Submitted:
06 June 2025
Posted:
09 June 2025
You are already at the latest version
Abstract
Pulmonary arterial hypertension (PAH) is a chronic disorder with high fatality rates, and its progression is highly associated with the genetic background. Alongside pathogenic variants in genes central to the BMP/TGF-β signaling pathway, recent evidence has linked aquaporin 1 (AQP1) gene variants to PAH. While BMP9 shows promise as a PAH therapy, emerging conflicting evidence challenges this prospect. Herein, we modulated the gene expression of AQP1 and TGFB1 and examined their effect, before and after BMP9 administration, on BMP9, BMP10, BMPR2, AQP1, TGFBR1, and TGFB1 in human pulmonary microvascular endothelial cells (HPMECs) in vitro. Our results demonstrated that silencing of the AQP1 gene resulted in decreased BMPR2 mRNA and protein, downregulated TGFB1 and TGFBR1 mRNA, while tended to reduce TGFBR1 protein levels. BMP9 exogenous administration affected only TGFB1 mRNA, restoring control levels. Silencing of the TGFB1 gene downregulated BMPR2 mRNA and protein levels and affected the expression of its ligands; BMP9 mRNA and protein increased, while BMP10 mRNA levels decreased. Exogenous BMP9 treatment of TGFB1-silenced cells decreased AQP1 mRNA and protein levels. Our results indicate that modulation of AQP1 and TGFB1 genes could possibly disrupt the complex signaling pathway, and that the effects of BMP9 may be cell- and context-dependent. Together, these findings could provide a novel perspective on the interactions of the BMP/TGF-β signaling pathway.
Keywords:
PAH
; BMP9
; BMPR2
; AQP1
; TGF-β
1. Introduction
Pulmonary arterial hypertension (PAH) is a progressive and often fatal disorder characterized by structural changes in the pulmonary vascular bed, including muscularization of pre-capillary pulmonary arteries, formation of plexiform lesions, and increased pulmonary arterial pressure [1]. The pulmonary endothelium plays a central role in PAH pathogenesis [2]. Disease initiation likely stems from endothelial cell (EC) apoptosis, triggered by genetic susceptibility or injury (e.g., shear stress, chronic hypoxia, or inflammation) [3]. Bone morphogenetic protein receptor type II (BMPR2) loss-of-function variants or deficient BMP9 signaling disrupt the BMP/transforming growth factor β (TGF-β) balance, and the EC survival pathways [4,5]. This selects for ECs with hyperproliferative, apoptosis-resistant phenotypes, driving obstructive vascular remodeling [6]. Endothelial-to-mesenchymal transition (EndMT) may further amplify remodeling, though its clinical significance remains under investigation [7]. Concurrent endothelial dysfunction, manifested by vasoconstriction, prothrombotic states, and impaired angiogenesis, propagates disease progression [8,9,10].
Dysfunctional BMPR2-dependent signaling pathways [11,12] and overactive TGF-β pathways are considered hallmarks of PAH [12,13,14]. However, while pathogenic variants of genes involved in these signaling pathways represent a major causative factor in PAH, other genomic studies have identified novel genes potentially contributing to the disease. Recently, the task force of the 7th “World Symposium on Pulmonary Hypertension genetics and genomics” published their conclusions and recommendations on the level of evidence supporting PAH gene-disease associations. Among the genes associated with PAH, aquaporin 1 (AQP1) was classified as having limited evidence, which is consistent with the fact that only within the past 5 years has the AQP1 gene been linked to PAH [15]. AQP1 is membrane protein within the aquaporin family, with a significant role in water transport regulation across the cell membrane [16]. It has gained attention in the setting of PAH for its role in regulating endothelial cell permeability, migration, and proliferation [17]. Notably, our group recently described a link between AQP1 and components of the BMP signaling pathway, namely BMPR2 and TGF-β1 and demonstrated that in human pulmonary microvascular endothelial cells (HPMECs), silencing of either the BMPR2 or the AQP1 gene results in decreased expression of TGFΒ1 [18,19].
Elucidating the genetic landscape of PAH could represent potential targets for the development of effective therapeutic strategies [20]. The traditional PAH treatments primarily target the endothelin 1, nitric oxide, and prostacyclin pathways to control pulmonary vascular tone and proliferation. Although these therapies improved the patients’ symptoms and reduced the risk of deterioration, unfortunately, they have not yet provided a cure [1]. Among the novel therapeutic strategies explored, the therapeutic administration of recombinant BMP9, the main ligand of BMPR2, has been presented as a potential strategy for enhancing endothelial BMP signaling in PAH [4]. However, the downstream effects of exogenous BMP9 administration on BMP signaling molecules could depend on the expression levels of BMPR2 [21]. Recently, sotatercept, an activin-binding molecule, has demonstrated significant effectiveness and has received approval for clinical use. Unlike previous therapies, sotatercept does not target the vascular tone, but partially restores the BMPR2/TGF-β signaling imbalance, and appears to be involved in reversing the remodeling seen in PAH [22]. It becomes evident that fully elucidating the underlying molecular interactions within these pathways is crucial.
It remains unclear whether the presence of co-occurring genetic variants or the interaction between proteins involved in the BMP signaling pathway are responsible for the establishment of PAH. To date, genetic studies have identified several pathogenic variants as causal factors for the development of PAH, however, there are no studies examining the interaction of the proteins involved. In this study, we examined the effect of exogenous administration of BMP9 in regulating the BMP/TGF-β pathways in AQP1- and TGFB1-silenced HPMECs, aiming to understand their involvement in the pathological mechanisms in PAH.
2. Results
2.1. Effects of AQP1-Silencing and Exogenous Administration of BMP9 on AQP1 and BMP/TGF-β Signaling Molecules in Human Pulmonary Microvascular Endothelial Cells
Initially, our experimental model was tested for transfection specificity and efficiency employing quantitative PCR. HPMECs were transfected using a universal scrambled negative control siRNA duplex (siRNA controls). Relative AQP1 mRNA expression levels in the siRNA controls [0.64 (0.45-0.90)] did not differ to those of the non-transfected control cells [1.00 (0.72-1.16)] (p > 0.05). Cell transfection with the AQP1-specific siRNA resulted in significant decrease of AQP1 mRNA expression compared to the non-transfected control cells [0.34 (0.20-0.52) vs. 1.00 (0.72-1.16), respectively, p < 0.01; Figure 1a]. Exogenous administration of BMP9 failed to restore the reduced AQP1 mRNA levels observed in the AQP1-silenced cells [0.26 (0.17-0.45), p < 0.01, compared to the non-transfected control cells; Figure 1a]. Regarding AQP1 protein levels, silencing of the AQP1 gene resulted in decreased protein levels compared to the non-transfected control cells (0.61 ± 0.04, p < 0.05; Figure 1b), while BMP9 administration did not reverse this effect on AQP1 protein levels (0.62 ± 0.18, p < 0.05; Figure 1b compared to the non-transfected control cells).
In previously published research, we have demonstrated the effect of AQP1-silencing on BMPR2 mRNA expression [19]. At first, we reconfirmed our findings demonstrating that silencing of the AQP1 gene results in decreased BMPR2 mRNA expression compared to non-transfected control cells [0.70 (0.67-0.80) vs. 1.00 (0.85-1.04), respectively, p < 0.05; Figure 1c]. BMPR2 mRNA expression remained low even after the exogenous administration of BMP9 [0.65 (0.59-0.85), p < 0.05, compared to the non-transfected control cells; Figure 1c]. BMPR2 protein levels demonstrated a similar pattern to that of mRNA. AQP1-silenced cells presented decreased BMPR2 protein expression compared to the non-transfected control cells (0.73 ± 0.05, p < 0.05; Figure 1d). BMPR2 levels remained stable in the AQP1-silenced cells following exogenous BMP9 administration (0.52 ± 0.13, p < 0.05, compared to the non-transfected control cells; Figure 1d).
Next, we proceeded to evaluate the effect of silencing the AQP1 gene on both TGFBR1 and TGFB1 expression, a key receptor-ligand duo in the BMP/TGF-β signaling pathway. TGFBR1 mRNA expression decreased significantly in comparison to the non-transfected control cells [0.57 (0.38-0.72) vs. 1.00 (0.88-1.08), respectively, p < 0.01; Figure 1e]. Exposure to exogenous BMP9 did not affect TGFBR1 mRNA expression in the AQP1-silenced cells compared to the non-transfected control cells [0.66 (0.45-0.81), p < 0.01; Figure 1e]. A similar trend was discovered for the TGFBR1 protein levels, although the statistical significance was marginal. More specifically, in AQP1-silenced cells, TGFBR1 protein levels were decreased compared to the non-transfected control cells both pre- and post-exogenous BMP9 administration (0.53 ± 0.15, p = 0.057; Figure 1f and 0.47 ± 0.14, respectively, p = 0.057; Figure 1f). A significant decrease in TGFB1 mRNA expression was discovered following AQP1-silencing compared to non-transfected control cells [0.79 (0.76-0.85) vs. 1.00 (0.87-1.10), respectively, p < 0.05; Figure 1g]. Administration of exogenous BMP9 restored TGFB1 mRNA expression to levels comparable with the non-transfected control cells [0.90 (0.75-1.05), p > 0.05; Figure 1g]. TGFB1 protein levels following AQP1-silencing did not differ compared to the non-transfected control cells (0.94 ± 0.10, p > 0.05; Figure 1h). Exogenous administration of BMP9 did not alter TGFB1 protein levels in the AQP1-silenced cells (0.87 ± 0.21, p > 0.05; Figure 1h).
2.2. Effects of TGFB1-Silencing and Exogenous Administration of BMP9 on BMP/TGF-β Signaling Molecules and AQP1 in Human Pulmonary Microvascular Endothelial Cells
In a prior study, our group demonstrated the effect of exogenous BMP9 administration on the dysregulated BMP/TGF-β signaling pathway of HPMECs silenced for the BMPR2 gene [21]. Considering these results, we decided to explore the effects of silencing the TGFB1 gene and the administration of BMP9 on key molecules of the BMP/TGF-β signaling pathway. A universal scrambled negative control siRNA duplex (siRNA controls) was utilized for transfection specificity testing. Transfection of HPMECs with the TGFB1-specific siRNA decreased TGFB1 mRNA expression significantly in comparison to the non-transfected control cells [0.04 (0.03-0.09) vs. 1.00 (0.95-1.05), respectively, p < 0.01; Figure 2a], while administration of exogenous BMP9 failed to impact TGFB1 mRNA levels in transfected cells, when compared against non-transfect control cells [0.10 (0.04-0.25), p < 0.01; Figure 2a]. It should be noted that the siRNA control group also exhibited decreased expression levels compared to the non-transfected control group, however higher than those of the transfected cells [0.56 (0.28-0.67), p < 0.01 vs. non-transfected control cells and p < 0.01 vs TGFB1-silenced cells)]. In contrast to the mRNA expression, TGFB1 protein levels remained unaltered following TGFB1-silencing (0.94 ± 0.10, p > 0.05; Figure 2b) and after BMP9 treatment (0.87 ± 0.21, p > 0.05; Figure 2b) compared to the non-transfected control cells.
TGFB1 gene silencing did not impact TGFBR1 mRNA expression, when compared to non-transfected control cells [0.98 (0.76-1.09) vs. 1.00 (0.88-1.15), respectively, p > 0.05; Figure 2c]. Furthermore, TGFBR1 mRNA levels in the TGFB1-silenced cells remained unaltered following BMP9 administration, in comparison to the non-transfected control cells [0.88 (0.77-0.98), p > 0.05; Figure 2d]. Similarly, no changes were discovered in TGFBR1 protein levels in the TGFB1-silenced cells either pre- or post-BMP9 treatment (0.74 ± 0.12, p > 0.05; Figure 2d and 0.96 ± 0.32, p > 0.05; Figure 2d, respectively) compared to the non-transfected control cells.
Silencing the TGFB1 gene did not affect AQP1 mRNA levels compared to the non-transfected control cells [0.83 (0.44-1.01) vs. 1.00 (0.82-1.32), respectively, p > 0.05; Figure 2e], BMP9 administration decreased AQP1 mRNA expression significantly in the TGFB1-silenced cells, compared to the non-transfected control cells [0.34 (0.08-0.57), p < 0.05; Figure 2e]. In the TGFB1-silenced cells, AQP1 protein levels remained stable (0.98 ± 0.19, p > 0.05; Figure 2f). Exogenous administration of BMP9 resulted in a significant decrease of AQP1 protein compared to the non-transfected control cells (0.61 ± 0.09, p < 0.05; Figure 2f).
HPMECs silenced for TGFB1 gene expression, demonstrated a significant decrease in BMPR2 mRNA levels compared to the non-transfected control cells [0.79 (0.39-0.85) vs. 1.00 (0.80-1.17), respectively, p < 0.05; Figure 2g]. Compared to the non-transfected control cells, treatment of TGFB1-silenced cells with exogenous BMP9 did not alter BMPR2 mRNA levels [0.62 (0.43-1.08), p < 0.05; Figure 2g]. BMPR2 protein levels followed a similar pattern to that of the mRNA. Initially, TGFB1-silenced cells demonstrated significantly lower levels of the BMPR2 protein (0.71 ± 0.09, p < 0.01; Figure 2h) compared to the non-transfected control cells. BMPR2 protein levels remained low post-exogenous administration of BMP9 (0.65 ± 0.08, p < 0.01; Figure 2h).
We then proceeded to evaluate the expression of BMP9 and BMP10, the main ligands of BMPR2, following silencing of the TGFB1 gene. BMP9 mRNA levels of the TGFB1-silenced cells increased nearly 3-fold compared to the non-transfected control cells [3.06 (1.35-5.58) vs. 1.01 (0.52-1.46), respectively, p < 0.05; Figure 2i], while BMP9 protein demonstrated an increase equivalent to 29% in the TGFB1-silenced cells (1.29 ± 0.16, p < 0.05; Figure 2j). On the other hand, BMP10 mRNA levels were significantly decreased in the TGFB1-silenced cells compared to the non-transfected control cells [0.53 (0.48-0.72) vs. 1.00 (0.85-1.16), respectively, p < 0.05; Figure 2k]. Exogenous BMP9 administration did not demonstrate any effect on BMP10 mRNA levels [0.63 (0.44-0.76), p < 0.05, compared to the non-transfected control cells; Figure 2k]. In contrast to mRNA expression, BMP10 protein levels in the TGFB1-silenced cells remained comparable to those of the non-transfected control cells pre- and post-exogenous BMP9 administration (1.09 ± 0.29, p > 0.05 and 1.22 ± 0.30, p > 0.05, respectively; Figure 2l).
3. Discussion
In this study, our goal was to delve deeper into the complex interactions between AQP1 and key molecules involved in the BMP/TGF-β pathways, and also shed light into the effect of exogenous BMP9 treatment in HPMECs. We demonstrated that silencing of AQP1 downregulated the mRNA expression of BMPR2, TGFB1, and TGFBR1, and BMPR2 protein levels. On the other hand, TGFB1 silencing did not affect TGFBR1 and AQP1 expression, while it decreased BMPR2 mRNA and protein levels, and BMP10 mRNA expression. Interestingly, TGFB1-silencing increased the mRNA and protein levels of BMP9, a main ligand of BMPR2. BMP9 treatment failed to restore the reduced AQP1 and BMPR2 levels in AQP1-silenced HPMECs, while was successful in restoring TGFB1 mRNA to control levels. In contrast, in TGFB1-silenced HPMECs, BMP9 treatment did not restore the reduced BMPR2 levels, though it led to reduced AQP1 levels.
As mentioned above, endothelial dysfunction is a hallmark in PAH pathophysiology. ECs are characterized by high morphological and functional heterogeneity across vascular beds [23]. They aid lung vascular remodeling either directly by increasing cell proliferation and induction of antiapoptotic pathways, or indirectly by releasing growth factors that stimulate the proliferation and migration of pulmonary artery smooth muscle cells (PASMCs) [24,25]. Recently, Zang et al. proposed a new mechanism regarding small vessel muscularization in pulmonary hypertension, suggesting that apoptosis and proliferation occur concurrently, however in different EC subsets [26].
In this study, we performed our experiments on HPMECs, since our group has previously explored the interplay between AQP1, BMPR2, and TGFB1 in this cell type. In 2021, we were the first to report that in AQP1-silenced HPMECs, BMPR2 mRNA expression levels were reduced, suggesting a possible crosstalk between the two pathways [19]. In the same study, we also observed reduced TGFB1 mRNA levels in AQP1-silenced HPMECs [19]. We have also demonstrated a dysfunctional AQP1 in terms of reduced levels and permeability function in BMPR2-silenced HPMECs [18]. Aiming to broaden our understanding of their interaction patterns, we further explored the effect of AQP1-silencing on BMPR2, and the key receptor-ligand duo, TGFB1 and TGFBR1. Subsequently, we examined the expression patterns of BMP9, BMP10, AQP1, BMPR2, TGFB1, and TGFBR1 in TGFB1-silenced HPMECs.
Recently, a whole genome sequencing analysis of over 1,000 patients with idiopathic and heritable forms of PAH found rare and likely causal heterozygous variants in the AQP1 gene [12]. Apart from fluid hemostasis, AQP1 plays a key role in endothelial cell migration and angiogenesis [27]. Studies on various hypoxia-induced pulmonary hypertension (HPH) rodent models have demonstrated the role of AQP1 in the proliferation and migration of PASMCs. More specifically, AQP1 expression was induced in response to hypoxia, while depletion of the AQP1 gene reduced hypoxia-induced proliferation, PASMCs’ migratory potential, and induced apoptosis, suggesting that AQP1 could be a potential target for the treatment of HPH [28,29,30]. Similar findings were also reported in pulmonary microvascular endothelial cells (MVECs) in a Sugen/Hypoxia rat model of severe PH, a model that better replicated the pathophysiological features of human PAH [31]. Herein, besides reconfirming that silencing of the AQP1 gene results in reduced BMPR2 mRNA [19,21], we were able to demonstrate that it also reduces its protein levels. These findings strengthen the evidence for a previously unknown interdependent relationship between AQP1 and BMPR2, at both the mRNA and protein level in HPMECs. We also observed that silencing of the AQP1 gene resulted in reduced TGFBR1 and TGFB1 mRNA expression, while it tended to reduce TGFBR1 protein levels. These results combined with our group’s previous findings that silencing of the BMPR2 gene negatively affects TGFB1 expression [18,21], suggest that silencing of both the AQP1 and BMPR2 genes contributes to the dysregulated expression of TGFB1 mRNA.
Until now, most studies have focused on the downstream signaling of BMPR2, however, there is a need to include the role of signaling through TGF-β in PAH. Although BMP ligands and their receptors play an important role in the progression of the disease and could serve as therapeutic targets [4], agents that effectively reduce TGF-β1 activity and selective ligand traps, open up new therapeutic approaches [32,33,34,35,36]. Increased TGF-β1 levels have been found systemically and locally in both PAH patients and animal models [37]. Furthermore, studies have demonstrated that TGF-β signaling can directly inhibit the BMP-Smad pathway in PASMCs and that TGF-β ligands can function as antagonists by competing for type II receptor binding [38,39]. It has therefore been suggested that inhibition of the TGF-β signaling pathways may constitute a novel therapeutic approach in PAH.
Herein, we silenced HPMECs for the TGFB1 gene and explored the modulated expression of BMP/TGF-β-related signaling molecules. Silencing of the TGFB1 gene negatively affected BMPR2 mRNA and protein expression, while it didn’t affect AQP1 and TGFBR1 expression. These findings suggest that TGFB1 could possibly participate in the regulation of BMPR2, without possessing a significant regulatory effect on AQP1. Regarding the BMPR2 ligands, BMP9 expression levels demonstrated a significant increase in TGFB1-silenced cells, which was similar to the expression pattern described previously in HPMECs silenced for the AQP1 and BMPR2 genes. Following silencing of the TGFB1 gene, BMP10 mRNA expression decreased, in contrast to its increased mRNA expression observed in AQP1- and BMPR2-silenced HPMECs [21].
It is well established that imbalanced BMP signaling is a causative factor of PAH progression. BMP9 is a growth factor belonging to the TGF-β superfamily. It is one of the main ligands of BMPR2 and a known circulating vascular-inducing factor. Depending on the dose and vascular context, BMP9 exhibits both pro- and anti-angiogenic roles, and these functions may further differ in quiescence and dysregulated vascular beds [40]. BMP9 has been suggested as a possible therapeutic strategy for PAH based on the hypothesis that supplementation of exogenous BMP9 could enhance endothelial BMPR2 signaling and reverse PAH in in vivo models [4]. However, findings from in vivo and in vitro PAH models are thus far conflicting [41,42,43,44,45,46]. These conflicting results could suggest that its effects may vary at different stages of PAH; it might have a beneficial role in early disease stages, while its role seems to be more complicated in later stages of the disease [1].
Previously published data from our group suggested that in HPMECs silenced for the BMPR2 gene, the effect of the exogenous administration of BMP9 on molecules participating in the BMP/TGF-β signaling pathways seems to depend on BMPR2 mRNA expression levels [21]. In the present study, we observed that the exogenous administration of BMP9 in AQP1-silenced HPMECs did not revert the decreased mRNA and protein levels of AQP1, BMPR2, and TGFBR1, while BMP9 treatment restored TGFB1 mRNA levels. It is plausible that BMP9’s effect on AQP1 is likely a result of the complex interplay between BMPR2 expression levels and an unknown molecular mechanism regulating its expression. On the other hand, in the TGFB1-silenced HPMECs, exogenous administration of BMP9 reduced the pre-BMP9 treatment AQP1 mRNA and protein levels, while it did not reverse the reduced BMPR2 levels. Hence, BMP9 does not restore BMPR2 expression when either AQP1 or TGFB1 genes are silenced in HPMECs, thus raising the possibility that its effect on BMPR2 is independent of the molecular mechanism by which BMPR2 expression is downregulated. It is possible that the effects of BMP9 could depend on the expression levels of the genes silenced, and that these effects apply in the cell type studied in our work.
Interestingly, Szulcek et al. examined the effect of BMP9 in HPMECs isolated from pleura-free peripheral lung tissues of PAH patients [47]. They found that HPMECs from PAH patients responded to BMP9 stimulation, exhibiting an 8-fold increase in BMPR2 levels, differing from the response observed in PAECs and endothelial colony-forming cells (ECFCs), thus suggesting that the lung tissue microenvironment could affect activation and outcome of BMP-dependent signaling pathways in PAH. More specifically, in PAH MVECs, BMP9 activated a persistent EndMT signaling, leading to the loss of endothelial-specific markers, gaining of mesenchymal characteristics, and decreased barrier integrity [47].
As stated above, the focus of our study was to explore the effects of silencing the AQP1 and TGFB1 genes in HPMECs on the BMP/TGF-β signaling pathway. While AQP1 mRNA and protein expression were decreased post-transfection, the reduction of TGFB1 expression levels was evident only at the mRNA level. Knowing that TGFB1 is secreted in an inactive form, bound to TGF binding proteins, we hypothesize that its effects could persist even after mRNA expression is silenced [48]. Though a decrease in TGFB1 protein expression could be observed at 48-72 h post-transfection, in our set of experiments, cells were harvested at 48 h due to increased cytotoxicity. Furthermore, among the limitations of this study is that our experiments mainly focused on the genetic and protein expression of molecules known to interact with the BMP/TGF-β signaling pathway in a single in vitro cell model. Additional studies performed on established animal PAH models or cell samples extracted from PAH patients could be beneficial. While our study provides novel insights into how BMP9 treatment affects the BMP/TGF-β signaling pathway, it should be stated that these results reflect specifically the ECs of the pulmonary microvasculature. Further studies exploring the effect of BMP9 on other cell types known to participate in the pathophysiology of PAH could demonstrate a differential effect on the genetic level. Importantly, additional clinical studies should be employed to further investigate the therapeutic potential of BMP9. It must be noted that one of the key aspects of our study is the fact that it provides novel insight into the established interaction of AQP1 and BMPR2. To the best of our knowledge, the present study is the first to demonstrate the effect of BMP9 on the complex interactions between AQP1 and key molecules of the BMP/TGF-β signaling pathway, at a molecular and protein level in HPMECs in the setting of either hindered AQP1 or TGFB1 gene expression.
4. Materials and Methods
4.1. Cell Culture, Transfection, and Treatment with BMP9
All experiments were performed using the HPMEC-ST1.6R cell line, originated from normal lung tissue distant from the tumor site of a 63-year-old male donor [49]. Cells were cultured in a controlled environment at a standard temperature of 37°C with stable 5% CO2 levels, using a fully supplemented M199 medium (Biosera, Cholet, France), as described previously [21]. Cell transfection for the two siRNAs was carried out as previously described in detail [21]. In 70% confluent cells, the specific siTran/siRNA (Origene) mixture was added, and cells were cultured in serum-free medium for 5 h before being replaced with fully supplemented M199 medium. HPMECs were treated with BMP9 (5 ng/mL) (OriGene, Rockville, MD, USA) 24 h post-transfection. Following a 24 h incubation, cells were harvested for total RNA and total protein extraction.
4.2. RNA and Protein Extraction
To ensure transfection efficiency, total RNA and total protein were extracted from cells cultured in the same plate. Total RNA was extracted using the TRI reagent (Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer’s instructions. The concentration and quality of the total extracted RNA were determined at 260 and 280 nm, using the NanodropTM spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). For protein extraction, samples were sonicated with an ultrasonic lab homogenizer (Thermo Fisher Scientific, Waltham, MA, USA), following pellet resuspension in lysis buffer consisting of 250 mM sucrose, 10 mM Tris-HCl pH 7.4, 10 mM NaCl, 1 mM Ethylenediaminetetraacetic acid (EDTA), 0.1 mM phenylmethylsulfonyl fluoride (PMSF), and 1% v/v Triton-X 100. Cell homogenates were centrifuged at 10,000 g at 4 °C for 20 min, and the supernatants containing the extracted proteins were collected. Total protein concentration was determined using the bicinchoninic acid (BCA) method [50] (Thermo Fisher Scientific, Waltham, MA, USA).
4.3. Reverse Transcription and Quantitative Real-Time PCR
From each sample, 100ng of total RNA were reverse transcribed into single stranded cDNA (Nippon Genetics, Duren, Germany), following the manufacturer’s instructions. Quantitative real-time polymerase chain reaction (qPCR) method was employed, using Kapa SYBR® Green PCR Master Mix (Sigma-Aldrich, St Louis, MO, USA), to measure AQP1, BMP9, BMP10, BMPR2, TGFB1, TGFBR1, and GAPDH mRNA expression levels. The analysis was carried out on a CFX Connect thermocycler (Bio-Rad Laboratories). The specific primer sets designed for the targeted genes are listed in Table 1. Non-transfected control cells were used as a calibrator, and the relative gene expression levels were determined through the comparative CT method 2-ΔΔCT [51]. Expression of the GAPDH housekeeping gene was used for normalization purposes.
4.4. SDS-Polyacrylamide Gel Electrophoresis (PAGE) and Immunoblotting
SDS–PAGE was performed using polyacrylamide slab gels on a “Biorad Mini Protean II” electrophoresis apparatus (Bio-Rad Laboratories, Inc., Hercules, CA, USA), as previously described [52]. Subsequently, samples were transferred onto an 0.45 μL pore size Immobilon-P PVDF membrane (MilliporeSigma, Burlington, MA, USA), using a wet transfer apparatus (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Immunological detection followed using specific antibodies for AQP1 (MilliporeSigma, Burlington, MA, USA), BMP9 (MilliporeSigma, Burlington, MA, USA), BMP10 (Affinity Biosciences, Cincinnati, OH, USA), BMPR2 (Cell Signaling Technology, MA, USA), TGFB1 (Affinity Biosciences, Cincinnati, OH, USA), and TGFBR1 (Cell Signaling Technology, MA, USA). Actin (MilliporeSigma, Burlington, MA, USA) and b-tubulin (Santa Cruz Biotechnology, TX, USA) were used as loading controls. Relative protein expression was estimated by densitometry with the use of the iBright Analysis Software (Thermo Fisher Scientific, Waltham, MA, USA).
4.5. Statistical Analysis
Data are presented as box plots, presenting median values with interquartile range (IQR), or as bar plots with mean ± SEM. The non-parametric Mann-Witney test was employed for statistical analysis. All tests were carried out using the GraphPad Prism 9 software (GraphPad Software, San Diego, CA, USA). All p-values are two-sided; statistical significance was set at p < 0.05.
5. Conclusions
Our findings suggest that in HPMECs, silencing of either BMPR2, AQP1, or TGFB1 genes, along with BMP9 administration alters the expression of key molecules in the BMP/TGF-β signaling pathway. Disruptions in this complex pathway may directly impact endothelial function, though this finding seems to be cell- and context-dependent. While our results further support the proposed interplay between AQP1 and BMPR2 in the canonical BMP/TGF-β signaling pathway, the specifics of this interaction are unknown. Additionally, though the function of BMP9 has yet to be fully understood in different PAH models and stages of the disease, its effect seems to differ depending on the genetic background and/or the condition of the vascular bed. Further studies across different cell types known to participate in PAH development and progression could unravel the complex interactions, leading to a better understanding of BMP9’s therapeutic potential.
Author Contributions
Conceptualization, S.E.O. and A.G.V.; methodology, C.K., N.S.L., K.A.P., and A.G.V.; validation, I.D., A.K. and A.G.V.; formal analysis, C.K., N.S.L., K.A.P., and A.G.V.; investigation, C.K., N.S.L., and A.G.V.; resources, S.E.O., and A.G.V.; data curation, C.K. and N.S.L.; writing—original draft preparation, C.K., N.S.L. and K.A.P.; writing—review and editing, I.D., A.K., D.L., S.E.O. and A.G.V.; visualization, D.L., S.E.O. and A.G.V.; supervision, D.L., S.E.O. and A.G.V.; project administration, I.D., A.K., D.L., S.E.O. and A.G.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Acknowledgments
The HPMEC-ST1.6R cell line used in the experiments was kindly gifted from Dr. Ronald E Unger, Johannes Gutenberg-Universität Mainz. The authors acknowledge support from the “Stavros Niarchos Foundation” (C.K., N.S.L. and A.G.V.).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Li, W.; Quigley, K. Bone morphogenetic protein signalling in pulmonary arterial hypertension: revisiting the BMPRII connection. Biochemical Society transactions 2024, 52, 1515–1528. [Google Scholar] [CrossRef] [PubMed]
- Huertas, A.; Guignabert, C.; Barberà, J.A.; Bärtsch, P.; Bhattacharya, J.; Bhattacharya, S.; Bonsignore, M.R.; Dewachter, L.; Dinh-Xuan, A.T.; Dorfmüller, P.; et al. Pulmonary vascular endothelium: the orchestra conductor in respiratory diseases: Highlights from basic research to therapy. The European respiratory journal 2018, 51. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.E.; Cober, N.D.; Dai, Z.; Stewart, D.J.; Zhao, Y.Y. Endothelial cells in the pathogenesis of pulmonary arterial hypertension. The European respiratory journal 2021, 58. [Google Scholar] [CrossRef]
- Long, L.; Ormiston, M.L.; Yang, X.; Southwood, M.; Gräf, S.; Machado, R.D.; Mueller, M.; Kinzel, B.; Yung, L.M.; Wilkinson, J.M.; et al. Selective enhancement of endothelial BMPR-II with BMP9 reverses pulmonary arterial hypertension. Nature medicine 2015, 21, 777–785. [Google Scholar] [CrossRef]
- Morrell, N.W.; Aldred, M.A.; Chung, W.K.; Elliott, C.G.; Nichols, W.C.; Soubrier, F.; Trembath, R.C.; Loyd, J.E. Genetics and genomics of pulmonary arterial hypertension. The European respiratory journal 2019, 53. [Google Scholar] [CrossRef] [PubMed]
- Marinho, Y.; Villarreal, E.S.; Loya, O.; Oliveira, S.D. Mechanisms of lung endothelial cell injury and survival in pulmonary arterial hypertension. American journal of physiology. Lung cellular and molecular physiology 2024, 327, L972–l983. [Google Scholar] [CrossRef]
- Gorelova, A.; Berman, M.; Al Ghouleh, I. Endothelial-to-Mesenchymal Transition in Pulmonary Arterial Hypertension. Antioxidants & redox signaling 2021, 34, 891–914. [Google Scholar] [CrossRef]
- Humbert, M.; Guignabert, C.; Bonnet, S.; Dorfmüller, P.; Klinger, J.R.; Nicolls, M.R.; Olschewski, A.J.; Pullamsetti, S.S.; Schermuly, R.T.; Stenmark, K.R.; et al. Pathology and pathobiology of pulmonary hypertension: state of the art and research perspectives. The European respiratory journal 2019, 53. [Google Scholar] [CrossRef]
- Kurakula, K.; Smolders, V.F.E.D.; Tura-Ceide, O.; Jukema, J.W.; Quax, P.H.A.; Goumans, M.-J. Endothelial Dysfunction in Pulmonary Hypertension: Cause or Consequence? Biomedicines 2021, 9. [Google Scholar] [CrossRef]
- Cober, N.D.; VandenBroek, M.M.; Ormiston, M.L.; Stewart, D.J. Evolving Concepts in Endothelial Pathobiology of Pulmonary Arterial Hypertension. Hypertension 2022, 79, 1580–1590. [Google Scholar] [CrossRef]
- Evans, J.D.; Girerd, B.; Montani, D.; Wang, X.J.; Galiè, N.; Austin, E.D.; Elliott, G.; Asano, K.; Grünig, E.; Yan, Y.; et al. BMPR2 mutations and survival in pulmonary arterial hypertension: an individual participant data meta-analysis. The Lancet. Respiratory medicine 2016, 4, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Gräf, S.; Haimel, M.; Bleda, M.; Hadinnapola, C.; Southgate, L.; Li, W.; Hodgson, J.; Liu, B.; Salmon, R.M.; Southwood, M.; et al. Identification of rare sequence variation underlying heritable pulmonary arterial hypertension. Nature communications 2018, 9, 1416. [Google Scholar] [CrossRef]
- Andre, P.; Joshi, S.R.; Briscoe, S.D.; Alexander, M.J.; Li, G.; Kumar, R. Therapeutic Approaches for Treating Pulmonary Arterial Hypertension by Correcting Imbalanced TGF-β Superfamily Signaling. Frontiers in medicine 2021, 8, 814222. [Google Scholar] [CrossRef] [PubMed]
- Bousseau, S.; Sobrano Fais, R.; Gu, S.; Frump, A.; Lahm, T. Pathophysiology and new advances in pulmonary hypertension. BMJ medicine 2023, 2, e000137. [Google Scholar] [CrossRef]
- Austin, E.D.; Aldred, M.A.; Alotaibi, M.; Gräf, S.; Nichols, W.C.; Trembath, R.C.; Chung, W.K. Genetics and precision genomics approaches to pulmonary hypertension. The European respiratory journal 2024, 64. [Google Scholar] [CrossRef]
- Lotsios, N.S.; Keskinidou, C.; Dimopoulou, I.; Kotanidou, A.; Orfanos, S.E.; Vassiliou, A.G. Aquaporin Expression and Regulation in Clinical and Experimental Sepsis. International journal of molecular sciences 2023, 25. [Google Scholar] [CrossRef] [PubMed]
- Meli, R.; Pirozzi, C.; Pelagalli, A. New Perspectives on the Potential Role of Aquaporins (AQPs) in the Physiology of Inflammation. Frontiers in physiology 2018, 9, 101. [Google Scholar] [CrossRef]
- Vassiliou, A.G.; Keskinidou, C.; Kotanidou, A.; Frantzeskaki, F.; Dimopoulou, I.; Langleben, D.; Orfanos, S.E. Knockdown of bone morphogenetic protein type II receptor leads to decreased aquaporin 1 expression and function in human pulmonary microvascular endothelial cells. Canadian Journal of Physiology and Pharmacology 2020, 98, 834–839. [Google Scholar] [CrossRef]
- Vassiliou, A.G.; Keskinidou, C.; Kotanidou, A.; Frantzeskaki, F.; Dimopoulou, I.; Langleben, D.; Orfanos, S.E. Decreased bone morphogenetic protein type II receptor and BMP-related signalling molecules’ expression in aquaporin 1-silenced human pulmonary microvascular endothelial cells. Hellenic journal of cardiology : HJC = Hellenike kardiologike epitheorese 2021, 62, 84–86. [Google Scholar] [CrossRef]
- Adu-Amankwaah, J.; You, Q.; Liu, X.; Jiang, J.; Yang, D.; Liu, K.; Yuan, J.; Wang, Y.; Hu, Q.; Tan, R. Pulmonary Hypertension: Molecular Mechanisms and Clinical Studies. MedComm 2025, 6, e70134. [Google Scholar] [CrossRef]
- Lotsios, N.S.; Keskinidou, C.; Dimopoulou, I.; Kotanidou, A.; Langleben, D.; Orfanos, S.E.; Vassiliou, A.G. Effects of Modulating BMP9, BMPR2, and AQP1 on BMP Signaling in Human Pulmonary Microvascular Endothelial Cells. International journal of molecular sciences 2024, 25. [Google Scholar] [CrossRef]
- Preston, I.R.; Lewis, D.; Gomberg-Maitland, M. Using Sotatercept in the Care of Patients With Pulmonary Arterial Hypertension. Chest 2024, 166, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Kalucka, J.; de Rooij, L.; Goveia, J.; Rohlenova, K.; Dumas, S.J.; Meta, E.; Conchinha, N.V.; Taverna, F.; Teuwen, L.A.; Veys, K.; et al. Single-Cell Transcriptome Atlas of Murine Endothelial Cells. Cell 2020, 180, 764–779.e720. [Google Scholar] [CrossRef] [PubMed]
- Masri, F.A.; Xu, W.; Comhair, S.A.; Asosingh, K.; Koo, M.; Vasanji, A.; Drazba, J.; Anand-Apte, B.; Erzurum, S.C. Hyperproliferative apoptosis-resistant endothelial cells in idiopathic pulmonary arterial hypertension. American journal of physiology. Lung cellular and molecular physiology 2007, 293, L548–554. [Google Scholar] [CrossRef]
- Guignabert, C.; Alvira, C.M.; Alastalo, T.P.; Sawada, H.; Hansmann, G.; Zhao, M.; Wang, L.; El-Bizri, N.; Rabinovitch, M. Tie2-mediated loss of peroxisome proliferator-activated receptor-gamma in mice causes PDGF receptor-beta-dependent pulmonary arterial muscularization. American journal of physiology. Lung cellular and molecular physiology 2009, 297, L1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yaoita, N.; Tabuchi, A.; Liu, S.; Chen, S.H.; Li, Q.; Hegemann, N.; Li, C.; Rodor, J.; Timm, S.; et al. Endothelial Heterogeneity in the Response to Autophagy Drives Small Vessel Muscularization in Pulmonary Hypertension. Circulation 2024, 150, 466–487. [Google Scholar] [CrossRef]
- Saadoun, S.; Papadopoulos, M.C.; Hara-Chikuma, M.; Verkman, A.S. Impairment of angiogenesis and cell migration by targeted aquaporin-1 gene disruption. Nature 2005, 434, 786–792. [Google Scholar] [CrossRef]
- Leggett, K.; Maylor, J.; Undem, C.; Lai, N.; Lu, W.; Schweitzer, K.; King, L.S.; Myers, A.C.; Sylvester, J.T.; Sidhaye, V.; et al. Hypoxia-induced migration in pulmonary arterial smooth muscle cells requires calcium-dependent upregulation of aquaporin 1. American journal of physiology. Lung cellular and molecular physiology 2012, 303, L343–353. [Google Scholar] [CrossRef]
- Schuoler, C.; Haider, T.J.; Leuenberger, C.; Vogel, J.; Ostergaard, L.; Kwapiszewska, G.; Kohler, M.; Gassmann, M.; Huber, L.C.; Brock, M. Aquaporin 1 controls the functional phenotype of pulmonary smooth muscle cells in hypoxia-induced pulmonary hypertension. Basic research in cardiology 2017, 112, 30. [Google Scholar] [CrossRef]
- Liu, M.; Liu, Q.; Pei, Y.; Gong, M.; Cui, X.; Pan, J.; Zhang, Y.; Liu, Y.; Liu, Y.; Yuan, X.; et al. Aqp-1 Gene Knockout Attenuates Hypoxic Pulmonary Hypertension of Mice. Arteriosclerosis, Thrombosis, and Vascular Biology 2019, 39, 48–62. [Google Scholar] [CrossRef]
- Yun, X.; Philip, N.M.; Jiang, H.; Smith, Z.; Huetsch, J.C.; Damarla, M.; Suresh, K.; Shimoda, L.A. Upregulation of Aquaporin 1 Mediates Increased Migration and Proliferation in Pulmonary Vascular Cells From the Rat SU5416/Hypoxia Model of Pulmonary Hypertension. Frontiers in physiology 2021, Volume 12, 2021. [Google Scholar] [CrossRef]
- Ogo, T.; Chowdhury, H.M.; Yang, J.; Long, L.; Li, X.; Torres Cleuren, Y.N.; Morrell, N.W.; Schermuly, R.T.; Trembath, R.C.; Nasim, M.T. Inhibition of overactive transforming growth factor-β signaling by prostacyclin analogs in pulmonary arterial hypertension. American journal of respiratory cell and molecular biology 2013, 48, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Zuo, C.; He, Y.; Chen, G.; Piao, L.; Zhang, J.; Xiao, B.; Shen, Y.; Tang, J.; Kong, D.; et al. EP3 receptor deficiency attenuates pulmonary hypertension through suppression of Rho/TGF-β1 signaling. The Journal of clinical investigation 2015, 125, 1228–1242. [Google Scholar] [CrossRef]
- Yung, L.M.; Nikolic, I.; Paskin-Flerlage, S.D.; Pearsall, R.S.; Kumar, R.; Yu, P.B. A Selective Transforming Growth Factor-β Ligand Trap Attenuates Pulmonary Hypertension. American journal of respiratory and critical care medicine 2016, 194, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Bellaye, P.S.; Yanagihara, T.; Granton, E.; Sato, S.; Shimbori, C.; Upagupta, C.; Imani, J.; Hambly, N.; Ask, K.; Gauldie, J.; et al. Macitentan reduces progression of TGF-β1-induced pulmonary fibrosis and pulmonary hypertension. The European respiratory journal 2018, 52. [Google Scholar] [CrossRef]
- Zabini, D.; Granton, E.; Hu, Y.; Miranda, M.Z.; Weichelt, U.; Breuils Bonnet, S.; Bonnet, S.; Morrell, N.W.; Connelly, K.A.; Provencher, S.; et al. Loss of SMAD3 Promotes Vascular Remodeling in Pulmonary Arterial Hypertension via MRTF Disinhibition. American journal of respiratory and critical care medicine 2018, 197, 244–260. [Google Scholar] [CrossRef]
- Rol, N.; Kurakula, K.B.; Happé, C.; Bogaard, H.J.; Goumans, M.J. TGF-β and BMPR2 Signaling in PAH: Two Black Sheep in One Family. International journal of molecular sciences 2018, 19. [Google Scholar] [CrossRef]
- Upton, P.D.; Davies, R.J.; Tajsic, T.; Morrell, N.W. Transforming growth factor-β(1) represses bone morphogenetic protein-mediated Smad signaling in pulmonary artery smooth muscle cells via Smad3. American journal of respiratory cell and molecular biology 2013, 49, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Aykul, S.; Martinez-Hackert, E. Transforming Growth Factor-β Family Ligands Can Function as Antagonists by Competing for Type II Receptor Binding. The Journal of biological chemistry 2016, 291, 10792–10804. [Google Scholar] [CrossRef]
- García de Vinuesa, A.; Abdelilah-Seyfried, S.; Knaus, P.; Zwijsen, A.; Bailly, S. BMP signaling in vascular biology and dysfunction. Cytokine & growth factor reviews 2016, 27, 65–79. [Google Scholar] [CrossRef]
- Tu, L.; Desroches-Castan, A.; Mallet, C.; Guyon, L.; Cumont, A.; Phan, C.; Robert, F.; Thuillet, R.; Bordenave, J.; Sekine, A.; et al. Selective BMP-9 Inhibition Partially Protects Against Experimental Pulmonary Hypertension. Circulation Research 2019, 124, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Upton, P.D.; Davies, R.J.; Trembath, R.C.; Morrell, N.W. Bone morphogenetic protein (BMP) and activin type II receptors balance BMP9 signals mediated by activin receptor-like kinase-1 in human pulmonary artery endothelial cells. The Journal of biological chemistry 2009, 284, 15794–15804. [Google Scholar] [CrossRef]
- Nikolic, I.; Yung, L.M.; Yang, P.; Malhotra, R.; Paskin-Flerlage, S.D.; Dinter, T.; Bocobo, G.A.; Tumelty, K.E.; Faugno, A.J.; Troncone, L.; et al. Bone Morphogenetic Protein 9 Is a Mechanistic Biomarker of Portopulmonary Hypertension. American journal of respiratory and critical care medicine 2019, 199, 891–902. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Rice, M.; Swist, S.; Kubin, T.; Wu, F.; Wang, S.; Kraut, S.; Weissmann, N.; Böttger, T.; Wheeler, M.; et al. BMP9 and BMP10 Act Directly on Vascular Smooth Muscle Cells for Generation and Maintenance of the Contractile State. Circulation 2021, 143, 1394–1410. [Google Scholar] [CrossRef]
- Bouvard, C.; Tu, L.; Rossi, M.; Desroches-Castan, A.; Berrebeh, N.; Helfer, E.; Roelants, C.; Liu, H.; Ouarné, M.; Chaumontel, N.; et al. Different cardiovascular and pulmonary phenotypes for single- and double-knock-out mice deficient in BMP9 and BMP10. Cardiovascular Research 2022, 118, 1805–1820. [Google Scholar] [CrossRef] [PubMed]
- Theilmann, A.L.; Hawke, L.G.; Hilton, L.R.; Whitford, M.K.M.; Cole, D.V.; Mackeil, J.L.; Dunham-Snary, K.J.; Mewburn, J.; James, P.D.; Maurice, D.H.; et al. Endothelial BMPR2 Loss Drives a Proliferative Response to BMP (Bone Morphogenetic Protein) 9 via Prolonged Canonical Signaling. Arterioscler Thromb Vasc Biol 2020, 40, 2605–2618. [Google Scholar] [CrossRef]
- Szulcek, R.; Sanchez-Duffhues, G.; Rol, N.; Pan, X.; Tsonaka, R.; Dickhoff, C.; Yung, L.M.; Manz, X.D.; Kurakula, K.; Kiełbasa, S.M.; et al. Exacerbated inflammatory signaling underlies aberrant response to BMP9 in pulmonary arterial hypertension lung endothelial cells. Angiogenesis 2020, 23, 699–714. [Google Scholar] [CrossRef]
- Krafft, E.; Lybaert, P.; Roels, E.; Laurila, H.P.; Rajamäki, M.M.; Farnir, F.; Myllärniemi, M.; Day, M.J.; Mc Entee, K.; Clercx, C. Transforming growth factor beta 1 activation, storage, and signaling pathways in idiopathic pulmonary fibrosis in dogs. Journal of veterinary internal medicine 2014, 28, 1666–1675. [Google Scholar] [CrossRef]
- Krump-Konvalinkova, V.; Bittinger, F.; Unger, R.E.; Peters, K.; Lehr, H.A.; Kirkpatrick, C.J. Generation of human pulmonary microvascular endothelial cell lines. Laboratory investigation; a journal of technical methods and pathology 2001, 81, 1717–1727. [Google Scholar] [CrossRef]
- Lleu, P.L.; Rebel, G. Interference of Good’s buffers and other biological buffers with protein determination. Analytical biochemistry 1991, 192, 215–218. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (San Diego, Calif.) 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of silencing the AQP1 gene and exogenous administration of BMP9 on AQP1 and BMP/TGF-β signaling molecules in human pulmonary microvascular endothelial cells. HPMECs were silenced for the AQP1 gene, and the relative mRNA and protein expression of AQP1 (a,b), BMPR2 (c,d), TGFBR1 (e,f), and TGFB1 (g,h) were estimated before and after the exogenous administration of BMP9. Relative mRNA expression is shown in box plots (line in the middle, median values; box edges, 25th and 75th percentiles; whiskers, range of values), and relative protein expression with bar plots (mean ± SEM). The data presented are from at least three independent experiments. Specificity (siRNA negative control), efficiency (AQP1 siRNA), and the effect of BMP9 exogenous administration on the non-transfected controls were tested each time as to ensure the consistency and reproducibility across the independent experiments. Statistical analysis was performed using the Mann–Whitney test. *, p < 0.05; **, p < 0.01 compared to non-transfected control HPMECs.
Figure 1.
Effects of silencing the AQP1 gene and exogenous administration of BMP9 on AQP1 and BMP/TGF-β signaling molecules in human pulmonary microvascular endothelial cells. HPMECs were silenced for the AQP1 gene, and the relative mRNA and protein expression of AQP1 (a,b), BMPR2 (c,d), TGFBR1 (e,f), and TGFB1 (g,h) were estimated before and after the exogenous administration of BMP9. Relative mRNA expression is shown in box plots (line in the middle, median values; box edges, 25th and 75th percentiles; whiskers, range of values), and relative protein expression with bar plots (mean ± SEM). The data presented are from at least three independent experiments. Specificity (siRNA negative control), efficiency (AQP1 siRNA), and the effect of BMP9 exogenous administration on the non-transfected controls were tested each time as to ensure the consistency and reproducibility across the independent experiments. Statistical analysis was performed using the Mann–Whitney test. *, p < 0.05; **, p < 0.01 compared to non-transfected control HPMECs.
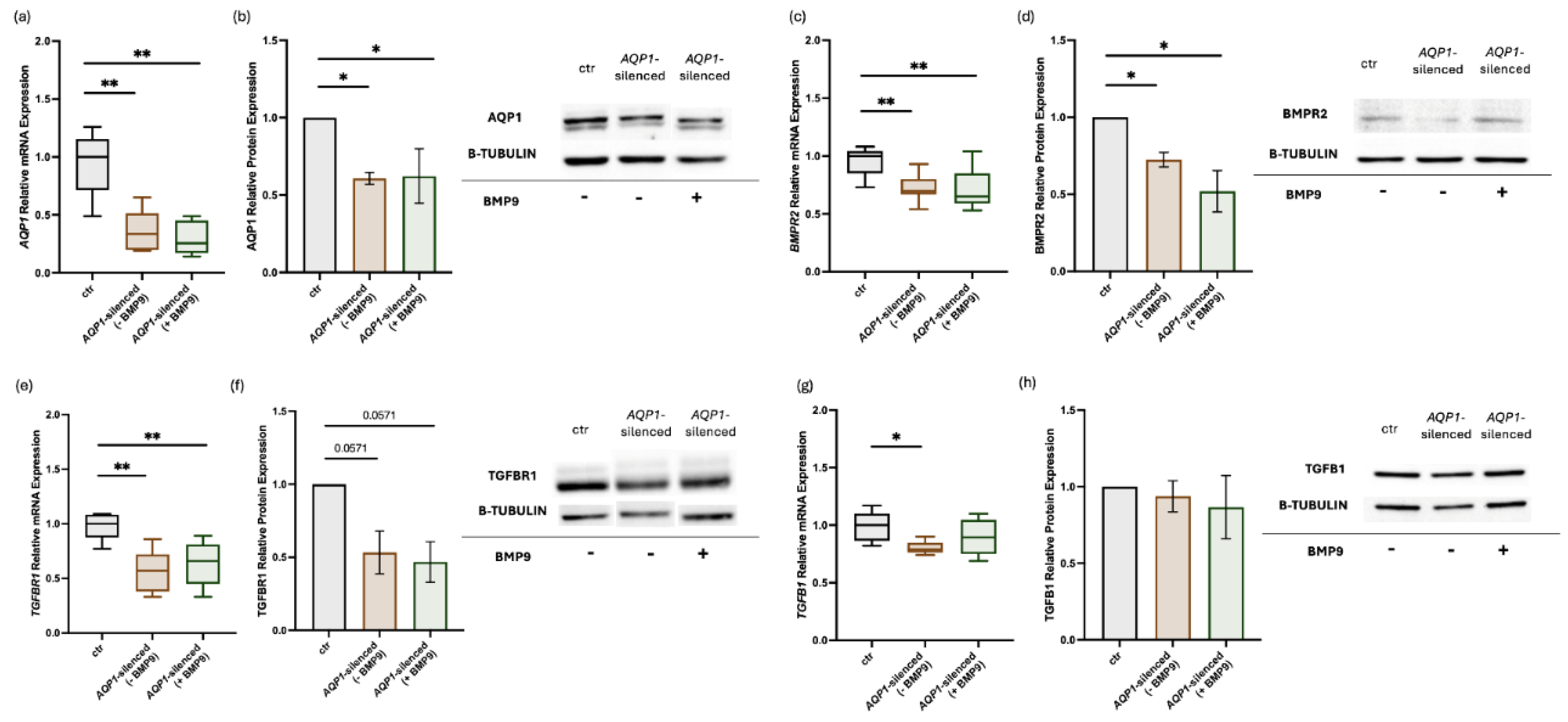
Figure 2.
Effects of silencing the TGFB1 gene and exogenous administration of BMP9 on BMP/TGF-β signaling molecules and AQP1 in human pulmonary microvascular endothelial cells. HPMECs were silenced for the TGFB1 gene, and the relative mRNA and protein expression of TGFB1 (a,b), TGFBR1 (c,d), AQP1 (e,f), BMPR2 (g,h), BMP9 (i,j), and BMP10 (k,l) were estimated before and after the exogenous administration of BMP9. Relative mRNA expression is shown in box plots (line in the middle, median values; box edges, 25th and 75th percentiles; whiskers, range of values), and relative protein expression with bar plots (mean ± SEM). Findings from at least three independent experiments are presented. Specificity (siRNA negative control), efficiency (AQP1 siRNA), and the effect of BMP9 exogenous administration on the non-transfected controls were tested each time as to ensure the consistency and reproducibility across the independent experiments. Statistical analysis was performed using the Mann–Whitney test. *, p < 0.05; **, p < 0.01 compared to non-transfected control HPMECs.
Figure 2.
Effects of silencing the TGFB1 gene and exogenous administration of BMP9 on BMP/TGF-β signaling molecules and AQP1 in human pulmonary microvascular endothelial cells. HPMECs were silenced for the TGFB1 gene, and the relative mRNA and protein expression of TGFB1 (a,b), TGFBR1 (c,d), AQP1 (e,f), BMPR2 (g,h), BMP9 (i,j), and BMP10 (k,l) were estimated before and after the exogenous administration of BMP9. Relative mRNA expression is shown in box plots (line in the middle, median values; box edges, 25th and 75th percentiles; whiskers, range of values), and relative protein expression with bar plots (mean ± SEM). Findings from at least three independent experiments are presented. Specificity (siRNA negative control), efficiency (AQP1 siRNA), and the effect of BMP9 exogenous administration on the non-transfected controls were tested each time as to ensure the consistency and reproducibility across the independent experiments. Statistical analysis was performed using the Mann–Whitney test. *, p < 0.05; **, p < 0.01 compared to non-transfected control HPMECs.
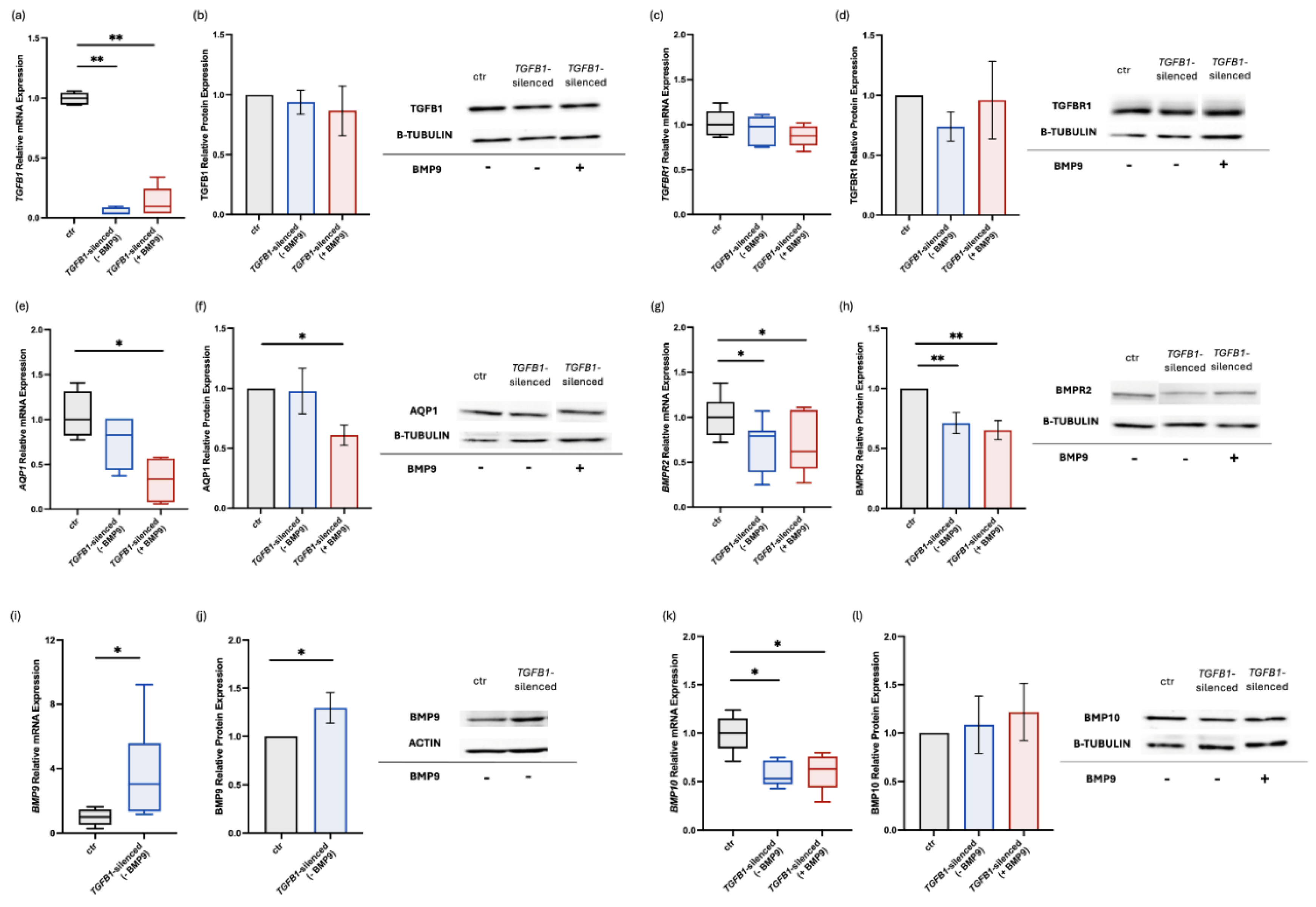

Table 1.
Gene-specific primer pairs used in quantitative real-time PCR experiments.
| Gene | Sequence (5’-3’) | nt | |
|---|---|---|---|
| AQP1 | F | 5’-TATGCGTGCTGGCTACTACCGA-3’ | 22 |
| R | 5’-GGTTAATCCCACAGCCAGTGTAG-3’ | 23 | |
| BMP9 | F | 5’-CCTGCCCTTCTTTGTTGTCTTCTC-3’ | 24 |
| R | 5’-TGACTGCTCTCACCTGCCTCTGTG-3’ | 24 | |
| BMP10 | F | 5’-AAGCCTATGAATGCCGTGGTG-3’ | 21 |
| R | 5’-AGGCCTGGATAATTGCATGCTT-3’ | 22 | |
| BMPR2 | F | 5’-CCACCTCCTGACACAACACC-3’ | 20 |
| R | 5’-TGTGAAGACCTTGTTTACGGT-3’ | 21 | |
| GAPDH | F | 5’-ATGGGGAAGGTGAAGGTCG-3’ | 19 |
| R | 5’-GGGGTCATTGATGGCAACAATA-3’ | 22 | |
| TGFB1 | F | 5’-GCGTGCTAATGGTGGAAAC-3’ | 19 |
| R | 5’-CGGTGACATCAAAAGATAACCAC-3’ | 23 | |
| TGFBR1 | F | 5’-GACAACGTCAGGTTCTGGCTCA-3’ | 22 |
| R | 5’-CCGCCACTTTCCTCTCCAAACT-3’ | 22 |
Abbreviations: AQP1, aquaporin 1; BMP9, bone morphogenetic protein 9; BMP10, bone morphogenetic protein 10; BMPR2, bone morphogenetic protein receptor type 2; GAPDH, glyceraldehyde-3-phospahte dehydrogenase; TGFB1, transforming growth factor beta 1; TGFBR1, transforming growth factor beta receptor 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



