
Submitted:
28 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
Pipefishes (Family Syngnathidae) are representative fishes in many European coastal areas asso-ciated with vegetal assemblages and other shallow habitats. The polyandrous pipefishes Syngnathus abaster and S. acus co-occur in Metruk Tuzla (Bargilya Wetland, Aegean Sea, Turkey), a protected but threatened wetland due to anthropogenic disturbances. Both species were sam-pled during the breeding season (April-2018 and June-2019) for the assessment of growth and trophic features revealed from stable isotope profiles (δ13C and δ15N) and gut content. Both species showed positive allometric growth, with similar length-weight relationships. The isotope approach indicated that both species occupy similar trophic niches, with mean δ13C and δ15N values of -17.0 ± 1.2 ‰ for δ13C and 13.2 ± 0.6 ‰ for δ15N in S. abaster, and -17.3 ± 1.2 ‰ for δ13C and 13.3 ± 0.7 ‰ for δ15N in S. acus. The niche area for all sex types (undifferentiated specimens, males and females) in S. acus was slightly broader than in S. abaster. The simulation analysis of the trophic position (TP) in both species agreed with the isotopic assessment (TP = 3.67 ± 0.61 in S. abaster and 3.46 ± 0.66 in S. acus). Despite the high trophic resemblance between S. abaster and S. acus, the gut content assessment revealed the large contribution of copepods (especially Calanipeda aquaedulcis) to the dietary regime in both pipefishes. However, while the diet in S. abaster seemed to be almost monospecific, cladocerans were also an important resource in S. acus. Hence, we suggest that potential inter-specific competition for dietary resources would be minimized likely due to the occupancy of different microhabitats, a hypothesis that needs to be confirmed by further research.
Keywords:
rophic
; niche
; diet
; syngnathidae
; pipefish
; Syngnathus abaster
; Syngnathus acus
; aegean sea
1. Introduction
Although wetlands only comprise 6% of the Earth’s surface, they are important ecosystems with numerous tasks, in addition to maintaining biodiversity. As such, many wetlands are often recognized as important conservation or restoration targets [1]. Hydrology, water conditions and oxygen availability in wetlands are largely variable over time, depending on seasonal changes in the sources of inflows and outflows, among other factors. However, wetlands are one of the most productive ecosystems in the world [2].
Turkey is a very rich country considering lakes and wetlands. However, a large area of Turkish wetlands has deteriorated or been irreversibly lost in the last decades. This decline is due to increased anthropogenic disturbances and it will likely accelerate a negative scenario including climate change [3,4,5].
Boğaziçi Lagoon (Turkey), also known as Bargylia Tuzla coastal Wetland (BTW), connects to the heavily damaged Güllük Bay (Aegean Sea) by a submarine strait [6]. It is one of the last lagoons in Turkey to survive and a permanent shallow marine lagoon less than 6 m deep at low tide, which includes sea bays and straits and intertidal salt flats [6].
BTW was declared as "Metruk Tuzlası Wetland Protection Area" by the Republic of Turkey (Ministry of Environment and Forestry, General Directorate of Nature Conservation and National Parks) in 2004 [7]. The region was then taken under the protection area by the National Wetland Commission and considered a wetland candidate for the Ramsar Convention [8], an international treaty aimed at conserving wetlands [9,10].
BTW is a shallow seawater area, with less than 6 m in depth, surrounded by several bays and inlets. The wetland is a fertile and sensitive oligo-hypersaline area, mainly fed by rainfall, streams, and karstic brackish springs. It is one of the most important national wetlands of Turkey in terms of bird species and biodiversity, covering an important part of the Güllük coastal wetland (GCWS), known for its hypersalinity. The substrate is characterized by silt-sandy and muddy sediments [11].
BTW, as part of Güllük Bay, is under threat due to fishing and aquaculture activities, agriculture runoff, domestic waste pollution, port activities and tourism, including a “Tourism City” project approved by the Ministry of Environment and Urbanization. Güllük Bay contributes about 70% of total Turkish aquaculture production, mainly for the production of seabass and seabream [12,13]. Those factors put the wetland under threat, as the impact will be far above the carrying capacity of the area, disrupting irreversibly the ecological balance [13].
Wetlands provide nurseries for fish and other organisms, hosting important fish communities that support small local fishing. Thus, economically important species such as Mugil cephalus, Liza aurata, Dicentrarchus labrax, Sparus aurata, Cyprinus carpio, Anguilla anguilla and Solea solea, among others, have been reported in Güllük Delta [8,14-16]. Güllük Delta is also inhabited by fish juveniles or non-economic small fish species that are prey for economical fishes [15]. In addition, Güllük Delta is also the habitat for other non-commercial fish species such as Atherina boyeri, Aphanius fasciatus, Neogobius fluviatilis, Syngnathus spp. (present study) and Chelon spp. [16]. Some of these species are strongly associated with vegetal assemblages [17], including Posidonia oceanica and Zostera marina meadows [18]. That is the case of pipefishes, a group of teleosts included in the Family Syngnathidae together with seahorses and seadragons.
Although European pipefishes are classified as Data Deficient or Least Concern by IUCN [19], many species are considered vulnerable from an ecological and biological point of view. There are two main nearshore representative pipefishes in BTW: the black-striped pipefish Syngnathus abaster Risso, 1827 and the greater pipefish Syngnathus acus Linnaeus, 1758 [20]. Both species are considered Least Concern by IUCN but their population trends are currently unknown [21,22]. The euryhaline black-striped pipefish occupies a wide variety of habitats, and there are no known threats at a large scale. It is a freshwater and estuarine species occurring in coastal waters and lower reaches of rivers from the Caspian Sea to the Bay of Biscay, as well as several rivers in northern Europe and Russia. The species has been progressively colonizing pure freshwater ecosystems in Eastern Europe [23-25]. Syngnathus acus shows a wider distribution as it has been identified in waters from the Baltic Sea to the Black Sea, and in other southern areas (Morocco, Senegal, Gambia, and South Africa). It is usually found on vegetated substrates from the intertidal zone and down to 20 meters, and on sandy or muddy areas in the mouths of estuaries [20].
Studies focusing on biological or ecological features in S. abaster and S. acus are limited, especially in S. acus. Some of them provided information on morphometrics and meristics [26-30], while others evaluated distribution patterns [24,31,32], early life history [33], reproduction traits [34,35] or trophic patterns [34,36-40] in local populations. While many European pipefish species are known to be associated with macroalgal assemblages or seagrass habitats and co-occur in the same meadows, the assessment of potential interspecific competition for resources has been poorly investigated. As in most syngnathids, it is known that pipefishes commonly forage on small prey, such as crustaceans [38,41]. S. abaster and S. acus are secondary consumers preferring amphipods, copepods, chironomids or cladocerans as well as carideans and isopods, depending on the species, size, developmental stage, region and season [25,34,39,40].
Based on their distinct morphology, mobility, gape biometrics and foraging skills, we investigated whether the co-occurring pipefishes S. abaster and S. acus might compete for food resources on a microhabitat scale. In this regard, it is important to notice that the larger mouth width in S. abaster compared to S. acus might play an important role in prey selection [27,30,40]. Accordingly, the main aim of this study was to extend the understanding of syngnathid ecology by describing, for the first time from an isotopic approach, the trophic niche characteristics and potential competition for resources in two sympatric pipefishes in the Aegean Sea.
2. Materials and Methods
2.1. Study site and fish collection
The study was carried out in Metruk Tuzla, positioned in Boğaziçi Tuzla lagoon (SW Turkey) (37°11'27'' N 27°35'20" E), as part of Bargilya Tuzla Wetland (BTW), classified as a Wetland of International Importance according to Ramsar criteria [44]. The lagoon is a shallow coastal area (maximum depth: 1.5 m; average depth: 45 cm) located in the southern part of the Bargilya wetland (Figure 1) and separated by the road built on the embankment from Bargil Cove, which is directly connected to Güllük Bay. The water level decreases with summer and fall evaporation, while the inward-flowing seawater from these channels prevents from drying up [17].
Specimens of black-striped pipefish Syngnathus abaster Risso, 1827 and greater pipefish S. acus Linnaeus, 1758 (Figure 2) were collected in Metruk Tuzla in April-2018 and June-2019, corresponding to the breeding season [34,36,43-45], by using a shore seine net (10 m long; 2 m height; 1.2 x 2 mm mesh size) (Figure 1 and Figure 2) operated several times for 5 minutes. At the end of each sampling operation, the pipefishes were removed from the net, manually sorted, anaesthetized with tricaine methane-sulfonate (MS-222; Sigma-Aldrich, Germany; 1 g ml-1), transferred to plastic containers containing 70% ethanol and transported to the laboratory of the Department of Hydrobiology at Ege University (Turkey). Since the syngnathids have a relatively undifferentiated gastrointestinal tract, stretched gut length of the pipefish specimens was measured (± 0,01mm) between the oesophagus and the intestine using a digital calliper.
Temperature, salinity, pH and oxygen saturation were also recorded on the sampling dates (YSI Pro Plus Multiparameter, YSI Inc., Yellow Springs, USA).
2.2. Fish sampling and analyses
The collected fishes were analyzed for wet weight, standard length (SL), sex and maturity. The reproductive condition was recorded, considering the trunk shape in females (i.e., full gonads with hydrated eggs) and pregnancy in males (i.e., brood pouches on the tail carrying fertilized eggs/embryos) [34]. Individuals that did not show those characteristics were considered undifferentiated, the most corresponding to preadults or immature fishes.
Since pipefishes do not have a fully differentiated tubular gastrointestinal tract, the entire guts were dissected out, weighed and identified under a stereomicroscope to the lowest possible taxonomic level. The fish with empty and full guts were determined. Gut contents’ frequency (F%) and numerical presence (N%) of prey were analyzed [46].
Fish dorsal fins were removed, rinsed with distilled water, dried in an oven (50°C), sent to the Institute of Marine Research (IIM-CSIC, Spain) and maintained frozen (-20°C) for further analysis of stable isotopes (SIA) (δ13C and δ15N), total C and total N.
2.3. Stable isotopes
Dried fin samples were homogenized with a pestle and mortar until fine powder, transferred (aliquot of 1-2 mg dry weight) to tin capsules, and sent to Servizos de Apoio á Investigación (SAI) of the University of A Coruña (Spain). Fin samples were analyzed by continuous-flow isotope ratio mass spectrometry using a FlashEA1112 elemental analyser (Thermo Finnigan, Italy) coupled to a DeltaV Advantage (Thermo Scientific) through a ConfloIV interface. Carbon and nitrogen stable isotope abundance was expressed as permil (‰) relative to VPDB (Vienna Pee Dee Belemnite) and Atmospheric Air, according to the following equation:
where X is 13C or 15N and R is the corresponding ratio of 13C/12C or 15N/14N. As part of an analytical batch run, a set of international reference materials for δ15N (USGS 40 (-4.52‰), USGS41a (+47.55‰), (IAEA-N-1 (+0.4‰), IAEA-N-2 (+20.3‰) y USGS-25 (-30.4‰) and δ13C (USGS 40 (-26.39‰), USGS41a (+36.55‰) NBS 22 (-30.031‰) e USGS 24 (-16.049‰)) were analyzed. The precision (standard deviation) for the analysis of δ13C and δ15N of the laboratory standard (acetanilide) was ± 0.15‰ (1-sigma, n=10). Standards were run every 10 biological samples. The isotopic analysis procedure fulfils the requirements of the ISO 9001 standard. The laboratory is submitted to annual inter-calibration exercises (Forensic isotope ratio mass spectrometry scheme – FIRMS, LGC Standards, UK).
δX = (Rsample / Rreference) - 1,
Due to potential alterations in δ13C values and, to a lesser extent, in δ15N values, normalized for lipids is highly recommended in tissues with high lipid content (i.e., C:N > 3.56) [47-51]. In the present study, C:N ratios in fin samples of ethanol-preserved specimens ranged from 3.07 to 3.46 (3.07-3.31 in S. abaster; 3.11-3.46 in S. acus). Previous studies in syngnathids demonstrated that the preservation in ethanol does not affect δ15N values in fins [52,53]. However, δ13C values might result slightly affected [53]. Consequently, δ13C data were normalised using the regression equation provided specifically for dorsal fins in syngnathids [53].
2.4. Data treatment
Data and statistical analyses were conducted in R v.3.6.1 [54]. Data normality and heteroscedasticity were checked with Shapiro-Wilk normality test and Levene’s test, respectively (stats v3.6.2 R package) [55].
One- or two-way analyses of variance (ANOVA) (unbalanced design) were applied for mean comparison. When significant, a Tukey HSD posthoc test was applied to check the significance of differences between pairs of group means.
Length-weight relationship (LWR) was estimated as follows [56]:
where W is the wet weight of fish (g), TL is the total length of fish (cm), a is the intercept of the regression, and b is the coefficient expressing the relationship between length and weight. Using the linear regression of the log-transformed equation:
W = aTLb,
log (W) = log (a) + b log (TL),
The parameters a and b were calculated with a representing the intercept and b the slope of the relationship. The increase in weight is isometric when b=3. When the value of b is other than 3, the weight increase is allometric (positive if b>3, negative if b<3) [57]. A positive allometry denotes that fish are heavier reflecting optimum conditions for growth.
The statistical significance of LWR when p < 0.05 was analysed with ANOVA. The student’s t-test was used to verify if the slopes (b) were significantly different from 3.
When comparing both species, we pooled the isotopic data from both sampling periods, as the sample size for males and females was not large enough to analyse both periods separately. The packages SIBER v.2.1.4 [58] and NicheRover v.1.1.0 [59] were used to assess differences in the trophic niche features between S. abaster and S. acus. The niche region was defined as the joint probability density function of the multidimensional niche indicators at a probability alpha of 95%. Uncertainty was accounted for in a Bayesian framework. The analysis provides directional estimates of niche overlap, accounts for species-specific distributions in multivariate niche space, and produces unique and consistent bivariate projections of the multivariate niche region [60].
Total convex hull areas (TA) and core trophic niche breadths were estimated using SIBER (Stable Isotope Bayesian Ellipses) while correcting for variable sample sizes (SEAc). Total trophic overlap values for 95% TA were estimated using nicheROVER, a method that is insensitive to sample size and incorporates statistical uncertainty using Bayesian methods [60].
Differences in gut content across species and seasons were assessed by t-test and one-way ANOVA.
Significance levels were set at p < 0.05 and the results are given as mean ± standard deviation. The graphics were constructed using ggplot2 v3.3.0 in R [61].
2.5. Bioethics
Animal sampling and manipulation practices were conducted in compliance with all bioethics standards on animal experimentation (Burdur Mehmet Akif Ersoy University, Animal Experiments Local Ethics Committee, Decision number: 1148)
3. Results
3.1. Abiotic data
Temperature and salinity recorded on the sampling dates were 14°C and 15 in spring and 20°C and 21 in summer, respectively. Oxygen saturation and pH levels were 6.8 mg L-1 in spring and 6.3 mg L-1 in summer, and 8.12 in spring and 8.37 in summer, respectively.
3.2. Fish populations
The general information on the pipefish collected is provided in Figure S1 and Tables S1 and S2.
Average TL was 5.9 ± 1.6 cm in S. abaster (n: 51; range: 3.3 – 10.9 cm) and 6.3 ± 1.9 cm in S. acus (n: 78; range: 3.1 – 12.0 cm) (Table 1; Figure S2). Mean individual weight was 0.115 ± 0.118 g in S. abaster (range: 0.015 – 0.669 g) and 0.124 ± 0.125 g in S. acus (range: 0.010 – 0.610 g).
Mean TL in both species did not differ significantly (Kruskal-Wallis test; Χ2(1) = 1.31; p = 0.251) but TL across sex groups were significantly different (undifferentiated < females < males) (K-W; Χ2(2) = 1.31; p < 0.0001). The minimum recorded TL in mature females was 5.4 cm in S. abaster and 6.1 cm in S. acus, whereas pregnant males were collected with a minimum TL of 7.7 cm in S. abaster and 6.3 cm in S. acus (Table 1).
3.3. Gut content
Only 7.8% of S. abaster individuals showed empty guts, whereas the percentage increased to 22.8% in S. acus. In S. abaster, the dominant prey group was the pseudodiaptomid copepod Calanipeda aquaedulcis, both in numerical (%N; 71.2%) and frequency of occurrence (%F = 42.6%). The contribution of unidentified both copepods and fish eggs was also important in the diet of the species. In S. acus, C. aquaedulcis was also dominant together with cladocera (pieces) both in %N (47.5% and 45.7 %, respectively) and %F (35.3% and 40%, respectively) (Table 3; Figure S3). The inter-specific difference for %N was not significant (t-value = -0.019, d.f. =10, p>0.05).
Regarding seasonal changes in gut contents (Table 3), the most noticeable findings were recorded in summer for S. acus, with a numerical increase in the cyclopoida copepod Halicyclops biscupidatus (%N = 54.2%). Cladoceran pieces also increased their frequency in summer (%F = 35.0%). However, inter-seasonal differences in the numerical prey content within each species were not significant (p>0.05).
3.4. Isotopic profiles
Globally, the isotopic profiles were highly similar in both species (two-way ANOVA, type III; p>0.05) and sex (two-way ANOVA, type III; p>0.05), with mean δ13C and δ15N values of -17.2 ± 1.2 ‰ and 13.3 ± 0.7 ‰, respectively (Table 4; Figure S4). However, the average δ15N value in females was significantly lower (13.1 ± 0.7 ‰) than in undifferentiated specimens (13.4 ± 0.6 ‰) (Tukey HSD test; Padj =0.036).
Mean isotopic values were -17.0 ± 1.2 ‰ for δ13C and 13.2 ± 0.6 ‰ for δ15N in S. abaster, and -17.3 ± 1.2 ‰ for δ13C and 13.3 ± 0.7 ‰ for δ15N in S. acus (Table 4).
Spearman’s correlations indicated that δ15N values were negatively correlated with TL in both species (S. abaster: r(51) = -0.482, p <0.001; S. acus: r(78) = -0.458, p <0.001), whereas the correlation between δ13C and TL was statistically not significant (Figure S1). The relationship between δ15N and Tl was modelled by linear regression. The resulting coefficients b in the models were significant, especially for the greater pipefish (b = -0.0123, P = 0.022 in S. abaster; b = -0.0125, p = 0.007 in S. acus), denoting a small progressive decrease in δ15N values with fish growth.
3.5. Trophic structure and niche overlap
The average niche area, SEAc, in S. abaster (2.06) was slightly smaller than in S. acus (2.51) in all sex types (undifferentiated specimens, males and females) (Figure 4 and Figure S6). The smallest and larger niche areas occurred in females and males, respectively (Table 5).
The isotopic niche overlap analysis with nicheROVER package revealed a high probability (96%) that a specimen of S. abaster will be included within the niche of S. acus. The probability decreased to 89% in the reverse situation. Considering inter- and intra-specific niche overlaps concerning the sexual condition (Table 6), the lower posterior probabilities regarding niche overlap estimates occurred in females and undifferentiated individuals of S. abaster, especially compared to males of the same species. Regarding S. acus, the probabilities that an individual will be found within the trophic niche of the species were rather high (73 - 95%).
4. Discussion
There is evidence that the size of the studied pipefishes S. abaster and S. acus depends on the geographical region considered (see the review by Gurkan and Innal [30]. The size of the specimens from Metruk Tuzla (5.9 ± 1.6 cm TL in S. abaster; 6.3 ± 1.9 cm TL in S. acus) was similar to those in the nearby Lake Bafa (Turkey) [30] but considerably lower than S. acus from Cíes Archipelago (NW Spain) (34.4 ± 6.8 cm SL) [34]. Both species showed comparable LWR and positive allometry, similar to the findings in other regions [28,34,62].
Pipefishes are ovoviviparous teleosts with parental care, with females depositing eggs in a specialized brooding structure of males, located on the tail. In the majority of European pipefishes the breeding extends from February to September, depending on the region and the species [34,36,63,64]. In addition, some species (e.g. S. acus) show temporal migratory events to breeding areas [34]. In the present study, the pipefishes were collected during the breeding season. In Metruk Tuzla, the minimum-recorded size in mature males and females was similar in both species (8.4 – 8.8 cm in males; 7.2 – 7.8 cm in females). This finding agrees with others on the Greek coastline, where 50% maturity of males and females of S. abaster measured more than 7 cm in TL [65]. However, the largest mature S. acus specimens collected in Metruk Tuzla were dramatically smaller than in Cíes Archipelago (19.1 - 47.4 cm in ovigerous females; 24.0 – 49.8 cm in pregnant males), where the largest specimen ever observed for the species has been recorder (49.8 cm). Hence, at least in the greater pipefish, the size at first maturity also seems to be geographically dependent. Contrarily, the ranges in size for S. abaster seem to be rather conservative and not related to the location.
Despite the limited number of males collected, we observed a significant relationship between male size and male brood pouch in both species (Figure S6), which is in agreement with the length-dependent reproductive fitness (i.e., with larger males accommodating more offspring) hypothesized by Silva et al. [35] in S. abaster.
Concerning their habitat, most native European pipefish species usually prefer shallow marine coastal habitats but a few exhibits a progressive colonization of freshwater habitats. The black-striped pipefish S. abaster is a euryhaline fish, found among detritus or vegetation, over sand or mud in marine, brackish and, more recently, in freshwater environments [23-25]. The greater pipefish S. acus is common in shallow and, less frequently, in brackish waters and mouths of estuaries (commonly at 3-15 m depth), among seaweed or macroalgae, and more rarely on non-vegetated areas. Still, both species co-occur in Metruk Tuzla, an area submitted to major seasonal fluctuations in the abiotic environment, depending largely on freshwater inputs. Unfortunately, historical data on both species are lacking in the studied site and nearby areas.
The co-occurrence of species, also known as sympatry, might rely on abiotic and/or biotic environmental conditions. Whereas some pipefishes are widespread over large marine areas where they co-occur reproductively isolated, others have shorter ranges or inhabit specific sites [66,67]. Areas of co-occurrence between two species are often thought to arise in regions where abiotic conditions are favourable to both species and might therefore be intermediate between regions where either species might occur alone (allopatry) [68]. Thus, under certain environmental conditions, hybridization may foster sympatry, retaining populations in areas where pure species would otherwise be unable to persist. For example, eutrophication (i.e., turbidity or increased algal cover) may hamper species-specific sexual signalling, leading to hybridization [69-70].
The fishes of the genus Syngnathus show sophisticated mating [35,44], with males protecting and brooding developing eggs in specialized brood pouches. The lack of premating isolating mechanisms suggests that hybridization may be common in sympatric species of Syngnathus (e.g., in southern Californian). However, hybrids might not be viable in wild populations [71] or under experimental conditions due to postzygotic reproductive isolation between hybrids and their parental species [72]. Similarly, there is not any evidence of hybridization between S. abaster and S. acus.
Both abiotic and biotic interactions in extreme environments may facilitate co-occurrence; however, simple responses to the abiotic environment might not be the primary factors mediating co-occurrence in animals. Sympatry can be fostered by the presence of food resources that minimize competition between ecologically similar species [68]. Because of the sympatry of S. abaster and S. acus and the specialized parental care by males, we were interested in assessing the effect of species and sexual condition on their trophic features and the potential interspecific competition for food resources.
Most studies have reported that many syngnathids forage upon small crustaceans, including different taxa in the diet depending on species, snout and gape morphometrics, seasonal prey availability, size and maturity, among other factors (see review [41]). Consequently, all those factors might significantly affect foraging and the resulting isotopic profiles in pipefish tissues [34,37,73], which will reflect the dietary regime of the consumer [74-76]. There are several studies available focusing on the life cycle and dietary regime in sympatric pipefishes. Franzoi et al. (1993) [36] analyzed the gut content in the co-occurring species S. abaster and S. taenionotus in Po River Delta (Adriatic Sea) suggesting that both pipefishes probably do not compete for food resources, remaining almost completely segregated in different foraging microhabitats during most of their life span.
Similar to the findings by Franzoi et al. [36] and Vizzini and Mazzola (2004) [37], our isotope results revealed that both species occupy highly similar trophic niches, foraging almost totally or partially on highly similar prey types along the breeding season. However, the complementary gut content analysis suggests that both species would not compete largely for the same resources as they very likely exploit different micro-habitats. Both species would forage upon the copepod Calanipeda aquaedulcis, which is the main and almost the unique resource for S. abaster, but cladocerans were highly important in the dietary regime of S. acus. How pipefish species are spatially distributed along the water column or on the substrate seems to play a pivotal role in inter-specific competition for food resources. It has been hypothesized that the shorter-snouted S. abaster feed near the bottom whereas S. acus forage upon prey above algal beds [36]. This pattern has been also reported in S. abaster and S. acus from Lake Bafa [40] but the spatial occupancy of the habitat in both species in Metruk Tuzla lagoon is unknown.
The foraging upon different types of prey in syngnathids varies markedly depending on head and mouth morphology and biometrics, and the type and use of the habitat, among other factors [38,41,77]. Snout length as a proportion of head length is a widely used morphometric character for species identification in syngnathids but information on the relationship between mouth size and prey selection is fragmented [41]. It has been reported in Lake Bafa that S. abaster and S. acus preferred similar, but not always identical, prey groups [40]. The inter-specific differences in feeding habits would rely on morphological differences in gape morphometry, as mouth width in the S. abaster was larger.
Alluvian areas like Lake Bafa and Metruk Tuzla are submitted to significant changes in their environmental parameters, mainly salinity and temperature. This variability would affect prey abundances, resulting in a seasonal shift in prey preference (as reflected in the increase in %F in guts from S. acus collected in summer). The present study was carried out with specimens collected in the breeding season when the abundance of copepods, cladocerans and other small prey (e.g., chironomids) increased compared to winter [78]. It is feasible that the trophic features in the pipefishes studied will change accordingly. The calanoid copepod Calanipeda aquaedulcis is a euryhaline, eurythermal and nonspecialized filter feeder (phyto–detritophage) that is dominant in confined and relatively oligotrophic Mediterranean wetlands [79-81]. The species attain high abundances from spring to autumn, with peaks in summer [82]. In Güllük Bay, copepoda and cladocerans are the main zooplankton groups, accounting for 73 and 20% of total zooplankton, respectively [78].
Specimens of S. abaster from the western Mediterranean Stagnone di Marsala mainly fed on Mysidacea and Isopoda [37]. The dietary regime of S. acus from Cíes Archipelago seemed to prey primarily upon amphipods with varying proportions of carideans, isopods, copepods and mysidaceans [34,83]. However, Gurkan and Taskavak [84] reported the importance of cladocerans in the diet of the greater pipefish in the Turkish Aegean Sea, whereas Gurkan et al. [40] pointed out the preference for amphipods and copepods. Previous investigations in S. abaster revealed ontogenetic dietary changes, shifting from harpacticoid copepods to larger prey [39]. Ontogenic changes in dietary regimes were also reported in S. acus and S. abaster from Lake Bafa (Turkey), with an increasing preference for amphipods in larger fishes [40]. The isotopic profiles and the trophic assessment in the present study also support the occurrence of slight ontogenic dietary changes in both species, which is also in agreement with ontogenetic switching to larger food items pointed out in other pipefishes [85,86].
An interesting finding in our study was the depletion observed in δ13C (-17.0 ± 1.2 ‰) compared to specimens from other regions [37,73]. This finding supports gut content data [40,84, present study] indicating preference, especially in small pipefishes, for small and less mobile crustaceans, such as copepods or cladocerans, with low δ13C values and positioned in lower levels on the food web compared to larger crustaceans such as mysidaceans or amphipods. The δ13C depletion in those prey would be related to the ingestion of particles of terrestrial origin [87].
The isotopic characterization of an adequate baseline (i.e., typically particulate organic matter, primary producers or secondary producers) is imperative in the assessment of the trophic positions (TP) of species. Due to the lack of stable isotope data of organisms of the food web (i.e., more importantly, the lack of a known baseline) in the study area, the analysis of the TP occupied by both species could not be properly conducted. Nevertheless, tentative TP estimates were obtained for S. abaster and S. acus in Metruk Tuzla based on isotopic data using a hypothetical baseline (Mytilidae Musculus costulatus (Risso, 1826); trophic level = 2) from previous studies in Cíes Archipelago [34,88]. This approach does not necessarily provide accurate TPs estimates but allowed us to estimate inter-specific differences in TP. Details on the procedure, trophic discrimination factors applied [88] and other considerations on the baseline are provided in Appendix A.
The estimated TP (empirical mean ± s.d.) for S. abaster and S. acus populations in Metruk Tuzla was 3.67 ± 0.61 and 3.46 ± 0.66, respectively. The estimated trophic position for S. acus was highly similar to that in Cíes populations (TP = 3.40 ± 0.06) [34]. To our knowledge, other references to TP estimates in S. abaster and S. acus are lacking, impeding further comparisons. Anyway, our approach also suggests that both species are secondary consumers that occupy similar TP on the food web, confirming the inter-specific similarity revealed by the niche structure and overlap assessment.
Even though S. abaster and S. acus are currently listed in the Least Concern category by IUCN, the black-striped pipefish is included in Appendix III (Protected species) of Bern Convention and is considered critically endangered in some regions [89,90]. Due to their vulnerability considering biological and ecological features (i.e., limited dispersal ability, reduced home ranges, reproduction with parental care, susceptibility to anthropogenic disturbances and other factors), the protection of areas inhabited by these species must be taken into account, especially in disturbed coastal zones.
5. Conclusions
In summary, the present study provides new insights into the ecology of the sympatric pipefishes S. abaster and S. acus. From our results in Metruk Tuzla, it can be highlighted that both polyandrous species seem to have developed similar trophic features in the area, feeding on isotopically similar prey. The feeding regime in S. abaster seemed to be almost monospecific, foraging exclusively upon copepods, whereas both copepods and cladocerans were the main dietary sources in S. acus. Hence, both species can be considered selective but flexible carnivorous consumers in which the inter-specific competition for dietary resources seemed to be minimized likely due to the occupancy of different microhabitats. Considering future negative impacts resulting from anthropogenic activities on the area, future investigations should be conducted, including nearby populations, to analyze the genetic structure and local adaptations driven by variable environmental and physicochemical conditions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Global information on Syngnathus abaster and S. acus specimens collected in Metruk Tuzla (Boğaziçi Tuzla Lagoon, Turkey) in 2019; Figure S2: Length and weight in S. abaster and S. acus specimens collected in Metruk Tuzla (Boğaziçi Tuzla Lagoon, Turkey) in 2019; Figure S3: Isotopic values (δ13C and δ15N) in S. abaster and S. acus specimens collected in Metruk Tuzla (Boğaziçi Tuzla Lagoon, Turkey) in 2019; Figure S4: Numerical occurrence (%N) of prey items in the gusts of S. abaster and S. acus specimens collected in Metruk Tuzla (Boğaziçi Tuzla Lagoon, Turkey) in 2019. Figure S5: Niche plots (95% alpha) (A - δ15N; D - δ13C), density distributions (B), and raw data (C) for each pairwise combination of isotope (δ13C and δ15N) data in S. abaster and S. acus specimens collected in Metruk Tuzla (Boğaziçi Tuzla Lagoon, Turkey) in 2019. Figure S6: Relationship between total length and pouch length in S. acus and S. acus males from Metruk Tuzla (Boğaziçi Tuzla Lagoon, Turkey). Table S1: General information of S. acus specimens collected in Metruk Tuzla (Boğaziçi Tuzla Lagoon, Turkey) in 2019. Table S2: General information of S. abaster specimens collected in Metruk Tuzla (Boğaziçi Tuzla Lagoon, Turkey) in 2019. Table S3: δ13C and δ15N values used as baseline 1 and baseline 2 (Musculus costulatus sampled in Cíes Archipelago, NW Spain). Data from [34].
Author Contributions
Conceptualization, S.G. and M.P.; methodology, S.G. and M.P.; software, M.P.; validation, S.G. and M.P.; formal analysis, S.G., P.A., C.MS. and M.P.; investigation, S.G. and M.P.; resources, S.G., D.I. and M.P.; data curation, S.G. and M.P.; writing—original draft preparation, M.P.; writing—review and editing, S.G., P.A., D.I., C.MS. and M.P.; visualization, S.G. and M.P.; supervision, M.P.; project administration, S.G and M.P.; funding acquisition, M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by Contrato Programa Xunta de Galicia-CSIC (Spain).
Data Availability Statement
The data presented in this manuscript are available in Supplementary Material.
Acknowledgments
We would like to thank Dr. Metin Türker, General Manager of Agricultural Research and Policies of the Ministry of Agriculture and Forestry of the Republic of Turkey, for his approval and contribution in obtaining the necessary legal permits and documents for this study at an international level.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A
The procedure followed to estimate the trophic positions in S. abaster and S. acus is described in [34]. The analyses were performed using tRophicPosition v. 0.7.7 package in R [91]. For that, we applied experimentally derived TDF (Trophic Discriminant Factors) values for syngnathids (2.5 for δ13C; 3.9 for δ15N) [88].
Two baselines (spring-summer 2017 and spring-summer-2018) were selected considering available isotopic data in the scarce isotopic studies available on S. abaster and S. acus in European populations. Vizzini and Mazola (2004) [37] reported mean δ13C and δ15N values of -10.5 ‰ and 11.9 ‰ in S. abaster from western Mediterranean Stagnone di Marsala (Italy), whereas Planas (2022) [34] provided mean values of -15.4 ‰ for δ13C and 11.1 ‰ for δ15N in S. acus from Cíes Archipelago (NW Spain). Considering the higher similarity between our data in Metruk Tuzla (-17.0 ± 1.2 ‰ for δ13C and 13.2 ± 0.6 ‰ for δ15N in S. abaster, and -17.3 ± 1.2 ‰ for δ13C and 13.3 ± 0.7 ‰ for δ15N in S. acus) and those from Planas (2022) [34], we used the isotopic data of the baseline organism (Mytilidae Musculus costulatus) in Cíes Archipelago. Baseline isotopic values are provided in Table S3.
The TP analysis was carried out using “twoBaselinesFull” model, which included δ13C and δ15N values, under the following conditions:
- -
- Iterations: 20210:40000
- -
- Thinning interval: 10
- -
- Number of chains: 5
- -
- Sample size per chain: 2000
References
- Cherry, J. A. Ecology of wetland ecosystems: Water, substrate, and life. Nat. Educ. Know. 2011. 3(10), 16. [Google Scholar]
- Whittaker, R.H.; Likens, G.E. Carbon in the biota. In Carbon in the Biosphere; Woodwell, G. M., Peacan, E. R. Peacan, Eds.; National Technical Information Service: Springfield, VA, 1973; pp. 281–302. [Google Scholar]
- Curebal, I. , Efe, R., Soykan, A.; Sonmez, S. Impacts of anthropogenic factors on land degradation during the Anthropocene in Turkey. J. of Environ. Bio. 2015. 36(1), 51-58. [Google Scholar]
- Murat, A.; Ortaç, O. Wetland loss in Turkey over a hundred years: implications for conservation and management. Ecosyst. Health Sustain. 2021; 7(1),1930587.
- Çolak, M.A.; Öztaş, B.; Özgencil, İ.K.; Soyluer, M.; Korkmaz, M.; Ramírez-García, A.; Metin, M.; Yılmaz, G.; Ertuğrul, S.; Tavşanoğlu, Ü.N.; et al. Increased water abstraction and climate change have substantial effect on morphometry, salinity, and biotic communities in lakes: Examples from the semi-arid Burdur Basin (Turkey). Water 2022, 14, 1241. [Google Scholar] [CrossRef]
- Somay-Altaş, A.M. Hydrogeochemical fingerprints of a mixohaline wetland in the Mediterranean: Güllük coastal wetland systems- GCWS (Muğla, Turkey). Turkish J Earth Sci 2021, 30, 38–58. [Google Scholar] [CrossRef]
- Anonymous. Gediz Delta Wetland Management Plan. T.R. Ministry of Environment and Forestry, General Directorate of Nature Conservation and National Parks, Department of Nature Conservation, Directorate of Wetlands Branch, 2007, 424 p. (in Turkish).
- Tosunoğlu, Z.; Ünal, V.; Kaykaç, M.H. Ege Dalyanları. SÜR-KOOP Su Ürünleri Kooperatifleri Merkez Birliği Yayınları No: 03, ISBN:978-605-60880-2-5, Ankara, 2017, 322 pp. (in Turkish).
- Davis, T.; Blasco, D.; Carbonell, M. The Ramsar Convention manual: a guide to the Convention on wetlands (Ramsar, Iran, 1971);Convention on Wetlands of International Importance Especially as Waterfowl Habitat, Ramsar Convention Bureau.; IUCN: International Union for Conservation of Nature: Gland, Switzerland, 1997.
- Ramsar Convention Secretariat. An Introduction to the Convention on Wetlands (previously The Ramsar Convention Manual). Ramsar Convention Secretariat: Gland, Switzerland. 2016; pp. 107.
- Demirak, A.; Balcı, A.; Demirhan, H.; Tüfekçi, M. The reasons of the pollution in the Güllük Bay, in Proceedings of the 4th National Ecology and Environment Congress, 2001; pp. 383–388 (in Turkish).
- Demirak, A.; Balci, A.; Tüfekçi, M. Environmental impact of the marine aquaculture in Güllük Bay, Turkey. Environ Monit Assess. 2006, 123(1-3), 1-12.
- Yucel-Gier, G.; Pazi, I.; Kucuksezgin, F. Spatial analysis of fish farming in the Gulluk Bay (Eastern Aegean). Turk. J. Fish. Aquat. Sci. 2013, 13, 737–744. [Google Scholar] [CrossRef]
- MARA (Ministry of Agriculture and Rural Affairs) Report, 1988. Research project of Gulluk Lagun. Institute of Fishery in Bodrum, Turkey, 1988, 41 pp (In Turkish).
- Tokaç, A., Ünal V., Tosunoğlu, Z., Akyol O., Özbilgin H., Gökçe G. Fisheries of Aegean Sea; IMEAK Deniz Ticaret Odası İzmir Şubesi Yayınları: İzmir, 2010; 371 pp. (in Turkish).
- Sağlam, C.; Akyol, O.; Ceyhan, T. Fisheries in Güllük Lagoon. Ege J Fish Aqua Sci. 2015, 32(3), 145–149. [Google Scholar]
- Altınsaçlı, S.; Perçin-Paçal, F.; Altınsaçlı, S.. Assessments on diversity, spatiotemporal distribution and ecology of the living ostracod species (Crustacea) in oligo-hypersaline coastal wetland of Bargilya (Milas, Muğla, Turkey). Int. J. Fish. Aquat. Stud. 2015, 3(2), 357-373.
- Cerim, H. Determination of some biological and population parameters of common sole (Solea solea Linnaeus, 1758) in Güllük Bay trammel net fishery. Doctoral dissertation, Muğla Sıtkı Koçman University, Muğla, Turkey, 2017 (in Turkish).
- IUCN. The IUCN Red List of threatened species. 2023, https://www.iucnredlist.org/ (accessed on 12 June 2023).
- Dawson, C.E. Syngnathidae. In Fishes of the North-eastern Atlantic and the Mediterranean, Whitehead, P.J.P., Bauchot, M.-L., Hureau, J.-C., Nielsen, J., Tortonese. E., Eds.; Unesco: Paris, 1986; Volume 2., p. 628-639.
- Smith-Vaniz, W.F. Syngnathus acus. The IUCN Red List of Threatened Species 2015: E.T198765A44933898. Available online: 10.2305/\T1\iucn.uk.20154 (accessed on 12 June 2023).
- Pollom, R.. Syngnathus abaster. The IUCN Red List of Threatened Species 2016: e.T21257A19423178. https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T21257A19423178.en (accessed on 12 June 2023).
- Vasileva, E.D. Main alterations in ichthyofauna of the largest rivers of the northern coast of the Black Sea in the last 50 years: A review. Folia Zoo.a 2003, 52, 337–358. [Google Scholar]
- Monteiro, N.M.; Vieira, M.N. Rendez-vous at the Baltic? The ongoing dispersion of the black-striped pipefish, Syngnathus abaster. Oceanogr. Fish. Open Access J. 2017, 3, 2. [Google Scholar]
- Didenko, A.; Kruzhylina, S.; Gurbyk, A. Feeding patterns of the black-striped pipefish Syngnathus abaster in an invaded freshwater habitat. Environ. Biol. Fishes 2018, 101, 917–931. [Google Scholar] [CrossRef]
- Cakić, P.M.; Lenhardt, D.; Mićkovıć, N.; Sekulıć, L.; Budakov, J. Biometric analysis of Syngnathus abaster populations. J. Fish Biol. 2002, 60, 1562–1569.
- Kolangi-Miandare, H.; Askari, G.; Fadakar1, D.; Aghilnegad, M.; Azizah, S. The biometric and cytochrome oxidase sub unit I (COI) gene sequence analysis of Syngnathus abaster (Teleostei: Syngnathidae) in Caspian Sea. Mol. Bio. Res. Commun. 2013, 2(4),133–142. [Google Scholar]
- Vieira, R.P.; Monteiro, P.; Ribeiro, J.; Bentes, L.; Oliveira, F.; Erzini, K.; Gonçalves, J.M.S. Length-weight relationships of six syngnathid species from Ria Formosa, SW Iberian coast. Cah. Biol. Mar. 2014, 55, 9–12. [Google Scholar]
- Yildiz, T.; Uzer, U.; Firdes Saadet, K. Preliminary report of a biometric analysis of greater pipefish Syngnathus acus Linnaeus, 1758 for the western Black Sea. Turk. J. Zool. 2015. 39(5), 19. [Google Scholar] [CrossRef]
- Gurkan, S.; Innal, D. Some morphometric features of congeneric pipefish species (Syngnathus abaster Risso 1826, Syngnathus acus Linnaeus, 1758) distributed in Lake Bafa (Turkey). Oceanol. Hydrobiol. Stud. 2018, 47(3), 239–247. [Google Scholar] [CrossRef]
- Dawson, C.E. Syngnathidae. InFishes of the north-eastern Atlantic and the Mediterranean, Whitehead, P.J.P. Ed.; Unesco, Paris, France, 1986;, pp. 628-639.
- Planas, M.; Piñeiro-Corbeira, C.; Bouza, C.; Castejón-Silvo, I.; Vera, M.; Regueira, M.; Ochoa, V.; Bárbara, I.; Terrados, J.; Chamorro, A.; et al. A multidisciplinary approach to identify priority areas for the monitoring of a vulnerable family of fishes in Spanish Marine National Parks. BMC Ecol. Evol. 2021, 21, 4. [Google Scholar] [CrossRef]
- Silva, K.; Monteiro, N.M.; Almada, V.C.; Vieira, M.N. Early life history of Syngnathus abaster (Pisces: syngnathidae). J. Fish Biol. 2006, 68, 80–86. [Google Scholar] [CrossRef]
- Planas, M. Ecological traits and trophic plasticity in the greater pipefish Syngnathus acus in the NW Iberian Peninsula. Biology 2022, 11(5), 712. [Google Scholar] [CrossRef]
- Silva, K,; Monteiro, N. M.; Vieira, M.N.; Almada, V.C. Reproductive behaviour of the black-striped pipefish, Syngnathus abaster (Pisces; Syngnathidae). J. Fish Biol. 2006, 69, 1860–1869. [Google Scholar] [CrossRef]
- Franzoi, P.R.; Maccagnani, R.R.; Ceccherelli, V.U. Life cycles and feeding habits of Syngnathus taenionotus and Syngnathus abaster (Pisces, Syngnathidae) in brackish bay of the Po River delta (Adriatic Sea). Mar. Ecol. Prog. Ser. 1993, 97, 71–81. [Google Scholar] [CrossRef]
- Vizzini, S.; Mazzola, A. The structure of pipefish community (Pisces: Synganthidae) from a western Mediterranean sea grass meadow based on stable isotope analysis. Estuaries 2004, 27(2), 325–333. [Google Scholar] [CrossRef]
- Kendrick, A.J.; Hydnes, G.A. Variations in the dietary compositions of morphologically diverse syngnathid fishes. Environ. Biol. Fish., 2005, 72, 415–427. [Google Scholar] [CrossRef]
- Taşkavak, E.; Gürkan, Ş.; Severa, T.M.; Akalına, S.; Özaydına, O. Gut contents and feeding habits of the great pipefish, Syngnathus acus Linnaeus, 1758, in Izmir Bay (Aegean Sea, Turkey). Zool. Middle East 2010, 50, 75–82. [Google Scholar] [CrossRef]
- Gurkan, Ş.; Innal, D.; Gulle, I. Monitoring of the trophic ecology of pipefish species (Syngnathus abaster, Syngnathus acus) in an alluvial lake habitat (Lake Bafa, Turkey). Oceanol. Hydrobiol. Stud. 2021, 50, 24–32. [Google Scholar] [CrossRef]
- Manning, C.G.; Foster, S.J.; Vincent, A.C.J. A review of the diets and feeding behaviours of a family of biologically diverse marine fishes (Family Syngnathidae). Rev. Fish Biol. Fisheries 2019, 29, 197–221. [Google Scholar] [CrossRef]
- MEF, 2011. General Directory of Nature Protection and National Parks. http://gis2.cevreorman.gov.tr/mp/ (accessed 2 June 2023). 2 June.
- Tomasini, J.A.; Quignard, P.; Capapé, C.; Bouchereau, L. Facteurs du succès reproductif de Syngnathus abaster Risso, 1826 (Pisces, Teleostei, Syngnathidae) en milieu lagunaire méditerranéen (lagune de Mauguio, France). Acta Oecol. 1991, 12, 331–355. [Google Scholar]
- Vincent, A.; Berglund, A.; Ahnesjö, I. Reproductive ecology of five pipefish species in one eelgrass meadow. Environ. Biol. Fish 1995, 44, 347–361. [Google Scholar] [CrossRef]
- Riccato, F.; Fiorin, R.; Franco, A.; et al. Population structure and reproduction of three pipefish species (Teleostei, Syngnathidae) in a seagrass meadow of the Venice lagoon. Biol. Mar. Medit. 2003, 10, 138–145. [Google Scholar]
- Liao, H.; Pierce, C.L.; Larscheid, J.G. Empirical assessment of indices of prey importance in the diets of predacious fish. T. Am. Fısh Soc. 2001, 130, 583–591. [Google Scholar] [CrossRef]
- Pinnegar, J.K.; Polunin, N.V.C. Differential fractionation of δ13C and δ15N among fish tissues: implications for the study of trophic interactions. Funct. Ecol. 1999, 13, 225–231. [Google Scholar] [CrossRef]
- Sotiropoulos, M.A.; Tonn, W.M.; Wassenaar, L.I. Effects of lipid extraction on stable carbon and nitrogen isotope analyses of fish tissues: potential consequences for food web studies. Ecol. Freshw. Fish 2004, 13, 155–160. [Google Scholar] [CrossRef]
- Sweeting, C.J.; Polunin, N.V.C.; Jennings, S. Effects of chemical lipid extraction and arithmetic lipid correction on stable isotope ratios of fish tissues. Rapid Commun. Mass Spectrom. 2006, 20, 595–601. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montaña, C.G. Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecol. 2007, 152, 179–189. [Google Scholar] [CrossRef]
- Logan, J.M.; Jardine, T.D.; Miller, T.J.; Bunn, S.E.; Cunjak, R.A.; Lutcavage, M.E. Lipid corrections in carbon and nitrogen stable isotope analyses: comparison of chemical extraction and modelling methods. J. Anim. Ecol. 2008, 77, 838–846. [Google Scholar] [CrossRef]
- Valladares, S.; Planas, M. Non-lethal dorsal fin sampling for stable isotope analysis in seahorses. Aquat. Ecol. 2012, 46, 363–370. [Google Scholar] [CrossRef]
- Planas, M.; Paltrinieri, A.; Carneiro, M.D.D.; Hernández-Urcera, J. Effects of tissue preservation on carbon and nitrogen stable isotope signatures in Syngnathid fishes and prey. Anim. 2020, 10, 2301. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. The R Project for Statistical Computing. Available online: http://www.R-project.org (accessed on 23 April 2020).
- R Core Team. Stats: The R stats package. Available online: https:// https://www.rdocumentation.org/packages/stats/versions/3.6.2 (accessed on 23 February 2018).
- Le Cren, E.D. The length-weight relationship and seasonal cycle in gonad weight and condition in the Perch (Perca fluviatilis). J. Ani. Ecol., 1951, 20, 201–219. [Google Scholar] [CrossRef]
- Ricker, W.E.; Carter, N.M. Handbook of computations for biological statistics of fish populations, No. 119. The Fisheries Research Board of Canada, Queen’s printer and controller of stationary: Ottawa, 1958.
- Jackson, A. L.; Inger, R.; Parnell, A. C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER – Stable isotope Bayesian ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Lysy, M.; Stasko, A.D.; Swanson, H.K. nicheROVER: (Niche) (R)egion and Niche(Over)lap metrics for multidimensional ecological niches. R package version 1.1.0. https://CRAN.R-project.org/package=nicheROVER (accessed 17 September 2019). 17 September.
- Swanson, H.K.; Lysy, M.; Power, M.; Stasko, A.D.; Johnson, J.D.; Reist, J.D. A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap. Ecol. 2015, 96, 318–324. [Google Scholar] [CrossRef]
- Wickham, H.; Chang, W. Create elegant data visualisations using the grammar of graphics. https://CRAN.R-project.org/package=ggplot2 (accessed 17 September 2019). 17 September.
- Veiga, P.; Machado, D.; Almeida, C.; Bentes, L.; Monteiro, P.; Oliveira, F.; Ruano, M.; Erzini, K.; Gonçalves, J.M.S. Weight-length relationships for 54 species of the Arade estuary, southern Portugal. J. Appl. Ichthyol. 2009, 25, 493–496. [Google Scholar] [CrossRef]
- Gürkan, Ş.; Taşkavak, E.; Hoşsucu, B. The reproductive biology of the Great Pipefish Syngnathus acus (Family: Syngnathidae) in the Aegean Sea. North˗West J. Zool. 2009, 5(1), 179˗190.
- Simal Rodríguez, A.; Grau, A.; Castro-Fernández, J.; et al. Reproductive biology of pipefish Syngnathus typhle and S. abaster (Syngnathidae) from Western Mediterranean Sea. J. Ichthyol. 2021, 61, 608–615. [Google Scholar] [CrossRef]
- Liousia, V. Study on the biology of the syngnathidae family in Greece. PhD Thesis, Dissertation, University of Ioannina, Greece, 2015. [Google Scholar]
- Fritzsche, R. A. Revision of the Eastern Pacific Syngnathidae (Pisces: Syngnathiformes), including both recent and fossil forms. Proceedings of the California Academy of Sciences, San Francisco, USA, 1980; 42, 181–227.
- Garcia, E.; Rice, C.A.; Eernisse, D.J.; Forsgren, K.L.; Quimbayo, J.P.; Rouse, G.W. Systematic relationships of sympatric pipefishes (Syngnathus spp.): A mismatch between morphological and molecular variation. J. Fish Biol. 2019, 95(4), 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Chunco, A.J.; Jobe, T.; Pfennig, K.S. Why do species co-occur? A test of alternative hypotheses describing abiotic differences in sympatry versus allopatry using spadefoot toads. PLoS One 2012. 7(3), e32748. [Google Scholar] [CrossRef] [PubMed]
- Seehausen, O.; van Alphen, J.J.M.; Witte, F. Cichlid fish diversity threatened by eutrophication that curbs sexual selection. Sci. 1977, 277, 1808–1811. [Google Scholar] [CrossRef]
- Sundin, J.; Aronsen, T.; Rosenqvist, G. et al. Sex in murky waters: algal-induced turbidity increases sexual selection in pipefish. Behav. Ecol. Sociobiol. 2017, 71-78.
- Wilson, A. B. Interspecies mating in sympatric species of Syngnathus pipefish. Mol. Ecol. 2006, 15, 809–824. [Google Scholar] [CrossRef] [PubMed]
- Moser, F.N.; Wilson, A.B. Reproductive isolation following hybrid speciation in Mediterranean pipefish (Syngnathus spp.). Anim. Behav. 2020, 161, 77–87. [Google Scholar] [CrossRef]
- Piñeiro-Corbeira, C.; Iglesias, L.; Nogueira, R.; Campos, S.; Jiménez, A.; Regueira, M.; Barreiro, R.; Planas, M. Structure and trophic niches in mobile epifauna assemblages associated with seaweeds and habitats of syngnathid fishes in Cíes Archipelago (Atlantic Islands Marine National Park, NW Iberia). Front. Mar. Sci. 2021, 8, 773367. [Google Scholar] [CrossRef]
- Tieszen, L.L.; Boutton, T.W.; Tesdahl, K.G.; Slade, N.A. Fractionation and turnover of stable carbon isotopes in animal tissues: implications for δ13C analysis of diet. Oecol. 1983, 57, 32–37. [Google Scholar] [CrossRef]
- Fry, B. Food web structure on Georges Bank from stable C, N and S isotopic compositions. Limnol. Oceanogr. 1988, 33, 1182–1190. [Google Scholar] [CrossRef]
- Hobson, K. A.; Ambrose, Jr. W. G.; Renaud, P. E. Sources of primary production, benthic-pelagic coupling, and trophic relationships within the Northeast Water Polynya: insights from δ13C and δ15N analysis. Mar. Ecol. Prog. Ser. 1995, 128, 1–10. [Google Scholar] [CrossRef]
- de Lussanet Marc, H.E; Muller, M. The smaller your mouth, the longer your snout: predicting the snout length of Syngnathus acus, Centriscus scutatus and other pipette feeders. J. R. Soc. Interface. 2007, 4561–573. [Google Scholar] [CrossRef]
- Killi, N. Spatio-temporal variation in the distribution and abundance of marine cladocerans in relation to environmental factors in a productive lagoon (Güllük Bay, SW Aegean Sea, Turkey)" Oceanol. Hydrobiol. Stud. 2020, 49(4), 374–382. [Google Scholar] [CrossRef]
- Boix, D.; Gascón, S.; Sala, J.; Martinoy, M.; Gifre, J.; Quintana, X.D. A new index of water quality assessment in Mediterranean wetlands based on crustacean and insect assemblages: the case of Catalunya (NE Iberian Peninsula). Aquatic Conserv. Mar. Freshw. Ecosyst. 2005, 15, 635–651. [Google Scholar] [CrossRef]
- Brucet, S.; Compte, J.; Boix, D.; López-Flores, R.; Quintana, X. D. Feeding of nauplii, copepodites and adults of Calanipeda aquaedulcis (Calanoida) in Mediterranean salt marshes. Mar. Ecol. Prog. Ser. 2008, 355, 183–191. [Google Scholar] [CrossRef]
- Lazareva, V.I. The Mediterranean copepod Calanipeda aquaedulcis Kritschagin, 1873 (Crustacea, Calanoida) in the Volga River reservoirs. Inland Water Biol. 2018, 11(3), 303–309. [Google Scholar] [CrossRef]
- Uriarte, I.; Villate, F.; Iriarte, A. Zooplankton recolonization of the inner estuary of Bilbao: influence of pollution abatement, climate and non-indigenous species. J. Plankton Res. 2016, 38(3), 718–731. [Google Scholar] [CrossRef]
- Planas, M. Was that my meal? Uncertainty from source sampling period in diet reconstruction based on stable isotopes in a syngnathid fish. Front. Mar. Sci. 2022, 9, 982883. [Google Scholar] [CrossRef]
- Gurkan, Ş.; Taşkavak, E. The relationships between gut length and prey preference of three pipefish (Syngnathus acus, Syngnathus typhle, Nerophis ophidion Linnaeus, 1758) species distributed in Aegean Sea, Turkey. Iran. J. Fish. Sci. 2019, 18, 1093–1100. [Google Scholar]
- Tipton, K.; Bell, S.S. Foraging patterns of two syngnathid fishes: importance of harpacticoid copepods. Mar. Ecol. Prog. Ser. 1988, 47, 31–43. [Google Scholar] [CrossRef]
- Oliveira, F.; Erzini, K.; Gonçalves, J.M.S. Feeding habits of the deep-snouted pipefish Syngnathus typhle in a temperate coastal lagoon. Estuar. Coast. Shelf Sci. 2007, 72, 337–347. [Google Scholar] [CrossRef]
- Cole, J.J.; Carpenter, S.R.; Kitchell, J.; Weidel, B. Strong evidence for terrestrial support of zooplankton in small lakes based on stable isotopes of carbon, nitrogen, and hydrogen. Biol. Sci. 2020, 108(5), 1975–1980. [Google Scholar] [CrossRef]
- Planas, M.; Chamorro, A.; Paltrinieri, A.; Campos, S.; Nedelec, K.; Hernández-Urcera, J. Effect of diet on breeders and inheritance in Syngnathids: Application of isotopic experimentally derived data to field studies. Mar. Ecol. Prog. Ser. 2020, 650, 107–123. [Google Scholar] [CrossRef]
- Mayol, J.; Grau, A.; Riera, F.; Oliver, J. Llista vermella dels peixos de les Balears; Conselleria de Medi Ambient-Conselleria d’Agricultura i Pesca, Palma de Mallorca, España, 2000.
- Manent Sintes, P.; Abella-Gutiérrez, J. Aspectos ecológicos y biológicos de Syngnathus abaster (Risso, 1826) en la bahía de Fornells. Revista de Menorca 2004, 88, 89–105. [Google Scholar]
- Quezada-Romegialli, C.; Jackson, A. L.; Hayden, B.; Kahilainen, K. K.; Lopes, C.; Harrod, C. tRophicPosition, an R package for the Bayesian estimation of trophic position from consumer stable isotope ratios. Methods Ecol. Evol. 2018, 9, 1592–1599. [Google Scholar] [CrossRef]
Figure 1.
Study site and shore seine net used for pipefish collection.
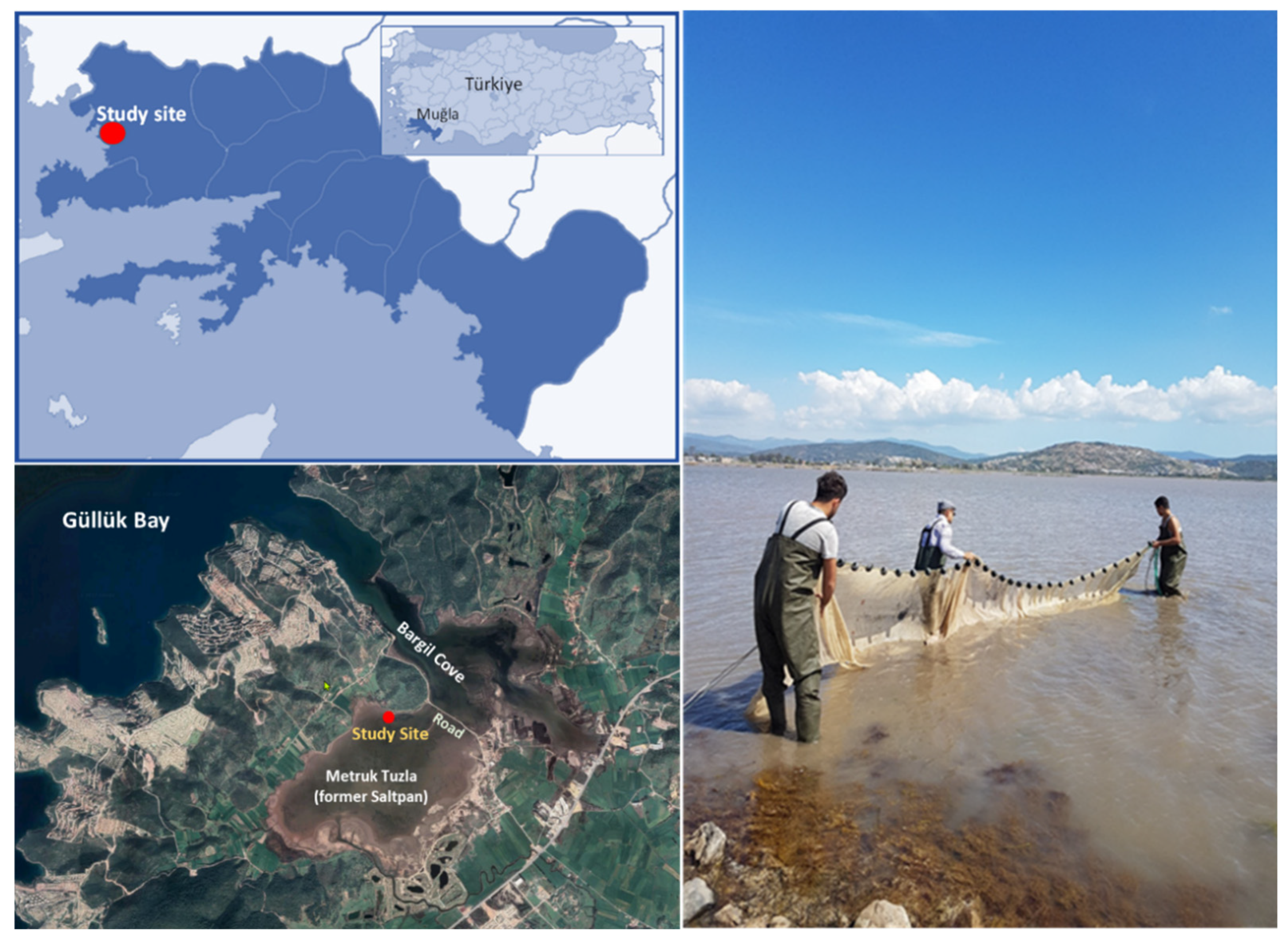
Figure 2.
Syngnathus acus collected by the shore seine net (left) and adults (right) of S. abaster and S. acus (females and pregnant males).
Figure 2.
Syngnathus acus collected by the shore seine net (left) and adults (right) of S. abaster and S. acus (females and pregnant males).

Figure 3.
Relationship between total length (TL) and wet weight (W) (W=aTLb) in the pipefishes S. abaster and S. acus collected in Metruk Tuzla.
Figure 3.
Relationship between total length (TL) and wet weight (W) (W=aTLb) in the pipefishes S. abaster and S. acus collected in Metruk Tuzla.
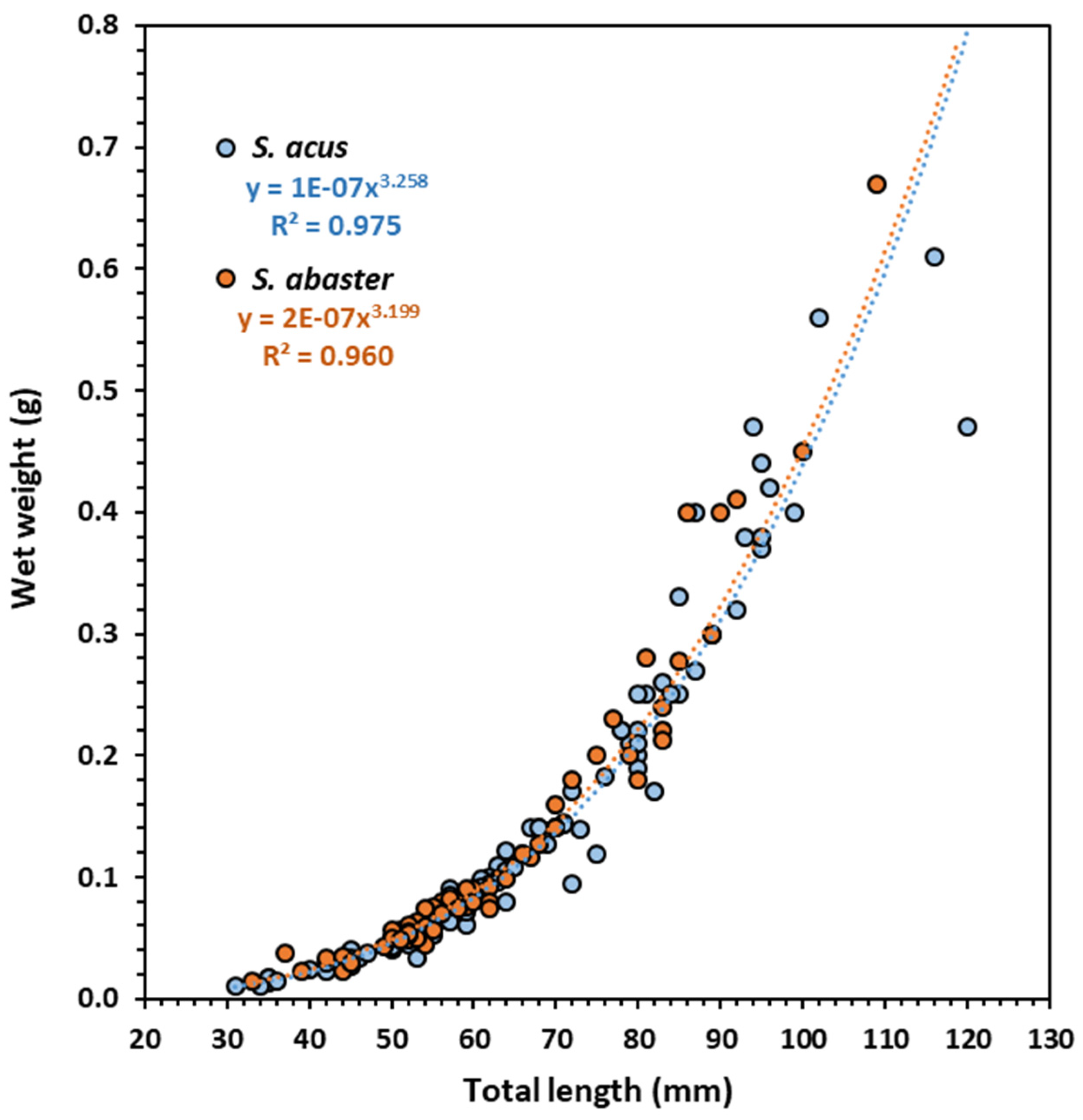
Figure 4.
Stable isotope Bayesian ellipses showing trophic niche widths and overlaps in S. abaster and S. acus specimens collected in Metruk Tuzla. Ellipses with 95% credible intervals for the means are based on standard ellipses corrected for small sample sizes (SEAc; isotopic niche metrics; SIBER package). Centroids and credible intervals are displayed for each ellipse.
Figure 4.
Stable isotope Bayesian ellipses showing trophic niche widths and overlaps in S. abaster and S. acus specimens collected in Metruk Tuzla. Ellipses with 95% credible intervals for the means are based on standard ellipses corrected for small sample sizes (SEAc; isotopic niche metrics; SIBER package). Centroids and credible intervals are displayed for each ellipse.
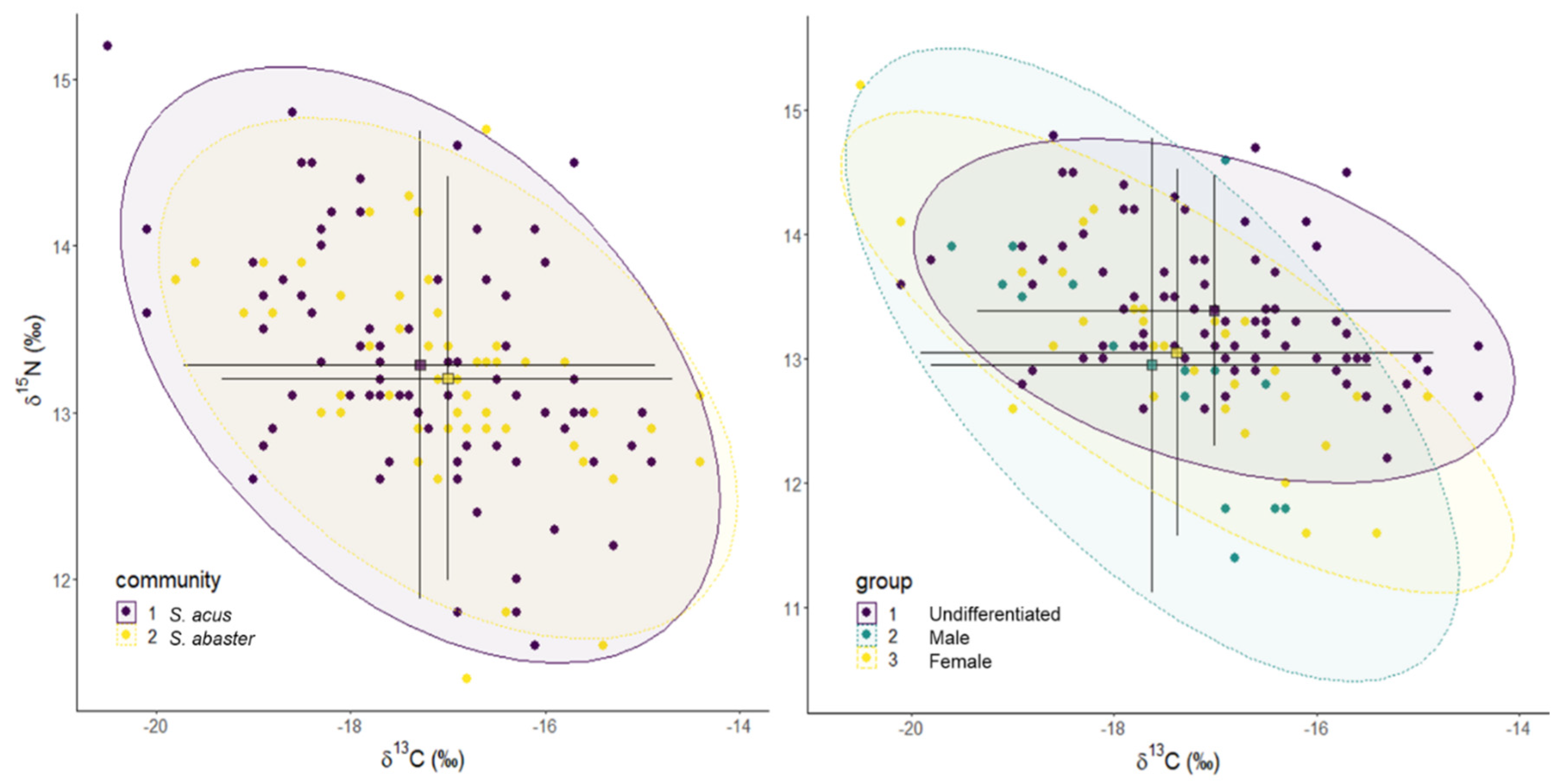

Table 1.
Total length (TL; cm) and wet weight (g) in specimens of S. abaster and S. acus collected in Metruk Tuzla (n: sample size; sd: standard deviation).
Table 1.
Total length (TL; cm) and wet weight (g) in specimens of S. abaster and S. acus collected in Metruk Tuzla (n: sample size; sd: standard deviation).
| Total length (cm) | Wet weight (g) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Sex condition | n | Mean | sd | Max | Min | Mean | sd | Max | Min |
| Syngnathus abaster | Pooled | 51 | 5.9 | 1.6 | 10.9 | 3.3 | 0.115 | 0.128 | 0.669 | 0.015 |
| Undifferentiated | 37 | 5.2 | 0.7 | 6.6 | 3.3 | 0.058 | 0.023 | 0.119 | 0.015 | |
| Male | 7 | 8.8 | 1.0 | 10.9 | 7.7 | 0.358 | 0.155 | 0.669 | 0.230 | |
| Female | 7 | 7.2 | 1.5 | 10.0 | 5.4 | 0.168 | 0.134 | 0.450 | 0.069 | |
| Syngnathus acus | Pooled | 78 | 6.3 | 1.9 | 12.0 | 3.1 | 0.124 | 0.125 | 0.610 | 0.010 |
| Undifferentiated | 45 | 5.0 | 0.9 | 6.4 | 3.1 | 0.051 | 0.025 | 0.096 | 0.010 | |
| Male | 9 | 8.4 | 1.9 | 12.0 | 6.3 | 0-272 | 0.154 | 0.470 | 0.098 | |
| Female | 24 | 7.8 | 1.4 | 11.6 | 6.1 | 0.206 | 0.130 | 0.610 | 0.095 | |
Table 2.
LWR parameters for S. abaster and S. acus collected in Metruk Tuzla. Equation: log (W) = log a + b log (TL). n: sample size; a: intercept; b: slope; se: standard error; CI: confidence interval.
Table 2.
LWR parameters for S. abaster and S. acus collected in Metruk Tuzla. Equation: log (W) = log a + b log (TL). n: sample size; a: intercept; b: slope; se: standard error; CI: confidence interval.
| Species | n | a | b | se b | CI b | R2 | p | t-test p | Growth type |
|---|---|---|---|---|---|---|---|---|---|
| Syngnathus abaster | 51 | 0.000294 | 3.179 | 0.095 | 2.989 – 3.369 | 0.958 | < 0.0001 | < 0.0001 | Allometric (+) |
| Syngnathus acus | 78 | 0.000269 | 3.182 | 0.057 | 3.069 – 3.296 | 0.976 | < 0.0001 | < 0.0001 | Allometric (+) |
Table 3.
Gut contents (pooled, spring and summer samples) in S. abaster and S. acus individuals (%F: frequency percentage; %N: prey number percentage) from Metruk Tuzla. n.d.: not detected.
Table 3.
Gut contents (pooled, spring and summer samples) in S. abaster and S. acus individuals (%F: frequency percentage; %N: prey number percentage) from Metruk Tuzla. n.d.: not detected.
| S. abaster | S. acus | S. abaster | S. acus | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pooled | Pooled | Spring | Summer | Spring | Summer | |||||||
| Prey Groups | %F | %N | %F | %N | %F | %N | %F | %N | %F | %N | %F | %N |
| Copepoda | ||||||||||||
| Halicyclops biscupidatus | n.d. | n.d. | 1.2 | 0.2 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 1.0 | 54.2 |
| Calanipeda aquaedulcis | 42.6 | 71.8 | 35.3 | 47.5 | 37.0 | 70.5 | 47.3 | 75.5 | 87.7 | 44.9 | 53.6 | 53.6 |
| Other copepods | 37.7 | 15.5 | 11.8 | 4.7 | 73.0 | 29.6 | 18.9 | 10.2 | 5.2 | 4.1 | 6.2 | 6.3 |
| Cladocera | ||||||||||||
| Daphnia longispina | n.d. | n.d. | 3.5 | 0.6 | n.d. | n.d. | n.d. | n.d. | 0.6 | 0.5 | 1.0 | 0.7 |
| Ceriodaphnia quadrangula | n.d. | n.d. | 1.2 | 0.2 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 1.0 | 1.0 |
| Pleopis polyphemoides | n.d. | n.d. | 2.3 | 0.4 | n.d. | n.d. | n.d. | n.d. | 0.6 | 0.3 | 1.0 | 0.7 |
| Cladocera pieces | n.d. | n.d. | 40.0 | 45.7 | n.d. | n.d. | n.d. | n.d. | 4.5 | 49.6 | 35.0 | 35.9 |
| Insecta | n.d. | n.d. | 1.2 | 0.2 | n.d. | n.d. | n.d. | n.d. | 0.6 | 0.3 | n.d. | 0.7 |
| Mollusca | n.d. | n.d. | 1.2 | 0.2 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 1.0 | n.d. |
| Fish scale | 4.9 | 0.7 | 1.2 | 0.2 | 7.4 | 0.5 | 1.3 | 1.4 | 0.6 | 0.3 | n.d. | 0.7 |
| Unidentified eggs | 14.7 | 12.0 | n.d. | n.d. | 25.9 | 11.7 | 32.4 | 12.9 | n.d. | n.d. | n.d. | n.d. |
Table 4.
Isotopic values (δ13C and δ15N) in S. abaster and S. acus specimens from Metruk Tuzla.
| δ13C ‰ | δ15N ‰ | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Sex condition | n | Mean | sd | Max | Min | Mean | sd | Max | Min |
| Syngnathus abaster | Pooled | 51 | -17.0 | 1.2 | -14.4 | -19.8 | 13.2 | 0.6 | 14.7 | 11.4 |
| Undifferentiated | 37 | -17.0 | 1.2 | -14.4 | -19.8 | 13.4 | 0.5 | 14.7 | 12.6 | |
| Male | 7 | -17.6 | 1.2 | -16.4 | -19.6 | 12.8 | 0.9 | 13.9 | 11.4 | |
| Female | 7 | -16.6 | 0.9 | -15.4 | -17.8 | 12.9 | 0.6 | 13.4 | 11.6 | |
| Syngnathus acus | Pooled | 75 | -17.3 | 1.2 | -14.9 | -20.5 | 13.3 | 0.7 | 15.2 | 11.6 |
| Undifferentiated | 43 | -17.1 | 1.2 | -15.0 | -22.1 | 13.4 | 0.6 | 14.8 | 12.2 | |
| Male | 8 | -17.6 | 1.6 | -16.3 | -21.4 | 13.4 | 0.6 | 14.8 | 12.2 | |
| Female | 24 | -17.6 | 1.1 | -16.3 | -19.0 | 13.1 | 0.8 | 15.2 | 11.6 | |
Table 5.
Estimated niche areas in S. abaster and S. acus collected in Metruk Tuzla. TA, SEA, and SEAc (SIBER package): Total area of the convex hull, standard ellipse area, and corrected standard ellipse with a correction for small sample sizes, respectively.
Table 5.
Estimated niche areas in S. abaster and S. acus collected in Metruk Tuzla. TA, SEA, and SEAc (SIBER package): Total area of the convex hull, standard ellipse area, and corrected standard ellipse with a correction for small sample sizes, respectively.
| Syngnathus abaster | Syngnathus acus | |||||
|---|---|---|---|---|---|---|
| Metrics | Undifferentiated | Males | Females | Undifferentiated | Males | Females |
| TA | 6.65 | 2.19 | 1.60 | 8.79 | 4.11 | 7.98 |
| SEA | 1.66 | 1.69 | 1.06 | 2.11 | 2.87 | 2.05 |
| SEAc | 1.70 | 2.03 | 1.28 | 2.16 | 3.35 | 2.14 |
Table 6.
Niche overlap estimates (NicheROVER package) showing posterior probabilities (ɑ = 0.95) that individuals from rows will be found within the niches indicated by the column header. Results (%) are provided for reproductive stages in S. abaster and S. acus specimens from Metruk Tuzla.
Table 6.
Niche overlap estimates (NicheROVER package) showing posterior probabilities (ɑ = 0.95) that individuals from rows will be found within the niches indicated by the column header. Results (%) are provided for reproductive stages in S. abaster and S. acus specimens from Metruk Tuzla.
| S. abaster | S. acus | ||||||
|---|---|---|---|---|---|---|---|
| Species | Sex condition | Female | Male | Undif. | Female | Male | Undif. |
| Syngnathus abaster | Female | - | 51 | 85 | 94 | 88 | 90 |
| Male | 34 | - | 58 | 87 | 86 | 71 | |
| Undifferentiated | 59 | 40 | - | 77 | 81 | 95 | |
| Syngnathus acus | Female | 53 | 70 | 74 | - | 87 | 82 |
| Male | 44 | 59 | 65 | 78 | - | 76 | |
| Undifferentiated | 52 | 40 | 86 | 73 | 80 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



