
Submitted:
04 June 2025
Posted:
04 June 2025
You are already at the latest version
Abstract
Glypican-1 (GPC1) is a heparan sulfate proteoglycan that plays a critical role in regulating various signaling pathways and tumor development. Overexpression of GPC1 promotes tumor cell proliferation and invasiveness, and is associated with poor clinical outcomes. Therefore, anti-GPC1 monoclonal antibodies (mAbs) have been developed in various modalities for tumor therapy. Here, we developed novel anti-GPC1 mAbs using a flow cytometry-based high-throughput screening approach, the Cell-Based Immunization and Screening (CBIS) method. A clone G1Mab-28 (IgG1, κ) reacted with GPC1-overexpressed Chinese hamster ovary-K1 (CHO/GPC1), but not parental CHO-K1, in flow cytometry. Furthermore, G1Mab-28 recognizes endogenous GPC1-expressing human esophageal squamous cell carcinoma KYSE770 cell line. Furthermore, G1Mab-28 specifically recognized only CHO/GPC1, but not other GPC family-overexpressed CHO-K1. The dissociation constant values of G1Mab-28 for CHO/GPC1 and KYSE770 were determined to be 3.3 × 10⁻8 M and 4.6 × 10⁻⁹ M, respectively. Moreover, G1Mab-28 is suitable for western blotting and immunohistochemistry. G1Mab-28, established by the CBIS method, is versatile for basic research and is expected to contribute to the antibody-based tumor therapy.
Keywords:
Glypican-1
; monoclonal antibody
; Cell-Based Immunization and Screening
; flow cytometry
; immunohistochemistry
1. Introduction
Glypicans are a significant class of extracellular matrix-associated heparan sulfate (HS) proteoglycans [1]. In mammals, the glypican family consists of six members from glypican-1 (GPC1) to glypican-6 (GPC6), all of which share highly conserved amino acid sequences between species [1]. Despite this conservation, individual glypicans exhibit distinct structural and functional characteristics [2]. The glypican family has attracted significant scientific attention due to its involvement in various signaling pathways [3] and tumor development [4].
The first identified member of the glypican family, GPC1, is present in both a membrane-bound form and an enzymatically cleaved soluble form by the metalloprotease ADAM17 [5] or the lipase Notum [6]. Human GPC1 consists of a core protein comprising 558 amino acids, which is anchored to the cell membrane via a glycosylphosphatidylinositol linkage at the serine-530 (S530) residue. Additionally, three HS chains are attached to the core protein at serine residues, S486, S488, and S490 [7]. These HS moieties recruit several signaling molecules, including hepatocyte growth factor (HGF), fibroblast growth factor-2 (FGF-2), vascular endothelial growth factor A (VEGF-A), and transforming growth factor-β (TGF-β), and facilitate their binding to corresponding receptors [3]. Consequently, GPC1 serves essential functions in regulating various signaling pathways involved in tumor cell proliferation, invasiveness, and tumorigenesis [8,9].
GPC1 expression is significantly upregulated in various tumors. Overexpression of GPC1 is associated with reduced overall survival, relapse-free survival, and/or disease-free survival, and chemoresistance in esophageal squamous cell carcinoma (SCC) [10]. Furthermore, the GPC1 overexpression has been reported in gliomas, lung SCC, pancreatic, prostate, and breast cancers. In these studies, a strong correlation has been observed between GPC1 overexpression and poor clinical outcomes. [11,12,13,14,15]. Therefore, GPC1 is an important predictive factor for tumor diagnosis.
GPC1 has been evaluated as a potential target for antibody-based therapies, including anti-GPC1 monoclonal antibodies (mAbs), antibody-drug conjugates (ADCs), radiotherapy, photoimmunotherapy, chimeric antigen receptor (CAR) T cell therapy, and bispecific T-cell engager (BiTE) in preclinical and clinical studies [7]. To develop these formats, anti-GPC1 mAbs with high reactivity and specificity in flow cytometry are essential.
Using the Cell-Based Immunization and Screening (CBIS) method, various mAbs against membrane proteins, such as chemokine receptors [16,17,18] and receptor tyrosine kinases [19,20] have been developed. The CBIS method includes immunizing antigen-overexpressed cells and high-throughput screening using flow cytometry. Therefore, mAbs obtained by the CBIS method are suitable for flow cytometry and tend to recognize conformational epitopes. Furthermore, some of these mAbs also apply to western blotting and immunohistochemistry (IHC). This study employed the CBIS method to develop highly versatile anti-GPC1 mAbs.
2. Materials and Methods
2.1. Cell Lines
Chinese hamster ovary (CHO)-K1, mouse myeloma P3X63Ag8.U1 (P3U1), and human glioblastoma LN229 were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). KYSE770 was obtained from the Japanese Collection of Research Bioresources (Osaka, Japan). These cells were maintained as described previously [21].
2.2. Plasmid Construction and Establishment of Stable Transfectants
The cDNAs of GPC3v2 (NM_001164618) and GPC6 (NM_005708) were synthesized by Eurofins Genomics KK (Tokyo, Japan). The cDNAs of GPC2 (NM_152742), GPC4 (NM_001448), and GPC5 (NM_004466) were obtained from RIKEN RBC. The cDNAs of GPC2, GPC3v2 (also known as GPC3), GPC4, and GPC5 were cloned into a pCAG-ble vector. A GPC6 cDNA was cloned into a pCAGzeo-ssnPA16 vector [22]. The expression vector of GPC1 (pCMV6_GPC1, NM_002081) was purchased from OriGene Technologies, Inc. (Rockville, MD, USA). The plasmids were transfected into CHO-K1 cells and stable transfectants were established by sorting with an anti-PA16 tag mAb (clone NZ-1 for GPC6) [22], an anti-GPC5 mAb (clone 297716; R&D Systems, Inc., Minneapolis, MN, USA), an anti-GPC4 mAb (clone A21050B; BioLegend, San Diego, CA, USA), an anti-GPC3 mAb (clone SP86; Abcam, Cambridge, UK), an anti-GPC2 mAb (clone CT3; Cell Signaling Technology, Inc., Danvers, MA, USA), and an anti-GPC1 mAb (clone 1019718; R&D systems) using a cell sorter (SH800, Sony Corp., Tokyo, Japan). After sorting, cultivation was performed in a medium containing 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA, USA) or 0.5 mg/mL of G418 (Nacalai Tesque, Inc.). These GPC-overexpressed CHO-K1 (e.g., CHO/GPC1) clones were finally established.
2.3. Hybridoma Production
The female BALB/cAJcl mice were purchased from CLEA Japan (Tokyo, Japan). The mice were intraperitoneally immunized with LN229/GPC1 cells (1 × 108 cells/injection) and Alhydrogel adjuvant 2% (InvivoGen). After three additional immunizations per week (1 × 108 cells/injection), a booster injection (1 × 108 cells/injection) was administered two days before harvesting the spleen cells from immunized mice. The hybridomas were generated as described previously [16].
2.4. Flow Cytometry
Cells were harvested using 1 mM ethylenediaminetetraacetic acid. The cells were washed with 0.1% bovine serum albumin (BSA) in phosphate-buffered saline (PBS, blocking buffer) and treated with mAbs for 30 minutes at 4°C. The cells were then stained with anti-mouse IgG or anti-rat IgG conjugated with Alexa Fluor 488 (Cell Signaling Technology, Inc., Danvers, MA, USA) for 30 minutes at 4°C. The data were collected using an SA3800 Cell Analyzer and analyzed using FlowJo software (BD Biosciences, Franklin Lakes, NJ, USA).
2.5. Determination of Dissociation Constant Values Using Flow Cytometry
CHO/GPC1 and KYSE770 were treated with serially diluted G1Mab-28. Subsequently, the cells were treated with anti-mouse IgG conjugated with Alexa Fluor 488 (200-fold dilution) for 30 minutes at 4°C. The dissociation constant (KD) values were determined as described previously [19].
2.6. Western Blotting
2.7. Immunohistochemistry (IHC) Using Cell Blocks
Cells were fixed with 4% paraformaldehyde, and the cell blocks were prepared using iPGell (Genostaff Co., Ltd., Tokyo, Japan) (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). The formalin-fixed paraffin-embedded (FFPE) cell sections were stained with G1Mab-28 (1 μg/mL) or 1019718 (1 μg/mL) using the ultraView Universal DAB Detection Kit and BenchMark ULTRA PLUS (Roche Diagnostics, Indianapolis, IN, USA).
3. Results
3.1. Development of Anti-GPC1 mAbs Using the CBIS Method
To develop anti-GPC1 mAbs, LN229/GPC1 was used as an antigen. Hybridomas were generated by fusing LN229/GPC1-immunized splenocytes with P3U1 cells. After forming colonies, the supernatants were screened for CHO/GPC1-positive and CHO-K1-negative. Subsequently, anti-GPC1 mAb-producing hybridomas were cloned by limiting dilution. A clone G1Mab-28 (IgG1, κ) was finally established (Figure 1).
3.2. Flow Cytometry Using Anti-GPC1 mAbs
We conducted flow cytometry using the G1Mab-28 and a commercially available anti-GPC1 mAb (clone 1019718) against CHO/GPC1 and CHO-K1 cells. The G1Mab-28 and 1019718 recognized CHO/GPC1 dose-dependently from 10 to 0.01 μg/mL (Figure 2A), but did not recognize CHO-K1 even at 10 μg/mL (Figure 2B). We next examined the reactivity of G1Mab-28 against an endogenous GPC1-expressing cell line, KYSE770. G1Mab-28 showed dose-dependent recognition of KYSE770 (Figure 2C). These results indicate that G1Mab-28 recognizes GPC1 in flow cytometry.
3.3. Specificity of G1Mab-28 Against GPC Family Members
We established other GPC2-6-overexpressed CHO-K1, and the specificity of G1Mab-28 to the GPC family members was determined. As shown in Figure 3, G1Mab-28 recognized CHO/GPC1 and did not react with other GPC-overexpressed CHO-K1. These results indicate that G1Mab-28 is a specific mAb against GPC1 among GPC family members.
3.4. Determination of KD Values of G1Mab-28 by Flow Cytometry
The binding affinity of G1Mab-28 was determined using a flow cytometry-based assay. The KD values of G1Mab-28 for CHO/GPC1 and KYSE770 were 3.3 × 10⁻8 M and 4.6 × 10⁻⁹ M, respectively (Figure 4). The KD values of 1019718 for CHO/GPC1 and KYSE770 were 6.3 × 10⁻8 M and 1.3 × 10⁻8 M, respectively (Figure 4). These results indicated that G1Mab-28 has high affinity to GPC1-positive cells.
3.5. Western Blotting Using G1Mab-28
We next investigated whether G1Mab-28 is suitable for western blotting. Whole-cell lysates of CHO-K1 and CHO/GPC1 were used. G1Mab-28 detected weak bands around 75 kDa in CHO/GPC1, but not in CHO-K1 (Figure 5A, left). 1019718 potently detected in same exposure conditions (Figure 5B). In a long exposure, G1Mab-28 could detect clear bands around 75 kDa in CHO/GPC1, but not in CHO-K1 (Figure 5A, right). IDH1 detected by RcMab-1 was used as an internal control (Figure 5C). These results indicate that G1Mab-28 can detect overexpressed GPC1 in western blotting.
3.6. IHC Using G1Mab-28 in FFPE Cell Sections
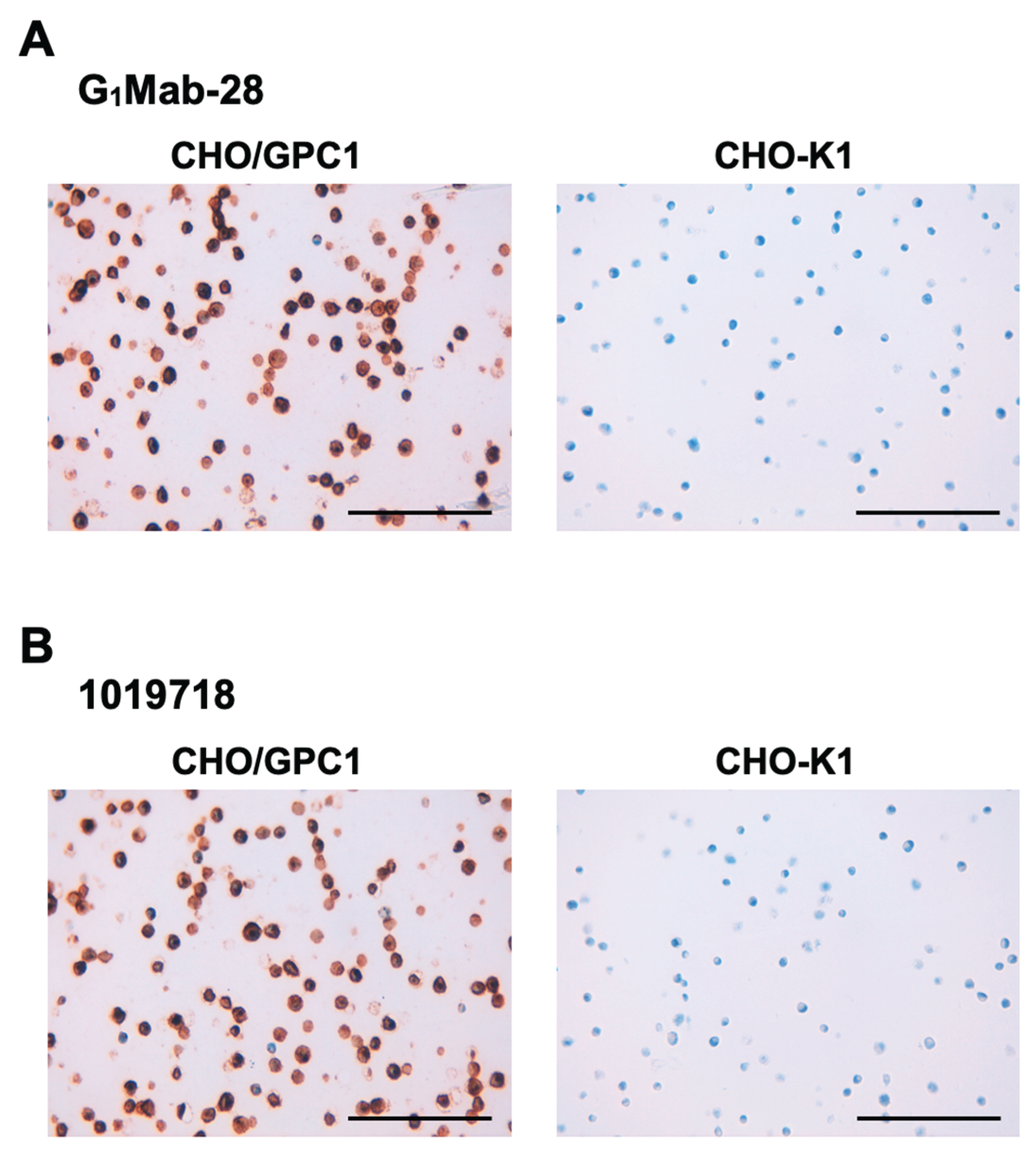
We examined whether G1Mab-28 is suitable for the IHC analysis of FFPE sections of CHO-K1 and CHO/GPC1. Both intense membranous and cytoplasmic staining by G1Mab-28 were detected in CHO/GPC1 but not in CHO-K1 (Figure 6A). 1019718 also reacted with CHO/GPC1, but not CHO-K1 (Figure 6B). These results indicate that G1Mab-28 can detect GPC1 in FFPE cell samples.
4. Discussion
This study demonstrated the property of a novel anti-GPC1 mAb clone G1Mab-28 established by the CBIS method. In flow cytometry, G1Mab-28 exhibited the superior reactivity (Figure 2) and affinity (Figure 4) compared to 1019718, a commercially available anti-GPC1 mAb. G1Mab-28 specifically recognized GPC1 among GPC family (Figure 3). Furthermore, G1Mab-28 can detect GPC1 in western blotting (Figure 5) and GPC1-positive cells in immunohistochemistry (Figure 6). Therefore, G1Mab-28 can detect GPC-1 on tumor cells and probably exosomes. It should be determined whether G1Mab-28 can detect an enzymatically cleaved soluble form. The combined detection of exosomal GPC1, exosomal CD82, and serum CA19-9 was reported as a promising method for pancreatic cancer detection [24]. Therefore, G1Mab-28 could contribute to the detection of exosomal GPC1 in early diagnosis of pancreatic cancer.
GPC1 acts as a co-receptor for HGF, FGF-2, some Wnt ligands, and TGF-β to enhance the signaling pathways, which plays essential roles in tumor cell proliferation, stemness, invasiveness, and epithelial-to-mesenchymal transition [3,9,25]. A GPC1-targeted immunotoxin inhibits pancreatic tumor growth via degradation of internalized GPC1 and downregulation of Wnt signaling [26]. Therefore, the antibody-mediated internalization of GPC1 could reduce the recruitment of those signaling molecules and suppress the tumorigenic signaling. We have developed other clones of anti-GPC1 mAbs (see below). The analyses of their internalizing activity and growth inhibitory effect are essential to evaluate the anti-proliferative effect and future development of ADC.
Several mAbs have been evaluated in preclinical and clinical studies [7]. A chimeric antibody Miltuximab is developed from a BLCA-38 subclone, MIL-38 [7]. The anti-GPC1 mAb, BLCA-38 was obtained by the immunization with human bladder cancer UCRU-BL-17CL cells [27]. A first-in-human trial of Miltuximab was conducted and has validated the safety and tolerability in patients with advanced pancreatic, prostate, and bladder cancer (ACTRN12616000787482) [28]. Furthermore, Miltuximab has been developed to an immunotheranostics ([67Ga]Ga-DOTA-Miltuximab) and evaluated the safety and tolerability in patients with advanced solid cancers [29]. 89Zr-DFO-Miltuximab has been developed as an effective immuno-positron emission tomography imaging agent for detecting GPC1-positive glioblastoma in a mouse model [30]. 225Ac- and 177Lu-labeled Miltuximab have been developed as α- and β-therapy, respectively [7,31]. A photoimmunotherapy, Miltuximab®-IR700 showed significant reduction in viability of GPC1-positive cancer cell lines [32]. MIL-38-CD3 BiTE possesses a tandem scFv format by combining the scFv of Miltuximab® linked to the anti-CD3 scFv. The MIL-38-CD3 BiTE redirects T cell cytolytic activity to kill GPC1-expressing prostate cancer cells in a preclinical model [33]. Other anti-GPC1 mAb-based format such as ADC has been evaluated and exhibited the antitumor efficacy in mouse models of glioblastoma, cholangiocarcinoma, uterine cervical cancer, and pancreatic cancer [34,35,36,37]. Anti-GPC1 CAR T cell therapy has been evaluated and exhibited the antitumor efficacy in mouse models of pancreatic cancer [38].
A significant proportion of patients exhibit elevated or detectable GPC1 expression, while the GPC1 expression is thought to be minimal or absent in normal tissues. The expression of GPC1 in normal tissue was evaluated by IHC [4,39]. However, our flow cytometry analysis revealed that both G1Mab-28 and 1019718 recognize non-tumorigenic keratinocyte HaCaT, embryonic fibroblast KMST-6, and embryonic kidney 293FT (Supplementary Figure S1). To attain a favorable therapeutic index while minimizing on-target toxicity, antigens that are highly expressed in tumors but exhibit minimal or no expression in normal tissues are considered ideal targets. Nevertheless, the scarcity of such ideal tumor-associated antigens poses a substantial challenge in the development of therapeutic mAbs for tumor therapy [40].
To address this challenge, we have developed cancer-specific mAbs (CasMabs) targeting various antigens, including human epidermal growth factor receptor 2 (HER2) [41,42], podoplanin [43,44,45], and podocalyxin [46], and have characterized both their cancer-specific epitopes and recognition mechanisms. In the development of anti-HER2 CasMabs, we generated over 300 anti-HER2 mAb clones by immunizing mice with HER2 expressed on cancer cells. These mAbs were screened for differential reactivity against HER2-expressing tumor versus normal cells using flow cytometry [41]. Among the clones, H2Mab-250 (also referred to as H2CasMab-2) selectively recognized HER2 in breast cancer cells, but not in normal epithelial cells derived from mammary gland, kidney proximal tubule, colon, or lung bronchus [41]. Furthermore, mouse IgG2a type or humanized H2Mab-250 exhibited ADCC, complement-dependent cytotoxicity (CDC), and in vivo antitumor activity against human breast cancer xenografts [47,48,49]. We have established more clones producing anti-GPC1 mAbs and listed our WEB page “Antibody bank (http://www.med-tohoku-antibody.com/topics/001_paper_antibody_PDIS.htm#GPC1)”. We will obtain more than one hundred clones and compare the reactivity to normal and cancer cell lines. The selection of anti-GPC1 CasMabs and identifying the epitopes are essential strategies for developing a novel therapeutic anti-GPC1 CasMab in the future.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Haruto Yamamoto: Investigation. Hiroyuki Suzuki: Writing—original draft. Tomohiro Tanaka: Investigation, Funding acquisition. Mika K. Kaneko: Conceptualization. Yukinari Kato: Conceptualization, Funding acquisition, Project administration, Writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding Information
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP25am0521010 (to Y.K.), JP25ama121008 (to Y.K.), JP25ama221339 (to Y.K.), and JP25bm1123027 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 24K18268 (to T.T.) and 25K10553 (to Y.K.).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- Filmus, J.; Capurro, M.; Rast, J. Glypicans. Genome Biol 2008;9(5): 224. [CrossRef]
- Filmus, J. The function of glypicans in the mammalian embryo. Am J Physiol Cell Physiol 2022;322(4): C694-c698.
- Pan, J.; Ho, M. Role of glypican-1 in regulating multiple cellular signaling pathways. Am J Physiol Cell Physiol 2021;321(5): C846-c858. [CrossRef]
- Li, N.; Spetz, M.R.; Ho, M. The Role of Glypicans in Cancer Progression and Therapy. J Histochem Cytochem 2020;68(12): 841-862. [CrossRef]
- Kawahara, R.; Granato, D.C.; Yokoo, S.; et al. Mass spectrometry-based proteomics revealed Glypican-1 as a novel ADAM17 substrate. J Proteomics 2017;151: 53-65. [CrossRef]
- Traister, A.; Shi, W.; Filmus, J. Mammalian Notum induces the release of glypicans and other GPI-anchored proteins from the cell surface. Biochem J 2008;410(3): 503-511. [CrossRef]
- Ghosh, S.; Huda, P.; Fletcher, N.; et al. Clinical development of an anti-GPC-1 antibody for the treatment of cancer. Expert Opin Biol Ther 2022;22(5): 603-613. [CrossRef]
- Lund, M.E.; Campbell, D.H.; Walsh, B.J. The Role of Glypican-1 in the Tumour Microenvironment. Adv Exp Med Biol 2020;1245: 163-176.
- Wang, S.; Qiu, Y.; Bai, B. The Expression, Regulation, and Biomarker Potential of Glypican-1 in Cancer. Front Oncol 2019;9: 614.
- Hara, H.; Takahashi, T.; Serada, S.; et al. Overexpression of glypican-1 implicates poor prognosis and their chemoresistance in oesophageal squamous cell carcinoma. Br J Cancer 2016;115(1): 66-75. [CrossRef]
- Kai, Y.; Amatya, V.J.; Kushitani, K.; et al. Glypican-1 is a novel immunohistochemical marker to differentiate poorly differentiated squamous cell carcinoma from solid predominant adenocarcinoma of the lung. Transl Lung Cancer Res 2021;10(2): 766-775. [CrossRef]
- Saito, T.; Sugiyama, K.; Hama, S.; et al. High Expression of Glypican-1 Predicts Dissemination and Poor Prognosis in Glioblastomas. World Neurosurg 2017;105: 282-288. [CrossRef]
- Duan, L.; Hu, X.Q.; Feng, D.Y.; Lei, S.Y.; Hu, G.H. GPC-1 may serve as a predictor of perineural invasion and a prognosticator of survival in pancreatic cancer. Asian J Surg 2013;36(1): 7-12. [CrossRef]
- Russell, P.J.; Ow, K.T.; Tam, P.N.; et al. Immunohistochemical characterisation of the monoclonal antibody BLCA-38 for the detection of prostate cancer. Cancer Immunol Immunother 2004;53(11): 995-1004. [CrossRef]
- Matsuda, K.; Maruyama, H.; Guo, F.; et al. Glypican-1 is overexpressed in human breast cancer and modulates the mitogenic effects of multiple heparin-binding growth factors in breast cancer cells. Cancer Res 2001;61(14): 5562-5569.
- Suzuki, H.; Tanaka, T.; Li, G.; et al. Development of a Sensitive Anti-Mouse CCR5 Monoclonal Antibody for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2024;43(4): 96-100. [CrossRef]
- Tanaka, T.; Nanamiya, R.; Takei, J.; et al. Development of Anti-Mouse CC Chemokine Receptor 8 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021;40(2): 65-70. [CrossRef]
- Nanamiya, R.; Takei, J.; Asano, T.; et al. Development of Anti-Human CC Chemokine Receptor 9 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021;40(3): 101-106. [CrossRef]
- Nanamiya, R.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of an Anti-EphB4 Monoclonal Antibody for Multiple Applications Against Breast Cancers. Monoclon Antib Immunodiagn Immunother 2023;42(5): 166-177. [CrossRef]
- Itai, S.; Yamada, S.; Kaneko, M.K.; et al. Establishment of EMab-134, a Sensitive and Specific Anti-Epidermal Growth Factor Receptor Monoclonal Antibody for Detecting Squamous Cell Carcinoma Cells of the Oral Cavity. Monoclon Antib Immunodiagn Immunother 2017;36(6): 272-281. [CrossRef]
- Goto, N.; Suzuki, H.; Tanaka, T.; et al. Development of a Novel Anti-CD44 Monoclonal Antibody for Multiple Applications against Esophageal Squamous Cell Carcinomas. Int J Mol Sci 2022;23(10). [CrossRef]
- Fujii, Y.; Kaneko, M.; Neyazaki, M.; et al. PA tag: a versatile protein tagging system using a super high affinity antibody against a dodecapeptide derived from human podoplanin. Protein Expr Purif 2014;95: 240-247. [CrossRef]
- Ikota, H.; Nobusawa, S.; Arai, H.; et al. Evaluation of IDH1 status in diffusely infiltrating gliomas by immunohistochemistry using anti-mutant and wild type IDH1 antibodies. Brain Tumor Pathol 2015;32(4): 237-244. [CrossRef]
- Xiao, D.; Dong, Z.; Zhen, L.; et al. Combined Exosomal GPC1, CD82, and Serum CA19-9 as Multiplex Targets: A Specific, Sensitive, and Reproducible Detection Panel for the Diagnosis of Pancreatic Cancer. Mol Cancer Res 2020;18(2): 300-310. [CrossRef]
- Tripathi, A.D.; Katiyar, S.; Mishra, A. Glypican1: A potential cancer biomarker for nanotargeted therapy. Drug Discov Today 2023;28(8): 103660. [CrossRef]
- Pan, J.; Li, N.; Renn, A.; et al. GPC1-Targeted Immunotoxins Inhibit Pancreatic Tumor Growth in Mice via Depletion of Short-lived GPC1 and Downregulation of Wnt Signaling. Mol Cancer Ther 2022;21(6): 960-973. [CrossRef]
- Walker, K.Z.; Russell, P.J.; Kingsley, E.A.; Philips, J.; Raghavan, D. Detection of malignant cells in voided urine from patients with bladder cancer, a novel monoclonal assay. J Urol 1989;142(6): 1578-1583. [CrossRef]
- Gurney, H.; Sabanathan, D.; Gillatt, D.; et al. MILGa-01: A first-in-human study assessing the safety and tolerability of chMIL-38 in metastatic prostate, bladder, and pancreatic cancers. Journal of Clinical Oncology 2017;35(6_suppl): e565-e565. [CrossRef]
- Sabanathan, D.; Campbell, D.H.; Velonas, V.M.; et al. Safety and tolerability of Miltuximab(®) - a first in human study in patients with advanced solid cancers. Asia Ocean J Nucl Med Biol 2021;9(2): 86-100. [CrossRef]
- Ghosh, S.; Fletcher, N.L.; Huda, P.; et al. Pharmacokinetics and Biodistribution of (89)Zr-Miltuximab and Its Antibody Fragments as Glypican-1 Targeting Immuno-PET Agents in Glioblastoma. Mol Pharm 2023;20(3): 1549-1563. [CrossRef]
- Yeh, M.C.; Tse, B.W.C.; Fletcher, N.L.; et al. Targeted beta therapy of prostate cancer with (177)Lu-labelled Miltuximab® antibody against glypican-1 (GPC-1). EJNMMI Res 2020;10(1): 46. [CrossRef]
- Polikarpov, D.M.; Campbell, D.H.; Lund, M.E.; et al. The feasibility of Miltuximab®-IRDye700DX-mediated photoimmunotherapy of solid tumors. Photodiagnosis Photodyn Ther 2020;32: 102064.
- Lund, M.E.; Howard, C.B.; Thurecht, K.J.; et al. A bispecific T cell engager targeting Glypican-1 redirects T cell cytolytic activity to kill prostate cancer cells. BMC Cancer 2020;20(1): 1214. [CrossRef]
- Uchida, S.; Serada, S.; Suzuki, Y.; et al. Glypican-1-targeted antibody-drug conjugate inhibits the growth of glypican-1-positive glioblastoma. Neoplasia 2024;50: 100982. [CrossRef]
- Yokota, K.; Serada, S.; Tsujii, S.; et al. Anti-Glypican-1 Antibody-drug Conjugate as Potential Therapy Against Tumor Cells and Tumor Vasculature for Glypican-1-Positive Cholangiocarcinoma. Mol Cancer Ther 2021;20(9): 1713-1722. [CrossRef]
- Nishigaki, T.; Takahashi, T.; Serada, S.; et al. Anti-glypican-1 antibody-drug conjugate is a potential therapy against pancreatic cancer. Br J Cancer 2020;122(9): 1333-1341.
- Matsuzaki, S.; Serada, S.; Hiramatsu, K.; et al. Anti-glypican-1 antibody-drug conjugate exhibits potent preclinical antitumor activity against glypican-1 positive uterine cervical cancer. Int J Cancer 2018;142(5): 1056-1066.
- Li, N.; Quan, A.; Li, D.; et al. The IgG4 hinge with CD28 transmembrane domain improves V(H)H-based CAR T cells targeting a membrane-distal epitope of GPC1 in pancreatic cancer. Nat Commun 2023;14(1): 1986.
- 39. Kato, D.; Yaguchi, T.; Iwata, T.; et al. GPC1 specific CAR-T cells eradicate established solid tumor without adverse effects and synergize with anti-PD-1 Ab. Elife 2020;9. [CrossRef]
- Paul, S.; Konig, M.F.; Pardoll, D.M.; et al. Cancer therapy with antibodies. Nat Rev Cancer 2024;24(6): 399-426.
- Kaneko, M.K.; Suzuki, H.; Kato, Y. Establishment of a Novel Cancer-Specific Anti-HER2 Monoclonal Antibody H(2)Mab-250/H(2)CasMab-2 for Breast Cancers. Monoclon Antib Immunodiagn Immunother 2024;43(2): 35-43.
- Arimori, T.; Mihara, E.; Suzuki, H.; et al. Locally misfolded HER2 expressed on cancer cells is a promising target for development of cancer-specific antibodies. Structure 2024;32(5): 536-549.e535. [CrossRef]
- Tanaka, T.; Suzuki, H.; Ohishi, T.; Kaneko, M.K.; Kato, Y. A Cancer-Specific Anti-Podoplanin Monoclonal Antibody, PMab-117-mG(2a) Exerts Antitumor Activities in Human Tumor Xenograft Models. Cells 2024;13(22).
- Suzuki, H.; Ohishi, T.; Kaneko, M.K.; Kato, Y. A Humanized and Defucosylated Antibody against Podoplanin (humLpMab-23-f) Exerts Antitumor Activities in Human Lung Cancer and Glioblastoma Xenograft Models. Cancers (Basel) 2023;15(20). [CrossRef]
- Kato, Y.; Kaneko, M.K. A cancer-specific monoclonal antibody recognizes the aberrantly glycosylated podoplanin. Sci Rep 2014;4: 5924. [CrossRef]
- Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. A Cancer-Specific Monoclonal Antibody against Podocalyxin Exerted Antitumor Activities in Pancreatic Cancer Xenografts. Int J Mol Sci 2023;25(1). [CrossRef]
- Kaneko, M.K.; Suzuki, H.; Ohishi, T.; et al. Antitumor Activities of a Humanized Cancer-Specific Anti-HER2 Monoclonal Antibody, humH(2)Mab-250 in Human Breast Cancer Xenografts. Int J Mol Sci 2025;26(3). [CrossRef]
- Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Anti-HER2 Cancer-Specific mAb, H(2)Mab-250-hG(1), Possesses Higher Complement-Dependent Cytotoxicity than Trastuzumab. Int J Mol Sci 2024;25(15). [CrossRef]
- Kaneko, M.K.; Suzuki, H.; Ohishi, T.; et al. A Cancer-Specific Monoclonal Antibody against HER2 Exerts Antitumor Activities in Human Breast Cancer Xenograft Models. Int J Mol Sci 2024;25(3). [CrossRef]
Figure 1.
Schematic representation of anti-GPC1 mAbs production. (A) LN229/GPC1 was injected intraperitoneally into BALB/cAJcl mice. (B) After five times immunizations per week, splenocytes were fused with P3U1. (C) A flow cytometry-based high-throughput screening was performed to select the CHO/GPC1-positive and CHO-K1-negative supernatants of hybridomas. (D) Anti-GPC1 specific mAb-producing hybridoma clones were established by limiting dilution.
Figure 1.
Schematic representation of anti-GPC1 mAbs production. (A) LN229/GPC1 was injected intraperitoneally into BALB/cAJcl mice. (B) After five times immunizations per week, splenocytes were fused with P3U1. (C) A flow cytometry-based high-throughput screening was performed to select the CHO/GPC1-positive and CHO-K1-negative supernatants of hybridomas. (D) Anti-GPC1 specific mAb-producing hybridoma clones were established by limiting dilution.
Figure 2.
Flow cytometry analysis of G1Mab-28 and 1019718 against CHO/GPC1, CHO-K1, and KYSE770. (A) CHO/GPC1 (A), CHO-K1 (B), and KYSE770 (C) were treated with G1Mab-28 or 1019718 at the indicated concentrations (red) or blocking buffer (black). The mAbs-treated cells were incubated with anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were collected using the SA3800 Cell Analyzer.
Figure 2.
Flow cytometry analysis of G1Mab-28 and 1019718 against CHO/GPC1, CHO-K1, and KYSE770. (A) CHO/GPC1 (A), CHO-K1 (B), and KYSE770 (C) were treated with G1Mab-28 or 1019718 at the indicated concentrations (red) or blocking buffer (black). The mAbs-treated cells were incubated with anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were collected using the SA3800 Cell Analyzer.
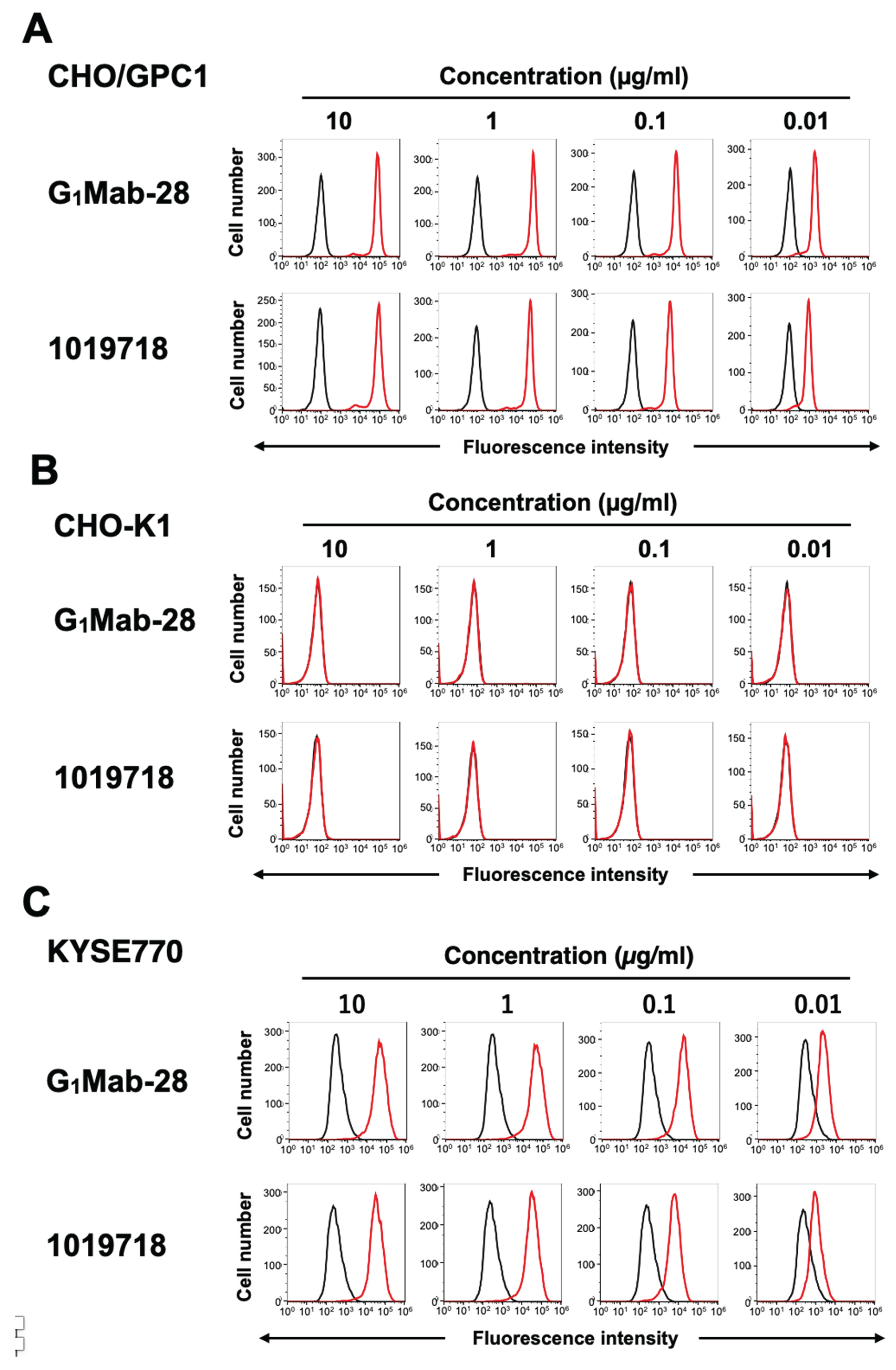
Figure 3.
Flow cytometry analysis of G1Mab-28 in GPC family members-expressed CHO-K1 cells. (A) The GPC family members (GPC1, GPC2, GPC3, GPC4, GPC5, and PA16-GPC6)-expressed CHO-K1 cells were treated with 1 µg/mL of G1Mab-28 (red) or control blocking buffer (black), followed by treatment with anti-mouse IgG conjugated with Alexa Fluor 488. (B) The expression of each GPC was confirmed by 1 µg/mL of an anti-GPC1 mAb (clone 1019718), 1 µg/mL of an anti-GPC2 mAb (clone CT3), 1 µg/mL of an anti-GPC3 mAb (clone SP86), 1 µg/mL of an anti-GPC4 mAb (clone A21050B), 1 µg/mL of an anti-GPC5 mAb (clone 297716), and 1 µg/mL of an anti-PA16 mAb, NZ-1.
Figure 3.
Flow cytometry analysis of G1Mab-28 in GPC family members-expressed CHO-K1 cells. (A) The GPC family members (GPC1, GPC2, GPC3, GPC4, GPC5, and PA16-GPC6)-expressed CHO-K1 cells were treated with 1 µg/mL of G1Mab-28 (red) or control blocking buffer (black), followed by treatment with anti-mouse IgG conjugated with Alexa Fluor 488. (B) The expression of each GPC was confirmed by 1 µg/mL of an anti-GPC1 mAb (clone 1019718), 1 µg/mL of an anti-GPC2 mAb (clone CT3), 1 µg/mL of an anti-GPC3 mAb (clone SP86), 1 µg/mL of an anti-GPC4 mAb (clone A21050B), 1 µg/mL of an anti-GPC5 mAb (clone 297716), and 1 µg/mL of an anti-PA16 mAb, NZ-1.
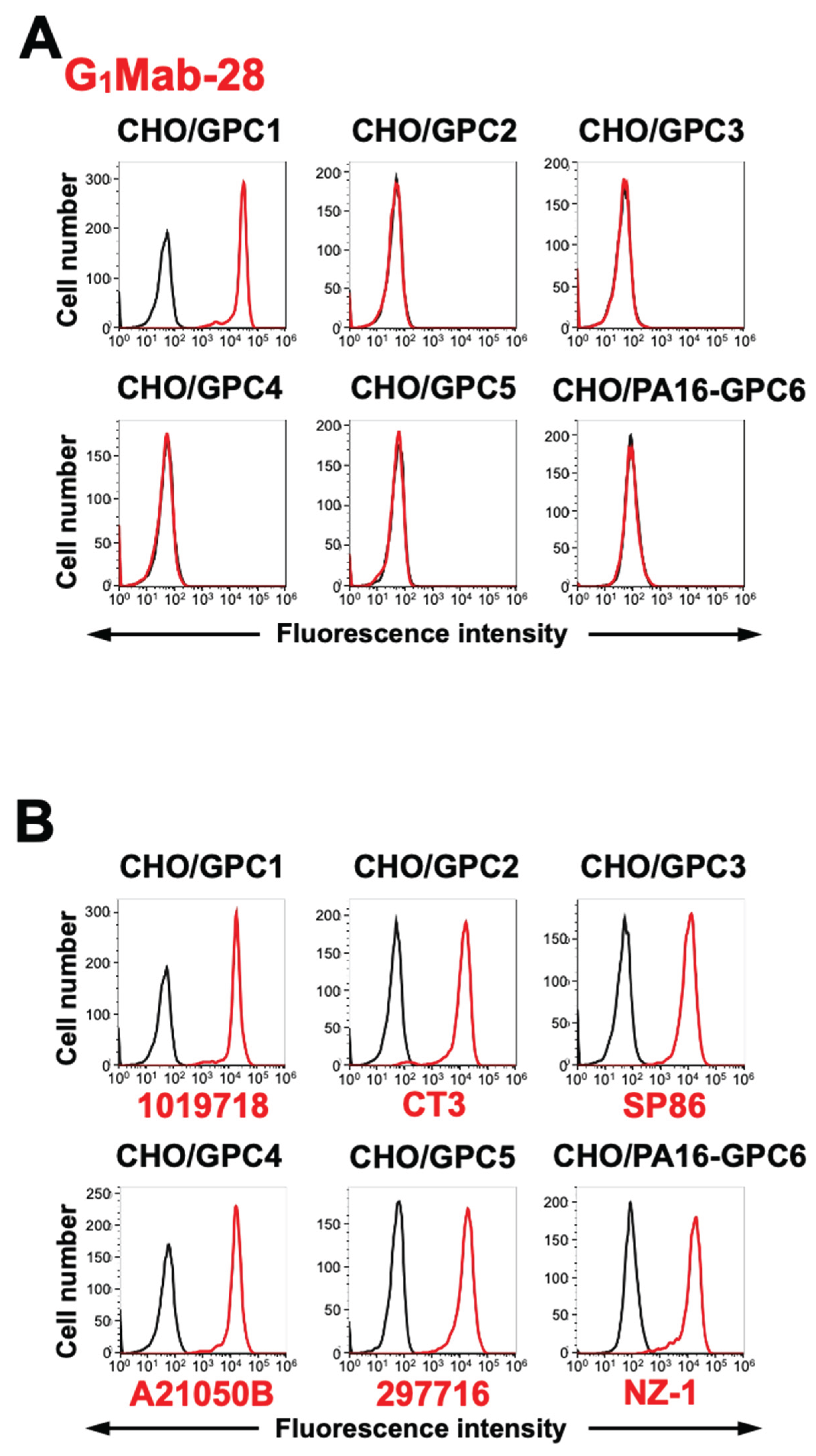
Figure 4.
Measurement of binding affinity of G1Mab-28. CHO/GPC1 (A) and KYSE770 (B) were treated with serially diluted G1Mab-28 or 1019718, followed by anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were analyzed using the BD FACSlylic. The KD values were determined using GraphPad PRISM 6.
Figure 4.
Measurement of binding affinity of G1Mab-28. CHO/GPC1 (A) and KYSE770 (B) were treated with serially diluted G1Mab-28 or 1019718, followed by anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were analyzed using the BD FACSlylic. The KD values were determined using GraphPad PRISM 6.
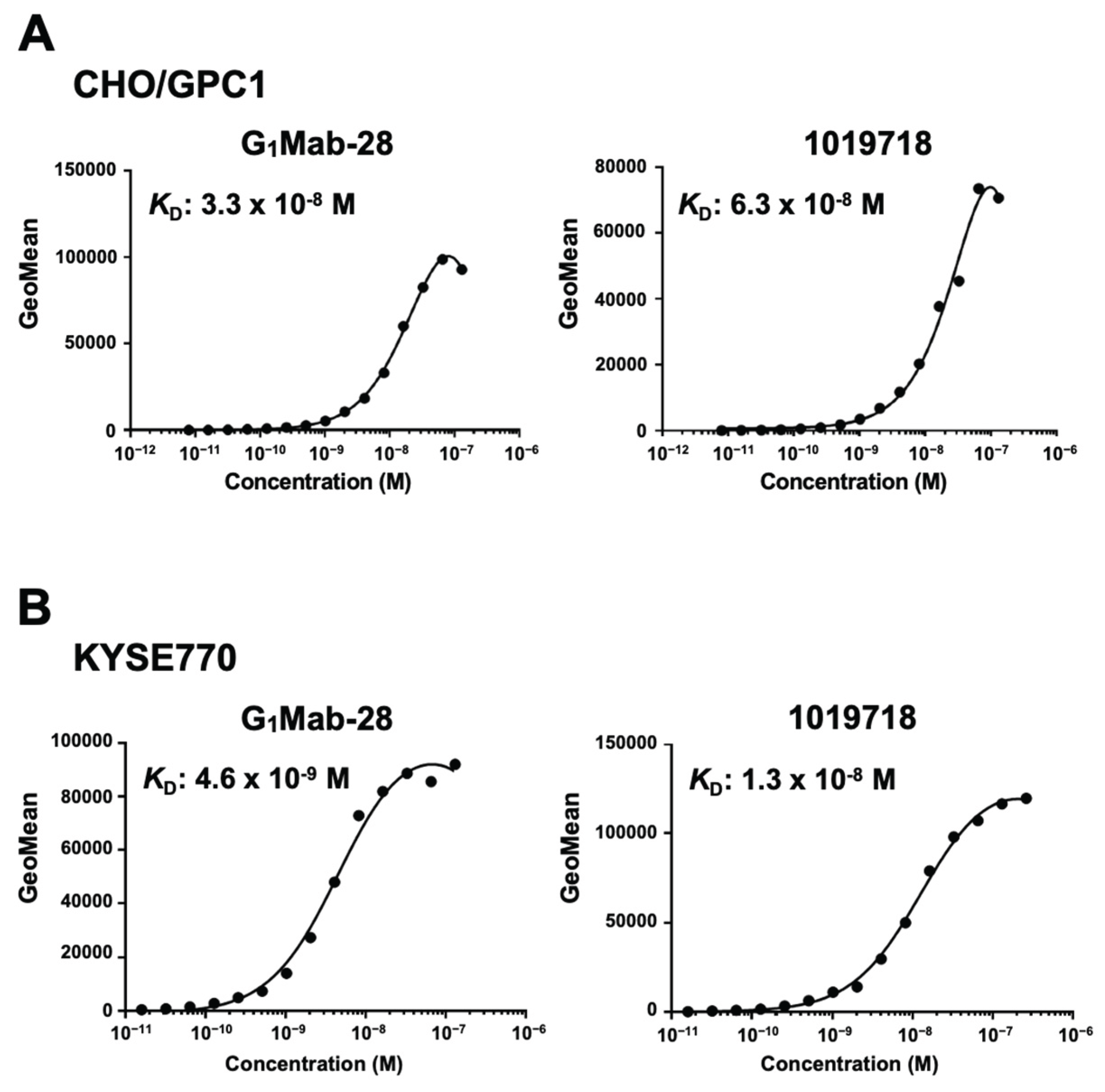
Figure 5.
Western blotting using G1Mab-28. The cell lysate of CHO-K1 and CHO/GPC1 was electrophoresed and transferred onto polyvinylidene difluoride membranes. The membranes were incubated with 1 μg/mL of G1Mab-28 (A), 1 μg/mL of 1019718 (B), or 1 μg/mL of RcMab-1 (an anti-isocitrate dehydrogenase 1, C), followed by treatment with anti-mouse or anti-rat IgG conjugated with horseradish peroxidase. Note that the exposure time of the left side of A and B was the same.
Figure 5.
Western blotting using G1Mab-28. The cell lysate of CHO-K1 and CHO/GPC1 was electrophoresed and transferred onto polyvinylidene difluoride membranes. The membranes were incubated with 1 μg/mL of G1Mab-28 (A), 1 μg/mL of 1019718 (B), or 1 μg/mL of RcMab-1 (an anti-isocitrate dehydrogenase 1, C), followed by treatment with anti-mouse or anti-rat IgG conjugated with horseradish peroxidase. Note that the exposure time of the left side of A and B was the same.
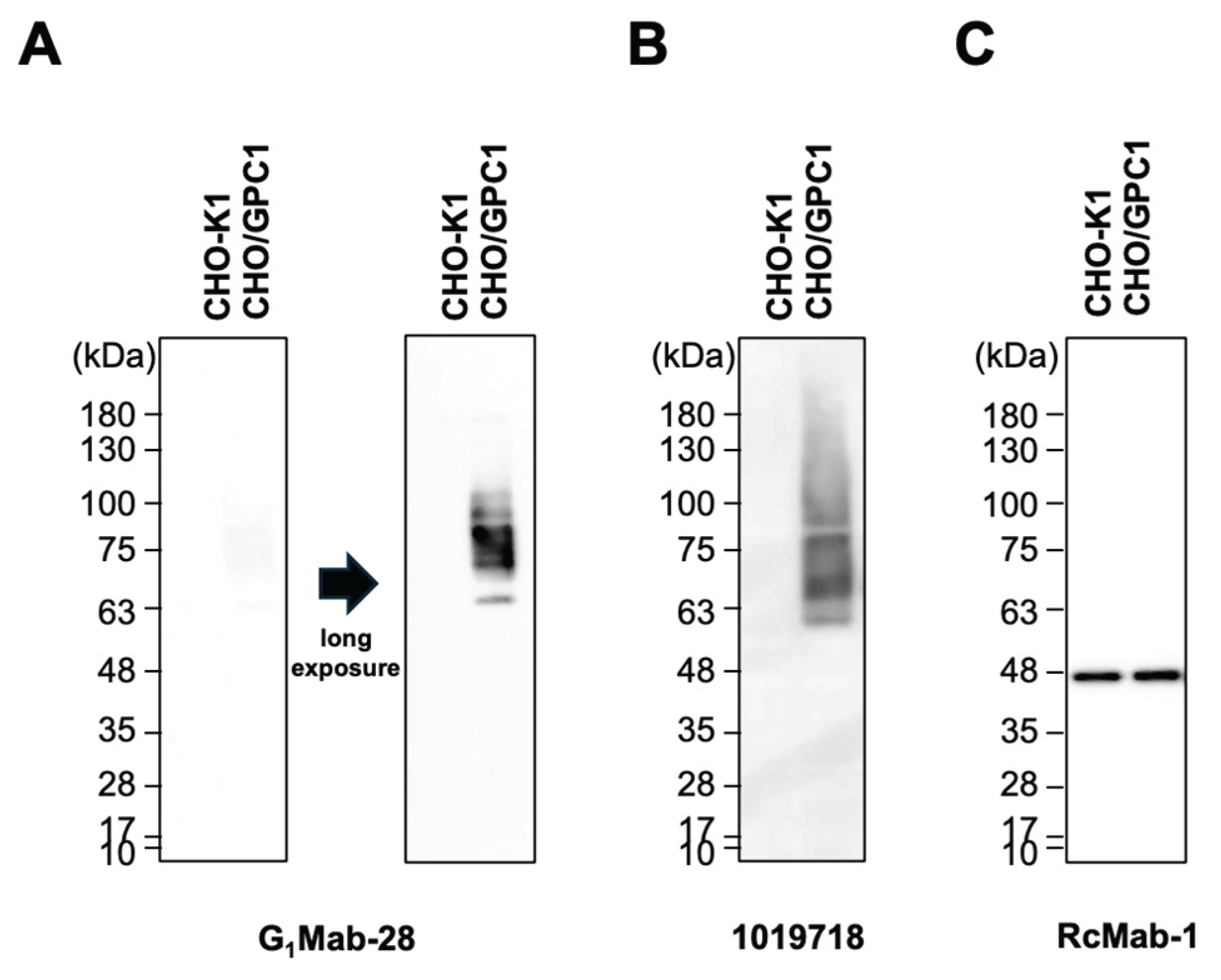
Figure 6.
Immunohistochemistry using G1Mab-28 in formalin-fixed paraffin-embedded cell blocks. CHO/GPC1 and CHO-K1 sections were treated with 1 μg/mL of G1Mab-28 (A) or 1 μg/mL of 1019718 (B). The staining was performed using ultraView Universal DAB Detection Kit and BenchMark ULTRA PLUS. Scale bar = 100 μm.
Figure 6.
Immunohistochemistry using G1Mab-28 in formalin-fixed paraffin-embedded cell blocks. CHO/GPC1 and CHO-K1 sections were treated with 1 μg/mL of G1Mab-28 (A) or 1 μg/mL of 1019718 (B). The staining was performed using ultraView Universal DAB Detection Kit and BenchMark ULTRA PLUS. Scale bar = 100 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



