Submitted:
10 June 2025
Posted:
11 June 2025
Read the latest preprint version here
Abstract
The kynurenine (KYN) metabolic pathway sits at the crossroads of immunity, metabolism, and neurobiology, yet its clinical translation remains fragmented. Emerging spatial-omics, wearable chronobiology, and synthetic-microbiota studies reveal that tryptophan (Trp) metabolism is regulated by distinct cellular “checkpoints” along the gut–brain axis, finely modulated by sex differences, circadian rhythms, and microbiome composition. However, current interventions tackle single levers in isolation, leaving a key gap in precision control of Trp fate. To address this, we drew upon an extensive body of primary literature and databases, mapping enzyme expression across tissues at single-cell resolution and linking these profiles to clinical trials investigating dual indoleamine 2,3-dioxygenase (IDO1)/tryptophan 2,3-dioxygenase (TDO2) inhibitors, engineered probiotics, and chrono-modulated dosing strategies. We then developed decision-tree algorithms that rank therapeutic combinations against biomarker feedback loops derived from real-time saliva, plasma, and stool metabolomics. This synthesis pinpoints microglial and endothelial KP hotspots, quantifies sex-specific chronotherapeutic windows, and identifies engineered Bifidobacterium consortia and dual inhibitors as synergistic nodes capable of reducing immunosuppressive KYN while preserving neuroprotective kynurenic acid. Here we highlight a framework that couples lifestyle levers, bio-engineered microbes, and adaptive pharmaco-regimens into closed-loop “smart protocols.” By charting these intersections, the study offers a roadmap for biomarker-guided, multidisciplinary interventions that could recalibrate KYN metabolic activity across cancer, mood, neurodegeneration, and metabolic disorders, appealing to clinicians, bioengineers, and systems biologists alike.
Keywords:
tryptophan
; kynurenine
; indoleamine 2
; 3-dioxygenase
; tryptophan 2
; microbiotal brain-gut axis
; circadian rhythm
; chronotherapy
; probiotics
; precision medicine
1. Introduction
Life on Earth has been inextricably linked to a dynamic coexistence with microbes [1]. From the earliest stages of development through aging, humans are continuously shaped by microbial signals encountered through environmental contact, diet, and respiration [2]. Nowhere is this symbiosis more profound than in the gastrointestinal tract, where a densely populated and metabolically diverse microbiota transforms dietary substrates into bioactive molecules with far-reaching systemic effects—including on the brain [3]. Among the complex network of metabolic interactions bridging the gut and distant organs, the tryptophan (Trp) metabolic axis has emerged as a pivotal regulator of immune homeostasis, neurophysiological integrity, and energy balance [2]. Beyond serving as a precursor for serotonin and niacin, Trp is enzymatically channeled by both host and microbial systems into the kynurenine (KYN) metabolic pathway, generating a suite of metabolites capable of modulating inflammatory responses—either dampening immune activation or exacerbating pathological inflammation [4]. Additionally, natural compounds targeting neuroinflammation are gaining attention for their antidepressant potential, offering a complementary pathway to modulate Trp metabolism [5].
Over the past decade, next-generation sequencing and metabolomics have mapped thousands of associations between altered Trp metabolism and diseases as diverse as depression, diabetes, cancer, and Alzheimer’s disease (AD) [6,7]. Over the past decade, groundbreaking research has redefined our understanding of neurological diseases and mental illnesses, laying the groundwork for precision interventions [8,9]. Moreover, the integration of biomarkers and imaging with neuroinflammatory markers offers promising diagnostic and therapeutic insights in AD and related disorders [10]. Similarly, phytochemicals such as phenols, alkaloids, and terpenoids have demonstrated notable neuroprotective effects against neurodegenerative disorders, offering complementary intervention strategies [11,12,13,14]. Yet association is not causation [15]. We still lack a coherent framework linking specific microbial consortia, host enzymes such as indoleamine-2,3-dioxygenase-1 (IDO1) and tryptophan-2,3-dioxygenase-2 (TDO2), and downstream metabolites like kynurenic acid (KYNA) or quinolinic acid (QUIN) to discrete physiological outcomes [16] (Table 1). This gap hampers the design of targeted interventions—whether they involve probiotics, small-molecule inhibitors, or lifestyle prescriptions—that aim to rebalance Trp flux toward health-promoting routes [17]. Recent reconceptualizations propose a paradigm shift in how Trp–KYN metabolism is targeted for innovative clinical interventions [18].
A second challenge is spatial. Traditional bulk assays average signals across tissues and cell types, obscuring metabolic micro-domains that may act as “checkpoints” for Trp fate [19] (Table 1). Recent spatial-omics and single-cell technologies have begun to reveal astrocytic and microglial niches in the human prefrontal cortex, endothelial “gates” at the blood–brain barrier (BBB), and perivascular hubs in peripheral organs where KYN metabolic activity is disproportionately high [20]. These findings demand a rewiring of our mental map: instead of a homogeneous pipeline, Trp metabolism resembles a switchboard with cell-type-specific levers that can be pharmacologically or genetically tuned [21].
Timing is the third frontier. Circadian biologists have long known that virtually every metabolic pathway oscillates over the 24-hour day [21]. Emerging evidence suggests that KYN metabolism is no exception and that sex hormones further modulate these rhythms [22] (Table 1). Clinical trials of chemotherapy and checkpoint inhibitors demonstrate that dosing time can double efficacy or halve toxicity, yet KYN-targeting agents have rarely been tested under chrono-pharmacological designs [23]. Without granular, time-stamped metabolite monitoring, we risk missing critical windows when interventions would be most effective—or least harmful [24].
Technological advances now offer tools to close these knowledge gaps [25]. Stable-isotope tracing in gnotobiotic mice can timestamp flux through IDO1 versus TDO2; single-cell proteomics in intestinal organoids can pinpoint which epithelial or immune subsets respond to specific “metabokines”; and CRISPR-based kill switches or inducible operons allow synthetic consortia to dial metabolite output like a volume knob [25,26,27]. Parallel progress in wearable biosensors, artificial intelligence (AI) feedback loops, and adaptive trial designs promises to link real-time biomarker readouts—such as saliva KYNA or morning KYN/TRP slopes—to dynamic adjustments in drug dose, exercise load, or probiotic composition [28,29,30].
Yet formidable obstacles remain. We lack validated, non-invasive biomarkers that faithfully mirror tissue-level KYN activity [31]. The ecological rules governing colonization by engineered microbes are not fully charted, and existing kill-switch circuits require rigorous containment testing [27]. Regulatory frameworks struggle to keep pace with live biotherapeutic products (LBPs) that blur the line between drugs and devices [32]. Finally, ethical and logistical challenges complicate the deployment of adaptive, time-randomized clinical trials that integrate molecular, behavioral, and environmental data streams [33].
Against this backdrop, the present review aims to synthesize cutting-edge insights across microbiology, neuroscience, immunology, and bioengineering to articulate a unified roadmap for precision modulation of the Trp–KYN axis. We first survey the spatial organization of KYN metabolism “checkpoints” in the brain and periphery, highlighting how localized enzyme activity interfaces with systemic immunity and neural circuitry. We then examine sex- and circadian-specific modifiers that dictate when and how the pathway tilts toward neurotoxicity or resilience. Next, we explore microbiota-based strategies—from designer consortia to encapsulated post-biotics—that act as precision switches for Trp flux, and we discuss their manufacturing, safety, and regulatory hurdles. Finally, we outline “Intervention 2.0,” an integrated platform combining dual-enzyme inhibitors, structured exercise, and AI-driven biosensing to create closed-loop therapeutics. By weaving these threads together, we seek to move the field beyond static snapshots toward dynamic, multi-scale models that can predict individual responses and guide adaptive interventions (Table 2). In doing so, we hope to catalyze collaborations between bench scientists, clinicians, data engineers, and regulatory experts, accelerating the translation of Trp-KYN biology into tangible health benefits.
The objectives of this review is to map the cellular and spatial heterogeneity of KYN metabolic activity and identify metabolic “checkpoints” amenable to intervention; to critically appraise evidence for sex- and circadian-dependent modulation of Trp metabolism and its clinical implications; to evaluate current and emerging microbiota-based tools for precision control of KYN flux, including synthetic consortia, kill switches, and post-biotic delivery systems; to assess the therapeutic promise and practical challenges of dual IDO1/TDO2 inhibition in conjunction with lifestyle levers such as exercise; and to propose a framework for adaptive, biomarker-guided clinical trials that integrate real-time metabolite monitoring with dosing algorithms. By addressing these aims, the review intends to illuminate fertile research avenues and provide a practical blueprint for next-generation strategies targeting the gut–brain–immune axis via the KYN metabolic pathway.
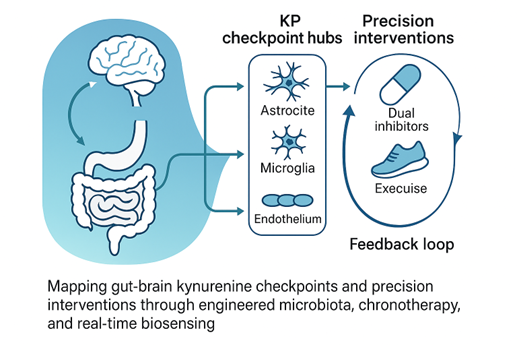
Specialized clusters of microglia, astrocytes, and endothelial cells, known as KYN checkpoints, regulate Trp catabolism and maintain immune balance throughout the brain–body axis. Spatial-omics pinpoints these hubs in the human prefrontal cortex and BBB, where KYN overload erodes neuronal resilience and fuels depression, stroke, and dementia. Peripheral checkpoints likewise calibrate T-cell vigor. Tumors hijack the circuit: chronic IDO1 activity floods the microenvironment with immunosuppressive KYN, yet IDO1 blockade alone falters, bypassed by TDO2 and alternative aryl-hydrocarbon-receptor ligands. Environmental toxins, infections, and diet further tilt these metabolic gates, whereas systemic checkpoints defend serotonin and niacin pools. Critical gaps stall translation. Cell-type-resolved maps of IDO1, TDO2, transporters, and System L remain incomplete; single-cell, longitudinal multi-omics in inflamed tissues and cancers are needed. Real-time biomarkers of KYN burden and transporter flux are lacking, complicating precision dosing. Optimal regimens for dual IDO1/TDO2 or aryhydorcarbon receptor (AhR) blockade—and for brain-penetrant microglial modulators—are undefined. Preclinical models rarely integrate microbiota composition, circadian rhythm, sex, or pollutant exposure, all strong modifiers of KYN metabolism. Bridging these voids will require integrated biosensing, adaptive trials, and machine-learning-guided strategies to tailor checkpoint-directed therapies to individual neuroimmune landscapes.
2. Microbiota-Driven Modulation of Indoleamine-2,3-Dioxygenase-1 (IDO1) and Tryptophan-2,3-Dioxygenase (TDO2) Signaling
Our intestinal microbiota function like molecular "remote controls," fine-tuning dietary Trp fate before it enters systemic circulation [34]. Through sophisticated fermentation and metabolic processing, gut bacteria generate bioactive molecules such as indole derivatives, short-chain fatty acids, and aryl-hydrocarbon-receptor ligands [35]. These small signaling molecules migrate to immune and hepatic tissues, influencing critical enzymes like IDO1 and TDO2 [36]. Consequently, Trp metabolism shifts: toward the KYN pathway when these enzymes are active, or towards serotonin, indole, and protein synthesis when their activity is inhibited [37]. Clinical and experimental studies have connected microbiota-driven dysbiosis and altered metabolite profiles to chronic inflammation and diseases including obesity, type 2 diabetes, human immunodeficiency virus (HIV), colorectal cancer, and AD [38,39,40,41,42]. An elevated KYN-to-Trp ratio, indicative of heightened IDO1 activity, is associated with compromised gut barrier integrity and systemic immune activation [43]. Conversely, higher levels of indole metabolites correlate with strengthened epithelial barriers and reduced inflammation [44]. Dietary interventions, particularly promoting fiber-rich taxa such as Bifidobacterium and Lactobacillus, enhance protective indole and short-chain fatty acid production, counteracting excessive KYN synthesis and promoting healthier metabolic pathways [45].
2.1. Literature Review: Microbial Metabolites as Modulators of Intestinal Integrity and Systemic Disease
Gut microbiota profoundly influence host health through their role in Trp metabolism [46]. Microbial metabolites like KYN, tryptamine, and indole-3-propionic acid regulate critical pathways related to immune, neurological, and metabolic health [47]. Notably, microbiota-produced KYN derivatives significantly impact intestinal permeability and obesity, highlighting their broad physiological relevance [48]. Additionally, these microbial metabolites demonstrate both pro- and anti-inflammatory effects, positioning them as promising therapeutic agents for managing intestinal inflammation and associated disorders, including inflammatory bowel disease and irritable bowel syndrome [49]. Dysbiosis-induced disruptions in Trp metabolism have also been implicated in autoimmune disorders and ischemic stroke, suggesting significant clinical implications and potential therapeutic benefits of dietary strategies involving food homologous plants [16]. Germ-free animal models further underline the essential role of microbiota in guiding host Trp metabolism, reinforcing the microbiome’s critical function as a regulator of health, aging processes, and disease progression [50].
2.2. Research Gaps: Gaps in Dosing Strategies, Longitudinal Efficacy, and Mechanistic Insights
Current research gaps include the need for clearer insights into optimal dosing and intervention strategies, particularly in clinical applications such as haploidentical transplantation, where long-term outcome data remain limited [51]. Additionally, the precise mechanisms underlying sustained benefits, as demonstrated in long-term diabetes remission trials, require further exploration to enhance replicability and scalability [52]. In educational and psychological contexts, while significant improvements in creativity through musical interventions are documented, there is still limited understanding of long-term impacts and the comparative effectiveness of different teaching methodologies [53]. Moreover, longitudinal health outcome studies, such as those examining coronavirus disease 2019 (COVID-19) survivors, underscore the necessity of extended follow-ups to better understand persistent symptoms and recovery trajectories, emphasizing gaps in comprehending chronic implications [54]. Taken together, these findings underscore the need for extended longitudinal, comparative, and mechanistically oriented studies across clinical, educational, and therapeutic domains to refine and empirically validate intervention efficacy [55].
2.3. Time-Stamped Isotope-Tracing in Gnotobiotic Mice Can Tag Flux Through Indoleamine-2,3-Dioxygenase-1 (IDO1) Versus Tryptophan-2,3-Dioxygenase (TDO2)
Emerging research highlights crucial insights into precisely distinguishing Trp metabolism fluxes mediated by IDO1 and TDO2, especially within the context of cancer progression and immune modulation [56]. Advanced techniques such as time-stamped isotope tracing in gnotobiotic mice offer significant promise for accurately mapping metabolic activity between these pathways [56]. Current evidence indicates dual inhibition of IDO1 and TDO2 surpasses single-target strategies, effectively reducing immunosuppressive KYN metabolites and enhancing anti-tumor immune responses across various cancers [57,58]. Nonetheless, substantial gaps remain in understanding precise metabolic dynamics, particularly in humanized mouse models designed to mimic human microbiota interactions [58]. Further insights have been gleaned from enzyme knockout studies demonstrating how disruption of KYN pathway genes impacts mitochondrial respiration and energy homeostasis in the brain [59]. Recent knockout models targeting kynurenine aminotransferase (KAT) enzymes have revealed heightened oxidative and excitatory stress responses, offering mechanistic insights into depression and post-traumatic stress disorder-like behaviors [60]. Meanwhile, rational drug design targeting the KYN pathway is being explored to fine-tune neuroprotective outcomes without tipping the balance toward neurotoxicity [61]. Future research should prioritize detailed mechanistic exploration through robust isotope tracing methodologies, enabling clearer delineation of the specific roles IDO1 and TDO2 play in disease pathology [57]. Additionally, further development and clinical validation of potent dual IDO1/TDO2 inhibitors are critical next steps to fully harness their therapeutic potential and optimize immunotherapeutic strategies across multiple malignancies [62].
2.4. Single-Cell Proteomics in Intestinal Organoids Could Reveal Which Epithelial or Immune Subsets Sense Each “Metabokine.”
Emerging research underscores the potential of single-cell proteomics in intestinal organoids as a powerful tool for clarifying which epithelial or immune subsets respond to specific "metabokines [63]." Single-cell resolution can unveil distinct proteomic signatures, enabling precise identification of cells sensing these metabolic signaling molecules [63]. Recent advancements demonstrate that intestinal organoids effectively model complex physiological responses, such as ischemia-reperfusion injury and gluten-induced inflammation seen in celiac disease, highlighting their suitability for detailed proteomic investigations [64]. Additionally, single-cell transcriptomics has successfully characterized specialized enteroendocrine cell populations, further emphasizing the feasibility and importance of single-cell analyses in organoid cultures [65]. Nevertheless, substantial research gaps persist in connecting specific metabolites or "metabokines" to their target cell types [63]. Future studies should employ single-cell proteomics to systematically map interactions between these bioactive metabolites and distinct cellular subsets within organoid models [63]. Crucial next steps involve refining protocols for isolating and analyzing pure populations of epithelial and immune cells, as well as developing robust analytical methods to integrate proteomic and transcriptomic data [65]. This integrated approach promises deeper insights into gut physiology and pathology, guiding targeted therapeutic interventions for intestinal and metabolic disorders [63].
2.5. Synthetic Consortia with Inducible Kyn Operons Would Let Us Dial Metabolite Output like a Volume Knob
Emerging research highlights the potential of synthetic microbial consortia with inducible KYN operons as sophisticated tools for precisely tuning metabolite production [66]. Utilizing inducible gene expression systems, these engineered microbial communities allow metabolite output to be modulated like a "volume knob," providing highly controlled, dynamic responses tailored to specific applications [67]. Studies employing synthetic consortia, particularly those leveraging cross-feeding and inducible metabolic pathways, demonstrate their capacity for robust and tunable synthesis of desired compounds, such as antioxidants and secondary metabolites [68]. For instance, yeast-based consortia effectively enhanced resveratrol production by precisely managing metabolic interactions. Further, advancements in understanding inducer-producer systems and integrating genomic, proteomic, and machine-learning approaches have improved consortium stability, efficiency, and predictability [69,70]. Despite promising initial results, substantial work remains in optimizing consortia dynamics, including refining inducer responsiveness, ensuring population stability, and developing standardized, scalable frameworks [71]. Next steps should focus on advanced computational modeling and experimental validation, employing genome-wide screens and emerging technologies to fine-tune induction mechanisms [72]. Ultimately, synthetic consortia with precisely controlled KYN operons hold transformative potential for targeted metabolic engineering and biomedical applications.
3. Kynurenine (KYN) Metabolic Pathway “Checkpoints” in the Brain’s Cellular Grid
The brain’s intricate cellular network is governed by distinct "checkpoints" that regulate critical metabolic pathways, notably the KYN metabolic pathway [73]. Recent advancements using spatially-resolved omics have uncovered specialized microglial and astrocytic niches within the human prefrontal cortex, which display notably elevated activity toward KYN metabolism [74]. Additionally, specific endothelial clusters located at neurovascular units show significant KYN metabolic signatures, suggesting their crucial roles as regulatory gates controlling permeability and transport functions at the BBB [75]. These novel insights provide a detailed spatial mapping of KYN metabolism modulation across the brain's cellular landscape, emphasizing its essential function in neuroimmune communication and neuronal homeostasis [76]. Advances in Parkinson’s disease research underscore how metabolic dysregulation, particularly in mitochondrial and KYN pathways, contributes to neurodegeneration [77]. Elucidating the precise locations and mechanisms of these KYN checkpoints enhances our understanding of how Trp metabolism contributes to maintaining brain health and how disruptions in this pathway might underpin various neurological and psychiatric disorders [78]. Disruptions in KYN metabolism have also been implicated in age-associated vascular dysfunction and sarcopenia, further linking metabolic decline to cognitive vulnerability [79]. Evidence from metabolic interventions, including AdipoRon treatment, supports the notion that correcting mitochondrial and inflammatory dysfunctions can mitigate neurodegenerative processes [80]. Future research focusing on these checkpoint regions promises innovative approaches to therapeutic interventions aimed at adjusting KYN activity, offering potential improvements in managing neurological health and disease [81,82,83,84].
3.1. Literature Review: Mapping Kynurenine (KYN) Dynamics Across Neurovascular and Immune Landscapes
KYN “checkpoints” distribute across the brain and periphery, steering Trp catabolism and dictating immune tone [85]. Spatial-omics now map microglial, astrocytic, and endothelial niches in the human prefrontal cortex where KYN enzymes are up-shifted, flagging BBB gates that couple metabolism with neuroimmune signaling [86]. At these hubs, surges in KYN dampen neuronal resilience, promote cerebrovascular inflammation, and correlate with depression, dementia, and stroke [87]. Outside the CNS, the same metabolic nodes calibrate T-cell fitness [88]. IDO1 overactivation floods tumors with immunosuppressive KYN, paralyzing cytotoxic responses; yet IDO1 inhibitors alone have disappointed, unveiling redundant escape circuits [89]. Environmental pollutants and viral infections further hack these checkpoints, amplifying cytokine storms by tipping regulatory T cell (Treg)/T helper 17 cell (Th17) balance [90]. Conversely, systemic checkpoints safeguard Trp homeostasis; deficiency or gene defects can trigger hypertryptophanemia, disturbing serotonin and niacin biosynthesis [88]. Next-wave interventions pair metabolic brakes with programmed cell death protein 1 (PD-1)/programmed cell death ligand 1 (PD-L1) or heat shock protein 90 (HSP90) blockade, deploy aryl-hydrocarbon-receptor antagonists, phytochemicals, or AhR inhibitors such as BAY2416964 to restore immunity and curb tumor growth [91]. These innovative models, complemented by optogenetic technologies, represent the forefront of translational neuroscience [92]. Brain-penetrant molecules that selectively damp microglial KYN flux while sparing peripheral metabolism could deliver precision therapy for neuropsychiatric disorders [93]. Integrating multi-omics with machine learning will rank checkpoint hierarchies, predict compensatory loops, and guide patient-specific cocktails that re-balance neuroimmune networks and prevent KYN-driven pathology [94].
3.2. Research Gaps: Mapping, Monitoring, and Modulating Kynurenine (KYN) Checkpoints Across Systems
KYN checkpoints—specialized microglial, astrocytic, and endothelial clusters—regulate Trp catabolism and immune tone across the brain–body axis [95]. Spatial-omics pinpoints these hubs in the human prefrontal cortex and BBB, where KYN overload erodes neuronal resilience and fuels depression, stroke, and dementia [96]. Peripheral checkpoints likewise calibrate T-cell vigor [95]. Tumors hijack the circuit: chronic IDO1 activity floods the microenvironment with immunosuppressive KYN, yet IDO1 blockade alone falters, bypassed by TDO2 and alternative AhR ligands [89]. Environmental toxins, infections, and diet further tilt these metabolic gates, whereas systemic checkpoints defend serotonin and niacin pools [97]. Critical gaps stall translation. Cell-type-resolved maps of IDO1, TDO2, transporters, and System L remain incomplete; single-cell, longitudinal multi-omics in inflamed tissues and cancers are needed [96]. Real-time biomarkers of KYN burden and transporter flux are lacking, complicating precision dosing [96]. Optimal regimens for dual IDO1/TDO2 or AhR blockade—and for brain-penetrant microglial modulators—are undefined [98]. Preclinical models rarely integrate microbiota composition, circadian rhythm, sex, or pollutant exposure, all strong modifiers of KYN tone [97]. Bridging these voids will require integrated biosensing, adaptive trials, and machine-learning-guided strategies to tailor checkpoint-directed therapies to individual neuroimmune landscapes [96,99,100].
3.3. CRISPRi “Zip-Codes” Delivered by Adeno-Associated Virus (AAV) Can Silence Kynurenine 3-Monooxygenase (KMO) or Kynureninase (KYNU) only in Perivascular Endothelium and Watch Downstream Glutamatergic Sync Crash—or Not
Picture an AAV vector outfitted with endothelial “zip-codes”—tight-junction promoters plus microRNA target sequences for off-cell clearance—delivering a zinc finger protein 3 (ZIM3)-Kruppel-associated box (KRAB) clustered regularly interspaced short palindromic repeats interference (CRISPRi) cassette that knocks down kynurenine 3-monooxygenase (KMO) or kynureninase (KYNU) exclusively in perivascular endothelium [101]. Two clues drive this strategy. First, perivascular cells are gatekeepers for KYN flux into the brain: endothelium converts circulating KYN into 3-HK and QUIN, feeding astrocyte–microglia glutamatergic coupling [102]. Second, KMO overactivity fuels cancer growth, neurodegeneration, and immune escape, yet systemic inhibition risks off-target nicotinamide adenine dinucleotide (NAD) imbalance [103]. Cell-restricted CRISPRi sidesteps that liability, and the ZIM3 domain offers potent, reversible silencing [104]. Sex- and circadian-modulated shifts in neurodegeneration also modulate vulnerability to cognitive and mood disturbances [105]. If endothelial KMO/KYNU expression drops, 3-HK and QUIN output should collapse, starving downstream N-methyl-D-aspartate (NMDA) -sensitizing signals and “crashing” the synchronized glutamatergic surge that drives excitotoxicity and tumor immune evasion [106]. Next steps include build bar-coded AAV libraries to refine endothelial specificity; validate knock-down efficiency and KYN metabolites flux in brain-slice co-culture; monitor glutamate dynamics with optogenetic reporters in vivo; and assess effects on tumor infiltration and behavior in KMO-high breast-cancer metastasis models [102]. Parallel safety screens must chart NAD pools and mitochondrial stress in non-target tissues [107] (Table 3). Success could open a precision channel to modulate neurovascular KYN flux without systemic collateral damage [101].
3.4. Light-Addressable Riboswitches Could Let Us Pulse KP Enzymes in Astrocytes and Read Real-Time Calcium Waves
Light-addressable riboswitches offer a precision dimmer for the KYN metabolic pathway inside astrocytes [108]. By coupling a photocleavable aptamer to KMO or KYNU transcripts, pulsed infrared or nanosecond-visible light can switch enzyme translation on or off within milliseconds, riding on the same wavelengths already proven to trigger or image astrocytic Ca2+ waves [109]. The concept rests on two converging clues: riboswitches endowed with the Z-lock or photocleavable linker reliably gate gene expression in living cells, and pulsed light drives robust, tunable calcium oscillations in astrocytes without overt photodamage, letting us read the metabolic consequences in real time [110] (Table 4). A burst of light thus delivers a double payload—inducing a KYN metabolism surge while simultaneously recording its impact through genetically encoded calcium indicator based on calmodulin and M13 peptide fused to green fluorescent protein (GCaMP) -tagged calcium reporters or two-photon glutamate sensors [110]. Key next steps include engineering astrocyte-specific AAVs carrying the light-gated riboswitch, benchmarking translation kinetics versus calcium rise in organotypic slices, and mapping how localized KYN pulses propagate through extracellular gliotransmitter waves [111]. In vivo, fiber-coupled two-photon uncaging combined with wide-field Ca2+ imaging can reveal whether transient KMO activation amplifies or quenches network excitability during sleep, seizure, or learning [110]. Ultimately, this optogenetic-metabolic fusion could dissect causality between KYN flux and astrocyte-driven neurophysiology with unprecedented temporal resolution [108].
4. Sex and the Circadian City: Hidden Modifiers
Circadian rhythms and biological sex weave an under-appreciated backdrop to KYN metabolism, subtly steering mood and immune tone across the day [112]. Sex hormones and circadian rhythms likewise modulate migraine vulnerability, further emphasizing the need for personalized neuroimmune interventions [113]. Immune challenges such as interferon-α therapy consistently lower circulating Trp, boost KYN, and heighten depressive symptoms, but these shifts peak at distinct clock phases and manifest more severely in women [114]. Preclinical lipopolysaccharide models echo this dimorphism, with female mice displaying protracted neuroimmune and behavioral scars that fluoxetine can erase only when given at their active phase [115] Human imaging studies add another layer: reduced Trpand skewed KYN metabolites track hippocampal-subfield atrophy, yet the correlation tightens in early-morning scans, hinting at chronobiological gating [116]. Rapid-acting antidepressants such as ketamine appear to reset both the monoaminergic–glutamatergic interface and downstream KYN flux, but again response rates diverge by sex and time of dosing [114]. Post-mortem data reveal anterior cingulate cortex KYN signatures that segregate by sex and suicide status, underscoring personalized vulnerability windows [116]. Neuroanatomical findings from autism research similarly reveal that circadian and neuroimmune factors may influence structural brain development [117]. Collectively, these findings suggest that sex hormones and the molecular clock act as “hidden modifiers,” dictating when and how immune activation tilts KYN balance toward neurotoxicity or resilience [118]. Similar to the evolving understanding in mental health research, recognizing sex- and circadian-dependent variability supports a dimensional view of disease vulnerability and resilience [119]. Parsing these temporal-sexual intersections could unlock chronotherapy strategies for mood disorders.
4.1. Literature Review: Circadian Misalignment, Mood Vulnerability, and Emerging Chronotherapeutics
Longitudinal wearables reveal that phase delays in core-body temperature and melatonin release often precede mood dips in major depression and bipolar disorder, supporting circadian realignment as a digital-medicine target [120]. In healthy adults, endogenous 24-hour rhythms modulate anxiety- and depression-like affect, with later dim-light melatonin onset and compressed phase-angle differences predicting poorer mood [121]. Disruption of the molecular clock by hypercortisolism in Cushing’s syndrome flattens immune-cell oscillations and skews steroid profiles, illustrating endocrine–immune crosstalk [122]. Similar circadian misalignment exacerbates autoimmunity in lupus, acting as a prodromal biomarker for flares [123]. Non-pharmacologic interventions show promise: group music therapy realigns autonomic and melatonin rhythms in depressed women, while optimal circadian–circasemidian coupling buffers morning blood-pressure surges and fosters resilience [121]. Nutritional modulation is equivocal; omega-3 supplementation dampens inflammation yet leaves KYN metabolism and mood unchanged in healthy men [124]. Experimental LPS infusion acutely activates KYN metabolism, linking immune challenge, chronobiology, and affect in real time [124]. Adolescents display bidirectional pathways: anxiety forecasts future sleep disruption, whereas a genetically longer free-running period in males predicts later mood vulnerability [123]. Collectively, these studies underscore circadian alignment as a modifiable lever for mood regulation and highlight the need for personalized chrono-therapeutic strategies across lifespan and disease contexts.
4.2. Research Gaps: Timing, Sex, and Biomarker Integration for Precision Kynurenine (KYN) Intervention
Despite compelling evidence that dosing time and sex strongly influence chemotherapy and immunotherapy outcomes, major knowledge gaps limit translation to KYN-targeting agents [125]. First, no clinical trials have yet tested chrono-pharmacology for IDO1/TDO2 inhibitors, KMO blockers, or KAT activators, leaving optimal schedules unexplored [126]. Second, existing studies rarely stratify by both circadian phase and biological sex; pharmacokinetic data disaggregated for women are virtually absent, obscuring why afternoon regimens reduce toxicity in female lymphoma patients [125]. Third, mechanistic links between peripheral clock genes, immune cell oscillations, and drug metabolism remain undefined, particularly how estrogen and glucocorticoids modulate daily fluctuations in KYN enzyme activity [127]. Fourth, wearable-derived chronotypes are not integrated into trial design, preventing personalized timing algorithms that could harmonize drug exposure with individual rhythms [125]. Fifth, preclinical tumor models often use male rodents housed in static light cycles, overlooking sex- and circadian-dimorphic responses seen in patients [128]. Finally, validated real-time biomarkers that couple KYN metabolite swings to treatment efficacy are lacking, hindering adaptive dosing [129] (Table 5). Addressing these gaps will require multi-omic chronopharmacology studies, sex-balanced animal models, and adaptive clinical trials embedding circadian sensors to craft precision schedules for KYN-focused therapies.
4.3. Multi-Time-Point Plasma Kynurenine (KYN) Profiles Stratified by Sex and Hormonal Phase
Current evidence shows that plasma KYN levels fluctuate with sex, age, and endocrine status, yet most datasets rely on single fasting draws [130]. Parkinson’s and breast-cancer cohorts reveal disease-specific shifts toward neurotoxic QUIN, but without high-resolution sampling it is unclear whether these changes reflect trait abnormalities or time-of-day/hormone-phase effects [131]. Metabolomic screens (KarMeN) can classify sex and age, implying a strong biological signal, while ambulatory microdialysis (U-RHYTHM) now permits hourly hormone/metabolite capture—an underused tool for KYN kinetics [131]. Feminizing gender-affirming hormone therapy reshapes amino-acid pools after six months; however, acute diurnal patterns and menstrual-cycle nuances remain unmapped [132]. Animal work shows estrous-dependent adipokine oscillations, hinting that luteal-phase progesterone surges might also bias KYN flux [132]. No study yet aligns luteal sub-phases, cytokine pulses, and KYN derivatives in healthy women, nor measures phase-specific KYN shifts during immunotherapy or antiandrogen treatment [130]. Next steps: deploy wearable-triggered, multi-time-point plasma collection across 24-h and menstrual cycles; integrate liquid chromatography–mass spectrometry (LC-MS) panels for KYN, Trp, QUIN, and KYNA with sex-hormone, cortisol, and brain-derived neurotrophic factor (BDNF) profiles; and model trajectories using mixed-effects chronopharmacology frameworks. Such datasets will clarify whether timing and hormonal milieu confound or mediate KYN biomarkers, guiding precise sampling windows and sex-hormone-aware interventions.
4.4. Wearable Light-Exposure + Metabolite Logging to See If Circadian Misalignment Exaggerates the Quinolinic Spike
Circadian misalignment (CM) rewires metabolic, cardiovascular, and immune circuits, yet whether it magnifies neurotoxic (QUIN) surges remains untested [133]. Night-shift paradigms reveal clock-driven insulin resistance, lipidome disruption, and blood-pressure creep, while murine models link chronic CM to immune senescence and shortened lifespan [134]. Sex adds complexity—females show partial protection against CM on a high-fat diet—suggesting divergent QUIN trajectories [135]. Light timing is the master zeitgeber; continuous lux logging via smartwatches can quantify misalignment magnitude, whereas ambulatory microdialysis or dried-blood-spot kits now enable multi-time-point KYN metabolite sampling [123]. Pairing these technologies would let researchers correlate light-phase offsets with 24-h QUIN area-under-curve, stratified by sex and feeding rhythms [136]. Key next steps includes pilot a cross-over study where shift-workers wear light- and activity-trackers plus collect hourly capillary samples across two work cycles; model QUIN dynamics versus lux-derived phase angle using mixed-effects chronobiology; overlay cortisol and melatonin rhythms to disentangle stress versus circadian effects; and test whether timed blue-light blockers, melatonin, or time-restricted feeding blunt QUIN spikes [136] (Table 6). Such integrative phenotyping could identify high-risk chronotypes and guide precision-timed KP interventions to mitigate CM-induced neuroinflammation.
4.5. Adaptive Trial Designs that Randomize Dose-Timing Rather Than Just Dose Size
Bayesian and response-adaptive frameworks have revolutionized dose-finding, yet virtually all published trials modulate quantity, not clock time [137]. Radiation for pancreatic cancer, ketamine infusions for late-life depression, and molnupiravir for early COVID-19 show how real-time efficacy–toxicity trade-offs hone dose size, but none test whether morning versus evening delivery alters those curves [138]. Chronotherapy evidence from chemotherapy and immune checkpoint inhibitors indicates that timing can double efficacy or halve toxicity, with sex as a major moderator, underscoring a missed opportunity [139]. Key gaps include statistical models that treat dosing time as a continuous, circadian-anchored covariate; real-time biomarkers (e.g., actigraphy-derived phase angle) to guide allocation; and operational logistics for pharmacy and nursing around-the-clock interventions [137]. Next steps include simulate Bayesian hierarchical designs where dose level and dosing hour are co-randomized, borrowing strength across adjacent time bins; integrate wearable-captured chronotype to stratify randomization and inform priors; embed rolling interim analyses that drop unfavorable time windows rather than doses; and pilot such designs in drugs with known chrono-toxicities, using point-of-care melatonin or cortisol assays as safety triggers [140] (Table 7). Developing regulatory guidance and EHR-linked scheduling tools will be crucial to mainstream adaptive chrono-trials, paving the way for precision-timed KP inhibitors and beyond [139].
5. Microbiota Engineering as a Precision Switch
Engineering the gut microbiome now offers a “precision switch” for neuroimmune circuits, with strain-level interventions poised to toggle mood and metabolic health on demand [141]. In murine models of chronic stress, supplementation with Bifidobacterium pseudonumeratum W112 reversed anhedonia and hepatic injury by recalibrating the gut-liver-brain axis, whereas B. breve and B. longum strains restored brain-derived neurotrophic factor signalling and serotonergic precursors through 5-hydroxy-tryptophan synthesis [142]. These psychobiotic effects align with peripheral data: patients with major depressive disorder show depleted Bifidobacterium and Lactobacillus counts, implicating taxa scarcity as a modifiable risk [143]. Importantly, anti-depressive benefits persist even with heat-killed cells, underscoring that microbial metabolites—not colonization per se—drive behavioural rescue [142]. Precision editing could therefore install synthetic operons for tryptophanase, KATs, or short-chain-fatty-acid production to steer host Trp flux toward neuroprotective pathways and away from QUIN toxicity [144]. Key next steps include high-resolution metagenomics to map strain-specific metabolic fingerprints, CRISPR-guided promoter tuning for inducible output, and gnotobiotic “plug-and-play” consortia to test combinatorial logic [145]. Adjunctive use of compounds like curcumin has been associated with improvements in microbiota composition and mood, reinforcing its role in gut–brain axis modulation [146]. Longitudinal trials integrating faecal metabolomics with mood and liver biomarkers will validate whether engineered consortia can act as dosage-controlled switches, heralding tailor-made microbiota therapeutics for depression and comorbid metabolic disease [147].
5.1. Literature Review: Microbiota-Targeted Strategies for Modulating Mood and Inflammation
Clinical and preclinical data increasingly connect gut-directed interventions to mood improvement [148]. A randomized trial showed Bifidobacterium breve CCFM1025 alleviates major depression while normalizing Trp-KYN balance and microbial diversity [149]. Complementary protocols—PRO-DEMET and PROVIT—deploy B. longum, L. helveticus, and biotin to probe combined metabolic-microbiome effects; preliminary findings suggest inflammation drops even when mood shifts are modest [149]. Beyond bifidobacteria, Akkermansia muciniphila and Lactobacillus reuteri mitigate stress-induced anhedonia in mice, accompanied by restored neurotrophins and lipid metabolism. A single FMT case demonstrates symptom remission in refractory depression, echoing murine transfers where microbiota from depressed women induce affective and fatty-acid perturbations in recipients. Non-bacterial strategies also show promise: low-dose linoleic acid rescues serotonin and microbial diversity, while Banxia Xiexin decoction adjusts the gut–lipid axis to relieve comorbid atherosclerosis and depression. Psychosocial immersion programs reveal that Prevotella-rich shifts correlate with lowered inflammatory tone and mood gains, underscoring environment–microbiome interplay. Collectively, these studies depict a converging theme: targeted modulation—whether via single strains, multi-strain consortia, diet, or behavioral change—can rebalance microbial metabolites, dampen inflammation, and lift mood. Future trials must standardize endpoints, stratify by metabolic comorbidity, and incorporate multi-omics to pinpoint responders and refine dosing regimens.
5.2. Research Gaps: Live Biotherapeutic Products Against Multi-Drug Resistant (MDR) Enteric Pathogens: Research Gaps
Proof-of-concept studies show engineered or defined consortia can decolonize carbapenem-resistant Klebsiella and other MDR organisms without disrupting resident flora, yet translation stalls on several fronts [150]. First, colonization remains unpredictable: VE303 engrafts only after antibiotic conditioning, while VE707’s murine efficacy lacks human pharmacokinetic analogues [151]. No head-to-head trials compare synthetic consortia with FMT, leaving efficacy drivers—bacteriocins, phages, or niche competition—unclear [152]. Second, durability data are scarce; longitudinal sequencing in patients post-FMT hints at phage-mediated suppression, but mechanistic dissection of phage–bacteria–host interplay is missing [153]. Third, safety and horizontal-gene-transfer risks are under-characterized—engineered E. coli secreting microcins could acquire resistance cassettes in vivo [154]. Fourth, manufacturing and QC frameworks lag behind pharmaceutical standards; batch-to-batch metabolite output is unverified for multi-strain LBPs [151]. Fifth, adaptive trial designs that modulate dose timing relative to antibiotics or diet are untapped, despite evidence that feeding rhythms and bile acids gate colonization resistance [155]. Finally, regulatory pathways for genetically modified LBPs remain fragmented across jurisdictions, deterring investment [151] (Table 8). Addressing these gaps demands standardized engraftment biomarkers, phage-omics integration, gnotobiotic validation of mechanism, and harmonized GMP guidelines to accelerate safe, predictable, and durable microbiota-based decolonization strategies.
5.3. Designer Strains with Kill-Switches and Inducible Kynurenine Aminotransferase (KAT) Expression
Recent CRISPR-based kill switches in E. coli Nissle prove that dual chemical-temperature triggers can achieve >105-fold clearance in the murine gut while retaining plasmid integrity over weeks—establishing a benchmark for biocontainment [27]. Parallel work in Mycobacterium tuberculosis, Bacillus Calmette-Guérin (BCG), and Pseudomonas fluorescens underscores two recurring clues: circuit simplicity boosts genetic stability, and layered toxins curb escape [156]. Yet open questions persist. First, evolutionary pressure within complex microbiomes may select for cryptic recombination events; long-read metagenomics of shed strains is needed to quantify real-world escape rates [157]. Second, few designs test function across variable pH, bile acids, or host temperatures; standardized “gut stress” challenge panels should precede human dosing [158]. Third, redundancy remains underused—stacking orthogonal CRISPRi, toxin-antitoxin, and auxotrophy modules could create multi-lock systems that tolerate single-node failure [159]. Fourth, kill-switch burden on therapeutic payloads is rarely measured; metabolic toggle libraries and inducible promoters from xenogeneic silencing studies could minimize fitness costs [160]. Finally, regulatory pathways are hazy—agencies require validated shutoff diagnostics and environmental fate data [161]. Next steps include head-to-head comparisons of toxin cassettes, adaptive evolution assays in bioreactors seeded with human stool, and digital-polymerase chain reaction (PCR)-based field tests to monitor persistence post-shedding [162]. Such work will convert kill switches from academic prototypes into deployable safeguards for live biotherapeutic products.
5.4. Encapsulated “Post-Biotics” (e.g., Stabilized KYNA) to Bypass Colonization
Skipped-colonization formulations—enteric capsules, pH-responsive hydrogels, and microfluidic microparticles—now allow direct delivery of stabilized KYNA and other post-biotics to the colon, sidestepping the variability of live engraftment [163]. Hydrogel and chitosan-alginate matrices protect labile metabolites from gastric acid, while spray-dried enteric microspheres and 3-D-printed capsules release cargo only at ≥pH 7, mirroring colonic transit [164]. Encapsulation reviews highlight that payload stability, release kinetics, and mucus penetration govern in vivo bioavailability, yet KYNA has never been loaded into these systems, and its solubility profile may demand excipient optimization [165]. Phage and enzyme encapsulation studies suggest co-packaging metabolite pumps or β-glucuronidases could boost local concentration, but dose-response curves for post-biotic neuroprotection remain undefined [166]. Next steps include screen generally recognized as safe (GRAS)-grade polymers for KYNA compatibility under accelerated aging; map release profiles in simulated gastrointestinal fluids and pig colonic explants; employ near-infrared–triggered nanocaps to test on-demand bursts during inflammation; and quantify systemic versus luminal KYNA using LC-MS in gnotobiotic mice, benchmarking against Bifidobacterium-produced levels [167]. Parallel human pilot studies can deploy encapsulated KYNA alongside wearable pH sensors to correlate release timing with mood and sleep metrics [168] (Table 9). Success would validate post-biotic capsules as a low-variance, regulatory-friendly alternative to live psychobiotics [169].
5.5. Cloud-Linked Stool Metabolomics Dashboards to Guide Weekly Probiotic Titration
Remote probiotic trials in COVID-19 and depression show patients can self-collect and courier fecal samples, while digital platforms already monitor calprotectin, zonulin, and secretory immunoglobulin A (sIgA) [170]. Yet strain-specific responses vary and optimal titration schedules remain guesswork [171]. Cloud-linked dashboards that stream LC-MS metabolite panels—short-chain fatty acids, indoles, bile-acid ratios—could provide weekly feedback loops to adjust probiotic dose or strain composition [172]. Key clues are multispecies probiotics shorten gut transit time and shift microbiota in constipation meta-analyses, but responders stratify by baseline diet and Lactobacillus colonization predictors such as cheese and n-3 fatty acid intake; probiotics modulate clock-gene expression and the gut–lung axis, implying time-of-day and symptom-phase windows for dosing; large-scale genome scans map foodborne lactic acid bacteria that could seed personalized consortia [170] (Table 10). Gaps include lack of validated metabolite thresholds that signal “dose-too-low” versus “overshoot,” and absence of algorithms that incorporate diet, sleep, and medication logs alongside omics [173]. Next steps include build a reference library of weekly stool metabolomes from diverse cohorts on fixed probiotic regimens; train adaptive Bayesian models that recommend titration when variance-normalized short-chain fatty acids (SCFAs) or indole scores drift beyond control limits; integrate wearable-captured feeding rhythms to schedule capsule timing; and run N-of-1 crossover trials to benchmark dashboard-guided titration against static dosing [174]. A successful system would transform probiotics from one-size-fits-all supplements into dynamic, biomarker-steered therapies [175] (Table 7).

Table 10.
Key Clues for Personalized Probiotic Development. This table summarizes critical insights into probiotic responsiveness and personalization, emphasizing diet, circadian timing, and foodborne bacterial strain selection as key factors for optimizing gut microbiome interventions.
Table 10.
Key Clues for Personalized Probiotic Development. This table summarizes critical insights into probiotic responsiveness and personalization, emphasizing diet, circadian timing, and foodborne bacterial strain selection as key factors for optimizing gut microbiome interventions.
| Key Clues | Implication | |
|---|---|---|
| 1. Multispecies probiotics shorten gut transit time and shift microbiota in constipation meta-analyses, with response stratified by baseline diet and Lactobacillus colonization predictors like cheese and n-3 fatty acid intake | Personalize probiotic therapy based on individual dietary patterns and colonization potential | |
| 2. Probiotics modulate clock gene expression and the gut–lung axis, suggesting time of day and symptom phase windows for optimized dosing | Incorporate circadian timing in probiotic administration to enhance therapeutic efficacy | |
| 3. Large-scale genome scans map foodborne lactic acid bacteria, offering candidates to seed personalized probiotic consortia | Develop customized probiotic blends from identified foodborne strains for targeted microbiome modulation |
Table 11.
Next Steps for Adaptive Probiotic Titration Using Stool Metabolomics. This table outlines a strategic roadmap for developing adaptive probiotic titration systems, leveraging stool metabolomics, Bayesian modeling, wearable-derived feeding rhythms, and N-of-1 trial designs to optimize precision microbiome therapies.
Table 11.
Next Steps for Adaptive Probiotic Titration Using Stool Metabolomics. This table outlines a strategic roadmap for developing adaptive probiotic titration systems, leveraging stool metabolomics, Bayesian modeling, wearable-derived feeding rhythms, and N-of-1 trial designs to optimize precision microbiome therapies.
| Next Steps | Purpose | |
|---|---|---|
| 1. Build a reference library of weekly stool metabolomes from diverse cohorts on fixed probiotic regimens | Establish a baseline for microbiome metabolite variation under controlled probiotic interventions | |
| 2. Train adaptive Bayesian models to recommend titration when normalized SCFA or indole scores drift beyond control limits | Enable dynamic adjustment of probiotic dosing based on real-time metabolic biomarkers | |
| 3. Integrate wearable-captured feeding rhythms to optimize capsule timing | Personalize probiotic administration schedules to individual feeding patterns | |
| 4. Run N-of-1 cross-over trials to benchmark dashboard-guided titration against static dosing | Validate the superiority of adaptive, dashboard-driven interventions over conventional static dosing | |
| 5. Build a reference library of weekly stool metabolomes from diverse cohorts on fixed probiotic regimens | Establish a baseline for microbiome metabolite variation under controlled probiotic interventions |
SCFA, short-chain fatty acids.
6. Intervention 2.0: Dual Inhibitors, Exercise, and Real-Time Biosensing
Next-generation therapeutics are converging on a three-pronged strategy to disarm KYNs in cancer and immune dysregulation [176]. First, dual inhibitors such as RY103 and ZC0101 simultaneously block IDO1 and TDO2 (or IDO1 and thioredoxin reductase), outperforming single-agent IDO1 blockade by collapsing redundant enzyme circuits and lowering intratumoral KYN to neuroprotective baselines [177]. Preclinical models show these agents stunt glioma and pancreatic tumor growth by quenching Kyn-AhR signaling and restoring cytotoxic T-cell traction [177]. Second, structured endurance exercise—a potent physiological lever—amplifies KYN re-balancing by accelerating peripheral KYN clearance via skeletal-muscle KAT up-regulation, creating a metabolic “sink” that synergizes with pharmacological inhibition [30]. Third, wearable biosensors now capture continuous lactate, glucose, and microvascular O₂ data, while emerging implantable electrochemical probes detect real-time KYN-to-Try ratios in interstitial fluid [178]. Linking these feeds to adaptive dosing algorithms could personalize dual-inhibitor schedules around exercise bouts, maximizing metabolic windowing and minimizing toxicity [176]. Immediate next steps include human-machine-interface trials that pair IDO1/TDO2 inhibitors with supervised exercise and real-time KYNs monitoring, testing whether algorithm-guided titration enhances tumor response and rescues mood or fatigue [176]. Such “Intervention 2.0” fuses pharmacology, lifestyle, and biosensing into a closed-loop platform poised to redefine KYNs-targeted precision medicine. Newly designed KYNA derivatives, refined by C-3 side chain alterations, are being evaluated for their ability to fine-tune motor and cognitive outcomes [179].
6.1. Literature Review. Dual Inhibition and KP Modulation
Early dual-inhibitor work centres on M4112, the first oral agent targeting both IDO1 and TDO2 [58]. Preclinically, M4112 halved tumor KYN-to-Trp ratios across xenograft panels; phase I results confirmed on-target plasma suppression with manageable fatigue and nausea, yet a compensatory rise in systemic KYN hinted at peripheral sinks that may blunt efficacy [58]. Parallel chemistry screens generated RY103 (IDO1/TDO2) and ZC0101 (IDO1/TrxR), each extending the dual-hit concept to pancreatic and oxidative-stress contexts, respectively [177,180]. These compounds curtailed migration, invasion, and colony formation in cell-line models, supporting broad antitumor potential [177]. Outside cancer, pathway modulation remains complex. N-acetylcysteine attenuates cisplatin-induced cognitive decline by normalizing brain KYNA, whereas TNF-α blockade in rheumatoid arthritis leaves KYN flux largely untouched despite clinical remission, underscoring disease-specific regulation [181,182,183]. Human endotoxin challenge acutely spikes both neurotoxic QUIN and protective KYNA without correlating to sickness behavior, questioning causal links [184]. Observational cohorts add nuance: higher downstream metabolites (xanthurenic and picolinic acids) associate with better survival and reduced fatigue in colorectal-cancer survivors, yet KYNs activation predicts mortality in acute respiratory distress syndrome [185,186]. Collectively, the literature depicts a heterogeneous KYN landscape where dual enzyme blockade is promising but must contend with systemic metabolite reservoirs, context-dependent feedback loops, and variable host outcomes—parameters future trials must quantify.
6.2. Research Gaps: Adaptive Dose-Timing and Real-Time Monitoring
Although adaptive designs are reshaping oncology and metabolic drug development, most trials still optimise dose magnitude while ignoring circadian or activity-linked timing [187]. Continuous glucose-monitoring data show that SGLT2 inhibitors lower glycaemic variability yet raise ketoacidosis risk in type 1 diabetes, but no study dynamically shifts dosing around exercise or nocturnal hypoglycaemia [188]. Therapeutic drug-monitoring platforms for kinase inhibitors confirm exposure–toxicity windows, yet do not integrate wearable-captured vital signs or KYN biomarkers to guide intraday titration [187]. Intermittent schedules for phosphoinositide 3-kinase (PI3K), Bruton's tyrosine kinase (BTK), and Kirsten rat sarcoma viral oncogene homolog (KRAS) inhibitors reduce adverse events preclinically, but statistical frameworks treating “time-on” and “time-off” as co-randomized variables remain undeveloped [189,190]. Next-generation maturation and ropomyosin receptor kinase (TRK) inhibitors collect rich pharmacokinetic data but lack algorithms that pair those curves with real-time biosensor feeds [191,192]. Key gaps therefore include adaptive randomization models that incorporate dosing clock-time as a modifiable arm; validated software bridges between continuous glucose monitoring (CGM), lactate, or KYN sensors and electronic trial master files; safety rules for rapid dose-time shifts in outpatient settings; and patient-reported outcome measures sensitive to circadian toxicity [193,194,195,196] (Table 12). Addressing these deficits will require multidisciplinary consortia linking chronobiologists, bioinformaticians, and trialists to pilot fully time-adaptive protocols for next-gen inhibitors and metabolic adjuvants.
6.3. Phase-Ib “Smart Protocols”: Micro-Dosed Dual Inhibitors Guided by Saliva KYNA Sensors
Phase Ib studies already test dual phosphoinositide 3-kinase delta/gamma isoforms (PI3Kδ/γ) or spleen tyrosine kinase (SYK)/Fms-like tyrosine kinase 3 (FLT3) inhibitors with Bayesian dose-finding, yet none exploit non-invasive biosignatures to steer real-time titration [197]. Portable electrochemical strips now quantify KYNA in saliva within minutes and correlate with plasma levels, creating an opportunity for closed-loop dosing [198]. Conceptually, micro-doses of IDO1/TDO2 inhibitors could be administered every 6–8 h; if post-dose saliva KYNA falls below a personalized threshold, the algorithm triggers the next micro-dose, otherwise delays it—minimizing over-suppression and toxicity [199]. Thermal- and pH-responsive microspheres from dual-stimuli delivery research provide a vehicle for sub-gram, subcutaneous depots that release inhibitor pulses when externally warmed or after a salivary cue [200]. Trial design would integrate Bayesian optimal interval design for Phase 1/2 Trials (BOIN12) utility-based intervals, defining acceptable KYNA suppression windows rather than traditional maximum tolerated dose (MTD) [201]. Key gaps include validation of saliva–tumor KYNA concordance, stability of sensor reagents under recurrent sampling, and pharmacokinetic modelling of micro-pulse dynamics [202,203]. Immediate next steps are run a crossover pharmacokynetics study comparing saliva, plasma, and tumor microdialysate KYNA after RY103 micro-dosing; calibrate the Bayesian control algorithm using simulated patient data; and embed patient-reported fatigue and cognitive scores to test whether KYNA-targeted pacing improves tolerability relative to fixed bis in die (BID) regimens [204,205] (Table 13). Success would pioneer biomarker-responsive phase-Ib frameworks, setting a precedent for precision chronopharmacology.
6.4. AI-Driven Feedback Loops That Auto-Adjust Evening Treadmill Sessions or Probiotic Cocktails Based on Morning KYN/TRP Slope.AI-Driven KYN/TRP Feedback Loops
Emerging hardware and informatics can now pair morning finger-stick KYN-to-TRP slopes with adaptive lifestyle prescriptions that update nightly [206]. AI-controlled treadmills and exoskeletons already modulate belt speed or torque in real time from gait and heart-rate inputs; integrating cloud-fed metabolite data would allow the algorithm to lengthen or intensify an evening run only when the day’s KYN/TRP ratio signals pro-inflammatory drift [87]. Likewise, modular probiotic cocktails that shift gut indole and short-chain-fatty-acid production could be titrated each afternoon, with doses nudged up when the biomarker slope exceeds a personalized threshold [45]. Clues supporting feasibility include: clinical artificial intelligence operations (ClinAIOps) frameworks for continuous therapeutic monitoring in hypertension and diabetes; Kinect- or sensor-driven treadmills that already auto-pace speed by user position; and murine and human studies where tailored probiotic blends reduce intestinal inflammation and modulate TRP metabolism [36] (Table 14). Gaps remain. No longitudinal dataset links daily KYN/TRP excursions with exercise intensity or probiotic-induced metabolomic shifts. Sensor validation for dried-blood-spot or saliva TRP/KYN needs real-world robustness, and reinforcement-learning models must balance metabolic targets against user fatigue and adherence [206]. Next steps: deploy a 12-week N-of-1 crossover where participants collect morning KYN/TRP, receive algorithm-set treadmill or probiotic adjustments, and stream compliance plus mood, glucose, and heart-rate metrics. Success would prototype a closed-loop “exercise-microbiome” medicine that personalizes both movement and microbes to biochemical feedback.
7. Conclusion
Harnessing the gut–KYN axis demands an integrative framework that unites microbiology, neuro-immunology, and chronomedicine. This review demonstrates how spatial “checkpoints,” circadian and sex modifiers, and engineered microbiota collectively steer Trp fate toward either neuroprotection or pathology. By weaving enzymatic dual-inhibition with lifestyle levers—exercise, diet, and real-time biosensing—the authors chart a precision-medicine roadmap that transcends siloed approaches. Theoretical insight lies in reframing the KYN metabolism as a dynamically gated network rather than a linear cascade; practical value emerges in the proposed closed-loop trials that titrate inhibitors, probiotics, or treadmill load to biomarker feedback. Yet critical questions remain: Which cell-specific KYN fluxes truly drive disease? How stable are engineered consortia in the complex gut ecosystem? What statistical architectures best randomize dosing time as well as dose? Addressing these gaps will require single-cell multi-omics, longitudinal metabolite dashboards, and adaptive, sex-balanced clinical designs. These advances are underpinned by AI integration and the development of human-specific platforms, which promise to transform the personalization of neuropsychiatric care [207]. Advancing these fronts will not only refine KYN-targeted therapeutics but also provide a template for biomarker-guided interventions across metabolic and neuroimmune disorders.
Author Contributions
Conceptualization, M.T.; methodology, M.T.; software, M.T.; validation, M.T.; formal analysis, M.T.; investigation, M.T.; resources, M.T.; data curation, M.T.; writing—original draft preparation, M.T.; writing—review and editing, M.T. and L.V.; visualization, M.T.; supervision, M.T. and L.V.; project administration, M.T. and L.V.; funding acquisition, M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research, Development, and Innovation Office—NKFIH K138125, SZTE SZAOK-KKA No: 2022/5S729, and the HUN-REN Hungarian Research Network.
Acknowledgments
The graphical abstract was created using biorender.com.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AAV | adeno-associated virus |
| AD | Alzheimer’s disease |
| AhR | aryhydorcarbon receptor |
| AI | artificial intelligence |
| BBB | blood–brain barrier |
| CM | Circadian misalignment |
| COVID-19 | coronavirus disease 2019 |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| CRISPRi | clustered regularly interspaced short palindromic repeats interference |
| IDO1 | indoleamine-2,3-dioxygenase 1 |
| KMO | kynurenine 3-monooxygenase |
| KYN | kynurenine |
| KYNU | kynureninase |
| KYNA | kynurenic acid |
| KAT | kynurenine aminotransferase |
| LBPs | biotherapeutic products |
| LC-MS | liquid chromatography–mass spectrometry |
| LD | Linear dichroism |
| NAD | nicotinamide adenine dinucleotide |
| QUIN | quinolinic acid |
| TDO2 | tryptophan-2,3-dioxygenase-2 |
| TLA | Three letter acronym |
| Trp | tryptophan |
| ZIM3 | zinc finger protein 3 |
References
- Sun, M.; Ma, N.; He, T.; Johnston, L.J.; Ma, X. Tryptophan (Trp) modulates gut homeostasis via aryl hydrocarbon receptor (AhR). Critical reviews in food science and nutrition 2020, 60, 1760-1768.
- Roth, W.; Zadeh, K.; Vekariya, R.; Ge, Y.; Mohamadzadeh, M. Tryptophan metabolism and gut-brain homeostasis. International journal of molecular sciences 2021, 22, 2973.
- Su, X.; Gao, Y.; Yang, R. Gut microbiota-derived tryptophan metabolites maintain gut and systemic homeostasis. Cells 2022, 11, 2296.
- Hyland, N.P.; Cavanaugh, C.R.; Hornby, P.J. Emerging effects of tryptophan pathway metabolites and intestinal microbiota on metabolism and intestinal function. Amino Acids 2022, 54, 57-70.
- Figueiredo Godoy, A.C.; Frota, F.F.; Araújo, L.P.; Valenti, V.E.; Pereira, E.; Detregiachi, C.R.P.; Galhardi, C.M.; Caracio, F.C.; Haber, R.S.A.; Fornari Laurindo, L.; et al. Neuroinflammation and Natural Antidepressants: Balancing Fire with Flora. Biomedicines 2025, 13. [CrossRef]
- Chen, X.; Xu, D.; Yu, J.; Song, X.-J.; Li, X.; Cui, Y.-L. Tryptophan metabolism disorder-triggered diseases, mechanisms, and therapeutic strategies: a scientometric review. Nutrients 2024, 16, 3380.
- Vécsei, L.; Szalárdy, L.; Fülöp, F.; Toldi, J. Kynurenines in the CNS: recent advances and new questions. Nat Rev Drug Discov 2013, 12, 64-82. [CrossRef]
- Tanaka, M.; Vécsei, L. A Decade of Dedication: Pioneering Perspectives on Neurological Diseases and Mental Illnesses. Biomedicines 2024, 12. [CrossRef]
- Tanaka, M.; Battaglia, S.; Giménez-Llort, L.; Chen, C.; Hepsomali, P.; Avenanti, A.; Vécsei, L. Innovation at the Intersection: Emerging Translational Research in Neurology and Psychiatry. Cells 2024, 13. [CrossRef]
- Tanaka, M.; Battaglia, S.; Liloia, D. Navigating Neurodegeneration: Integrating Biomarkers, Neuroinflammation, and Imaging in Parkinson's, Alzheimer's, and Motor Neuron Disorders. Biomedicines 2025, 13. [CrossRef]
- de Lima, E.P.; Laurindo, L.F.; Catharin, V.C.S.; Direito, R.; Tanaka, M.; Jasmin Santos German, I.; Lamas, C.B.; Guiguer, E.L.; Araújo, A.C.; Fiorini, A.M.R.; et al. Polyphenols, Alkaloids, and Terpenoids Against Neurodegeneration: Evaluating the Neuroprotective Effects of Phytocompounds Through a Comprehensive Review of the Current Evidence. Metabolites 2025, 15. [CrossRef]
- Barbalho, S.M.; Leme Boaro, B.; da Silva Camarinha Oliveira, J.; Patočka, J.; Barbalho Lamas, C.; Tanaka, M.; Laurindo, L.F. Molecular Mechanisms Underlying Neuroinflammation Intervention with Medicinal Plants: A Critical and Narrative Review of the Current Literature. Pharmaceuticals (Basel) 2025, 18. [CrossRef]
- Pagotto, G.L.O.; Santos, L.; Osman, N.; Lamas, C.B.; Laurindo, L.F.; Pomini, K.T.; Guissoni, L.M.; Lima, E.P.; Goulart, R.A.; Catharin, V.; et al. Ginkgo biloba: A Leaf of Hope in the Fight against Alzheimer's Dementia: Clinical Trial Systematic Review. Antioxidants (Basel) 2024, 13. [CrossRef]
- Valotto Neto, L.J.; Reverete de Araujo, M.; Moretti Junior, R.C.; Mendes Machado, N.; Joshi, R.K.; Dos Santos Buglio, D.; Barbalho Lamas, C.; Direito, R.; Fornari Laurindo, L.; Tanaka, M.; et al. Investigating the Neuroprotective and Cognitive-Enhancing Effects of Bacopa monnieri: A Systematic Review Focused on Inflammation, Oxidative Stress, Mitochondrial Dysfunction, and Apoptosis. Antioxidants (Basel) 2024, 13. [CrossRef]
- Xue, C.; Li, G.; Zheng, Q.; Gu, X.; Shi, Q.; Su, Y.; Chu, Q.; Yuan, X.; Bao, Z.; Lu, J. Tryptophan metabolism in health and disease. Cell Metabolism 2023, 35, 1304-1326.
- Miyamoto, K.; Sujino, T.; Kanai, T. The tryptophan metabolic pathway of the microbiome and host cells in health and disease. International immunology 2024, 36, 601-616.
- Cellini, B.; Zelante, T.; Dindo, M.; Bellet, M.M.; Renga, G.; Romani, L.; Costantini, C. Pyridoxal 5′-phosphate-dependent enzymes at the crossroads of host–microbe tryptophan metabolism. International Journal of Molecular Sciences 2020, 21, 5823.
- Tanaka, M.; Szabó, Á.; Vécsei, L. Redefining Roles: A Paradigm Shift in Tryptophan-Kynurenine Metabolism for Innovative Clinical Applications. Int J Mol Sci 2024, 25. [CrossRef]
- Vandereyken, K.; Sifrim, A.; Thienpont, B.; Voet, T. Methods and applications for single-cell and spatial multi-omics. Nature Reviews Genetics 2023, 24, 494-515.
- Bressan, D.; Battistoni, G.; Hannon, G.J. The dawn of spatial omics. Science 2023, 381, eabq4964.
- Hrovatin, K.; Fischer, D.S.; Theis, F.J. Toward modeling metabolic state from single-cell transcriptomics. Molecular metabolism 2022, 57, 101396.
- Lok, R.; Qian, J.; Chellappa, S.L. Sex differences in sleep, circadian rhythms, and metabolism: implications for precision medicine. Sleep medicine reviews 2024, 101926.
- Lévi, F.A.; Okyar, A.; Hadadi, E.; Innominato, P.F.; Ballesta, A. Circadian regulation of drug responses: toward sex-specific and personalized chronotherapy. Annual review of pharmacology and toxicology 2024, 64, 89-114.
- Weger, M.; Weger, B.D.; Gachon, F. Understanding circadian dynamics: current progress and future directions for chronobiology in drug discovery. Expert Opinion on Drug Discovery 2023, 18, 893-901.
- Liang, Y.; Truong, T.; Saxton, A.J.; Boekweg, H.; Payne, S.H.; Van Ry, P.M.; Kelly, R.T. HyperSCP: combining isotopic and isobaric labeling for higher throughput single-cell proteomics. Analytical chemistry 2023, 95, 8020-8027.
- Fernández-García, J.; Altea-Manzano, P.; Pranzini, E.; Fendt, S.-M. Stable isotopes for tracing mammalian-cell metabolism in vivo. Trends in biochemical sciences 2020, 45, 185-201.
- Rottinghaus, A.G.; Ferreiro, A.; Fishbein, S.R.; Dantas, G.; Moon, T.S. Genetically stable CRISPR-based kill switches for engineered microbes. Nature communications 2022, 13, 672.
- Zhang, Y.; Hu, Y.; Jiang, N.; Yetisen, A.K. Wearable artificial intelligence biosensor networks. Biosensors and Bioelectronics 2023, 219, 114825.
- Qiu, Y.; Li, M. A Bayesian Dynamic Model-Based Adaptive Design for Oncology Dose Optimization in Phase I/II Clinical Trials. Pharm Stat 2025, 24, e2451. [CrossRef]
- Joisten, N.; Kummerhoff, F.; Koliamitra, C.; Schenk, A.; Walzik, D.; Hardt, L.; Knoop, A.; Thevis, M.; Kiesl, D.; Metcalfe, A.J.; et al. Exercise and the Kynurenine pathway: Current state of knowledge and results from a randomized cross-over study comparing acute effects of endurance and resistance training. Exerc Immunol Rev 2020, 26, 24-42.
- Jankovskaja, S.; Engblom, J.; Rezeli, M.; Marko-Varga, G.; Ruzgas, T.; Björklund, S. Non-invasive skin sampling of tryptophan/kynurenine ratio in vitro towards a skin cancer biomarker. Sci Rep 2021, 11, 678. [CrossRef]
- Cordaillat-Simmons, M.; Rouanet, A.; Pot, B. Live biotherapeutic products: the importance of a defined regulatory framework. Exp Mol Med 2020, 52, 1397-1406. [CrossRef]
- Cerqueira, F.P.; Jesus, A.M.C.; Cotrim, M.D. Adaptive Design: A Review of the Technical, Statistical, and Regulatory Aspects of Implementation in a Clinical Trial. Ther Innov Regul Sci 2020, 54, 246-258. [CrossRef]
- Hou, Y.; Li, J.; Ying, S. Tryptophan Metabolism and Gut Microbiota: A Novel Regulatory Axis Integrating the Microbiome, Immunity, and Cancer. Metabolites 2023, 13. [CrossRef]
- Su, X.; Gao, Y.; Yang, R. Gut Microbiota-Derived Tryptophan Metabolites Maintain Gut and Systemic Homeostasis. Cells 2022, 11. [CrossRef]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front Cell Infect Microbiol 2018, 8, 13. [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716-724. [CrossRef]
- Mostafavi Abdolmaleky, H.; Zhou, J.R. Gut Microbiota Dysbiosis, Oxidative Stress, Inflammation, and Epigenetic Alterations in Metabolic Diseases. Antioxidants (Basel) 2024, 13. [CrossRef]
- Peña-Durán, E.; García-Galindo, J.J.; López-Murillo, L.D.; Huerta-Huerta, A.; Balleza-Alejandri, L.R.; Beltrán-Ramírez, A.; Anaya-Ambriz, E.J.; Suárez-Rico, D.O. Microbiota and Inflammatory Markers: A Review of Their Interplay, Clinical Implications, and Metabolic Disorders. Int J Mol Sci 2025, 26. [CrossRef]
- Hand, T.W.; Vujkovic-Cvijin, I.; Ridaura, V.K.; Belkaid, Y. Linking the Microbiota, Chronic Disease, and the Immune System. Trends Endocrinol Metab 2016, 27, 831-843. [CrossRef]
- Potrykus, M.; Czaja-Stolc, S.; Stankiewicz, M.; Kaska, Ł.; Małgorzewicz, S. Intestinal Microbiota as a Contributor to Chronic Inflammation and Its Potential Modifications. Nutrients 2021, 13. [CrossRef]
- Rizzetto, L.; Fava, F.; Tuohy, K.M.; Selmi, C. Connecting the immune system, systemic chronic inflammation and the gut microbiome: The role of sex. J Autoimmun 2018, 92, 12-34. [CrossRef]
- Wang, Q.; Liu, D.; Song, P.; Zou, M.H. Tryptophan-kynurenine pathway is dysregulated in inflammation, and immune activation. Front Biosci (Landmark Ed) 2015, 20, 1116-1143. [CrossRef]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc Natl Acad Sci U S A 2020, 117, 19376-19387. [CrossRef]
- Gasaly, N.; de Vos, P.; Hermoso, M.A. Impact of Bacterial Metabolites on Gut Barrier Function and Host Immunity: A Focus on Bacterial Metabolism and Its Relevance for Intestinal Inflammation. Front Immunol 2021, 12, 658354. [CrossRef]
- Mayengbam, S.; Chleilat, F.; Reimer, R.A. Dietary Vitamin B6 Deficiency Impairs Gut Microbiota and Host and Microbial Metabolites in Rats. Biomedicines 2020, 8. [CrossRef]
- Hou, C.; Shi, H.; Xiao, J.; Song, X.; Luo, Z.; Ma, X.; Shi, L.; Wei, H.; Li, J. Pomegranate Juice Supplemented with Inulin Modulates Gut Microbiota and Promotes the Production of Microbiota-Associated Metabolites in Overweight/Obese Individuals: A Randomized, Double-Blind, Placebo-Controlled Trial. J Agric Food Chem 2024, 72, 14663-14677. [CrossRef]
- Hiel, S.; Gianfrancesco, M.A.; Rodriguez, J.; Portheault, D.; Leyrolle, Q.; Bindels, L.B.; Gomes da Silveira Cauduro, C.; Mulders, M.; Zamariola, G.; Azzi, A.S.; et al. Link between gut microbiota and health outcomes in inulin -treated obese patients: Lessons from the Food4Gut multicenter randomized placebo-controlled trial. Clin Nutr 2020, 39, 3618-3628. [CrossRef]
- Zhang, J.; Zhu, S.; Ma, N.; Johnston, L.J.; Wu, C.; Ma, X. Metabolites of microbiota response to tryptophan and intestinal mucosal immunity: A therapeutic target to control intestinal inflammation. Med Res Rev 2021, 41, 1061-1088. [CrossRef]
- Gupta, S.K.; Vyavahare, S.; Duchesne Blanes, I.L.; Berger, F.; Isales, C.; Fulzele, S. Microbiota-derived tryptophan metabolism: Impacts on health, aging, and disease. Exp Gerontol 2023, 183, 112319. [CrossRef]
- Meghani, S.; Frishkopf, M.; Park, T.; Montgomery, C.L.; Norris, C.; Papathanassoglou, E. Music-based interventions and theoretical mechanisms in post-ICU survivors: A critical narrative synthesis. Intensive Crit Care Nurs 2025, 86, 103777. [CrossRef]
- Dimitriadis, T.; Della Porta, D.; Perschl, J.; Evers, A.W.M.; Magee, W.L.; Schaefer, R.S. Motivation and music interventions in adults: A systematic review. Neuropsychol Rehabil 2024, 34, 649-678. [CrossRef]
- Kuuse, A.K.; Paulander, A.S.; Eulau, L. Characteristics and impacts of live music interventions on health and wellbeing for children, families, and health care professionals in paediatric hospitals: a scoping review. Int J Qual Stud Health Well-being 2023, 18, 2180859. [CrossRef]
- Pakdeesatitwara, N.; Clark, I.; Tamplin, J. A mixed-studies systematic review of self-administered music interventions (SAMIs) for psychological wellbeing in people with chronic health conditions: Meta-analysis and narrative summary. Patient Educ Couns 2024, 118, 108006. [CrossRef]
- Chang, E.X.; Brooker, J.; Hiscock, R.; O'Callaghan, C. Music-based intervention impacts for people with eating disorders: A narrative synthesis systematic review. J Music Ther 2023, 60, 202-231. [CrossRef]
- Aslamkhan, A.G.; Xu, Q.; Loughlin, A.; Vu, H.; Pacchione, S.; Bhatt, B.; Garfinkel, I.; Styring, T.G.; LaFranco-Scheuch, L.; Pearson, K.; et al. Characterization of indoleamine-2,3-dioxygenase 1, tryptophan-2,3-dioxygenase, and Ido1/Tdo2 knockout mice. Toxicol Appl Pharmacol 2020, 406, 115216. [CrossRef]
- Tijono, S.M.; Palmer, B.D.; Tomek, P.; Flanagan, J.U.; Henare, K.; Gamage, S.; Braun, L.; Ching, L.M. Evaluation of Novel Inhibitors of Tryptophan Dioxygenases for Enzyme and Species Selectivity Using Engineered Tumour Cell Lines Expressing Either Murine or Human IDO1 or TDO2. Pharmaceuticals (Basel) 2022, 15. [CrossRef]
- Naing, A.; Eder, J.P.; Piha-Paul, S.A.; Gimmi, C.; Hussey, E.; Zhang, S.; Hildebrand, V.; Hosagrahara, V.; Habermehl, C.; Moisan, J.; et al. Preclinical investigations and a first-in-human phase I trial of M4112, the first dual inhibitor of indoleamine 2,3-dioxygenase 1 and tryptophan 2,3-dioxygenase 2, in patients with advanced solid tumors. J Immunother Cancer 2020, 8. [CrossRef]
- Juhász, L.; Spisák, K.; Szolnoki, B.Z.; Nászai, A.; Szabó, Á.; Rutai, A.; Tallósy, S.P.; Szabó, A.; Toldi, J.; Tanaka, M.; et al. The Power Struggle: Kynurenine Pathway Enzyme Knockouts and Brain Mitochondrial Respiration. J Neurochem 2025, 169, e70075. [CrossRef]
- Szabó, Á.; Galla, Z.; Spekker, E.; Szűcs, M.; Martos, D.; Takeda, K.; Ozaki, K.; Inoue, H.; Yamamoto, S.; Toldi, J.; et al. Oxidative and Excitatory Neurotoxic Stresses in CRISPR/Cas9-Induced Kynurenine Aminotransferase Knockout Mice: A Novel Model for Despair-Based Depression and Post-Traumatic Stress Disorder. Front Biosci (Landmark Ed) 2025, 30, 25706. [CrossRef]
- Tanaka, M.; Szatmári, I.; Vécsei, L. Quinoline Quest: Kynurenic Acid Strategies for Next-Generation Therapeutics via Rational Drug Design. Pharmaceuticals (Basel) 2025, 18. [CrossRef]
- Capochiani de Iudicibus, R.; Tomek, P.; Palmer, B.D.; Tijono, S.M.; Flanagan, J.U.; Ching, L.M. Parallel discovery of selective and dual inhibitors of tryptophan dioxygenases IDO1 and TDO2 with a newly-modified enzymatic assay. Bioorg Med Chem 2021, 39, 116160. [CrossRef]
- MacCannell, A.D.; Roberts, L.D. Metabokines in the regulation of systemic energy metabolism. Curr Opin Pharmacol 2022, 67, 102286. [CrossRef]
- Choi, M.J.; Jung, S.B.; Chang, J.Y.; Shong, M. Cellular and Intercellular Homeostasis in Adipose Tissue with Mitochondria-Specific Stress. Endocrinol Metab (Seoul) 2021, 36, 1-11. [CrossRef]
- Whitehead, A.; Krause, F.N.; Moran, A.; MacCannell, A.D.V.; Scragg, J.L.; McNally, B.D.; Boateng, E.; Murfitt, S.A.; Virtue, S.; Wright, J.; et al. Brown and beige adipose tissue regulate systemic metabolism through a metabolite interorgan signaling axis. Nat Commun 2021, 12, 1905. [CrossRef]
- Qian, X.; Chen, L.; Sui, Y.; Chen, C.; Zhang, W.; Zhou, J.; Dong, W.; Jiang, M.; Xin, F.; Ochsenreither, K. Biotechnological potential and applications of microbial consortia. Biotechnol Adv 2020, 40, 107500. [CrossRef]
- Hampton, T. From the Literature. Circulation 2020, 142, 1491-1493.
- Benninghaus, L.; Schwardmann, L.S.; Jilg, T.; Wendisch, V.F. Establishment of synthetic microbial consortia with Corynebacterium glutamicum and Pseudomonas putida: Design, construction, and application to production of γ-glutamylisopropylamide and l-theanine. Microb Biotechnol 2024, 17, e14400. [CrossRef]
- Peng, H.; Darlington, A.P.S.; South, E.J.; Chen, H.H.; Jiang, W.; Ledesma-Amaro, R. A molecular toolkit of cross-feeding strains for engineering synthetic yeast communities. Nat Microbiol 2024, 9, 848-863. [CrossRef]
- Gasparek, M.; Steel, H.; Papachristodoulou, A. Deciphering mechanisms of production of natural compounds using inducer-producer microbial consortia. Biotechnol Adv 2023, 64, 108117. [CrossRef]
- Alnahhas, R.N.; Sadeghpour, M.; Chen, Y.; Frey, A.A.; Ott, W.; Josić, K.; Bennett, M.R. Majority sensing in synthetic microbial consortia. Nat Commun 2020, 11, 3659. [CrossRef]
- Bodapati, S.; Daley, T.P.; Lin, X.; Zou, J.; Qi, L.S. A benchmark of algorithms for the analysis of pooled CRISPR screens. Genome Biol 2020, 21, 62. [CrossRef]
- Li, Y.; Tang, J.; Jiang, J.; Chen, Z. Metabolic checkpoints and novel approaches for immunotherapy against cancer. International Journal of Cancer 2022, 150, 195-207.
- Yang, S.; Yuan, Z.; Zhu, Y.; Liang, C.; Chen, Z.; Zhang, J.; Leng, L. Multi-omics analysis reveals GAPDH posttranscriptional regulation of IFN-γ and PHGDH as a metabolic checkpoint of microglia polarization. Brain, behavior, and immunity 2024, 117, 155-166.
- Fang, J.; Lu, Y.; Zheng, J.; Jiang, X.; Shen, H.; Shang, X.; Lu, Y.; Fu, P. Exploring the crosstalk between endothelial cells, immune cells, and immune checkpoints in the tumor microenvironment: new insights and therapeutic implications. Cell death & disease 2023, 14, 586.
- Liu, M.; Hong, L.; Sridhar, S.; Jaynes, P.; Tipgomut, C.; Poon, L.; De Mel, S.; Lee, J.S.X.; Ng, S.-B.; Tan, C.L. Spatial-Resolved Transcriptomics Reveals Immune Landscape Variations in Primary Central Nervous System Lymphoma (PCNSL) and Diffuse Large B-Cell Lymphoma (DLBCL). Blood 2024, 144, 3004.
- Tanaka, M.; Vécsei, L. Revolutionizing our understanding of Parkinson's disease: Dr. Heinz Reichmann's pioneering research and future research direction. J Neural Transm (Vienna) 2024, 131, 1367-1387. [CrossRef]
- Abdel-Rahman, S.A.; Gabr, M. Small molecule immunomodulators as next-generation therapeutics for glioblastoma. Cancers 2024, 16, 435.
- de Lima, E.P.; Tanaka, M.; Lamas, C.B.; Quesada, K.; Detregiachi, C.R.P.; Araújo, A.C.; Guiguer, E.L.; Catharin, V.; de Castro, M.V.M.; Junior, E.B.; et al. Vascular Impairment, Muscle Atrophy, and Cognitive Decline: Critical Age-Related Conditions. Biomedicines 2024, 12. [CrossRef]
- Barbalho, S.M.; Laurindo, L.F.; de Oliveira Zanuso, B.; da Silva, R.M.S.; Gallerani Caglioni, L.; Nunes Junqueira de Moraes, V.B.F.; Fornari Laurindo, L.; Dogani Rodrigues, V.; da Silva Camarinha Oliveira, J.; Beluce, M.E.; et al. AdipoRon's Impact on Alzheimer's Disease-A Systematic Review and Meta-Analysis. Int J Mol Sci 2025, 26. [CrossRef]
- Kim, J.E.; Patel, K.; Jackson, C.M. The potential for immune checkpoint modulators in cerebrovascular injury and inflammation. Expert opinion on therapeutic targets 2021, 25, 101-113.
- Battaglia, S.; Fazio, C.D.; Borgomaneri, S.; Avenanti, A. Cortisol Imbalance and Fear Learning in PTSD: Therapeutic Approaches to Control Abnormal Fear Responses. Curr Neuropharmacol 2025, 23, 835-846. [CrossRef]
- Battaglia, S.; Di Fazio, C.; Mazzà, M.; Tamietto, M.; Avenanti, A. Targeting Human Glucocorticoid Receptors in Fear Learning: A Multiscale Integrated Approach to Study Functional Connectivity. Int J Mol Sci 2024, 25. [CrossRef]
- Tortora, F.; Hadipour, A.L.; Battaglia, S.; Falzone, A.; Avenanti, A.; Vicario, C.M. The Role of Serotonin in Fear Learning and Memory: A Systematic Review of Human Studies. Brain Sci 2023, 13. [CrossRef]
- Tanaka, M.; Tóth, F.; Polyák, H.; Szabó, Á.; Mándi, Y.; Vécsei, L. Immune influencers in action: metabolites and enzymes of the tryptophan-kynurenine metabolic pathway. Biomedicines 2021, 9, 734.
- Fujigaki, H.; Yamamoto, Y.; Saito, K. L-Tryptophan-kynurenine pathway enzymes are therapeutic target for neuropsychiatric diseases: Focus on cell type differences. Neuropharmacology 2017, 112, 264-274.
- Cervenka, I.; Agudelo, L.Z.; Ruas, J.L. Kynurenines: Tryptophan’s metabolites in exercise, inflammation, and mental health. Science 2017, 357, eaaf9794.
- Klaessens, S.; Stroobant, V.; De Plaen, E.; Van den Eynde, B.J. Systemic tryptophan homeostasis. Frontiers in molecular biosciences 2022, 9, 897929.
- Cheong, J.E.; Sun, L. Targeting the IDO1/TDO2–KYN–AhR pathway for cancer immunotherapy–challenges and opportunities. Trends in pharmacological sciences 2018, 39, 307-325.
- Stone, T.W.; Williams, R.O. Tryptophan metabolism as a ‘reflex’feature of neuroimmune communication: sensor and effector functions for the indoleamine-2, 3-dioxygenase kynurenine pathway. Journal of Neurochemistry 2024, 168, 3333-3357.
- Labadie, B.W.; Bao, R.; Luke, J.J. Reimagining IDO pathway inhibition in cancer immunotherapy via downstream focus on the tryptophan–kynurenine–aryl hydrocarbon axis. Clinical Cancer Research 2019, 25, 1462-1471.
- Tanaka, M.; Vécsei, L. From Lab to Life: Exploring Cutting-Edge Models for Neurological and Psychiatric Disorders. Biomedicines 2024, 12. [CrossRef]
- Platten, M.; von Knebel Doeberitz, N.; Oezen, I.; Wick, W.; Ochs, K. Cancer immunotherapy by targeting IDO1/TDO and their downstream effectors. Frontiers in immunology 2015, 5, 673.
- Triplett, T.A.; Garrison, K.C.; Marshall, N.; Donkor, M.; Blazeck, J.; Lamb, C.; Qerqez, A.; Dekker, J.D.; Tanno, Y.; Lu, W.-C. Reversal of indoleamine 2, 3-dioxygenase–mediated cancer immune suppression by systemic kynurenine depletion with a therapeutic enzyme. Nature biotechnology 2018, 36, 758-764.
- Stone, T.W.; Williams, R.O. Modulation of T cells by tryptophan metabolites in the kynurenine pathway. Trends in Pharmacological Sciences 2023, 44, 442-456.
- Williams, H.L.; Frei, A.L.; Koessler, T.; Berger, M.D.; Dawson, H.; Michielin, O.; Zlobec, I. The current landscape of spatial biomarkers for prediction of response to immune checkpoint inhibition. NPJ precision oncology 2024, 8, 178.
- Ala, M. The footprint of kynurenine pathway in every cancer: a new target for chemotherapy. European Journal of Pharmacology 2021, 896, 173921.
- Liu, M.; Wang, X.; Wang, L.; Ma, X.; Gong, Z.; Zhang, S.; Li, Y. Targeting the IDO1 pathway in cancer: from bench to bedside. J Hematol Oncol 2018, 11, 100. [CrossRef]
- Battaglia, S.; Nazzi, C.; Di Fazio, C.; Borgomaneri, S. The role of pre-supplementary motor cortex in action control with emotional stimuli: A repetitive transcranial magnetic stimulation study. Ann N Y Acad Sci 2024, 1536, 151-166. [CrossRef]
- Battaglia, S.; Nazzi, C.; Fullana, M.A.; di Pellegrino, G.; Borgomaneri, S. 'Nip it in the bud': Low-frequency rTMS of the prefrontal cortex disrupts threat memory consolidation in humans. Behav Res Ther 2024, 178, 104548. [CrossRef]
- Krolak, T.; Chan, K.Y.; Kaplan, L.; Huang, Q.; Wu, J.; Zheng, Q.; Kozareva, V.; Beddow, T.; Tobey, I.G.; Pacouret, S.; et al. A High-Efficiency AAV for Endothelial Cell Transduction Throughout the Central Nervous System. Nat Cardiovasc Res 2022, 1, 389-400. [CrossRef]
- Chen, X.; Wolfe, D.A.; Bindu, D.S.; Zhang, M.; Taskin, N.; Goertsen, D.; Shay, T.F.; Sullivan, E.E.; Huang, S.F.; Ravindra Kumar, S.; et al. Functional gene delivery to and across brain vasculature of systemic AAVs with endothelial-specific tropism in rodents and broad tropism in primates. Nat Commun 2023, 14, 3345. [CrossRef]
- Bolanos-Palmieri, P.; Kotb, A.; Schenk, H.; Bähre, H.; Schroder, P.; Schiffer, M. MO006 CHANGES IN THE KYNURENINE PATHWAY LEAD TO ALTERATIONS IN NAD BALANCE AND BIOENERGETICS PARAMETERS IN GLOMERULAR CELLS IN VITRO AND CONTRIBUTE TO PROTEINURIA IN A ZEBRAFISH MODEL. Nephrology Dialysis Transplantation 2021, 36, gfab079. 002.
- Kampmann, M. CRISPRi and CRISPRa Screens in Mammalian Cells for Precision Biology and Medicine. ACS Chem Biol 2018, 13, 406-416. [CrossRef]
- Battaglia, S.; Avenanti, A.; Vécsei, L.; Tanaka, M. Neurodegeneration in Cognitive Impairment and Mood Disorders for Experimental, Clinical and Translational Neuropsychiatry. Biomedicines 2024, 12. [CrossRef]
- Späth, M.R.; Hoyer-Allo, K.J.R.; Seufert, L.; Höhne, M.; Lucas, C.; Bock, T.; Isermann, L.; Brodesser, S.; Lackmann, J.W.; Kiefer, K.; et al. Organ Protection by Caloric Restriction Depends on Activation of the De Novo NAD+ Synthesis Pathway. J Am Soc Nephrol 2023, 34, 772-792. [CrossRef]
- Flickinger, K.M.; Cantor, J.R. Uncovering the Conditionally Essential Roles of NAD Kinases in Human Cells. The FASEB Journal 2022, 36.
- Walsh, S.; Gardner, L.; Deiters, A.; Williams, G.J. Intracellular light-activation of riboswitch activity. Chembiochem 2014, 15, 1346-1351. [CrossRef]
- Borrachero-Conejo, A.I.; Adams, W.R.; Saracino, E.; Mola, M.G.; Wang, M.; Posati, T.; Formaggio, F.; De Bellis, M.; Frigeri, A.; Caprini, M.; et al. Stimulation of water and calcium dynamics in astrocytes with pulsed infrared light. Faseb j 2020, 34, 6539-6553. [CrossRef]
- Spennato, D.; Leone, J.; Gundhardt, C.; Varnavski, O.; Fabbri, R.; Caprini, M.; Zamboni, R.; Benfenati, V.; Goodson, T., 3rd. Investigations of Astrocyte Calcium Signaling and Imaging with Classical and Nonclassical Light. J Phys Chem B 2024, 128, 7966-7977. [CrossRef]
- Qiao, L.; Niu, L.; Wang, M.; Wang, Z.; Kong, D.; Yu, G.; Ye, H. A sensitive red/far-red photoswitch for controllable gene therapy in mouse models of metabolic diseases. Nature Communications 2024, 15, 10310.
- Walton, J.C.; Bumgarner, J.R.; Nelson, R.J. Sex Differences in Circadian Rhythms. Cold Spring Harb Perspect Biol 2022, 14. [CrossRef]
- Tanaka, M.; Tuka, B.; Vécsei, L. Navigating the Neurobiology of Migraine: From Pathways to Potential Therapies. Cells 2024, 13. [CrossRef]
- Minbay, M.; Khan, A.; Ghasemi, A.R.; Ingram, K.K.; Ay, A.A. Sex-specific associations between circadian-related genes and depression in UK Biobank participants highlight links to glucose metabolism, inflammation and neuroplasticity pathways. Psychiatry Res 2024, 337, 115948. [CrossRef]
- Abo, S.M.; Layton, A.T. Modeling the circadian regulation of the immune system: Sexually dimorphic effects of shift work. PLoS computational biology 2021, 17, e1008514.
- Logan, R.W.; Xue, X.; Ketchesin, K.D.; Hoffman, G.; Roussos, P.; Tseng, G.; McClung, C.A.; Seney, M.L. Sex Differences in Molecular Rhythms in the Human Cortex. Biol Psychiatry 2022, 91, 152-162. [CrossRef]
- Liloia, D.; Zamfira, D.A.; Tanaka, M.; Manuello, J.; Crocetta, A.; Keller, R.; Cozzolino, M.; Duca, S.; Cauda, F.; Costa, T. Disentangling the role of gray matter volume and concentration in autism spectrum disorder: A meta-analytic investigation of 25 years of voxel-based morphometry research. Neurosci Biobehav Rev 2024, 164, 105791. [CrossRef]
- Bailey, M.; Silver, R. Sex differences in circadian timing systems: implications for disease. Front Neuroendocrinol 2014, 35, 111-139. [CrossRef]
- Tanaka, M. Beyond the boundaries: Transitioning from categorical to dimensional paradigms in mental health diagnostics. Adv Clin Exp Med 2024, 33, 1295-1301. [CrossRef]
- Wu, F.; Langer, P.; Shim, J.; Fleisch, E.; Barata, F. Comparative Efficacy of Commercial Wearables for Circadian Rhythm Home Monitoring From Activity, Heart Rate, and Core Body Temperature. IEEE J Biomed Health Inform 2025, 29, 900-908. [CrossRef]
- Zisapel, N. New perspectives on the role of melatonin in human sleep, circadian rhythms and their regulation. Br J Pharmacol 2018, 175, 3190-3199. [CrossRef]
- Lightman, S.L.; Conway-Campbell, B.L. Circadian and ultradian rhythms: Clinical implications. J Intern Med 2024, 296, 121-138. [CrossRef]
- Steele, T.A.; St Louis, E.K.; Videnovic, A.; Auger, R.R. Circadian Rhythm Sleep-Wake Disorders: a Contemporary Review of Neurobiology, Treatment, and Dysregulation in Neurodegenerative Disease. Neurotherapeutics 2021, 18, 53-74. [CrossRef]
- Tordjman, S.; Chokron, S.; Delorme, R.; Charrier, A.; Bellissant, E.; Jaafari, N.; Fougerou, C. Melatonin: Pharmacology, Functions and Therapeutic Benefits. Curr Neuropharmacol 2017, 15, 434-443. [CrossRef]
- Lévi, F.A.; Okyar, A.; Hadadi, E.; Innominato, P.F.; Ballesta, A. Circadian Regulation of Drug Responses: Toward Sex-Specific and Personalized Chronotherapy. Annu Rev Pharmacol Toxicol 2024, 64, 89-114. [CrossRef]
- Karaboué, A.; Innominato, P.F.; Wreglesworth, N.I.; Duchemann, B.; Adam, R.; Lévi, F.A. Why does circadian timing of administration matter for immune checkpoint inhibitors' efficacy? Br J Cancer 2024, 131, 783-796. [CrossRef]
- Ohdo, S.; Koyanagi, S.; Matsunaga, N. Chronopharmacological strategies focused on chrono-drug discovery. Pharmacol Ther 2019, 202, 72-90. [CrossRef]
- Wang, C.; Zeng, Q.; Gül, Z.M.; Wang, S.; Pick, R.; Cheng, P.; Bill, R.; Wu, Y.; Naulaerts, S.; Barnoud, C.; et al. Circadian tumor infiltration and function of CD8(+) T cells dictate immunotherapy efficacy. Cell 2024, 187, 2690-2702.e2617. [CrossRef]
- Ye, Y.; Xiang, Y.; Ozguc, F.M.; Kim, Y.; Liu, C.J.; Park, P.K.; Hu, Q.; Diao, L.; Lou, Y.; Lin, C.; et al. The Genomic Landscape and Pharmacogenomic Interactions of Clock Genes in Cancer Chronotherapy. Cell Syst 2018, 6, 314-328.e312. [CrossRef]
- Ephraim, A.; Leatheng, C.; Lu, Z.E.; Xia, X.; Pirruccello, J.P.; Marotti, J.D.; MacKenzie, T.; Chamberlin, M.D. Association of plasma kynurenine (KYN) with plasma osteopontin (OPN) in patients with locally invasive breast cancer. 2024.
- Chang, K.H.; Cheng, M.L.; Tang, H.Y.; Huang, C.Y.; Wu, Y.R.; Chen, C.M. Alternations of Metabolic Profile and Kynurenine Metabolism in the Plasma of Parkinson's Disease. Mol Neurobiol 2018, 55, 6319-6328. [CrossRef]
- Chantrapanichkul, P.; Stevenson, M.O.; Suppakitjanusant, P.; Goodman, M.; Tangpricha, V. SERUM HORMONE CONCENTRATIONS IN TRANSGENDER INDIVIDUALS RECEIVING GENDER-AFFIRMING HORMONE THERAPY: A LONGITUDINAL RETROSPECTIVE COHORT STUDY. Endocr Pract 2021, 27, 27-33. [CrossRef]
- Kervezee, L.; Cermakian, N.; Boivin, D.B. Individual metabolomic signatures of circadian misalignment during simulated night shifts in humans. PLoS Biol 2019, 17, e3000303. [CrossRef]
- Skene, D.J.; Skornyakov, E.; Chowdhury, N.R.; Gajula, R.P.; Middleton, B.; Satterfield, B.C.; Porter, K.I.; Van Dongen, H.P.A.; Gaddameedhi, S. Separation of circadian- and behavior-driven metabolite rhythms in humans provides a window on peripheral oscillators and metabolism. Proc Natl Acad Sci U S A 2018, 115, 7825-7830. [CrossRef]
- Whittaker, D.S.; Akhmetova, L.; Carlin, D.; Romero, H.; Welsh, D.K.; Colwell, C.S.; Desplats, P. Circadian modulation by time-restricted feeding rescues brain pathology and improves memory in mouse models of Alzheimer's disease. Cell Metab 2023, 35, 1704-1721.e1706. [CrossRef]
- Emens, J.S.; Burgess, H.J. Effect of Light and Melatonin and Other Melatonin Receptor Agonists on Human Circadian Physiology. Sleep Med Clin 2015, 10, 435-453. [CrossRef]
- Chen, X.; He, R.; Chen, X.; Jiang, L.; Wang, F. Optimizing dose-schedule regimens with bayesian adaptive designs: opportunities and challenges. Front Pharmacol 2023, 14, 1261312. [CrossRef]
- Lu, M.; Yuan, Y.; Liu, S. A Bayesian pharmacokinetics integrated phase I-II design to optimize dose-schedule regimes. Biostatistics 2024, 26. [CrossRef]
- Ballesta, A.; Innominato, P.F.; Dallmann, R.; Rand, D.A.; Lévi, F.A. Systems Chronotherapeutics. Pharmacol Rev 2017, 69, 161-199. [CrossRef]
- Jiang, L.; Li, R.; Yan, F.; Yap, T.A.; Yuan, Y. Shotgun: A Bayesian seamless phase I-II design to accelerate the development of targeted therapies and immunotherapy. Contemp Clin Trials 2021, 104, 106338. [CrossRef]
- Bai, X.; Huang, Z.; Duraj-Thatte, A.M.; Ebert, M.P.; Zhang, F.; Burgermeister, E.; Liu, X.; Scott, B.M.; Li, G.; Zuo, T. Engineering the gut microbiome. Nature Reviews Bioengineering 2023, 1, 665-679.
- Sorbara, M.T.; Pamer, E.G. Microbiome-based therapeutics. Nat Rev Microbiol 2022, 20, 365-380. [CrossRef]
- Mousa, W.K.; Al Ali, A. The Gut Microbiome Advances Precision Medicine and Diagnostics for Inflammatory Bowel Diseases. Int J Mol Sci 2024, 25. [CrossRef]
- Dodd, D.; Spitzer, M.H.; Van Treuren, W.; Merrill, B.D.; Hryckowian, A.J.; Higginbottom, S.K.; Le, A.; Cowan, T.M.; Nolan, G.P.; Fischbach, M.A.; et al. A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature 2017, 551, 648-652. [CrossRef]
- Han, S.; Van Treuren, W.; Fischer, C.R.; Merrill, B.D.; DeFelice, B.C.; Sanchez, J.M.; Higginbottom, S.K.; Guthrie, L.; Fall, L.A.; Dodd, D.; et al. A metabolomics pipeline for the mechanistic interrogation of the gut microbiome. Nature 2021, 595, 415-420. [CrossRef]
- Nunes, Y.C.; Mendes, N.M.; Pereira de Lima, E.; Chehadi, A.C.; Lamas, C.B.; Haber, J.F.S.; Dos Santos Bueno, M.; Araújo, A.C.; Catharin, V.C.S.; Detregiachi, C.R.P.; et al. Curcumin: A Golden Approach to Healthy Aging: A Systematic Review of the Evidence. Nutrients 2024, 16. [CrossRef]
- Lamichhane, S.; Sen, P.; Dickens, A.M.; Orešič, M.; Bertram, H.C. Gut metabolome meets microbiome: A methodological perspective to understand the relationship between host and microbe. Methods 2018, 149, 3-12. [CrossRef]
- Guo, H.; Liu, X.; Chen, T.; Wang, X.; Zhang, X. Akkermansia muciniphila Improves Depressive-Like Symptoms by Modulating the Level of 5-HT Neurotransmitters in the Gut and Brain of Mice. Mol Neurobiol 2024, 61, 821-834. [CrossRef]
- Ding, Y.; Bu, F.; Chen, T.; Shi, G.; Yuan, X.; Feng, Z.; Duan, Z.; Wang, R.; Zhang, S.; Wang, Q.; et al. A next-generation probiotic: Akkermansia muciniphila ameliorates chronic stress-induced depressive-like behavior in mice by regulating gut microbiota and metabolites. Appl Microbiol Biotechnol 2021, 105, 8411-8426. [CrossRef]
- Liu, J.Y.; Lin, T.L.; Chiu, C.Y.; Hsieh, P.F.; Lin, Y.T.; Lai, L.Y.; Wang, J.T. Decolonization of carbapenem-resistant Klebsiella pneumoniae from the intestinal microbiota of model mice by phages targeting two surface structures. Front Microbiol 2022, 13, 877074. [CrossRef]
- Medlock, G.; Felix, C.; Alsharif, W.; Cornacchione, L.; Schinn, M.; Watson, A.; Bedard-Shurtleff, S.; Norman, J.; Faith, J.; Kuijper, E.J. 2521. VE707, a defined live biotherapeutic product for prevention of infection by multidrug-resistant gram-negative bacteria. In Proceedings of the Open Forum Infectious Diseases, 2023; p. ofad500. 2139.
- Tavoukjian, V. Faecal microbiota transplantation for the decolonization of antibiotic-resistant bacteria in the gut: a systematic review and meta-analysis. J Hosp Infect 2019, 102, 174-188. [CrossRef]
- Macareño-Castro, J.; Solano-Salazar, A.; Dong, L.T.; Mohiuddin, M.; Espinoza, J.L. Fecal microbiota transplantation for Carbapenem-Resistant Enterobacteriaceae: A systematic review. J Infect 2022, 84, 749-759. [CrossRef]
- Mortzfeld, B.M.; Palmer, J.D.; Bhattarai, S.K.; Dupre, H.L.; Mercado-Lubio, R.; Silby, M.W.; Bang, C.; McCormick, B.A.; Bucci, V. Microcin MccI47 selectively inhibits enteric bacteria and reduces carbapenem-resistant Klebsiella pneumoniae colonization in vivo when administered via an engineered live biotherapeutic. Gut Microbes 2022, 14, 2127633. [CrossRef]
- Osbelt, L.; Wende, M.; Almási, É.; Derksen, E.; Muthukumarasamy, U.; Lesker, T.R.; Galvez, E.J.C.; Pils, M.C.; Schalk, E.; Chhatwal, P.; et al. Klebsiella oxytoca causes colonization resistance against multidrug-resistant K. pneumoniae in the gut via cooperative carbohydrate competition. Cell Host Microbe 2021, 29, 1663-1679.e1667. [CrossRef]
- Agarwal, S.; Tiwari, P.; Deep, A.; Kidwai, S.; Gupta, S.; Thakur, K.G.; Singh, R. System-Wide Analysis Unravels the Differential Regulation and In Vivo Essentiality of Virulence-Associated Proteins B and C Toxin-Antitoxin Systems of Mycobacterium tuberculosis. J Infect Dis 2018, 217, 1809-1820. [CrossRef]
- Lin, M.; Kussell, E. Inferring bacterial recombination rates from large-scale sequencing datasets. Nat Methods 2019, 16, 199-204. [CrossRef]
- Martin, G.; Kolida, S.; Marchesi, J.R.; Want, E.; Sidaway, J.E.; Swann, J.R. In Vitro Modeling of Bile Acid Processing by the Human Fecal Microbiota. Front Microbiol 2018, 9, 1153. [CrossRef]
- Cheng, F.; Wu, A.; Liu, C.; Cao, X.; Wang, R.; Shu, X.; Wang, L.; Zhang, Y.; Xiang, H.; Li, M. The toxin-antitoxin RNA guards of CRISPR-Cas evolved high specificity through repeat degeneration. Nucleic Acids Res 2022, 50, 9442-9452. [CrossRef]
- Wiechert, J.; Gätgens, C.; Wirtz, A.; Frunzke, J. Inducible Expression Systems Based on Xenogeneic Silencing and Counter-Silencing and Design of a Metabolic Toggle Switch. ACS Synth Biol 2020, 9, 2023-2038. [CrossRef]
- Schwarz, S.; Gildemeister, D.; Hein, A.; Schröder, P.; Bachmann, J. Environmental fate and effects assessment of human pharmaceuticals: lessons learnt from regulatory data. Environmental Sciences Europe 2021, 33, 68.
- Richard, E.; Darracq, B.; Littner, E.; Vit, C.; Whiteway, C.; Bos, J.; Fournes, F.; Garriss, G.; Conte, V.; Lapaillerie, D.; et al. Cassette recombination dynamics within chromosomal integrons are regulated by toxin-antitoxin systems. Sci Adv 2024, 10, eadj3498. [CrossRef]
- Yu, L.; Sun, Q.; Hui, Y.; Seth, A.; Petrovsky, N.; Zhao, C.X. Microfluidic formation of core-shell alginate microparticles for protein encapsulation and controlled release. J Colloid Interface Sci 2019, 539, 497-503. [CrossRef]
- Omer, A.M.; Ahmed, M.S.; El-Subruiti, G.M.; Khalifa, R.E.; Eltaweil, A.S. pH-Sensitive Alginate/Carboxymethyl Chitosan/Aminated Chitosan Microcapsules for Efficient Encapsulation and Delivery of Diclofenac Sodium. Pharmaceutics 2021, 13. [CrossRef]
- George, M.; Abraham, T.E. Polyionic hydrocolloids for the intestinal delivery of protein drugs: alginate and chitosan--a review. J Control Release 2006, 114, 1-14. [CrossRef]
- Qu, Q.; Yang, A.; Wang, J.; Xie, M.; Zhang, X.; Huang, D.; Xiong, R.; Pei, D.; Huang, C. Responsive and biocompatible chitosan-phytate microparticles with various morphology for antibacterial activity based on gas-shearing microfluidics. J Colloid Interface Sci 2023, 649, 68-75. [CrossRef]
- Feng, R.; Wang, L.; Zhou, P.; Luo, Z.; Li, X.; Gao, L. Development of the pH responsive chitosan-alginate based microgel for encapsulation of Jughans regia L. polyphenols under simulated gastrointestinal digestion in vitro. Carbohydr Polym 2020, 250, 116917. [CrossRef]
- Liao, P.; Dai, S.; Lian, Z.; Tong, X.; Yang, S.; Chen, Y.; Qi, W.; Peng, X.; Wang, H.; Jiang, L. The Layered Encapsulation of Vitamin B(2) and β-Carotene in Multilayer Alginate/Chitosan Gel Microspheres: Improving the Bioaccessibility of Vitamin B(2) and β-Carotene. Foods 2021, 11. [CrossRef]
- Liu, X.; Liu, L.; Huang, F.; Meng, Y.; Chen, Y.; Wang, J.; Wang, S.; Luo, Y.; Li, J.; Liang, Y. pH-sensitive chitosan/sodium alginate/calcium chloride hydrogel beads for potential oral delivery of rice bran bioactive peptides. Food Chem 2025, 470, 142618. [CrossRef]
- Tian, Y.; Ran, H.; Wen, X.; Fu, G.; Zhou, X.; Liu, R.; Pan, T. Probiotics improve symptoms of patients with COVID-19 through gut-lung axis: a systematic review and meta-analysis. Front Nutr 2023, 10, 1179432. [CrossRef]
- Schaub, A.C.; Schneider, E.; Vazquez-Castellanos, J.F.; Schweinfurth, N.; Kettelhack, C.; Doll, J.P.K.; Yamanbaeva, G.; Mählmann, L.; Brand, S.; Beglinger, C.; et al. Clinical, gut microbial and neural effects of a probiotic add-on therapy in depressed patients: a randomized controlled trial. Transl Psychiatry 2022, 12, 227. [CrossRef]
- Horvath, A.; Habisch, H.; Prietl, B.; Pfeifer, V.; Balazs, I.; Kovacs, G.; Foris, V.; John, N.; Kleinschek, D.; Feldbacher, N.; et al. Alteration of the Gut-Lung Axis After Severe COVID-19 Infection and Modulation Through Probiotics: A Randomized, Controlled Pilot Study. Nutrients 2024, 16. [CrossRef]
- Xu, L.; Yang, C.S.; Liu, Y.; Zhang, X. Effective Regulation of Gut Microbiota With Probiotics and Prebiotics May Prevent or Alleviate COVID-19 Through the Gut-Lung Axis. Front Pharmacol 2022, 13, 895193. [CrossRef]
- Kazemi, A.; Noorbala, A.A.; Azam, K.; Eskandari, M.H.; Djafarian, K. Effect of probiotic and prebiotic vs placebo on psychological outcomes in patients with major depressive disorder: A randomized clinical trial. Clin Nutr 2019, 38, 522-528. [CrossRef]
- Rudzki, L.; Ostrowska, L.; Pawlak, D.; Małus, A.; Pawlak, K.; Waszkiewicz, N.; Szulc, A. Probiotic Lactobacillus Plantarum 299v decreases kynurenine concentration and improves cognitive functions in patients with major depression: A double-blind, randomized, placebo controlled study. Psychoneuroendocrinology 2019, 100, 213-222. [CrossRef]
- Platten, M.; Nollen, E.A.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat Rev Drug Discov 2019, 18, 379-401. [CrossRef]
- Liang, H.; Li, T.; Fang, X.; Xing, Z.; Zhang, S.; Shi, L.; Li, W.; Guo, L.; Kuang, C.; Liu, H.; et al. IDO1/TDO dual inhibitor RY103 targets Kyn-AhR pathway and exhibits preclinical efficacy on pancreatic cancer. Cancer Lett 2021, 522, 32-43. [CrossRef]
- Agudelo, L.Z.; Ferreira, D.M.S.; Dadvar, S.; Cervenka, I.; Ketscher, L.; Izadi, M.; Zhengye, L.; Furrer, R.; Handschin, C.; Venckunas, T.; et al. Skeletal muscle PGC-1α1 reroutes kynurenine metabolism to increase energy efficiency and fatigue-resistance. Nat Commun 2019, 10, 2767. [CrossRef]
- Martos, D.; Lőrinczi, B.; Szatmári, I.; Vécsei, L.; Tanaka, M. The Impact of C-3 Side Chain Modifications on Kynurenic Acid: A Behavioral Analysis of Its Analogs in the Motor Domain. Int J Mol Sci 2024, 25. [CrossRef]
- Fan, Q.Z.; Zhou, J.; Zhu, Y.B.; He, L.J.; Miao, D.D.; Zhang, S.P.; Liu, X.P.; Zhang, C. Design, synthesis, and biological evaluation of a novel indoleamine 2,3-dioxigenase 1 (IDO1) and thioredoxin reductase (TrxR) dual inhibitor. Bioorg Chem 2020, 105, 104401. [CrossRef]
- Di Gregorio, F.; Steinhauser, M.; Maier, M.E.; Thayer, J.F.; Battaglia, S. Error-related cardiac deceleration: Functional interplay between error-related brain activity and autonomic nervous system in performance monitoring. Neurosci Biobehav Rev 2024, 157, 105542. [CrossRef]
- Nazzi, C.; Avenanti, A.; Battaglia, S. The Involvement of Antioxidants in Cognitive Decline and Neurodegeneration: Mens Sana in Corpore Sano. Antioxidants (Basel) 2024, 13. [CrossRef]
- Battaglia, S.; Nazzi, C.; Lonsdorf, T.B.; Thayer, J.F. Neuropsychobiology of fear-induced bradycardia in humans: progress and pitfalls. Mol Psychiatry 2024, 29, 3826-3840. [CrossRef]
- Heyes, M.P. Metabolism and neuropathologic significance of quinolinic acid and kynurenic acid. Biochemical Society Transactions 1993, 21, 83-89.
- Damerell, V.; Klaassen-Dekker, N.; Brezina, S.; Ose, J.; Ulvik, A.; van Roekel, E.H.; Holowatyj, A.N.; Baierl, A.; Böhm, J.; Bours, M.J.L.; et al. Circulating tryptophan-kynurenine pathway metabolites are associated with all-cause mortality among patients with stage I-III colorectal cancer. Int J Cancer 2025, 156, 552-565. [CrossRef]
- Chiu, L.C.; Tang, H.Y.; Fan, C.M.; Lo, C.J.; Hu, H.C.; Kao, K.C.; Cheng, M.L. Kynurenine Pathway of Tryptophan Metabolism Is Associated with Hospital Mortality in Patients with Acute Respiratory Distress Syndrome: A Prospective Cohort Study. Antioxidants (Basel) 2022, 11. [CrossRef]
- Hoong, C.W.S.; Chua, M.W.J. SGLT2 Inhibitors as Calorie Restriction Mimetics: Insights on Longevity Pathways and Age-Related Diseases. Endocrinology 2021, 162. [CrossRef]
- Palmer, B.F.; Clegg, D.J. Euglycemic Ketoacidosis as a Complication of SGLT2 Inhibitor Therapy. Clin J Am Soc Nephrol 2021, 16, 1284-1291. [CrossRef]
- Hoeflich, K.P.; Merchant, M.; Orr, C.; Chan, J.; Den Otter, D.; Berry, L.; Kasman, I.; Koeppen, H.; Rice, K.; Yang, N.Y.; et al. Intermittent administration of MEK inhibitor GDC-0973 plus PI3K inhibitor GDC-0941 triggers robust apoptosis and tumor growth inhibition. Cancer Res 2012, 72, 210-219. [CrossRef]
- Yu, M.; Chen, J.; Xu, Z.; Yang, B.; He, Q.; Luo, P.; Yan, H.; Yang, X. Development and safety of PI3K inhibitors in cancer. Arch Toxicol 2023, 97, 635-650. [CrossRef]
- Wu, T.; Zhang, C.; Lv, R.; Qin, Q.; Liu, N.; Yin, W.; Wang, R.; Sun, Y.; Wang, X.; Sun, Y.; et al. Design, synthesis, biological evaluation and pharmacophore model analysis of novel tetrahydropyrrolo[3,4-c]pyrazol derivatives as potential TRKs inhibitors. Eur J Med Chem 2021, 223, 113627. [CrossRef]
- Yan, W.; Lakkaniga, N.R.; Carlomagno, F.; Santoro, M.; McDonald, N.Q.; Lv, F.; Gunaganti, N.; Frett, B.; Li, H.Y. Insights into Current Tropomyosin Receptor Kinase (TRK) Inhibitors: Development and Clinical Application. J Med Chem 2019, 62, 1731-1760. [CrossRef]
- Altzerinakou, M.A.; Paoletti, X. An adaptive design for the identification of the optimal dose using joint modeling of continuous repeated biomarker measurements and time-to-toxicity in phase I/II clinical trials in oncology. Stat Methods Med Res 2020, 29, 508-521. [CrossRef]
- Pinsker, J.E.; Dassau, E.; Deshpande, S.; Raghinaru, D.; Buckingham, B.A.; Kudva, Y.C.; Laffel, L.M.; Levy, C.J.; Church, M.M.; Desrochers, H.; et al. Outpatient Randomized Crossover Comparison of Zone Model Predictive Control Automated Insulin Delivery with Weekly Data Driven Adaptation Versus Sensor-Augmented Pump: Results from the International Diabetes Closed-Loop Trial 4. Diabetes Technol Ther 2022, 24, 635-642. [CrossRef]
- Iasonos, A.; O'Quigley, J. Adaptive dose-finding studies: a review of model-guided phase I clinical trials. J Clin Oncol 2014, 32, 2505-2511. [CrossRef]
- Visser, M.M.; Charleer, S.; Fieuws, S.; De Block, C.; Hilbrands, R.; Van Huffel, L.; Maes, T.; Vanhaverbeke, G.; Dirinck, E.; Myngheer, N.; et al. Comparing real-time and intermittently scanned continuous glucose monitoring in adults with type 1 diabetes (ALERTT1): a 6-month, prospective, multicentre, randomised controlled trial. Lancet 2021, 397, 2275-2283. [CrossRef]
- Perry, M.W.D.; Abdulai, R.; Mogemark, M.; Petersen, J.; Thomas, M.J.; Valastro, B.; Westin Eriksson, A. Evolution of PI3Kγ and δ Inhibitors for Inflammatory and Autoimmune Diseases. J Med Chem 2019, 62, 4783-4814. [CrossRef]
- Bornaei, M.; Khajehsharifi, H.; Shahrokhian, S.; Sheydaei, O.; Zarnegarian, A. Differential pulse voltammetric quantitation of kynurenic acid in human plasma using carbon-paste electrode modified with metal-organic frameworks. Materials Chemistry and Physics 2023, 295, 127016.
- Beatty, G.L.; O'Dwyer, P.J.; Clark, J.; Shi, J.G.; Bowman, K.J.; Scherle, P.A.; Newton, R.C.; Schaub, R.; Maleski, J.; Leopold, L.; et al. First-in-Human Phase I Study of the Oral Inhibitor of Indoleamine 2,3-Dioxygenase-1 Epacadostat (INCB024360) in Patients with Advanced Solid Malignancies. Clin Cancer Res 2017, 23, 3269-3276. [CrossRef]
- Wang, S.; Liu, H.; Wu, D.; Wang, X. Temperature and pH dual-stimuli-responsive phase-change microcapsules for multipurpose applications in smart drug delivery. J Colloid Interface Sci 2021, 583, 470-486. [CrossRef]
- Lin, R.; Zhou, Y.; Yan, F.; Li, D.; Yuan, Y. BOIN12: Bayesian Optimal Interval Phase I/II Trial Design for Utility-Based Dose Finding in Immunotherapy and Targeted Therapies. JCO Precis Oncol 2020, 4. [CrossRef]
- Liu, D.; Xin, Z.; Guo, S.; Li, S.; Cheng, J.; Jiang, H. Blood and Salivary MicroRNAs for Diagnosis of Oral Squamous Cell Carcinoma: A Systematic Review and Meta-Analysis. J Oral Maxillofac Surg 2021, 79, 1082.e1081-1082.e1013. [CrossRef]
- Zahran, F.; Ghalwash, D.; Shaker, O.; Al-Johani, K.; Scully, C. Salivary microRNAs in oral cancer. Oral Dis 2015, 21, 739-747. [CrossRef]
- Hare, S.M.; Adhikari, B.M.; Mo, C.; Chen, S.; Wijtenburg, S.A.; Seneviratne, C.; Kane-Gerard, S.; Sathyasaikumar, K.V.; Notarangelo, F.M.; Schwarcz, R.; et al. Tryptophan challenge in individuals with schizophrenia and healthy controls: acute effects on circulating kynurenine and kynurenic acid, cognition and cerebral blood flow. Neuropsychopharmacology 2023, 48, 1594-1601. [CrossRef]
- Huang, J.; Tong, J.; Zhang, P.; Zhou, Y.; Li, Y.; Tan, S.; Wang, Z.; Yang, F.; Kochunov, P.; Chiappelli, J.; et al. Elevated salivary kynurenic acid levels related to enlarged choroid plexus and severity of clinical phenotypes in treatment-resistant schizophrenia. Brain Behav Immun 2022, 106, 32-39. [CrossRef]
- Badawy, A.A. Tryptophan availability for kynurenine pathway metabolism across the life span: Control mechanisms and focus on aging, exercise, diet and nutritional supplements. Neuropharmacology 2017, 112, 248-263. [CrossRef]
- Tanaka, M. From Serendipity to Precision: Integrating AI, Multi-Omics, and Human-Specific Models for Personalized Neuropsychiatric Care. Biomedicines 2025, 13. [CrossRef]
Table 1.
Three Translational Frontiers in Tryptophan (Trp)–Kynurenine (KYN) Research. This table outlines three key challenges impeding the development of precision interventions targeting tryptophan metabolism: (1) the lack of causal frameworks linking microbial-host-enzyme interactions to downstream metabolites and clinical outcomes; (2) insufficient spatial resolution masking critical cell- and tissue-specific metabolic hotspots; and (3) the neglect of circadian and hormonal influences on metabolic flux, limiting chronotherapeutic optimization.
Table 1.
Three Translational Frontiers in Tryptophan (Trp)–Kynurenine (KYN) Research. This table outlines three key challenges impeding the development of precision interventions targeting tryptophan metabolism: (1) the lack of causal frameworks linking microbial-host-enzyme interactions to downstream metabolites and clinical outcomes; (2) insufficient spatial resolution masking critical cell- and tissue-specific metabolic hotspots; and (3) the neglect of circadian and hormonal influences on metabolic flux, limiting chronotherapeutic optimization.
| Challenge | Core Issue | Implication | ||
|---|---|---|---|---|
| 1. Causal Mapping (Conceptual Challenge) |
Despite thousands of disease associations, we lack a coherent causal framework linking microbiota, host enzymes (e.g., IDO1, TDO2), and metabolites (e.g., KYNA, QUIN) to physiological outcomes. | Limits precision design of interventions (e.g., probiotics, enzyme inhibitors, lifestyle prescriptions). | ||
| 2. Spatial Resolution (Anatomical Challenge) | Bulk assays obscure localized metabolic activity in specialized niches (e.g., astrocytes, microglia, BBB endothelial cells). | Demands a paradigm shift: from homogeneous pathways to cell-type-specific “switchboards” requiring targeted modulation. | ||
| 3. Temporal Dynamics (Chronobiological Challenge) | Trp–KYN flux oscillates with circadian rhythms and is modulated by sex hormones, but time-sensitive dosing is understudied. | Missing chronotherapeutic windows may reduce efficacy or increase toxicity of interventions. |
BBB, blood-brain barrier; IDO1, indoleamine-2,3-dioxygenase-1; KYN, kynurenine; QUIN, quinolinic acid; TDO2, tryptophan-2,3-dioxygenase-2; Trp, tryptophan.
Table 2.
Key Objectives for a Unified Roadmap to Precision Modulation of the Tryptophan (Trp)–Kynurenine (KYN) Axis. This table outlines five core objectives of the review: integrating spatial insights, sex and circadian modifiers, microbiota-based strategies, next-generation intervention platforms, and dynamic, multiscale modeling with cross-disciplinary collaboration to advance translational outcomes in Trp–KYN biology.
Table 2.
Key Objectives for a Unified Roadmap to Precision Modulation of the Tryptophan (Trp)–Kynurenine (KYN) Axis. This table outlines five core objectives of the review: integrating spatial insights, sex and circadian modifiers, microbiota-based strategies, next-generation intervention platforms, and dynamic, multiscale modeling with cross-disciplinary collaboration to advance translational outcomes in Trp–KYN biology.
| Objective | Description | |
|---|---|---|
| 1. Map Spatial “Checkpoints” | Survey localized KYN metabolism niches in brain and periphery, detailing how enzyme activity in astrocytes, microglia, BBB, and peripheral hubs interfaces with immunity and circuitry. | |
| 2. Characterize Sex & Circadian Modifiers | Examine how sex hormones and circadian rhythms influence Trp–KYN flux, identifying periods tilting toward neurotoxicity or resilience to inform time- and sex-specific interventions. | |
| 3. Develop Microbiota-Based Precision Switches | Explore designer microbial consortia and encapsulated post-biotics to steer Trp flux, addressing manufacturing, safety, and regulatory considerations for clinical translation. | |
| 4. Outline Integrated “Intervention 2.0” Platform | Propose a closed-loop therapeutic framework combining dual enzyme inhibitors, structured exercise regimens, and AI-driven biosensing for adaptive modulation of the Trp–KYN axis. | |
| 5. Build Dynamic, Multiscale Predictive Models & Networks | Weave spatial, temporal, and microbiome data into predictive tools; foster collaborations among bench scientists, clinicians, data engineers, and regulators to guide personalized interventions. |
Table 3.
Next steps in precision modulation of neurovascular kynurenine (KYN) flux. This table outlines the sequential experimental steps to refine endothelial targeting, validate functional outcomes, monitor real-time neurotransmitter dynamics, evaluate effects on cancer metastasis, and ensure systemic safety, aiming to open new avenues for precise modulation of the neurovascular KYN metabolism.
Table 3.
Next steps in precision modulation of neurovascular kynurenine (KYN) flux. This table outlines the sequential experimental steps to refine endothelial targeting, validate functional outcomes, monitor real-time neurotransmitter dynamics, evaluate effects on cancer metastasis, and ensure systemic safety, aiming to open new avenues for precise modulation of the neurovascular KYN metabolism.
| Next Steps | Purpose | |
|---|---|---|
| 1. Build bar-coded AAV libraries to refine endothelial specificity | Enhance targeting precision | |
| 2. Validate knock-down efficiency and KYN metabolite flux in brain-slice co-culture | Confirm functional impact on Trp metabolism | |
| 3. Monitor glutamate dynamics with optogenetic reporters in vivo | Track real-time neurotransmitter changes | |
| 4. Assess effects on tumor infiltration and behavior in KMO-high breast-cancer metastasis models | Evaluate therapeutic impact on metastasis | |
| 5. Conduct parallel safety screens to chart NAD pools and mitochondrial stress in non-target tissues | Ensure systemic safety and minimize off-target effects |
AAV, adeno-associated virus; KMO, kynurenine 3-monooxygenase KYN, kynurenine; NAD, nicotinamide adenine dinucleotide; Trp, tryptophan.
Table 4.
Converging Strategies for Real-Time Control and Monitoring of Cellular Function. Summary of complementary approaches integrating riboswitch-mediated gene regulation and pulsed light-induced calcium dynamics to enable real-time metabolic readouts without cellular damage.
Table 4.
Converging Strategies for Real-Time Control and Monitoring of Cellular Function. Summary of complementary approaches integrating riboswitch-mediated gene regulation and pulsed light-induced calcium dynamics to enable real-time metabolic readouts without cellular damage.
| Key Concept | Mechanism | Application | Advantages | |||
|---|---|---|---|---|---|---|
| Riboswitches with Z-lock or Photocleavable Linker | Light-controlled conformational changes in RNA elements regulate gene expression. | Precise gating of target gene expression in living cells | High specificity; reversible; minimal background activation | |||
| Pulsed Light-Driven Calcium Oscillations in Astrocytes | Tunable light pulses induce calcium signaling cascades without causing photodamage. | Real-time monitoring of cellular metabolism and signaling | Non-invasive; tunable frequency/amplitude; low cytotoxicity |
Table 5.
Critical Research Gaps in Chrono-pharmacology of Kynurenine (KYN)-Targeting Agents. This table delineates six principal gaps hindering the integration of circadian and sex-specific considerations into KYN-modulating therapies. Addressing these deficits will necessitate longitudinal, multi-omic chrono-pharmacology studies, sex-balanced preclinical models, and adaptive clinical trial designs incorporating real-time circadian monitoring and biomarker-driven dosing.
Table 5.
Critical Research Gaps in Chrono-pharmacology of Kynurenine (KYN)-Targeting Agents. This table delineates six principal gaps hindering the integration of circadian and sex-specific considerations into KYN-modulating therapies. Addressing these deficits will necessitate longitudinal, multi-omic chrono-pharmacology studies, sex-balanced preclinical models, and adaptive clinical trial designs incorporating real-time circadian monitoring and biomarker-driven dosing.
| Research Gap | Description | |
|---|---|---|
| 1. Absence of Chrono-pharmacology Trials | No clinical studies have evaluated time-of-day effects for IDO1/TDO2 inhibitors, KMO inhibitors, or KAT activators. Optimal administration schedules remain unexplored, impeding evidence-based chrono-dosing strategies. | |
| 2. Inadequate Stratification by Circadian Phase and Sex | Trials and PK/PD analyses seldom disaggregate data by both circadian timing and biological sex. Female-specific pharmacokinetic profiles are largely unreported, obscuring mechanistic reasons for sex-dependent toxicity reductions (e.g., afternoon regimens in lymphoma). | |
| 3. Undefined Mechanistic Links between Clock Genes, Immune Oscillations, and KYN Enzyme Activity | The interactions among peripheral circadian regulators, immune cell rhythmicity, and drug metabolism—especially modulation by estrogen and glucocorticoids on daily fluctuations of KYN pathway enzymes—remain mechanistically uncharacterized. | |
| 4. Lack of Wearable-Derived Chronotype Integration in Trial Design | Personalized chrono-type metrics from wearable sensors are not incorporated into clinical protocols. Without individual rhythm profiling, dosing algorithms cannot be tailored to synchronize drug exposure with each patient’s endogenous rhythm. | |
| 5. Sex and Circadian Biases in Preclinical Models | Preclinical experiments predominantly use male rodents under fixed light–dark schedules, neglecting sex-dimorphic and circadian-variant responses. This undermines translational relevance for patients exhibiting divergent chrono-biological profiles. | |
| 6. Absence of Validated Real-Time Biomarkers Coupling KYN Dynamics to Efficacy | There is a shortage of dynamic biomarkers that track KYN metabolite oscillations in real time and correlate these fluctuations with therapeutic outcomes. This gap limits the implementation of adaptive dosing regimens based on metabolic feedback. |
IDO1, indoleamine-2,3-dioxygenase-1; KYN, kynurenine; TDO2, tryptophan-2,3-dioxygenase-2; KAT, kynurenine aminotransferase: KMO, kynurenine 3-monooxygenase KYN, kynurenine; PD; pharmacodynamics; PK, pharmacodynamics.
Table 6.
Key Next Steps for Circadian Misalignment and Quinolinic Acid (QUIN) Dynamics Intervention Studies. This table summarizes the proposed strategies to investigate how circadian misalignment influences QUIN dynamics and to evaluate interventions that may mitigate these effects in shift workers through real-time monitoring and targeted therapies.
Table 6.
Key Next Steps for Circadian Misalignment and Quinolinic Acid (QUIN) Dynamics Intervention Studies. This table summarizes the proposed strategies to investigate how circadian misalignment influences QUIN dynamics and to evaluate interventions that may mitigate these effects in shift workers through real-time monitoring and targeted therapies.
| Next Steps | Purpose | |
|---|---|---|
| 1. Pilot a cross-over study where shift workers wear light and activity trackers and collect hourly capillary samples across two work cycles | Capture real-time physiological and circadian data in shift workers | |
| 2. Model QUIN dynamics versus lux-derived phase angle using mixed-effects chronobiology | Analyze the relationship between light exposure patterns and QUIN fluctuations | |
| 3. Overlay cortisol and melatonin rhythms to disentangle stress versus circadian effects | Separate stress-induced effects from circadian-driven changes in biomarkers | |
| 4. Test whether timed blue light blockers, melatonin, or time-restricted feeding blunt QUIN spikes | Evaluate interventions to mitigate QUIN elevations linked to circadian misalignment | |
| 5. Pilot a cross-over study where shift workers wear light and activity trackers and collect hourly capillary samples across two work cycles | Capture real-time physiological and circadian data in shift workers |
QUIN, quinolinic acid.
Table 7.
Next Steps in Chronopharmacology-Informed Adaptive Trial Designs. This table outlines the strategic steps to develop and pilot advanced Bayesian hierarchical models that integrate dosing time and level, wearable-derived chronotype data, and real-time safety biomarkers, aiming to optimize precision chronotherapy in clinical trials.
Table 7.
Next Steps in Chronopharmacology-Informed Adaptive Trial Designs. This table outlines the strategic steps to develop and pilot advanced Bayesian hierarchical models that integrate dosing time and level, wearable-derived chronotype data, and real-time safety biomarkers, aiming to optimize precision chronotherapy in clinical trials.
| Next Steps | Purpose | |
|---|---|---|
| 1. Simulate Bayesian hierarchical designs co-randomizing dose level and dosing hour, borrowing strength across adjacent time bins | Enhance efficiency and robustness in chrono-dose finding | |
| 2. Integrate wearable-captured chronotype to stratify randomization and inform priors | Personalize treatment timing based on individual circadian profiles | |
| 3. Embed rolling interim analyses that drop unfavorable time windows rather than doses | Optimize trial adaptation by focusing on optimal dosing windows | |
| 4. Pilot such designs in drugs with known chronotoxicities, using point-of-care melatonin or cortisol assays as safety triggers | Validate design feasibility and safety in chronotherapy-prone drugs | |
| 5. Simulate Bayesian hierarchical designs co-randomizing dose level and dosing hour, borrowing strength across adjacent time bins | Enhance efficiency and robustness in chrono-dose finding |
Table 8.
Critical Research Gaps in Translating Engineered Consortia for Multi-Drug Resistant Organisms Decolonization. This table highlights six principal gaps stalling the clinical translation of defined or engineered microbial consortia for decolonizing multidrug-resistant organisms (MDROs). Addressing these gaps requires standardized engraftment biomarkers, mechanistic phage–bacteria–host studies in gnotobiotic models, robust safety assessments of horizontal gene transfer, GMP-aligned manufacturing/QC, adaptive trial frameworks integrating antibiotics and diet rhythms, and harmonized regulatory pathways.
Table 8.
Critical Research Gaps in Translating Engineered Consortia for Multi-Drug Resistant Organisms Decolonization. This table highlights six principal gaps stalling the clinical translation of defined or engineered microbial consortia for decolonizing multidrug-resistant organisms (MDROs). Addressing these gaps requires standardized engraftment biomarkers, mechanistic phage–bacteria–host studies in gnotobiotic models, robust safety assessments of horizontal gene transfer, GMP-aligned manufacturing/QC, adaptive trial frameworks integrating antibiotics and diet rhythms, and harmonized regulatory pathways.
| Next Steps | Purpose | |
|---|---|---|
| 1. Unpredictable Engraftment and Lack of Comparative Trials | Engraftment remains erratic (e.g., VE303 only after antibiotic conditioning; VE707’s murine efficacy lacks human PK analogues). No head-to-head studies versus FMT)exist, so key efficacy drivers (bacteriocins, phages, niche competition) are unclear. | |
| 2. Scarce Durability and Mechanistic Data | Longitudinal sequencing hints at phage-mediated suppression post-FMT, but detailed mechanistic dissection of phage–bacteria–host interactions over time is missing, leaving durability of decolonization unpredictable. | |
| 3. Undercharacterized Safety and Horizontal Gene Transfer Risks | Potential for engineered strains (e.g., E. coli secreting microcins) to acquire resistance cassettes in vivo is insufficiently studied, so safety profiles and mitigation strategies for HGT remain undefined. |
|
| 4. Manufacturing and Quality Control Deficits | Current frameworks lag pharmaceutical standards: multi- LBPs lack verified batch-to-batch consistency in metabolite outputs and stability, hindering reproducibility and scale-up. | |
| 5. Absence of Adaptive Trial Designs Integrating Timing Factors | Trials rarely modulate dosing relative to antibiotic schedules, feeding rhythms, or bile acid fluctuations, despite evidence these factors gate colonization resistance; person-alized timing algorithms remain unexplored. | |
| 6. Fragmented Regulatory Pathways for Genetically Modified LBPs | Regulatory requirements differ across jurisdictions for engineered or defined consortia, deterring investment and delaying standardization; clear, harmonized guidelines are needed to accelerate safe, predictable, and durable applications. |
FMT, fecal microbiota transplant; HGT, horizontal gene transfers; LBPs, biotherapeutic products; PK, pharmacokinetics;.
Table 9.
Next Steps for kynurenic acid (KYNA) Encapsulation and Delivery Research. This table outlines the proposed experimental approaches for improving KYNA delivery systems, including stability testing, release profiling, triggered delivery, and bioavailability assessments in gnotobiotic models.
Table 9.
Next Steps for kynurenic acid (KYNA) Encapsulation and Delivery Research. This table outlines the proposed experimental approaches for improving KYNA delivery systems, including stability testing, release profiling, triggered delivery, and bioavailability assessments in gnotobiotic models.
| Next Steps | Purpose | |
|---|---|---|
| 1. Screen GRAS-grade polymers for KYNA compatibility under accelerated aging | Identify stable encapsulation materials suitable for KYNA under storage conditions | |
| 2. Map release profiles in simulated gastrointestinal fluids and pig colonic explants | Characterize release dynamics in physiologically relevant gut environments | |
| 3. Employ near-infrared–triggered nanocapsules to test on-demand bursts during inflammation | Enable controlled KYNA release in response to inflammation using light-triggered mechanisms | |
| 4. Quantify systemic versus luminal KYNA using LC-MS in gnotobiotic mice, benchmarking against Bifidobacterium-produced levels | Evaluate bioavailability and distribution of KYNA compared to natural microbial production | |
| 5. Screen GRAS-grade polymers for KYNA compatibility under accelerated aging | Identify stable encapsulation materials suitable for KYNA under storage conditions |
KYNA, kynurenic acid; LC-MS, liquid chromatography–mass spectrometry.
Table 12.
Key Gaps in Chronotherapeutic Clinical Trial Methodologies. Summary of critical gaps hindering the integration of circadian biology into clinical trial designs, highlighting areas needing methodological innovation and validation for effective translational application.
Table 12.
Key Gaps in Chronotherapeutic Clinical Trial Methodologies. Summary of critical gaps hindering the integration of circadian biology into clinical trial designs, highlighting areas needing methodological innovation and validation for effective translational application.
| Key Gap Description | Implication | |
|---|---|---|
| 1. Adaptive randomization models that incorporate dosing clock time as a modifiable arm | Enables dynamic optimization of treatment timing to enhance therapeutic efficacy and reduce toxicity | |
| 2. Validated software bridges between CGM, lactate, or KYN sensors and electronic trial master files | Facilitates seamless integration of real-time biomarker data into clinical trial records, improving data fidelity and regulatory compliance | |
| 3. Safety rules for rapid dose time shifts in outpatient settings | Ensures patient safety during temporal treatment adjustments, especially outside controlled environments | |
| 4. Patient-reported outcome measures (PROMs) sensitive to circadian toxicity | Captures subjective patient experiences related to time-dependent side effects, enhancing the assessment of tolerability and quality of life |
1 Tables may have a footer. CGM, continuous glucose monitoring; KYN, kynurenine;.
Table 13.
Immediate Next Steps for KYNA-Targeted Pacing and Dosing Optimization. This table summarizes the next phase in developing precision KYNA-targeted dosing strategies, focusing on pharmacokinetic profiling, Bayesian algorithm calibration, and integration of patient-reported outcomes to improve therapeutic tolerability.
Table 13.
Immediate Next Steps for KYNA-Targeted Pacing and Dosing Optimization. This table summarizes the next phase in developing precision KYNA-targeted dosing strategies, focusing on pharmacokinetic profiling, Bayesian algorithm calibration, and integration of patient-reported outcomes to improve therapeutic tolerability.
| Immediate Next Steps | Objective | |
|---|---|---|
| 1. Run a crossover pharmacokinetic study comparing saliva, plasma, and tumor microdialysate KYNA after RY103 micro-dosing | Evaluate the relationship between KYNA levels in different biological matrices post micro-dosing | |
| 2. Calibrate the Bayesian control algorithm using simulated patient data | Optimize the Bayesian algorithm for real-time adaptive control of dosing schedules | |
| 3. Embed patient-reported fatigue and cognitive scores to test whether KYNA-targeted pacing improves tolerability compared to fixed BID regimens | Assess the clinical benefit of KYNA-guided dosing on patient fatigue and cognitive outcomes relative to standard dosing practices |
KYNA, kynurenic acid.
Table 14.
Clues Supporting Feasibility of AI-Driven, Adaptive Gut–Brain Modulation. This table summarizes key lines of evidence that support the feasibility of integrating AI-driven therapeutic feedback loops with sensor-driven exercise platforms and probiotic modulation strategies, laying the groundwork for dynamic, real-time gut–brain axis interventions.
Table 14.
Clues Supporting Feasibility of AI-Driven, Adaptive Gut–Brain Modulation. This table summarizes key lines of evidence that support the feasibility of integrating AI-driven therapeutic feedback loops with sensor-driven exercise platforms and probiotic modulation strategies, laying the groundwork for dynamic, real-time gut–brain axis interventions.
| Clues Supporting Feasibility | Objective | |
|---|---|---|
| 1. Clinical artificial intelligence operations (ClinAIOps) frameworks for continuous therapeutic monitoring in hypertension and diabetes | Demonstrate the capability of AI frameworks to adaptively manage complex biological feedback loops | |
| 2. Kinect- or sensor-driven treadmills that auto-adjust speed based on user position | Validate the use of real-time biomechanical data to modulate physical activity interventions | |
| 3. Murine and human studies showing tailored probiotic blends reduce intestinal inflammation and modulate TRP metabolism | Provide proof of principle that microbiome-based therapies can influence gut–brain biochemical pathways |
AI, artificial intelligence.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



