with dry sourdough fermented with P. pentosaceus TI6, which was experimentally infected with A. flavus and Penicillium verrucosum. In turn, Teixeira et al. [67] showed an extension of the shelf life of bread (especially from whole grain flour) using 10% type III sourdough with starter cultures of F. sanfranciscensis and L. plantarum. The water-soluble sourdough extract inhibited P. roqueforti, P. chrysogenum and A. niger. The best effect (shelf life of 10 days) was shown by sourdough from a mixture of wheat and flaxseed flour fermented with F. sanfranciscensis. In other studies [68], probiotic cultures of Lactobacillus acidophilus, Lacticaseibacillus casei, Bifidobacterium spp., and Bacillus coagulans were used in combination with a spray-dried sourdough fermented with LAB and propionibacteria. Its application of 1% (w/w) extended the shelf life of pizza and focaccia bases by an additional 10 days.
2.2.4. Type IV Sourdough
Type IV sourdough is a mixture of traditional type I and type II sourdoughs, where the starter cultures are added to a mature spontaneous sourdough [69]. Type IV sourdough can also be dried [70]. Another option for preparing a type IV sourdough is adding starter based on pure LAB cultures to the dough with subsequent daily renewal [71].
3. The Effect of Sourdough LAB on the Shelf Life of Bread
3.1. Antifungal Compounds
Sourdough LAB have been shown to inhibit the following mold fungi: A. fumigatus, A. niger, A. oryzae, Aspergillus versicolor, Aspergillus japonicus, A. clavatus, Aspergillus chevalieri, Aspergillus montevidensis, P. roqueforti, P. chrysogenum, Penicillium expansum, Penicillium crustosum, Penicillium olsonii, Penicillium polonicum, P. corylophilum, F. oxysporum, F. moniliforme, F. culmorum, Mucor spp., Rhizopus spp., Cladosporium spp., Neurospora spp., Mortierella spp., and others [2,72,73,74,75,76].
Various researchers have shown that the addition of 20-30% LAB-based sourdough to the dough extends the shelf life by 1 to 25 days, depending on the selected microbial strains [7,77,78]. Along with the shelf life extension of bread baked using sourdough, an improvement in texture and other consumer and production-relevant attributes is often noted [43]. Sensory properties are assessed in various studies in the range from slightly below control to above control, which is associated with an increase in the diversity and content of volatile compounds and consumer individual preferences [79,80].
3.1.1. Organic Acids
Many researchers note that the main mechanism of LAB that exerts an antifungal effect is the production of organic acids. A mixture of lactic, phenyllactic, and acetic acids is usually reported [47,65,73,76,78]. For example, aqueous extracts of wheat flour fermented with L. acidophilus and L. casei produced 1–2% lactic acid, which reduced the growth rate of the molds P. crysogenum and P. corylophilum [73]. In another study, cell-free supernatants of several LAB (W. cibaria, L. plantarum subsp. plantarum, L. pseudomesenteroides, F. sanfranciscensis, L. brevis, and L. pentosus) contained 4.33–8.41 g/L lactic acid that inhibited A. flavus, A. niger, and P. expansum [81]. Other researchers estimated an increase in lactic and acetic acid concentrations in sourdough bread by 4.5 and 1.6 times, respectively, compared to the control [82]. In the studies of Wu et al. [76], L. plantarum produced 60 times more phenyllactic acid than P. pentosaceus, which provided better suppression of A. niger and P. polonicum. In turn, L. plantarum P10 inhibited spore and mycelial growth of A. niger in Chinese steamed bread [83], while panettone with L. fermentum contained 5–10.4 mmol/kg phenyllactic acid, 1.17–8.85 mmol/kg acetic acid and 3.5–4.3 mmol/kg propionic acid [84].
Some researchers use standard unmodified de Mann, Rogosa, and Sharpe (MRS) medium in in vitro studies of LAB antagonism against molds [85,86], which may contribute to obtaining overestimated results regarding the antifungal activity of the isolated strains. It has been shown that sodium acetate of the MRS medium contributes greatly to the antifungal effect [59,87,88]. Studies by Stiles et al. [87] revealed an inhibitory effect of sodium acetate on the growth of 33 out of 42 mold strains from the genera Fusarium, Penicillium, Aspergillus, and Rhizopus. The combination of L. rhamnosus VT1 and sodium acetate exerted a synergistic inhibitory effect against 39 mold strains. Fraberger et al. [89] suggested that the MRS medium is a good stimulator of the production of antifungal compounds. In our work (unpublished data), the exclusion of sodium acetate from the MRS medium abolished the antagonistic activity of most LAB that inhibited fungi on the standard medium.
The effect of sodium acetate can be explained by the displacement of weak acids from their salts by stronger acids. It is known that the strength of organic acids depends on the length of their hydrocarbon radical, decreasing with its elongation. However, the acidic properties of organic acids are enhanced by electron-withdrawing substituents [90]. This is why lactic acid is more acidic than acetic acid, although its radical is longer. The pKa value of acetic acid is 4.76, while the pKa value of lactic acid is 3.86, and strong acids have pKa values less than zero [91. These data indicate that the production of lactic acid in a medium containing sodium acetate will lead to the accumulation of undissociated acetic acid. In our studies (unpublished data), GS-MC of homofermentative LAB volatiles in MRS did not reveal lactic acid but indicated the accumulation of acetic acid. Anyway, high production of organic acids helps to suppress the growth of fungal microorganisms, so the selection of active acid formers on a medium with sodium acetate can also be effective, which was confirmed by extended storage of bread for a week in the experiments of Bartkiene et al. [92] without intentional contamination.
Debonne et al. [25] demonstrated the importance of undissociated acids in improving the shelf life of bread. In their studies, air-packed bread prepared with 30% sourdough, fermented by L. sanfranciscensis and S. cerevisiae, did not deteriorate over the entire 7-week observation period. Sourdough bread contained 36 mmol undissociated lactic acid and 220 mmol undissociated acetic acid. It was also shown that the main effect depended on undissociated acetic acid and lactic acid was insignificant in inhibiting fungal growth, only contributing to the overall acidification. A concentration of 150-200 mmol undissociated acetic acid per liter of the aqueous phase increased the shelf life of the bread. It has also been shown that different fungi are sensitive to the effects of acetic acid to varying degrees. Thus, the growth of A. niger was completely inhibited in chemically acidified bread at an acetic acid concentration above 33 mmol/kg dough (which corresponds to 165 mmol undissociated acetic acid in one liter of the aqueous phase of bread), while the growth of Penicillium paneum was inhibited only at a concentration above 100 mmol/kg dough [93]. In another study by Debonne et al. [94], the effect of undissociated acetic acid and phenyllactic acid on the growth of P. paneum and A. niger fungi and the shelf life of semi-baked bread was assessed. The minimum inhibitory concentration (MIC) of undissociated acetic acid in bread was 110–169 mM, which was sufficient to prevent mold growth for 45 days. The MIC of phenyllactic acid was 39–84 mmol/L aqueous phase. It was concluded that the naturally occurring phenyllactic acid in sourdough bread is not sufficient to extend the shelf life of bread and that acetic acid is the most important antifungal agent in sourdough bread. The MIC value for phenyllactic acid is consistent with the previously reported MIC value of 7.5 mg ml-1 (45 mmol/l) against A. fumigatus and P. roqueforti by Ström et al. [95]. In the study by Lavermicocca et al. [96] MIC of phenyllactic acid was 8.3 mg/disk against P. roqueforti IBT18687 and 2.5 mg/disk against Endomyces fibuliger. Unfortunately, MIC data of phenyllactic acid against other fungi are not presented in the work, despite the information on the suppression of a wide range of mold fungi (Eurotium repens, Eurotium rubrum, P. corylophilum, P. roqueforti, P. expansum, Endomyces fibuliger, A. niger, A. flavus, Monilia sitophila, and F. graminearum) by 10-fold-concentrated L. plantarum 21B culture filtrate in wheat flour hydrolysate. Maximum antifungal activity was found in the ethyl acetate extract of the culture filtrate, which contained the highest concentration of phenyllactic acid. However, the authors reported no synergistic effect between the MIC of phenyllactic acid and non-inhibitory concentrations of p-hydroxyphenyllactic and palmitic acids.
Formic acid at 24.7 mM, formed during fermentation of wheat germ by LAB L. plantarum LB1 and F. rossiae LB5, showed high antifungal activity and acted in a mixture with four peptides [97]. Propionic acid is determined among LAB metabolites much less frequently than lactic and acetic acids.
However, a stronger inhibitory effect of propionic acid (MIC from 8 to 20 mmol/liter) was shown, compared to acetic acid (MIC from 23 to 72 mmol/liter) against A. niger, P. corylophilum, and E. repens [98]. Acetate and propionate produced by Lentilactobacillus buchneri and Lentilactobacillus diolivorans inhibited bread spoilage by A. clavatus, Cladosporium spp., and Mortierella spp. in the study by Zhang et al. [99]. At the same time, increasing the proportion of sourdough from 10% to 20% also inhibited the growth of baker's yeast S. cerevisiae in dough. Therefore, the dosage of antifungal compounds should be adjusted to take into account the effect not only on molds but also on the yeast used in bread production.
The list of antifungal organic acids produced by LAB in soudfough includes lactic, acetic, formic, propanoic, hydrocinnamic acid, phenyllactic and hydroxyphenyllactic, butyric, n-valeric, 2-hydroxyisocaproic, capric, ρ-coumaric, (E)-2-methylcinnamic, azelaic, gallic, caffeic, ferulic, hydrocaffeic, hydroferulic, syringic, sinapic, monohydroxy octadecenoic, phloretic, salicylic, vanillic, and other acids [2,6,7,14,47,100,101,102] (Table 1).
However, despite the wide range of LAB antifungal compounds, their detected concentrations are often lower than the known MICs [47]. At the same time, combinations of LAB organic acids can have a synergistic antifungal effect. However, the contribution of different acids to the overall synergistic effect is not the same. Thus, Corsetti et al. [103] showed that F. sanfranciscensis suppressed A. niger, F. graminearum, P. expansum, and M. sitophila to varying degrees. The antifungal effect was synergistic between acetic, caproic, formic, propionic, butyric, and n-valeric acids. These acids did not inhibit the growth of the studied fungus F. graminearum individually in the produced concentrations, but at a dose corresponding to the sum of all acids (16 mM), each acid gave inhibition halos. The inhibition zones of F. graminearum were maximum when using all six acids. However, the decisive role in the manifestation of the antifungal effect was played by caproic acid, followed by acetic acid. The input of other acids in increasing antifungal activity was weak.
The water-soluble extract of L. bulgaricus contained phenyllactic and sinapic acids [101], while L. plantarum produced several phenolic acids, namely gallic, caffeic, chlorogenic (an ester of caffeic and quinic acids), and syringic acids. Chlorogenic acid was also detected in wheat-whey dough fermented by L. bulgaricus. Both species produced vanillic acid in wheat-whey dough. Water-soluble extracts from wheat dough fermented by L. plantarum inhibited P. expansum, P. roqueforti, P. camemberti, F. moniliformis, F. graminearum, F. verticillioides, A. niger, and A. parasiticus. L. bulgaricus extracts were active only against Fusarium species and P. expansum. However, the MICs of the organic acids were lower than those of L. plantarum. A. flavus was also inhibited by both LAB when whey powder was added to the dough. Bread contaminated with P. expansum baked with L. bulgaricus had a shelf life one day longer than that baked with L. plantarum. These results suggest that phenyllactic and sinapic acids were more effective in inhibiting P. expansum, while the other identified phenolic acids contained in the L. plantarum extract were better inhibitors of P. camemberti, P. roqueforti, and A. parasiticus.
Often, the most significant factor in inhibiting the growth of fungal microorganisms is the production of organic acids. Thus, neutralization of LAB cell-free supernatants led to a reduction in the zones of inhibition of mold fungi P. chrysogenum, F. graminearum, for Rhizopus stolonifer and Aspergillus nidulans by 67.0–77.4% [108]. The use of neutralization of cell-free supernatants to exclude the effect of medium pH [72] allows the selection of microorganisms with other mechanisms of action. Axel et al. [47] also showed that chemical acidification with a mixture of lactic and acetic acids does not allow achieving the activity level of sourdough fermented by L. amilovorus DSM19280, L. brevis R2Δ, and L. reuteri R29.
At the same time, medium-chain fatty acids may affect fungal cells not by lowering the pH, but mainly due to the fatty acid radical, which, probably having surface activity, has a direct damaging effect on cell membranes. Thus, sodium decanoate had the lowest MIC among all the studied compounds of L. amylovorous DSM 19280, which also included carboxylic acids, cyclic-dipetides, and nucleosides [105], and Sadeghi et al. [104] showed the suppression of A. niger by the active fraction of L. reuteri containing n-decanoic, 3-hydroxydecanoic, and 3-hydroxydodecanoic acids.
Thus, LAB associated with bread sourdoughs were found to produce several organic acids, primarily saturated aliphatic acids with a short carbon chain, and a large set of acids containing hydroxy and phenolic substituents, including those with an unsaturated carbon skeleton. Benzoic acids were also noted.
Suppression of bread spoilage fungi by organic acids strongly depends on the level of their LAB production. As studies show, the levels of LAB organic acid production vary significantly. Some studies have shown the leading contribution to the suppression of mold fungi by such acids as acetic, 3-phenyllactic, lactic, caproic, propionic, and formic. Other acids were more often produced in concentrations below MICs and had only an auxiliary effect.
3.1.2. Other Compounds
Except for organic acids, LAB antifungal compounds also include hydrogen peroxide, reuterin, diacetyl, peptides and cyclic dipeptides, proteinaceous compounds, and phenolic compounds [6,7,14,47,100,102,106]. The main effect of LAB is synergistic between pH and other antifungal metabolites [7,14]. Bacteriocins produced by LAB can also affect fungi. Thus, Ai et al. [109] showed that nisin of L. lactis was retained during baking and reduced fungal contamination. Other researchers have also demonstrated the antifungal activity of nisin [110,111]. L. plantarum is known to produce a two-peptide bacteriocin, plantaricin [112], which may be effective against Gram-positive bacteria of the genus Bacillus [113,114] and molds of the genera Aspergillus, Fusarium, Mucor, and Penicillium [115].
Reuterin (3-hydroxypropionaldehyde) is specific to L. reuteri and likely the reason for the more extensive study of this microorganism (Section 2.1, Figure 2). However, in the studies of Schmidt et al. [107], the antifungal activity of reuterin from L. reuteri R29 was not transferred to the bread system. On the other hand, the increased accumulation of phenyllactic acid by this microorganism affected the shelf life of bread. Another factor explaining the extensive study of L. reuteri in bread making is the production of the exopolysaccharide reuteran, which improves bread volume and texture [116].
Diacetyl was also not detected in sourdoughs at levels above the MIC [77].
Studies have shown [117] that short-chain organic acids produced by LAB, such as formic, acetic, propionic, oxalic, and lactic acids, exert a synergistic effect with H2O2 accumulated by LAB due to their lack of catalase [118].
The accumulation of acids in sourdough is often noted as a factor acting in concert with protein and peptide compounds. Thus, Illueca et al. [9] showed increased concentrations of lactic and phenolic acids in breads fermented with L. plantarum with simultaneous hydrolysis of proteins (50 kDa). In turn, Hernandez-Figueroa et al. [119] identified protein fractions in wheat sourdough fermented with L. plantarum NRRL B-4496, in addition to lactic and acetic acids, with high antifungal activity against P. chrysogenum and P. corylophilum. The molecular weight of the antagonistically active fraction was greater than 30 kDa. It has been shown [120] that L. plantarum-based sourdough is a good natural alternative to sodium propionate as an antifungal agent in bread. Some protein-like compounds of L. fermentum and P. pentosaceus prevented Aspergillus spoilage of bread [51], while compounds less than 10 kDa were responsible for the antifungal activity of L. mesenteroides and L. plantarum at pH 3 [121].
In another study [122], antifungal metabolites of P. pentosaceus from wheat sourdough included a fatty acid ester, a hydroxylated fatty acid ester, 3-isobutyl 2,5 piperazinedione, and a cyclic dipeptide. It was previously shown [95] that L. plantarum can produce antifungal cyclic dipeptides (diketopiperazines) cyclo(L-Phe–L-Pro) and cyclo(L-Phe–trans-4-OH-L-Pro). MIC for cyclo(L-Phe–L-Pro) against A. fumigatus and P. roqueforti was 20 mg ml-1. This cyclic dipeptide revealed a weak synergistic effect in combination with phenyllactic acid, with both compounds at concentrations below their MICs. In turn, Dal Bello et al. [123] isolated two L. plantarum antifungal dipeptides: cyclo (L-Leu-L-Pro) and cyclo (L-Phe-L-Pro). These peptides were active against bread spoilage Fusarium culmorum and F. graminearum. Later, Ryan et al. (2011) identified five antifungal cyclic dipeptides in L. amylovorous DSM 19280 broth that were effective against A. fumigatus at concentrations >25–50 mg mL−1. All compounds contained proline as one of the amino acids. In addition to cyclo (L-Leu-L-Pro), the cyclic dipeptides also included cyclo(L-His-L-Pro), cyclo (L-Pro-L-Pro), cyclo (L-Met-L-Pro), and cyclo (L-Tyr-L-Pro). The study by Ryan et al. [105] simultaneously demonstrated antifungal activity of high doses (>200 mg mL−1) of cytidine nucleosides and 2′-Deoxycytidine of L. amylovorous against A. fumigatus. In earlier work, Ryan et al. [124] showed that cyclic dipeptides are not LAB metabolic products, since the amount of diketopiperazines in acidified dough did not differ from that in dough obtained using sourdough. Axel et al. [100] also showed that L. brevis R2Δ also produces cyclic dipeptides such as cyclo(Leu-Pro), cyclo(ProPro) and cyclo(Phe-Pro).
Nionelli et al. [125] detected nine antifungal peptides with a molecular weight of about 1.1-1.2 kDa and 10-17 amino acid residues during the hydrolysis of bread residues by L. brevis AM7. All these peptides were encrypted into the sequences of wheat proteins. Previously, Coda et al. [126] also detected nine antifungal peptides in a water-soluble extract obtained from dough fermented by L. plantarum 1A7 (S1A7) and effective against P. roqueforti. The identified peptides with a molecular weight from 1.5 to 5.6 kDa were encrypted into sequences of Oryza sativa proteins and contained 14-53 amino acid residues. The MICs of identified peptides varied from 2.5 to 11.2 mg mL−1.
It is known that endogenous wheat proteinases have an optimum pH of 3.0–4.0 [127]. Therefore, acidification of dough to the optimum value by LAB promotes activation of endogenous proteinases and hydrolysis of wheat proteins with the formation of biologically active peptides. The wheat gluten is a complex mixture of hundreds of proteins, the main ones being gliadin and glutenin [128]. Garofalo et al. [77] found several peptides related to wheat α-gliadin in the active fractions of dough extracts obtained using F. rossiae LD108. L. rossiae LD108 and Lactobacillus paralimentarius PB127 exhibited antagonism against the bread spoilage fungi A. japonicus, E. repens, and Penicillium roseopurpureum. At the same time, the detected acetic acid, phenyllactic acid, and diacetyl were in concentrations below MICs. In the studies of Rizzello et al. [97] in the fermentation of wheat germs, antifungal peptides were found in various proteins (formin-like protein 4, homeobox-leucine zipper protein HOX2, expansin-B4, and probable cation transporter HKT6) with sequences from 7 to 52 amino acids. The MICs of defined peptides ranged from 2.5 to 15.2 mg mL−1.
Considering the above information, it can be assumed that a significant reason for the accelerated spoilage of gluten-free bread [2] may be the absence of proteins capable of being hydrolyzed to antifungal peptides.
Thus, LAB produce a whole list of antifungal compounds, mainly a wide range of organic acids, which can either have a direct damaging effect in an undissociated state inside the cell or damage cell membranes, or activate flour proteinases by lowering the pH with subsequent hydrolysis of its proteins and the production of antifungal peptides.
At the same time, there are also results showing the lack of effectiveness of 10% sourdough fermented with F. sanfranciscensis in extending the shelf life of bread contaminated with A. clavatus and P. roqueforti [129]. Only a decrease in the aW value to 0.92 and the addition of propionate contributed to an increase in the shelf life of contaminated bread. There is also evidence that the visual absence of mold growth is not always accompanied by complete suppression and can simultaneously increase the production of mycotoxins [130]. The research results show the need for careful selection of starter microorganisms to receive positive results and control the level of mycotoxins in the environment.
3.1.3. Mycotoxin Removal
Despite the reduction of mycotoxin levels during bread baking, their concentrations may still exceed the maximum permissible levels at high contamination [130,131]. At high bread consumption, mycotoxin exposure in adults and children may exceed the guideline values [132]. In these cases, the use of sourdough is a good option to improve the safety of contaminated flour. Several studies have shown the possibility of reducing mycotoxin levels by sourdough LAB [9,130,131,133]. For example, Cao et al. [134] showed that L. plantarum AR524 inhibited the growth of F. graminearum by 60.19% compared to the control, while removing up to 40.9% of deoxynivalenol (DON). Mycotoxin removal occurred primarily through cell wall binding. In the studies of Badji et al. [135], in vitro removal of aflatoxin B1 and ochratoxin A was reported using both viable and nonviable LAB cells of L. lactis ssp. lactis, L. paracasei ssp. paracasei, Enterococcus faecium, and Enterococcus durans. The mycotoxin removal efficiency was strain-specific, with aflatoxin B1 being better bound by nonviable cells. In the studies of Illueca et al. [9], L. reuteri was also effective in aflatoxin removal.
The duration of fermentation has been shown to affect the removal of mycotoxins. After 48 h, the level of DON in the sourdough was reduced by 44–69% in highly contaminated grain [136]. At the same time, toxins such as 15-acetyldeoxynivalenol, alternariol, deoxynivalenol-3-glucoside, H-2, and HT-2 were completely removed, while the reduction of enniatin depended on its form.
In the investigation of Lafuente et al. [66], a high effect in reducing aflatoxin content in experimentally contaminated bread was shown by dry sourdough fermented with P. pentosaceus TI6. Although the main mechanism of mycotoxin removal by LAB is cell wall absorption, their biotransformation into non-toxic by-products is also possible [102,130]. At the same time, since the mycotoxin content is higher in the outer grain layers, bran breads may contain increased amounts of mycotoxins [137,138]. The studies by Vidal et al. [137] did not provide a clear answer regarding the effect of sourdough on the mycotoxin levels in bran breads. In some cases, an increase in toxin levels was noted. Therefore, further research in this area is needed.
3.2. Antibacterial Activity
Rope disease is caused by amylase-producing spore-forming bacteria belonging to Bacillus subtilis and other closely related genera and species that survive in bread during baking and degrade bread polysaccharides to form mucus, for example, Bacillus amyloliquefaciens, B. licheniformis, the B. cereus group, B. pumilus, B. sonorensis, Cytobacillus firmus, Niallia circulans, Paenibacillus spp., Lysinibacillus spp., and Priestia megaterium [10,139]. Due to consumer preference for preservative-free products, a re-emergence of the rope disease problem is expected. Global warming exacerbates this problem due to the abundant development of temperature- and desiccation-resistant endophytic microbiota in cereals [10].
The whole grain flour is a rich source of spore-forming bacteria, causing rope-like spoilage of bread. Thus, 45 out of 327 isolated bacterial strains produced amylase and caused rope-like spoilage in the study of Pereira et al. [140]. B. licheniformis accounted for 62% of the identified microorganisms, followed by B. sonorensis (20%) and B. cereus (11%), while B. pumilus and Paenibacillus polymyxa accounted for only 2% of isolates each.
Fraberger et al. [89] showed inhibition of B. subtilis, B. cereus, and B. licheniformis by most LAB. The antibacterial effect was strain-specific. Different strains of L. plantarum, F. sanfranciscensis, Loigolactobacillus coryniformis, and L. paracasei were effective to varying degrees against the studied Bacillus species. In other studies, L. paraplantarum and Pediococcus acidilactici from spontaneous fermentation of einkorn wheat showed high antagonism against B. subtilis and B. cereus [141], while W. cibaria from spontaneous fermentation of chia flour suppressed B. subtilis [142]. In turn, Iosca et al. [139] isolated L. plantarum, P. pentosaceus, and L. citreum, which were highly antagonistic against the agents of ropy bread spoilage. The studied LABs had operons for bacteriocins, but phenotypic confirmation of their production was not obtained. All LABs from 12 typical Bulgarian sourdoughs [108] also suppressed bread spoilage by B. subtilis. From them, L. plantarum, two strains of L. brevis, and E. durans showed the highest antagonism with growth inhibition zones of 65–74 mm. However, neutralization of CFS reduced antagonism by 33–52%. At the same time, denaturation of protein compounds by boiling and treatment with trypsin reduced antagonism in almost all LAB strains to an even greater extent. The results indicate that protein antimicrobial compounds make the greatest contribution to the antibacterial activity of LAB, followed by organic acids. Li et al. [143] showed that the use of sourdough inhibits the germination of Bacillus spores in bread, an effect enhanced by reutericyclin produced by L. reuteri.
4. Contribution of Other Microorganisms to Protecting Bread from Spoilage
4.1. Yeast
The yeasts used in sourdough are also capable of inhibiting the growth of mold fungi. The contribution of Wickerhamomyces anomalus yeast is most often noted. Thus, in the studies of Pahlavani et al. [144], W. anomalus inhibited the growth of A. flavus in challenge dough, especially on wheat bread with the addition of spontaneously fermented barley (type IV sourdough). In the studies of Coda et al. [126], W. anomalus yeast also contributed to the antifungal activity of sourdough with LAB by producing ethanol and ethyl acetate. Germination of P. roqueforti conidia was not observed at ethanol and ethyl acetate concentrations of 1.69 (36.6 mM) and 6.81 (77.0 mM) mg/ml, respectively. Syrokou et al. [72] also selected 12 out of 195 yeast strains from spontaneously fermented Greek wheat sourdoughs that inhibited the growth of P. chrysogenum by producing ethyl acetate. These yeasts were assigned to S. cerevisiae, W. anomalus, and Pichia fermentans. In turn, the yeast Pichia kudriavzevii, which inhibited the growth of A. niger and A. flavus, was isolated from buckwheat sourdough by Shahryari et al. [145]. There is also evidence of a reduction in mycotoxin levels when using yeast. Thus, compressed baker's yeast showed better results in reducing ochratoxin levels in bread compared to other forms [146].
4.2. Acetic Acid Bacteria
Since AAB are often found in the spontaneous microbiota of sourdoughs, the possibility of using them as part of sourdough microorganisms was investigated. Thus, Kozakia baliensis and Neoasaia chiangmaiensis were shown to produce polysaccharides in dough from wheat, whole grain, spelt, and rye flours [147]. The polysaccharide content in the dough exceeded 30 g/kg, which will allow the use of one-tenth to one-third of the sourdough in the dough. At the same time, the acetic acid yield of K. baliensis was 2-3 times higher (up to 100 mM/kg) than that of lactobacilli. The obtained data may indicate the prospects of using AAB in sourdoughs to improve the shelf life of bread. This assumption is supported by positive results in preventing mold formation and extending the shelf life of bread when using kombucha as a starter [148,149]. A. pasteurianus and Gluconobacter oxydans isolated from fermented cocoa pulp-bean mass also revealed promise as sourdough starter cultures [48]. Increased dough acidification when adding AAB to the starter [41] and an increase in acetic acid production [150] may contribute to improved shelf life of bread.
5. New Strategies for Using Bread Sourdoughs
In recent years, additives of various components that improve the quality and shelf life of sourdough bread have been actively studied. The potential of legumes, pseudo-legume pseudo-cereals, and other additives in the development of new bakery products is noted [21,151]. This area is particularly relevant in connection with the need to ensure a balanced diet and transition to a diet with a higher content of plant proteins to reduce greenhouse gas emissions [151,152,153].
Most often, additives are pre-fermented and used as sourdough in bread making. Sourdoughs based on non-standard flours have not yet been studied much due to their wide variety. In the study of such starters and additives, special attention is paid to increasing the nutritional value and digestibility of the final products, as well as the content of biologically active components in them [154,155,156]. For example, it was shown that pea sourdough increased the digestibility of protein in traditional durum wheat focaccia, reducing starch digestibility [157]. In turn, fermented mixtures of wheat and legume flours (green lentils, fava beans, chickpeas) were characterized by a significant increase in antioxidant activity [158]. The best effect was shown by sprouted lentils fermented with L. casei and Kluyveromyces marxianus. Increases in antioxidant activity and total phenolic content have been observed with replacement of up to 15% of wheat flour with flour derived from legume seeds (grass pea, yellow lupine, and narrow-leaf lupine) [159], with the addition of 25% yellow pea flour to gluten-free wholemeal rice flour [160], and in various mixtures of wheat flour and legumes [161].
There is evidence of antimicrobial activity in microorganisms isolated from spontaneously fermented substrates. Thus, W. cibaria was isolated from spontaneously fermented chia sourdough, inhibiting the growth of B. subtilis, P. roquefortii, and A. niger. Enterococcus casseliflavus, E. faecium, and L. lactis CH179 inhibited the growth of both fungi, and L. rhamnosus and other strains of L. lactis supressed the growth of only one mold [142].
A large body of data has been obtained on the effect of non-standard starters and additives on the preservation of bread. Sourdough based on einkorn, oat, hull-less barley, wheat germ, fermented sprouted mung bean and lentil, soya flour and rice bran, chickpea flour, faba bean flour, quinoa and red lentil, amaranth, pitaya fruit, cornelian cherry, chokeberry and pomegranate juices were used to improve the quality, nutritional value and preservation of bread (Table 2). Among the mechanisms of action studied, organic acids, including phenolic, and antifungal peptides were noted. Similar to the origin of the peptides encrypted into flour gliadin found during wheat flour fermentation, Verni's study showed the origin of antifungal peptides from vicilin and legumin of faba beans [162].
The use of various food wastes has been recently investigated to recycle waste and by-products and mitigate the impact on the environment, as well as for imparting new properties to bread, namely increased nutritional value and shelf life. Thus, the use of barley rootlets (a by-product of the malting and brewing industry) fermented with W. cibaria, L. citreum, L. plantarum, L. amylovorus, and L. reuteri significantly slowed down the spoilage of bread [183].
Dopazo et al. [184] showed that the use of 5% whey fermented with LAB L. plantarum increased the content of lactic and phenyllactic acids and allowed extension shelf life of bread contaminated with A. flavus and Penicillium verrucosum. The technological properties of the bread were not significantly changed. In the studies of Luz et al. [185], a higher amount of whey from Mozzarella di Bufala Campana was used (replacement of 50% and 100% water). Naturally contaminated bread with 100% water replaced by whey fermented with L. plantarum or L. ghanensis was characterized by an increase in shelf life by 15 days compared to the control. The fermentation of LAB whey significantly increased the amount of acetic, hexanoic and octanoic acids. In breads with complete replacement of water with whey, the content of total phenol compounds and antioxidant activity (in DPPH radical-scavenging assay) increased significantly, especially when fermenting whey with L. plantarum compared to L. ghanensis and unfermented whey. In another study [186], fermented L. plantarum whey was used to prepare pita bread. When inoculating the bread surface with P. expansum and Penicillium brevicompactum, the shelf life of the bread increased to 8 days, and with natural inoculation to 19 days. However, an untrained taste panel detected no differences between the control and whey-produced pita samples.
The use of edible coatings to protect against microbial spoilage in bread baking is a new trend. Coatings include LAB in some carrier or plant materials with antimicrobial and antioxidant properties. Research by Bartkiene et al. [187] showed that the additional use of apple pomace coatings further extended the shelf life of sourdough bread with Loigolactobacillus coryniformis, L. curvatus, Lentilactobacillus farraginis, and L. mesenteroides. In another study, cranberry coating of bread with P. pentosaceus, P. acidilactici, L. paracasei, L. brevis, Lactobacillus plantarum, and L. mesenteroides increased antifungal activity against Aspergillus fischeri, Penicillium oxalicum, Penicillium funiculosum, Fusarium poae, Alternaria alternate, and F. graminearum [188].
Also shown to be highly effective in extending the shelf life of pan breads and muffins is the use of chitosan nanoenzymes of LAB L. plantarum, L. helveticus, and L. rhamnosus in the last stage of fermentation and as a coating after baking [189]. Streptococcus salivarius subsp. thermophilus, Lactobacillus delbrueckii subsp. bulgaricus, L. acidophilus were used in studies with alginate and whey, which protected bread from contamination by filamentous fungi of the genera Aspergillus and Penicillium [190]. The addition of LAB to a pectin-based coating to protect bread from A. flavus, A. niger, and P. paneum was also used by Iosca et al. [191]. L. plantarum, F. rossiae, and P. pentosaceus were sprayed onto the coating surface in this study
Recent trends based on more detailed studies of sourdough microbiomes include the construction of sourdough communities accounting for the metabolic contributions of individual microorganisms. This is a very new approach attempting to identify dominant, subdominant, and satellite species that can jointly provide gene and transcript redundancy in the sourdough [192]. At present, the theoretical foundations for a new generation of sourdoughs, combining multiple microorganisms that are maximally adapted to specific food matrices, are only just being laid.
6. Conclusions
Bread protection from spoilage remains an unsolved problem for humanity at this stage. The desire for clean label products forces researchers to search for and constantly improve safe ways to protect bread. The use of sourdoughs in protecting bread from spoilage is widely studied, but the huge variety of fermentable substrates and local features of the microorganisms inhabiting them, including those causing spoilage, do not allow us to put an end to these studies. The development of modern research methods reveals new features of the interactions of sourdough microorganisms and the mechanisms of their action. To date, the antimicrobial and especially antifungal effects of lactic acid bacteria in starters have been linked to their production of a wide range of organic acids, both aliphatic and those with hydroxyl- and phenyl- substituents, as well as benzoic acids, which often act in synergy and promote the hydrolysis of flour proteins to form antifungal peptides. Moreover, such peptides have been identified both in wheat sourdoughs and in starters based on non-traditional substrates, being sequences embedded in specific proteins of various types of flour. However, data on antifungal peptides of sourdoughs based on various types of flour are still very limited, their study is a promising direction. The mechanisms of action of antifungal compounds in sourdoughs are still poorly understood. The influence of various non-traditional additives and substrates on the antimicrobial activity of sourdoughs is widely studied. There are still many gaps in this area due to the wide variety of substrates and their microbiota. A current trend is the use of food waste and by-products for baking. Thus, the use of whey in sourdough fermentation has shown encouraging results. There is also a clear trend to expand the list of used starter microorganisms using non-traditional genera of LAB and yeast, such as Weissella and Wickerhamomyces, and AAB, which are constant and almost unstudied companions of LAB and yeast in the fermentations they carry out. The selection of a variety of microorganisms that best perform significant functions in dough matrices from different types of flour, taking into account the metabolic contributions of dominant, subdominant, and satellite species, is a completely new and promising direction in the research and development of sourdoughs.
Author Contributions
Conceptualization, writing—original draft preparation, project administration, Y.O.; writing—review and editing, M.S., A.S., A.A., A. Z.; formal analysis, M.S.; A.S.: funding acquisition; data curation, A.A., A.Z.
Funding
The research was supported by the Science Committee of the Ministry of Science and Higher Education of the Republic of Kazakhstan (Grant No. AP19674612).
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- The Federation of Bakers. European Bread Market. Available online: https://www.fob.uk.com/about-the-bread-industry/industry-facts/european-bread-market/ (accessed on 28 June 2025).
- Axel, C.; Zannini, E.; Arendt, E.K. Mold spoilage of bread and its biopreservation: A review of current strategies for bread shelf life extension. Crit. Rev. Food Sci. Nutr. 2017, 57(16), 3528–3542. [CrossRef]
- Nasrollahzadeh, A.; Mokhtari, S.; Khomeiri, M.; Saris, P.E.J. Antifungal Preservation of Food by Lactic Acid Bacteria. Foods 2022, 11, 395. [CrossRef]
- Garcia, M.V.; Bernardi, A.O.; Parussolo, G.; Stefanello, A.; Lemos, J.G., Copetti, M.V. Spoilage fungi in a bread factory in Brazil: Diversity and incidence through the bread-making process. Food Res. Int. 2019a, 126, 108593. [CrossRef]
- Garcia, M.V.; Bernardi, A.O.; Copetti, M.V. The fungal problem in bread production: Insights of causes, consequences, and control methods. Curr. Opin. Food Sci. 2019b, 29, 1–6. [CrossRef]
- Awulachew, M.T. Bread deterioration and a way to use healthful methods. CyTA J. Food, 2024, 22(1): 2424848. [CrossRef]
- Hernández-Figueroa, R.H.; Mani-López, E.; Palou, E.; López-Malo, A. Sourdoughs as Natural Enhancers of Bread Quality and Shelf Life: A Review. Fermentation 2024a, 10, 7. [CrossRef]
- Lemos, J.G.; Silva, L.P.; Mahfouz, M.A.A.R.; Cazzuni, L.A.F.; Rocha, L.O.; Steel, C.J. Use of dielectric-barrier discharge (DBD) cold plasma for control of bread spoilage fungi. Int. J. Food Microbiol. 2025, 430, 111034. [CrossRef]
- Illueca, F.; Moreno, A.; Calpe, J.; Nazareth, T.d.M.; Dopazo, V.; Meca, G.; Quiles, J.M.; Luz, C. Bread biopreservation through the addition of lactic acid bacteria in sourdough. Foods 2023, 12, 864. [CrossRef]
- Pacher, N.; Burtscher, J.; Johler, S.; Etter, D.; Bender, D.; Fieseler, L.; Domig, K.J. Ropiness in Bread—A Re-Emerging Spoilage Phenomenon. Foods 2022, 11, 3021. [CrossRef]
- EFSA. Scientific Opinion on the maintenance of the list of QPS biological agents intentionally added to food and feed (2012 update). EFSA J. 2012, 10:3020. [CrossRef]
- Zheng J.; Wittouck S.; Salvetti E.; Franz C.M.A.P.; Harris H.M.B.; Mattarelli P.; O’Toole, P.W.; Pot B.; Vandamme P.; Walter, J.; Watanabe, K.; Wuyts, S.; Felis, G.E.; Gänzle, M.G.; Lebeer, S. A taxonomic note on the genus Lactobacillus: description of 23 novel genera, emendeddescription of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [CrossRef]
- Lau, S.W.; Chong, A.Q.; Chin, N.L.; Talib, R.A.; Basha, R.K. Sourdough Microbiome Comparison and Benefits. Microorganisms 2021, 9, 1355. [CrossRef]
- De Vero, L.; Iosca, G.; Gullo, M.; Pulvirenti, A. Functional and Healthy Features of Conventional and Non-Conventional Sourdoughs. Appl. Sci. 2021, 11, 3694. [CrossRef]
- De Bondt, Y.; Verdonck, C.; Brandt, M.J.; De Vuyst, L.; Gänzle, M.G.; Gobbetti, M.; Zannini, E.; Courtin, C.M. Wheat sourdough breadmaking: a scoping review. Ann. Rev. Food Sci. Technol. 2024, 15: 265–282. [CrossRef]
- De Angelis, M.; Minervini, F.; Siragusa, S.; Rizzello, C.G.; Gobbetti, M. Wholemeal wheat flours drive the microbiome and functional features of wheat sourdoughs. Int. J. Food Microbiol. 2019, 302, 35–46. [CrossRef]
- Boreczek, J.; Litwinek, D.; Żylińska-Urban, J.; Izak, D.; Buksa, K.; Gawor, J.; Gromadka, R.; Bardowski, J.K.; Kowalczyk, M. Bacterial community dynamics in spontaneous sourdoughs made from wheat, spelt, and rye wholemeal flour. MicrobiologyOpen, 2020, 9(4), e1009. [CrossRef]
- Oshiro, M.; Tanaka, M.; Zendo, T.; Nakayama, J. Impact of pH on succession of sourdough lactic acid bacteria communities and their fermentation properties. Biosci. Microbiota Food Health, 2020, 39(3), 152–159. [CrossRef]
- Suo, B., Chen, X., Wang, Y. Recent research advances of lactic acid bacteria in sourdough: origin, diversity, and function. Curr. Opin. Food Sci. 2021, 37, 66–75. [CrossRef]
- Khlestkin, V.K.; Lockachuk, M.N.; Savkina, O.A.; Kuznetsova, L.I.; Pavlovskaya, E.N.; Parakhina, O.I. Taxonomic structure of bacterial communities in sourdoughs of spontaneous fermentation. Vavilov J. Genetics Breeding, 2022, 26(4), 385. [CrossRef]
- Lee, S.H.; Jung, J.Y.; Jeon, C.O. Source tracking and succession of kimchi lactic acid bacteria during fermentation. J. Food Sci. 2015, 80(8), M1871–M1877. [CrossRef]
- Romi, W.; Ahmed, G.; Jeyaram, K. Three-phase succession of autochthonous lactic acid bacteria to reach a stable ecosystem within 7 days of natural bamboo shoot fermentation as revealed by different molecular approaches. Mol. Ecol. 2015, 24(13), 3372–3389. [CrossRef]
- Wang, Y.; Zhang, C.; Liu, F.; Jin, Z.; Xia, X. Ecological succession and functional characteristics of lactic acid bacteria in traditional fermented foods. Crit. Rev. Food Sci. Nutr. 2023, 63(22), 5841–5855. [CrossRef]
- Baev, V.; Apostolova, E.; Gotcheva, V.; Koprinarova, M.; Papageorgiou, M.; Rocha, J.M.; Yahubyan, G.; Angelov, A. 16S-rRNA-Based Metagenomic Profiling of the Bacterial Communities in Traditional Bulgarian Sourdoughs. Microorganisms 2023, 11, 803. [CrossRef]
- Debonne, E.; Van Schoors, F.; Maene, P.; Van Bockstaele, F.; Vermeir, P.; Verwaeren, J.; Eeckhout, M.; Devlieghere, F. Comparison of the antifungal effect of undissociated lactic and acetic acid in sourdough bread and in chemically acidified wheat bread. Int. J. Food Microbiol. 2020a, 321, 108551. [CrossRef]
- Weckx, S.; Van der Meulen, R.; Maes, D.; Scheirlinck, I.; Huys, G.; Vandamme, P.; De Vuyst, L. Lactic acid bacteria community dynamics and metabolite production of rye sourdough fermentations share characteristics of wheat and spelt sourdough fermentations. Food Microbiol. 2010, 27(8), 1000–1008. [CrossRef]
- Sanmartín, G.; Sánchez-Adriá, I.E.; Prieto, J.A.; Estruch, F.; Randez-Gil, F. Bioprospecting of sourdough microbial species from artisan bakeries in the city of Valencia. Food Microbiol. 2024, 120, 104474. [CrossRef]
- Minervini F.; Lattanzi A.; De Angelis M.; Di Cagno R.; Gobbetti M. Influence of artisan bakery- or laboratory-propagated sourdoughs on the diversity of lactic acid bacterium and yeast microbiotas. Appl. Environ. Microbiol. 2012, 78(15), 5328–5340. [CrossRef]
- Landis E.A.; Oliverio A.M.; McKenney E.A.; Nichols L.M.; Kfoury N.; Biango-Daniels M.; Shell L.K.; Madden A.A.; Shapiro L.; Sakunala S.; Drake K.; Robbat A.; Booker M.; Dunn R.R.; Fierer N.; Wolfe B.E. The diversity and function of sourdough starter microbiomes. eLife 2021, 10, e61644. [CrossRef]
- Leathers, T.D.; Bischoff, K.M. Biofilm formation by strains of Leuconostoc citreum and L. mesenteroides. Biotechnol. Lett. 2011, 33, 517–523. [CrossRef]
- Yang, Q.; Wang, Y.; An, Q., Sa, R.; Zhang, D.; Xu, R. Research on the role of LuxS/AI-2 quorum sensing in biofilm of Leuconostoc citreum 37 based on complete genome sequencing. 3 Biotech, 2021, 11, 189. [CrossRef]
- Chen, Y.; Gong, X.; Song, J.; Peng, Y.; Zeng, Y.; Chen, J.; Wang, Z.; Li, Z.; Zhu, Y. A novel bio-based film-forming helper derived from Leuconostoc mesenteroides: A promising alternative to chemicals for the preparation of biomass film. Chem. Eng. J. 2024, 493, 152436. [CrossRef]
- Minervini, F.; Celano, G.; Lattanzi, A.; Tedone, L.; De Mastro, G.; Gobbetti, M.; De Angelis, M. Lactic Acid Bacteria in Durum Wheat Flour Are Endophytic Components of the Plant during Its Entire Life Cycle. Appl. Environ. Microbiol. 2015, 81(19), 6736–6748. [CrossRef]
- Gänzle M.G.; Zheng J. Lifestyles of sourdough lactobacilli – Do they matter for microbial ecology and bread quality? Int. J. Food Microbiol. 2019, 302, 15–23. [CrossRef]
- Vogel, R.F.; Pavlovic, M.; Ehrmann, M.; Wiezer, A.; Liesegang, H.; Offschanka, S.; Voget, S.; Angelov, A.; Böcker, G.; Liebl, W. Genomic analysis reveals Lactobacillus sanfranciscensis as stable element in traditional sourdoughs. Microb. Cell. Fact. 2011, 10(Suppl 1), S6. [CrossRef]
- Boiocchi, F.; Porcellato, D.; Limonta, L.; Picozzi, C.; Vigentini, I.; Locatelli, D. P.; Foschino, R. Insect frass in stored cereal products as a potential source of Lactobacillus sanfranciscensis for sourdough ecosystem. J. Appl. Microbiol. 2017, 123(4), 944–955. [CrossRef]
- Comasio, A.; Verce, M.; Van Kerrebroeck, S.; De Vuyst, L. Diverse Microbial Composition of Sourdoughs from Different Origins. Front Microbiol. 2020a, 11, 1212. [CrossRef]
- Xing, X.; Ma, J.; Fu, Z.; Zhao, Y.; Ai, Z.; Suo, B. Diversity of bacterial communities in traditional sourdough derived from three terrain conditions (mountain, plain and basin) in Henan Province, China. Food Res. Int. 2020, 133, 109139. [CrossRef]
- Han, D.; Yang, Y.; Guo, Z.; Dai, S.; Jiang, M.; Zhu, Y.; Wang, Y.; Yu, Z.; Wang, K.; Rong, C.; et al. A Review on the Interaction of Acetic Acid Bacteria and Microbes in Food Fermentation: A Microbial Ecology Perspective. Foods 2024, 13, 2534. [CrossRef]
- Costa, L.F.; Kothe, C.I.; Grassotti, T.T.; Garske, R.P.; Sandoval, B.N.; Varela, A.P.M.; Prichula. J.; Frazzon, J.; Mann, M.B.; Thys, R.C.S.; Frazzon, A.P.G. Evolution of the spontaneous sourdoughs microbiota prepared with organic or conventional whole wheat flours from South Brazil. Anais da Academia Brasileira de Ciências, 2022, 94(suppl 4), e20220091. [CrossRef]
- Rappaport, H.B.; Senewiratne, N.P.; Lucas, S.K.; Wolfe, B.E.; Oliverio, A.M. Genomics and synthetic community experiments uncover the key metabolic roles of acetic acid bacteria in sourdough starter microbiomes. Msystems, 2024, 9(10), e00537-24. [CrossRef]
- Semumu, T.; Zhou, N.; Kebaneilwe, L.; Loeto, D.; Ndlovu, T. Exploring the Microbial Diversity of Botswana’s Traditional Sourdoughs. Fermentation, 2024, 10(8), 417. 10.3390/fermentation10080417.
- Arora, K.; Ameur, H.; Polo, A.; Di Cagno, R.; Rizzello, C.G.; Gobbetti, M. Thirty years of knowledge on sourdough fermentation: A systematic review. Trends Food Sci. Technol. 2021, 108, 71–83. [CrossRef]
- Canesin, M.R.; Betim Cazarin, C.B. Nutritional quality and nutrient bioaccessibility in sourdough bread. Curr. Opin. Food Sci., 2021, 40, 81–86. [CrossRef]
- Pétel, C.; Onno, B.; Prost, C. Sourdough volatile compounds and their contribution to bread: A review. Trends Food Sci. Technol. 2017, 59. 105–123. [CrossRef]
- Warburton, A.; Silcock, P.; Eyres, G.T. Impact of sourdough culture on the volatile compounds in wholemeal sourdough bread. Food Res. Int. 2022, 161, 111885. [CrossRef]
- Axel, C.; Brosnan, B.; Zannini, E.; Peyer, L.; Furey, A.; Coffey, A.; Arendt, E. Antifungal activities of three different Lactobacillus species and their production of antifungal carboxylic acids in wheat sourdough. Appl. Microbiol. Biotechnol. 2016, 100, 1701–1711. [CrossRef]
- Comasio, A.; Van Kerrebroeck, S.; Harth, H.; Verté, F.; De Vuyst, L. Potential of Bacteria from Alternative Fermented Foods as Starter Cultures for the Production of Wheat Sourdoughs. Microorganisms 2020b, 8, 1534. [CrossRef]
- Zhang, C.; Brandt, M.J.; Schwab, C.; Gänzle, M.G. Propionic acid production by cofermentation of Lactobacillus buchneri and Lactobacillus diolivorans in sourdough. Food Microbiol. 2010, 27(3), 390–395. [CrossRef]
- Sun, L.; Li, X.; Zhang, Y.; Yang, W.; Ma, G.; Ma, N.; Hu, Q.; Pei, F. A novel lactic acid bacterium for improving the quality and shelf life of whole wheat bread. Food Control 2020, 109, 106914. [CrossRef]
- Muhialdin, B.J.; Hassan, Z.; Sadon, S.K. Antifungal Activity of Lactobacillus fermentum Te007, Pediococcus pentosaceus Te010, Lactobacillus pentosus G004, and L. paracasi D5 on Selected Foods. J. Food Sci. 2011, 76(7), M493–M499. [CrossRef]
- Zhao, S.; Hao, X.; Yang, F.; Wang, Y.; Fan, X.; Wang, Y. Antifungal Activity of Lactobacillus plantarum ZZUA493 and Its Application to Extend the Shelf Life of Chinese Steamed Buns. Foods 2022, 11, 195. [CrossRef]
- Plessas, S.; Mantzourani, I.; Bekatorou, A. Evaluation of Pediococcus pentosaceus SP2 as Starter Culture on Sourdough Bread Making. Foods 2020, 9, 77. [CrossRef]
- Kazakos, S.; Mantzourani, I.; Plessas, S. Quality Characteristics of Novel Sourdough Breads Made with Functional Lacticaseibacillus paracasei SP5 and Prebiotic Food Matrices. Foods 2022, 11, 3226. [CrossRef]
- Li, H.; Fu, J.; Hu, S.; Li, Z.; Qu, J.; Wu, Z.; Chen, S. Comparison of the effects of acetic acid bacteria and lactic acid bacteria on the microbial diversity of and the functional pathways in dough as revealed by high-throughput metagenomics sequencing. Int. J. Food Microbiol. 2021, 346, 109168. [CrossRef]
- Li H.; Hu S.; Fu J. Effects of acetic acid bacteria in starter culture on the properties of sourdough and steamed bread. Grain Oil Sci. Technol. 2022, 5, 13–21. [CrossRef]
- Chang J.M.; Fang T.J. Survival of Escherichia coli O157:H7 and Salmonella enterica serovars Typhimurium in iceberg lettuce and the antimicrobial effect of rice vinegar against E. coli O157:H7. Food Microbiol. 2007, 24(7-8), 745–751. [CrossRef]
- Ezz Eldin H.M.; Sarhan R.M.; Khayyal A.E. The impact of vinegar on pathogenic Acanthamoeba astronyxis isolate. J. Parasit. Dis. 2019, 43(3), 351–359. [CrossRef]
- Aitzhanova, A.; Oleinikova, Y.; Mounier, J.; Hymery, N.; Leyva Salas, M.; Amangeldi, A.; Saubenova, M.; Alimzhanova, M.; Ashimuly, K.; Sadanov, A. Dairy associations for the targeted control of opportunistic Candida. World J. Microbiol. Biotechnol. 2021, 37, 143. [CrossRef]
- Oleinikova, Y.; Alybayeva, A.; Daugaliyeva, S.; Alimzhanova, M.; Ashimuly, K.; Yermekbay, Zh.; Khadzhibayeva, I.; Saubenova, M. Development of an antagonistic active beverage based on a starter including Acetobacter and assessment of its volatile profile. Int. Dairy J. 2024a, 148, 105789. [CrossRef]
- Oleinikova, Y., Daugaliyeva, S., Mounier, J., Saubenova M., Aitzhanova A. Metagenetic analysis of the bacterial diversity of Kazakh koumiss and assessment of its anti-Candida albicans activity. World J. Microbiol. Biotechnol. 2024b, 40, 99. [CrossRef]
- Taheur, F.B.; Fedhila, K.; Chaieb, K.; Kouidhi, B.; Bakhrouf, A.; Abrunhosa, L. Adsorption of aflatoxin B1, zearalenone and ochratoxin A by microorganisms isolated from Kefir grains. Int. J. Food Microbiol. 2017, 251, 1–7. [CrossRef]
- Adebiyi, J.A.; Kayitesi, E.; Adebo, O.A.; Changwa, R.; Njobeh, P.B. Food fermentation and mycotoxin detoxification: An African perspective. Food Control 2019, 106, 106731. [CrossRef]
- Afshar, P.; Shokrzadeh, M.; Raeis, S.N.; Saraei, A.G-H.; Nasiraii, L.R. Aflatoxins biodetoxification strategies based on probiotic bacteria. Toxicon 2020, 178, 50–58. [CrossRef]
- Lafuente, C.; Calpe, J.; Musto, L.; Nazareth, T.d.M.; Dopazo, V.; Meca, G.; Luz, C. Preparation of Sourdoughs Fermented with Isolated Lactic Acid Bacteria and Characterization of Their Antifungal Properties. Foods 2023, 12, 686. [CrossRef]
- Lafuente, C.; de Melo Nazareth, T.; Dopazo, V.; Meca, G.; Luz, C. Enhancing Bread Quality and Extending Shelf Life Using Dried Sourdough. LWT, 2024, 116379. [CrossRef]
- Teixeira, L.B.; Campos, J.Z.; Kothe, C.I.; Welke, J.E.; Rodrigues, E.; Frazzon, J.; Thys, R.C.S. Type III sourdough: Evaluation of biopreservative potential in bakery products with enhanced antifungal activity. Food Res. Int. 2024, 189, 114482. [CrossRef]
- Calasso, M.; Marzano, M.; Caponio, G.R.; Celano, G.; Fosso, B.; Calabrese, F.M.; De Palma. D.; Vacca. M.; Notario, E.; Pesole, G.; De Angelis, M.; De Leo, F. Shelf-life extension of leavened bakery products by using bio-protective cultures and type-III sourdough. LWT, 2023, 177, 114587. [CrossRef]
- Siepmann, F.B.; Ripari, V.; Waszczynskyj, N.; Spier, M.R. Overview of sourdough technology: From production to marketing. Food Bioproc. Tech. 2018, 11, 242–270. [CrossRef]
- Fekri, A.; Abedinzadeh, S.; Torbati, M.; Azadmard-Damirchi, S.; Savage, G.P. Considering sourdough from a biochemical, organoleptic, and nutritional perspective. J. Food Compos. Anal. 2024, 125, 105853. [CrossRef]
- Mūrniece, R.; Kļava, D. Impact of Long-Fermented Sourdough on the Technological and Prebiotical Properties of Rye Bread. Proc.Latv. Acad. Sci. 2022, 76(1), 1–8. [CrossRef]
- Syrokou, M.K.; Tziompra, S.; Psychogiou, E.-E.; Mpisti, S.-D.; Paramithiotis, S.; Bosnea, L.; Mataragas, M.; Skandamis, P.N.; Drosinos, E.H. Technological and Safety Attributes of Lactic Acid Bacteria and Yeasts Isolated from Spontaneously Fermented Greek Wheat Sourdoughs. Microorganisms 2021, 9, 671. [CrossRef]
- Hernández-Figueroa, R.H.; Mani-López, E.; López-Malo, A. Antifungal activity of wheat-flour sourdough (Type II) from two different Lactobacillus in vitro and bread. Appl. Food Res. 2023, 3(2), 100319. [CrossRef]
- Garcia, M.V.; Stefanello, R.F.; Pia, A.K.; Lemos, J.G.; Nabeshima, E.H.; Bartkiene, E.; Rocha, J.M.; Copetti, M.V.; Sant'Ana, A.S. Influence of Limosilactobacillus fermentum IAL 4541 and Wickerhamomyces anomalus IAL 4533 on the growth of spoilage fungi in bakery products. Int. J. Food Microbiol. 2024, 413, 110590. [CrossRef]
- Sani A.R.; Hamza, T. M.; Legbo I.M.; Jubril F.I.; Ibrahim A.; Mohammed, A.; Buru, A.S. Isolation and Identification of Fungi spp Associated with Bread Spoilage in Lapai, Niger State. Nigerian Health J. 2025, 25(1), 365–370. [CrossRef]
- Wu, X.; Liu, Y.; Guo, Z.; Ji, N.; Sun, Q.; Liu, T.; Li, Y. Antifungal activities of Pediococcus pentosaceus LWQ1 and Lactiplantibacillus plantarum LWQ17 isolated from sourdough and their efficacies on preventing spoilage of Chinese steamed bread. Food Control, 2025, 168, 110940. [CrossRef]
- Garofalo, C.; Zannini, E.; Aquilanti, L.; Silvestri, G.; Fierro, O.; Picariello, G.; Clementi, F. Selection of Sourdough Lactobacilli with Antifungal Activity for Use as Biopreservatives in Bakery Products. J. Agric. Food Chem. 2012, 60(31), 7719–7728. [CrossRef]
- Hernández-Figueroa, R.H.; Mani-López, E.; Ramírez-Corona, N.; López-Malo, A. Optimizing Lactic Acid Bacteria Proportions in Sourdough to Enhance Antifungal Activity and Quality of Partially and Fully Baked Bread. Foods 2024b, 13, 2318. [CrossRef]
- Hernández-Figueroa, R.H.; Mani-López, E.; Ramírez-Corona, N.; López-Malo, A. Optimizing Lactic Acid Bacteria Proportions in Sourdough to Enhance Antifungal Activity and Quality of Partially and Fully Baked Bread. Foods 2024c-f, 13, 2318. [CrossRef]
- Mou, T.; Xu, R.; Li, Q.; Li, J.; Liu, S.; Ao, X.; Chen, S.; Liu, A. Screening of Antifungal Lactic Acid Bacteria and Their Impact on the Quality and Shelf Life of Rye Bran Sourdough Bread. Foods 2025, 14, 1253. [CrossRef]
- Arsoy, E.S.; Gül, L.B.; Çon, A.H. Characterization and selection of potential antifungal lactic acid bacteria isolated from Turkish spontaneous sourdough. Curr. Microbiol. 2022, 79(5), 148. [CrossRef]
- Jin, J.; Nguyen, T.T.H.; Humayun, S.; Park, S.; Oh, H.; Lim, S.; Mk, I.K.; Li, Y.; Pal, K.; Kim, D. Characteristics of sourdough bread fermented with Pediococcus pentosaceus and Saccharomyces cerevisiae and its bio-preservative effect against Aspergillus flavus. Food Chem. 2021, 345, 128787. [CrossRef]
- Ma, M.; Li, A.; Feng, J.; Wang, Z.; Jia, Y.; Ma, X.; Ning, Y. Antifungal mechanism of Lactiplantibacillus plantarum P10 against Aspergillus niger and its in-situ biopreservative application in Chinese steamed bread. Food Chem. 2024, 449, 139181. [CrossRef]
- Stefanello, R.F.; Vilela, L.F.; Margalho, L.P.; Nabeshima, E.H.; Matiolli, C.C.; da Silva, D.T.; Schwan, R.F.; Emanuelli T.; Noronha, M.F.; Cabral, L.; Sant’Ana, A.S.; Copetti, M.V. Dynamics of microbial ecology and their bio-preservative compounds formed during the panettones elaboration using sourdough-isolated strains as starter cultures. Food Biosci. 2024, 60, 104279. [CrossRef]
- Mota-Gutierrez, J.; Franciosa, I.; Ruggirello, M.; Dolci, P. Technological, functional and safety properties of lactobacilli isolates from soft wheat sourdough and their potential use as antimould cultures. World J. Microbiol. Biotechnol. 2021, 37(9), 146. [CrossRef]
- EL Boujamaai, M.; Mannani, N.; Aloui, A.; Errachidi, F.; Salah-Abbès, J.B.; Riba, A.; Abbès, S.; Rocha, J.M.; Bartkiene, E.; Brabet, C.; Zinedine, A. Biodiversity and biotechnological properties of lactic acid bacteria isolated from traditional Moroccan sourdoughs. World J. Microbiol. Biotechnol. 2023, 39, 331 . [CrossRef]
- Stiles, J.; Penkar, S.; Plocková, M.; Chumchalová, J.; Bullerman, L.B. Antifungal activity of sodium acetate and Lactobacillus rhamnosus. J. Food Prot. 2002, 65(7), 1188e1191. [CrossRef]
- Le Lay, C.; Mounier, J.; Vasseur, V.; Weill, A.; Le Blay, G.; Barbier, G.; Coton E. In vitro and in situ screening of lactic acid bacteria and propionibacteria antifungal activities against bakery product spoilage molds. Food Control 2016, 60, 247–255. [CrossRef]
- Fraberger, V.; Ammer, C.; Domig, K.J. Functional Properties and Sustainability Improvement of Sourdough Bread by Lactic Acid Bacteria. Microorganisms 2020, 8, 1895. [CrossRef]
- Kennepohl, D.; Farmer, S.; Reusch, W.; Neils, T. Acidity of carboxylic acids. In Organic Chemistry. LibreTextsTM Chemistry. 1118–1121. Available online: https://chem.libretexts.org/Bookshelves/Organic_Chemistry/Supplemental_Modules_(Organic_Chemistry)/Carboxylic_Acids/Properties_of_Carboxylic_Acids/Physical_Properties_of_Carboxylic_Acids/Acidity_of_Carboxylic_Acids (accessed on 16 May 2025).
- Williams, R. pKa Values. Available online: https://chem.libretexts.org/@go/page/169800 (accessed on 16 May 2025).
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.J.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G.; et al. Lactic Acid Bacteria Isolation from Spontaneous Sourdough and Their Characterization Including Antimicrobial and Antifungal Properties Evaluation. Microorganisms 2020, 8, 64. [CrossRef]
- Debonne, E.; Maene, P.; Vermeulen, A.; Van Bockstaele, F.; Depredomme, L.; Vermeir, P.; Eeckhout, M.; Devlieghere, F. Validation of in-vitro antifungal activity of the fermentation quotient on bread spoilage moulds through growth/no-growth modelling and bread baking trials. LWT, 2020b, 117, 108636. [CrossRef]
- Debonne, E.; Vermeulen, A.; Bouboutiefski, N.; Ruyssen, T.; Van Bockstaele, F.; Eeckhout, M.; Devlieghere, F. Modelling and validation of the antifungal activity of DL-3-phenyllactic acid and acetic acid on bread spoilage moulds. Food Microbiol. 2020c, 88, 103407. [CrossRef]
- Ström, K.; Sjögren, J.; Broberg, A.; Schnürer, J. Lactobacillus plantarum MiLAB 393 Produces the Antifungal Cyclic Dipeptides Cyclo(L-Phe-L-Pro) and Cyclo(L-Phe-trans-4-OH-L-Pro) and 3-Phenyllactic Acid. Appl. Environ. Microbiol. 2002, 68(9), 4322–4327. [CrossRef]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and Characterization of Novel Antifungal Compounds from the Sourdough Lactobacillus plantarum Strain 21B. Appl. Environ. Microbiol. 2000, 66(9), 4084–4090. [CrossRef]
- Rizzello, C.G.; Cassone, A.; Coda, R.; Gobbetti, M. Antifungal activity of sourdough fermented wheat germ used as an ingredient for bread making. Food Chem. 2011, 127:952–959. [CrossRef]
- Dagnas, S.; Gauvry, E.; Onno, B.; Membré, J.M. Quantifying effect of lactic, acetic, and propionic acids on growth of molds isolated from spoilesd bakery products. J. Food Prot. 2015, 78(9), 1689–1698. [CrossRef]
- Zhang, C.; Brandt, M.J.; Schwab, C.; Gänzle, M.G. Propionic acid production by cofermentation of Lactobacillus buchneri and Lactobacillus diolivorans in sourdough. Food Microbiol. 2010, 27(3): 390–395. [CrossRef]
- Axel, C.; Zannini, E.; Arendt, E.K.; Waters, D.M.; Czerny, M. Quantification of cyclic dipeptides from cultures of Lactobacillus brevis R2D by HRGC/MS using stable isotope dilution assay. Anal. Bioanal. Chem. 2014, 406, 2433–2444. [CrossRef]
- Luz, C.; D’Opazo, V.; Mañes, J.; Meca, G. Antifungal activity and shelf life extension of loaf bread produced with sourdough fermented by Lactobacillus strains. J. Food Process. Preserv. 2019, 43, e14126. [CrossRef]
- Houssni, I.E.; Khedid, K.; Zahidi, A.; Hassikou, R. The inhibitory effects of lactic acid bacteria isolated from sourdough on the mycotoxigenic fungi growth and mycotoxins from wheat bread. Biocatal. Agric. Biotechnol. 2023, 50, 102702. [CrossRef]
- Corsetti, A.; Gobbetti, M.; Rossi, J.; Damiani, P. Antimould activity of sourdough lactic acid bacteria: identification of a mixture of organic acids produced by Lactobacillus sanfrancisco CB1. Appl. Microbiol. Biotechnol. 1998, 50(2), 253–256. [CrossRef]
- Sadeghi, A.; Ebrahimi, M.; Mortazavi, S.A.; Abedfar, A. Application of the selected antifungal LAB isolate as a protective starter culture in pan whole-wheat sourdough bread. Food Control 2018, 95, 298–307. [CrossRef]
- Ryan, L.A.M.; Zannini, E.; Dal Bello, F.; Pawlowska, A.; Koehler, P.; Arendt, E.K. Lactobacillus amylovorus DSM 19280 as a novel food-grade antifungal agent for bakery products. Int. J. Food Microbiol. 2011, 146, 276–283. 10.1016/j.ijfoodmicro.2011.02.036.
- Axel, C.; Röcker, B.; Brosnan, B.; Zannini, E.; Furey, A.; Coffey, A.; Arendt, E.K. Application of Lactobacillus amylovorus DSM19280 in gluten-free sourdough bread to improve the microbial shelf life. Food Microbiol. 2015, 47, 36–44. [CrossRef]
- Schmidt, M.; Lynch, K.M.; Zannini, E.; Arendt, E.K. Fundamental study on the improvement of the antifungal activity of Lactobacillus reuteri R29 through increased production of phenyllactic acid and reuterin. Food Control, 2018, 88, 139–148. [CrossRef]
- Petkova, M.; Stefanova, P.; Gotcheva, V.; Angelov, A. Isolation and Characterization of Lactic Acid Bacteria and Yeasts from Typical Bulgarian Sourdoughs. Microorganisms 2021, 9, 1346. [CrossRef]
- Ai, Y.; Kang, N.; Montalbán-López, M.; Wu, X.; Li, X.; Mu, D. Enhanced stability, quality and flavor of bread through sourdough fermentation with nisin-secreting Lactococcus lactis NZ9700. Food Biosci. 2024, 62, 105484. [CrossRef]
- Thanjavur, N.; Sangubotla, R.; Lakshmi, B.A.; Rayi, R.; Mekala, C.D.; Reddy, A.S.; Viswanath, B. Evaluating the antimicrobial and apoptogenic properties of bacteriocin (nisin) produced by Lactococcus lactis. Process Biochem. 2022, 122, 76–86. [CrossRef]
- Abd-Elhamed, E.Y.; El-Bassiony, T.A.E.R.; Elsherif, W.M.; Shaker, E.M. Enhancing Ras cheese safety: antifungal effects of nisin and its nanoparticles against Aspergillus flavus. BMC Vet. Res. 2024, 20(1), 493. [CrossRef]
- Yilmaz, B.; Bangar, S.P.; Echegaray, N.; Suri, S.; Tomasevic, I.; Manuel Lorenzo, J.; Melekoglu, E.; Rocha, J.M.; Ozogul, F. The Impacts of Lactiplantibacillus plantarum on the Functional Properties of Fermented Foods: A Review of Current Knowledge. Microorganisms 2022, 10, 826. [CrossRef]
- Wen, L.S.; Philip, K.; Ajam, N. Purification, characterization and mode of action of plantaricin K25 produced by Lactobacillus plantarum. Food Control 2016, 60, 430–439. [CrossRef]
- Zhao, S.; Han, J.; Bie, X.; Lu, Z.; Zhang, C.; Lv, F. Purification and characterization of plantaricin JLA-9: a novel bacteriocin against Bacillus spp. produced by Lactobacillus plantarum JLA-9 from Suan-Tsai, a traditional Chinese fermented cabbage. J. Agric.Food Chem. 2016, 64(13), 2754–2764. [CrossRef]
- Digaitiene, A.; Hansen, Å.S.; Juodeikiene, G.; Eidukonyte, D.; Josephsen, J. Lactic acid bacteria isolated from rye sourdoughs produce bacteriocin-like inhibitory substances active against Bacillus subtilis and fungi. J. Appl. Microbiol. 2012, 112(4), 732–742. [CrossRef]
- Chen, X. Y.; Levy, C.; Gänzle, M.G. Structure-function relationships of bacterial and enzymatically produced reuterans and dextran in sourdough bread baking application. Int. J. Food Microbiol. 2016, 239, 95–102. [CrossRef]
- Martin, H.; Maris, P. Synergism between hydrogen peroxide and seventeen acids against five Agri-food-borne fungi and one yeast strain. J. Appl. Microbiol. 2012, 113, 1451–1460. [CrossRef]
- Liu, A.; Xu, R.; Zhang, S.; Wang, Y.; Hu, B.; Ao, X.; Li, Q.; Li, J.; Hu, K.; Yang, Y.; Liu, S. Antifungal mechanisms and application of lactic acid bacteria in bakery products: a review. Front. Microbiol. 2022, 13, 924398. [CrossRef]
- Hernández-Figueroa, R.H.; Morales-Camacho, J.I.; Mani-López, E.; López-Malo, A. Assessment of antifungal activity of aqueous extracts and protein fractions from sourdough fermented by Lactiplantibacillus plantarum. Future Foods 2024c, 9, 100314. [CrossRef]
- Hernández-Figueroa, R.H.; Mani-López, E.; López-Malo, A. Antifungal Capacity of Poolish-Type Sourdough Supplemented with Lactiplantibacillus plantarum and Its Aqueous Extracts In Vitro and Bread. Antibiotics 2022, 11, 1813. [CrossRef]
- Muhialdin, B.J.; Hassan, Z.; Saari, N. In vitro antifungal activity of lactic acid bacteria low molecular peptides against spoilage fungi of bakery products. Ann. Microbiol. 2018, 68(9), 557–567. [CrossRef]
- Ebrahimi, M.; Sadeghi, A.; Mortazavi, S.A. The use of cyclic dipeptide producing LAB with potent anti-aflatoxigenic capability to improve techno-functional properties of clean-label bread. Ann. Microbiol. 2020, 70, 24. [CrossRef]
- Dal Bello, F.; Clarke, C.I.; Ryan, L.A.M.; Ulmer, H.; Schober, T.J.; Ström, K., Sjögren, J.; van Sinderen, D.; Schnürer, J.; Arendt, E.K. Improvement of the quality and shelf life of wheat bread by fermentation with the antifungal strain Lactobacillus plantarum FST 1.7. J. Cereal Sci. 2007. 45(3), 309–318. [CrossRef]
- Ryan, L.A.M.; Dal Bello, F.; Arendt, E.K.; Koehler, P. Detection and Quantitation of 2,5-Diketopiperazines in Wheat Sourdough and Bread. J. Agric. Food Chem. 2009, 57(20), 9563–9568. [CrossRef]
- Nionelli, L.; Wang, Y.; Pontonio, E.; Immonen, M.; Rizzello, C.G.; Maina, H. N.; Katina, K.; Coda, R. Antifungal effect of bioprocessed surplus bread as ingredient for bread-making: Identification of active compounds and impact on shelf-life. Food Control 2020, 118, 107437. [CrossRef]
- Coda, R.; Cassone, A.; Rizzello, C.G.; Nionelli, L.; Cardinali, G.; Gobbetti, M. Antifungal Activity of Wickerhamomyces Anomalus and Lactobacillus Plantarum during Sourdough Fermentation: Identification of Novel Compounds and Long-Term Effect during Storage of Wheat Bread. Appl. Environ. Microbiol. 2011, 77, 3484–3492. [CrossRef]
- Thiele, C.; Gänzle, M.G.; Vogel, R.F. Contribution of sourdough lactobacilli, yeast, and cereal enzymes to the generation of amino acids in dough relevant for bread flavour. Cereal Chem. 2002, 79(1), 45–51. [CrossRef]
- Biesiekierski, J.R. What is gluten? J. Gastroenterol. Hepatol. 2017, 32, 78–81. [CrossRef]
- Rahimi, D.; Auld, K.; Sadeghi, A.; Kashaninejad, M.; Ebrahimi, M.; Zhang, J.; Gänzle, M.G. Optimising bread preservation: use of sourdough in combination with other clean label approaches for enhanced mould-free shelf life of bread. Eur. Food Res. Technol. 2025, 1–10. [CrossRef]
- Hassan, Y.I.; Zhou, T.; Bullerman, L.B. Sourdough lactic acid bacteria as antifungal and mycotoxin-controlling agents. Food Sci. Technol. Int. 2015, 22(1), 79–90. [CrossRef]
- Özdemir, N.; Gül, H. Effects of fermentation time, baking, and storage on ochratoxin A levels in sourdough flat bread. Food Sci. Nutr. 2024. 12(10), 7370–7378. [CrossRef]
- Noroozi, R.; Kobarfard, F.; Rezaei, M.; Ayatollahi, S.A.; Paimard, G.; Eslamizad, S.; Razmjoo, F.; Sadeghi, E. Occurrence and exposure assessment of aflatoxin B1 in Iranian breads and wheat-based products considering effects of traditional processing. Food Control, 2022, 138, 108985. [CrossRef]
- Hu, L.; Koehler, P.; Rychlik, M. Effect of sourdough processing and baking on the content of enniatins and beauvericin in wheat and rye bread. Eur. Food Res. Technol. 2014, 238(4), 581–587. [CrossRef]
- Cao, H.; Meng, D.; Zhang, W.; Ye, T.; Yuan, M.; Yu, J.; Wu, X.; Li, Y.; Yin, F.; Fu, C.; Xu, F. Growth inhibition of Fusarium graminearum and deoxynivalenol detoxification by lactic acid bacteria and their application in sourdough bread. Int. J. Food Sci. Technol. 2021, 56(5), 2304–2314. [CrossRef]
- Badji, T.; Durand, N.; Bendali, F.; Piro-Metayer, I.; Zinedine, A.; Salah-Abbès, J.B.; Montet, D.; Riba, A.; Brabet, C. In vitro detoxification of aflatoxin B1 and ochratoxin A by lactic acid bacteria isolated from Algerian fermented foods. Biol. Control, 2023, 179, 105181. [CrossRef]
- Zadeike, D.; Vaitkeviciene, R.; Bartkevics, V.; Bogdanova, E.; Bartkiene, E.; Lele, V.; Juodeikiene, G.; Cernauskas, D.; Valatkeviciene, Z. The expedient application of microbial fermentation after whole-wheat milling and fractionation to mitigate mycotoxins in wheat-based products. LWT, 2021, 137, 110440. [CrossRef]
- Vidal, A.; Marín, S.; Morales, H.; Ramos, A.J.; Sanchis, V. The fate of deoxynivalenol and ochratoxin A during the breadmaking process, effects of sourdough use and bran content. Food Chem. Toxicol. 2014, 68, 53–60. 10.1016/j.fct.2014.03.006.
- Banu, I.; Dragoi, L.; Aprodu, I. From wheat to sourdough bread: A laboratory scale study on the fate of deoxynivalenol content. Quality Assurance and Safety of Crops & Foods, 2014, 6(1), 53–60. [CrossRef]
- Iosca, G.; Fugaban, J.I.I.; Özmerih, S.; Wätjen, A.P.; Kaas, R.S.; Hà, Q.; Shetty, R.; Pulvirenti, A.; De Vero, L.; Bang-Berthelsen, C.H. Exploring the Inhibitory Activity of Selected Lactic Acid Bacteria against Bread Rope Spoilage Agents. Fermentation 2023, 9, 290. [CrossRef]
- Pereira, A.P.M.; Stradiotto, G.C.; Freire, L.; Alvarenga, V.O.; Crucello, A.; Morassi, L.L.; Silva, F.P.; Sant’Ana, A.S. Occurrence and enumeration of rope-producing spore forming bacteria in flour and their spoilage potential in different bread formulations. Lwt 2020, 133, 110108. [CrossRef]
- Çakır, E.; Arıcı, M.; Durak, M.Z.; Karasu, S. The molecular and technological characterization of lactic acid bacteria in einkorn sourdough: Effect on bread quality. J. Food Meas. Charact. 2020, 14, 1646–1655. [CrossRef]
- Maidana, S.D.; Ficoseco, C.A.; Bassi, D.; Cocconcelli, P.S.; Puglisi, E.; Savoy, G.; Vignolo, G.; Fontana, C. Biodiversity and technological-functional potential of lactic acid bacteria isolated from spontaneously fermented chia sourdough. Int. J. Food Microbiol. 2020, 316, 108425. [CrossRef]
- Li, Z.; Siepmann, F.B.; Tovar, L.E.R.; Chen, X.; Gänzle, M.G. Effect of copy number of the spoVA2mob operon, sourdough and reutericyclin on ropy bread spoilage caused by Bacillus spp. Food Microbiol. 2020, 91, 103507. [CrossRef]
- Pahlavani, M.; Sadeghi, A.; Ebrahimi, M.; Kashaninejad, M.; Moayedi, A. Application of the selected yeast isolate in type IV sourdough to produce enriched clean-label wheat bread supplemented with fermented sprouted barley. J. Agric. Food Res. 2024, 15, 101010. [CrossRef]
- Shahryari, S.; Sadeghi, A.; Ebrahimi, M.; Mahoonak, A.S.; Moayedi, A. Evaluation of probiotic and antifungal properties of the yeast isolated from buckwheat sourdough. Iranian Food Science & Technology Research Journal/Majallah-i Pizhūhishhā-yi ̒Ulūm va Sanāyi̒-i Ghaz̠āyī-i Īrān, 2022, 18(5), 575. [CrossRef]
- Milani, J.; Heidari, S. Stability of ochratoxin A during bread making process. J. Food Saf. 2017, 37(1), e12283. [CrossRef]
- Hermann, M.; Petermeier, H.; Vogel, R.F. Development of novel sourdoughs with in situ formed exopolysaccharides from acetic acid bacteria. European Food Research and Technology, 2015, 241(2), 185–197. [CrossRef]
- Mohd Roby, B. H.; Muhialdin, B.J.; Abadl, M.M.T.; Mat Nor, N.A.; Marzlan, A.A.; Lim, S.A.H., N.A. Mustapha; Meor Hussin, A.S. Physical properties, storage stability, and consumer acceptability for sourdough bread produced using encapsulated kombucha sourdough starter culture. J. Food Sci. 2020, 85(8), 2286–2295. [CrossRef]
- Kilmanoglu, H.; Akbas, M.; Cinar, A.Y.; Durak, M.Z. Kombucha as alternative microbial consortium for sourdough fermentation: Bread characterization and investigation of shelf life. Int. J. Gastron. Food Sci. 2024, 35, 100903. [CrossRef]
- Calvert, M.D.; Madden, A.A.; Nichols, L.M.; Haddad, N.M.; Lahne, J.; Dunn, R.R.; McKenney, E.A. A review of sourdough starters: Ecology, practices, and sensory quality with applications for baking and recommendations for future research. PeerJ 2021, 9, e11389. [CrossRef]
- Pontonio, E., Verni, M., Montemurro, M., Rizzello, C.G. Sourdough: A Tool for Non-conventional Fermentations and to Recover Side Streams. In Handbook on Sourdough Biotechnology; Gobbetti, M., Gänzle, M., Eds.; Springer; Cham, Switzerland, 2023, 257–302. [CrossRef]
- Banovic, M.; Arvola, A.; Pennanen, K.; Duta, D.E.; Brückner-Gühmann, M., Lähteenmäki, L., Grunert, K.G. Foods with increased protein content: A qualitative study on European consumer preferences and perceptions. Appetite 2018, 125, 233–243. [CrossRef]
- Oleinikova, Y.; Maksimovich, S.; Khadzhibayeva, I.; Khamedova, E.; Zhaksylyk, A.; Alybayeva, A. Meat quality, safety, dietetics, environmental impact, and alternatives now and ten years ago: a critical review and perspective. Food Prod. Process. Nutr. 2025, 7(1), 18. [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Polo, A.; Rizzello, C.G. The sourdough fermentation is the powerful process to exploit the potential of legumes, pseudo-cereals and milling by-products in baking industry. Crit. Rev. Food Sci. Nutr. 2020, 60(13), 2158–2173. [CrossRef]
- Ameur, H.; Arora, K.; Polo, A.; Gobbetti, M. The sourdough microbiota and its sensory and nutritional performances. In Good Microbes in Medicine, Food Production, Biotechnology, Bioremediation, and Agriculture; de Bruijn, F.J., Smidt, H., Cocolin, L.S., Sauer, M., Dowling, D., Thomashow, L., Eds.; John Wiley & Sons: Hoboken, New Jersey, USA, 2022, 169–184. 10.1002/9781119762621.ch14.
- Aspri, M., Mustač, N. Č., Tsaltas, D. Non-cereal and Legume Based Sourdough Metabolites. In Sourdough Innovations; Garcia-Vaquero, M., Rocha, J.M.F., Eds.; CRC Press, Boca Raton, USA, 2023, 63–86. 10.1201/9781003141143-4.
- Vurro, F.; Santamaria, M.; Summo, C.; Pasqualone, A.; Rosell, C.M. Exploring the functional potential of pea-based sourdough in traditional durum wheat focaccia: Role in enhancing bioactive compounds, in vitro antioxidant activity, in vitro digestibility and aroma. J. Funct. Foods, 2024, 123, 106607. [CrossRef]
- Patrascu, L.; Vasilean, I.; Turtoi, M.; Garnai, M.; Aprodu, I. Pulse germination as tool for modulating their functionality in wheat flour sourdoughs. Qual. Assur. Saf. Crops Foods, 2019, 11(3), 269–282. [CrossRef]
- Cacak-Pietrzak, G.; Sujka, K.; Księżak, J.; Bojarszczuk, J.; Dziki, D. Sourdough wheat bread enriched with grass pea and lupine seed flour: Physicochemical and sensory properties. Appl. Sci. 2023, 13(15), 8664. [CrossRef]
- Drakula, S.; Novotni, D.; Mustač, N.Č.; Voučko, B.; Krpan, M.; Hruškar, M.; Ćurić, D. Alteration of phenolics and antioxidant capacity of gluten-free bread by yellow pea flour addition and sourdough fermentation. Food Biosci. 2021, 44, 101424. [CrossRef]
- González-Montemayor, A.M.; Solanilla-Duque, J.F.; Flores-Gallegos, A.C.; López-Badillo, C.M.; Ascacio-Valdés, J.A.; Rodríguez-Herrera, R. Green Bean, Pea and Mesquite Whole Pod Flours Nutritional and Functional Properties and Their Effect on Sourdough Bread. Foods 2021, 10, 2227. [CrossRef]
- Verni, M., Wang, Y., Clement, H., Koirala, P., Rizzello, C. G., Coda, R. Antifungal peptides from faba bean flour fermented by Levilactobacillus brevis AM7 improve the shelf-life of composite faba-wheat bread. Int. J. Food Microbiol. 2023, 407, 110403. [CrossRef]
- Hajinia, F.; Sadeghi, A.; Sadeghi Mahoonak, A. The use of antifungal oat-sourdough lactic acid bacteria to improve safety and technological functionalities of the supplemented wheat bread. J. Food Safety, 2021, 41(1), e12873. [CrossRef]
- Çakır, E.; Arıcı, M.; Durak, M.Z. Biodiversity and techno-functional properties of lactic acid bacteria in fermented hull-less barley sourdough. J. Biosci. Bioeng. 2020, 130(5), 450-456. [CrossRef]
- Korcari, D.; Secchiero, R.; Laureati, M.; Marti, A.; Cardone, G.; Rabitti, N. S.; Ricci, G.; Fortina, M. G. Technological properties, shelf life and consumer preference of spelt-based sourdough bread using novel, selected starter cultures. LWT 2021, 151, 112097. [CrossRef]
- Rizi, A.Z.; Sadeghi, A., Jafari, S. M., Feizi, H., & Purabdolah, H. Controlled fermented sprouted mung bean containing ginger extract as a novel bakery bio-preservative for clean-label enriched wheat bread. J. Agric. Food Res. 2024, 16, 101218. [CrossRef]
- Rizi A.Z.; Sadeghi A, Feizi H, Jafari S M, Purabdolah H. Evaluation of textural, sensorial and shelf-life characteristics of bread produced with mung bean sourdough and saffron petal extract. FSCT 2024, 21(148), 141–153. URL: http://fsct.modares.ac.ir/article-7-72055-en.html.
- Aryashad, M.; Sadeghi, A.; Nouri, M.; Ebrahimi, M.; Kashaninejad, M.; Aalami, M. Use of fermented sprouted mung bean (Vigna radiata) containing protective starter culture LAB to produce clean-label fortified wheat bread. Int. J. Food Sci. Technol. 2023, 58(6), 3310–3320. [CrossRef]
- Rasoulifar, M.; Sadeghi, A.; Hajinia, F.; Ebrahimi, M.; Ghorbani, M. Effect of the Controlled Fermented Sprouted Lentil Containing Fennel Extract on the Characteristics of Wheat Bread. Iran. Food Sci. Technol. Res. J., 2024, 20(4), 433–446. [CrossRef]
- Dopazo, V.; Musto, L.; de Melo Nazareth, T.; Lafuente, C.; Meca, G.; Luz, C. Revalorization of rice bran as a potential ingredient for reducing fungal contamination in bread by lactic acid bacterial fermentation. Food Biosci. 2024, 58, 103703. [CrossRef]
- Wang, Y.; Xie, C.; Pulkkinen, M.; Edelmann, M.; Chamlagain, B.; Coda, R.; Sandell, M.; Piironen, V.; Maina, N.H.; Katina, K. In situ production of vitamin B12 and dextran in soya flour and rice bran: A tool to improve flavour and texture of B12-fortified bread. LWT, 2022, 161, 113407. [CrossRef]
- Păcularu-Burada, B.; Georgescu, L.A.; Vasile, M.A.; Rocha, J.M.; Bahrim, G.-E. Selection of Wild Lactic Acid Bacteria Strains as Promoters of Postbiotics in Gluten-Free Sourdoughs. Microorganisms 2020, 8, 643. [CrossRef]
- Schettino, R.; Pontonio, E.; Gobbetti, M.; Rizzello, C.G. Extension of the Shelf-Life of Fresh Pasta Using Chickpea Flour Fermented with Selected Lactic Acid Bacteria. Microorganisms 2020, 8, 1322. [CrossRef]
- Hoehnel, A.; Bez, J.; Sahin, A.W.; Coffey, A.; Arendt, E.K.; Zannini, E. Leuconostoc citreum TR116 as a Microbial Cell Factory to Functionalise High-Protein Faba Bean Ingredients for Bakery Applications. Foods 2020, 9, 1706. [CrossRef]
- Rouhi, E.; Sadeghi, A.; Jafari, S.M.; Abdolhoseini, M.; Assadpour, E. Effect of the controlled fermented quinoa containing protective starter culture on technological characteristics of wheat bread supplemented with red lentil. J. Food Sci. Technol. 2023, 60(8), 2193–2203. [CrossRef]
- Kia, P.S.; Sadeghi, A.; Kashaninejad, M.; Zarali, M.; Khomeiri, M. Application of controlled fermented amaranth supplemented with purslane (Portulaca oleracea) powder to improve technological functionalities of wheat bread. Appl. Food Res. 2024, 4(1), 100395. [CrossRef]
- Ebrahimi, M.; Sadeghi, A.; Sarani, A.; Purabdolah, H. Enhancement of technological functionality of white wheat bread using wheat germ sourdough along with dehydrated spinach puree. J. Agric. Sci. Technol. 2021, 23(4), 839–851. URL: http://jast.modares.ac.ir/article-23-37775-en.html.
- Omedi, J.O.; Huang, J.; Huang, W.; Zheng, J.; Zeng, Y.; Zhang, B.; Zhou, L.; Zhao, F.; Li, N.; Gao, T. Suitability of pitaya fruit fermented by sourdough LAB strains for bread making: its impact on dough physicochemical, rheo-fermentation properties and antioxidant, antifungal and quality performance of bread. Heliyon, 2021, 7(11), e08290. [CrossRef]
- Zarali, M.; Sadeghi, A.; Ebrahimi, M.; Jafari, S.M.; Mahoonak, A.S. Techno-nutritional capabilities of sprouted clover seeds sourdough as a potent bio-preservative against sorbate-resistant fungus in fortified clean-label wheat bread. Food Measure 2024, 18, 5577–5589. [CrossRef]
- Mantzourani, I.; Daoutidou, M.; Plessas, S. Impact of Functional Supplement Based on Cornelian Cherry (Cornus mas L.) Juice in Sourdough Bread Making: Evaluation of Nutritional and Quality Aspects. Appl. Sci. 2025, 15, 4283. [CrossRef]
- Mantzourani, I.; Daoutidou, M.; Terpou, A.; Plessas, S. Novel Formulations of Sourdough Bread Based on Supplements Containing Chokeberry Juice Fermented by Potentially Probiotic L. paracasei SP5. Foods 2024, 13, 4031. [CrossRef]
- Plessas, S.; Mantzourani, I.; Alexopoulos, A.; Alexandri, M.; Kopsahelis, N.; Adamopoulou, V.; Bekatorou, A. Nutritional Improvements of Sourdough Breads Made with Freeze-Dried Functional Adjuncts Based on Probiotic Lactiplantibacillus plantarum subsp. plantarum and Pomegranate Juice. Antioxidants 2023, 12, 1113. [CrossRef]
- Neylon, E.; Nyhan, L.; Zannini, E.; Sahin, A.W.; Arendt, E.K. From Waste to Taste: Application of Fermented Spent Rootlet Ingredients in a Bread System. Foods 2023, 12, 1549. [CrossRef]
- Dopazo, V.; Illueca, F.; Luz, C.; Musto, L.; Moreno, A.; Calpe, J.; Meca, G. Evaluation of shelf life and technological properties of bread elaborated with lactic acid bacteria fermented whey as a bio-preservation ingredient. LWT, 2023, 174, 114427. [CrossRef]
- Luz, C.; Quiles, J. M.; Romano, R.; Blaiotta, G.; Rodríguez, L.; Meca, G. Application of whey of Mozzarella di Bufala Campana fermented by lactic acid bacteria as a bread biopreservative agent. Int. J. Food Sci. Technol. 2021, 56(9), 4585–4593. [CrossRef]
- Izzo, L.; Luz, C.; Ritieni, A.; Mañes, J.; Meca, G. Whey fermented by using Lactobacillus plantarum strains: A promising approach to increase the shelf life of pita bread. J. Dairy Sci. 2020, 103(7), 5906–5915. [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Lele, V.; Pugajeva, I.; Zavistanaviciute, P.; Zadeike, D.; Juodeikiene, G. Application of antifungal lactobacilli in combination with coatings based on apple processing by-products as a bio-preservative in wheat bread production. J. Food Sci.Technol. 2019, 56, 2989–3000. [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Lele, V.; Pugajeva, I.; Zavistanaviciute, P.; Mickiene, R.; Zadeike, D.; Juodeikiene, G. A concept of mould spoilage prevention and acrylamide reduction in wheat bread: Application of lactobacilli in combination with a cranberry coating. Food Control, 2018, 91, 284–293. [CrossRef]
- Sharaf, O.; Ibrahim, G.A.; Mahammad, A.A. Prevention of mold spoilage and extend the shelf life of bakery products using modified mixed nano fermentate of Lactobacillus sp. Egypt. J. Chem. 2023, 66(12), 207–215. [CrossRef]
- Gregirchak, N.; Stabnikova, O.; Stabnikov, V. Application of lactic acid bacteria for coating of wheat bread to protect it from microbial spoilage. Plant Foods Hum. Nutr. 2020, 75(2), 223–229. [CrossRef]
- Iosca, G.; Turetta, M.; De Vero, L.; Bang-Berthelsen, C. H.; Gullo, M.; Pulvirenti, A. Valorization of wheat bread waste and cheese whey through cultivation of lactic acid bacteria for bio-preservation of bakery products. LWT, 2023, 176, 114524. [CrossRef]
- Calabrese, F.M.; Ameur, H.; Nikoloudaki, O.; Celano, G.; Vacca, M.; Lemos Junior, W.J.F.; Manzari, C.; Vertè, F.; Di Cagno, R.; Pesole, G.; De Angelis, M.; Gobbetti, M. Metabolic framework of spontaneous and synthetic sourdough metacommunities to reveal microbial players responsible for resilience and performance. Microbiome 2022, 10, 148 . [CrossRef]
Figure 1.
VOSviewer mapping of terms repeated at least five times in articles with the term “sourdough” in the abstract, based on OpenAlex API text data. (a) the group containing the bulk of the species names occupies a central position among other groups of terms; (b) Lactiplantibacillus plantarum dominates among terms directly related to sourdoughs.
Figure 1.
VOSviewer mapping of terms repeated at least five times in articles with the term “sourdough” in the abstract, based on OpenAlex API text data. (a) the group containing the bulk of the species names occupies a central position among other groups of terms; (b) Lactiplantibacillus plantarum dominates among terms directly related to sourdoughs.
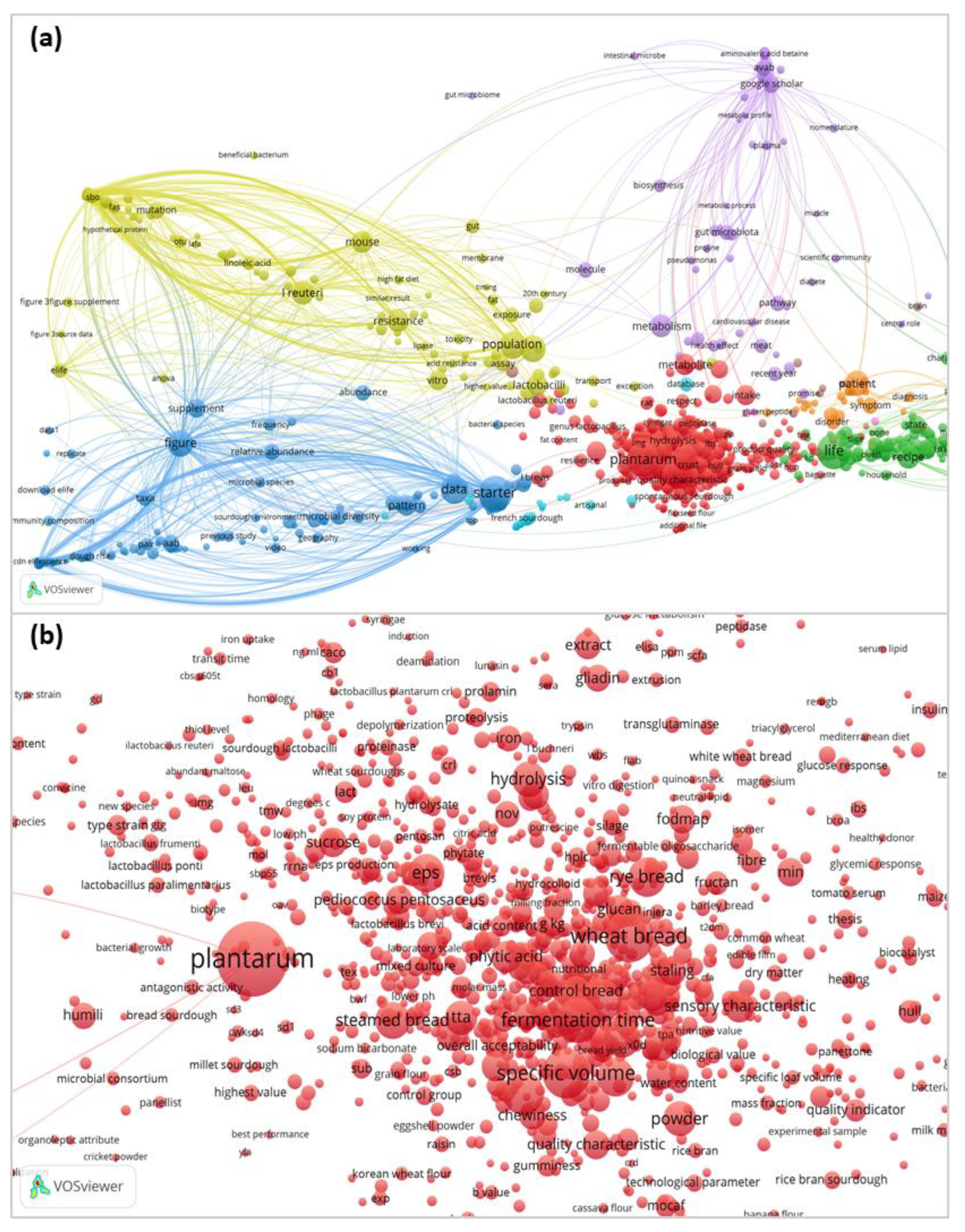
Figure 2.
Occurrence of different bacterial species in publications on bread sourdoughs. The diagram is based on VOSviewer mapping (OpenAlex API text data) of terms occurring at least five times in the articles with “sourdough” in the abstract.
Figure 2.
Occurrence of different bacterial species in publications on bread sourdoughs. The diagram is based on VOSviewer mapping (OpenAlex API text data) of terms occurring at least five times in the articles with “sourdough” in the abstract.
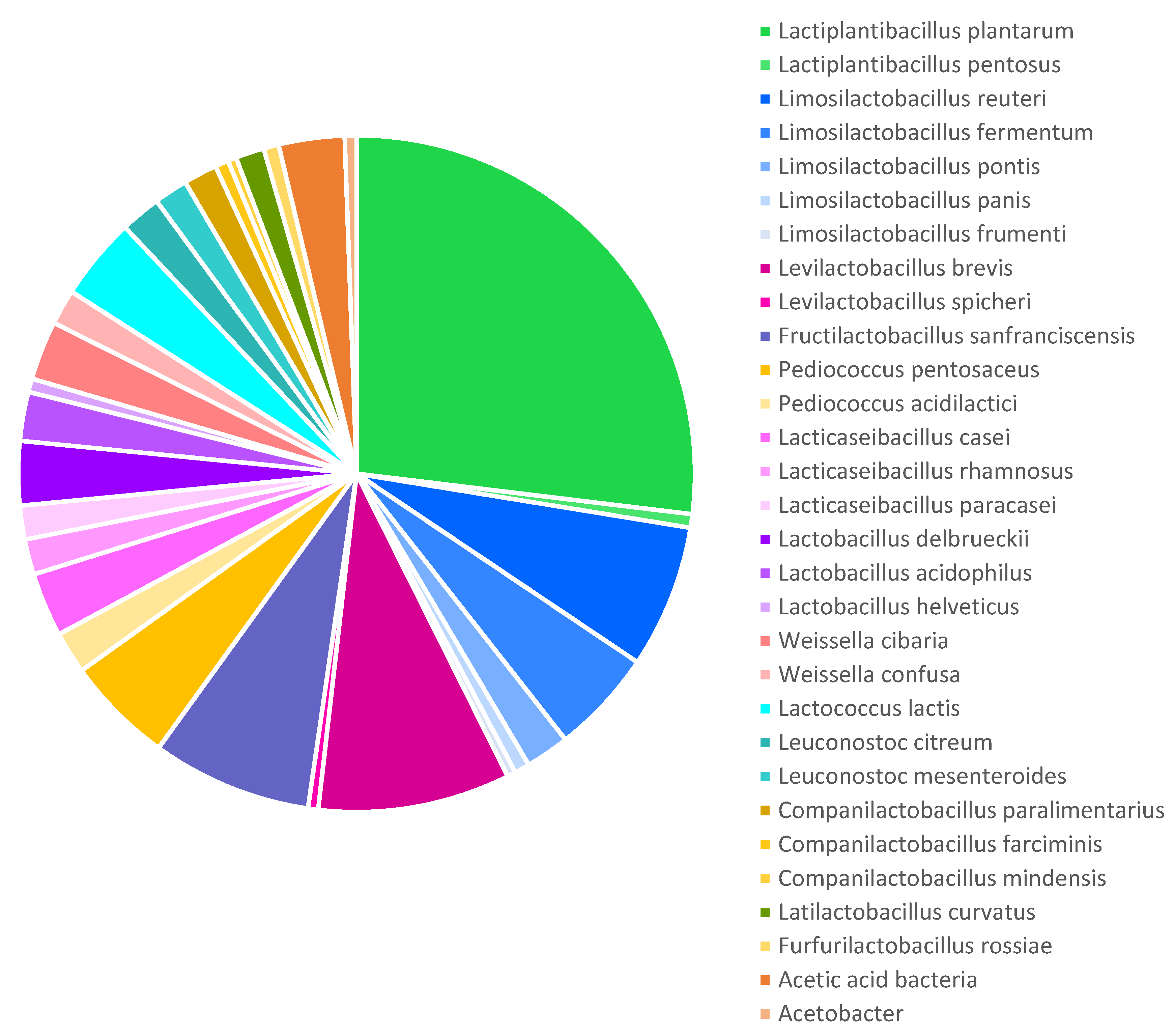
Figure 3.
Sourdough fungal microbiota based on VOSviewer mapping (OpenAlex API text data) of terms occurring at least in five articles with the term “sourdough” in the abstract.
Figure 3.
Sourdough fungal microbiota based on VOSviewer mapping (OpenAlex API text data) of terms occurring at least in five articles with the term “sourdough” in the abstract.
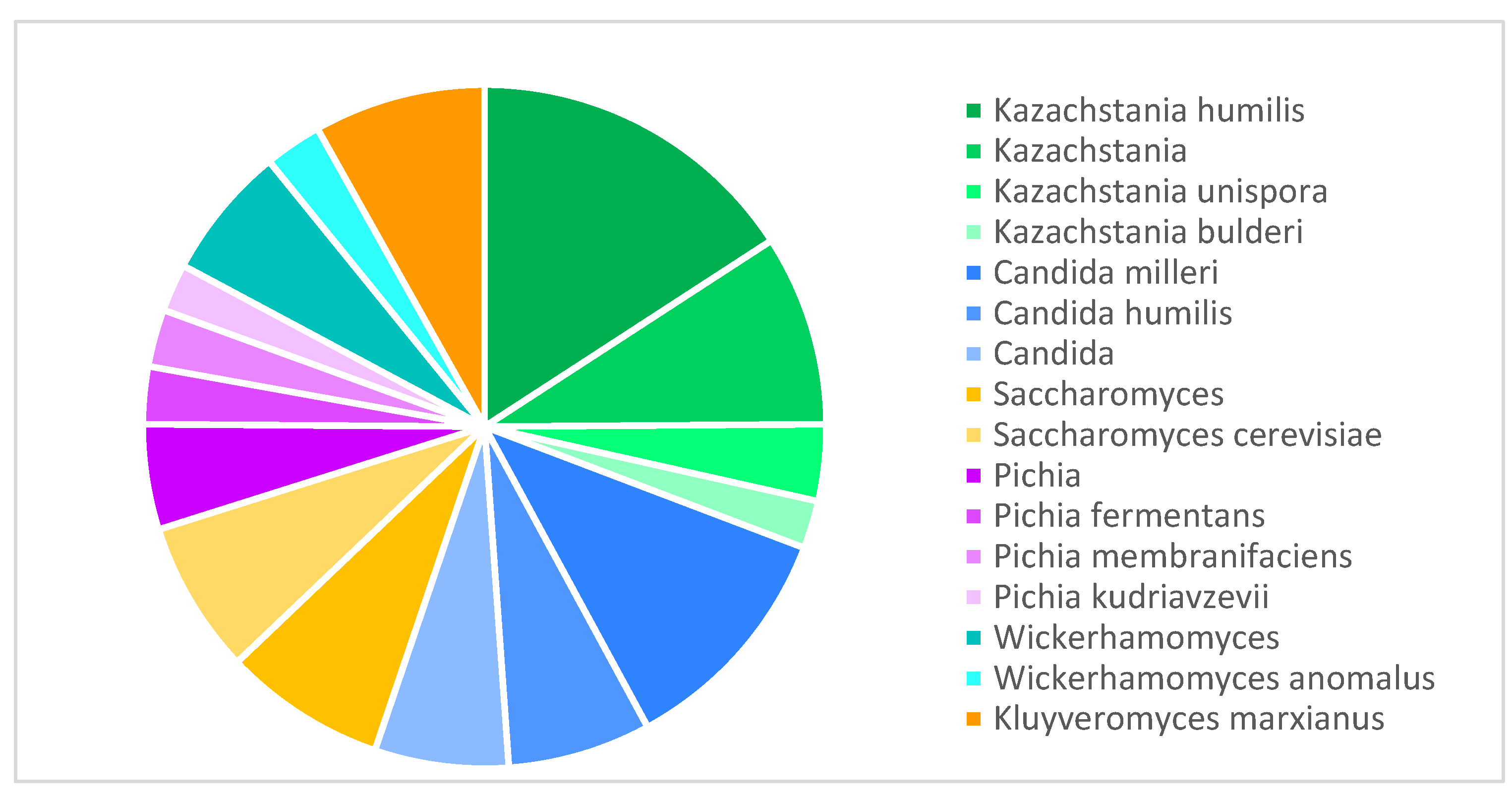

Table 1.
Antifungal organic acids produced by lactic acid bacteria for bread preservation.
| Name | Systematic name | Producing LAB species | Affected molds | References |
|---|---|---|---|---|
| Saturated aliphatic fatty acids | ||||
| Formic* | formic |
F. sanfranciscensis; L. plantarum and F. rossiae |
A. niger, F. graminearum, P. expansum, and M. sitophila; P. roqueforti |
[103]; [97] |
| Acetic* | acetic | F. sanfranciscensis | A. niger, P. paneum | [25,93,94] |
| Propionic* | propanoic |
F. sanfranciscensis; L. buchneri and L. diolivorans |
A. niger, F. graminearum, P. expansum, and M. sitophila; A. clavatus, Cladosporium spp., Mortierella spp. and P. roquefortii |
[103]; [99] |
| Butyric | butanoic | F. sanfranciscensis | A. niger, F. graminearum, P. expansum, and M. sitophila | [103] |
| n-Valeric | pentanoic | F. sanfranciscensis | A. niger, F. graminearum, P. expansum, and M. sitophila | [103] |
| Caproic* | hexanoic | F. sanfranciscensis | A. niger, F. graminearum, P. expansum, and M. sitophila | [103] |
| Capric | decanoic | L. reuteri | A. niger | [104] |
| Hydroxy acids and phenyl substituted acids | ||||
| Lactic* | 2-hydroxypropanoic |
L. acidophilus, L. casei; W. cibaria, L. plantarum subsp. plantarum, L. pseudomesenteroides, F. sanfranciscensis, L. brevis, and L. pentosus |
P. crysogenum, P. corylophilum; A. flavus, A. niger, P. expansum |
[73]; [81] |
| Hydro-cinnamic | 3-phenylpropanoic | L. amylovorus | A. fumigatus | [105] |
| Phenyl-lactic* | 2-hydroxy-3-phenylpropanoic |
L. plantarum; L. bulgaricus; L. amylovorus; L. reuteri and, L. brevis |
E. repens, E. rubrum, P. corylophilum, P. roqueforti, P. expansum, E. fibuliger, A. niger, A. flavus, M. sitophila, and F. graminearum; A. niger and P. polonicum; F. moniliformis, F. graminearum, F. verticillioides and P. expansum; environmental moulds; Fusarium culmorum and environmental moulds |
[96]; [76]; [101]; [106]; [47]; [107] |
| Phloretic | 3-(4-hydroxyphenyl)propanoic | L. amylovorus | environmental moulds | [47,106] |
| Hydroxy-phenyl-lactic | 2-hydroxy-3-(4-hydroxyphenyl)propanoic | L. amylovorus |
A. fumigatus; environmental moulds |
[105]; [47,106] |
| Hydro-caffeic | 3-(3,4-dihydroxyphenyl)propanoic |
L. amylovorus |
F. culmorum, environmental moulds | [47] |
| Hydro-ferulic | 3-(4-hydroxy-3-methoxyphenyl)propanoic | L. amylovorus | F. culmorum, environmental moulds | [47,106] |
| 2-Hydroxyiso-caproic | 2-hydroxy-4-methylpentanoic | L. amylovorus, L. reuteri, and L. brevis | F. culmorum, environmental moulds | [47] |
| 3-Hydroxy-capric | 3-hydroxydecanoic | L. reuteri | A. niger | [104] |
| 3-Hydroxy-lauric | 3-hydroxydodecanoic | L. reuteri | A. niger | [104] |
| Unsaturated phenyl-substituted acids | ||||
| ρ-Coumaric | (2E)-3-(4-Hydroxyphenyl)prop-2-enoic) | L. amylovorus | A. fumigatus | [105] |
| Caffeic | (2E)-3-(3,4-dihydroxyphenyl)prop-2-enoic | L. plantarum | P. expansum, P. roqueforti, P. camemberti, F. moniliformis, F. graminearum, F. verticillioides, A. niger, and A. parasiticus | [101] |
| Ferulic | (2E)-3-(4-hydroxy-3-methoxyphenyl)prop-2-enoic | L. brevis | F. culmorum, environmental moulds | [47] |
| 2-Methyl-cinnamic | 2e-3-2-methylphenylprop-2-enoic acid | L. amylovorus | A. fumigatus | [105] |
| Sinapic | 3-(4-hydroxy-3,5-dimethoxyphenyl)prop-2-enoic acid | L. bulgaricus | F. moniliformis, F. graminearum, F. verticillioides, and P. expansum | [101] |
| Polyphenol compound | ||||
| Chlorogenic | (1S,3R,4R,5R)-3-{[(2E)-3-(3,4-Dihydroxyphenyl)prop-2-enoyl]oxy}-1,4,5-trihydroxycyclohexane-1-carboxylic acid | L. plantarum and L. bulgaricus | P. expansum, P. roqueforti, P. camemberti, F. moniliformis, F. graminearum, F. verticillioides, A. niger, and A. parasiticus | [101] |
| Dicarboxylic acids | ||||
| Azelaic | nonanedioic | L. amylovorus and L. reuteri | F. culmorum, environmental moulds | [47] |
| Uronic acids | ||||
| D-Glucuronic acid | (2S,3S,4S,5R,6R)-3,4,5,6-Tetrahydroxyoxane-2-carboxylic acid | L. amylovorus | A. fumigatus | [105] |
| Benzoic (aromatic) acids | ||||
| Salicylic | 2-hydroxybenzoic acid | L. amylovorus | A. fumigatus | [105] |
| Gallic | 3,4,5-trihydroxybenzoic | L. plantarum | P. expansum, P. roqueforti, P. camemberti, F. moniliformis, F. graminearum, F. verticillioides, A. niger, and A. parasiticus | [101] |
| Vanillic | 4-hydroxy-3-methoxybenzoic | L. plantarum and L. bulgaricus | P. expansum, P. roqueforti, P. camemberti, F. moniliformis, F. graminearum, F. verticillioides, A. niger, and A. parasiticus | [101] |
| Syringic | 4-hydroxy-3,5-dimethoxybenzoic | L. plantarum | P. expansum, P. roqueforti, P. camemberti, F. moniliformis, F. graminearum, F. verticillioides, A. niger, and A. parasiticus | [101] |
* At least one study has shown the greatest significance of this acid in the manifestation of the antifungal effect among the mixture of acids produced.
Table 2.
Using sourdoughs with non-standard components.
| Component | LAB | Target | Mode of action | Refference |
| Einkorn sourdough | L. paraplantarum and P. acidilactici | B. subtilis ATCC6633, B. cereus ATCC11778 | Not studied | [141] |
| L. crustorum and L. brevis | Penicillium carneum, A. flavus and A. niger. | |||
| Oat-sourdough | P. pentosaceus | A. flavus | Increased total phenolic content and antioxidant activity | [163] |
| Hull-less barley sourdough | P. acidilactici and L. plantarum | P. carneum, A. flavus, and A. niger | Not studied | [164] |
| Spelt-based sourdough | W. cibaria and P. pentosaceus | F. verticillioides, A. flavus | Not studied | [165], |
| Fermented sprouted mung bean sourdough | L. brevis | A. niger | Not studied | [166,167] |
| Fermented sprouted mung bean sourdough | P. pentosaceus | A. niger | Not studied | [168] |
| Fermented sprouted lentil with fennel extract | P. acidilactici | A. niger | Not studied | [169] |
| 20 % of rice bran | L. plantarum |
Penicillium commune and A. flavus; aflatoxin |
Lactic and phenyllactic acids | [170] |
| 50% of fermented soya flour and rice bran | Propionibacterium freudenreichii and W. confusa | Environmental molds | Acetic and propionic acids | [171] |
| Fermented extracts of chickpea, quinoa, and buckwheat flour | Lactobacillus spp., Leuconostoc spp. | A. niger, A. flavus, Penicillium spp., Bacillus spp. | Organic acids |
[172] |
| Fermented chickpea flour | L. plantarum and F. rossiae | P. roqueforti, P. paneum, and P. carneum | Peptides of 12-20 amino residues | [173] |
| Fermented faba bean flour (30%) | L. brevis | P. roqueforti | Peptides of 11–22 amino acid residues, encrypted into sequences of vicilin and legumin type B; defensin-like protein (8,792 Da) and a non-specific lipid-transfer protein (11,588 Da) | [162] |
| Fermented faba bean flour (15%) | L. citreum | Potentially antifungal | Lactic, acetic, 4-hydroxybenzoic, caffeic, coumaric, ferulic, phenyllactic acids | [174] |
| Fermented quinoa and red lentil supplement | Enterococcus hirae | Environmental molds | Not studied | [175] |
| Fermented amaranth sourdough supplemented with purslane powder | L. brevis | A. niger | Not studied | [176] |
| Wheat germ sourdough along with dehydrated spinach puree | L. lactis | A. flavus | Not studied | [177] |
| 20% of fermented pitaya fruit | L. plantarum and P. pentosaceus | A. niger, Cladosporium sphaerospermum, and P. chrysogenum | Phenolic acids: gallic, caffeic, protocatechuic; increased antioxidant activity | [178] |
| Sprouted clover seeds sourdough | Lacticaseibacillus rhamnosus | Aspergillus brasiliensis | Increased antioxidant activity | [179] |
| Fermented cornelian cherry supplement | L. plantarum ATCC 14917 | Environmental molds, Bacillus spp. | Increased total phenolic content and antioxidant activity; lactic, acetic, formic, n-valeric, and caproic acids | [180] |
| Fermented chokeberry juice supplement | L. paracasei | Environmental molds, Bacillus spp. | Lactic and acetic acids; increased total phenolic content and antioxidant activity | [181] |
| Fermented pomegranate juice supplement | L. plantarum | Environmental molds, Bacillus spp. | Increased total phenolic content; lactic acid, acetic acid | [182] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



