Submitted:
06 June 2025
Posted:
10 June 2025
You are already at the latest version
Abstract
The slender snipe eel Nemichthys scolopaceus Richardson, 1848 is cosmopolitan in tropical and temperate seas, inhabiting the mesopelagic and bathypelagic zone between 200 and 1000 m depth. It is known to be an active predator in DSL (Deep Scattering Layer) and NBA (Near Bottom Assemblage), feeding mostly decapods and euphausiids crustaceans, and playing a central role in carbon fluxes through meso and bathypelagic ecosystems. Despite its potential importance in deep trophic web ecosystem, Nemichthys scolopaceus has been little investigated about its trophic ecology. The aim of this paper is start to fill this knowledge gap. A total of 35 specimens of N. scolopaceus caught through bottom trawling in Mediterranean Sea were analyzed in laboratory for stomach content. As expected, mainly decapod crustaceans were found, in particular Plesionika martia, Pasiphaea multidentata, Funchalia woodwardi, Robustosergia robusta species. The degree of digestion observed was in all cases rather high. Our findings seem to confirm the specialist diet of Nemichthys scolopaceus based on shrimp-like crustaceans.
Keywords:
deep-water species
; feeding habits
; morphological prey counts
; pelagic eel
; Nemichthys scolopaceus
1. Introduction
The slender snipe eel Nemichthys scolopaceus Richardson, 1848 is a cosmopolitan Anguilliformes, reported in the Atlantic, Indian, and Pacific Oceans and in the Mediterranean Sea [1]. N. scolopaceus is known to inhabit the mesopelagic and bathypelagic depth strata [2,3], mainly between 200 and 1000 m [4]. Differently from the other Nemichthyidae species, which don't seem to be strong vertical migrants, N. scolopaceus, conversely, seems to carry out wide vertical displacements along the water column to capture prey [5,6]. According to Gartner et al. [7], N. scolopaceus was frequently observed in both the DSL (Deep Scattering Layer) and NBA (Near Bottom Aggregation) layers, over the North Carolina and Virginia continental slopes, in the western North Atlantic Ocean.
This species shows marked sexual and ontogenetic dimorphism. At the juvenile stage, all individuals have non-occlusible, beak-like jaws armed with very small villiform teeth. As soon as reaching sexual maturity, males are subjected to jaws radically shortening and complete loss of teeth [8] while females keep a morphology similar to juveniles. These deep changes in the physical features of male specimens are likely due to semelparity, a biological phenomenon in which reproduction is rapidly followed by death [9]. Moreover, the male N. scolopaceus will not be able to catch their preys, concentrating their energy on reproduction to the detriment of their life [8]. Adult males were found rarely in the Mediterranean Sea [10].
Nemichthyid eels may have several predation strategies, including ambushing prey that passes near them while they maintain a vertical body position [11], or actively chasing [7,12]. In the Gulf of Maine (North-Atlantic) N. scolopaceus fed exclusively on crustaceans, mostly decapods, and euphausiids, avoiding potential prey that were very abundant in that habitat (‘oceanic rim’ ecosystem, [13], such as fishes, cephalopods, and other crustaceans (e.g. amphipods, crabs, copepods). This prey selectivity, linked to the high biomass of decapod crustaceans and large euphausiids in this oceanic zone and their ecological importance, and the fact that N. scolopaceus is one of the few mesopelagic fish predators of shrimps and euphausiids in the area, underlined the central role in carbon fluxes through meso and bathypelagic ecosystems [6].
On the other hand, the metabarcoding analysis of gut contents carried out by Bucklin et al. [14] highlighted that the slender snipe eel N. scolopaceus, though foraged mostly on large crustaceans, occasionally preyed smaller zooplankton (e.g. copepods), pelagic mollusks, and fish.
The aims of this study were to expand the knowledge about Nemichthys scolopaceus, fill the gap of information about its trophic ecology, especially in the Mediterranean Sea, and compare our findings with other studies carried out in other zones of the global ocean. In particular, we tried to understand if also in the Central Mediterranean Sea this Nemichthydae feed exclusively (or almost exclusively) large shrimp-like crustaceans, as just highlighted [6,14,15].
2. Materials and Methods
2.1. Study Area and Samples Collection
A total of 35 N. scolopaceus specimens were collected through bottom trawling (Table 1). 23 of these as bycatch of commercial fishing in the Gulf of Patti (Southern Tyrrhenian Sea, Central Mediterranean) during the summer of the years 2022-23. The remaining 12 specimens were collected during three oceanographic cruises of scientific fishing (NAUCRATES and MEDINEA in 2022, MEDITS in 2024), off the Calabria and Campania coast in the Southern Tyrrhenian Sea, and between Sardinia Island and Tunisian coast in the Sardinia Channel, respectively (Figure 1). A standard bottom trawl is the equipment that is normally used in commercial fishing. The net is made up of two panels with a 40 mm square mesh net. The sampling equipment utilized both in MEDINEA22 and NAUCRATES22 is quite similar to a standard commercial bottom trawl, with the same square mesh size, but has some experimental features like the adding of a metallic grid in the NAUCRATES22 cruise. The sampling gear used during the MEDITS campaign is totally different: it consists of a four-panel bottom trawl (model IFREMER GOC73) capable of operating between 10-800 m deep. The net has a mesh side of 10 mm, which means it has a mesh opening of 20 mm and a selectivity as low as possible. The trawl generally operates with a vertical opening of 2-3 m (although the maximum vertical opening is 7.40 m), and a horizontal opening between 35-40 m. In most cases, N. scolopaceus were captured near the seabed, at depths between 18 and 700 m. The specimens were frozen on board at -20°C.
2.2. Laboratory Analysis
In the laboratory, the specimens of N. scolopaceus were thawed, and the biomass and morphometric measurements were carried out. The Wet Weight (WW, ±0,01 g) was taken operating analytical balance (Sartorius-ENTRIS32O2-1S 0,01g) after drying the fish body with blotting paper. Measurements of Total Lenght (TL to the nearest 0.5 cm) were carried out. Moreover, because the peculiar morphology of this Nemichthydae, with its long and very thin body, can cause fragmentation of the tail or snout, the distance between the eye and gill cover was measured (EGD: Eye-Gill cover Distance, ±0.1 mm, Figure 2)
The stomachs were extracted and preserved in ethanol/seawater solution (70/30). Subsequently, the stomachs were dried with blotting paper, weighted (WW) by mean analytical balance (Mettler-Toledo AG204 0.1 mg) and carefully opened operating with scissors and forceps under stereomicroscope (ZEISS STEMI SV 8 and Leica WILD M10) observation.
Stomach fullness degree was estimated from 0 to 4, with 0 = empty stomach, 1 =only traces of prey, 2 = stomach filled from partly to half, 3 = stomach full but not stretched, and 4 = a full and stretched stomach. The degree of digestion of prey items was evaluated according to the following scale: 1 = no evident signs of digestion, prey whole and complete; 2 = prey partly digested with missing portions; 3 = prey from enough to strongly digested, only pieces remaining; and 4 = almost totally digested, only traces remaining [6]. Prey identification was carried out to species if possible, or to the lowest feasible taxonomic level, by the following taxonomic manuals: [16] for decapods crustaceans; [17] for fish otoliths and [18,19] for fish scales). Prey items were preserved in ethanol/seawater solution (70/30). After prey extraction, the stomach tissues were weighted to obtain the stomach content’s weight (subtracting this weight from full stomach weight). Subsequently, the biomass of preys was calculated according to [20,21,22,23,24,25].
2.3. Trophic Indexes
Trophic indexes were calculated for individuals with stomachs containing food, with the aim to evaluate the dietary composition of N. scolopaceus. First of all, Percentage of Vacuity: , where nes is the number of empty stomachs, while nt is the total number of stomachs. The degree of stomach fullness was estimated by the index %SCI (Stomach Content Index) as follows:, where the numerator reports the sum of the weights of the prey found in the stomach of the predator and the denominator reports the value resulting from the difference between the weight of the predator and that of the prey ingested by it. The average value of %SCI was subsequently calculated.
The importance of each prey i was assessed by calculating the following food indices. Percentage in number: × 100, where ni is the total number of individuals of prey i found in the stomachs, while nt is the total number of prey; Weight percentage: %Wi = x 100, where wi is the total weight of individuals of prey i found in the stomachs, while wt is the total weight of all prey; Percentage frequency: x 100, where nsi is the number of stomachs containing prey i, while ns is the total number of stomachs containing prey. The Relative Importance Index [26,27,28] was also estimated for each prey i, which takes into account and integrates the values of the previous food indices (%N, %W, %F): . The percentage contribution of the Relative Importance Index for each prey i (%IRIi) was then estimated for each prey i (%IRIi) on the total prey: x 100.
The feeding strategy of N. scolopaceus was evaluated by applying the graphic method of [29], modified by [30], which plots on a two-dimensional graph the specific abundance of each prey category with respect to the frequency of finding of the prey itself in the stomach contents. This method allows us to deduce information on the importance of the prey and the feeding strategy used by the predator. In this graphical representation, the specific abundance of each prey i (Pi) is calculated as follows: × 100, where Si is the total abundance (as weight or number) of prey i; Sti is the total stomach content only of the samples in whose stomachs prey i is present.
The trophic level (TROPH; [31] of N. scolopaceus was estimated by mean the weight participation and the trophic level of each prey species to the diet: TROPH=1+∑DCj∗TROPHj where TROPHj is the fractional trophic level of prey j and DCj represents the fraction of j in the diet of N. scolopaceus. The TROPH can range from 2.0, for herbivorous/detritivorous, to 5.0, for piscivorous/carnivorous organisms [32,33,34]. Trophic level values of each prey item were evaluated according to [33].
3. Results
3.1. Trophic Ecology
The 35 N.s scolopaceus specimens examined for stomach contents ranged from 500 to 1325 mm Total Lenght (TL), having a mean value of 807.97 mm (Standard deviation: ±189.95 mm). The individuals were assigned, according their TL, to size classes of 100 mm: from 500 to 590 mm TL (6 specimens), from 610 to 690 mm TL (6 specimens), from 730 to 785 mm TL (5 specimens), from 815 to 890 mm TL (6 specimens), from 900 to 995 mm TL (6 specimens), and from 1000 to 1088 mm TL (5 specimens) with only one specimen that measured 1325 mm (Figure 3).
The Wet Weight resulted from 2.94 to 76 g (Mean WW: 31.29 g; standard deviation ±23.23 g), while the Eye-Gill cover Distance varied from 5.8 to 25.5 mm (Mean EGD: 16.5 mm; standard deviation ±5.45 mm, Figure 4). Between individuals here investigated, no adult males were present, given the presence of beaks, that are absent in males after metamorphosis.
Of the 35 stomachs analyzed, 4 were empty, while 31 contained prey items (% vacuity= 11.43%). The Stomach Content Index (SCI) ranged from 0.0028% to 14.5629%, with a mean value of 1.65%. A total of 34 preys were found in stomachs of N. scolopaceus (Figure 5), with a mean number of 1.097 prey per specimen. The list of food items of N. scolopaceus is reported in Table 2, together with the values of dietary indexes (%N, %W, %F, %IRI) for each prey. Among the 31 stomachs containing food items, 12 contained only unidentifiable traces because of the high degree of digestion. In the remaining ones, we observed mainly decapod crustaceans, in particular: Plesionika martia (A. Milne-Edwards, 1883), Pasiphaea multidentata (Esmark, 1866), Funchalia woodwardi (Johnson, 1868), Robustosergia robusta (Smith, 1882), the genera Funchalia and Plesionika, and unidentified crustaceans. Moreover, we found traces of Teleostei: one otolith of Stomias boa (Risso, 1810), and scales of Macrouridae. Some sand was found in one stomach.
The degree of digestion observed was in all cases rather high (2-4; preys partially, highly, or almost completely digested). We did not find any prey whole and complete. Unidentified preys resulted the most numerically abundant (%N=35.29) and frequently found (%F=38.71) in stomachs of N. scolopaceus in the present study, followed by unidentified crustaceans (%N= 17.76; %F=19.35) and decapods (%N=14.71; %F=12.90). On the biomass level, has been confirmed the prevalence of unidentified preys (%W=35.52%), unidentified crustaceans (%W=15.81%), and decapods (%W=13.18%), but was possible starting to notice the importance of Teleostei Macrouridae (%W=11.05%), Plesionika martia (%W=9.31%), and Plesionika sp. (%W= 8.33%). Calculation of %IRI further highlighted the relevance of Teleostei Macrouridae (%IRI= 18.66%), Plesionika sp. (%IRI=15.19%), and Plesionika martia (%IRI= 10.46%), with standing out of Pasiphaea multidentata (%IRI=10.47%).
The estimated value of the index of trophic level (TROPH) for N. scolopaceus is 4.40.
3.2. Feeding Strategy
The graphic method of Costello [29], modified by Amundsen [30], plots on a two-dimensional graph the specific abundance, for the number (Figure 6a), and for weight (Figure 6b) of each prey category with respect to the frequency of finding the prey itself in the stomach contents, showed that the diet of N. scolopaceus was dominated by unidentified preys both in numerical and biomass point of view of biomass, while other prey items if considered at species levels, don’t stand out for abundance either weight. Nevertheless, the predominance of crustaceans was highlighted by considering them to the group level, regardless of single species, genera, and family (hence including unidentified crustaceans; Figure 7).
4. Discussion
The slender snipe eel Nemichthys scolopaceus has been poorly studied in general, especially from the trophic point of view. In particular, in the Mediterranean Sea, the food habits of this Nemichthydae have never been investigated. Only three recent studies focused on the diet of N. scolopaceus, all carried out in the NW Atlantic [6,14,15]. The present paper aims to contribute to filling the gap of knowledge about the trophic ecology of N. scolopaceus, particularly in our study area. Of the 35 specimens of N. scolopaceus examined by us, 31 had stomachs containing preys and 4 were empty, resulting in a vacuity percentage of 11%, decisively lower than that detected by [6] (30%), in turn even less of other mesopelagic predator fishes [12], that proved to be until 60%. This finding could confirm that N. scolopaceus has a higher consumption rate with respect to other mesopelagic fish. However, the high percentage of stomachs containing preys might be due to the slow digestion of this Nemichthydae [6,35,36], which carries food to remain in the stomach for a longer time. In addition, the long digestion time did not permit an evaluation of the feeding chronology in an accurate way [6]. Moreover, the degree of digestion of preys found by us in the stomachs was in all cases rather high (2-4; preys partially, highly, or almost completely digested). We did not find any prey whole and complete. Prey might have been ingested more than 24 hours before fish sampling [6,12,35,36], also because of the relatively big size of preys found in this study, as soon as in previously [6,15].
According to some latest studies [6,15], in the stomachs of N. scolopaceus mainly decapod crustaceans are found, as we did in this study. However, we did not detect any traces of euphausiids, which conversely represented a relevant portion of diets of N. scolopaceus [6,15]. Nevertheless, given the considerable percentage of unidentified preys because of their high degree of digestion, we did not totally exclude that also in the Mediterranean Sea N. scolopaceus feeds on euphausiids, in addition to decapod crustaceans. Further studies will be necessary to investigate this aspect of food habits of this Nemichthydae in the Mediterranean Sea. Moreover, in the stomach contents of N. scolopaceus we found traces of teleosts, as soon as stated by [14]. However, given also that in the studies of [6,15] no traces of fish have been found, and given that we found only one otolith (Stomias boa) and three scales (Macrouridae), we could hypothesize secondary ingestion, in other words, the possibility that otolith and scales found by us in the stomachs of N. scolopaceus originated in turn by stomach contents of crustaceans eaten by N. scolopaceus itself. In support of this hypothesis, it must be said that at least two of preys found by us in eels’s stomach contents, the decapods Pasiphaea multidentata and Plesionika martia, are known to be active predators of small fishes, as well as scavengers [37,38]. Moreover, either in the study of [14] no whole fish specimen was found in the stomach contents of N. scolopaceus, but rather the presence of teleosts has been detected by mean DNA metabarcoding [14], which could not exclude the possibility of secondary ingestion. Further reinforcement to this hypothesis is the predation strategy of N. scolopaceus. In fact, this eel is highly selective for shrimp-like crustaceans, also in function of its morphology [5,6,15,39]. The thin and elongated jaws of juvenile and adult female individuals of N. scolopaceus are covered with innumerable and very little villiform teeth, which can stick to the crustaceans’s antennae; this fact, joined to the extension of this latter, helps the mechanism of crustacean entrapment by entanglement of antennae [5,6,15,39]. According to [6,15] this peculiar feeding way, together with non-occlusible jaws, could contribute to making this Nemichthydae extremely selective for large crustaceans, avoiding other potential prey present in the environment, such as fishes, cephalopods, and other crustaceans (e.g. amphipods, crabs, copepods). Further studies may serve to shed light on this aspect as well, in addition to the apparent absence of euphausiids that we detected.
Prey found by us in the stomach contents of Nemichthys scolopaceus are benthic (Plesionika martia, Macrouridae) and benthopelagic (Pasiphaea multidentata) organisms linked to muddy bottoms between 300 and 700 m of depth [12,16,18,20,21,22,23,25,37,38] or mesopelagic (Robustosergia robusta, Funchalia woodwardi, Stomias boa) organisms that can be found near the bottom [7,12,16,21,25] at the same depths. Most specimens of this Anguilliformes investigated in the present study were caught exactly at these depths, near the bottom, where, as just stated, the crustacean and fish species detected in their stomachs can frequently be found. Therefore, the type of preys found in the present study can only confirm the statement of [7] that, in NW Atlantic, observed Nemichthys scolopaceus in the Near Bottom Aggregations. The finding of some sand in the stomach of one specimen of N. scolopaceus examined by us was a further sign that this eel fed frequently near the bottom. On the other hand, two specimens (on 35 sampled) of Nemichthys scolopaceus investigated in the present paper were caught at low depth during daytime, in one case at 83-86 m few hours after sunrise (around h 7:00), in the other case at the very low depth of 18-26 m, with the sun around noon (h 12-12:30). This fact could confirm that N. scolopaceus, despite living habitually between 200 and 1000 m (or deeper) [40], is able to carry out wide displacements along water column to catch preys [5,6].
Author Contributions
Conceptualization, A.GE., A.S., A.P., R.M., L.G. and A.GR; Methodology, Y.G., F.V. and D.M.; Software, F.V. and D.D.P.; Validation, A.GE., A.S. and A.GR.; Formal Analysis, A.GE., A.S., C.L. and A.GR; Investigation, A.GE., A.S., A.P., F.V., R.M., L.G., P.C. and A.GR; Data Curation, A.P., F.V., D.M. and L.G.; Writing – Original Draft Preparation, A.GE., A.S., C.L., A.P., R.M., F.V., D.D.P., L.G., P.C. and A.GR; Writing – Review & Editing, A.GE., A.S., R.M., L.G. and A.GR; Supervision, A.GR.;
Funding
No funds are currently available for this research.
Data Availability Statement
Data are contained within the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bilecenoglu, M.; Kaya, M.; Irmak, E. First records of the slender snipe eel, Nemichthys scolopaceus (Nemichthyidae), and the robust cusk-eel, Benthocometes robustus (Ophidiidae), from the Aegean Sea. Acta ichthyologica et piscatoria 2006, 36, 85–88. [Google Scholar] [CrossRef]
- Nielsen, J.G.; Smith, D.G. The eel family Nemichthyidae (Pisces, Anguilliformes). Dana Report 1978, 88. [Google Scholar]
- Fishelson, L. Comparative morphology of deep-sea eels, with particular emphasis on gonads and gut structure. J. Fish. Biol. 1994, 44, 75–101. [Google Scholar] [CrossRef]
- Smith, D.G.; Tighe, K.A. Snipe eels. Family Nemichthyidae. In: Collette, B.B.; Klein-MacPhee G (eds) Fishes of the Gulf of Maine 3rd edn; Smithsonian Institute Press, Washington, D.C. 2002, pp. 100–101.
- Karmovskaya, E.S. Systematics and some ecology of the snipe eels of the family Nemichthyidae. Proc. PP Shirshov Inst. Oceanol. 1982, 118, 151–161. [Google Scholar]
- Feagans-Bartow, J. N.; Sutton, T. T. Ecology of the oceanic rim: Pelagic eels as key ecosystem components. Marine Ecology Progress Series 2014, 502, 257–266. [Google Scholar] [CrossRef]
- Gartner, J.V. Jr.; Sulak, K.J.; Ross, S.W.; Necaise, A.M. Persistent near-bottom aggregations of mesopelagic animals along the North Carolina and Virginia continental slopes. Mar. Biol. 2008, 153, 825–841. [Google Scholar] [CrossRef]
- Smith, D.G.; Nielsen, J.G. Family Nemichthyidae: Snipe eels. In: Böhlke, E.B. (ed) Fishes of the western North Atlantic, Part 9, Vol 1. Sears Foundation for Marine Research, New Haven, CT 1989, pp 441-459. [CrossRef]
- Charter, S.R. Nemichthyidae: Snipe eels. In: H.G. Moser (ed.) The early stages of fishes in the California Current region. California Cooperative Oceanic Fisheries Investigations (CalCOFI) Atlas 1996, 33, 122–129.
- Bayhan, Y. K.; Ergüden, D.; Ayas, D. The first occurrence of male specimen of Nemichthys scolopaceus (Richardson, 1848) from Eastern Mediterranean. Turkish Journal of Maritime and Marine Sciences 2020, 6, 137–143. [Google Scholar]
- Miller, M.; Koyama, S.; Mochioka, N.; Aoyama, J.; Watanabe, S.; Tsukamoto, K. Vertical body orientation by a snipe eel (Nemichthyidae, Anguilliformes) in the deep mesopelagic zone along the West Mariana Ridge. Marine and Freshwater Behaviour and Physiology 2014, 47, 265–272. [Google Scholar] [CrossRef]
- Gartner, J.V. Jr.; Crabtree, R.E.; Sulak, K.J. Feeding at depth. In: Randall, D.J.; Farrell, A.P. (eds). Deep-sea fishes. New York (NY), Academic Press; pp. 115–193. Marine and Freshwater Behaviour and Physiology 1997, 271. [CrossRef]
- Merrett, N.R. Biogeography and the oceanic rim: A poorly known zone of ichthyofaunal interaction. Pelagic Biogeography, Proc of an Int Symposium. UNESCO Tech. Pap. Mar. Sci 1986, 49, 201–209. [Google Scholar]
- Bucklin, A.; Batta-Lona, P.G.; Questel, J.M.; McMonagle, H.; Wojcicki, M.; Llopiz, J.K.; Wiebe, P.H. Metabarcoding and morphological analysis of diets of mesopelagic fishes in the NW Atlantic Slope Water. Frontiers in Marine Science 2024, 11, 1411996. [Google Scholar] [CrossRef]
- Feagans, J. N. Trophic ecology of the slender snipe eel, Nemichthys scolopaceus (Anguilliformes: Nemichthyidae). M.S. thesis 2008. Boca Raton (FL) Florida Atlantic University, p. 30.
- Falciai, L.; Minervini, R. Guida ai crostacei decapodi d'Europa Seconda edizione. Ricca editore . 2023. [Google Scholar]
- Tuset, V.M.; Lombarte, A.; Assis, C.A. Otolith atlas for the western Mediterranean, north and central eastern Atlantic. Scientia Marina 2008, 72(S1), 7–198. [Google Scholar] [CrossRef]
- Cohen, D.M.; Inada, T.; Iwamoto, T.; Scialabba. Gadiform Fishes of the World. FAO Fisheries Synopsis 1990, 125 (10), pp. 453.
- Bräger, Z.; Moritz, T. A scale atlas for common Mediterranean teleost fishes. Vertebrate zoology 2016, 66, 275–386. [Google Scholar] [CrossRef]
- Apollonio, S. Breeding Fecundity of the Glass Shrimp, Pasiphaea multidentata (Decapoda, Caridea), in the Gulf of Maine. Journal Fisheries Research Board of Canada 1969, 26, 1969–1983. [Google Scholar] [CrossRef]
- Company, J. B.; Sardà, F. Growth parameters of deep-water decapod crustaceans in the Northwestern Mediterranean Sea: A comparative approach. Marine Biology 2000, 136, 79–90. [Google Scholar] [CrossRef]
- Chilari, A.; Thessalou-Legaki, M.; Petrakis, G. Population structure and reproduction of the deep-water shrimp Plesionika martia (decapoda: Pandalidae) from the eastern Ionian Sea. Journal of Crustacean Biology 2005, 25, 233–241. [Google Scholar] [CrossRef]
- Başusta, N.; Başusta, A. Length- Weight Relationship and Condition Factor of Hollowsnout grenadier (Coelorinchus caelorhincus, (Rinso, 1810) From Iskenderun Bay, northeastern Mediterranean, Turkey. International Marine & Freshwater Sciences Symposium Proceedings (MARFRESH2018). In: Özcan G, Tarkan A.S, Özcan T. (Eds.). 2018. Proceeding Book, International Marine & Freshwater Sciences Symposium, 18-21 October 2018, Kemer-Antalya/Turkey 2018, 300-302.
- Sousa, R.; Gouveia, L.; Pinto, A. R.; Timóteo, V.; Delgado, J.; Henriques, P. Weight-length relationships of six shrimp species caught off the Madeira Archipelago, Northeastern Atlantic. Brazilian Journal of Biology 2018, 79, 133–138. [Google Scholar] [CrossRef]
- Guerra-Marrero, A.; Caballero-Méndez, C.; Espino-Ruano, A.; Couce-Montero, L.; Jiménez-Alvarado, D.; Castro, J. J. Length-weight relationships of 15 mesopelagic shrimp species caught during exploratory surveys off the Canary Islands (central eastern Atlantic). Scientia Marina 2024, 88, e081. [Google Scholar] [CrossRef]
- Pinkas, L.; Oliphant, M.S.; Iverson, I.L.K. Food habits of albacore, bluefin tuna and bonito in California waters. California Department of Fish and Game’s Fish Bulletin 1971, 152, 1–105. [Google Scholar]
- Hyslop, E.J. Stomach content analysis: A review of methods and their application. Journal of Fish Biology 1980, 17, 411–422. [Google Scholar] [CrossRef]
- Hacunda, J.S. Trophic relationships among demersal fishes in a coastal area of the Gulf of Maine. Fishery Bulletin 1981, 79, 775–788. [Google Scholar]
- Costello, M.J. Predator feeding strategy and prey importance: A new graphical analysis. Journal of Fish Biology 1990, 36, 261–263. [Google Scholar] [CrossRef]
- Amundsen, P.A.; Gabler, H.M.; Staldvik, F.J. A new approach to graphical analysis of feeding strategy from stomach contents data modification of the Costello (1990) method. J. Fish. Biol. 1996, 48, 607–614. [Google Scholar] [CrossRef]
- Pauly, D.; Christensen, V. Primary production required to sustain global fisheries. Nature 1995, 374, 255–257. [Google Scholar] [CrossRef]
- Pauly, D.; Trites, A.W.; Capuli, E.V.; Christensen, V. Diet composition and trophic levels of marine mammals. ICES J. Mar. Sci 1998, 55, 467–481. [Google Scholar] [CrossRef]
- Pauly, D.; Froese, R.; Sa-a, P.; Palomares, M.; Christensen, V.; Rius, J. TrophLab Manual. ICLARM 2000, Manila, Philippines.
- Pauly, D.; Palomares, M.L. Approaches for dealing with three sources of bias when studying the fishing down marine food web phenomenon. CIESM Work. Ser. 2000, 12, 61–66. [Google Scholar]
- Pandian, T.J. Intake, digestion, absorption, and conversion of food in the fishes Megalops cyprinoides and Ophiocephalus striatus. Mar. Biol. 1967, 1, 16–32. [Google Scholar] [CrossRef]
- Pandian, T.J. Transformation of food in the fish Megalops cyprinoides. I. Influence of quality of food. Mar Biol 1967, 1, 60–64. [Google Scholar] [CrossRef]
- Cartes, J. E. Diets of deep-water pandalid shrimps on the Western Mediterranean slope. Marine Ecology Progress Series 1993, 96, 49–61. [Google Scholar] [CrossRef]
- Cartes, J. E. Feeding habits of pasiphaeid shrimps close to the bottom on the western Mediterranean slope. Marine Biology 1993, 117, 459–468. [Google Scholar] [CrossRef]
- Mead, G.W.; Earle, S.A. Notes on the natural history of snipe eels. Proc. Calif. Acad. Sci. 1970, 99–103. [Google Scholar]
- Smith, D.G.; Tighe, K.A. Snipe eels. Family Nemichthyidae. In: Collette B.B., Klein-MacPhee G (eds) Fishes of the Gulf of Maine. 3rd edn. Smithsonian Institute Press, Washington, DC 2002, pp. 100–101.
Figure 1.
Study area in the Central Mediterranean Sea.
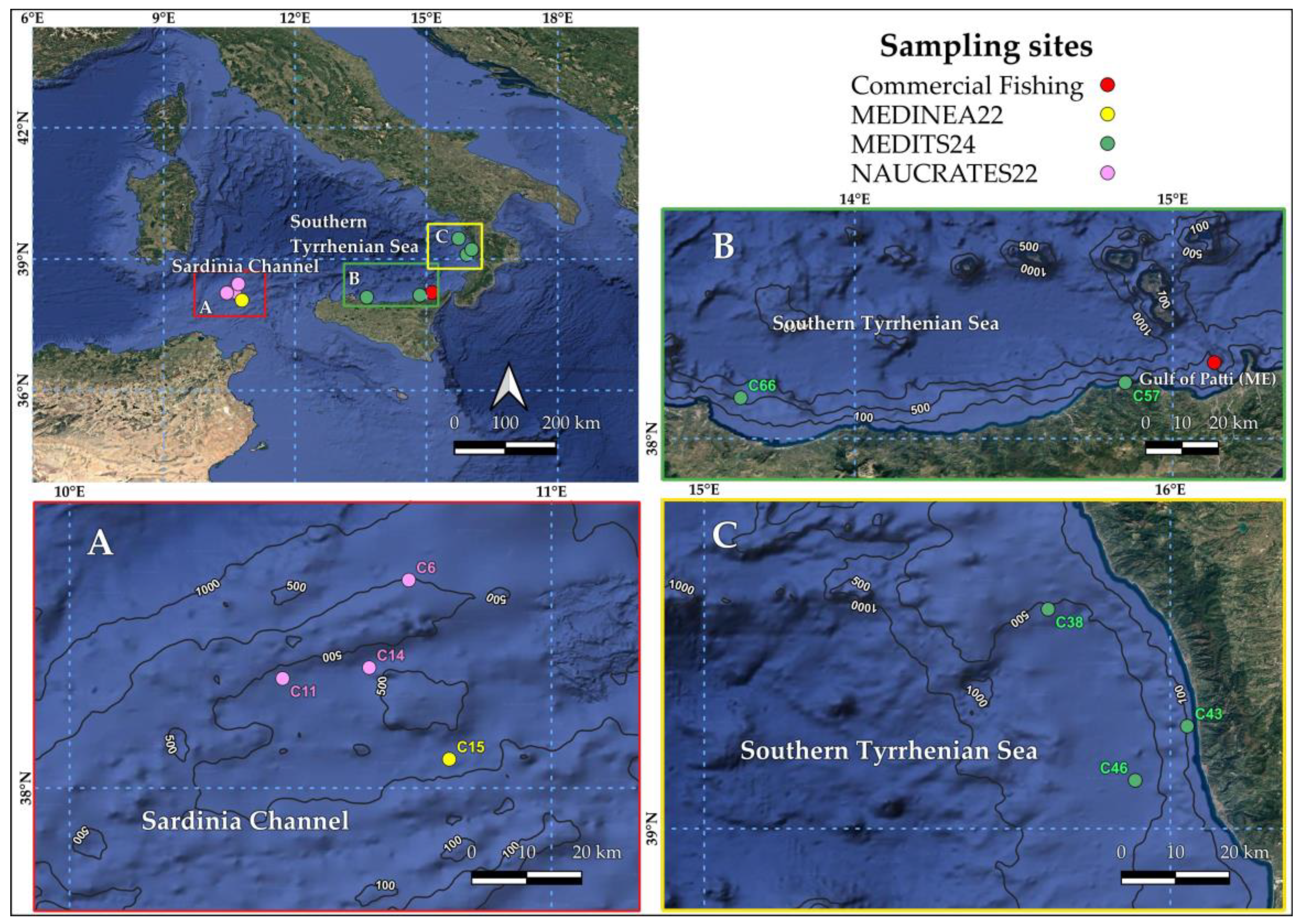
Figure 2.
Eye-Gill cover Distance (EGD).
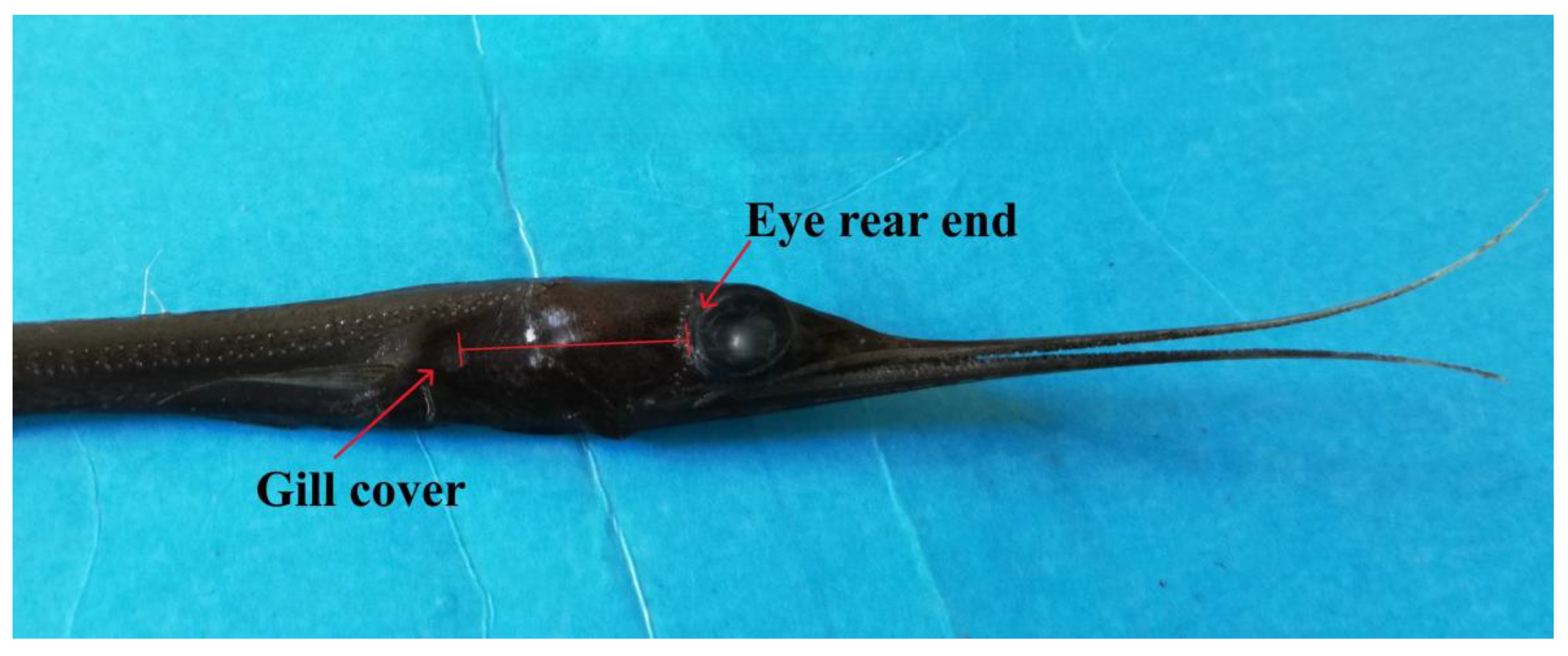
Figure 3.
Size classes and seasons of Nemichthys scolopaceus sampled in the study area.
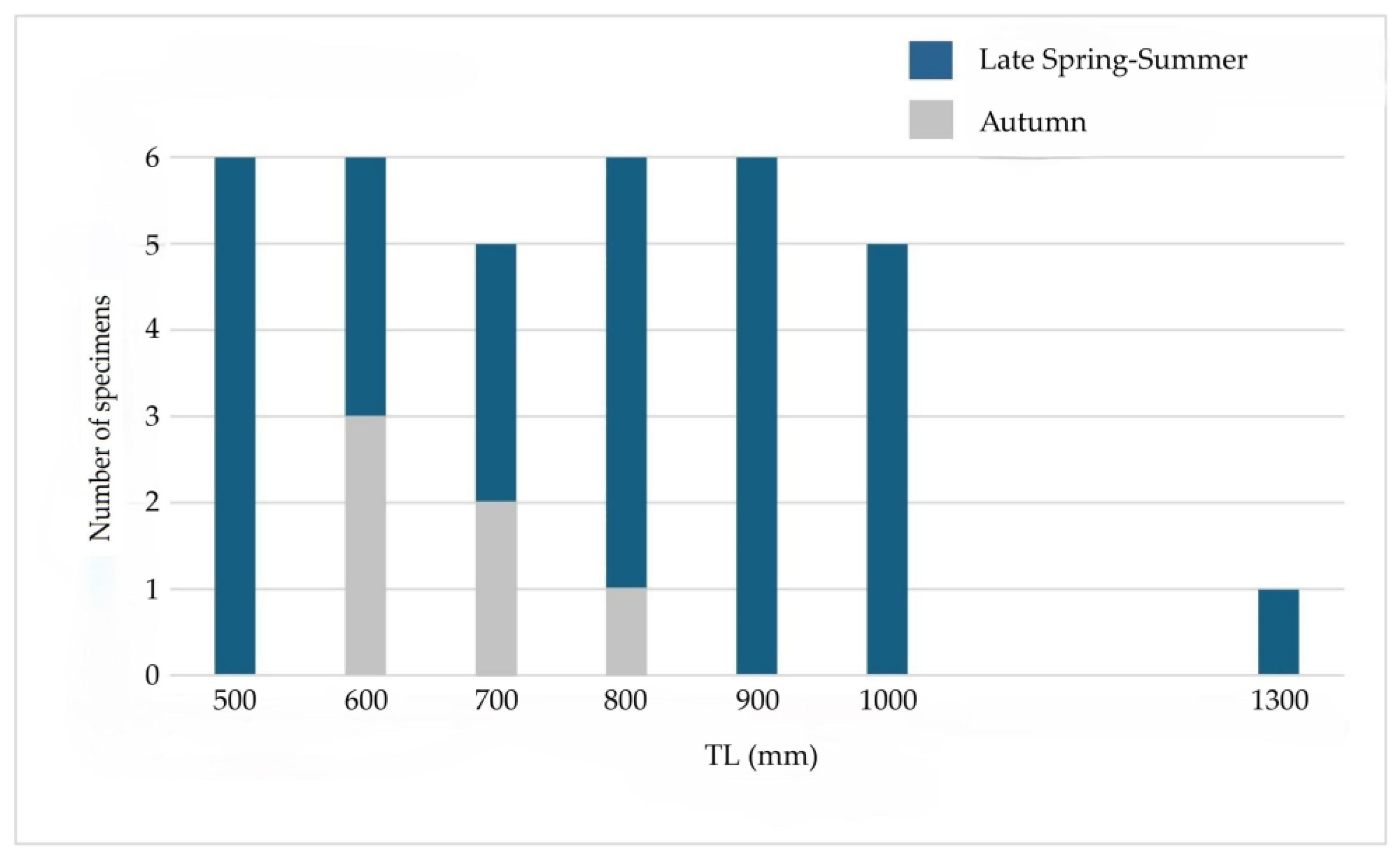
Figure 4.
Relationship between TL (Total Length), EGD (Eye Gill cover Distance) and WW (Wet Weight) of N. scolopaceus examined specimens.
Figure 4.
Relationship between TL (Total Length), EGD (Eye Gill cover Distance) and WW (Wet Weight) of N. scolopaceus examined specimens.
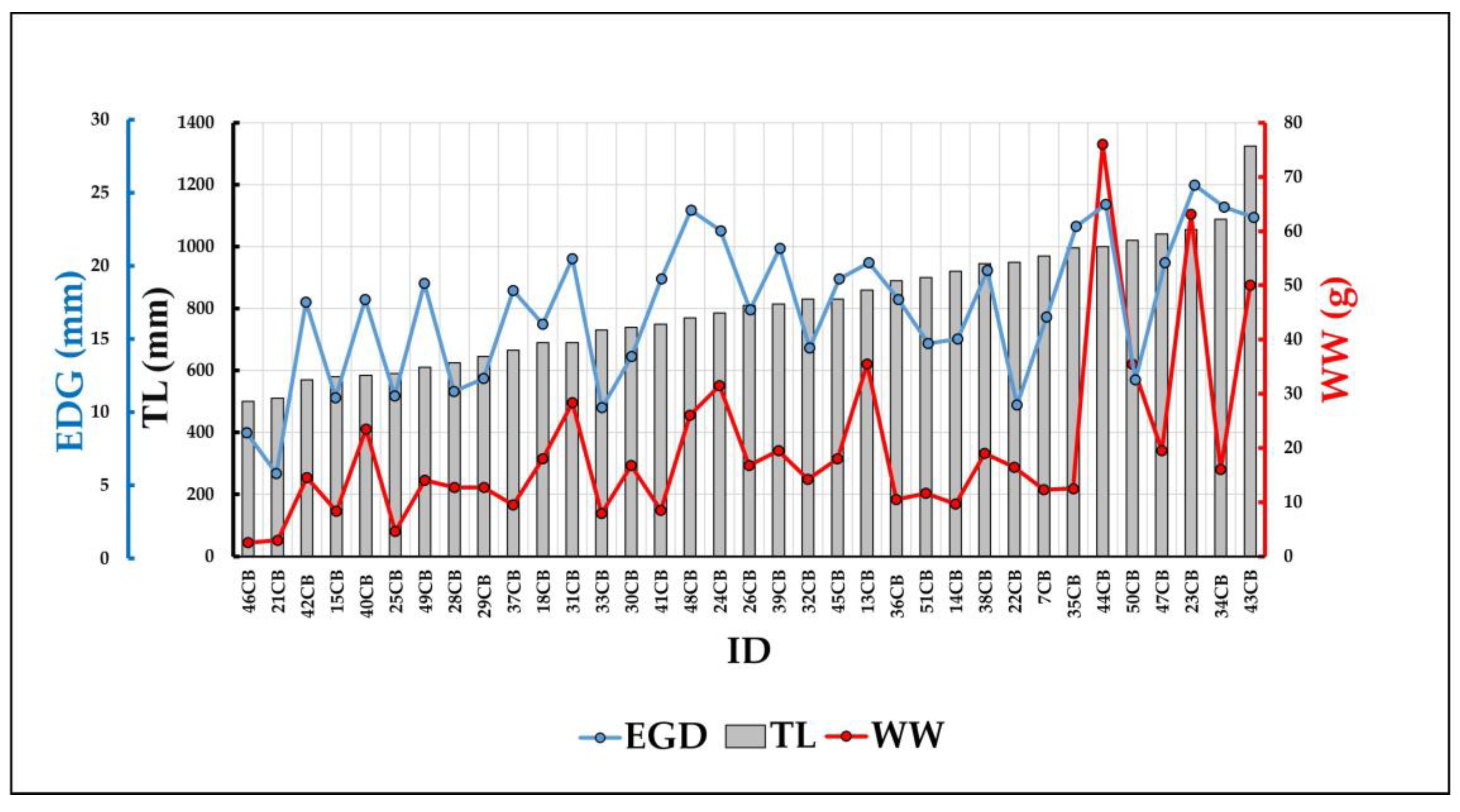
Figure 5.
Number of preys found in stomachs of Nemichthys scolopaceus in the study area.
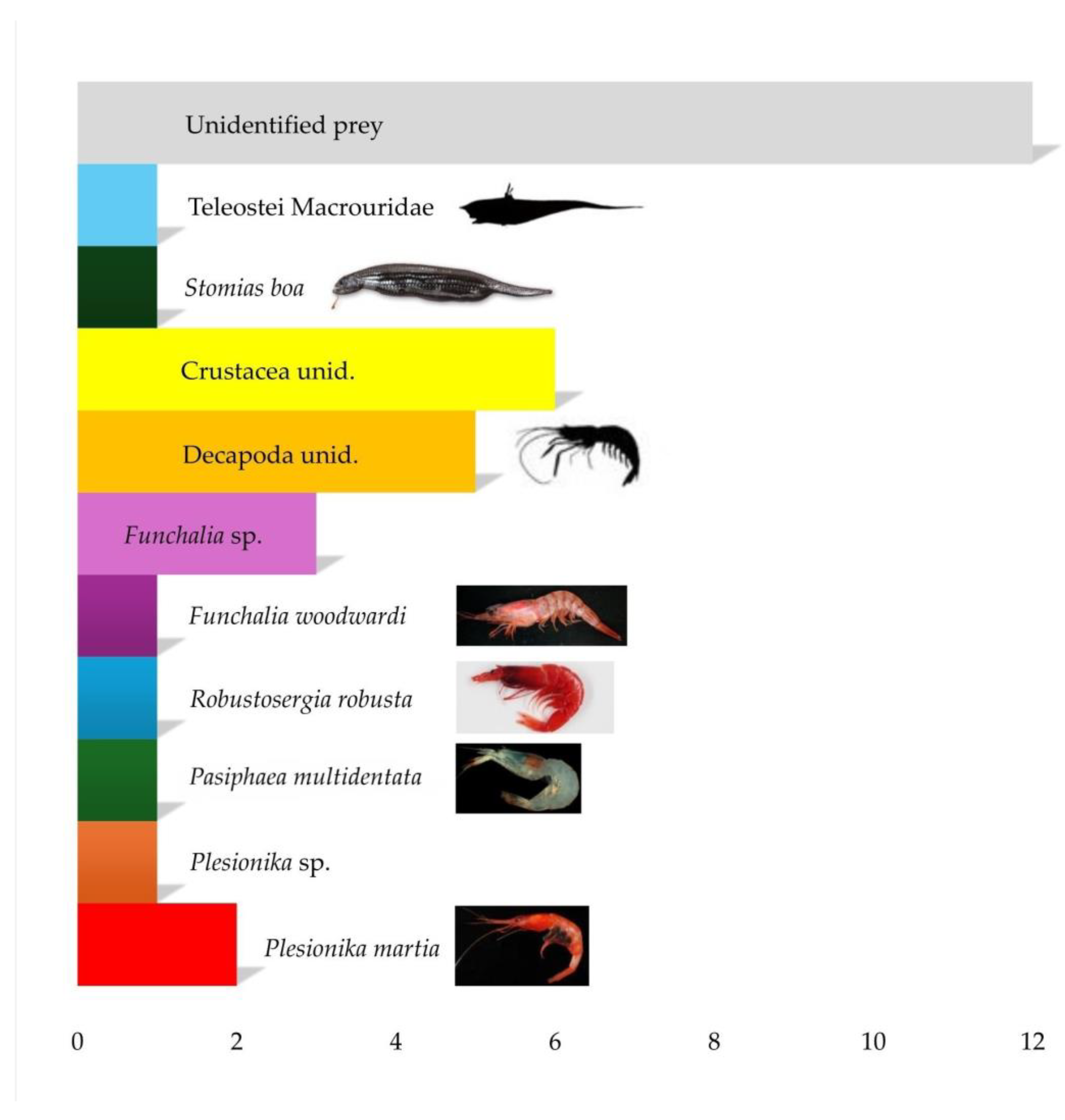
Figure 6.
The graphic method of Costello (1990), modified by Amundsen et al. (1996), which plots on a two-dimensional graph the specific abundance, for number (a), and for weight (b) of each prey category with respect to the frequency of finding of the prey itself in the stomach contents. BPC: between-phenotype component; WPC: within-phenotype component. FUNC: Funchalia sp., FUNCWOO: Funchalia woodwardii, PASIMUL: Pasiphaea multidentata, PLES: Plesionika sp., PLESMAR: Plesionika martia, ROBUROB: Robustosergia robusta, STOMBOA: Stomias boa.
Figure 6.
The graphic method of Costello (1990), modified by Amundsen et al. (1996), which plots on a two-dimensional graph the specific abundance, for number (a), and for weight (b) of each prey category with respect to the frequency of finding of the prey itself in the stomach contents. BPC: between-phenotype component; WPC: within-phenotype component. FUNC: Funchalia sp., FUNCWOO: Funchalia woodwardii, PASIMUL: Pasiphaea multidentata, PLES: Plesionika sp., PLESMAR: Plesionika martia, ROBUROB: Robustosergia robusta, STOMBOA: Stomias boa.
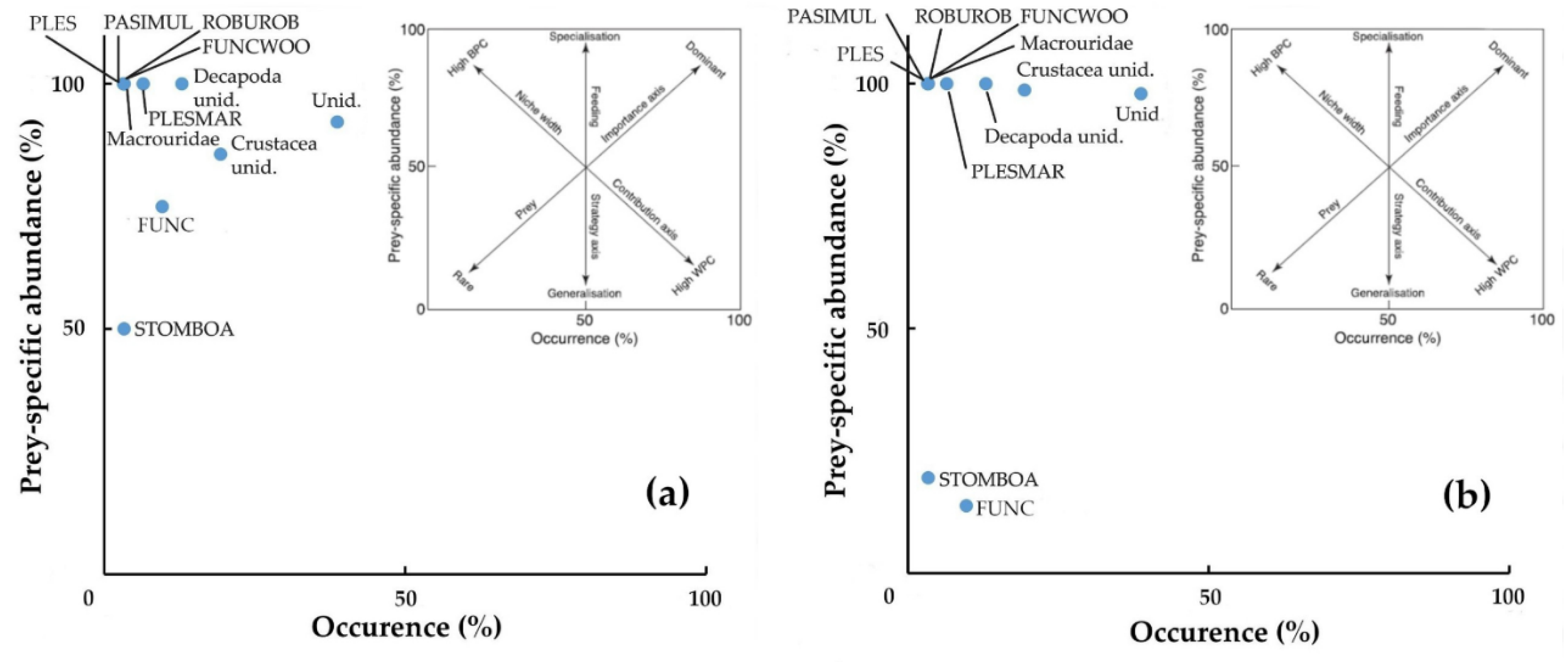
Figure 7.
Percentage in number (a), and in weight (b) of preys of Nemichthys scolopaceus, considering them to group level, regardless of single species, genera and family.
Figure 7.
Percentage in number (a), and in weight (b) of preys of Nemichthys scolopaceus, considering them to group level, regardless of single species, genera and family.
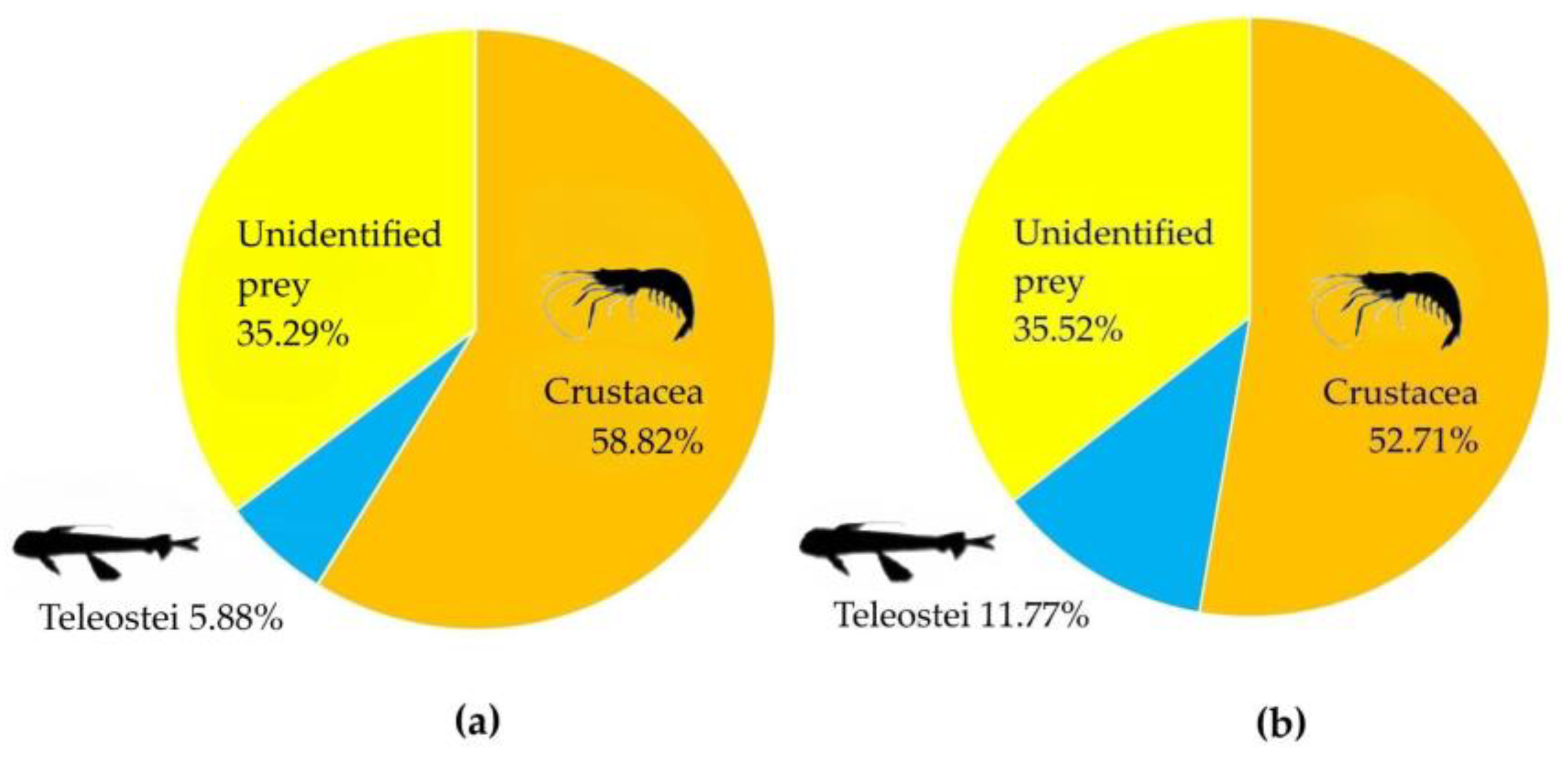

Table 1.
Collection of metadata for Nemicthys scolopaceus samples from all the cruises.
| Campaign | Fishing Area | Local Date | Lat. N | Long. E | Sampling depth (m) | Local time (Start-Finish) |
Specimens caught (per time) |
Specimen ID number |
| Commercial fishing | Gulf of Patti (ME) | May-June 2022 | 38°14.346' | 15°7.818' | 300-700 | - | 11 | 7CB, 13CB, 14CB, 15CB, 18CB, 21CB, 22CB, 23CB, 24CB, 25CB, 26CB |
| May-June 2023 | - | 12 | 34CB, 35CB, 36CB, 37CB, 38CB, 39CB, 40CB, 41CB, 42CB, 47CB, 48CB, 49CB | |||||
| NAUCRATES22 | Pa-Ca Canyon | 12/10/2022 | 38°25.91' | 10°42.24' | 309-305 | 12:53-17:35 | 1 | 28CB |
| 19/10/2022 | 38°13.65' | 10°37.32' | 323-358 | 22:20-1:47 | 1 | 31CB | ||
| 14/10/2022 | 38°15.00' | 10°37.32' | 370-311 | 12:30-16:33 | 2 | 32CB, 33CB | ||
| MEDINEA22 | Pa-Ca Canyon | 14/10/2022 | 38°03.60' | 10°47.28' | 332-349 | 19:07-23:07 | 2 | 29CB, 30CB |
| MEDITS24 | GSA10 | 21/06/2024 | 39° 28.24' | 15° 44.27' | 583-626 | 07:13-08:13 | 2 | 44CB, 45CB |
| 22/06/2024 | 39° 13.18' | 16° 02.19' | 83-86 | 06:51-07:21 | 1 | 50CB | ||
| 22/06/2024 | 39° 06.21' | 15° 55.49' | 634-619 | 14:46-15:46 | 1 | 46CB | ||
| 25/06/2024 | 38° 10.56' | 14° 51.02' | 26-18 | 12:00-12:30 | 1 | 51CB | ||
| 26/06/2024 | 38° 07.70' | 13° 38.44' | 568-529 | 16:15-17:15 | 1 | 43CB |
Table 2.
Preys of Nemichthys scolopaceus with trophic indexes.
| Phylum/ Subphylum/ Class/Order |
Family | Species/Taxa | %Ni | %Wi | %Fi | %IRIi |
|---|---|---|---|---|---|---|
| ARTROPODA | ||||||
| CRUSTACEA | ||||||
| Malacostraca | ||||||
| Decapoda | Pandalidae |
Plesionika martia (A. Milne-Edwards, 1883) |
5.88 | 9.31 | 6.45 | 10.46 |
| Plesionika sp. | 2.94 | 8.33 | 3.23 | 15.19 | ||
| Pasiphaeidae | Pasiphea multidentata Esmark, 1866 | 2.94 | 4.65 |
3.23 | 10.47 |
|
| Sergestidae | Robustosergia robusta (Smith, 1882) | 2.94 | 0.78 |
3.33 | 5.53 |
|
| Peneidae | Funchalia woodwardi Johnson, 1868 | 2.94 | 0.23 |
3.33 | 4.82 |
|
| Funchalia sp. | 8.82 | 0.43 | 9.68 | 4.70 | ||
| Decapoda unid. | 14.71 | 13.18 | 12.90 | 8.10 | ||
| Crustacea unid. | 17.65 | 15.81 | 19.35 | 7.45 | ||
| CHORDATA | ||||||
| VERTEBRATA | ||||||
| Teleostei | ||||||
| Stomiiformes | Stomiidae |
Stomias boa (Risso, 1810) |
2.94 | 0.72 |
3.33 | 5.44 |
| Gadiformes | Macrouridae | Macrouridae unid. | 2.94 | 11.05 | 3.33 | 18.66 |
| Unidentified | 35.29 | 35.42 | 40.00 | 9.17 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



