
Submitted:
02 June 2025
Posted:
02 June 2025
You are already at the latest version
Abstract
Copper (Cu) is an essential micronutrient for cyanobacteria, where it functions as a cofactor in key proteins involved in photosynthesis and antioxidant defense. However, at elevated concentrations, Cu becomes toxic, exhibiting algicidal effects by disrupting metal homeostasis and competing for metal-binding sites on critical cellular proteins. Due to the considerable morphological and physiological diversity within the phylum Cyanobacteria, the thresholds for Cu deficiency or toxicity vary considerably among strains. Maintaining Cu homeostasis in cyanobacterial cells is a complex process involving multiple layers of regulation. It begins at the extracellular polysaccharide layer, involves specialized membrane-bound proteins (in the outer, plasma, and thylakoid membranes), and results in transcriptional regulation in response to intracellular Cu status. This review summarizes the current understanding of Cu uptake and efflux pathways in cyanobacteria and explores how these mechanisms contribute to maintaining cellular Cu balance. The knowledge gained may contribute to the application of cyanobacteria in bioremediation strategies and/or the targeted use of Cu in the control of harmful cyanobacterial blooms.
Keywords:
Copper homeostasis
; cyanobacterial stress response
; copper toxicity.
1. Introduction
Cyanobacteria, formerly known as blue-green algae, are a monophyletic group of oxygenic photosynthetic prokaryotes [1]. They exhibit extensive physiological and morphophysiological diversity, which enables them to colonize nearly all photic environments on Earth [2]. Due to their metabolic versatility, cyanobacteria have gained attention for their potential in various biotechnological and industrial applications, including the production of biodiesel and biofertilizers [3,4,5,6,7,8,9]. Cyanobacterial strains also exhibit variable yet consistent bioabsorption and bioaccumulation capacities for a range of toxic metals, including copper (Cu), nickel (Ni), cadmium (Cd), lead (Pb), mercury (Hg), zinc (Zn), chromium (Cr), and manganese (Mn) [10]. These traits make them promising candidates for targeted bioremediation and the removal of organic and inorganic contaminants from wastewater [7,10,11,12]. Furthermore, the potential to repurpose cyanobacterial biomass for industrial applications creates new opportunities for integrated waste treatment in effluents from sugarcane processing [13], dairy industries [14], rice [15], maize [16], paper [17], and textiles [8].
Cu is an essential micronutrient for cyanobacteria, playing a critical role in fundamental metabolic pathways. It acts as a cofactor in several enzymes involved in redox reactions, notably in photosynthesis, where it is crucial for the proper function of plastocyanin (PC) [18]. In the respiratory chain, Cu is a critical component of cytochrome c oxidase enzyme, which catalyzes the reduction of oxygen to water [19]. In addition, Cu contributes to the oxidative stress response through its association with Cu/Zn-superoxide dismutase (Cu/Zn-SOD), which catalyzes the dismutation of superoxide radicals into oxygen and hydrogen peroxide [20]. However, despite its biological importance, at elevated concentrations, Cu can excert toxic effects on cyanobacteria [21]. Anthropogenic activities, including mining and the paint manufacturing, have significantly increased Cu levels in aquactic environments [22]. At elevated concentrations, Cu competes with other ions, particularly iron, for binding sites within the context of Fe-S containing enzymes. This can result in metabolic imbalances and subsequent cellular stress [11].
Here, we explored the molecular and physiological mechanisms underlying Cu uptake and response in cyanobacteria, with an emphasis on their potential contributions and application in environmental remediation strategies.
2. Cu Essentiality, Deficiency, and Toxicity
Given its classification as a micronutrient, minimal quantities of Cu are required for its supply to cyanobacteria. Due to their important role in redox reactions, the absence of Cu can result in deficiencies in processes such as photosynthesis and cellular respiration in cyanobacterial cells. Furthermore, there is the possibility that it will influence responses to environmental conditions due to the disruption of the homeostasis of this element.
To date, the paucity of research in this area is evident, with the majority of studies conducted on Synechocystis sp. PCC 6803. For example, Castielli et al. (2009) observed that in the absence of Cu, the strain reduced the expression of PC, which contains Cu ions coordinated by amino acid residues, and increased the expression of cytochrome c6 (Cyt c6), which contains iron [23]. This substitution ensures that the cell can maintain the electron transport chain in Cu deficiency by using Cyt c6 as an alternative electron donor to photosystem I (PSI), growing at rates similar to those observed under Cu-sufficient conditions. García-Canas et al. (2021) further explored the regulation of the PC/Cyt c6 switch in the absence of Cu in Synechocystis sp. PCC 6803, which is regulated by the BlaI/CopY-family transcription factor PetR and the BlaR-membrane protease PetP [24]. PetP is a Cu-sensing membrane protease that regulates PetR. In the presence of Cu, PetP degrades PetR, thereby enabling elevated expression of PC. Conversely, in the absence of Cu, PetP becomes inactive and can no longer degrade PetR. Thus, PetR represses the expression of petE (the gene encoding PC) under limiting Cu, while simultaneously activating the transcription of petJ (the gene encoding Cyt c6) [24]. By varying its concentration, copper can be used in metal-regulated promoters responsive to this element to induce or repress the expression of specific genes, making it a highly functional tool for research routines. In addition to the 'pet' system in cyanobacteria, copper-inducible genes have also been described in fungi and Escherichia coli [24,25,26].
The absence of literature on suboptimal doses of Cu indicates a potential area for further exploration in the cyanobacteria field. Furthermore, the doses of Cu utilized in various media for culturing cyanobacteria can provide valuable insights into the optimal utilization of suboptimal doses. The BG-11 medium, the most well-known and commonly used medium for culturing freshwater, non-nitrogen-fixing cyanobacteria, contains 0,316 μM of CuSO₄·5H₂O [27]. The ASN-III medium, utilized to replicate the marine environment for cyanobacteria from such environments, also contains 0,316 μM of CuSO₄·5H₂O [28]. The Z8 medium, which does not contain NaCl and is ideal for cyanobacteria that do not tolerate salinity, contains 5,01 µM of CuSO4·5H2O [29]. Finally, the Zarrouk medium, an alkaline medium used as a standard for Arthrospira platensis to promote growth and maximum biomass gain, also contains 0,316 μM of CuSO₄·5H₂O [30].
In contrast to the paucity of research on Cu deficiency in cyanobacteria, there is a substantial body of literature on the effects of Cu excess, most likely due to the algicidal potential of Cu [31]. The several detrimental physiological impacts of elevated Cu levels in cyanobacteria encompass, but are not limited to, growth inhibition, diminished photosynthetic rates, elevated reactive oxygen species (ROS) generation, and morphological and protein destabilization [31].
In the strain Leptolyngbya sp. GUEco1015, Kalita and Baruah (2024) emphasised the capacity for Cu bioaccumulation and documented the consequences of elevated Cu doses [21]. The LC50 of Cu2+ for this strain was determined to be 0.9 ppm (or 14.17 μM). Excessive doses led to severe chlorosis, filament disorganization, and a twofold increase in cellular H2O2 and MDA concentrations when comparing control (standard BG-11) with a dose of 1.5 ppm (or 23.6 µM) [21]. Moreover, Ahad & Siyem (2018) tested 10 µM Cu2+ in the strain Nostoc muscorum and observed effects similar to those described above, including chlorosis and filament disorganization. The aforementioned treatment resulted in a ~38% reduction in chlorophyll content, a ~20% decrease in total protein content, and a ~40% decline in photosynthetic potential [32].
The development of new technologies for harmful cyanobacterial bloom control could be based on the impact of Cu on harmful species such as Microcystis aeruginosa. Wu et al. (2007) demonstrated that, after 24 hours of exposure to 0.25 mg (or 3.93 µM) of Cu, the cyanobacterial phenotype plays a crucial role in Cu tolerance. Strains exhibiting unicellular morphology demonstrated a loss of total cell viability exceeding 95%, while colonial strains exhibited viability levels exceeding 70% under all treatment conditions [33]. Previous data observed for Microcystis aeruginosa show that, at least, strains with unicellular morphology are substantially more sensitive to copper than many microalgae [34], reinforcing the potential of this element for controlling this potentially harmful species.
Furthermore, a decrease in total protein content and an increase in free amino acids were also observed in Spirulina platensis. In a separate study, Mohy El-Din (2017) found that exposure to 0.5, 1.0, and 3.0 mg (or 7.87 µM, 15.74 µM and 47.21 µM) of Cu2+ resulted in a continuous decline in total protein content (from 493.63 mg/L in the control to 182.47 mg/L at 3.0 mg) and a steady increase in free amino acids (~12.5 mg/L in the control to ~30 mg/L at 3.0 mg) [35]. Finally, in the model strain Synechocystis sp. PCC 6803, Giner-Lamia et al. (2014) observed that at a concentration of 3 µM Cu²⁺, ROS production is nearly five times higher than at the standard concentration found in BG-11 medium [36].
In summary, while the toxic effects of Cu on cyanobacterial strains are broadly similar in nature, the concentration threshold that elicits these effects varies considerably. This variability reflects the substantial morphological and metabolic diversity within cyanobacterial phylum, making it likely that different strains exhibit distinct responses to equivalent Cu concentration. As such, determining a specific LD50 for each strain may be required to accurately characterize their sensitivity to Cu toxicity. Importantly, this review highlights an apparent trend of increased Cu sensitivity among unicellular strains compared to filamentous ones. Evidence suggests that Cu concentrations up to 10 µM can induce pronounced metabolic disruptions in unicellular cyanobacteria, whereas filamentous strains typically require higher concentrations to exhibit comparable physiological impairments.
3. Cu Transport: Extracellular to Intracellular Space
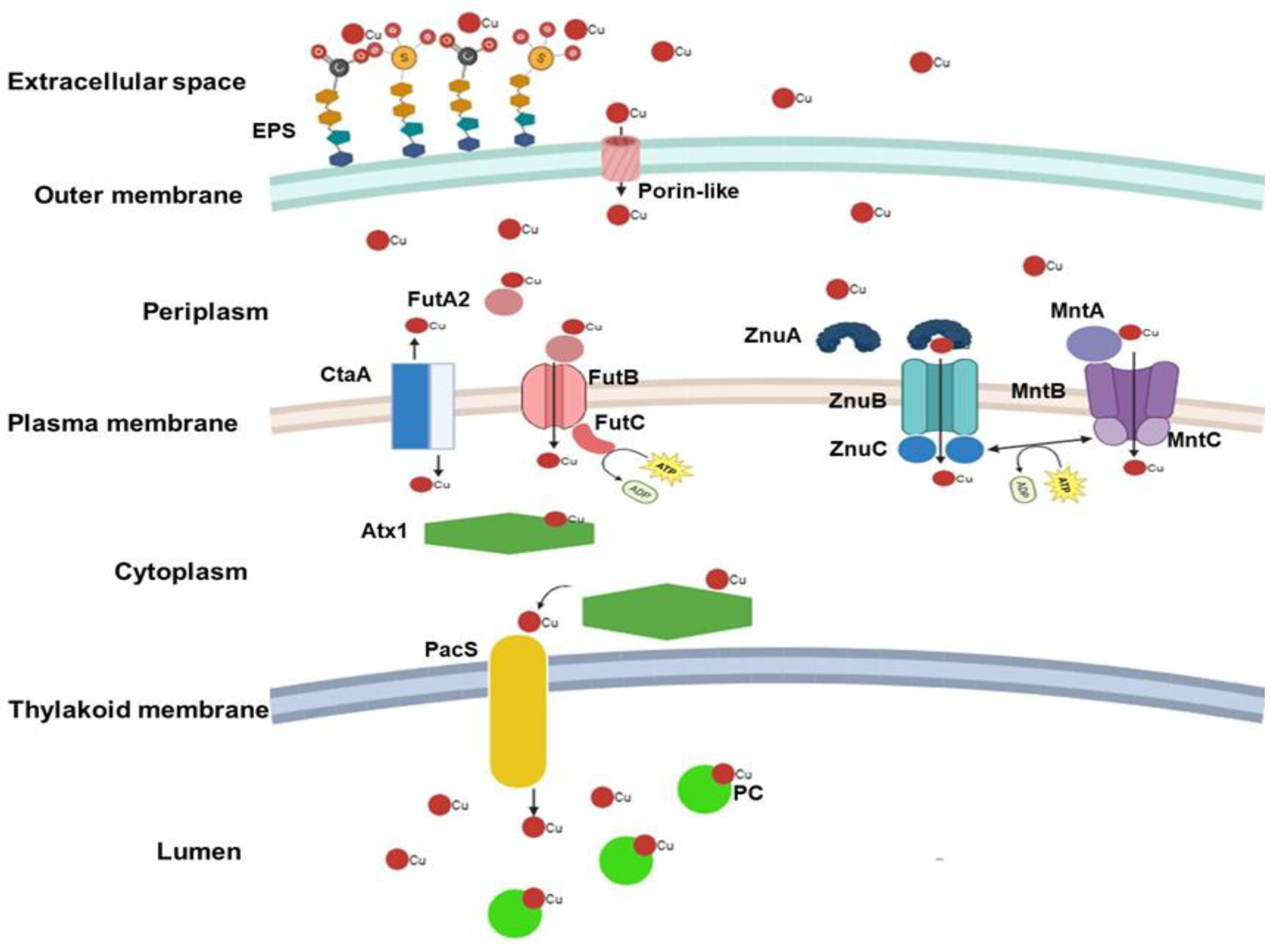
Porins are well-known for facilitating the passive entry of Cu²⁺ from the extracellular space to the periplasm in bacteria [37,38]. The outer membrane of strain Synechocystis sp. PCC 6803 contains abundant porin-like proteins such as Slr1841, Slr1908, and Slr0042 that have been reported as the main mediators of iron transport; however, they also exhibit high permeability to other inorganic ions. This suggests that ions such as Cu²⁺ may have rapid access to the periplasm through these protein pores [39]. These proteins contain an N-terminal S-layer homologous (SLH) domain bound to the polysaccharide moiety of the peptidoglycan and a C-terminal transmembrane region that is predicted to form a β-barrel structure [39,40].
The OprB domain of porins in bacteria was initially described as a specific domain for the recognition and transport of sugars [41]. However, in seven mutants of Anabaena sp. PCC 7120 missing different OprB-domain porins, most putative porins appear to play a role in Cu transport, as these mutants exhibited higher resistance to elevated Cu concentrations [42]. Furthermore, the OprB domain has been demonstrated to facilitate interaction and selection in the entry of carbohydrates from the extracellular space into the periplasm in cyanobacteria [43,44].
Cu could also enter cyanobacterial cells as Cu2+ through competition mechanisms with other metal transporters, using ABC-type metal transport systems such as the Zn/Mn transporter (ZnuABC/MntABC). The Zn system is well-documented and relatively conserved across several bacterial groups. It is particularly well-investigated in Synechocystis sp. PCC 6803 [45,46]. Once in the periplasm, Cu2+ binds to the high-affinity protein ZnuA and is subsequently delivered to the inner membrane complex ZnuBC, which then transports Cu2+ into the cytoplasm [45]. In Synechocystis sp. PCC 6803 and E. coli, Cu2+ has been observed to bind to the histidine-rich site of the ZnuA protein, indicating at least one periplasmic interaction between Cu2+ and Zn-importing proteins [45,47,48]. The transfer of Cu2+ from the ZnuA to the membrane permease protein ZnuB is required for the translocation of Cu2+ into the cytosol. The function of ZnuC, an ATPase subunit, is to provide energy to the complex to facilitate active transport [49]. Further studies are required to elucidate the interaction of Cu2+ with the ZnuB and ZnuC proteins. However, expression data from Nostoc punctiforme have demonstrated that the entire operon is responsible under Cu and other metal treatments [50], suggesting that the range of substrates for the Znu system is broad and includes Cu.
It has been established that, due to the chemical similarity between Cu2+ and Mn2+ ions, Cu2+ may also enter cyanobacterial cells through the high-affinity Mn2+ import system, MntABC. This system functions via a binding and transport mechanism analogous to that of ZnuABC, as has been previously reported and studied in Synechocystis sp. PCC 6803 [51]. While studies quantifying the interaction between Cu2+ and MntABC are lacking in cyanobacteria, the entry of Cu into the cytoplasm through this system has been demonstrated in Staphylococcus aureus and Zn entry in Neisseria gonorrhoeae [52,53].
The only exclusive, well-known protein that facilitates the import of Cu2+ from the periplasmic space through the plasma membrane into cyanobacterial cells is CtaA [54]. CtaA belongs to the P1B-type ATPase family, which comprises specialized proteins that facilitate the translocation of transition metal ions, including Cu²⁺, Zn²⁺, Cd²⁺, and other essential or potentially toxic heavy metals [55]. The structure of the CtaA protein is characterized by the presence of 6 or 8 membrane-spanning alpha-helices, which form specific channels for the passage of metal ions. It also contains conserved residues (mostly histidine) in the N-terminal region that coordinate metal ions, acting as sensors or facilitators of transport, as well as ATP-binding domains [56,57,58]. While certain studies suggest that CtaA's primary function is the efflux of Cu2+ to the periplasm [59], others provide evidence, including in cyanobacteria, that this protein also contributes to Cu import and not solely to export for detoxification purposes. Generally, mutants missing these proteins exhibit a phenotype resistant to excess Cu [46,60,61].
In addition to the CtaA protein, the FutABC iron transport system has also been demonstrated to participate in Cu transfer to the cytoplasm [62]. Notably, the FutABC system is recognized as a factor in iron and Cu homeostasis in cyanobacterial cells. In this context, the FutABC transport system (primarily responsible for importing iron from the periplasm into the cytoplasm) located in the plasma membrane of cyanobacteria, actively influences the cellular Fe/Cu ratio. The FutABC system is composed of the proteins FutA1 and FutA2, which are responsible for Fe binding in the periplasm, FutB, which is a pore in the inner membrane, and FutC, which is an ATPase [63]. It has been demonstrated that Synechocystis PCC 6803 futA2 mutants accumulate Cu in the periplasm [64]. Interestingly, futB expression is altered under high Cu concentrations in Anabaena sp. PCC 7120 [65].
The cytoplasmic space is typically a predominantly reducing environment, and metal ions such as Cu2+ tend to be reduced to the Cu+ form by molecules such as glutathiones and thioredoxins [66,67]. Once in the cytosol, Cu+ binds to the metallochaperone Atx1, which 'delivers' this ion to another P1-type ATPase protein complex, PacS. Atx1 features a ferredoxin-like βαββαβ fold in its structure, with a CXXC motif (two cysteine residues separated by any two amino acids) responsible for Cu⁺ binding. The β-α-β-β-α-β structure has been shown to stabilize the protein core and create the environment for metal binding, while the CXXC motif has been demonstrated to form covalent bonds with metal ions such as Cu+ [68,69,70].
PacS is responsible for transporting Cu⁺ into the thylakoid lumen and supplying Cu+ where it is required for plastocyanin [11,54]. Notably, studies on Synechococcus PCC 7942 indicate that Atx1 interacts with the amino-terminal domains of both PacS and CtaA and is designed to shuttle Cu between them, with evidence suggesting that Atx1 may also positively influence Cu-dependent thylakoid proteins [46].
A final factor that may impact Cu uptake by cyanobacterial cells is the presence of metallothioneins in the cytoplasm and their interaction with Cu ions. Metallothioneins are metal-binding peptides rich in cysteine and capable of binding a wide range of metal ions [71,72]. Although the classical role of these peptides is the sequestration of metal ions in general to prevent their excess from becoming harmful [73], it has been demonstrated in cyanobacteria that they may also participate in Cu uptake, probably in a "secondary" manner by sequestering free Cu in the cytoplasm and thus creating a favorable gradient for the entry of more Cu into the cell [74].
Figure 1.
Schematic representation of the passage of Cu ions from the extracellular environment into cyanobacteria cell. Interaction of Cu ions with negative charges present in the EPS layer. Entry by diffusion through porin-like structures into the periplasm and via the CtaA and FutABC (ATP-Binding Cassette transporter – iron ABC) proteins into the cytoplasm. Alternatively, passage into the cytoplasm by competing with other metal import systems such as Zn (ZnABC) and Mn (MntABC) - ATP-Binding Cassette transporters – ABC.
Figure 1.
Schematic representation of the passage of Cu ions from the extracellular environment into cyanobacteria cell. Interaction of Cu ions with negative charges present in the EPS layer. Entry by diffusion through porin-like structures into the periplasm and via the CtaA and FutABC (ATP-Binding Cassette transporter – iron ABC) proteins into the cytoplasm. Alternatively, passage into the cytoplasm by competing with other metal import systems such as Zn (ZnABC) and Mn (MntABC) - ATP-Binding Cassette transporters – ABC.
4. Response to Excess Cu
As elevated levels of Cu are toxic to cyanobacteria cells, they have evolved an active Cu extrusion system, the expression of which is regulated by the Cu concentration inside the cell [36]. In Nostoc calcicola, it was observed that the absorption process of Cu2+ occurs in two phases: 1) a rapid binding of ions to the cell wall through charge interactions (around 10 minutes), and 2) the slower entry of ions into the intracellular environment (40 minutes to 1 hour) [75]. The uptake and transport of ions across membranes are enhanced in photosynthetically active cells once they are exposed to light, ATP, and a proton gradient, as has been proven that the ATP responsible for Cu transport is derived from PSII reactions [75].
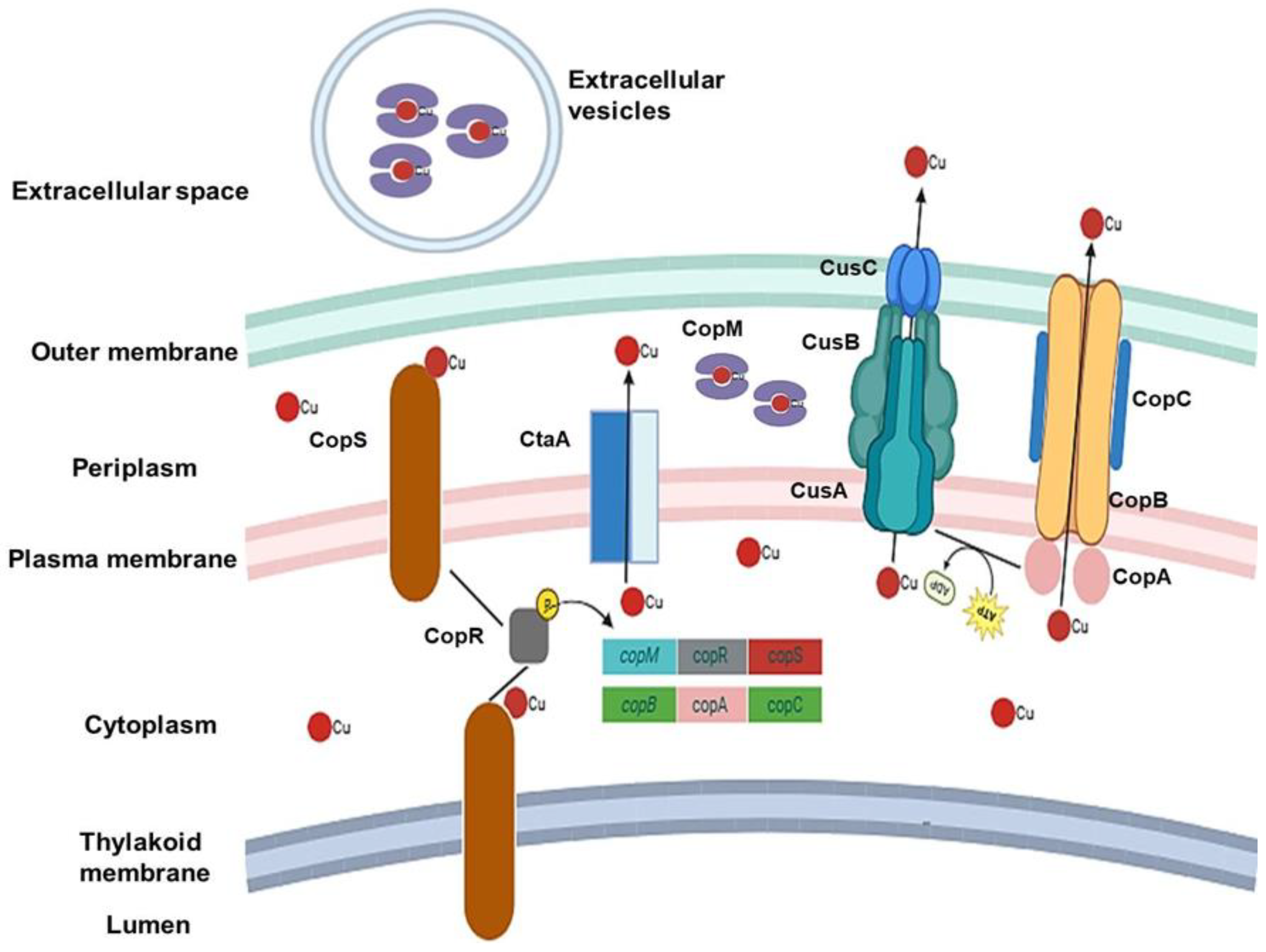
As demonstrated in the case of Synechocystis sp. PCC 6803, the subject of most studies, the presence of excess Cu²⁺ ions within the cytoplasm instigates the activation of a two-component system designated as CopRS, which belongs to the Resistance-Nodulation-Cell Division (RND) family. This system plays a pivotal role in the export of toxic metals and is responsible for Cu resistance [36,54]. Initially, CopS, a histidine kinase (HK) protein associated with the plasma and thylakoid membranes, detects Cu ions and binds them with high affinity [54]. Studies on the chromosome of Escherichia coli K-12 suggest that CopS has two transmembrane domains, with peptide loops extending into the periplasmic space [76]. CopS appears to be involved in detecting Cu ions throughout the intermembrane region and serves as the first step in regulating homeostasis in cases of Cu excess [11,54]. As the concentration of Cu increases, it is hypothesized that the kinase domain of the CopS protein phosphorylates its cognate response regulator, CopR, thereby converting it into a transcriptional activator for its respective target operons [76,77,78]. The activation of CopR appears to be directly associated with increased transcription of two closely related Cu homeostasis operons, copMRS and copBAC. In Synechocystis sp. PCC 6803, the operons are located on the native plasmid pSYSX, with copMRS genes also present in the chromosomal genome of the strain [79]. Studies have shown that in Synechocystis sp. PCC 6803 mutants lacking the copR gene have reduced expression of these operons and heightened sensitivity to Cu excess [36]. As mentioned above, the action of the transcriptional activator CopR on the copMRS operon not only increases the transcription of the copRS system but also enhances the expression of the protein CopM [36]. CopM is a metallochaperone, already identified in cytosolic, periplasmic and extracellular spaces of cyanobacteria, capable of interacting with both Cu²⁺ and Cu⁺. It appears that CopM can help prevent Cu stress in the extracellular environment, suggesting an interaction that immobilizes Cu ions outside the cell [36]. The structure of the CopM protein was resolved by Zhao et al. (2016), revealing it to be a dimeric periplasmic protein forming a bundle of six helices [80]. These helical bundles can bind multiple Cu ions through their abundant methionine and histidine residues [81]. Although previously completely unknown, the mechanism of CopM export to the extracellular space has recently been partially uncovered by the identification of the presence of multiple extracellular vesicles isolated from Synechocystis sp. PCC 6803 containing CopM interacting with Cu ions [82].
A higher intracellular concentration of Cu is required for CopR to activate the transcription of the copBAC operon compared to the concentration required to enhance copMRS expression [36]. The copBAC operon encodes a heavy metal efflux system, the CopBAC, classified as an HME-RND (heavy metal efflux/Resistance-Nodulation-Cell Division) type system [36]. This complex spans both the plasma and outer membranes of cyanobacterial cells, extending across the periplasmic space [78].
Although detailed subcellular localization studies of each CopBAC subunit are limited, research on Synechocystis sp. PCC 6803 has identified CopB as a membrane fusion protein located in the periplasm, while CopA is a P-type ATPase RND associated with the inner membrane [79]. While not fully elucidated in cyanobacteria, CopC is recognized as a periplasmic Cu-binding protein with distinct binding sites for both Cu⁺ and Cu²⁺, likely involved in shuttling Cu to the efflux channel [83]. Comparative studies of Cu resistance mechanisms in other bacterial species have contributed to our understanding of such systems; however, the degree of homology between these bacterial and cyanobacterial proteins involved in Cu transport and resistance remains unclear and warrants further investigation [84,85,86,87,88].
The components responsible for Cu response and efflux discussed in this section until here have been well characterized only in Synechocystis sp. PCC 6803 within the Cyanobacteria phylum, but it was possible to demonstrate that CopA (and probably CopB and CopC) is present throughout the phylum (Figure 4). Transcripts highly similar to copM have already been identified in Anabaena sp. PCC 7120 [89]. Other studies have shown that the CopR, CopS, and CopM components also exist in other bacterial phyla [81,90,91]. Therefore, while the presence of all these proteins requires experimental verification in other cyanobacterial strains, it is plausible that this system is conserved throughout the group.
Another system poorly studied in cyanobacteria but likely wide-spread in the phylum is the RND Cu and silver efflux system CusCBA, which is well described in the literature for E. coli [92]. The CusA protein is a proton-motive-force-dependent inner membrane RND efflux pump. CusC is a trimeric outer membrane porin that allows Cu to be directly expelled into the extracellular medium, while CusB is a hexameric membrane fusion protein capable of forming a structural bridge between CusA and CusC [93]. Studies in Anabaena sp. PCC 7120 have recorded the expression of genes homologous to cusA and cusB from E. coli, indicating that this system may also be present and play a role in Cu homeostasis in cyanobacterial cells [89,94,95].
Metallothioneins, already briefly discussed in the previous section, also play the classical role of Cu detoxification in cyanobacteria. Their binding to metal ions helps the cell reduce the excess of free ions. An example of a metallothionein in cyanobacteria is SmtA, which is composed of 56 amino acids. In Synechococcus PCC 7942, its expression is upregulated in response to excess Zn, Cd, and Cu, showing the highest responsiveness to elevated Zn levels [46].
Figure 2.
Schematic representation of copper (Cu) ion response and efflux mechanisms in cyanobacteria. This diagram illustrates the cellular processes involved in sensing and transporting Cu ions from the intracellular environment to the extracellular space. CopR/S (copper regulator/sensor proteins) are responsible for sensing Cu excess and increasing the transcription of related operons (copMRS and copBAC). The CopM (copper binding protein), CopBAC and CusABC (ABC efflux systems) complexes are directly responsible for the efflux of Cu ions.
Figure 2.
Schematic representation of copper (Cu) ion response and efflux mechanisms in cyanobacteria. This diagram illustrates the cellular processes involved in sensing and transporting Cu ions from the intracellular environment to the extracellular space. CopR/S (copper regulator/sensor proteins) are responsible for sensing Cu excess and increasing the transcription of related operons (copMRS and copBAC). The CopM (copper binding protein), CopBAC and CusABC (ABC efflux systems) complexes are directly responsible for the efflux of Cu ions.
5. Exopolysaccharides (EPS) Interactions
EPS function as a pivotal interface between cyanobacterial cells and their surrounding environment. While glucose predominates as the sugar-building block in EPS, other sugars, including rhamnose, xylose, arabinose, fucose, mannose, and uronic acids, have also been observed in certain cyanobacterial strains [96]. The classification of EPS is typically performed into two categories: capsular exopolysaccharides (CPS) and released exopolysaccharides (RPS). CPS have been observed to maintain a tight association with the cell surface, manifesting as sheaths, capsules, or slime layers. In contrast, RPS are known to be excreted into the extracellular space as free-floating polymers [97]. It has been demonstrated that both CPS and RPS exhibit distinct chemical and morphological characteristics. However, RPS has garnered greater industrial interest due to their larger production yields and greater independence in environmental interactions, facilitating metal biosorption [98,99,100,101,102].
It has been demonstrated that EPS are implicated in several essential biological functions, including the provision of structural integrity, facilitation of motility, and protection from various abiotic stresses [96]. The matrix of EPS is rich in charged functional groups, including carboxyl and hydroxyl side chains from deoxy-sugars, sulfate, and uronic acids [103]. This property enables EPS to interact with cationic species in solution. This interaction suggests that EPS may serve as a site for metal ion sequestration [104]. The interaction of EPS from various strains (whether bound to cells or isolated) with a wide range of environmental metal ions, including Cu, Pb, As, Cd, Cr, and Zn, has been shown to result in a significant reduction of these ions in the environment [105]. The overall negative charge of cyanobacterial EPS is particularly significant for the chelation of essential metal cations, which are present in trace amounts in the environment, as well as for mitigating direct contact with toxic heavy metals [98,103]. It has been demonstrated that an increase in Cu2+ concentrations can stimulate the production of EPS in Nostoc spongiaeforme [106], while Microcystis aeruginosa exhibits a high affinity for Cu2+ ion adsorption, primarily through complexation [100]. Furthermore, substantial Cu²⁺ removal via EPS has been reported in Cyanospira capsulata and Nostoc PCC 7936 [107].
Cu has been found to be one of the most efficiently adsorbed metals by cyanobacteria, surpassing other metal ions such as Ni and Zn [102]. Cu2+ ions have been observed to form complexes with anions such as CO32-, OH-, and Cl-, as well as organic complexes with ligands including thiols, EPS, and humic substances [108]. The phenomenon of biosorption is characterised by the rapid, reversible, and passive binding of metal ions to functional groups present on the EPS surface, thereby protecting the cells themselves from the high concentration of metal ions [102]. However, it is important to note that the dynamics of the Cu-EPS interaction may be modified upon interaction with Cu2+ or other metal ions. For instance, alterations in pH have been demonstrated to influence the adsorption capacity and the specific binding sites between Cu2+ and EPS, as evidenced by studies involving EPS extracted from sludge in wastewater treatment [109].
6. Phylogeny and Structure of the CopA Protein
The structure of the CopA protein acting as P-type ATPase in Cu transport is probably the best studied among all the systems discussed in this review, as its primary structure is the most represented in online repositories, such as the NCBI (National Center for Biotechnology Information), and spans a diverse range of prokaryotic groups.
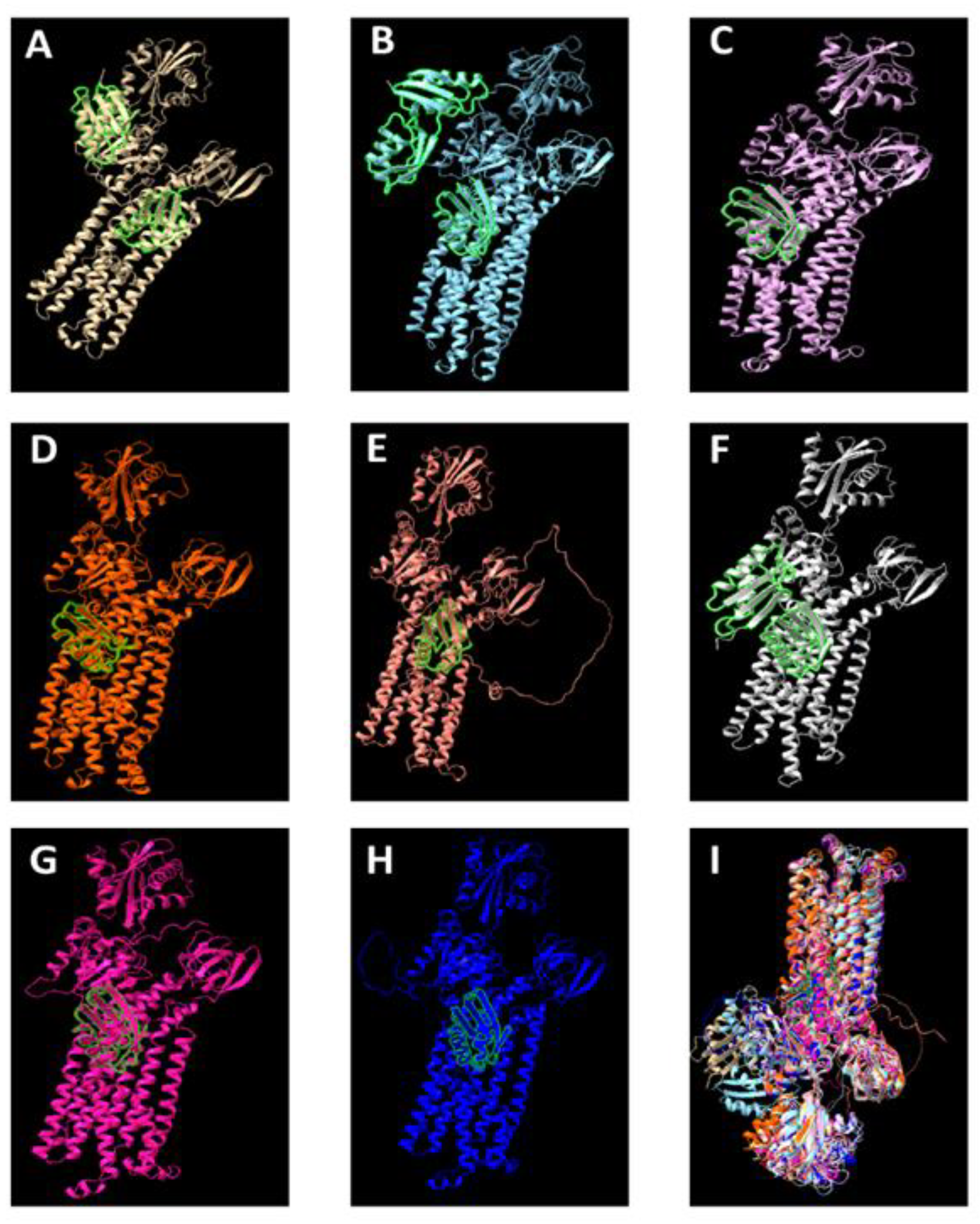
The three-dimensional structure of the protein revealed that CopA is slightly smaller in the cyanobacterial phylum compared to other prokaryotic groups (Figure 3). This is because in cyanobacteria, CopA presents a Heavy-Metal-Associated domain and a P-type ATPase Cu-like domain, whereas in other groups, CopA contains two Heavy-Metal-Associated domains and one P-type ATPase Cu-like domain. Whether this is a unique and universal characteristic of the cyanobacterial phylum still requires further investigation.
The phylogeny of the P-type ATPase Cu-like domain of the CopA protein showed a clear division into three branches: cyanobacteria, other prokaryotes, and Archaeoglobus (Archaea) (Figure 4). This division is consistent with the phylogenetic history of these groups. Moreover, within the cyanobacterial branch, the phylogeny appeared to follow a pattern similar to that observed in trees based on the 16S rRNA molecular clock, indicating that this system is vertically distributed throughout the phylum as an ancient Cu homeostasis mechanism. This view is further supported by its well conserved structure.
Figure 4.
Phylogeny of the functional domain P-type ATPase Cu exportation CopA protein in cyanobacteria. Sequences with a single chain of approximately 750 amino acids and an identity > 65% among them. Archaea in blue, cyanobacteria in green, and other bacteria in red.
Figure 4.
Phylogeny of the functional domain P-type ATPase Cu exportation CopA protein in cyanobacteria. Sequences with a single chain of approximately 750 amino acids and an identity > 65% among them. Archaea in blue, cyanobacteria in green, and other bacteria in red.

7. Conclusions
This review underscores the critical role of copper (Cu) homeostasis in cyanobacterial physiology, with a focus on the molecular components that mediate Cu uptake—from the extracellular environment to the thylakoid lumen—and Cu efflux under conditions of excess. We highlight key proteins involved in these pathways, their subcellular localization, and the genes regulated in response to both Cu deficiency and toxicity. Particular attention is given to the potential involvement of extracellular polymeric substances (EPS) in Cu binding and detoxification. These insights provide a more integrated understanding of the complex Cu transport network in cyanobacteria, offering a foundation for future applications in bioremediation and the development of Cu-based strategies for controlling harmful cyanobacterial blooms in aquatic ecosystems.
Additionally, we identified key gaps in current knowledge, including the need for further investigation into Cu limitation and the effects of sub-toxic concentrations, as well as the functional characterization of poorly studied systems such as CusABC. Our phylogenetic and structural analysis of the CopA protein reveals notable findings, including the widespread presence of the CopBAC system across the phylum and the unique feature of cyanobacterial CopA harbouring a single functional metal-binding domain—contrasting with the dual-domain structure observed in many other prokaryotes. Importantly, here, we compiled data from a wide range of cyanobacterial strains beyond the commonly used model organisms, aiming to offer a more comprehensive overview of Cu homeostasis across the phylum.
Author Contributions
Conceptualization J.C.O.; investigation J.C.O. and L.A.L.; resources and data curation L.A.L., W.L.A., A.N.N., L.B.S. and T.E.S; writing—original draft preparation J.C.O.; writing—review and editing J.C.O., L.A.L., W.L.A., A.N.N., L.B.S. and T.E.S; supervision L.A.L. and W.L.A.; funding acquisition L.A.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Institüt für Biowissenschaften, Abteilung Pflanzenphysiologie, Universität Rostock.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank Dr. Marcelo Gomes Marçal Vieira Vaz and Dr. João Antônio Batista de Siqueira for their insights in the development of Figures 3 and 4, MSc. Ana Luiza Viana Silva for her assistance in the revision of the text, and the Department of Plant Physiology, University of Rostock, for funding and contribution.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Cu | copper |
| EPS | exopolysaccharides |
| Ni | nickel |
| Cd | cadmium |
| Pb | lead |
| Hg | mercury |
| Zn | zinc |
| Cr | chromium |
| Mn | manganese |
| PC | plastocyanin |
| SOD | superoxide dismutase |
| Fe-S | iron-sulfur |
| PCC | Pasteur Culture Collection |
| Cyt | c6 cytochrome c6 |
| PSI | photosystem I |
| ROS | reactive oxygen species |
| LC50 | lethal concentration 50 |
| SLH | S-layer Homologous |
| PSII | photosystem II |
| RND | Nodulation-Cell Division |
| HK | histidine kinase |
| CPS | capsular exopolysaccharides |
| RPS | released exopolysaccharides |
| NCBI | National Center for Biotechnology Information |
References
- Sinha, R.P.; Häder, D.-P. UV-protectants in cyanobacteria. Plant Sci. 2008, 174, 278–289. [Google Scholar] [CrossRef]
- De Los Ríos, A.; et al. Ultrastructural and genetic characteristics of endolithic cyanobacterial biofilms colonizing Antarctic granite rocks. FEMS Microbiol. Ecol. 2007, 59, 386–395. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Chittora, D.; et al. Cyanobacteria as a source of biofertilizers for sustainable agriculture. Biochem. Biophys. Rep. 2020, 22, 100737. [Google Scholar] [CrossRef] [PubMed]
- Żymańczyk-Duda, E.; et al. Versatile applications of cyanobacteria in biotechnology. Microorganisms 2022, 10, 2318. [Google Scholar] [CrossRef]
- Zhang, H.; et al. Characteristics of γ-hexachlorocyclohexane biodegradation by a nitrogen-fixing cyanobacterium, Anabaena azotica. J. Appl. Phycol. 2012, 24, 221–225. [Google Scholar] [CrossRef]
- Cepoi, L.; et al. Removal of organic pollutants from wastewater by cyanobacteria. In: Cyanobacteria for Bioremediation of Wastewaters 2016, 27–43.
- Parikh, A.; Madamwar, D. Textile dye decolorization using cyanobacteria. Biotechnol. Lett. 2005, 27, 323–326. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Tiwari, B.S. Cyanotherapeutics: An emerging field for future drug discovery. Appl. Phycol. 2020, 1, 44–57. [Google Scholar] [CrossRef]
- Kalita, N.; Baruah, P.P. Cyanobacteria as a potent platform for heavy metals biosorption: Uptake, responses and removal mechanisms. J. Hazard. Mater. Adv. 2023, 100349. [Google Scholar] [CrossRef]
- Huertas, M.J.; et al. Metals in cyanobacteria: analysis of the copper, nickel, cobalt and arsenic homeostasis mechanisms. Life 2014, 4, 865–886. [Google Scholar] [CrossRef]
- Chevalier, P.; et al. Nitrogen and phosphorus removal by high latitude mat-forming cyanobacteria for potential use in tertiary wastewater treatment. J. Appl. Phycol. 2000, 12, 105–112. [Google Scholar] [CrossRef]
- Lynch, F.; et al. Screening native isolates of cyanobacteria and a green alga for integrated wastewater treatment, biomass accumulation and neutral lipid production. Algal Res. 2015, 11, 411–420. [Google Scholar] [CrossRef]
- Kabariya, J.H.; Ramani, V.M. Dairy wastewater treatment by cyanobacteria for removal of nutrients with extraction of high value compounds from biomass. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1527–1538. [Google Scholar] [CrossRef]
- Mukherjee, C.; et al. Parboiled rice effluent: A wastewater niche for microalgae and cyanobacteria with growth coupled to comprehensive remediation and phosphorus biofertilization. Algal Res. 2016, 19, 225–236. [Google Scholar] [CrossRef]
- Del Valle-Real, M.; et al. Wastewater from maize lime-cooking as growth media for alkaliphilic microalgae–cyanobacteria consortium to reduce chemical oxygen demand and produce biomass with high protein content. Int. J. Food Sci. Technol. 2023, 58, 6775–6783. [Google Scholar] [CrossRef]
- Nagasathya, A.; Thajuddin, N. Decolourization of paper mill effluent using hypersaline cyanobacterium. 2008.
- Blaby-Haas, C.E. Cyanobacteria provide a new paradigm in the regulation of cofactor dependence. Proceedings of the National Academy of Sciences. 2021, 118, e2100281118. [Google Scholar] [CrossRef]
- Kong, L. Copper requirement and acquisition by marine microalgae. Microorganisms. 2022, 10, 1853. [Google Scholar] [CrossRef]
- Boden, J.S.; et al. Timing the evolution of antioxidant enzymes in cyanobacteria. Nature Communications. 2021, 12, 4742. [Google Scholar] [CrossRef] [PubMed]
- Kalita, N.; Baruah, P.P. Copper removal efficacy and stress tolerance potential of Leptolyngbya sp. GUEco1015. Heliyon. 2024, 10, 8. [Google Scholar] [CrossRef]
- National Research Council et al. Copper in drinking water. 2000.
- Castielli, O.; et al. Proteomic analyses of the response of cyanobacteria to different stress conditions. FEBS letters. 2009, 583, 1753–1758. [Google Scholar] [CrossRef]
- García-Cañas, R.; et al. A protease-mediated mechanism regulates the cytochrome c 6/plastocyanin switch in Synechocystis sp. PCC 6803. Proceedings of the National Academy of Sciences. 2021, 118, e2017898118. [Google Scholar] [CrossRef]
- Bellemare, D.R.; et al. A novel copper-regulated promoter system for expression of heterologous proteins in Schizosaccharomyces pombe. Gene. 2001, 273, 191–198. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ishihama, A. Characterization of copper-inducible promoters regulated by CpxA/CpxR in Escherichia coli. Bioscience, biotechnology, and biochemistry. 2006, 70, 1688–1695. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R.; et al. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Waterbury, J.B.; Stanier, R.Y. Isolation and growth of cyanobacteria from marine and hypersaline environments. The prokaryotes: a handbook on habitats, isolation, and identification of bacteria. Berlin, Heidelberg: Springer Berlin Heidelberg. 1981, 221-223.
- Kotai, J. Instruction of preparations of modified nutrient medium Z8 for algae. Norw. Inst. Water Res. Blind. Oslo. 1972, 32–39. [Google Scholar]
- Zarrouk, C. Contribution a l'etude d'une Cyanophycee. Influence de Divers Facteurs Physiques et Chimiques sur la croissance et la photosynthese de Spirulina mixima. Thesis. University of Paris, France. 1966.
- Nongrum, N.A.; Syiem, M.B. Effects of copper ion (Cu2+) on the physiological and biochemical activities of the cyanobacterium Nostoc ANTH. Environmental Engineering Research. 2012, 17, 63–67. [Google Scholar] [CrossRef]
- Ahad, R.I.A.; Syiem, M.B. Copper and cadmium-induced toxicity on the cyanobacterium Nostoc muscorum Meg 1: a comparative study. EurAsian Journal of BioSciences. 2018, 12, 333–345. [Google Scholar]
- Wu, Z.; et al. Response of Microcystis to copper stress–do phenotypes of Microcystis make a difference in stress tolerance? Environmental Pollution. 2007, 147, 324–330. [Google Scholar] [CrossRef]
- Wang, H.; Ebenezer, V.; Ki, J. Photosynthetic and biochemical responses of the freshwater green algae Closterium ehrenbergii Meneghini (Conjugatophyceae) exposed to the metal coppers and its implication for toxicity testing. Journal of Microbiology. 2018, 56, 426–434. [Google Scholar] [CrossRef]
- Mohy El Din, S. Effect of copper and lead on growth and some metabolic activities of cyanobacterium Spirulina platensis (Nordstedt). Egyptian Journal of Botany. 2017, 57, 445–456. [Google Scholar] [CrossRef]
- Giner-Lamia, J.; López-Maury, L.; Florencio, F.J. Global transcriptional profiles of the copper responses in the cyanobacterium Synechocystis sp. PCC 6803. PLoS One. 2014, 9, e108912. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, R.; Kenney, G.E.; Rosenzweig, A.C. Dual pathways for copper uptake by methanotrophic bacteria. Journal of biological chemistry. 2011, 286, 37313–37319. [Google Scholar] [CrossRef] [PubMed]
- Andrei, A.; et al. Cu homeostasis in bacteria: the ins and outs. Membranes. 2020, 10, 9–242. [Google Scholar] [CrossRef]
- Kowata, H.; et al. Outer membrane permeability of cyanobacterium Synechocystis sp. strain PCC 6803: studies of passive diffusion of small organic nutrients reveal the absence of classical porins and intrinsically low permeability. Journal of bacteriology. 2017, 199. [Google Scholar] [CrossRef]
- Kojima, S.; Muramoto, K.; Kusano, T. Outer membrane proteins derived from non-cyanobacterial lineage cover the peptidoglycan of Cyanophora paradoxa cyanelles and serve as a cyanelle diffusion channel. Journal of Biological Chemistry. 2016, 291, 20198–20209. [Google Scholar] [CrossRef]
- Wylie, J.L.; Worobec, E.A. The OprB porin plays a central role in carbohydrate uptake in Pseudomonas aeruginosa. Journal of bacteriology. 1995, 177, 3021–3026. [Google Scholar] [CrossRef] [PubMed]
- Schätzle, H.; et al. Comparative phenotypic analysis of Anabaena sp. PCC 7120 mutants of porinlike genes. Journal of Microbiology and Biotechnology. 2021, 31, 645. [Google Scholar] [CrossRef]
- Cardoso, D.; et al. The role of outer membrane protein (s) harboring SLH/OprB-domains in extracellular vesicles’ production in Synechocystis sp. PCC 6803. Plants. 2021, 10, 2757. [Google Scholar] [CrossRef]
- Coines, J.; et al. Glucose transport via the pseudomonad porin OprB: Implications for the design of Trojan Horse anti-infectives. Physical Chemistry Chemical Physics. 2019, 21, 8457–8463. [Google Scholar] [CrossRef]
- Pederick, V.G.; et al. ZnuA and zinc homeostasis in Pseudomonas aeruginosa. Scientific reports. 2015, 5, 13139. [Google Scholar] [CrossRef]
- Cavet, J.S.; Borrelly, G.P.M.; Robinson, N.J. Zn, Cu and Co in cyanobacteria: selective control of metal availability. FEMS Microbiology Reviews. 2003, 27, 165–181. [Google Scholar] [CrossRef]
- Banerjee, S.; et al. Structural determinants of metal specificity in the zinc transport protein ZnuA from Synechocystis 6803. Journal of molecular biology. 2003, 333, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Yatsunyk, L.A.; et al. Structure and metal binding properties of ZnuA, a periplasmic zinc transporter from Escherichia coli. JBIC Journal of Biological Inorganic Chemistry. 2008, 13, 271–288. [Google Scholar] [CrossRef] [PubMed]
- Patzer, S.I.; Hantke, K. The ZnuABC high-affinity zinc uptake system and its regulator Zur in Escherichia coli. Molecular microbiology. 1998, 28, 1199–1210. [Google Scholar] [CrossRef] [PubMed]
- Hudek, L.; et al. Molecular and cellular characterisation of the zinc uptake (Znu) system of Nostoc punctiforme. FEMS microbiology ecology. 2013, 86, 149–171. [Google Scholar] [CrossRef]
- Bartsevich, V.V.; Pakrasi, H.B. Manganese transport in the cyanobacterium Synechocystis sp. PCC 6803. Journal of Biological Chemistry. 1996, 271, 26057–26061. [Google Scholar] [CrossRef]
- Al-tameemi, H.; et al. Staphylococcus aureus lacking a functional MntABC manganese import system has increased resistance to copper. Molecular microbiology. 2021, 115, 554–573. [Google Scholar] [CrossRef]
- Lim, K. HL.; et al. Metal binding specificity of the MntABC permease of Neisseria gonorrhoeae and its influence on bacterial growth and interaction with cervical epithelial cells. Infection and immunity. 2008, 76, 3569–3576. [Google Scholar] [CrossRef]
- Giner-Lamia, J.; et al. The CopRS two-component system is responsible for resistance to copper in the cyanobacterium Synechocystis sp. PCC 6803. Plant physiology. 2012, 159, 1806–1818. [Google Scholar] [CrossRef]
- Rosenzweig, A.C.; Argüello, J.M. Toward a molecular understanding of metal transport by P1B-Type ATPases. In: Current topics in membranes. Academic Press. 2012, 113-136.
- Williams, L.E.; Mills, R.F. P1B-ATPases–an ancient family of transition metal pumps with diverse functions in plants. Trends in plant science. 2005, 10, 491–502. [Google Scholar] [CrossRef]
- Salustros, Nina et al. Structural basis of ion uptake in copper-transporting P1B-type ATPases. Nature Communications. 2022, 13, 5121.
- Hederstedt, L.; Lewin, A.; Throne-Holst, M. . Heme A synthase enzyme functions dissected by mutagenesis of Bacillus subtilis CtaA. Journal of bacteriology. 2005, 187, 8361–8369. [Google Scholar] [CrossRef] [PubMed]
- Raimunda, D.; González-Guerrero, M.; Leeber, B., III; Argüello, J. The transport mechanism of bacterial Cu+-ATPases: Distinct efflux rates adapted to different function. BioMetals. 2011, 24, 467–475. [Google Scholar] [CrossRef]
- Phung, L.T.; Ajlani, G.; Haselkorn, R. P-type ATPase from the cyanobacterium Synechococcus 7942 related to the human Menkes and Wilson disease gene products. Proceedings of the National Academy of Sciences. 1994, 91, 9651–9654. [Google Scholar] [CrossRef]
- Yuan, D.S.; Dancis, A.; Klausner, R.D. Restriction of copper export in Saccharomyces cerevisiae to a late Golgi or post-Golgi compartment in the secretory pathway. Journal of Biological Chemistry. 1997, 272, 25787–25793. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Z.-H.; et al. Reciprocal effect of copper and iron regulation on the proteome of Synechocystis sp. PCC 6803. Frontiers in Bioengineering and Biotechnology. 2021, 9, 673402. [Google Scholar] [CrossRef]
- Katoh, H.; Hagino, N.; Ogawa, T. Iron-binding activity of FutA1 subunit of an ABC-type iron transporter in the cyanobacterium Synechocystis sp. strain PCC 6803. Plant and Cell Physiology. 2001, 42, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Waldron, K.J.; et al. A periplasmic iron-binding protein contributes toward inward copper supply. Journal of Biological Chemistry. 2007, 282, 3837–3846. [Google Scholar] [CrossRef]
- Stevanovic, M.; et al. The components of the putative iron transport system in the cyanobacterium Anabaena sp. PCC 7120. Environmental Microbiology. 2012, 14, 1655–1670. [Google Scholar] [CrossRef] [PubMed]
- Aliaga, M.E.; et al. Redox-implications associated with the formation of complexes between copper ions and reduced or oxidized glutathione. Journal of Inorganic Biochemistry. 2016, 154, 78–88. [Google Scholar] [CrossRef]
- Rakshit, A.; et al. Cu²⁺ selective chelators relieve copper-induced oxidative stress in vivo. Chemical Science. 2018, 9, 7916–7930. [Google Scholar] [CrossRef]
- Markossian, K.A.; Kurganov, B.I. Copper chaperones, intracellular copper trafficking proteins. Function, structure, and mechanism of action. Biochemistry (Moscow). 2003, 68, 827–837. [Google Scholar] [CrossRef]
- Arnesano, F.; et al. Solution structure of the Cu(I) and apo forms of the yeast metallochaperone, Atx1. Biochemistry. 2001, 40, 1528–1539. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; et al. Transition metal transporting P-type ATPases: terminal metal-binding domains serve as sensors for autoinhibitory tails. The FEBS Journal. 2024. [Google Scholar] [CrossRef] [PubMed]
- Gounder, P.E.; et al. Metallothioneins: Diverse Protein Family to Bind Metallic Ions. Chapters. 2021. [Google Scholar]
- Bourdieunaud, J.-P.; et al. Challenging the model for induction of metallothionein gene expression. Biochimie. 2006, 88, 1787–1792. [Google Scholar] [CrossRef] [PubMed]
- Dziegiel, P.; et al. Metallothioneins: structure and functions. Metallothioneins in Normal and Cancer Cells. 2016, 3, 20. [Google Scholar]
- Robinson, N.J.; Wilson, J.R.; Turner, J.S. Expression of the type 2 metallothionein-like gene MT2 from Arabidopsis thaliana in Zn²⁺-metallothionein-deficient Synechococcus PCC 7942: putative role for MT2 in Zn²⁺ metabolism. Plant Molecular Biology. 1996, 30, 1169–1179. [Google Scholar] [CrossRef]
- Verma, S.K.; Singh, S.P. Factors regulating copper uptake in a cyanobacterium. Current Microbiology. 1990, 21, 33–37. [Google Scholar] [CrossRef]
- Munson, G.P.; et al. Identification of a copper-responsive two-component system on the chromosome of Escherichia coli K-12. Journal of Bacteriology. 2000, 182, 5864–5871. [Google Scholar] [CrossRef]
- Rouch, D.A.; Brown, N.L. Copper-inducible transcriptional regulation at two promoters in the Escherichia coli copper resistance determinant pco. Microbiology. 1997, 143, 1191–1202. [Google Scholar] [CrossRef]
- Giner-Lamia, J.; et al. Extracellular proteins: novel key components of metal resistance in cyanobacteria? Frontiers in Microbiology. 2016, 7, 878. [Google Scholar] [CrossRef] [PubMed]
- Gittins, J.R. Cloning of a copper resistance gene cluster from the cyanobacterium Synechocystis sp. PCC 6803 by recombineering recovery. FEBS Letters. 2015, 589, 1872–1878. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; et al. Structural basis for copper/silver binding by the Synechocystis metallochaperone CopM. Biological Crystallography. 2016, 72, 997–1005. [Google Scholar]
- Yang, J.; et al. Structural basis of copper binding by a dimeric periplasmic protein forming a six-helical bundle. Journal of Inorganic Biochemistry. 2022, 229, 111728. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.; et al. Extracellular vesicles as an alternative copper-secretion mechanism in bacteria. Journal of Hazardous Materials. 2022, 431, 128594. [Google Scholar] [CrossRef]
- Lawton, T.J.; et al. The CopC family: structural and bioinformatic insights into a diverse group of periplasmic copper binding proteins. Biochemistry. 2016, 55, 2278–2290. [Google Scholar] [CrossRef]
- Gautam, P.; Erill, I.; Cusick, K.D. Linking Copper-Associated Signal Transduction Systems with Their Environment in Marine Bacteria. Microorganisms. 2023, 11, 1012. [Google Scholar] [CrossRef]
- Rensing, C.; et al. CopA: an Escherichia coli Cu(I)-translocating P-type ATPase. Proceedings of the National Academy of Sciences. 2000, 97, 652–656. [Google Scholar] [CrossRef]
- Gómez-Santos, N.; et al. CorE from Myxococcus xanthus is a copper-dependent RNA polymerase sigma factor. PLoS Genetics. 2011, 7, e1002106. [Google Scholar] [CrossRef]
- Cha, J.-S.; Cooksey, D.A. Copper resistance in Pseudomonas syringae mediated by periplasmic and outer membrane proteins. Proceedings of the National Academy of Sciences. 1991, 88, 8915–8919. [Google Scholar] [CrossRef]
- Cha, J.-S.; Cooksey, D.A. Copper hypersensitivity and uptake in Pseudomonas syringae containing cloned components of the copper resistance operon. Applied and Environmental Microbiology. 1993, 59, 1671–1674. [Google Scholar] [CrossRef] [PubMed]
- Olivan-Muro, I.; et al. Unbalancing Zur (FurB)-mediated homeostasis in Anabaena sp. PCC7120: Consequences on metal trafficking, heterocyst development and biofilm formation. Environmental Microbiology. 2023, 25, 2142–2162. [Google Scholar] [CrossRef] [PubMed]
- Behlau, F.; et al. Molecular characterization of copper resistance genes from Xanthomonas citri subsp. citri and Xanthomonas alfalfae subsp. citrumelonis. Applied and Environmental Microbiology. 2011, 77, 4089–4096. [Google Scholar] [PubMed]
- Mills, S.D.; Jasalavich, C.A.; Cooksey, D.A. A two-component regulatory system required for copper-inducible expression of the copper resistance operon of Pseudomonas syringae. Journal of Bacteriology. 1993, 175, 1656–1664. [Google Scholar] [CrossRef]
- Franke, S.; et al. Molecular analysis of the copper-transporting efflux system CusCFBA of Escherichia coli. Journal of Bacteriology. 2003, 185, 3804–3812. [Google Scholar] [CrossRef]
- Su, C.-C.; et al. Crystal structure of the CusBA heavy-metal efflux complex of Escherichia coli. Nature. 2011, 470, 558–562. [Google Scholar] [CrossRef]
- Hahn, A.; et al. The outer membrane TolC-like channel HgdD is part of tripartite resistance-nodulation-cell division (RND) efflux systems conferring multiple-drug resistance in the cyanobacterium Anabaena sp. PCC7120. Journal of Biological Chemistry. 2013, 288, 31192–31205. [Google Scholar] [CrossRef]
- Nicolaisen, K.; et al. The interplay between siderophore secretion and coupled iron and copper transport in the heterocyst-forming cyanobacterium Anabaena sp. PCC 7120. Biochimica et Biophysica Acta (BBA)-Biomembranes. 2010, 1798, 2131–2140. [Google Scholar] [CrossRef]
- Cruz, D.; et al. Exopolysaccharides from cyanobacteria: strategies for bioprocess development. Applied Sciences. 2020, 10, 3763. [Google Scholar] [CrossRef]
- Tiwari, O.N.; et al. Biosynthesis, purification and structure-property relationships of new cyanobacterial exopolysaccharides. Polymer Testing. 2020, 89, 106592. [Google Scholar] [CrossRef]
- De Philippis, R.; Vincenzini, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiology Reviews. 1998, 22, 151–175. [Google Scholar] [CrossRef]
- Nogueira, P.F.M.; et al. The effects of Anabaena spiroides exopolysaccharides on copper accumulation in an aquatic food chain. Aquatic Toxicology. 2009, 93, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Kuang, X.; et al. Enhancement mechanisms of copper(II) adsorption onto kaolinite by extracellular polymeric substances of Microcystis aeruginosa (cyanobacterium). International Biodeterioration & Biodegradation. 2019, 138, 8–14. [Google Scholar]
- Paperi, R.; Micheletti, E.; De Philippis, R. Optimization of copper sorbing–desorbing cycles with confined cultures of the exopolysaccharide-producing cyanobacterium Cyanospira capsulata. Journal of Applied Microbiology. 2006, 101, 1351–1356. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; et al. Semi-continuous cultivation of EPS-producing marine cyanobacteria: A green biotechnology to remove dissolved metals obtaining metal-organic materials. New Biotechnology. 2024, 82, 33–42. [Google Scholar] [CrossRef]
- Pereira, S.; et al. Complexity of cyanobacterial exopolysaccharides: composition, structures, inducing factors and putative genes involved in their biosynthesis and assembly. FEMS Microbiology Reviews. 2009, 33, 917–941. [Google Scholar] [CrossRef]
- Pagli, C.; et al. Isolation of biocrust cyanobacteria and evaluation of Cu, Pb, and Zn immobilisation potential for soil restoration and sustainable agriculture. Science of the Total Environment. 2024, 946, 174020. [Google Scholar] [CrossRef]
- Gupta, P.; Diwan, B. Bacterial exopolysaccharide mediated heavy metal removal: a review on biosynthesis, mechanism and remediation strategies. Biotechnology Reports. 2017, 13, 58–71. [Google Scholar] [CrossRef]
- Singh, N.; et al. Thiol and exopolysaccharide production in a cyanobacterium under heavy metal stress. Process Biochemistry. 1999, 35, 63–68. [Google Scholar] [CrossRef]
- De Philippis, R.; Paperi, R.; Sili, C. Heavy metal sorption by released polysaccharides and whole cultures of two exopolysaccharide-producing cyanobacteria. Biodegradation. 2007, 18, 181–187. [Google Scholar] [CrossRef]
- Leal, P.P.; et al. Copper pollution exacerbates the effects of ocean acidification and warming on kelp microscopic early life stages. Scientific Reports. 2018, 8, 14763. [Google Scholar] [CrossRef] [PubMed]
- Comte, S.; Guibaud, G.; Baudu, M. Biosorption properties of extracellular polymeric substances (EPS) towards Cd, Cu and Pb for different pH values. Journal of Hazardous Materials. 2008, 151, 185–193. [Google Scholar] [CrossRef] [PubMed]
Figure 3.
3D model of the CopA protein in Cyanobacteria and other groups of prokaryotes constructed using ChimeraX. A. Escherichia coli O157:H7 (model obtained from Swiss-model; ~85.9 kDa); B. Vibrio cholerae serotype O1 strain ATCC 39315 (model obtained from Swiss-model; ~96.4 KDa); C. Hydrococcus rivularis NIES-593 (model obtained from Swiss-model; ~80.2 kDa); D. Gloeobacter violaceus (model obtained from Swiss-model; ~77.2 kDa); E. Acidovorax sp. 99 (model obtained from Swiss-model; ~86.3 kDa); F. Staphylococcus aureus strain JH9 (model obtained from Swiss-model; ~86.7 kDa); G. Nodularia spumigena UHCC0039 (model built using AlphaFold2 based on the sequence used for phylogeny; ~82.8 kDa); H. Nostoc muscorum FACHB395 (model built using AlphaFold2 based on the sequence used for phylogeny; ~83.0 kDa); I. Superposition of the proteins from a different perspective. *The green markings highlight the Heavy-Metal-Associated domain of the proteins. The rest of the protein includes the P-type ATPase Cu-like domain.
Figure 3.
3D model of the CopA protein in Cyanobacteria and other groups of prokaryotes constructed using ChimeraX. A. Escherichia coli O157:H7 (model obtained from Swiss-model; ~85.9 kDa); B. Vibrio cholerae serotype O1 strain ATCC 39315 (model obtained from Swiss-model; ~96.4 KDa); C. Hydrococcus rivularis NIES-593 (model obtained from Swiss-model; ~80.2 kDa); D. Gloeobacter violaceus (model obtained from Swiss-model; ~77.2 kDa); E. Acidovorax sp. 99 (model obtained from Swiss-model; ~86.3 kDa); F. Staphylococcus aureus strain JH9 (model obtained from Swiss-model; ~86.7 kDa); G. Nodularia spumigena UHCC0039 (model built using AlphaFold2 based on the sequence used for phylogeny; ~82.8 kDa); H. Nostoc muscorum FACHB395 (model built using AlphaFold2 based on the sequence used for phylogeny; ~83.0 kDa); I. Superposition of the proteins from a different perspective. *The green markings highlight the Heavy-Metal-Associated domain of the proteins. The rest of the protein includes the P-type ATPase Cu-like domain.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



