Submitted:
02 June 2025
Posted:
04 June 2025
You are already at the latest version
Abstract
A novel evidence-based theory is offered that proposes microbial D-amino acids as evolutionarily-conserved interkingdom stress signals by lower life forms such as soil or gut microbiota serving as front-line ecological microsensors for plants and humans. These D-amino acids seem to act as metabolic switches that biochemically link xenobiotic stress to pathological disruptions in plant and human metabolisms and defense/immune systems. D-amino acids and peptides, by impairing chirally-sensitive hormones/phytohormones and mitochondrial and photosynthetic complexes, are able to disrupt the Krebs and photosynthetic cycles and switch cellular metabolism in higher life forms from orderly, oxidative and ATP-efficient, to microbial, reductive and ATP-deficient with low quantum yield. The hypothesis links the lower electron transfer efficiency in heterochiral peptides, where D-amino acids partially replace L-analogs, to photosynthetic/mitochondrial dysfunctions and diseases.
Keywords:
soil
; microbiology
; microbiome
; metabolism
; Amino acids
; peptides
; nutrition
Introduction
Although several integrative models such as the original framework proposed by Husson et al. [1] study soil-plant interactions, there are currently no unifying models to study the effect of interkingdom microbial crosstalk on plant and human metabolism and diseases.
Amino acids, as precursors of peptides (including neuropeptides and enzymes), immune cells and hormones/phytohormones, have key intersectional and interkingdom functions in metabolic and immune systems. As building blocks of life, Amino acids owe their ubiquitous functions in living organisms to their zwitterionic nature, making their polarity and stereo configuration a sensitive function of environmental conditions such as pH and oxidative (redox) state. This environmentally-sensitive polarity and configurational stereospecificity of amino acids makes them ideal as metabolic signaling molecules and precursors for biologically active peptides such as enzymes, ligands, receptors, substrates or hormones.
It has long been established that biologically significant amino acids in higher life forms such as plants and humans are homochiral molecules that rotate the plane of polarized light to the left (counterclockwise) and are categorized as L-amino acids (L is for Levorotatory, Latin for left-rotating). Conversely, micro organisms and primitive life forms have the ability to produce and metabolize D (Dextrorotatory)-amino acids. As shown in this paper, these microbial D-amino acids seem to universally act as xenobiotic ecological stress signals to higher life forms and alter the “directionality” of their metabolic cycles and immune/stress responses. The paper also proposes a novel integrative hypothesis that defines microbial D-amino acids as interkingdom modulators of homeostasis and redox reactions. This unifying conceptual framework can cogently explain the impact of environmental stressors on mitochondrial and chloroplastic dysfunctions responsible for diseases in plants and humans.
Discussion
The Role of D-Amino Acids in Stress Signaling and Defense Responses in Nature
Microbial colonies are known to convert L-amino acids to D-amino acids as a primary defense mechanism against invasive species and harsh environments. Enzymes that convert L-amino acids to D-amino acids mainly exist in lower life forms such as fungi, bacteria and lower plants [2] like mosses and green algae. D-amino acids are known to inhibit biofilm formation [3] and quorum sensing, a process in which hostile bacterial colonies coordinate their biofilm formation. Bacterial cell walls and plastids in moss and angiosperms also use simple D-amino acids such as D-alanine, D-glutamate, and D-aspartate as building blocks of peptidoglycan proteins that are resistant to enzymatic breakdown. Biochemical reactions involving D-amino acids in plants could be viewed as remnants of cyanobacterial metabolism that reflect ancestral peptidoglycan chemistry. Other than their role in building defensive peptidoglycans, D-amino acids seem to be of little anabolic value in plants. D-amino acids reportedly constitute on average about 0.7% of the plant’s total amino acid content [4] so it is reasonable to regard them mainly as xenobiotic and signaling compounds and not as an essential source of organic nitrogen.
The Plantae and Fungi kingdoms utilize D-amino acids as stress signals and defense mechanisms by harming the predator’s nervous system. For instance, ergot fungi and seeds of morning glories naturally produce D-Lysergic acid, the amino acid precursor humans use to make the psychedelic drug LSD (Lysergic acid diethylamide). LSD’s hallucinogenic effect on the nervous system could be related to the source plant’s D-amino acid neurotoxic stress signal and defense strategy against herbivores.
In some aquatic invertebrates, D-alanine levels increase under osmotic (high salinity) or anaerobic stress, suggesting D-amino acids can have a metabolic role that signals ecological stress. Some amphibians and invertebrates also use D-amino acids in their toxic defense systems, for example by corrupting the predator’s RNAs [5] and protein translation processes. Venoms from spiders contain D-amino acids which are used in manufacturing antibiotics and ion channel blockers. Frogs, in their skin secretions, produce peptides containing D-amino acids, often with opioid [6] or antimicrobial properties. Neurotoxic peptides in snail venom, called conotoxins, which disrupt neuromuscular transmissions, contain D-tryptophan or D-leucine [7] and are fatal to fish and in high doses to mammals. Conotoxins are used, in small doses, in pharmaceutical analgesics for numbing and relieving pain.
D-amino acid peptides are also shown [8] to disrupt the binding of natural peptides to heat shock proteins which function as molecular chaperones that prevent protein misfolding and aggregations during environmentally stressful conditions.
In mammals, D-forms of simple amino acids such as glutamic, serine, alanine and aspartic are utilized in small concentrations as agonists to excitatory glutamate receptors in the central nervous system. When overactivated, these receptors are associated with neurotoxicity in the brain, inflammation and damage of the spinal cord, dyskinesias, Parkinson’s disease, [9] demyelinating diseases [10] such as multiple sclerosis (MS), transverse myelitis and polio. The excitatory function of D-amino acids in the mammalian nervous system points out to their putative function as both responders to stress stimuli and activators of stress response.
D-Amino Acids in Food Chains: From Soil to Plants, Animals and Humans
While plants mostly lack enzymes to synthesize or metabolize D-amino acids, bacterial broad spectrum racemases and Dextrorotatory amino acid aminotransferases can convert L-amino acids to D-amino acids when subjected to biotic or abiotic stressors such as heat, high pH or salinity, anaerobic conditions, UV or microwave radiations, oxidative stress and free radicals (high Eh), and chemicals like aldehydes formed from microbial or enzymatic degradation of pesticides. As a result, high concentrations of D-amino acids in soil often indicate high stress conditions and decomposition of bacterial cell walls, synthetic pesticides, antibiotics and veterinary medicine leaching into agricultural soils via wastewater irrigation or animal manure. Because the decomposition of insects, plant residues and worms mainly produces L-amino acids, a high ratio of D- to L-amino acids in the soil correlates with a high density of stressed or dying microbes in necromass composition.
Generally, D-amino acids such as D-serine, D-alanine and D-tyrosine are mineralized at slower rates compared to the corresponding L-enantiomers and thus inhibit plant growth [11] and are considered phytotoxins when they overwhelm L-amino acids. Commensal plant microbiomes have a limited ability to metabolize toxic loads of D-amino acids into non-toxic products using D-amino acid oxidase (DAO).
In the Animalia kingdom, although simpler life forms like fruit flies seem to have formed symbiotic relationships with microbes that produce D-amino acids, mammals such as humans, have a limited capacity in metabolizing or even recognizing D-amino acids. The ability of viruses such as Bacillus anthracis that use D-amino acids in their capsules to evade mammalian immune systems is capitalized in new vaccine [12] developments. Denatured peptides synthesized from D-amino acids [13] which resist breakdown in human serum and gastric acid, and bypass the body’s innate immune system are also being researched as artificial T-cell immunogens in new vaccines. These same features of D-amino acid peptides could also activate autoimmune reactions, as explained later in this paper. Generally-speaking, in mammals, as in plants, the D-enantiomers of essential amino acids are utilised at very low levels [14] and when in excess, mark pathological environmental stress and act as inhibitors of cellular growth and metabolism.
Mutualistic gut bacteria (via fermentation) and some brain neurons can produce simple D-amino acids in free form and small quantities, in the gut as bactericides against pathogenic invasive microbes, and in the central nervous system as excitatory (stress) signaling molecules via NMDA (N-methyl-D-aspartate) receptors. Amino acid racemase and oxidases in the human brain, kidney, heart [15] and liver, and broad-spectrum racemases in gut bacteria can modulate D-amino acid signaling [16] and prevent accumulation of D-amino acids that can result in apoptosis and programmed cell death.
Foods fermented with Lactic acid bacteria produce L-amino acids and safe low levels of D-lactates [17], which can be metabolized by commensal lactic-acid gut bacteria. Other exogenous free D-amino acids from plant, animal or synthetic/processed sources, unlike D-amino acids produced by commensal bacteria, can challenge the endogenous mammalian D-amino acid oxidase system. Although we have limited data [18] on the effect of long-term consumption of processed D-amino acid proteins on the human organism, researchers have shown [19] that even a minor degree of racemisation to D-amino acids in food proteins can result in a major decrease in their proteolytic digestibility and, as discussed later, in allergic reactions and autoimmunity. Also, when overloaded with D-amino acids, the liver may become deficient in L-cysteine, which is required to make glutathione, a powerful antioxidant that metabolizes toxic loads of D-amino acids and alcohol metabolites like acetaldehyde.
High concentrations of D-lactate can also form as a by-product of hypoxic (oxygen-deprived), stressed and dysfunctional cellular metabolic pathways that lead to conditions such as diabetes and hyperglycemia. Studies have shown that even trauma, like gunshot or burn injuries, [20] can result in elevated endotoxic levels of serum D-lactate.
A Novel Hypothesis: D-amino Acids as Metabolic Switches in Plants and Humans
As their front-line response to foliar and root stressors, plants deploy phytochemicals such as salicylic acid, abscisic acid and jasmonic acid, which initiate crosstalk signaling to coordinate defense responses, osmoregulation and antioxidant (reactive oxygen species/ROS) homeostasis, impacting apoplastic and rhizosphere pH and redox potential (Eh). These hormonal responses to xenobiotic stresses originating outside the ecological biome are shown to be initiated and tightly regulated by microbial colonies [21], which, as shown earlier, primarily use D-amino acids in their own stress response. So it can be hypothesized that microbial D-amino acids play an interkingdom role in modulating a plant's redox signaling networks, cellular Eh–pH and response to environmental conditions that affect pest or pathogen-host interactions [1].
In humans, research data also underscore [22] the metabolic crosstalk between the gut microbiota and D-amino acids. The expression of D-amino acid oxidase enzymes in leukocytes (white blood cells) also indicates the ability of the human immune system to detect and neutralize D-amino acids as microbial stress signals.
The novel hypothesis shared in this paper proposes microbial D-amino acids as evolutionarily-conserved interkingdom stress signals and metabolic switches that allow plants and animals to exploit lower life forms such as soil or gut microbiota as their front-line ecological microsensors. This hypothesis is supported by the endosymbiotic theory [23], which postulates the bacterial evolutionary origin of cellular respiration and immune cells. Given the conserved prokaryotic biochemistry of metabolic organelles and immune cells, they are evolutionarily privileged to receive and decode D-amino acids as xenobiotic stress signals from their bacterial ancestors that act as ecological microbiosensors.
Furthermore, the fact that only with the help of commensal microbiomes and enzymes, plants and humans can metabolize D-amino acids and peptides, slowly and in small doses, may be indicative of the role of D-amino acids as interkingdom messengers of ecological stress.
Conversely, homochiral L-amino acids are essential in orchestrating immune/defense systems and the efficiency and directionality of metabolic cycles (flow of electrons) in plants and humans.
To maintain organism-level homeostasis in response to stressful conditions signaled by microbial D-amino acids, plants and humans deploy, in a similar and sometimes coordinated fashion [21], homochiral L-amino acid-containing hormones, neuropeptides and phytochemicals such as auxin (plants), serotonin/phytoserotonin and melatonin that regulate metabolic and physiological functions like growth, dormancy and circadian/seasonal rhythms. Higher life forms also rely heavily on L-amino acids like L-Cysteine, a precursor to many vitamins and antioxidants like glutathione, for adaptation to oxidative stress and bacterial redox signaling.
In addition, for optimized light-harvesting and metabolic adaptation to changing seasons and levels of oxidative stress, humans have evolved other neuropeptides like dopamine, hormones like adrenaline and cortisol, and pigments such as melanin, and plants have evolved phytochemicals such as salicylates, tocopherols (Vitamin E), flavonoids and anthocyanins. These protective plant phytonutrients seem to be also involved in interspecies hormesis (xenohormesis), which means activating and optimizing human stress responses. The majority of stress-sensitive metabolic hormones and xenohormetic phytonutrients, as well as light-harvesting and mitochondrial complexes and cytochromes, use homochiral L-amino acids such as L-phenylalanine, L-tyrosine, and L-tryptophan synthesized in the shikimate pathway (which pesticides like glyphosate can block.) [24]
Support by Quantum Physics and Thermodynamics
Higher forms of life are maintained by metabolic cycles that use light and oxygen to extract energy from the living organism’s ecosystem. Krebs and photosynthetic cycles rely on an “orderly and directional flow of electrons and protons” inside living cells in order to extract energy (ATP) from organic matter, water, oxygen and light. Thermodynamically, the ability of living organisms to impose order and organization over chaos is expressed as Gibbs Free Energy, a measure of vitality and work potential that increases with energy harvesting but decreases with chaos and temperature.
In Quantum physics, the efficiency of this energy-harvesting process in living cells, expressed as “quantum yield,” depends on the orderly flow of electrons inside organelles like mitochondria and chloroplasts, which in turn depends on L-amino acids and peptides because the chirality of amino acids determines their “directional” influence on harmonizing electron spins and the “quantum effectiveness” of electron transport and proton pumping processes. In higher life forms, health and vitality correlates with high metabolic “quantum yields” via the controlled “directional” flow of electrons. In other words, metabolic cycles in living cells of higher life forms act like semiconductor circuits that rely on L-amino acid switches for optimal performance.
In his paper "The Living State and Cancer" [25] Nobel laureate Albert Szent-Györgyi defined the “living state” as a dynamic (quasi-equilibrium) steady state achieved through the movement of electrons facilitated by the semiconductive role of amino-acids and proteins as the workhorses of life. In this process, proper foldings of chiral proteins, achieved through side chains such as L-Lysine, allow charge transfer reactions and mobility of electrons in the semi-conductive “desaturated” proteins, which build next to them aqueous “charge exclusion” zones with low dielectric constants.
This quantum view of metabolism is compatible with the chiral-specificity of olfactory receptors and the "vibration theory of olfaction,” [26] which point out to the importance of stereospecificity in both reward- and stress-signaling pathways.
Energy-harvesting organelles (mitochondria and chloroplasts) use biological semiconductors such as Light-harvesting Complex (LHC) proteins, cytochromes, mitochondrial membrane protein complexes and ATP synthase which rely on homochiral aromatic L-amino acid peptides. These stereospecific chiral peptides can function both as energy boosters by polarizing electron spins, or as dissipaters/quenchers (in times of stress) by transducing damaging excess photons or electrons. The L-configuration of amino acids allows proper protein folding and electron delocalization in these proteins. These functions are disrupted by the same stressful xenobiotic stimuli that also upregulate microbial formation of D-amino acids.
Once in the intercellular space, D-amino acids, as the chiral opposites of L-amino acids, can disrupt their binding affinity and electron transfer efficiency by slowing down or even reversing the protein desaturation process. This effect can act like a semiconductor switch and lead to a transition from a thermodynamically-efficient “living” organized state to what Szent-Györgyi called the disorganized “proliferative alpha state,” a chaotic “cancerous” disruption to cellular homeostasis. In fact, we now know that cancer cells, as well as primitive bacteria and archaea found in deep-sea vents facing extremely harsh hot or alkaline environments can indeed reverse the Krebs cycle to build lipids and other organic compounds for self-sufficient survival and rapid chaotic growth in metabolically hostile ecosystems (Figure 1).
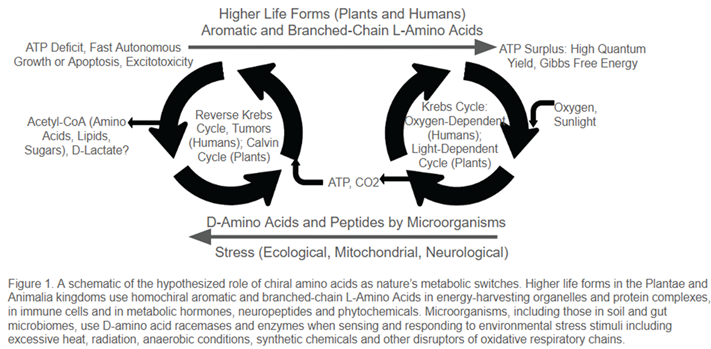
In humans, Szent-Györgyi had identified a D-amino acid (D-lactate) as the end product as well as a marker of the cancerous state that leads to reversal of Krebs cycle and damages to proteins and DNA structures. Recently, serum D-lactate levels have been suggested as a biomarker for diseases related to liver functions or intestinal [27] integrity.
In plants, abundance of D-amino acids can impair light-dependent photosynthesis, which relies on chiral L-amino acids (for light-harvesting complexes and cytochromes) and functions as the plant’s primary reductive mechanism in restoring the antioxidant pools of NADPH (Reduced Nicotinamide Adenine Dinucleotide Phosphate) and Glutathione, both essential in prevention of vicious metabolic/oxidative cycles under stress.
Based on these observations and the recently confirmed role of D-amino acids as biochemical modulators in the rhizosphere [28] and chemotactic stress signals [29] it is reasonable to hypothesize that D-amino acids from microbes, as sensitive front-line sensors of metabolically hostile ecosystems, play a key role in redox-signaling cascade networks in plants and humans by switching the directionality of electron spin and flow in the Krebs and photosynthetic cycles and ultimately causing a transition from an orderly metabolism in higher life forms to a microbial metabolism with low quantum yield. A recent study confirms [30] that the appearance of peptides, where L-amino acids are partially replaced by D-analogs, impacts the efficiency of the photoinduced electron transfer and is indeed a primary cause of some diseases. Another recent review [31] has provided evidence relating D-amino acids to mitochondrial dysfunctions and diabetic kidney disease.
Researchers have also recently demonstrated [32] that dysfunctions in Citrate Synthase, the first rate-limiting enzyme in the Krebs cycle (also called the Citric Acid Cycle), can switch the cellular metabolic direction from oxidative to reductive (glycolysis or anaerobic) Reverse-Krebs cycle. The enzyme relies on homochiral L-amino acids histidine and aspartate to distinguish pro-chiral carbons in the citrate molecule. Other enzymes in the citric acid cycle also rely on homochiral L-amino acids so one can make a cogent argument that D-amino acids are able to induce enzymatic dysfunctions which are linked to metabolic disruptions, diseases, cell death (apoptosis) and tumors.
Metabolic Diseases and Autoimmunity
High concentrations of D-amino acids found in certain plant toxins, antibiotics and pesticides are indeed shown to cause mitochondrial disruptions and induce apoptosis (programmed cell death). [33] Significantly elevated levels of D-aspartic and D-serine have been detected in human breast cancer cells. [19] Studies confirm that the level of D-amino acid oxidase (DAO) in the brain [5] or D-amino acids in blood, saliva, bone marrow or urine could serve as an early biomarker [34] for a wide range of conditions such as Alzheimer's disease, schizophrenia, chronic kidney disease, as well as leukemia, liver, colorectal and gastric cancers. We also now have experimental evidence [35] that malignant cancer cells have a much higher ability than healthy cells to metabolize and use specific D-amino acids. Even ascorbic acid (Vitamin C) which was shown by Szent-Györgyi to slow down the cancerous “alpha state” is bioactive in its natural L-ascorbic acid form and not the synthetic D-ascorbic acid. Generally, studies confirm [36] that the accumulation of D-amino acids and their metabolism by D-Amino acid oxidase (DAO), which produces hydrogen peroxide (a reactive oxygen species), is associated with age-related diseases and increased immune activation.
Human immune cells also rely on L-type Amino acid Transporters and homochiral L-amino acids such as L-arginine and L-tryptophan for proper activation, proliferation and balancing of proinflammatory cytotoxic activity and suppressive activity, which regulates inflammation and tissue repair. Dysregulations of amino acid metabolism have been linked to either immunological underactivity, as in tumor outgrowth and metastasis, or overactivity, as in autoimmune diseases like lupus erythematosus.
Because D-amino acid peptides in plant proteins resist proteolytic enzyme hydrolysis, they cannot be easily broken down in the digestive process and converted to L-amino acids in the body. The poor digestibility of denatured proteins and abundance of D-amino acids could act as a metabolic “denaturation” signal that activates the gut-associated lymphoid tissue (GALT), the largest immune system in the body, and cause inflammation. If the inflammation is chronic and the commensal gut bacteria are deficient or overwhelmed, it may result in a leaky gut, which allows undigested food proteins to enter the bloodstream. When the peptide’s amino acid types and sequence (regardless of chirality) are sufficiently similar to those in human cells, the immune system can produce antibodies against self-antigens and attack the body's own tissues and cells. The “molecular mimicry” of immunogens and toxins, which makes them indistinguishable from endogenous proteins to the immune cells, is established as a key trigger of severe allergic and autoimmune reactions. For example, D-aspartate and D-glutamate in both raw and roasted peanuts are shown to cause allergic reactions by inhibiting the immunoglobulin E (IgE) [37] in the consuming host’s immune system from binding to peanut allergens. Even the dreaded autoimmune condition categorized as “Gluten Resistance” is likely to be triggered by a combination of gut dysbiosis and ingestion of denatured D-amino acid peptides from stressed/sprayed crops. Gluten, the much-maligned protein in grains, when naturally composed of L-amino acids in living wheat germs, is clinically shown not to be responsible [38] for causing autoimmune symptoms in healthy individuals.
Autoimmune responses to D-amino acids may also be rooted in certain structural and functional similarities between microbial D-amino acid peptidoglycans and human/plant L-amino acid based glycoproteins, proteoglycans and glycine-based copolymers, which are denatured upon insertion of D-amino acids from microbial or pesticide actions. Increased levels of D-amino acids have been linked to motor neuron degeneration in the spinal cord [39] and abnormal locomotor activity. D-amino acids also interfere with the proteolytic function [40] of matrix metalloproteinases, which are involved in spinal tissue repair. D-amino acids could also have a putative role in denaturation of glycoproteins in myelin sheaths and triggering autoimmune reactions in the nervous system.
Conclusions
The convergence of literature reviewed here supports the role proposed for microbial D-amino acids as evolutionarily-conserved interkingdom ecological stress signals that are linked to numerous disruptions in photosynthetic and metabolic cycles in plants and humans, and to pathological disruptions in human neuroendocrine and immune systems. In this framework, the soil and gut microbiomes are front-line ecological biosensors of xenobiotic stressors and communicate via D-amino acids to higher life-forms.
Immunologically, D-amino acids not only disrupt L-type Amino acid Transporters but can also trigger self antigens and immune disruptions in the body of herbivorous animals and humans that consume products from such plants and animals because plant peptides that incorporate D-amino acids resist proteases.
Neurologically, D-amino acids, as agonists to excitatory NMDA receptors in the central nervous system, can overwhelm these glutamate receptors and cause neurotoxicity and neuropathic conditions.
Metabolically, D-amino acids can disrupt mitochondrial and photosynthetic cycles as well as the L-amino acid-containing hormones, neuropeptides and phytochemicals which have evolved as higher life forms’ response to biotic and abiotic stresses. It is plausible that D-amino acids are one of the key metabolites from rhizosphere microorganisms that act as stress signals priming or eliciting plants’ induced systemic resistance (ISR) against pathogens [41]. For example, D-amino acids in the rhizosphere are shown to stimulate ethylene production in some plants, which can counteract the effects of abscisic acid in promoting dormancy and inhibiting germination of seeds [42].
Because the activities of both bacteria-specific and broad-spectrum D-amino acid racemases/oxidases correlate with the intensity and duration of abiotic and biotic stresses, D-amino acids can register the spatio-temporal profile of ecological stress and hyper parasitism. For example, in low stress, mutualistic (diverse, non-pathogenic, non-parasitic) soil or gut microbial environments, the homeostatic (equilibrium) levels of D-amino acids metabolized by plant or human commensal bacteria remain low and not disruptive to photosynthetic and mitochondrial cycles or immune/defense systems. This is compatible with the properties of plant-disease suppressive soils, which as suggested by Husson et al., [1] support a range of Eh–pH niches and a diverse microbial Plant Soil community. We have also experimentally observed the same positive effect of microbial diversity and soil health in our own regenerative farm on the resistance of garlic and potato crops to enzymatic browning and fungal degradation [43].
In their classical 1987 textbook of Organic Chemistry, [44] Morrison and Boyd wrote: “We eat optically active bread & meat.. The proteins that make up our muscles, the glycogen in our liver and blood, the enzymes and hormones are all optically active.” Today, we know the chirality of these optical molecules is impacted by the chirality of metabolites of microbes that act as biosensors of ecological stress, and can electronically impact plant and human metabolism. When cued by microbial stress signals, higher life forms can switch from an orderly, efficient and surplus-driven metabolic state in times of peace, to one which is chaotic and deficit-driven in times of crisis, resulting in unregulated growth, immune reactions, inflammation and apoptosis. As such, chiral amino acids seem to control the evolutionary direction of life and tune “the curious harp of the human body” – the term coined by Francis Bacon– for cacophony or harmony.
References
- Husson, O. et al. Soil and plant health in relation to dynamic sustainment of Eh and pH homeostasis: A review. Plant and Cell (2021). [CrossRef]
- Porras-Dominguez, J., Lothier, J., Limami, A. M. & Tcherkez, G. d-amino acids metabolism reflects the evolutionary origin of higher plants and their adaptation to the environment. Plant, Cell & Environment (2024). [CrossRef]
- Aliashkevich, A., Alvarez, L. & Cava, F. New Insights Into the Mechanisms and Biological Roles of D-Amino Acids in Complex Ecosystems. Frontiers Microbiology (2018). [CrossRef]
- Vranová, V. et al. The significance of D-amino acids in soil, fate and utilization by microbes and plants: Review and identification of knowledge gaps. Plant and Soil (2011). [CrossRef]
- Grishin, D. V., Zhdanov, D. D., Pokrovskaya, M. V. & Sokolov, N. N. D-amino acids in nature, agriculture and biomedicine. All Life (2019). [CrossRef]
- Lazarus, L. H., et al. Frog skin opioid peptides: a case for environmental mimicry. Environmental Health Perspectives (1994). [CrossRef]
- Jimenez E. C., et al. Contryphan Is a D-Tryptophan-containing Conus Peptide. Journal of Biological Chemistry (1996). [CrossRef]
- Kho, J., Pham, P. C., Kwon, S., et al. De Novo Design, Synthesis, and Mechanistic Evaluation of Short Peptides That Mimic Heat Shock Protein 27 Activity. ACS Medicinal Chemistry Letters (2021). [CrossRef]
- Lovino, L., Tremblay, M. E. & Civiero L. Glutamate-induced excitotoxicity in Parkinson’s disease: The role of glial cells. Journal of Pharmacological Sciences (2020). [CrossRef]
- Scott, G.S., et al. Glutamate-stimulated peroxynitrite production in a brain-derived endothelial cell line is dependent on N-methyl-d-aspartate (NMDA) receptor activation. Biochemical Pharmacology (2006). [CrossRef]
- Forsum, O., et al. Capacities and constraints of amino acid utilization in Arabidopsis. New Phytologist Foundation (2008). [CrossRef]
- Joyce, J. G., Cook, J. C., Chabot, D. J., et al. Immunogenicity and Protective Efficacy of Bacillus anthracis Poly-γ-d-glutamic Acid Capsule Covalently Coupled to a Protein Carrier Using a Novel Triazine-based Conjugation Strategy. Journal of Biological Chemistry (2006). [CrossRef]
- Miles, J. J., Tan, M. P., Dolton G., et al. Peptide mimic for influenza vaccination using nonnatural combinatorial chemistry. Journal of Clinical Investigation (2018). [CrossRef]
- Csapo’, J., et al. The D-amino acid content of foodstuffs (A Review). Acta Univ. Sapientiae, Alimentaria, 2, 1 (2009) 5–30.
- Ariyoshi, M., Katane, M., Hamase, K., et al. D-Glutamate is metabolized in the heart mitochondria. Scientific Reports (2017). [CrossRef]
- Hernández, S. B. & Cava, F. Environmental roles of microbial amino acid racemases. Environmental Microbiology (2015). [CrossRef]
- Ewaschuk, J. B., Naylor, J. M. & Zello, G. A. D-Lactate in Human and Ruminant Metabolism. The Journal of Nutrition (2005). [CrossRef]
- Friedman, M. Chemistry, Biochemistry, Nutrition, and Microbiology of Lysinoalanine, Lanthionine, and Histidine Alanine in Food and Other Proteins. Journal of Agricultural and Food Chemistry (1999). [CrossRef]
- Hayashi, R. & Kameda, I. Racemization of Amino Acid Residues during Alkali-Treatment of Protein and Its Adverse Effect on Pepsin Digestibility. Agricultural and Biological Chemistry (1980). [CrossRef]
- Ewaschuk, J. B., Naylor, J. M. & Zello, G. A. D-Lactate in Human and Ruminant Metabolism. The Journal of Nutrition (2005). [CrossRef]
- Chanclud, E. & Lacombe, B. Plant Hormones: Key Players in Gut Microbiota and Human Diseases? Trends in Plant Science (2017). [CrossRef]
- Bastings, J. J. A. J. et al. D-amino Acids in Health and Disease: A Focus on Cancer. Nutrients (2019). [CrossRef]
- Sadeghi, M., Divangahi, M. Discovering adaptive features of innate immune memory. Immunological Reviews (2024). [CrossRef]
- Vivancos, P. D, Driscoll, S. P., Bulman, C. A, et al. Perturbations of Amino Acid Metabolism Associated with Glyphosate-Dependent Inhibition of Shikimic Acid Metabolism Affect Cellular Redox Homeostasis and Alter the Abundance of Proteins Involved in Photosynthesis and Photorespiration. Plant Physiology (2011). [CrossRef]
- Szent-Györgyi, A. The living state and cancer. Proc. Natl. Acad. Sci. (1977). [CrossRef]
- Turin, L. A Spectroscopic Mechanism for Primary Olfactory Reception. Chemical Senses (1996). [CrossRef]
- Remund, B., Yılmaz, B. & Sokollik C. D-Lactate: Implications for Gastrointestinal Diseases. Children (Basel) (2023). [CrossRef]
- Aliashkevich, A., et al. A D-amino acid produced by plant-bacteria metabolic crosstalk empowers interspecies competition. bioRxiv (2020). [CrossRef]
- Irazoki, O., et al. D-amino acids signal a stress-dependent run-away response in Vibrio cholerae. Nature Microbiology (2023). [CrossRef]
- Ageeva, A. A. et al. Chiral Linked Systems as a Model for Understanding D-Amino Acids Influence on the Structure and Properties of Amyloid Peptides. International Journal of Molecular Sciences (2022). [CrossRef]
- Thuy Linh, H., Nakade, Y., Wada, T. & Iwata, Y. The Potential Mechanism of D-Amino Acids - Mitochondria Axis in the Progression of Diabetic Kidney Disease. Kidney International Reports (2024). [CrossRef]
- Kang, W., Suzuki, M., Saito, T. & Miyado, K. Emerging Role of TCA Cycle-Related Enzymes in Human Diseases. International Journal of Molecular Sciences (2021). [CrossRef]
- Li, H., et al. Roles of d-Amino Acids on the Bioactivity of Host Defense Peptides. International Journal of Molecular Sciences (2016). [CrossRef]
- Pollegioni, L., et al. D-amino acids: New functional insights. The FEBS Journal (2025). [CrossRef]
- Du, S., Wang, Y., Alatrash, N., et al. Altered profiles and metabolism of l- and d-amino acids in cultured human breast cancer cells vs. non-tumorigenic human breast epithelial cells. Journal of Pharmaceutical and Biomedical Analysis (2019). [CrossRef]
- Yap, S. H., Lee, C. S., Zulkifli, N. D., et al. D-Amino acids differentially trigger an inflammatory environment in vitro. Amino Acids (2024). [CrossRef]
- Chung, S. Y. & Reed, S. IgE binding to peanut allergens is inhibited by combined D-aspartic and D-glutamic acids. Food Chemistry (2015). [CrossRef]
- Barnett, J.A. & Gibson, D. L., Separating the Empirical Wheat From the Pseudoscientific Chaff: A Critical Review of the Literature Surrounding Glyphosate, Dysbiosis and Wheat-Sensitivity. Frontiers in Microbiology (2020). [CrossRef]
- Bastings, J. J. A. J. et al. D-amino Acids in Health and Disease: A Focus on Cancer. Nutrients (2019). [CrossRef]
- Ndinguri, M. et al. Peptide-Based Selective Inhibitors of Matrix Metalloproteinase-Mediated Activities. Molecules (2012). [CrossRef]
- Thepbandit W., Athinuwat D. Rhizosphere Microorganisms Supply Availability of Soil Nutrients and Induce Plant Defense. Microorganisms (2024). [CrossRef]
- Kolukisaoglu Ü., D-amino Acids in Plants: Sources, Metabolism, and Functions. Int J Mol Sci. (2020). [CrossRef]
- Armat, R., Each Vegetable has a Unique Metabolic and Nutritional Fingerprint (2024) Farming/Health Blog Article on Simplescience.Substack.com.
- Morrison, R. T., Boyd, R. N. Organic Chemistry, Longman Higher Education (1987).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



