Submitted:
30 May 2025
Posted:
02 June 2025
You are already at the latest version
Abstract
PrimPol is a human DNA primase and DNA polymerase involved in DNA damage tolerance both in nuclei and mitochondria. PrimPol restarts stalled replication forks by synthesizing DNA primers de novo and also possesses DNA translesion activity (TLS activity). PrimPol efficiently and relatively accurately bypasses several DNA lesions including 8-oxoguanine and 5-formyluracil as well as carries out synthesis on DNA with thymine glycol in an error-prone manner. In this work, we showed that PrimPol possesses efficient and accurate TLS activity across 8-oxoadenine, another common DNA lesion caused by oxidative stress. The accuracy of PrimPol on DNA with 8-oxoA was significantly higher compared to DNA containing 8-oxoG. Replacement of Mg2+ ions with Mn2+ stimulated activity of PrimPol on DNA with 8-oxoA and 8-oxoG as well as undamaged A in a sequence-dependent manner by lesion skipping (or template scrunching) mechanism. Altogether, our data support the idea that PrimPol possesses efficient TLS activity across a wide range of DNA lesions caused by oxidative stress.
Keywords:
PrimPol
; 8-oxoadenine
; DNA translesion synthesis
1. Introduction
Human PrimPol possesses DNA primase and DNA polymerase activities and is present in both nucleus and mitochondria [1,2,3]. It is involved in DNA damage tolerance by restart of replication forks at the sites of DNA damage and non-B DNA structures such as G-quadruplexes [4,5,6,7,8,9]. Cells deficient in PrimPol are sensitive to DNA-damaging agents [3,10,11].
PrimPol also demonstrates DNA translesion activity (TLS activity) and can bypass a variety of small DNA lesions [1,12]. PrimPol is blocked on DNA with bulky N2-dG adducts [13] and a DNA-protein and AP site–peptide cross-links [14,15] but can bypass and incorporate complementary nucleotides opposite the cisplatin GG cross-link [15].
Previously, we and others demonstrated that PrimPol carries out efficient and relatively accurate synthesis past DNA lesions caused by oxidation such as 8-oxoguanine (8-oxoG) and 5-formyluracil [1,12,16]. Substitution of Mg2+ with Mn2+ ions also stimulated the synthesis on DNA with thymine glycol in an error-prone manner, likely by lesion skipping (or template scrunching) mechanism [12]. Such efficient TLS activity might facilitate PrimPol-mediated repriming on severely damaged DNA, e.g. containing clustered DNA damage.
Along with 8-oxoG, 8-oxoadenine (8-oxoA) is the most abundant oxidative lesion [17,18,19,20] with dual miscoding properties. These modified bases readily adopt the syn conformation: 8-oxoG forms stable 8-oxoG(syn):A(anti) Hoogsteen mispair while 8-oxoA efficiently forms the 8-oxoA(syn):G(anti) pair [21,22]. The majority of DNA polymerases preferentially incorporates non-complementary dAMP opposite 8-oxoG (so-called “A-rule”) leading to G:C→T:A transversions [22,23]. While 8-oxoA is not mutagenic in Escherichia coli [24], its mutagenic effect was demonstrated in mammalian cells [25,26]. The 8-oxoA lesion placed in the HRAS oncogene sequence stimulated A:T → C:G transversions and A:T → G:C transitions at a relatively high frequency of mutagenesis comparable [25] or 4-fold reduced [27] to that caused by 8-oxoG.
In this work, we for the first time report activity of PrimPol on DNA with 8-oxoA. We showed that PrimPol accurately bypasses 8-oxoA in vitro. We also analyzed the effect of metal ions and DNA sequence context on the TLS activity of PrimPol on DNA with 8-oxoA and compared these data with those obtained for 8-oxoG.
Altogether, these findings further support the possible role of PrimPol in replication of DNA with oxidative damage.
2. Results
2.1. Efficient and Accurate Bypass of 8-oxoA in Reactions in the Presence of Mg2+
First, we analyzed activity of PrimPol in reactions in the presence of Mg2+ on DNA substrates with the HRAS oncogene sequence context 5`-CCXA-3` containing A, G, 8-oxoA or 8-oxoG in the +1 position. Both DNA lesions, 8-oxoA and 8-oxoG, only slightly inhibited activity of PrimPol (Figure 1). Unlike many other DNA polymerases, PrimPol was more accurate and incorporated opposite 8-oxoG complementary dCMP with slight preference over non-complementary dAMP (Figure 1A, Table 1). However, PrimPol almost exclusively incorporated complementary dTMP opposite A and 8-oxoA (Figure 1, Table 1). PrimPol incorporated non-complementary dGMP with 3- to 4-fold reduced efficiency on DNA template with 8-oxoA and with a 10-fold reduction in efficiency on DNA template with undamaged A compared to dTMP (Figure 1C, Table 1). Interestingly, incorporation of dGMP was observed as a ladder in reactions in the presence of DNA templates containing 8-oxoA, 8-oxoG or undamaged A but not in reactions with template G (Figure 1A, lanes 4, 10, 16, 22). This activity can be a result of dGMP incorporation opposite C in the +2 and +3 template positions during template scrunching (or lesion skipping) leading to small deletions.
2.2. Mn2+ Ions Decrease Accuracy of PrimPol on DNA Substrates with 8-Oxopurines
The DNA polymerase activity of PrimPol as well as the template scrunching mechanism is stimulated by Mn2+ ions [12,13,28]. Indeed, replacement of Mg2+ with Mn2+ ions stimulated activity of PrimPol and reduced its accuracy on all DNA templates (Figure 2). PrimPol carried out error-prone synthesis on DNA with 8-oxoG. Enzyme incorporated dAMP slightly more efficient than complementary dCMP (Figure 2A, lane 21, Table 1).
Mn2+ ions also facilitated incorporation of dGMP and dTMP on DNA with 8-oxoG (Figure 2A, lanes 22, 23). Mn2+ stimulated the incorporation of dGMP on both templates A and 8-oxoA (Figure 2A, lanes 4 and 16, Тable 1). PrimPol was slightly more accurate on DNA with A compared to 8-oxoA in the presence of Mg2+ (Table 1). In reactions in the presence of Mn2+, PrimPol demonstrated similar accuracy by incorporating dGMP on both templates A and 8-oxoA with 4- to 7-fold reduced efficiency (Table 1). The dGMP incorporation was observed as prominent ladders on DNA with A, 8-oxoA and 8-oxoG and is likely a result of indels following alternative alignments with short +2-+3 CC template microhomology region.
2.3. The Effect of DNA Sequence Context on A and 8-oxoA Bypass
To test the effect of DNA sequence context on nucleotide incorporation, we replaced C in the +2 position of the HRAS oncogene sequence with A, G or T. All replacements increased the accuracy of PrimPol and reduced the incorporation of dGMP on DNA substrates with template undamaged A or 8-oxoA lesion in the presence of Mg2+ (Figure 3A,B). In reaction with Mn2+, PrimPol also incorporated dGMP on DNA templates with A and 8-oxoA in the HRAS 5`-CCXAG-3` sequence context only (Figure 3C,D). Moreover, PrimPol incorporated dCMP on both A- and 8-oxoA-containing DNA substrates after replacement +2 template C with G. Altogether, these data suggest that PrimPol carries out efficient and accurate DNA synthesis across 8-oxoA. PrimPol also induce microdeletions with low efficiency in the sequence-dependent manner.
3. Discussion
8-oxoG and 8-oxoA are the most common lesions caused by reactive oxygen species. 8-oxoG is ambiguously read by DNA polymerases leading to mutations after the next round of replication rather than showing blocking effects. The available evidence suggests that 8-oxoA in mammalian cells also has a moderate mutagenic potential and induces A:T → C:G transversions and A:T → G:C transitions in the HRAS 5`-CCXAG-3` sequence [25,26,27].
In contrast to 8-oxoG, the activity of eukaryotic DNA polymerases opposite 8-oxoA remains poorly characterized. Only three DNA polymerases, namely Pol α, Pol β and Pol η, have been studied to date [25,29,30]. These DNA polymerases preferentially incorporated opposite 8-oxoA dTMP and small amounts of dGMP. Pol α carried out quite accurate DNA synthesis by incorporating complementary dTMP 10-fold more efficiently compared to non-complementary dGMP [29], while translesion Pol η incorporated dTMP and dGMP with almost similar efficiency [30]. Accuracy of Pol β varied from 4- to 18-fold preference for dTMP over dGMP on DNA substrates with different sequence context [29,30].
Human PrimPol is a unique DNA primase involved in DNA damage tolerance pathways both in nuclei and mitochondria [1,16,31]. PrimPol can encounter DNA lesions during repriming events and also possesses the DNA translesion activity. PrimPol is also known by template scrunching activity which can generate small deletions [28,32]. This activity is stimulated by Mn2+ and DNA lesions (lesion skipping mechanism) [12,28].
In this work, we studied PrimPol bypass of 8-oxopurine lesions in the HRAS sequence context. We demonstrated that PrimPol bypasses 8-oxoA with high efficiency and relatively high fidelity. PrimPol preferentially incorporated complementary dTMP opposite 8-oxoA in reactions in the presence of both Mg2+ and Mn2+ cofactors. PrimPol also incorporated dGMP on DNA substrates with 8-oxoA with 3- to 4-fold reduced efficiency compared to the complementary dTMP. Unlike other DNA polymerases, dGMP-incorporation by PrimPol was observed on DNA with both 8-oxoA and undamaged A. It was sequence-dependent and was stimulated by short CC nucleotide repeats in the HRAS CCXAG sequence. Replacement of C in the +2 template position abrogated the dGMP incorporation suggesting that it is mediated by template scrunching mechanism and causes deletions.
The error-prone dGMP incorporation was observed in reactions with both Me2+ cofactors but the efficiency of dGMP incorporation was higher with Mn2+. In particular, PrimPol incorporated dGMP on DNA substrate with 8-oxoA ~ 3-fold less efficient compared to undamaged A in the presence of Mg2+ ions and with equal efficiencies for both templates in reactions with Mn2+. Another type of errors – dCMP incorporation on DNA templates with A or 8-oxoA – was exclusively stimulated by Mn2+ ions and was observed only in reactions with the CGXAG sequence context and guided by G in the +2 templates position. Altogether, these data are in agreement that PrimPol misincorporates nucleotides in a sequence-dependent manner utilizing the Mn2+-stimulated template scrunching mechanism which is not specific to the 8-oxoA lesion. Our data also suggest that the rate of PrimPol-mediated errors is relatively low in all tested sequence contexts and PrimPol unlikely contributes to the 8-oxoA-induced mutagenesis in living cells.
The 5`-flanking nucleotide near 8-oxoG affects the accuracy of TLS by Pol η [33]. Interestingly, in our work (in the HRAS oncogene sequence context 3'-A8-oxoGC-5'), PrimPol demonstrated lower accuracy on DNA with 8-oxoG than in previous studies and incorporated dCMP and dAMP with almost equal efficiencies. In contrast, PrimPol incorporated dCMP almost exclusively or about 6- to 8-fold more efficiently than non-complementary dAMP in other sequence contexts such as 3'-C8-oxoGT-5' [1], 3'-G8-oxoGC-5' [34], 3'-G8-oxoGA-5' [12] and 3'-G8-oxoGT-5' [16]. Crystallographic PrimPol studies demonstrated that 8-oxoG in DNA containing the 3'-C8-oxoGA-5' sequence in complex with both incoming dCTP and dATP adopts the anti or syn conformation, respectively, without significant structural hindrance within the active site which supports the relatively low accuracy of PrimPol opposite 8-oxoG [35]. Our results corroborate these observations. It is possible that stacking interactions of 8-oxoG with flanking nucleobases contribute to its positioning in the anti or syn conformation.
Since PrimPol efficiently bypasses several DNA lesions including 8-oxoA, 8-oxoG, 5-fU and thymine glycol, we suggest that PrimPol carries out efficient TLS across a wide range of DNA lesions caused by oxidative stress. Indeed, PrimPol attenuates response of A549 cells to oxidative damage [36]. Also PrimPol as a component of the MUS81-LIG4 axis takes part in replication fork restart during transcription-dependent replication stress under excessive reactive oxygen species action [37]. Recent study demonstrated that PrimPol can contribute to SBS-A mutational signature resembling the mutagenic effect of 8-oxoG due to PrimPol ability to bypass oxidized damage [38].
4. Materials and Methods
4.1. DNA Templates and Enzymes
PrimPol was purified from E. coli as described [15]. DNA oligonucleotides used in the study (Table 1) were synthetized as described previously [39]. 8-oxoA and 8-oxoG lesions were placed in the sequence context similar with the HRAS CCXAG mutagenesis hot spot [25] in TemplateXA. Templates TemplateXG, TemplateXT and TemplateXC differ from TemplateXA by the single substitution at the +2 position. To prepare DNA substrates, the 5’-Cy5 labeled primer Pr18-Cy5 of 32P-labelled primer Pr18 was annealed to the corresponding unlabeled template oligonucleotides at a molar ratio of 1:1.1 in 100 mM NaCl by heating to 97°C and slowly cooling to 4°C.

Table 1.
Oligonucleotides used in the study.
| Oligonucleotide | Sequence 5`-3` |
|---|---|
| Pr18-Cy5 | Сy5-AGGGCAGAGTATTCTTCT |
| Pr18 | AGGGCAGAGTATTCTTCT |
| TemplateXA | TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTATACCGCAGGCAXAGAAGAATACTCTGCCCT |
| TemplateXG | TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTATACCGCAGGCGXAGAAGAATACTCTGCCCT |
| TemplateXT | TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTATACCGCAGGCTXAGAAGAATACTCTGCCCT |
| TemplateXC | TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTATACCGCAGGCCXAGAAGAATACTCTGCCCT |
X = A, 8-oxoA, G or 8-oxoG.
4.2. DNA Polymerase Reactions for the Primer Extension Assay
Primer extension reactions were performed in 20 µl containing 100 nM DNA substrate, 200 μM dNTP, 30 mM HEPES рН 7.0, 10 mM MgCl2 or 1 mM MnCl2, 100 µg/ml BSA, 1 mM DTT, 4% glycerol and 100 – 200 nM PrimPol. Reactions were incubated at 37°C for 1 – 4 min, stopped by the addition of an equal volume of 2х loading buffer (20 mM EDTA, 0.001% bromophenol blue, 96% formamide) and heated for 5 min at 95°С. The reaction products were resolved on 21% polyacrylamide gels with 8 M urea, visualized on Typhoon 9400 (GE Healthcare, USA) and analyzed with ImageQuant software. All experiments were repeated three times. The percent of the extended primer (PrExt) was calculated for each reaction and the mean values of PrExt with the standard errors are shown in figures.
4.3. Steady-State Kinetics Analysis of dNMP Incorporation
To quantify the incorporation of individual dNMP opposite DNA lesions, we varied the dNTP concentration from 2.5 to 6000 μM in reactions in the presence of 10 nM PrimPol and 10 mM MgCl2 and from 0.25 to 1000 µM in reactions with 5 nM PrimPol and 1 mM MnCl2. Depending on the lesion, the reactions were incubated for 3–20 min with MgCl2 and for 1–10 min with MnCl2 to ensure that less than 40% of the primer is utilized. Calculations were made using GraFit software (Erithacus Soft-ware, UK). The data were fit to the Michaelis-Menten equation V = Vmax×[dNTP])/(KM + [dNTP]), where V and Vmax are the observed and the maximum rates of the reaction (in percentages of utilized primer per minute), respectively, and KM is the apparent Michaelis constant.
Author Contributions
methodology E.O.B., A.A.K., P.N.K., investigation, E.O.B., A.A.K., P.N.K..; resources, A.V.M., A.V.A..; data curation, E.O.B., A.A.K.; writing—original draft preparation, A.V.M..; writing—review and editing, A.V.A.; visualization, E.O.B..; supervision, A.V.M., A.V.A.; funding acquisition, A.V.M.
Funding
This research was funded by Russian Scientific Foundation, grant number 23-14-00209.
Data Availability Statement
All data are available under request.
Acknowledgments
We thank Timofei Zatsepin for oligonucleotide synthesis.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 8-oxoA | 8-oxoadenine |
| 8-oxoG | 8-oxoguanine |
References
- García-Gómez, S.; Reyes, A.; Martínez-Jiménez, M.I.; Chocrón, S.; Mourón, S.; Terrados, G.; Powell, C.; Salido, E.; Méndez, J.; Holt, I.J.; et al. PrimPol, an Archaic Primase/Polymerase Operating in Human Cells. Mol. Cell 2013, 52, 541–553. [Google Scholar] [CrossRef]
- Bianchi, J.; Rudd, S.G.; Jozwiakowski, S.K.; Bailey, L.J.; Soura, V.; Taylor, E.; Stevanovic, I.; Green, A.J.; Stracker, T.H.; Lindsay, H.D.; et al. Primpol Bypasses UV Photoproducts during Eukaryotic Chromosomal DNA Replication. Mol. Cell 2013, 52, 566–573. [Google Scholar] [CrossRef]
- Mourón, S.; Rodriguez-Acebes, S.; Martínez-Jiménez, M.I.; García-Gómez, S.; Chocrón, S.; Blanco, L.; Méndez, J. Repriming of DNA Synthesis at Stalled Replication Forks by Human PrimPol. Nat. Struct. Mol. Biol. 2013, 20, 1383–1389. [Google Scholar] [CrossRef]
- Piberger, A.L.; Bowry, A.; Kelly, R.; Walker, A.K.; Gonzalez, D.; Bailey, L.J.; Doherty, A.J.; Mendez, J.; Morris, J.R.; Bryant, H.E.; et al. PrimPol-Dependent Single-Stranded Gap Formation Mediates Homologous Recombination at Bulky DNA Adducts. Nat. Commun. 2020, 11, 5863. [Google Scholar] [CrossRef]
- González-Acosta, D.; Blanco-Romero, E.; Ubieto-Capella, P.; Mutreja, K.; Míguez, S.; Llanos, S.; García, F.; Muñoz, J.; Blanco, L.; Lopes, M.; et al. PrimPol-mediated Repriming Facilitates Replication Traverse of DNA Interstrand Crosslinks. EMBO J. 2021, 40, e106355. [Google Scholar] [CrossRef]
- Schiavone, D.; Jozwiakowski, S.K.; Romanello, M.; Guilbaud, G.; Guilliam, T.A.; Bailey, L.J.; Sale, J.E.; Doherty, A.J. PrimPol Is Required for Replicative Tolerance of G Quadruplexes in Vertebrate Cells. Mol. Cell 2016, 61, 161–169. [Google Scholar] [CrossRef]
- Li, T.; Tang, L.; Kou, H.; Wang, F. Primpol Competes with Rad51 to Resolve G-Quadruplexinduced Replication Stress via Its Interaction with Rpa. Acta Biochim. Biophys. Sin. (Shanghai). 2023, 55, 498–507. [Google Scholar] [CrossRef]
- Butler, T.J.; Estep, K.N.; Sommers, J.A.; Maul, R.W.; Moore, A.Z.; Bandinelli, S.; Cucca, F.; Tuke, M.A.; Wood, A.R.; Bharti, S.K.; et al. Mitochondrial Genetic Variation Is Enriched in G-Quadruplex Regions That Stall DNA Synthesis in Vitro. Hum Mol Genet. 2020, 29, 1292–1309. [Google Scholar] [CrossRef]
- Šviković, S.; Crisp, A.; Tan-Wong, S.M.; Guilliam, T.A.; Doherty, A.J.; Proudfoot, N.J.; Guilbaud, G.; Sale, J.E. R-loop Formation during S Phase Is Restricted by PrimPol-mediated Repriming. EMBO J. 2019, 38, e99793. [Google Scholar] [CrossRef]
- Kobayashi, K.; Guilliam, T.A.; Tsuda, M.; Yamamoto, J.; Bailey, L.J.; Iwai, S.; Takeda, S.; Doherty, A.J.; Hirota, K. Repriming by PrimPol Is Critical for DNA Replication Restart Downstream of Lesions and Chain-Terminating Nucleosides. Cell Cycle 2016, 15, 1997–2008. [Google Scholar] [CrossRef]
- Bailey, L.J.; Bianchi, J.; Doherty, A.J. PrimPol Is Required for the Maintenance of Efficient Nuclear and Mitochondrial DNA Replication in Human Cells. Nucleic Acids Res. 2019, 47, 4026–4038. [Google Scholar] [CrossRef]
- Makarova, A.V.; Boldinova, E.O.; Belousova, E.A.; Lavrik, O.I. In Vitro Lesion Bypass by Human PrimPol. DNA Repair (Amst). 2018, 70, 18–24. [Google Scholar] [CrossRef]
- Boldinova, E.O.; Ghodke, P.P.; Sudhakar, S.; Mishra, V.K.; Manukyan, A.A.; Miropolskaya, N.; Pradeepkumar, P.I.; Makarova, A. V Translesion Synthesis across the N2-Ethyl-Deoxyguanosine Adduct by Human PrimPol. ACS Chem. Biol. 2022, 17, 3238–3250. [Google Scholar] [CrossRef]
- Yudkina, A.V.; Barmatov, A.E.; Bulgakov, N.A.; Boldinova, E.O.; Shilkin, E.S.; Makarova, A.V.; Zharkov, D.O. Bypass of Abasic Site–Peptide Cross-Links by Human Repair and Translesion DNA Polymerases. Int. J. Mol. Sci. 2023, 24, 10877. [Google Scholar] [CrossRef]
- Boldinova, E.O.; Yudkina, A.V.; Shilkin, E.S.; Gagarinskaya, D.I.; Baranovskiy, A.G.; Tahirov, T.H.; Zharkov, D.O.; Makarova, A.V. Translesion Activity of PrimPol on DNA with Cisplatin and DNA-Protein Cross-Links. Sci. Rep. 2021, 11, 17588. [Google Scholar] [CrossRef]
- Stojkovic, G.; Makarova, A.V.; Wanrooij, P.H.; Forslund, J.; Burgers, P.M.J.; Wanrooij, S. Oxidative DNA Damage Stalls the Human Mitochondrial Replisome. Sci. Rep. 2016, 6, 28942. [Google Scholar] [CrossRef]
- Helbock, H.J.; Beckman, K.B.; Shigenaga, M.K.; Walter, P.B.; Woodall, A.A.; Yeo, H.C.; Ames, B.N. DNA Oxidation Matters: The HPLC-Electrochemical Detection Assay of 8-Oxo-Deoxyguanosine and 8-Oxo-Guanine. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 288–293. [Google Scholar] [CrossRef]
- Møller, P.; Cooke, M.S.; Collins, A.; Olinski, R.; Rozalski, R.; Loft, S. Harmonising Measurements of 8-Oxo-7,8-Dihydro-2′-Deoxyguanosine in Cellular DNA and Urine. Free Radic. Res. 2012, 46, 541–553. [Google Scholar] [CrossRef]
- Fuciarelli, A.F.; Wegher, B.J.; Gajewski, E.; Dizdaroglu, M.; Blakely, W.F. Quantitative Measurement of Radiation-Induced Base Products in DNA Using Gas Chromatography-Mass Spectrometry. Radiat. Res. 1989, 119, 219–231. [Google Scholar] [CrossRef]
- Tuo, J.; Jaruga, P.; Rodriguez, H.; Dizdaroglu, M.; Bohr, V.A. The Cockayne Syndrome Group B Gene Product Is Involved in Cellular Repair of 8-Hydroxyadenine in DNA. J. Biol. Chem. 2002, 277, 30832–30837. [Google Scholar] [CrossRef]
- Leonard, G.A.; Brown, T.; Guy, A.; Téoule, R.; Hunter, W.N. Conformation of Guanine-8-Oxoadenine Base Pairs in the Crystal Structure of d(CGCGAATT(08A)GCG). Biochemistry 1992, 31, 8415–8420. [Google Scholar] [CrossRef]
- Yudkina, A.V.; Shilkin, E.S.; Endutkin, A.V.; Makarova, A.V.; Zharkov, D.O. Reading and Misreading 8-Oxoguanine, a Paradigmatic Ambiguous Nucleobase. Crystals 2019, 9, 269. [Google Scholar] [CrossRef]
- Moriya, M. Single-Stranded Shuttle Phagemid for Mutagenesis Studies in Mammalian Cells: 8-Oxoguanine in DNA Induces Targeted G·C → T·A Transversions in Simian Kidney Cells. Proc. Natl. Acad. Sci. U. S. A. 1993, 90, 1122–1126. [Google Scholar] [CrossRef]
- Wood, M.L.; Esteve, A.; Morningstar, M.L.; Kuziemko, G.M.; Essigmann, J.M. Genetic Effects of Oxidative DNA Damage: Comparative Mutagenesis of 7,8-Dihydro-8-Oxoguanine and 7,8-Dihydro-8-Oxoadenine in Escherichia Coli. Nucleic Acids Res. 1992, 20, 6023–6032. [Google Scholar] [CrossRef]
- Kamiya, H.; Miura, H.; Murata-kamiya, N.; Ishikawa, H.; Sakaguchi, T.; Inoue, H.; Sasaki, T.; Masutanl, C.; Hanaoka, F.; Nishimura, S.; et al. 8-Hydroxyadenine (7, 8-Dihydro-8-Oxoadenine) Induces Misincorporation in in Vitro DNA Synthesis and Mutations in NIH 3t3 Cells. Nucleic Acids Res. 1995, 23, 2893–2899. [Google Scholar] [CrossRef]
- Kamiya, H.; Murata-kamiya, N.; Koizume, S.; Inoue, H.; Nishimura, S.; Ohtsuka, E. 8-Hydroxyguanine (7,8-Dihydro-8-Oxoguanine) in Hot Spots of the c-Ha-Ras Gene: Effects of Sequence Contexts on Mutation Spectra. Carcinogenesis 1995, 16, 883–889. [Google Scholar] [CrossRef]
- Tan, X.; Grollman, A.P.; Shibutani, S. Comparison of the Mutagenic Properties of 8-Oxo-7,8-Dihydro-2’-Deoxyadenosine and 8-Oxo-7,8-Dihydro-2’-Deoxyguanosine DNA Lesions in Mammalian Cells. Carcinogenesis 1999, 20, 2287–2292. [Google Scholar] [CrossRef]
- Martínez-Jiménez, M.I.; García-Gómez, S.; Bebenek, K.; Sastre-Moreno, G.; Calvo, P.A.; Díaz-Talavera, A.; Kunkel, T.A.; Blanco, L. Alternative Solutions and New Scenarios for Translesion DNA Synthesis by Human PrimPol. DNA Repair (Amst). 2015, 29, 127–138. [Google Scholar] [CrossRef]
- Shibutani, S.; Bodepudi, V.; Johnson, F.; Grollman, A.P. Translesional Synthesis on DNA Templates Containing 8-Oxo-7,8-Dihydrodeoxyadenosine. Biochemistry 1993, 32, 4615–4621. [Google Scholar] [CrossRef]
- Koag, M.C.; Jung, H.; Lee, S. Mutagenic Replication of the Major Oxidative Adenine Lesion 7,8-Dihydro-8-Oxoadenine by Human DNA Polymerases. J. Am. Chem. Soc. 2019, 141, 4584–4596. [Google Scholar] [CrossRef]
- Torregrosa-Muñumer, R.; Forslund, J.; Goffart, S.; Pfeiffer, A.; Stojkovic, G.; Carvalho, G.; Al-Furoukh, N.; Blanco, L.; Wanrooij, S.; Pohjoismäki, J.L.O. PrimPol Is Required for Replication Reinitiation after MtDNA Damage. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, 11398–11403. [Google Scholar] [CrossRef]
- Guilliam, T.A.; Jozwiakowski, S.K.; Ehlinger, A.; Barnes, R.P.; Rudd, S.G.; Bailey, L.J.; Skehel, J.M.; Eckert, K.A.; Chazin, W.J.; Doherty, A.J. Human PrimPol Is a Highly Error-Prone Polymerase Regulated by Single-Stranded DNA Binding Proteins. Nucleic Acids Res. 2015, 43, 1056–1068. [Google Scholar] [CrossRef]
- Yung, C.; Suzuki, T.; Okugawa, Y.; Kawakami, A.; Loakes, D.; Negishi, K.; Negishi, T. Nucleotide Incorporation against 7,8-Dihydro-8-Oxoguanine Is Influenced by Neighboring Base Sequences in TLS DNA Polymerase Reaction. In Proceedings of the Nucleic acids symposium series; 2007; pp. 49–50. [Google Scholar]
- Zafar, M.K.; Ketkar, A.; Lodeiro, M.F.; Cameron, C.E.; Eoff, R.L. Kinetic Analysis of Human PrimPol DNA Polymerase Activity Reveals a Generally Error-Prone Enzyme Capable of Accurately Bypassing 7,8-Dihydro-8-Oxo-2′-Deoxyguanosine. Biochemistry 2014, 53, 6584–6594. [Google Scholar] [CrossRef]
- Rechkoblit, O.; Johnson, R.E.; Gupta, Y.K.; Prakash, L.; Prakash, S.; Aggarwal, A.K. Structural Basis of DNA Synthesis Opposite 8-Oxoguanine by Human PrimPol Primase-Polymerase. Nat. Commun. 2021, 12, 4020. [Google Scholar] [CrossRef]
- Gromova, A.S.; Boldinova, E.O.; Kim, D.V.; Chuprov-Netochin, R.N.; Leonov, S.V.; Pustovalova, M.V.; Zharkov, D.O.; Makarova, A.V. Response of PRIMPOL-Knockout Human Lung Adenocarcinoma A549 Cells to Genotoxic Stress. Biochem. 2023, 88, 1933–1943. [Google Scholar] [CrossRef]
- Andrs, M.; Stoy, H.; Boleslavska, B.; Chappidi, N.; Kanagaraj, R.; Nascakova, Z.; Menon, S.; Rao, S.; Oravetzova, A.; Dobrovolna, J.; et al. Excessive Reactive Oxygen Species Induce Transcription-Dependent Replication Stress. Nat. Commun. 2023, 14. [Google Scholar] [CrossRef]
- Gyüre, Z.; Póti, Á.; Németh, E.; Szikriszt, B.; Lózsa, R.; Krawczyk, M.; Richardson, A.L.; Szüts, D. Spontaneous Mutagenesis in Human Cells Is Controlled by REV1-Polymerase ζ and PRIMPOL. Cell Rep. 2023, 42, 112887. [Google Scholar] [CrossRef]
- Petushkov, I.V.; Aralov, A.V.; Ivanov, I.A.; Baranov, M.S.; Zatsepin, T.S.; Kulbachinskiy, A.V. Effect of 8-Oxo-1,N6-Ethenoadenine Derivatives on the Activity of RNA Polymerases from SARS-CoV-2 and Escherichia Coli. Biochemistry. (Mosc). 2024, 89, 2263–2273. [Google Scholar] [CrossRef]
Figure 1.
DNA polymerase activity of PrimPol on DNA with 8-oxoA or 8-oxoG in the presence of Mg2+. (a). Primer extension reactions on DNA containing 8-oxoA or 8-oxoG with Mg2+. Reactions were carried out in the presence of 10 mM MgCl2, 200 nM PrimPol, 100 nM Cy5-DNA-substrate and 200 μM of all four dNTP (N) or individual nucleotide substrates (A – dATP, G – dGTP, T – dTTP, C – dCTP) for 4 min. (b). Diagram showing the percent of primer extension on DNA with 8-oxoA or 8-oxoG in reactions in the presence of Mg2+ (Figure 1A). The mean values of primer extension and standard errors are indicated. (c). Diagram showing the VMAX/KM ratio calculated for DNA containing 8-oxoA or 8-oxoG in reactions in the presence of Mg2+ (Table 1).
Figure 1.
DNA polymerase activity of PrimPol on DNA with 8-oxoA or 8-oxoG in the presence of Mg2+. (a). Primer extension reactions on DNA containing 8-oxoA or 8-oxoG with Mg2+. Reactions were carried out in the presence of 10 mM MgCl2, 200 nM PrimPol, 100 nM Cy5-DNA-substrate and 200 μM of all four dNTP (N) or individual nucleotide substrates (A – dATP, G – dGTP, T – dTTP, C – dCTP) for 4 min. (b). Diagram showing the percent of primer extension on DNA with 8-oxoA or 8-oxoG in reactions in the presence of Mg2+ (Figure 1A). The mean values of primer extension and standard errors are indicated. (c). Diagram showing the VMAX/KM ratio calculated for DNA containing 8-oxoA or 8-oxoG in reactions in the presence of Mg2+ (Table 1).
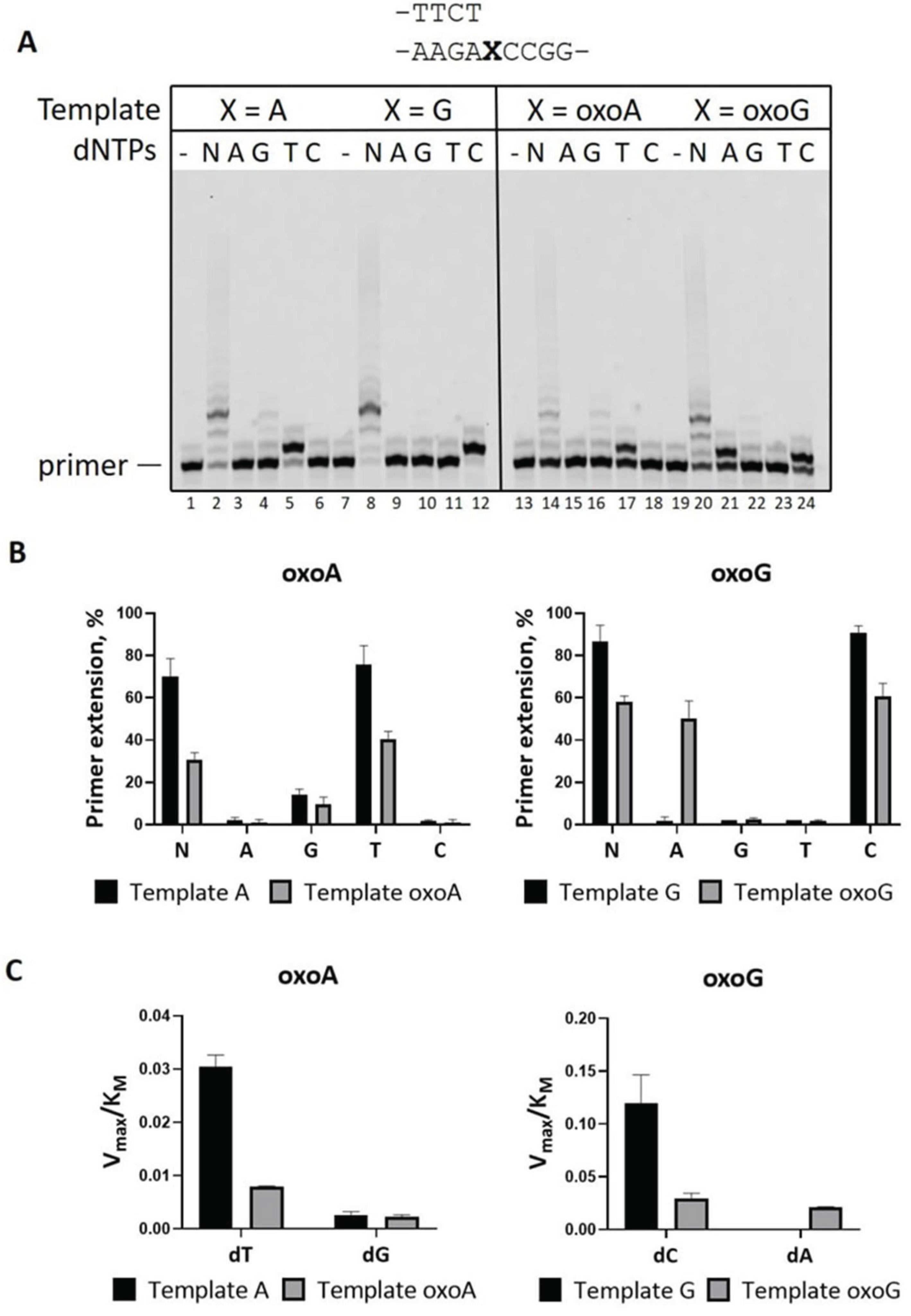
Figure 2.
DNA polymerase activity of PrimPol on DNA with 8-oxoA or 8-oxoG in the presence of Mn2+. (a). Primer extension reactions on DNA containing 8-oxoA or 8-oxoG with Mn2+. Reactions were carried out in the presence of 1 mM MnCl2, 200 nM PrimPol, 100 nM Cy5-DNA-substrate and 200 μM of all four dNTP (N) or individual nucleotide substrates (A – dATP, G – dGTP, T – dTTP, C – dCTP) for 1 min. (b). Diagram showing the percent of primer extension on DNA with 8-oxoA or 8-oxoG in reactions in the presence of Mn2+ (Figure 2A). The mean values of primer extension and standard errors are indicated. (c). Diagram showing the VMAX/KM ratio calculated for DNA containing 8-oxoA or 8-oxoG in reactions with Mn2+ (Table 1).
Figure 2.
DNA polymerase activity of PrimPol on DNA with 8-oxoA or 8-oxoG in the presence of Mn2+. (a). Primer extension reactions on DNA containing 8-oxoA or 8-oxoG with Mn2+. Reactions were carried out in the presence of 1 mM MnCl2, 200 nM PrimPol, 100 nM Cy5-DNA-substrate and 200 μM of all four dNTP (N) or individual nucleotide substrates (A – dATP, G – dGTP, T – dTTP, C – dCTP) for 1 min. (b). Diagram showing the percent of primer extension on DNA with 8-oxoA or 8-oxoG in reactions in the presence of Mn2+ (Figure 2A). The mean values of primer extension and standard errors are indicated. (c). Diagram showing the VMAX/KM ratio calculated for DNA containing 8-oxoA or 8-oxoG in reactions with Mn2+ (Table 1).
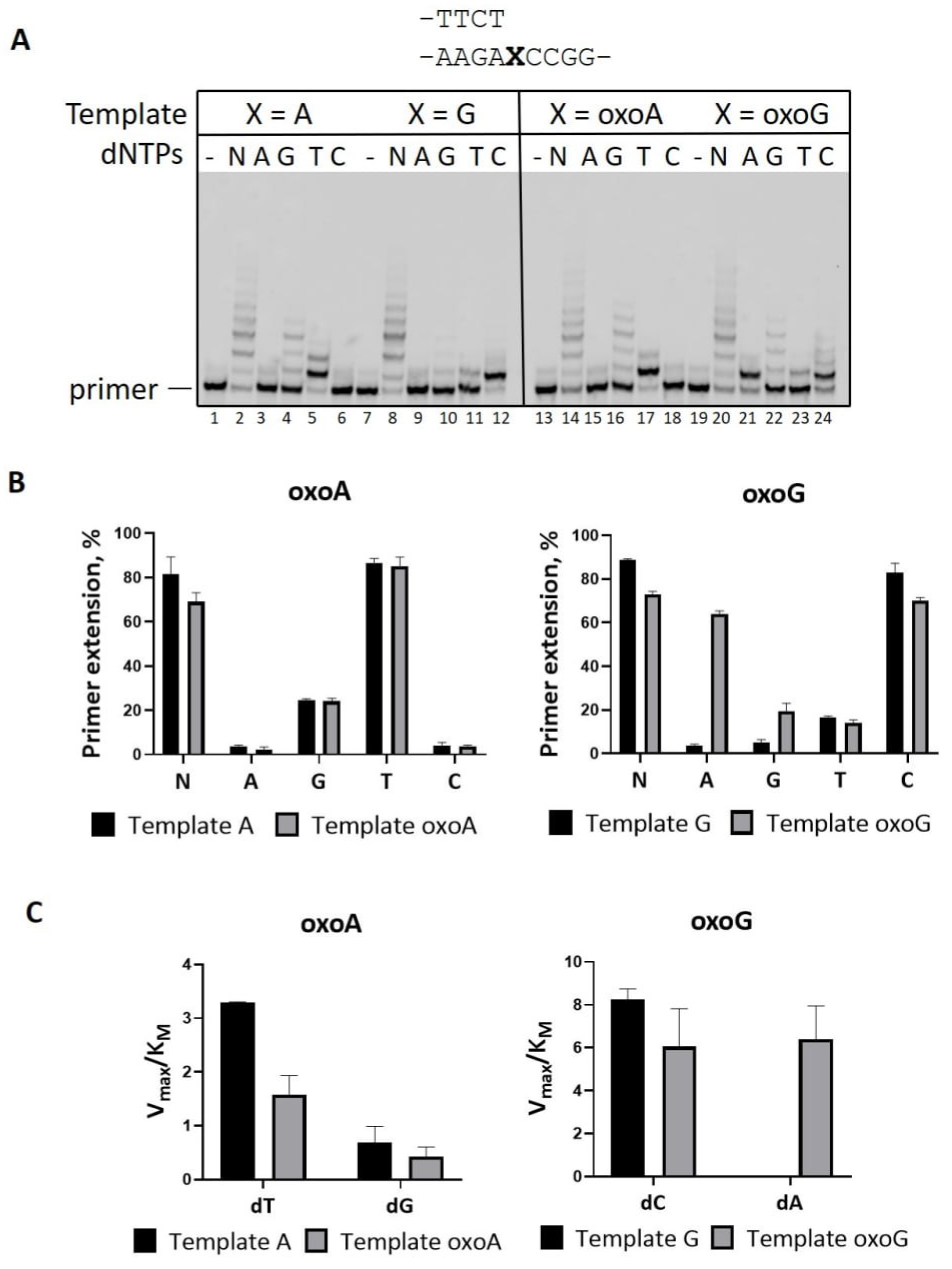
Figure 3.
DNA polymerase activity of PrimPol on DNA with A or 8-oxoA in different sequence context. (a). Primer extension reactions on DNA containing A or 8-oxoA with Mg2+. DNA substrates contain A, G, T or C at the +2 position of the template (N). Reactions were carried out in the presence of 10 mM MgCl2, 200 nM PrimPol, 100 nM Cy5-DNA-substrate and 200 μM of all four dNTP (N) or individual nucleotide substrates (A – dATP, G – dGTP, T – dTTP, C – dCTP) for 4 min. (c). Primer extension reactions on DNA containing A or 8-oxoA with Mn2+. DNA substrates contain A, G, T or C at the +2 position of the template. Reactions were carried out in the presence of 1 mM MnCl2, 200 nM PrimPol, 100 nM Cy5-DNA-substrate and 200 μM of all four dNTP (N) or individual nucleotide substrates (A – dATP, G – dGTP, T – dTTP, C – dCTP) for 1 min. (b, d). Diagrams showing the percent of primer extension on DNA with A or 8-oxoA in reactions in the presence of Mg2+ (Figure 3A) and Mn2+ (Figure 3C). The mean values of primer extension and standard errors are indicated.
Figure 3.
DNA polymerase activity of PrimPol on DNA with A or 8-oxoA in different sequence context. (a). Primer extension reactions on DNA containing A or 8-oxoA with Mg2+. DNA substrates contain A, G, T or C at the +2 position of the template (N). Reactions were carried out in the presence of 10 mM MgCl2, 200 nM PrimPol, 100 nM Cy5-DNA-substrate and 200 μM of all four dNTP (N) or individual nucleotide substrates (A – dATP, G – dGTP, T – dTTP, C – dCTP) for 4 min. (c). Primer extension reactions on DNA containing A or 8-oxoA with Mn2+. DNA substrates contain A, G, T or C at the +2 position of the template. Reactions were carried out in the presence of 1 mM MnCl2, 200 nM PrimPol, 100 nM Cy5-DNA-substrate and 200 μM of all four dNTP (N) or individual nucleotide substrates (A – dATP, G – dGTP, T – dTTP, C – dCTP) for 1 min. (b, d). Diagrams showing the percent of primer extension on DNA with A or 8-oxoA in reactions in the presence of Mg2+ (Figure 3A) and Mn2+ (Figure 3C). The mean values of primer extension and standard errors are indicated.
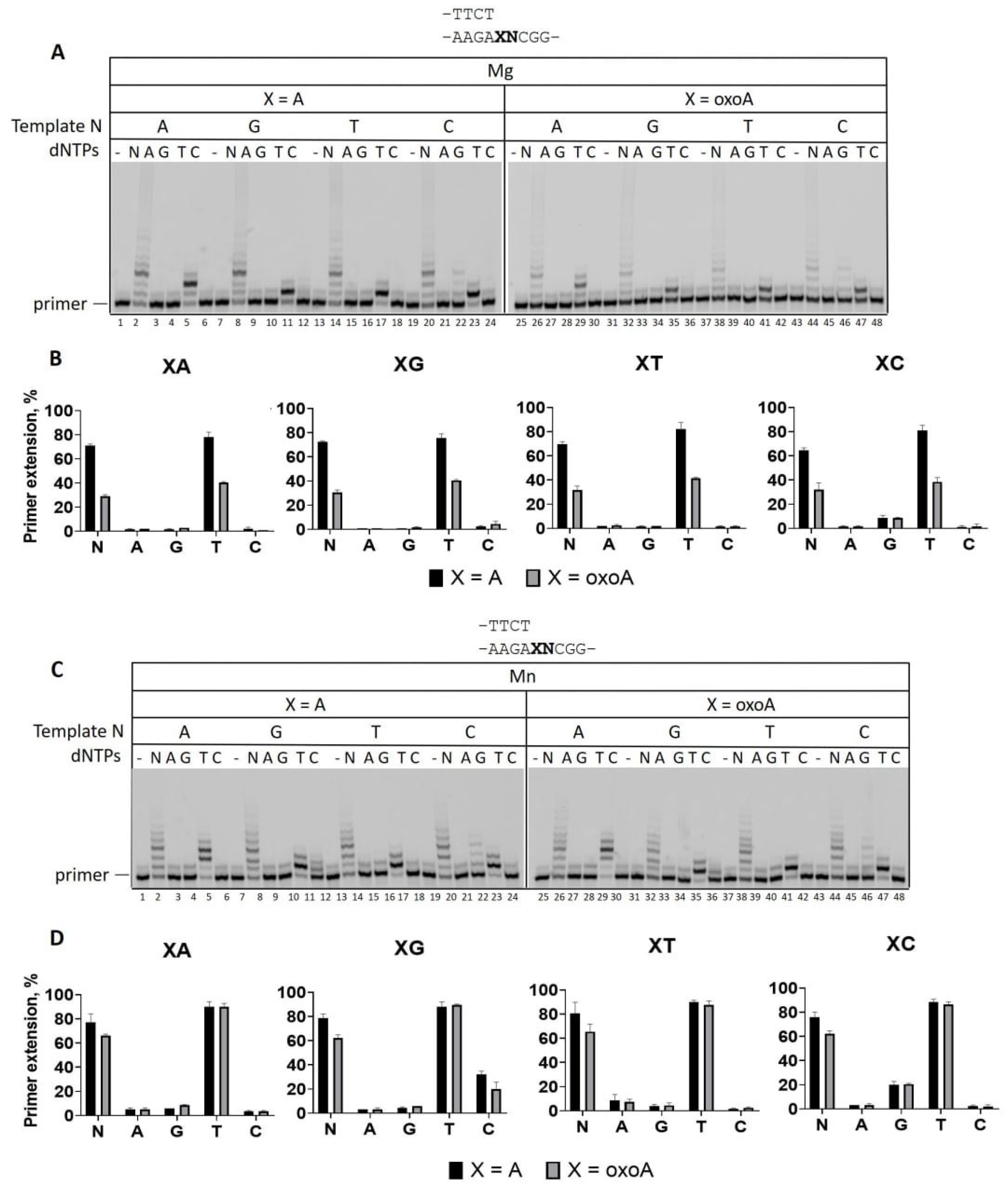
Table 1.
Steady-state kinetics analysis of dNMP incorporation opposite A, G, 8-oxoA and 8-oxoG.
| Template | dNMP | Vmax, % per min | KM, μM | Vmax/KM | Finc |
|---|---|---|---|---|---|
| Mg2+ | |||||
| Template A | dTradio | 12.9 ± 0.7 | 420 ± 37 | 0.031 ±0.003 | |
| dT | 12.2 ± 0.7 | 400 ± 42 | 0.031 ± 0.002 | 1 | |
| dG | 0.4 ± 0.004 | 184 ± 46 | 0.003 ± 0.0005 | 0.09 | |
| Template oxoA | dTradio | 3.6 ±0.3 | 610 ± 11 | 0.006 ±0.001 | |
| dT | 5.3 ± 0.1 | 677 ± 16 | 0.008 ± 0.0001 | 1 | |
| dG | 0.4 ± 0.005 | 182 ± 33 | 0.002 ± 0.0001 | 0.25 | |
| Template G | dC | 9 ± 0.9 | 73 ± 5 | 0.123 ± 0.019 | |
| dA | ND | ||||
| Template oxoG | dC | 3.8 ± 0.3 | 128 ± 3 | 0.029 ± 0.004 | 1 |
| dA | 2.9 ± 0.3 | 137 ± 9 | 0.021 ± 0.0005 | 0.72 | |
| Mn2+ | |||||
| Template A | dTradio | 50 ± 1.5 | 11 ± 1 | 4.5 ± 0.3 | |
| dT | 40 ± 0.9 | 12.2 ± 0.3 | 3.3 ± 0.01 | 1 | |
| dG | 2.7 ± 0.5 | 4 ± 0.3 | 0.7 ± 0.2 | 0.2 | |
| Template oxoA | dTradio | 28 ± 1.4 | 10 ± 0.5 | 2.7 ± 0.1 | |
| dT | 22.5 ± 0.02 | 14.6 ± 1.9 | 1.6 ± 0.2 | 1 | |
| dG | 2.4 ± 0.2 | 3.9 ± 0.8 | 0.4 ± 0.1 | 0.25 | |
| Template G | dC | 41.5 ± 0.1 | 5.1 ± 0.2 | 8.2 ± 0.3 | |
| dA | ND | ||||
| Template oxoG | dC | 23.5 ± 1 | 4 ± 0.5 | 6 ± 1 | 1 |
| dA | 15.9 ± 0.6 | 2.5 ± 0.3 | 6.4 ± 0.9 | 1.1 | |
dTradio – data calculated for 32P-labelled DNA substrate. Finc = Vmaxnon-complementary/KMnon-complementary/Vmaxcomplementary/KMcomplementary. *ND – not detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



