Submitted:
02 June 2025
Posted:
02 June 2025
You are already at the latest version
Abstract
Cancer immunotherapy has emerged as a revolutionary approach that harnesses the immune system to combat malignancies. While compelling evidence demonstrates that gut microbiota critically influences treatment efficacy through tumor microenvironment modulation, the role of vitamin D (VD) remains controversial yet promising. VD exerts dual immunomodulatory effects: it activates immune cells via vitamin D receptor (VDR) signaling while simultaneously shaping gut microbial composition to enhance antitumor immunity potentially. Clinical data present conflicting results - optimal VD levels correlate with improved immunotherapy response and reduced toxicity in some studies, yet paradoxically associate with immunosuppression and even poorer survival outcomes in others. While VD generally suppresses pro-inflammatory Th17/IL-17 pathways, emerging evidence reveals context-dependent Th17 activation that may promote tumor metastasis in specific populations. These contradictory findings underscore the urgent need to clarify VD's complex dose-dependent and microbiome-mediated immunoregulatory mechanisms. Future investigations should prioritize resolving these controversies to establish precise clinical applications of VD in immunotherapy regimens.
Keywords:
Vitamin D
; Gut microbiota
; Cancer immunotherapy
; Crosstalk
Introduction
Cancer immunotherapy represents a breakthrough treatment modality focused on activating or strengthening the host immune system to recognize and destroy malignant cells. Significant advances in this field have reinvigorated the therapeutic landscape for cancer patients [1,2]. However, immunotherapy efficacy varies substantially among patients, characterized by low response rates and significant side effects. Consequently, improving immunotherapy response rates through endogenous immune regulation mechanisms has become a critical direction in tumor immunology research. Within this context, vitamin D (VD), a hormone with both nutritional and immunomodulatory effects, has gradually demonstrated potential value.
VD is well-established for its classical functions in calcium homeostasis and bone metabolism and its pleiotropic effects, including cardiovascular protection, neuroregulation, and immunomodulation [3,4]. Beyond these roles, emerging evidence highlights VD as a potent immunomodulator with promising applications in cancer immunotherapy. The biological effects of VD are mediated through its binding to vitamin D receptors, which are widely expressed on various immune cells. This interaction modulates immune cell differentiation, functional activation, and cytokine secretion profiles [5,6]. Notably, the immunomodulatory effects of VD are also achieved through its impact on the gut microbiota. VD exhibits prebiotic-like properties by selectively enriching beneficial bacterial populations, thereby becoming a key regulator of gut microbial structure [7]. The gut microbiota itself plays a key role in regulating anti-tumor immune responses.
In recent years, the "gut-immuno-tumor" research framework has gradually taken shape. Multiple studies have confirmed associations between the abundance and diversity of specific microbiota and immunotherapy efficacy [8,9,10,11]. Growing evidence indicates that microbial diversity and taxonomic characteristics are linked to general health and significantly regulate tumorigenesis, progression, and treatment efficacy [12]. This microbiota-mediated immunomodulation may synergize with immune checkpoint inhibitor therapies, creating a favorable microenvironment for enhancing anti-tumor immunity [13,14].
In conclusion, although VD and intestinal microbiota independently regulate immune responses, the linkage between VD and its synergistic effect on cancer immunotherapy requires systematic exploration, particularly because VD also influences intestinal microbiota changes. Current studies generally suggest that VD can potentially promote tumor immunotherapy by regulating intestinal microbiota and immune function; however, its effect is inconsistent. Excessive VD supplementation can trigger immunosuppression, interfering with anti-tumor immune responses [15]. Concurrently, while substantial evidence supports vitamin D receptor (VDR) inhibiting the Th17/IL-17 pathway, reducing inflammation, and delaying tumor progression, it may also activate the Th17 response and promote tumor metastasis in specific populations or tumor types [16]. This contradiction indicates that VD's immunomodulatory effect involves complex dose-dependence and individual variation; its interaction with microbiota and immunotherapy still necessitates systematic study.
Therefore, this review aims to systematically explore the interaction between VD, gut microbiota, and cancer immunotherapy efficacy, analyze their potential mechanisms and influencing factors, clarify current research consensus and controversy, and evaluate the prospects and challenges of VD as an adjunct strategy for cancer immunotherapy.
VD Metabolism and Function
VD deficiency is a major public health problem for all ages worldwide, even in countries with perennial sun exposure [17,18,19]. The main source of VD in humans (90%) is the transformation of 7-dehydrocholesterol, which is developed in the skin after UVB radiation from the sun, into preVD. Only 10% of VD is obtained through dietary intake. VD is a fat-soluble vitamin in two forms, VD2 (ergocalciferol) and VD3 (cholecalciferol), both available through the diet. Vitamins D2 and D3 are transformed into 25-hydroxyVD (25(OH)D) by 25-hydroxylase enzymes such as CYP27A1 and CYP2R1 in the liver, which is the main circulating form of VD. Subsequently it is hydroxylated to 1,25 dihydroxyVD (1,25(OH)2D) in the kidney by the 25(OH)D-1α-hydroxylase CYP27B1 [3,20].
1,25(OH)2D served as the primary ligand of VDR. The VDR is a nuclear hormone receptor and transcription factor in virtually all cell types. The VDR forms a heterodimer within the nucleus with the retinoid X receptor (RXR). This enables the VDR/RXR complex to bind to VD-responsive elements (VDREs) in target genes and regulate their transcription. As a result, VD modulates numerous cellular processes, with one of its most significant effects being the regulation of calcium absorption in the intestine. Currently, 11,031 potential VDR target genes have been identified [21]. Among these, 43% are associated with metabolic processes, 19% with cell and tissue morphology, 10% with cell junctions and adhesion, another 10% with differentiation and development, 9% with angiogenesis, and 5% with epithelial-to-mesenchymal transition. Furthermore, the VDR governs various microRNAs (miRNAs) and long non-coding RNAs linked to directly or indirectly expressing a broad spectrum of proteins. These insights collectively highlight VD's vital role in numerous biological processes [20,22,23,24,25] (Figure 1).
As such, VD is vital for maintaining calcium and phosphate homeostasis and bone mineralization and plays an increasingly recognized role in metabolic regulation, immune modulation, and cellular differentiation [26,27]. For instance, VD exhibits significant immunomodulatory effects by enhancing the function of immune organs such as the spleen and lymph nodes, as well as boosting T-cell activity, thereby strengthening the body's antitumor immune defenses. VD exerts these effects through VDR expressed on immune cells, modulating genes related to cell proliferation and suppressing tumor cell survival, proliferation, migration, and metastasis [28]. In terms of inflammation regulation, clinical studies have confirmed that regular VD supplementation significantly reduces disease activity in patients with relapsing-remitting multiple sclerosis [29] and lowers the risk of cancer-associated inflammation, such as that linked to colorectal cancer [28]. Additionally, VD provides cardiovascular protection [30], with supplementation shown to reduce the risk of atherosclerotic cardiovascular diseases, particularly ischemic heart disease, suggesting it may serve as an effective preventive strategy in populations with VD deficiency [31,32]. Research also indicates that higher serum 25-hydroxyVD levels are significantly associated with a lower risk of venous thromboembolism, with this protective effect being more pronounced in individuals with diabetes [25]. VD also plays a vital protective role during pregnancy and fetal development [27,33].
Given these broad systemic functions, recent research has turned to exploring how VD status contributes to more complex physiological and pathological processes, particularly in the fields of cancer immunotherapy and host-microbiota interactions. The following sections will discuss these emerging domains of VD biology.
Effects of VD-Cancer Immunotherapy
Cancer remains a formidable global public health challenge in contemporary medicine[34,35]. Modern oncological interventions typically employ a multimodal approach, integrating surgical resection, chemotherapy, radiotherapy, hormone therapy, immunotherapy, and hematopoietic stem cell transplantation to combat malignant progression. Immunotherapy is one of the oncological treatments by harnessing the immune system to selectively target tumor cells [36,37,38,39]. However, treatment responses are often heterogeneous. Considering its established roles in calcium-phosphate metabolism and systemic homeostasis, VD has emerged as a key immunomodulatory agent. In this context, VD has attracted attention for its potential to both enhance antitumor immunity and mitigate immunotherapy-associated toxicity [40].
The association between VD and cancer presents complex and varied findings. While multiple observational studies have found that breast cancer patients generally exhibit lower serum 25(OH)D levels compared to healthy controls, with more severe VD deficiency observed in advanced-stage patients [41,42,43] , conclusions across studies differ significantly. Recent research has unveiled the population-specific nature of VD's anticancer effects: a meta-analysis showed that higher serum 25(OH)D levels demonstrated significant protective effects only in premenopausal women or at the time of diagnosis [44]. Conversely, a cohort study focusing on elderly European populations reached the opposite conclusion, finding that breast cancer risk increased with higher 25(OH)D concentrations [45]. More strikingly, research by Kanstrup et al. indicated that female breast cancer patients with excessively high serum 25(OH)D levels (exceeding 110 nmol/L) exhibited poorer survival outcomes [15]. At the mechanistic level, most evidence supports VD's role in inhibiting Th17 cell differentiation and IL-17 production via the VDR signaling pathway, thereby mitigating inflammatory responses and suppressing tumor progression [16]. However, under certain conditions (such as in younger individuals or high-estrogen environments), VD may paradoxically promote Th17 cell activation and tumor metastasis risk by upregulating osteopontin (OPN) or activating estrogen receptor pathways, among other mechanisms [16]. These conflicting findings suggest that VD's role in cancer development and progression may be influenced by various factors, including age, hormone levels, and genetic background. VD's precise mechanisms and clinical applications in oncology warrant further in-depth research.
Cancer immunotherapy represents a paradigm shift in oncology, fundamentally transforming cancer treatment strategies. This innovative approach harnesses and enhances the host's immune system to target and eliminate malignant cells specifically, establishing itself as a cornerstone in contemporary cancer management [1]. Conventional immunotherapeutic strategies primarily focus on immune cell activation and immune response potentiation through several mechanisms: immune checkpoint blockade targeting CTLA-4 and PD-1/PD-L1 pathways to counteract immune evasion; tumor vaccines for immune system priming; adoptive cell therapy involving ex vivo immune cell modification and expansion; and monoclonal antibody-mediated specific antigen targetin [2,39,46,47,48]. The therapeutic advantages of immunotherapy are substantial, characterized by its exceptional specificity and minimal off-target effects on normal tissues. Clinically, this modality has demonstrated remarkable efficacy, with some patients achieving durable remission or complete eradication of malignancies [49]. Furthermore, the immunological memory conferred by this approach provides sustained protection against tumor recurrence, representing a significant advancement in cancer therapeutics [50].
VD demonstrates synergistic potential in cancer immunotherapy, primarily mediated through its interaction with the VD receptor (VDR). It is broadly expressed across immune cell lineages, including T lymphocytes, dendritic cells, and macrophages [51,52]. Studies reveal that activation of the VDR signaling pathway enhances anti-tumor immune responses via dual mechanisms: it promotes the differentiation, maturation, and functional optimization of regulatory T cells (Tregs), while concurrently reducing immunosuppressive factor levels within the tumor microenvironment [35,53]. Notably, VD upregulates the expression of major histocompatibility complex (MHC) molecules, thereby significantly enhancing the immune system's recognition capacity [54,55,56]. Additionally, the crosstalk regulatory network between VD and key signaling pathways such as PPARγ, PI3K/AKT/mTOR can dynamically modulate the expression of immune checkpoint molecules like PD-L1, providing new targets for combination therapies. Emerging evidence also reveals the synergistic tumor-suppressive effects between VD signaling and the estrogen receptor (ESR) pathway. These findings suggest that VD supplementation, through multi-pathway synergistic effects, may serve as an ideal adjuvant to enhance the efficacy of immune checkpoint blockade therapies [6].
The tumor microenvironment's inflammatory milieu constitutes a pivotal driver of oncogenesis, facilitating neoplastic progression through sustained tumor cell proliferation, angiogenic induction, and metastatic dissemination [57] . In this pathological context, VD exerts multimodal anti-inflammatory effects, notably suppressing IL-6 and TNF-α production to disrupt the self-perpetuating cycle of inflammation-mediated tumorigenesis [58,59,60]. These immunomodulatory mechanisms collectively position VD as a potential adjuvant capable of recalibrating immune homeostasis to potentiate conventional immunotherapies.
In immunotherapy, particularly immune checkpoint inhibitors (ICIs) such as anti-PD-1/PD-L1 and anti-CTLA-4 therapies4. Studies have shown that maintaining VD levels within the normal range during anti-PD-1 immunotherapy is necessary to ensure treatment efficacy in patients with advanced melanoma [61,62]. Additional research has indicated that, for melanoma patients receiving PD-1, CTLA-4, or combined immune checkpoint inhibitors (ICIs), the administration of VD significantly decreases the likelihood of developing ICI-induced colitis [4]. The PROVIDENCE study highlights that early implementation of systematic VD supplementation may exert beneficial effects on clinical outcomes in advanced cancer patients undergoing immune checkpoint inhibitor (ICI) therapy, while also demonstrating that maintaining optimal VD sufficiency could serve as a preventive strategy against thyroid-related immune-related adverse events (irAEs) [63]. Clinical observations have also suggested that higher serum levels of VD are associated with improved responses to immunotherapy and better overall survival in cancer patients [4,64,65].
However, the relationship between VD (VD) and cancer immunotherapy is complex and context-dependent (Figure 4). While VD demonstrates immunomodulatory benefits that may enhance immune checkpoint blockade therapy, emerging evidence suggests it may also exert immunosuppressive effects on dendritic cells (DCs) and B cell function, which could potentially limit its therapeutic efficacy in certain contexts [15,66]. The current literature presents conflicting findings regarding VD's role in cancer immunotherapy, with variations in study outcomes potentially attributable to differences in dosing regimens, patient characteristics [27], and cancer types [64]. Further research is needed to establish optimal dosing protocols, determine the most effective timing for VD administration relative to treatment cycles, elucidate its precise mechanisms of action, and identify patient subgroups that may derive the most significant clinical benefit from VD supplementation in combination with immunotherapy.
In conclusion, VD constitutes a promising adjunctive therapy in cancer immunotherapy, demonstrating potential to bolster immune responses and enhance therapeutic efficacy. However, its complex and context-dependent effects require further clarification about its interplay with the gut microbiome, an emerging axis of immunoregulation in cancer.
VD Interacts with Gut Microbiota
Based on its role in modulating the immune system, VD also profoundly affects the gut microbiota. The interaction between VD and the gut microbiota may further illuminate VD's contribution to cancer immunotherapy efficacy.
The gut microbiota, a complex and dynamic ecosystem residing within the host's gastrointestinal tract, comprises various microorganisms, including bacteria, archaea, fungi, viruses, and bacteriophages. This intricate microbial community is pivotal in modulating multiple host physiological functions and is intimately involved in health maintenance and disease pathogenesis [67,68,69,70].
In the realm of nutritional metabolism, the gut microbiota plays an indispensable role in nutrient biotransformation, particularly through the fermentation of indigestible dietary fibers into biologically active short-chain fatty acids (SCFAs) [71], while simultaneously orchestrating lipid metabolism and energy homeostasis. Regarding immune regulation, commensal microorganisms are essential for the maturation and differentiation of immune cells, particularly in gut-associated lymphoid tissues [72]. Furthermore, these microbial communities contribute to maintaining intestinal epithelial barrier integrity and regulating mucosal immune homeostasis through complex host-microbe interactions [73]. Meanwhile, it plays a significant role in digesting food, regulating intestinal endocrine function and neural signaling [74], training host immunity [72], modifying drug action and metabolism [75], as well as detoxifying the body [68]. However, although the gut microbiota plays many key roles, it is complexly influenced by various physiological and environmental factors.
VD influences the gut microbiota apparently (Figure 2) [76,77]. The findings revealed a correlation between VD levels and the composition, diversity, or functionality of the gut microbiota [78,79,80]. In a double-blind randomized controlled trial, VD was found to improve gut microbiota and promote muscle anabolism [81]. The Mediterranean diet was applied to 91 patients with obesity and metabolic syndrome. After one year of dietary intervention, patients with low levels of 25(OH)D exhibited an increase in the diversity of their intestinal microbiota, which influenced their metabolic processes [82]. Additionally, individuals with the highest versus lowest concentrations of 1,25(OH)2D and its activation ratios tend to possess greater abundances of butyrate-producing bacteria, which have been linked to improved gut microbial health [83]. In particular, VD supplements can increase beneficial gut microbiota, including Ruminococcaceae, Akkermansia, Faecalibacterium, and Coccus, thereby modulating autoimmune responses [84]. In mice subjected to VD-deficient diets or genetic knockout models, the abundance of Bacteroidetes (or taxa within this phylum) in the gut microbiota was observed to increase [80]. VD induces the expression of antimicrobial peptides (AMPs) in the zebrafish intestine by activating microbiota-dependent IL-22 signaling. In VD-deficient zebrafish, the abundance of the acetate-producing bacterium Vibrio is reduced. This study demonstrates that VD regulates the composition of the gut microbiota in zebrafish and the production of short-chain fatty acids (SCFAs), thereby enhancing immunity [71]. VD deficiency syndrome can manifest as colonic hyperplasia and epithelial barrier dysfunction during C. rodentium attack [85].
VDR is also closely related to the gut microbiota [77]. The downregulation of VDR and the impaired ability to produce the active form of VD have been correlated with a reduction in Lactobacillus and an increase in Proteobacteria within the gut microbiota [86]. By the way, other research indicates that the induction of Cyp27b1 in mice colonic epithelial cells, which is anticipated to boost local production of 1,25(OH)2D, functions as a protective mechanism. This mechanism partially mitigates the downregulation of epithelial VDR during colonic inflammation. The elevated local levels of 1,25(OH)2D sustain the 1,25(OH)2D-VDR signaling pathway, which safeguards the mucosal barrier and diminishes colonic inflammation [87]. In parallel, fecal and cecal stool samples were collected from VDR knockout (Vdr-/-) and wild-type mice to extract bacterial DNA. Then, samples were subjected to 454 pyrosequencing to determine the bacterial composition present in the stool samples. The findings suggest that VDR status influences the gut microbiota at a taxonomic and functional level and correlates with VDR-associated bacterial changes in clinical disease [88].
Studies have shown that gut microbiota can modulate intestinal VD metabolism [89]. Specifically, the Bifidobacterium longum strain FSHHK13M significantly elevated 1,25-dihydroxy VD and osteocalcin serum, thereby alleviating osteoporosis in mice [90]. In addition, the study found that doubling the genetic liability associated with the abundance of Erysipelotrichia, Erysipelotrichaceae, and Erysipelotrichales reduced the concentration of 25(OH)D [91].
The literature thus suggests a potential pathogenic cascade: VD deficiency triggers gut microbiota imbalance, exacerbating microbial dysbiosis and systemic disease [79]. Furthermore, the gut microbiota reciprocally modulates intestinal VD levels. This bidirectional interplay between VD and gut microbiota may inform clinical strategies to optimize VD supplementation.
Given the central role of the gut microbiota in immune modulation and its tight interplay with VD signaling, it is reasonable to hypothesize that VD-microbiota interactions may significantly influence responses to cancer immunotherapy. The following section explores how gut microbiota composition and function affect the efficacy and safety of cancer immunotherapy, thereby complementing the immunoregulatory role of VD.
Gut Microbiota as a Determinant of Immunotherapy Efficacy
The gut microbiota has emerged as a key modulator of cancer immunotherapy efficacy. Research has demonstrated that gut microbiota profoundly influences the effectiveness of immunotherapy through multiple mechanisms [8], including modulation of immune cell function, production of immunologically active metabolites, maintenance of intestinal barrier integrity, and alteration of the tumor microenvironment (Figure 3). Firstly, concerning immune cell function, studies have demonstrated that secondary bile acids, which are products of primary bile acid metabolism by gut microbiota, enhance the activation and effector functions of T cells while reducing the accumulation and functionality of myeloid-derived suppressor cells (MDSCs) [92,93], thereby modulating immune cell activity. In terms of the production of immunologically active metabolites, gut microbiota such as Akkermansia muciniphila and Bifidobacterium enhance the anti-tumor activity of CD8+ T cells by producing short-chain fatty acids (SCFAs) and tryptophan metabolites, thus improving the efficacy of immunotherapy [13,14]. Analysis of fecal metagenomes from 112 melanoma patients and in vivo experiments in mice revealed that bacterial subpopulations encoding immunostimulatory hexa-acylated lipopolysaccharide (LPS) can enhance the anti-tumor efficacy of anti-PD-1 therapy [94]. Additionally, the gut microbiota supports intestinal barrier integrity, blocking bacterial and toxin entry into the bloodstream to reduce systemic inflammation and indirectly enhance anti-tumor immunity [95]. Finally, research has shown that the gut microbiota can alter the tumor microenvironment; For instance, Bacteroides fragilis enhances the efficacy of anti-CTLA-4 therapy by remodeling the tumor microenvironment through immunomodulatory mechanisms [9].
Many findings highlight the critical role of gut microbiota in modulating host immune responses and shaping clinical outcomes during cancer immunotherapy [10,11]. Emerging evidence positions the gut microbiota as a vital determinant in regulating therapeutic responses to immune checkpoint inhibitors (ICIs)> [13,14,96]. Fecal microbiota transplantation from humans to mice demonstrated that anti-CTLA-4 antibody treatment in melanoma patients promotes the expansion of Bacteroides fragilis, which possesses potent anticancer properties [9]. In previous studies, extensive research has shown that the gut microbiota composition serves as a reliable predictive biomarker for both the therapeutic efficacy of immune checkpoint blockade (ICB) therapy and the likelihood of associated adverse events.
In summary, the gut microbiota plays a critical role in cancer immunotherapy. Looking ahead, modulating the gut microbiota through approaches such as probiotics and fecal microbiota transplantation (FMT) may emerge as a pivotal strategy for enhancing therapeutic efficacy and reducing adverse effects in immunotherapy.
Importantly, VD is a potential co-regulator of this microbiota–immune axis to enhance responses in cancer immunotherapy. The following section thoroughly explores this emerging interplay between VD and gut microbiota.
Gut Microbiota and VD Synergy in Modulating Cancer Immunotherapy
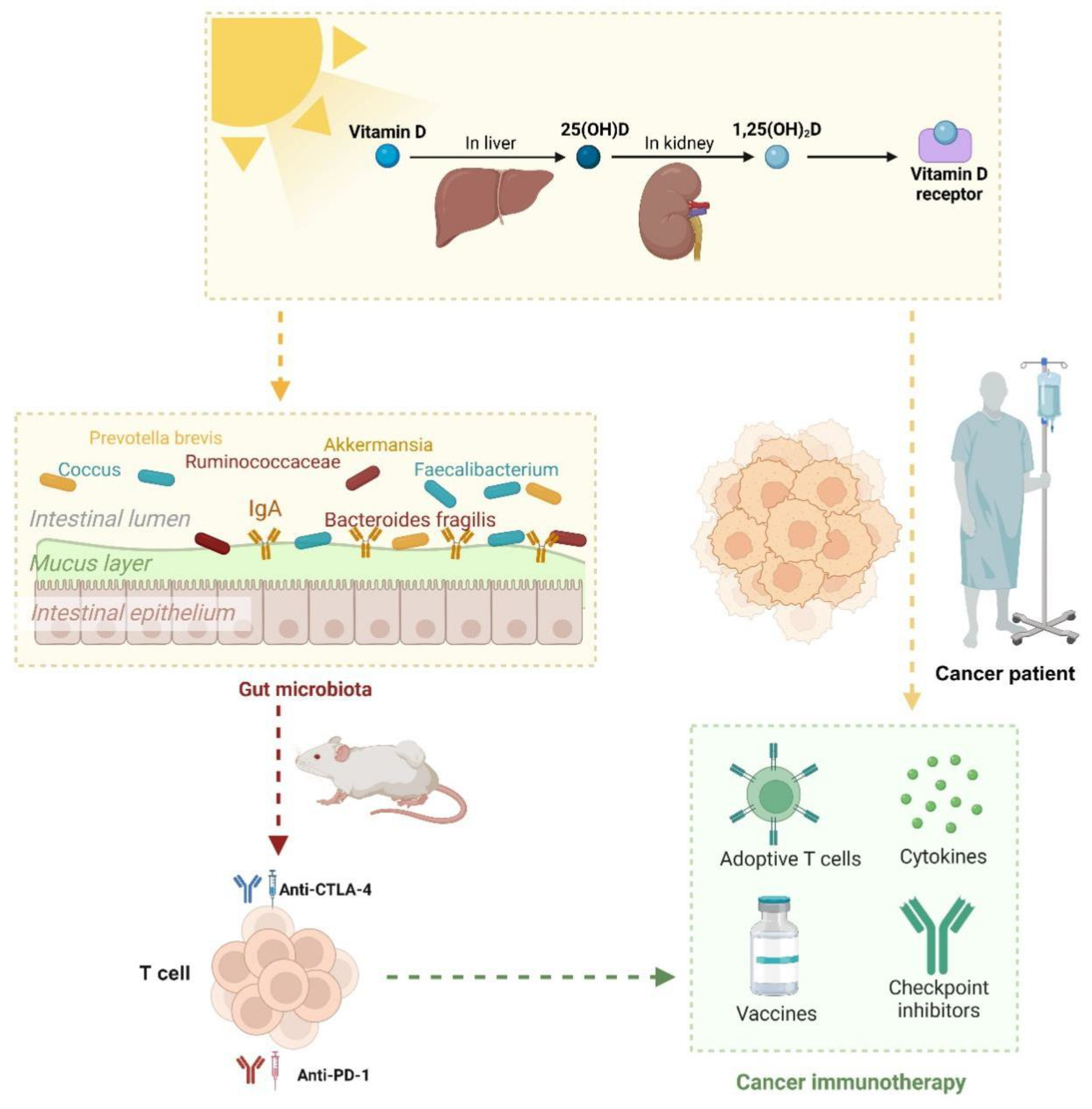
Recent studies have underscored the pivotal role of VD in immune regulation and its profound implications for cancer immunity [97,98,99]. Research by Giampazolias et al. [7] demonstrates that elevated VD levels enhance immune-mediated resistance to melanoma and improve responses to immune checkpoint blockade therapies. This immunomodulatory effect is mediated through VD's action on intestinal epithelial cells (IECs), which remodel the gut microbiota and promote the proliferation of Bacteroides fragilis, a bacterium known for its positive regulation of cancer immunity.
Animal studies reveal that VD deficiency or supplementation directly impacts gut microbiota composition, triggering significant immune response alterations. VD facilitates the expansion of Bacteroides fragilis and reduces the abundance of Prevotella brevis. This microbial shift markedly enhances anti-tumor immunity by boosting T cell activity and improving immunotherapy efficacy. These beneficial effects can be transferred via fecal microbiota transplantation (FMT), provided recipients maintain adequate dietary VD levels. This indicates that VD establishes a conducive microenvironment in the gut that supports the growth of beneficial bacteria like Bacteroides fragilis, thereby amplifying cancer immune responses. In human studies, VD levels and the expression of VD receptor (VDR) target genes correlate with improved cancer prognoses and enhanced efficacy of immune checkpoint inhibitor [7,97,98].
In summary, these findings suggest that VD status may serve as a predictive biomarker for immunotherapy outcomes and a potential therapeutic target to optimize cancer treatment [61,100]. This highlights the strategic potential of VD supplementation in modulating the microbiota and augmenting the effectiveness of cancer immunotherapies (Figure 5). Further clinical translational research is warranted to explore optimal approaches for VD supplementation in cancer patients.
Conclusions
The synergistic interaction among VD, gut microbiota, and cancer immunotherapy offers a novel research direction for tumor treatment. VD not only directly modulates the activity of immune cells through the VDR but also reshapes the gut microbiota structure, promoting the colonization of beneficial bacteria such as Bacteroides fragilis, thereby enhancing antitumor immune responses via the "microbiota-immune axis." Maintaining optimal VD levels can improve the efficacy of immune checkpoint inhibitors, reduce immune-related adverse events, and enhance patient survival rates. However, the role of VD is complex and paradoxical. While it generally suppresses the Th17/IL-17 pathway to mitigate inflammation and inhibit tumor progression, VD may unexpectedly enhance Th17 activity and even increase metastasis risk in specific populations or tumor types. Thus, the immunomodulatory effects of VD are highly dependent on individual variability, the tumor microenvironment, and gut microbiota composition.
Future Perspectives
Although the synergy between VD and immunotherapy shows great promise, several key issues remain to be addressed:
(1) Mechanistic Research: Further exploration is needed to understand how VD influences immune cell function via gut microbiota, particularly its bidirectional regulation of the Th17/Treg balance. Studies should also investigate VD's crosstalk with other critical signaling pathways (e.g., PPARγ, PI3K/AKT/mTOR) and its dynamic impact on PD-L1 expression.
(2) Personalized Treatment Strategies: Multi-omics data (e.g., metagenomics, metabolomics, immunomics) should be integrated to develop predictive models identifying patients who may benefit from VD supplementation. Precision intervention strategies, such as combining probiotics, prebiotics, or fecal microbiota transplantation (FMT), should be explored to optimize immunotherapy outcomes.
(3) Clinical Translation: Large-scale randomized controlled trials (RCTs) are required to determine the optimal VD dosage, timing, and target populations while avoiding the immunosuppressive risks of excessive supplementation. The combined use of VD with other immunomodulators should be investigated to develop more effective combination therapies.
(4) Technological Advancements: Rapid and cost-effective VD and gut microbiota detection methods should be developed to facilitate routine clinical monitoring. Organoid or humanized mouse models could help simulate VD-microbiota-immune system interactions, accelerating mechanistic research.
In summary, integrating VD and immunotherapy provides a promising new approach to cancer treatment, but its clinical application requires deeper mechanistic insights and rigorous validation. Combining basic research, multi-omics analysis, and personalized medicine, safer and more effective precision immunotherapy strategies may soon become a reality.
Author Contributions
All authors contributed to the conception and design of the study. Yan Yi-zhen and Guo Yi conducted the literature search, collation, interpretation, and wrote and revised the manuscript. Li Yi-ting and Zhao Li provided guidance and corrections for contents of article. Jiang Qing-rui and Yuan chen-hang provided guidance and corrections for the language of the article. The design, revision, and supervision of the dissertation were overseen by Shan-shan Mao. The final manuscript was read and approved by all authors.
Funding
This research was supported by the National Key R&D Program of China (No. 2022YFC3600201) and Chinese Universities Scientific Fund (No. 2024JNPD001).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We would like to thank all collaborators involved in this study for their valuable contributions to the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kennedy LB; Salama AKS. A review of cancer immunotherapy toxicity. CA Cancer J Clin. 2020, 70(2), 86-104. [CrossRef]
- Szeto GL; Finley SD. Integrative Approaches to Cancer Immunotherapy. Trends Cancer. 2019, 5(7), 400-410. [CrossRef]
- Bouillon R; Marcocci C; Carmeliet G; Bikle D; White JH; Dawson-Hughes B; Lips P; Munns CF; Lazaretti-Castro M; Giustina A; Bilezikian J. Skeletal and Extraskeletal Actions of Vitamin D: Current Evidence and Outstanding Questions. Endocr Rev. 2019, 40(4), 1109-1151. [CrossRef]
- Grover S; Dougan M; Tyan K; Giobbie-Hurder A; Blum SM; Ishizuka J; Qazi T; Elias R; Vora KB; Ruan AB; Martin-Doyle W; Manos M; Eastman L; Davis M; Gargano M; Haq R; Buchbinder EI; Sullivan RJ; Ott PA; Hodi FS; Rahma OE. Vitamin D intake is associated with decreased risk of immune checkpoint inhibitor-induced colitis. Cancer. 2020, 126(16), 3758-3767. [CrossRef]
- Ghaseminejad-Raeini A; Ghaderi A; Sharafi A; Nematollahi-Sani B; Moossavi M; Derakhshani A; Sarab GA. Immunomodulatory actions of vitamin D in various immune-related disorders: a comprehensive review. Front Immunol. 2023, 14, 950465. [CrossRef]
- Tsuji A; Yoshikawa S; Morikawa S; Ikeda Y; Taniguchi K; Sawamura H; Asai T; Matsuda S. Potential tactics with vitamin D and certain phytochemicals for enhancing the effectiveness of immune-checkpoint blockade therapies. Explor Target Antitumor Ther. 2023, 4(3), 460-473. [CrossRef]
- Giampazolias E; Pereira da Costa M; Lam KC; Lim KHJ; Cardoso A; Piot C; Chakravarty P; Blasche S; Patel S; Biram A; Castro-Dopico T; Buck MD; Rodrigues RR; Poulsen GJ; Palma-Duran SA; Rogers NC; Koufaki MA; Minutti CM; Wang P; Vdovin A; Frederico B; Childs E; Lee S; Simpson B; Iseppon A; Omenetti S; Kelly G; Goldstone R; Nye E; Suárez-Bonnet A; Priestnall SL; MacRae JI; Zelenay S; Patil KR; Litchfield K; Lee JC; Jess T; Goldszmid RS; Reis e Sousa C. Vitamin D regulates microbiome-dependent cancer immunity. Science. 2024, 384(6694), 428-437. [CrossRef]
- Fessler J; Matson V; Gajewski TF. Exploring the emerging role of the microbiome in cancer immunotherapy. J Immunother Cancer. 2019, 7(1), 108. [CrossRef]
- Vétizou M; Pitt JM; Daillère R; Lepage P; Waldschmitt N; Flament C; Rusakiewicz S; Routy B; Roberti MP; Duong CP; Poirier-Colame V; Roux A; Becharef S; Formenti S; Golden E; Cording S; Eberl G; Schlitzer A; Ginhoux F; Mani S; Yamazaki T; Jacquelot N; Enot DP; Bérard M; Nigou J; Opolon P; Eggermont A; Woerther PL; Chachaty E; Chaput N; Robert C; Mateus C; Kroemer G; Raoult D; Boneca IG; Carbonnel F; Chamaillard M; Zitvogel L. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science. 2015, 350(6264), 1079-84. [CrossRef]
- Chaput N; Lepage P; Coutzac C; Soularue E; Le Roux K; Monot C; Boselli L; Routier E; Cassard L; Collins M; Vaysse T; Marthey L; Eggermont A; Asvatourian V; Lanoy E; Mateus C; Robert C; Carbonnel F. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann Oncol. 2017, 28(6), 1368-1379. [CrossRef]
- Zhu X; Huang X; Hu M; Sun R; Li J; Wang H; Pan X; Ma Y; Ning L; Tong T; Zhou Y; Ding J; Zhao Y; Xuan B; Fang JY; Hong J; Hon Wong JW; Zhang Y; Chen H. A specific enterotype derived from gut microbiome of older individuals enables favorable responses to immune checkpoint blockade therapy. Cell Host Microbe. 2024, 32(4), 489-505.e5. [CrossRef]
- Park EM; Chelvanambi M; Bhutiani N; Kroemer G; Zitvogel L; Wargo JA. Targeting the gut and tumor microbiota in cancer. Nat Med. 2022, 28(4), 690-703. [CrossRef]
- Gopalakrishnan V; Spencer CN; Nezi L; Reuben A; Andrews MC; Karpinets TV; Prieto PA; Vicente D; Hoffman K; Wei SC; Cogdill AP; Zhao L; Hudgens CW; Hutchinson DS; Manzo T; Petaccia de Macedo M; Cotechini T; Kumar T; Chen WS; Reddy SM; Szczepaniak Sloane R; Galloway-Pena J; Jiang H; Chen PL; Shpall EJ; Rezvani K; Alousi AM; Chemaly RF; Shelburne S; Vence LM; Okhuysen PC; Jensen VB; Swennes AG; McAllister F; Marcelo Riquelme Sanchez E; Zhang Y; Le Chatelier E; Zitvogel L; Pons N; Austin-Breneman JL; Haydu LE; Burton EM; Gardner JM; Sirmans E; Hu J; Lazar AJ; Tsujikawa T; Diab A; Tawbi H; Glitza IC; Hwu WJ; Patel SP; Woodman SE; Amaria RN; Davies MA; Gershenwald JE; Hwu P; Lee JE; Zhang J; Coussens LM; Cooper ZA; Futreal PA; Daniel CR; Ajami NJ; Petrosino JF; Tetzlaff MT; Sharma P; Allison JP; Jenq RR; Wargo JA. Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients. Science. 2018, 359(6371), 97-103. [CrossRef]
- Matson V; Fessler J; Bao R; Chongsuwat T; Zha Y; Alegre ML; Luke JJ; Gajewski TF. The commensal microbiome is associated with anti–PD-1 efficacy in metastatic melanoma patients. Science. 2018, 359(6371), 104-108. [CrossRef]
- Kanstrup C; Teilum D; Rejnmark L; Bigaard JV; Eiken P; Kroman N; Tjønneland A; Mejdahl MK. 25-Hydroxyvitamin D at time of breast cancer diagnosis and breast cancer survival. Breast Cancer Res Treat. 2020, 179(3), 699-708. [CrossRef]
- Filip-Psurska B; Zachary H; Strzykalska A; Wietrzyk J. Vitamin D, Th17 Lymphocytes, and Breast Cancer. Cancers. 2022, 14(15), 3649. [CrossRef]
- Holick MF; Chen TC. Vitamin D deficiency: a worldwide problem with health consequences. Am J Clin Nutr. 2008, 87(4), 1080S-6S. [CrossRef]
- Palacios C; Gonzalez L. Is vitamin D deficiency a major global public health problem? J Steroid Biochem Mol Biol. 2014, 144 Pt A, 138-45. [CrossRef]
- Roth DE; Abrams SA; Aloia J; Bergeron G; Bourassa MW; Brown KH; Calvo MS; Cashman KD; Combs G; De-Regil LM; Jefferds ME; Jones KS; Kapner H; Martineau AR; Neufeld LM; Schleicher RL; Thacher TD; Whiting SJ. Global prevalence and disease burden of vitamin D deficiency: a roadmap for action in low- and middle-income countries. Ann N Y Acad Sci. 2018,1430(1),44-79. [CrossRef]
- Giustina A; Bilezikian JP; Adler RA; Banfi G; Bikle DD; Binkley NC; Bollerslev J; Bouillon R; Brandi ML; Casanueva FF; di Filippo L; Donini LM; Ebeling PR; Fuleihan GE; Fassio A; Frara S; Jones G; Marcocci C; Martineau AR; Minisola S; Napoli N; Procopio M; Rizzoli R; Schafer AL; Sempos CT; Ulivieri FM; Virtanen JK. Consensus Statement on Vitamin D Status Assessment and Supplementation: Whys, Whens, and Hows. Endocr Rev. 2024,45(5),625-654. [CrossRef]
- Saccone D; Asani F; Bornman L. Regulation of the vitamin D receptor gene by environment, genetics and epigenetics. Gene.2015,561(2),171-80. [CrossRef]
- Carlberg C; Raczyk M; Zawrotna N. Vitamin D: A master example of nutrigenomics. Redox Biol. 2023, 62,102695. [CrossRef]
- Delrue C; Speeckaert MM. Vitamin D and Vitamin D-Binding Protein in Health and Disease. International Journal of Molecular Sciences. 2023, 24(5),4642. [CrossRef]
- Thouvenot E; Laplaud D; Lebrun-Frenay C; Derache N; Le Page E; Maillart E; Froment-Tilikete C; Castelnovo G; Casez O; Coustans M; Guennoc AM; Heinzlef O; Magy L; Nifle C; Ayrignac X; Fromont A; Gaillard N; Caucheteux N; Patry I; De Sèze J; Deschamps R; Clavelou P; Biotti D; Edan G; Camu W; Agherbi H; Renard D; Demattei C; Fabbro-Peray P; Mura T; Rival M; D-Lay MS Investigators. High-Dose Vitamin D in Clinically Isolated Syndrome Typical of Multiple Sclerosis: The D-Lay MS Randomized Clinical Trial. JAMA. 2025,333(16),1413-1422. [CrossRef]
- Xiang H; Zhou C; Gan X; Huang Y; He P; Ye Z; Liu M; Yang S; Zhang Y; Zhang Y; Qin X. Relationship of Serum 25-Hydroxyvitamin D Concentrations, Diabetes, Vitamin D Receptor Gene Polymorphisms and Incident Venous Thromboembolism. Diabetes Metab Res Rev. 2025, 41(1),e70014. [CrossRef]
- Charoenngam N; Shirvani A; Holick MF. Vitamin D for skeletal and non-skeletal health: What we should know. J Clin Orthop Trauma. 2019, 10(6), 1082-1093. [CrossRef]
- Demay MB; Pittas AG; Bikle DD; Diab DL; Kiely ME; Lazaretti-Castro M; Lips P; Mitchell DM; Murad MH; Powers S; Rao SD; Scragg R; Tayek JA; Valent AM; Walsh JME; McCartney CR. Vitamin D for the Prevention of Disease: An Endocrine Society Clinical Practice Guideline. J Clin Endocrinol Metab. 2024,109(8):1907-1947. [CrossRef]
- Fekete M; Lehoczki A; Szappanos Á; Zábó V; Kaposvári C; Horváth A; Farkas Á; Fazekas-Pongor V; Major D; Lipécz Á; Csípő T; Varga JT. Vitamin D and Colorectal Cancer Prevention: Immunological Mechanisms, Inflammatory Pathways, and Nutritional Implications. Nutrients. 2025,17(8):1351. [CrossRef]
- Thouvenot E; Laplaud D; Lebrun-Frenay C; Derache N; Le Page E; Maillart E; Froment-Tilikete C; Castelnovo G; Casez O; Coustans M; Guennoc AM; Heinzlef O; Magy L; Nifle C; Ayrignac X; Fromont A; Gaillard N; Caucheteux N; Patry I; De Sèze J; Deschamps R; Clavelou P; Biotti D; Edan G; Camu W; Agherbi H; Renard D; Demattei C; Fabbro-Peray P; Mura T; Rival M; D-Lay MS Investigators. High-Dose Vitamin D in Clinically Isolated Syndrome Typical of Multiple Sclerosis. Jama 2025,333(16):1413-1422. [CrossRef]
- Lin K; Miao Y; Gan L; Zhao B; Fang F; Wang R; Chen X; Huang J. Associations of Serum 25-Hydroxyvitamin D Concentrations with Risks of Mortality and Cardiovascular Disease among Individuals with Psoriasis. J Am Acad Dermatol. 2025, S0190-9622(25)02118-8. [CrossRef]
- Sha S; Xie R; Gwenzi T; Wang Y; Brenner H; Schöttker B. Real-world evidence for an association of vitamin D supplementation with atherosclerotic cardiovascular disease in the UK Biobank. Clin Nutr. 2025,49,118-127. [CrossRef]
- Ruiz-García A; Pallarés-Carratalá V; Turégano-Yedro M; Torres F; Sapena V; Martin-Gorgojo A; Martin-Moreno JM. Vitamin D Supplementation and Its Impact on Mortality and Cardiovascular Outcomes: Systematic Review and Meta-Analysis of 80 Randomized Clinical Trials. Nutrients. 2023,15(8),1810. [CrossRef]
- Wang X; Li Q; Lyu Z; Wu Y. Supplementing with Vitamin D during Pregnancy Reduces Inflammation and Prevents Autism-Related Behaviors in Offspring Caused by Maternal Immune Activation. Biol Pharm Bull. 2025,48(5),632-640. [CrossRef]
- Mullard A. Addressing cancer's grand challenges. Nat Rev Drug Discov. 2020, 19(12), 825-826. [CrossRef]
- Elkin EB; Bach PB. Cancer's next frontier: addressing high and increasing costs. JAMA. 2010,303(11),1086-7. [CrossRef]
- Riley RS; June CH; Langer R; Mitchell MJ. Delivery technologies for cancer immunotherapy. Nature Reviews Drug Discovery. 2019,18(3),175-196. [CrossRef]
- Vanhooren J; Derpoorter C; Depreter B; Deneweth L; Philippé J; De Moerloose B; Lammens T. TARP as antigen in cancer immunotherapy. Cancer Immunol Immunother. 2021,70(11),3061-3068. [CrossRef]
- Paleari, L. Personalized Assessment for Cancer Prevention, Detection, and Treatment. Int J Mol Sci. 2024.25(15),8140. [CrossRef]
- Yu Q; Ding J; Li S; Li Y. Autophagy in cancer immunotherapy: Perspective on immune evasion and cell death interactions. Cancer Lett. 2024,590,216856. [CrossRef]
- Song S; Woo HD; Lyu J; Song BM; Lim JY; Park HY. Serum 25-hydroxyvitamin D levels and risk of overall and site-specific cancers in Korean adults: results from two prospective cohort studies. Nutr J. 2025, 24(1), 84. [CrossRef]
- Lim ST; Jeon YW; Suh YJ. Association Between Alterations in the Serum 25-Hydroxyvitamin D Status During Follow-Up and Breast Cancer Patient Prognosis. Asian Pac J Cancer Prev. 2015, 16(6),2507-13. [CrossRef]
- Shirazi L; Almquist M; Borgquist S; Malm J; Manjer J. Serum vitamin D (25OHD3) levels and the risk of different subtypes of breast cancer: A nested case–control study. Breast. 2016,28,184-90. [CrossRef]
- Li H; Liu H; Wang B; Jia X; Yu J; Zhang Y; Sang D; Zhang Y. Exercise Interventions for the Prevention and Treatment of Anthracycline-Induced Cardiotoxicity in Women with Breast Cancer: A Systematic Review. J. Sci. Sport Exerc. 2024, 7 (1), 14-27. [CrossRef]
- Estébanez N; Gómez-Acebo I; Palazuelos C; Llorca J; Dierssen-Sotos T. Vitamin D exposure and Risk of Breast Cancer: a meta-analysis. Sci Rep. 2018,8(1),9039. [CrossRef]
- Ordóñez-Mena JM; Schöttker B; Fedirko V; Jenab M; Olsen A; Halkjær J; Kampman E; de Groot L; Jansen E; Bueno-de-Mesquita HB; Peeters PH; Siganos G; Wilsgaard T; Perna L; Holleczek B; Pettersson-Kymmer U; Orfanos P; Trichopoulou A; Boffetta P; Brenner H. Pre-diagnostic vitamin D concentrations and cancer risks in older individuals: an analysis of cohorts participating in the CHANCES consortium. Eur J Epidemiol. 2016, 31(3),311-23. [CrossRef]
- Pouliliou S; Nikolaidis C; Drosatos G. Current trends in cancer immunotherapy: a literature-mining analysis. Cancer Immunol Immunother. 2020,69(12),2425-2439. [CrossRef]
- Yu WD; Sun G; Li J; Xu J; Wang X. Mechanisms and therapeutic potentials of cancer immunotherapy in combination with radiotherapy and/or chemotherapy. Cancer Lett. 2019,452,66-70. [CrossRef]
- Kaiser M; Semeraro MD; Herrmann M; Absenger G; Gerger A; Renner W. Immune Aging and Immunotherapy in Cancer. Int J Mol Sci 2021,22(13),7016. [CrossRef]
- Topalian SL; Weiner GJ; Pardoll DM. Cancer Immunotherapy Comes of Age. J Clin Oncol. 2011, 29(36),4828-36. [CrossRef]
- Rui R; Zhou L; He S. Cancer immunotherapies: advances and bottlenecks. Front Immunol. 2023,14,1212476. [CrossRef]
- Chun RF; Liu PT; Modlin RL; Adams JS; Hewison M. Impact of vitamin D on immune function: lessons learned from genome-wide analysis. Front Physiol. 2014, 5, 151. [CrossRef]
- Charoenngam N; Holick MF. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients. 2020, 12(7), 2097. [CrossRef]
- Feldman D; Krishnan AV; Swami S; Giovannucci E; Feldman BJ. The role of vitamin D in reducing cancer risk and progression. Nat Rev Cancer. 2014, 14(5), 342-57. [CrossRef]
- Martens PJ; Gysemans C; Verstuyf A; Mathieu AC. Vitamin D's Effect on Immune Function. Nutrients. 2020, 2(5), 1248. [CrossRef]
- Artusa P; White JH. Vitamin D and its analogs in immune system regulation. Pharmacol Rev. 2025, 77(2), 100032. [CrossRef]
- Sîrbe C; Rednic S; Grama A; Pop TL. An Update on the Effects of Vitamin D on the Immune System and Autoimmune Diseases. Int J Mol Sci. 2022, 23(17), 9784. [CrossRef]
- Das D; Karthik N; Taneja R. Crosstalk Between Inflammatory Signaling and Methylation in Cancer. Front Cell Dev Biol. 2021, 9, 756458. [CrossRef]
- Kim DH; Meza CA; Clarke H; Kim JS; Hickner RC. Vitamin D and Endothelial Function. Nutrients. 2020, 12(2), 575. [CrossRef]
- Holick MF. The vitamin D deficiency pandemic: Approaches for diagnosis, treatment and prevention. Rev Endocr Metab Disord. 2017, 18(2), 153-165. [CrossRef]
- Giulietti A; van Etten E; Overbergh L; Stoffels K; Bouillon R; Mathieu C. Monocytes from type 2 diabetic patients have a pro-inflammatory profile. 1,25-Dihydroxyvitamin D(3) works as anti-inflammatory. Diabetes Res Clin Pract. 2007, 77(1). 47-57. [CrossRef]
- Galus Ł; Michalak M; Lorenz M; Stoińska-Swiniarek R; Tusień Małecka D; Galus A; Kolenda T; Leporowska E; Mackiewicz J. Vitamin D supplementation increases objective response rate and prolongs progression-free time in patients with advanced melanoma undergoing anti-PD-1 therapy. Cancer. 2023, 129(13), 2047-2055. [CrossRef]
- Stucci LS; D'Oronzo S; Tucci M; Macerollo A; Ribero S; Spagnolo F; Marra E; Picasso V; Orgiano L; Marconcini R; De Rosa F; Di Guardo L; Galli G; Gandini S; Palmirotta R; Palmieri G; Queirolo P; Silvestris F. Italian Melanoma Intergroup (IMI). Vitamin D in melanoma: Controversies and potential role in combination with immune check-point inhibitors. Cancer Treat Rev. 2018, 69, 21-28. [CrossRef]
- Bersanelli M; Cortellini A; Leonetti A; Parisi A; Tiseo M; Bordi P; Michiara M; Bui S; Cosenza A; Ferri L; Giudice GC; Testi I; Rapacchi E; Camisa R; Vincenzi B; Caruso G; Rauti AN; Arturi F; Tucci M; Santo V; Ricozzi V; Burtet V; Sgargi P; Todeschini R; Zustovich F; Stucci LS; Santini D; Buti S. Systematic vitamin D supplementation is associated with improved outcomes and reduced thyroid adverse events in patients with cancer treated with immune checkpoint inhibitors: results from the prospective PROVIDENCE study. Cancer Immunol Immunother. 2023, 72(11), 3707-3716. [CrossRef]
- Mondul AM; Weinstein SJ; Layne TM; Albanes D. Vitamin D and Cancer Risk and Mortality: State of the Science, Gaps, and Challenges. Epidemiol Rev. 2017, 39(1), 28-48. [CrossRef]
- Jeffery LE; Burke F; Mura M; Zheng Y; Qureshi OS; Hewison M; Walker LS, Lammas DA; Raza K; Sansom DM. 1,25-Dihydroxyvitamin D3 and IL-2 combine to inhibit T cell production of inflammatory cytokines and promote development of regulatory T cells expressing CTLA-4 and FoxP3. J Immunol. 2009, 183(9), 5458-67. [CrossRef]
- Adorini L; Daniel KC; Penna G. Vitamin D receptor agonists, cancer and the immune system: an intricate relationship. Curr Top Med Chem. 2006, 6(12), 1297-301. [CrossRef]
- Wang J; Zhu N; Su X; Gao Y; Yang R. Gut-Microbiota-Derived Metabolites Maintain Gut and Systemic Immune Homeostasis. Cells. 2023, 12(5), 793. [CrossRef]
- Fan Y; Pedersen O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021, 19(1), 55-71. [CrossRef]
- Adak A; Khan MR. An insight into gut microbiota and its functionalities. Cell Mol Life Sci. 2019, 76(3), 473-493. [CrossRef]
- Zmora N; Suez J; Elinav E. You are what you eat: diet, health and the gut microbiota. Nat Rev Gastroenterol Hepatol. 2019, 16(1), 35-56. [CrossRef]
- Liao X; Lan Y; Wang W; Zhang J; Shao R; Yin Z; Gudmundsson GH; Bergman P; Mai K; Ai Q; Wan M. Vitamin D influences gut microbiota and acetate production in zebrafish (Danio rerio) to promote intestinal immunity against invading pathogens. Gut Microbes. 2023, 15(1), 2187575. [CrossRef]
- Fan L; Xia Y; Wang Y; Han D; Liu Y; Li J; Fu J; Wang L; Gan Z; Liu B; Fu J; Zhu C; Wu Z; Zhao J; Han H; Wu H; He Y; Tang Y; Zhang Q; Wang Y; Zhang F; Zong X; Yin J; Zhou X; Yang X; Wang J; Yin Y; Ren W. Gut microbiota bridges dietary nutrients and host immunity. Sci China Life Sci. 2023, 66(11), 2466-2514. [CrossRef]
- Góralczyk-Bińkowska A; Szmajda-Krygier D; Kozłowska E. The Microbiota-Gut-Brain Axis in Psychiatric Disorders. Int J Mol Sci. 2022, 23(19), 11245. [CrossRef]
- Wang Z; Wang Z; Lu T; Chen W; Yan W; Yuan K; Shi L; Liu X; Zhou X; Shi J; Vitiello MV; Han Y; Lu L. The microbiota-gut-brain axis in sleep disorders. Sleep Med Rev. 2022, 65, 101691. [CrossRef]
- Zuo WF; Pang Q; Yao LP; Zhang Y; Peng C; Huang W; Han B. Gut microbiota: A magical multifunctional target regulated by medicine food homology species. J Adv Res. 2023 ,52, 151-170. [CrossRef]
- Huang L; Lum D; Haiyum M; Fairbairn KA. Vitamin D Status of Elite Athletes in Singapore and Its Associations With Muscle Function and Bone Health. SSEJ. 2021, 3 (4), 385-393. [CrossRef]
- Ullah H. Gut-vitamin D interplay: key to mitigating immunosenescence and promoting healthy ageing. Immun Ageing. 2025, 22(1), 20. [CrossRef]
- Aggeletopoulou I; Tsounis EP; Mouzaki A; Triantos C. Exploring the Role of Vitamin D and the Vitamin D Receptor in the Composition of the Gut Microbiota. Front Biosci (Landmark Ed). 2023, 28(6), 116. [CrossRef]
- Sun J; Zhang YG. Vitamin D Receptor Influences Intestinal Barriers in Health and Disease. Cells. 2022, 11(7), 1129. [CrossRef]
- Waterhouse M; Hope B; Krause L; Morrison M; Protani MM; Zakrzewski M; Neale RE. Vitamin D and the gut microbiome: a systematic review of in vivo studies. Eur J Nutr. 2019, 58(7), 2895-2910. [CrossRef]
- Jamshidi S; Masoumi SJ; Abiri B; Vafa M. The effects of synbiotic and/or vitamin D supplementation on gut-muscle axis in overweight and obese women: a study protocol for a double-blind, randomized, placebo-controlled trial. Trials. 2022, 23(1), 631. [CrossRef]
- Boughanem H; Ruiz-Limón P; Pilo J; Lisbona-Montañez JM; Tinahones FJ; Moreno Indias I; Macías-González M. Linking serum vitamin D levels with gut microbiota after 1-year lifestyle intervention with Mediterranean diet in patients with obesity and metabolic syndrome: a nested cross-sectional and prospective study. Gut Microbes. 2023 ,15(2), 2249150. [CrossRef]
- Thomas RL; Jiang L; Adams JS; Xu ZZ; Shen J; Janssen S; Ackermann G; Vanderschueren D; Pauwels S; Knight R; Orwoll ES; Kado DM. Vitamin D metabolites and the gut microbiome in older men. Nat Commun. 2020 ,11(1), 5997. [CrossRef]
- Clark A; Mach N. Role of Vitamin D in the Hygiene Hypothesis: The Interplay between Vitamin D, Vitamin D Receptors, Gut Microbiota, and Immune Response. Front Immunol. 2016, 7, 627. [CrossRef]
- Assa A; Vong L; Pinnell LJ; Avitzur N; Johnson-Henry KC; Sherman PM. Vitamin D deficiency promotes epithelial barrier dysfunction and intestinal inflammation. J Infect Dis. 2014, 210(8), 1296-305. [CrossRef]
- Battistini C; Ballan R; Herkenhoff ME; Saad SMI; Sun J. Vitamin D Modulates Intestinal Microbiota in Inflammatory Bowel Diseases. Int J Mol Sci. 2020, 22(1), 362. [CrossRef]
- Du J; Wei X; Ge X; Chen; Li YC. Microbiota-Dependent Induction of Colonic Cyp27b1 Is Associated With Colonic Inflammation: Implications of Locally Produced 1,25-Dihydroxyvitamin D3 in Inflammatory Regulation in the Colon. Endocrinology. 2017,158(11),4064-4075. [CrossRef]
- Jin D; Wu S; Zhang YG; Lu R; Xia Y; Dong H; Sun J. Lack of Vitamin D Receptor Causes Dysbiosis and Changes the Functions of the Murine Intestinal Microbiome. Clin Ther. 2015, 7(5), 996-1009.e7. [CrossRef]
- Barbáchano A; Fernández-Barral A; Ferrer-Mayorga G; Costales-Carrera ; Larriba MJ; Muñoz A. The endocrine vitamin D system in the gut. Mol Cell Endocrinol. 2017, 453, 79-87. [CrossRef]
- Wang H; Tian G; Pei Z; Yu X; Wang Y; Xu F; Zhao J; Lu S; Lu W. Bifidobacterium longum increases serum vitamin D metabolite levels and modulates intestinal flora to alleviate osteoporosis in mice. mSphere. 2025, 10(3), e0103924. [CrossRef]
- Yang X; Zhu Q; Zhang L; Pei Y; Xu X; Liu X; Lu G; Pan J; Wang Y. Causal relationship between gut microbiota and serum vitamin D: evidence from genetic correlation and Mendelian randomization study. Eur J Clin Nutr. 2022,76(7),1017-1023. [CrossRef]
- Ma C; Han M; Heinrich B; Fu Q; Zhang Q; Sandhu M; Agdashian D; Terabe M; Berzofsky JA; Fako V; Ritz T; Longerich T; Theriot CM; McCulloch JA; Roy S; Yuan W; Thovarai V; Sen SK; Ruchirawat M; Korangy; Wang XW; Trinchieri G; Greten TF. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science. 2018, 360(6391), eaan5931. [CrossRef]
- Song X; Sun X; Oh SF; Wu M; Zhang Y; Zheng W; Geva-Zatorsky N; Jupp R; Mathis D; Benoist C; Kasper DL. Microbial bile acid metabolites modulate gut RORγ+ regulatory T cell homeostasis. Nature. 2020,577(7790),410-415. [CrossRef]
- Sardar P; Beresford-Jones BS; Xia W, Shabana O; Suyama S; Ramos RJF; Soderholm AT; Tourlomousis P; Kuo P; Evans AC; Imianowski CJ; Conti AG; Wesolowski AJ; Baker NM; McCord EAL; Okkenhaug K; Whiteside SK; Roychoudhuri R; Bryant CE; Cross JR; Pedicord VA. Gut microbiota-derived hexa-acylated lipopolysaccharides enhance cancer immunotherapy responses. Nat Microbiol. 2025,10(3),795-807. [CrossRef]
- Zitvogel L; Ma Y; Raoult D; Kroemer G; Gajewski TF. The microbiome in cancer immunotherapy: Diagnostic tools and therapeutic strategies. Science. 2018, 359(6382),1366-1370. [CrossRef]
- Routy B; Le Chatelier E; Derosa L; Duong CPM; Alou MT; Daillère R; Fluckiger A, Messaoudene M; Rauber C, Roberti MP; Fidelle M; Flament C; Poirier-Colame V; Opolon P; Klein C; Iribarren K; Mondragón L; Jacquelot N; Qu B; Ferrere G; Clémenson C; Mezquita L; Masip JR; Naltet C; Brosseau S; Kaderbhai C; Richard C; Rizvi H; Levenez F; Galleron N; Quinquis B; Pons N; Ryffel B; Minard-Colin V; Gonin P; Soria JC; Deutsch E; Loriot Y; Ghiringhelli F; Zalcman G; Goldwasser F; Escudier B; Hellmann MD; Eggermont A; Raoult D; Albiges L; Kroemer G; Zitvogel L. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science. 2018, 359(6371),91-97. [CrossRef]
- Izquierdo JM. Vitamin D-dependent microbiota-enhancing tumor immunotherapy. Cell Mol Immunol. 2024,21(10),1083-1086. [CrossRef]
- Franco F; McCoy KD. Microbes and vitamin D aid immunotherapy. Science.2024, 384(6694), 384-385. [CrossRef]
- Kanno K; Akutsu T; Ohdaira H; Suzuki Y; Urashima M. Effect of Vitamin D Supplements on Relapse or Death in a p53-Immunoreactive Subgroup With Digestive Tract Cancer: Post Hoc Analysis of the AMATERASU Randomized Clinical Trial. JAMA Netw Open. 2023, 6(8), e2328886. [CrossRef]
- Fernández-Barral A; Peña C; Pisano DG; Cantero R; Rojo F; Muñoz A; Larriba MJ. Vitamin D receptor expression and associated gene signature in tumour stromal fibroblasts predict clinical outcome in colorectal cancer. Gut.2017,66(8),1449-1462. [CrossRef]
Figure 1.
VD metabolism and mechanism of action.
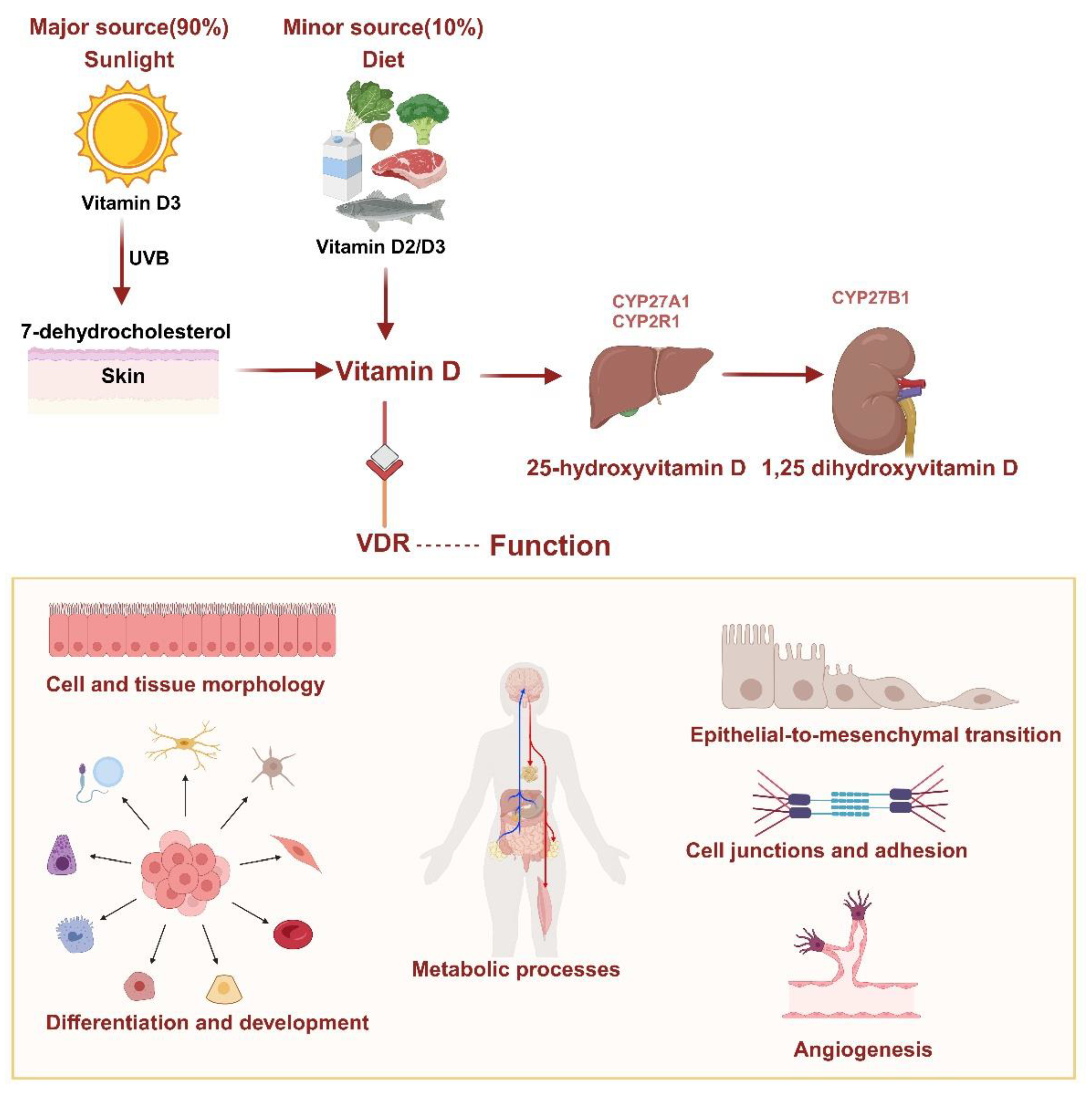
Figure 4.
VD can alleviate cancer progression and benefit the immune response of cancer immunotherapy.
Figure 4.
VD can alleviate cancer progression and benefit the immune response of cancer immunotherapy.
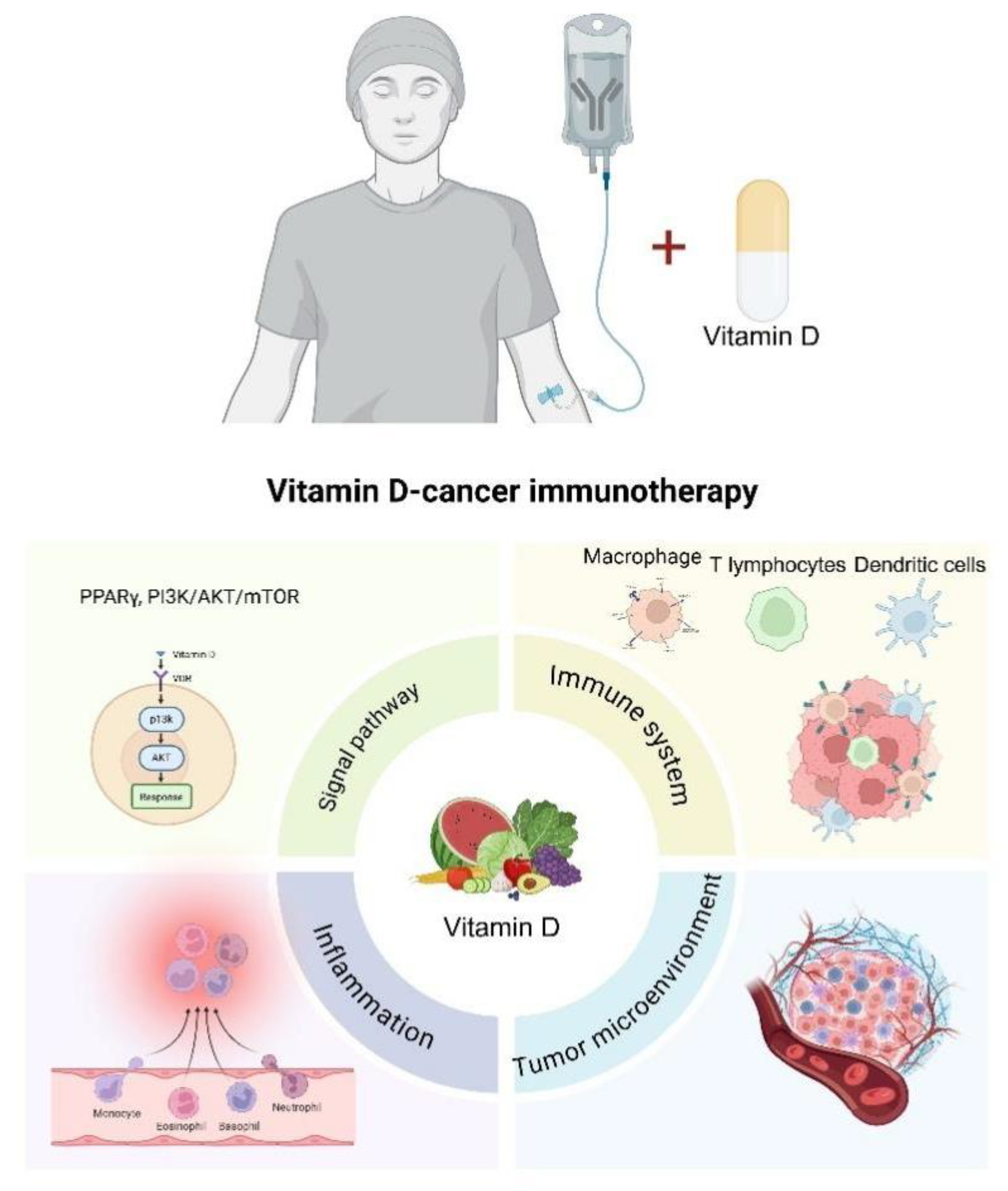
Figure 2.
VD serves as a key regulator of gut microbiota, with bidirectional interactions between them.
Figure 2.
VD serves as a key regulator of gut microbiota, with bidirectional interactions between them.
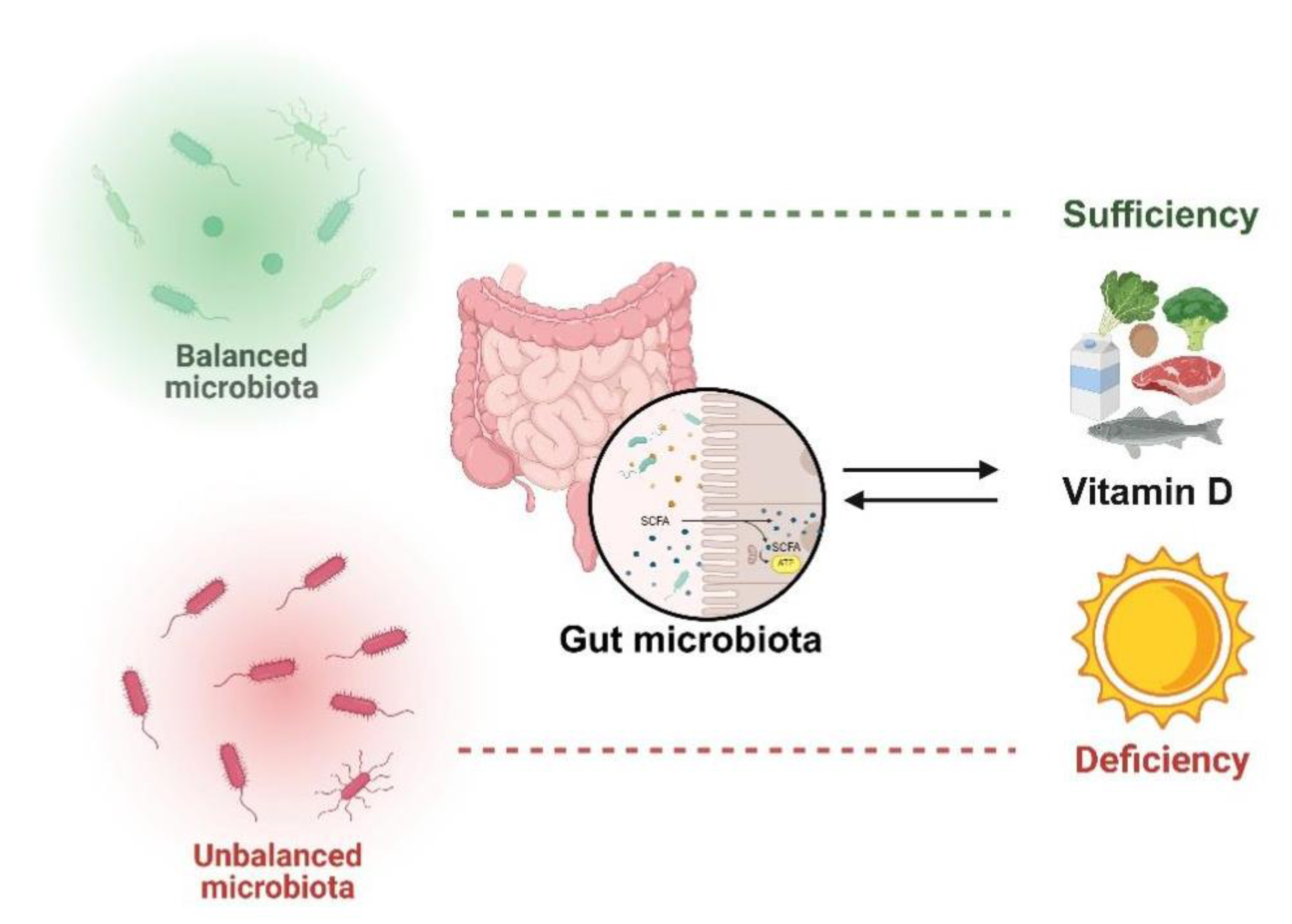
Figure 3.
Gut microbiota impact on cancer immunotherapy.
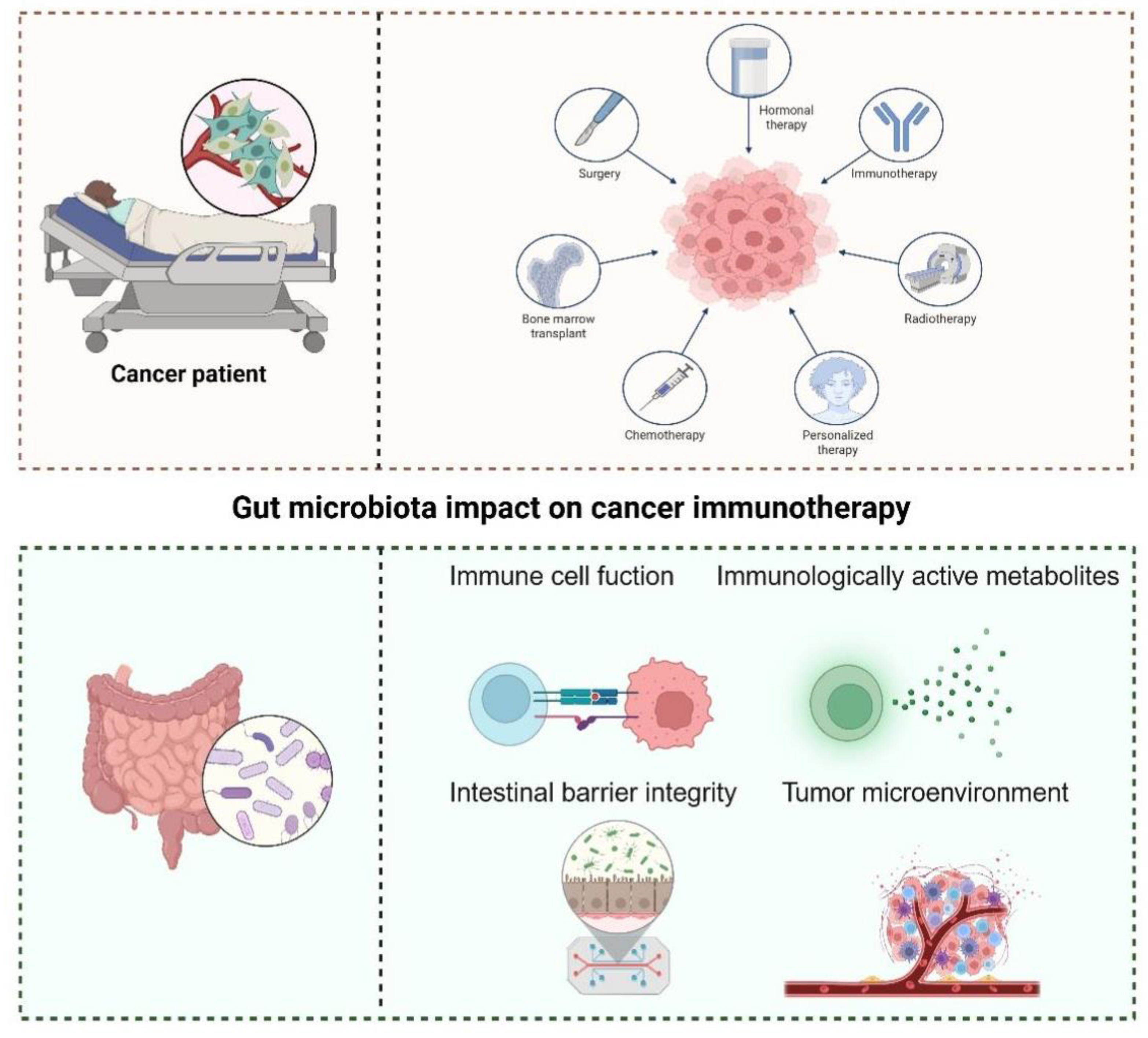
Figure 5.
VD-dependent microbiota-enhancing tumor immunotherapy.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



