Submitted:
09 June 2025
Posted:
11 June 2025
You are already at the latest version
Abstract
The AUG codon represents the canonical start site for translation initiation in eukaryotic cells, yet its proper function can be disrupted by a variety of regulatory mechanisms. In this review, conducted between April and June 2025, we comprehensively examined the most recent literature to identify key modulators that impair AUG-mediated initiation. These include suboptimal nucleotide contexts (e.g., weak Kozak consensus), RNA secondary structures adjacent to the start codon, and upstream open reading frames (uORFs) that compete with the primary initiation site. Furthermore, RNA-binding proteins and microRNAs were found to occlude ribosomal access, while variations in levels or activity of initiation factors—such as eIF1, eIF5, and related cofactors—can compromise start-site recognition. Collectively, these factors reduce the efficiency of 43S pre-initiation complex formation and accurate AUG selection, leading to decreased or misregulated protein synthesis with downstream effects on gene expression, stress response, development, and disease. By mapping structural, post-transcriptional, and initiation-factor–mediated regulatory pathways, this review highlights the multilayered control ordaining translation initiation. We also pinpoint gaps in existing knowledge and propose future research directions to enhance our understanding of AUG codon selection under both physiological and pathological contexts.
Keywords:
AUG initiation
; Kozak context
; uORF
; eIF1/eIF5
; RNA structure
; post-transcriptional regulation
1. Introduction
1.1. A Functional Overview of Translation and tRNA Binding Sites
Translation, or mRNA-directed protein synthesis, is a fundamental step in gene expression that shapes the composition and abundance of the cellular proteome. The ribosome, which catalyzes this process, contains three distinct tRNA-binding sites: the A-site (aminoacyl site), P-site (peptidyl site), and E-site (exit site). During the initiation phase, the ribosome binds the initiator methionyl-tRNA (Met-tRNAi) and the mRNA, positioning the start codon in the P-site where it base-pairs with the anticodon of Met-tRNAi. In the elongation phase, an aminoacyl-tRNA complementary to the next codon enters the A-site. The ribosome then catalyzes the transfer of the methionyl or peptidyl group from the P-site tRNA to the amino group of the A-site aminoacyl-tRNA, extending the polypeptide chain. This is followed by translocation, whereby the ribosome shifts the A-site tRNA—now carrying the growing peptide—to the P-site, while the deacylated tRNA is released from the E-site [1].
1.2. Molecular Insights into AUG Recognition During Translation Initiation
A start codon is a specific sequence of three nucleotides in messenger RNA (mRNA) that signals the beginning of protein synthesis. The most common start codon is AUG, which codes for the amino acid methionine. This codon plays a critical role in the initiation stage of translation, where ribosomes recognize the start codon to begin assembling amino acids into a polypeptide chain. Accurate translation of mRNA critically relies on the ability to initiate at the correct AUG codon. In most eukaryotic mRNAs, this is achieved through a scanning mechanism, whereby the small (40S) ribosomal subunit binds to the 5′ end of the mRNA and progressively examines the 5′ untranslated region (UTR) for an AUG codon situated within an appropriate sequence context. This process is guided primarily by base-pairing interactions between the AUG codon and the anticodon of the methionyl initiator tRNA (Met-tRNAi^Met). Over the past decade, significant advances in yeast genetics, reconstituted biochemical systems, and structural biology have collectively contributed to a deeper understanding of the molecular mechanisms underlying ribosomal scanning [2].
1.3. Mechanisms Contributing to the Inactivation of the AUG Codon as a Translation Initiation Site
1.3.1. Inadequate Kozak Sequence Context
Mechanism:
The Kozak consensus sequence, (GCC)GCC(A/G)CCAUGG, is critical for accurate recognition of the start codon. The purine at position -3 and the guanine at +4 relative to the "A" of AUG are particularly important.
- If these positions are mutated (e.g., A→U at -3), the recognition efficiency of AUG drops significantly.
- As a result, ribosomes may bypass this AUG and continue scanning for an alternative downstream AUG within a stronger context.
Experimental Evidence:
In the case of β-globin mRNA, mutations at positions -3 and +4 reduce translation initiation efficiency by over 90%, confirming the critical nature of these nucleotides [3].
1.3.2. Leaky Scanning Phenomenon
Mechanism:
Leaky scanning occurs when the ribosome fails to initiate translation at the first AUG codon due to a suboptimal Kozak sequence. Instead, it continues scanning and initiates at a downstream AUG or non-AUG codon.
- This mechanism allows the generation of multiple protein isoforms from a single mRNA.
- The frequency of leaky scanning can be modulated by translation initiation factors like eIF1 and eIF1A.
Biological Relevance:
In the CCAAT/enhancer-binding protein β (C/EBPβ), leaky scanning produces distinct isoforms (LAP, LAP, and LIP) with different functional roles in transcription regulation [4].
1.3.3. Phosphorylation of eIF2α Under Cellular Stress Conditions
Mechanism:
During stress responses (e.g., ER stress, amino acid starvation, oxidative stress), kinases such as PERK, GCN2, and PKR phosphorylate the α subunit of eIF2.
- Phosphorylated eIF2α sequesters the guanine nucleotide exchange factor eIF2B, reducing the recycling of eIF2-GDP to eIF2-GTP.
- This results in a global decrease in translation initiation from AUG codons.
- Some transcripts escape this repression through alternative translation initiation mechanisms.
Example:
The ATF4 mRNA contains multiple uORFs. Under normal conditions, these prevent its translation. Upon eIF2α phosphorylation, ribosomes bypass the inhibitory uORFs and initiate translation at the downstream main ORF[5].
1.3.4. RNA Secondary Structures in the 5' UTR
Mechanism:
Stable secondary structures in the 5′ untranslated region (5′UTR) of mRNAs, such as stem-loops and G-quadruplexes, can impede the ribosome’s ability to scan and identify the AUG start codon.
- Structures with a high thermodynamic stability (ΔG < -30 kcal/mol) near the 5′ cap or close to the AUG can inhibit initiation.
- eIF4A, an RNA helicase, is often required to resolve these structures to allow proper scanning.
Biological Relevance:
The ferritin mRNA contains an iron-responsive element (IRE) that forms a stem-loop structure in its 5′UTR, blocking translation when bound by iron regulatory protein (IRP) [6].
1.3.5. microRNA- and RBP-Mediated Repression of AUG Recognition
Mechanism:
microRNAs (miRNAs) and RNA-binding proteins (RBPs) can bind near or overlap the AUG codon and hinder ribosome recruitment or scanning.
- miRNAs often recruit the RNA-induced silencing complex (RISC) to block initiation.
- RBPs like TIA-1 or HuR can bind and remodel RNA conformation or directly compete with translation machinery.
Example:
miR-122 in hepatocytes regulates expression of multiple genes by binding 5′UTRs, thereby suppressing translation from canonical AUG start sites[7].
1.3.6. Utilization of Non-AUG Start Codons
Mechanism:
In some cases, non-canonical start codons such as CUG, GUG, and UUG are used to initiate translation.
- This usually requires the use of alternative initiation factors like eIF2A or eIF2D instead of the canonical eIF2.
- These codons can lead to the synthesis of N-terminally extended or truncated isoforms.
Examples:
- CUG is frequently used in the translation of transcription factor C/EBPα.
- Hox genes have shown evidence of non-AUG initiation, leading to protein variants with distinct localization or stability[8].
1.3.7. Upstream Open Reading Frames (uORFs) and Reinitiation
Mechanism:
uORFs are short coding sequences upstream of the main ORF that can regulate translation by trapping ribosomes or altering scanning behavior.
- After translating a uORF, the ribosome may dissociate or reinitiate at downstream sites depending on the availability of reinitiation factors.
- This regulation is sensitive to eIF2α phosphorylation and stress conditions.
Example:
The CHOP mRNA contains uORFs that prevent translation under normal conditions. Under stress, reinitiation allows expression of the pro-apoptotic CHOP protein [9].
1.3.8. Environmental Influences on AUG Codon Function
Mechanism:
Environmental factors such as temperature shifts, oxidative stress, hypoxia, viral infection, nutrient deprivation, and exposure to toxins or heavy metals can alter the molecular machinery that governs AUG codon recognition.
- These stressors often activate specific signaling pathways that lead to the phosphorylation or inactivation of key initiation factors (e.g., eIF2α, eIF4E).
- Some environmental conditions can also affect RNA structure by altering folding kinetics or stabilizing repressive elements near the AUG codon.
- Chronic exposure to environmental agents like arsenic or lead has been shown to induce long-term epigenetic changes, leading to altered ribosome behavior and start site selection.
Examples:
- Heat shock proteins (HSPs) are upregulated via alternative initiation mechanisms when canonical AUG recognition is impaired by temperature stress.
- Hypoxia-inducible factors (HIFs) are translated through cap-independent mechanisms like IRES when AUG-based scanning is inefficient.
Implications:
Environmental modulation of translation initiation contributes to adaptive gene expression in cancer, aging, development, and immune responses. It also plays a role in virus-host interactions, as many viruses hijack the host translational machinery to favor their own non-AUG start sites[10].
1.4. Non-Methionine Start Codons
1.4.1. Natural
Translation initiation via internal ribosome entry sites (IRES) represents an alternative mechanism that circumvents several canonical eukaryotic initiation pathways. Notably, IRES-mediated initiation does not necessarily require the conventional methionine start codon; instead, translation can commence at non-AUG codons such as GCU (alanine) or CAA (glutamine) [11].
Moreover, in mammalian cells, translation initiation can occur from the codon CUG, which is decoded by a specialized leucyl-tRNA, allowing for the incorporation of leucine as the initial amino acid. This non-canonical initiation mechanism operates independently of the eukaryotic initiation factor eIF2 and does not require the presence of an IRES-like secondary RNA structure. Instead, it relies on ribosomal scanning, with initiation efficiency enhanced by a favorable Kozak consensus sequence [12].
1.4.2. Engineered Start Codons
Engineered initiator tRNAs have been employed to direct translation initiation at the amber stop codon UAG in Escherichia coli. These synthetic tRNAs not only facilitate the incorporation of the canonical formylmethionine but also allow the insertion of formylglutamine. This is due to the ability of glutamyl-tRNA synthetase to recognize and charge the engineered tRNA with glutamine. It is important to recall that the bacterial initiation system primarily recognizes the formyl modification on the initiator tRNA rather than the identity of the amino acid itself [13].
Notably, a study has demonstrated that these amber initiator tRNAs do not support detectable levels of translation initiation from native genomic UAG codons. Instead, measurable initiation was observed exclusively in plasmid-based reporter constructs containing robust upstream Shine-Dalgarno sequences [14].
1.5. Met-tRNAiMet Is Generally, but Not Always, Used for Initiation in Eukaryotes
When translation is initiated from a non-AUG start codon, the identity of the aminoacyl-tRNA (aa-tRNA) utilized depends on both the nature of the start codon and the specific initiation mechanism involved. In the canonical eukaryotic initiation pathway—employed by the majority of protein-coding genes—the eukaryotic initiation factor 2 (eIF2) is responsible for delivering Met-tRNAi^Met to the ribosomal P site. Given the high specificity of eIF2 for Met-tRNAi^Met and its minimal affinity for alternative tRNAs, initiation through this pathway is presumed to involve Met-tRNAi^Met exclusively[15].
This assumption is supported by early in vitro experiments using [³⁵S]-labeled Met-tRNAi^Met, which revealed that reporter mRNAs containing non-AUG start codons such as CUG, GUG, ACG, UUG, AGG, AAG, AUA, AUC, or AUU still yielded full-length proteins incorporating radiolabeled methionine (Peabody, 1989). These findings strongly suggest that Met-tRNAi^Met can be utilized for initiation at these non-canonical start sites. More recently, mass spectrometry analyses have further corroborated the use of Met-tRNAi^Met at alternative start codons such as ACG and AUU [16].
While eukaryotic translation initiation typically relies on eIF2 to deliver Met-tRNAi^Met to the ribosomal P-site, accumulating evidence indicates that at least two additional initiation factors—eIF2A and eIF2D—are capable of mediating initiation at non-AUG start codons (Figure 1). Notably, eIF2A (distinct from the eIF2α subunit) can initiate translation in a GTP-independent manner and demonstrates a relaxed specificity toward tRNA binding. In vitro studies have shown that eIF2A binds both charged and uncharged forms of initiator Met-tRNAi^Met with comparable affinity [17,18].
Moreover, eIF2A may facilitate initiation at non-canonical codons such as CUG and UUG by delivering Leu-tRNA^Leu, although the precise affinity of eIF2A for aminoacylated Leu-tRNA remains unquantified [19,20]. It remains possible that eIF2A does not directly deliver Leu-tRNA but instead promotes its recruitment to the 40S ribosomal subunit. In support of this, Starck et al. (2012), using an ELISA-based reporter system, demonstrated that translation initiation at a CUG codon can occur up to ~40% more efficiently when initiated with Leu-tRNA^CUG than with Met-tRNAi^Met, implicating eIF2A in this process. Depletion of eIF2A specifically impaired initiation at the CUG codon without affecting AUG-initiated translation[12].
Despite these specialized roles, eIF2A appears to be non-essential under standard physiological conditions, as evidenced by the lack of overt phenotype in eIF2A knockout mice [21]. However, its expression is upregulated under stress conditions, and it has been implicated in IRES-mediated translation.These findings suggest that eIF2A may become particularly important in contexts where canonical eIF2-dependent translation is suppressed, thereby allowing non-AUG initiation to play a more prominent role in the cellular translational landscape[17,18,22].
Similar to eIF2A, the initiation factor eIF2D—also known as ligatin (LGTN)—has been demonstrated to mediate the delivery of Met-tRNAi^Met, including its uncharged form, to the P site of the 40S ribosomal subunit in a GTP-independent manner. Although eIF2D is primarily recognized for its essential role in ribosome recycling and reinitiation in vivo, it can also support translation initiation at non-AUG codons in vitro, albeit with selectivity depending on the RNA context[23,24].
For instance, eIF2D has been shown to initiate translation at a GUG codon on the 26S mRNA of Sindbis virus via delivery of Val-tRNA^GUG, but this activity was not observed on the hepatitis C virus (HCV) internal ribosome entry site (IRES) . This selectivity suggests that eIF2D’s ability to promote non-canonical initiation may be context-dependent and possibly influenced by the structural features of the 5′ untranslated region (UTR) of the mRNA[23,25].
Intriguingly, eIF2D has been identified as an autoantigen in patients with hepatocellular carcinoma, implying a potential role for this factor in oncogenesis or tumor-associated translational control mechanisms[26].
Moreover, the MCT-1/DENR heterodimer—which is structurally homologous to the N-terminal and C-terminal domains of eIF2D, respectively—has also been implicated in facilitating translation reinitiation and ribosome recycling[27,28]. In vitro studies indicate that MCT-1/DENR can promote the eIF2-independent recruitment of Met-tRNAi^Met to the ribosomal P site on select viral mRNAs. However, whether MCT-1/DENR can recruit other aminoacyl-tRNAs or initiate translation at non-AUG start codons remains an open question requiring further investigation[24,25].
In specific instances, such as those involving the internal ribosome entry sites (IRESs) of the cricket paralysis virus (CrPV) [29,30], Taura syndrome virus (TSV), and Plautia stali intestine virus [31].translation initiation proceeds via a mechanism that is fundamentally distinct from the canonical eukaryotic pathway. Notably, in these systems, neither an aminoacyl-tRNA (aa-tRNA) positioned in the ribosomal P site nor canonical eukaryotic initiation factors (eIFs) are required[32].
Instead, these viral IRES elements adopt highly structured conformations that directly associate with the 40S ribosomal subunit and facilitate the formation of an active 80S•IRES initiation complex. Within this complex, a specific pseudoknot motif of the IRES mimics the positioning of initiator Met-tRNAi^Met and the AUG start codon in the P site of the 40S subunit. This structural mimicry enables the recruitment of a cognate aa-tRNA to the ribosomal A site, thus initiating polypeptide synthesis from the A site, rather than the P site—a striking deviation from the canonical paradigm. To date, this unique initiation strategy has been exclusively described for a subset of viral IRESs. Nevertheless, these findings provide compelling evidence that translation initiation can be achieved in the complete absence of Met-tRNAi^Met and traditional initiation factors, thereby underscoring the remarkable versatility and adaptability of the translational machinery in response to distinct RNA architectures[33,34].
1.6. Usage of Non-AUG Initiation Codons Changes During Development and upon Stress
Ribosome profiling enables snapshots of translation to be taken, thereby revealing insights into how endogenous genes, including those containing non-AUG start codons, are regulated in response to developmental or external cues. Highlighting this approach, generated >25 ribosome profiling libraries to reveal temporal changes in translation as S. cerevisiae sporulate and transition from exponential vegetative growth to meiotic growth. As expected, there were broad changes in gene expression at both the mRNA and translation levels, with some genes displaying stage-specific meiotic translation. Furthermore, a number of previously annotated translation events, including from antisense RNAs, intergenic regions, and uORFs, were found to be specific to meiotic cells. Only 5% of ribosome footprints mapped outside annotated ORFs in vegetative cells, but this number increased to a staggering ∼30% in meiotic cells [35]. This suggests that there may be a profound shift in meiotic cells away from canonical translation, although functional roles (if any) for the majority of these novel ORFs remain unknown.
Many (but certainly not all) of the translated uORFs in meiotic cells use non-AUG start codons. uORFs have generally been thought to play an inhibitory role and act competitively to prevent translation of the downstream AUG-encoded ORF [36,37]. Indeed, about half of the AUG-encoded uORFs were associated with translational repression of their downstream ORFs in meiosis [35]. In stark contrast, translation of non-AUG-encoded uORFs was correlated primarily with increased translation of their downstream ORFs. The underlying mechanism responsible for this positive correlation remains unknown, but it is possible that these non-AUG uORFs may directly prime translation of their downstream ORF, or, alternatively, the uORF-encoded polypeptides may have functional roles [35]. Given that nearly 50% of RNA polymerase II transcription is devoted to production of ribosomal proteins when yeast are grown under vegetative conditions [38], it is tempting to speculate that changes in ribosomal protein (or eIF) stoichiometry and/or post-translational modifications may be responsible for the drastic changes in translation patterns observed in meiotic cells. Consistent with this idea, it has been shown recently that ribosome composition and ribosome protein stoichiometry can influence translation of particular classes of mRNAs [39].
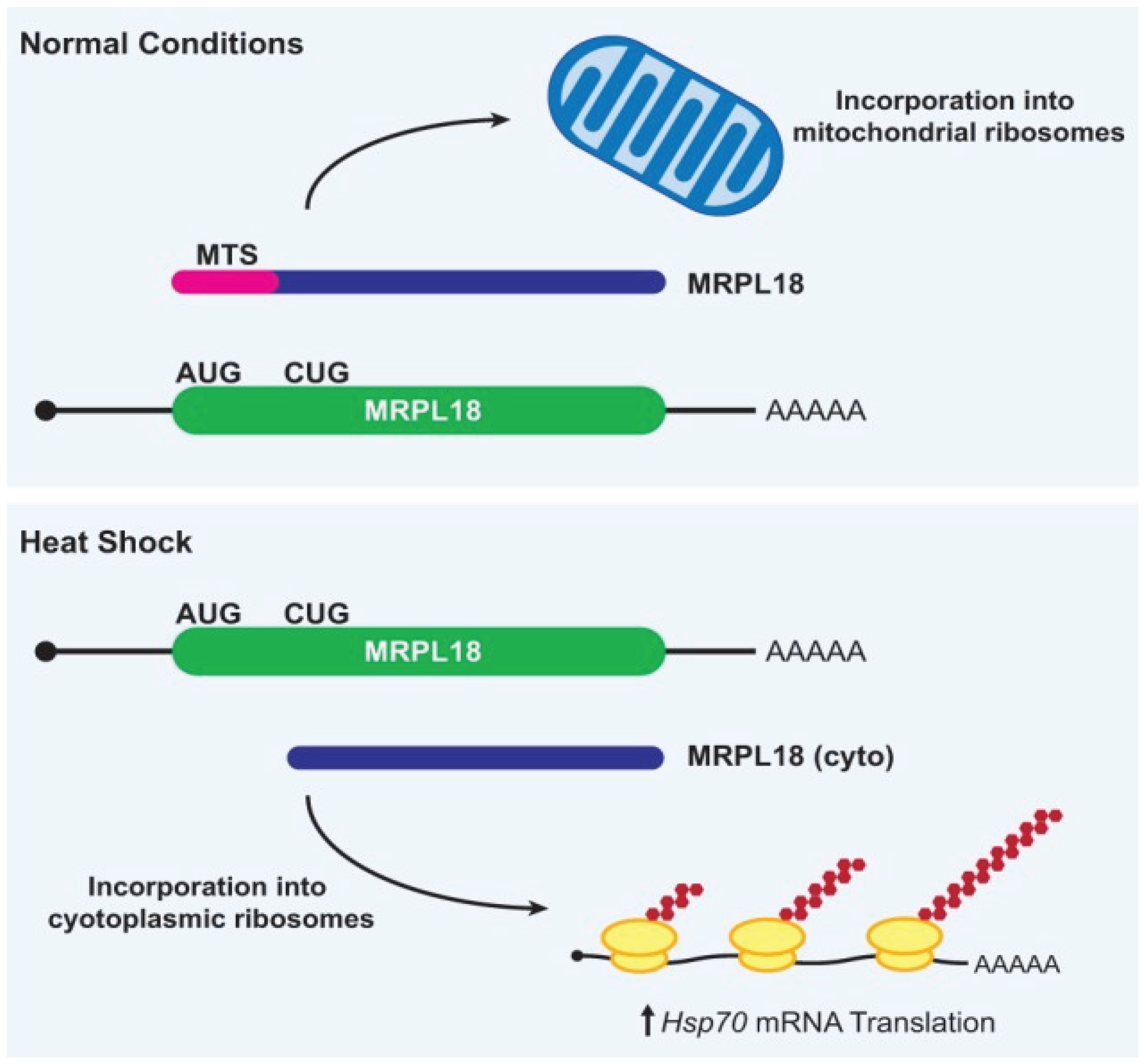
In mammalian cells, heat shock causes translation of MRPL18 (which encodes a mitochondrial large ribosomal subunit protein) to no longer initiate at the annotated AUG start codon but instead initiate at a downstream CUG codon (Figure 2). The truncated MRPL18 protein that is generated lacks the N-terminal mitochondrial targeting signal and thus is not incorporated into mitochondrial ribosomes. Instead, the truncated MRPL18 protein is incorporated into cytoplasmic ribosomes. These newly defined “hybrid” ribosomes are functional and are required for increased synthesis of the Hsp70 chaperone protein during heat shock [40]. This is perhaps because the presence of MRPL18 permits directed ribosome recruitment to Hsp70 mRNA, similar to what has been reported previously for RpL38 and the Hox genes [41].Regardless of the exact underlying mechanism, the MRPL18 locus beautifully demonstrates how changes in the efficiency of non-AUG start codon usage can have a profound impact on cell survival and homeostasis.
2. Conclusion
The AUG codon is the principal site for translation initiation in eukaryotes, yet its recognition is finely regulated by a complex interplay of molecular determinants. The efficiency of AUG recognition is influenced by factors such as the strength of the surrounding Kozak sequence, the presence of RNA secondary structures, the phosphorylation status of initiation factors, and the involvement of upstream open reading frames (uORFs) and microRNAs (miRNAs). Moreover, in specific cellular contexts, non-AUG codons can initiate translation, contributing to proteomic diversity and enabling adaptive responses. Together, these regulatory mechanisms highlight the evolutionary sophistication of translational control and emphasize the critical role of post-transcriptional regulation in cellular function, development, and disease.
References
- Asano, K. Why is start codon selection so precise in eukaryotes? Translation 2014, 2, e28387. [Google Scholar] [CrossRef] [PubMed]
- Hinnebusch, A.G. Molecular mechanism of scanning and start codon selection in eukaryotes. Microbiology and molecular biology reviews 2011, 75, 434–467. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M. Point mutations define a sequence flanking the AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell 1986, 44, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Hinnebusch, A.G. Molecular mechanism of scanning and start codon selection in eukaryotes. Microbiology and molecular biology reviews : MMBR 2011, 75, 434–467. [Google Scholar] [CrossRef] [PubMed]
- Spriggs, K.A.; Bushell, M.; Willis, A.E. Translational regulation of gene expression during conditions of cell stress. Molecular cell 2010, 40, 228–237. [Google Scholar] [CrossRef]
- Jackson, R.J.; Hellen, C.U.; Pestova, T.V. The mechanism of eukaryotic translation initiation and principles of its regulation. Nature reviews. Molecular cell biology 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Peabody, D.S. Translation initiation at non-AUG triplets in mammalian cells. The Journal of biological chemistry 1989, 264, 5031–5035. [Google Scholar] [CrossRef]
- Vattem, K.M.; Wek, R.C. Reinitiation involving upstream ORFs regulates ATF4 mRNA translation in mammalian cells. Proceedings of the National Academy of Sciences of the United States of America 2004, 101, 11269–11274. [Google Scholar] [CrossRef]
- Sonenberg, N.; Hinnebusch, A.G. Regulation of translation initiation in eukaryotes: mechanisms and biological targets. Cell 2009, 136, 731–745. [Google Scholar] [CrossRef]
- RajBhandary, U.L. More surprises in translation: initiation without the initiator tRNA. Proceedings of the National Academy of Sciences 2000, 97, 1325–1327. [Google Scholar] [CrossRef]
- Starck, S.R.; Jiang, V.; Pavon-Eternod, M.; Prasad, S.; McCarthy, B.; Pan, T.; Shastri, N. Leucine-tRNA initiates at CUG start codons for protein synthesis and presentation by MHC class I. Science 2012, 336, 1719–1723. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.; Shah, R.A.; Chembazhi, U.V.; Sah, S.; Varshney, U. Two highly conserved features of bacterial initiator tRNAs license them to pass through distinct checkpoints in translation initiation. Nucleic acids research 2017, 45, 2040–2050. [Google Scholar] [CrossRef] [PubMed]
- Vincent, R.M.; Wright, B.W.; Jaschke, P.R. Measuring amber initiator tRNA orthogonality in a genomically recoded organism. ACS synthetic biology 2019, 8, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Kolitz, S.E.; Lorsch, J.R. Eukaryotic initiator tRNA: finely tuned and ready for action. FEBS letters 2010, 584, 396–404. [Google Scholar] [CrossRef]
- Liang, H.; Chen, X.; Yin, Q.; Ruan, D.; Zhao, X.; Zhang, C.; McNutt, M.A.; Yin, Y. PTENβ is an alternatively translated isoform of PTEN that regulates rDNA transcription. Nature communications 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, S.M.; Park, J.H.; Keum, S.J.; Jang, S.K. eIF2A mediates translation of hepatitis C viral mRNA under stress conditions. The EMBO journal 2011, 30, 2454–2464. [Google Scholar] [CrossRef]
- Komar, A.A.; Gross, S.R.; Barth-Baus, D.; Strachan, R.; Hensold, J.O.; Kinzy, T.G.; Merrick, W.C. Novel characteristics of the biological properties of the yeast Saccharomyces cerevisiae eukaryotic initiation factor 2A. Journal of Biological Chemistry 2005, 280, 15601–15611. [Google Scholar] [CrossRef]
- Liang, H.; He, S.; Yang, J.; Jia, X.; Wang, P.; Chen, X.; Zhang, Z.; Zou, X.; McNutt, M.A.; Shen, W.H.; et al. PTENα, a PTEN isoform translated through alternative initiation, regulates mitochondrial function and energy metabolism. Cell metabolism 2014, 19, 836–848. [Google Scholar] [CrossRef]
- Sendoel, A.; Dunn, J.G.; Rodriguez, E.H.; Naik, S.; Gomez, N.C.; Hurwitz, B.; Levorse, J.; Dill, B.D.; Schramek, D.; Molina, H.; et al. Translation from unconventional 5′ start sites drives tumour initiation. Nature 2017, 541, 494–499. [Google Scholar] [CrossRef]
- Golovko, A.; Kojukhov, A.; Guan, B.-J.; Morpurgo, B.; Merrick, W.C.; Mazumder, B.; Hatzoglou, M.; Komar, A.A. The eIF2A knockout mouse. Cell Cycle 2016, 15, 3115–3120. [Google Scholar] [CrossRef]
- Starck, S.R.; Tsai, J.C.; Chen, K.; Shodiya, M.; Wang, L.; Yahiro, K.; Martins-Green, M.; Shastri, N.; Walter, P. Translation from the 5′ untranslated region shapes the integrated stress response. Science 2016, 351, aad3867. [Google Scholar] [CrossRef] [PubMed]
- Dmitriev, S.E.; Terenin, I.M.; Andreev, D.E.; Ivanov, P.A.; Dunaevsky, J.E.; Merrick, W.C.; Shatsky, I.N. GTP-independent tRNA delivery to the ribosomal P-site by a novel eukaryotic translation factor. Journal of Biological Chemistry 2010, 285, 26779–26787. [Google Scholar] [CrossRef] [PubMed]
- Skabkin, M.A.; Skabkina, O.V.; Hellen, C.U.; Pestova, T.V. Reinitiation and other unconventional posttermination events during eukaryotic translation. Molecular cell 2013, 51, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Skabkin, M.A.; Skabkina, O.V.; Dhote, V.; Komar, A.A.; Hellen, C.U.; Pestova, T.V. Activities of Ligatin and MCT-1/DENR in eukaryotic translation initiation and ribosomal recycling. Genes & development 2010, 24, 1787–1801. [Google Scholar]
- Wang, Y.; Han, K.-J.; Pang, X.-W.; Vaughan, H.A.; Qu, W.; Dong, X.-Y.; Peng, J.-R.; Zhao, H.-T.; Rui, J.-A.; Leng, X.-S.; et al. Large scale identification of human hepatocellular carcinoma-associated antigens by autoantibodies. The Journal of Immunology 2002, 169, 1102–1109. [Google Scholar] [CrossRef]
- Schleich, S.; Strassburger, K.; Janiesch, P.C.; Koledachkina, T.; Miller, K.K.; Haneke, K.; Cheng, Y.-S.; Küchler, K.; Stoecklin, G.; Duncan, K.E.; et al. DENR–MCT-1 promotes translation re-initiation downstream of uORFs to control tissue growth. Nature 2014, 512, 208–212. [Google Scholar] [CrossRef]
- Janich, P.; Arpat, A.B.; Castelo-Szekely, V.; Lopes, M.; Gatfield, D. Ribosome profiling reveals the rhythmic liver translatome and circadian clock regulation by upstream open reading frames. Genome research 2015, 25, 1848–1859. [Google Scholar] [CrossRef]
- Jan, E.; Sarnow, P. Factorless ribosome assembly on the internal ribosome entry site of cricket paralysis virus. Journal of molecular biology 2002, 324, 889–902. [Google Scholar] [CrossRef]
- Pestova, T.V.; Hellen, C.U. Translation elongation after assembly of ribosomes on the Cricket paralysis virus internal ribosomal entry site without initiation factors or initiator tRNA. Genes & development 2003, 17, 181–186. [Google Scholar]
- Cevallos, R.C.; Sarnow, P. Factor-independent assembly of elongation-competent ribosomes by an internal ribosome entry site located in an RNA virus that infects penaeid shrimp. Journal of virology 2005, 79, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, J.; Nakashima, N. Methionine-independent initiation of translation in the capsid protein of an insect RNA virus. Proceedings of the National Academy of Sciences 2000, 97, 1512–1515. [Google Scholar] [CrossRef] [PubMed]
- Fernández, I.S.; Bai, X.C.; Murshudov, G.; Scheres, S.H.; Ramakrishnan, V. Initiation of translation by cricket paralysis virus IRES requires its translocation in the ribosome. Cell 2014, 157, 823–831. [Google Scholar] [CrossRef]
- Koh, C.S.; Brilot, A.F.; Grigorieff, N.; Korostelev, A.A. Taura syndrome virus IRES initiates translation by binding its tRNA-mRNA–like structural element in the ribosomal decoding center. Proceedings of the National Academy of Sciences 2014, 111, 9139–9144. [Google Scholar] [CrossRef]
- Brar, G.A.; Yassour, M.; Friedman, N.; Regev, A.; Ingolia, N.T.; Weissman, J.S. High-resolution view of the yeast meiotic program revealed by ribosome profiling. science 2012, 335, 552–557. [Google Scholar] [CrossRef]
- Sachs, M.S.; Geballe, A.P. Downstream control of upstream open reading frames. Genes & development 2006, 20, 915–921. [Google Scholar]
- Chew, G.L.; Pauli, A.; Schier, A.F. Conservation of uORF repressiveness and sequence features in mouse, human and zebrafish. Nature communications 2016, 7, 11663. [Google Scholar] [CrossRef]
- Warner, J.R. The economics of ribosome biosynthesis in yeast. Trends in biochemical sciences 1999, 24, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Röst, H.L.; Teruel, M.N.; Barna, M. Heterogeneous ribosomes preferentially translate distinct subpools of mRNAs genome-wide. Molecular cell 2017, 67, 71–83. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, X.; Coots, R.A.; Conn, C.S.; Liu, B.; Qian, S.B. Translational control of the cytosolic stress response by mitochondrial ribosomal protein L18. Nature structural & molecular biology 2015, 22, 404–410. [Google Scholar]
- Xue, S.; Tian, S.; Fujii, K.; Kladwang, W.; Das, R.; Barna, M. RNA regulons in Hox 5′ UTRs confer ribosome specificity to gene regulation. Nature 2015, 517, 33–38. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Canonical and alternative initiator tRNAi-binding eIFs are differentially regulated and have different tRNAi-binding stringency. (Left) Canonical translation uses eIF2 in a GTP-dependent manner to deliver the canonical Met-tRNAiMet to the P site of the 40S ribosomal subunit. This allows initiation at AUG start codons as well as at non-AUG start codons, albeit at a reduced efficiency. (Right) eIF2A and eIF2D are somewhat similar to eIF2 but are GTP-independent and have the capability to bind both charged and uncharged forms of tRNAiMet. eIF2A can also use Leu-tRNACUG and initiate at CUG and UUG start codons in some mRNAs in vitro and in vivo. eIF2D, which regulates reinitiation, can initiate at AUG codons in vitro as well as use other aa-tRNAs for initiation in a selective manner.
Figure 1.
Canonical and alternative initiator tRNAi-binding eIFs are differentially regulated and have different tRNAi-binding stringency. (Left) Canonical translation uses eIF2 in a GTP-dependent manner to deliver the canonical Met-tRNAiMet to the P site of the 40S ribosomal subunit. This allows initiation at AUG start codons as well as at non-AUG start codons, albeit at a reduced efficiency. (Right) eIF2A and eIF2D are somewhat similar to eIF2 but are GTP-independent and have the capability to bind both charged and uncharged forms of tRNAiMet. eIF2A can also use Leu-tRNACUG and initiate at CUG and UUG start codons in some mRNAs in vitro and in vivo. eIF2D, which regulates reinitiation, can initiate at AUG codons in vitro as well as use other aa-tRNAs for initiation in a selective manner.
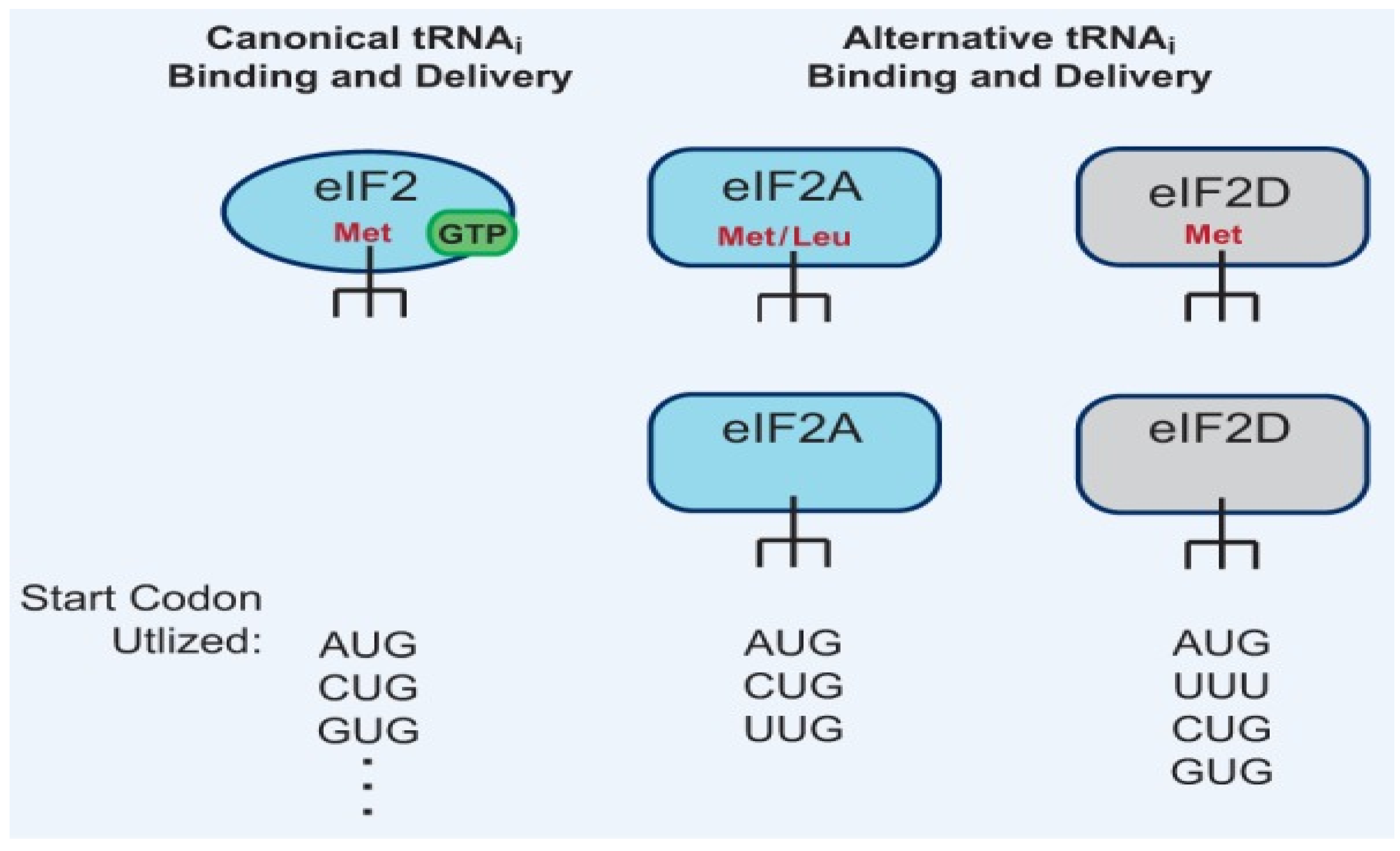
Figure 2.
Heat shock causes the production of a CUG-initiated MRPL18 protein that becomes incorporated into cytoplasmic ribosomes. (Top) Under normal growth conditions, the MRPL18 protein is synthesized from a canonical AUG start codon and includes the mitochondrial targeting signal (MTS; pink), which enables transport into mitochondria and subsequent incorporation into mitochondrial ribosomes. (Bottom) Heat shock causes a switch in the preferred start codon, resulting in initiation at a downstream CUG in the MRPL18 mRNA. The truncated MRPL18 (cyto) protein isoform lacks the MTS and is instead incorporated into cytoplasmic ribosomes, creating “hybrid ribosomes” that are required for increased Hsp70 mRNA translation during heat shock.
Figure 2.
Heat shock causes the production of a CUG-initiated MRPL18 protein that becomes incorporated into cytoplasmic ribosomes. (Top) Under normal growth conditions, the MRPL18 protein is synthesized from a canonical AUG start codon and includes the mitochondrial targeting signal (MTS; pink), which enables transport into mitochondria and subsequent incorporation into mitochondrial ribosomes. (Bottom) Heat shock causes a switch in the preferred start codon, resulting in initiation at a downstream CUG in the MRPL18 mRNA. The truncated MRPL18 (cyto) protein isoform lacks the MTS and is instead incorporated into cytoplasmic ribosomes, creating “hybrid ribosomes” that are required for increased Hsp70 mRNA translation during heat shock.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



