Submitted:
30 May 2025
Posted:
30 May 2025
You are already at the latest version
Abstract
Soil samples taken in the Puglia region, Southern Italy, around the entrance of dismissed bauxite mines at San Giovanni Rotondo, Otranto and Spinazzola, were assayed at 1% in Potato Dextrose Agar (PDA) and as liquid cultures in half-strength Potato Dextrose Broth against strains of Fusarium oxysporum f. sp. lycopersici (FOL), Sclerotinia minor (SM), Verticillium dahliae (VD) and Trichoderma harzianum (TH). In PDA experiments, all soil samples limited VD (21.9-32.2%), and SM (4.3-65.2%) or enhanced FOL (73.9-86.6%) radial growth, while the TH growth was differently altered. In solid media, soil samples lowered the FOL sporulation rate (23.53-62.75%), improved TH (5.4-109.3%) and VD (27.5-41.37%) conidia production, and had no effect on SM microsclerotia differentiation. In liquid cultures, almost all soil samples affected the mycelia biomass production, reduced VD (85-90.2%) and TH (20-70%) conidia production, and increased FOL (35-188%) sporulation. The SM strain did not produce microsclerotia in any liquid media tested. Soil samples in the liquid media also affected T. harzianum antagonistic activity mediated by secondary metabolites. Pearson correlation coefficient correlates these biological effects with pollutant total concentration, modifies contamination degree index and Nemerow integrated risk index. Data recommends soil surveys considering the impact on the biota, especially in sites of naturalistic, geological, and tourist interest.
Keywords:
radial growth
; sporulation
; biological activity
; toxic metals
; rare earth elements
; natural source
; risk of environmental contamination
1. Introduction
Dismissed mines are an important component of local cultural heritage worldwide. More ex-mining sites become museums or are equipped for educational and/or tourist purposes as diffused geoparks or archaeo-industrial sites [1]. An ever-increasing number of people visit refurbished old mines and are subject to the pilgrimage and recreative activities of curious people of all ages.
Soils around active or dismissed mines contain mainly toxic metals, Rare Earth Elements (REEs), eventually associated with polycyclic aromatic hydrocarbons (PAH), C10–C40 hydrocarbons and other organic contaminants [2]. Toxic metals, commonly indicated as “heavy metals”, include naturally occurring metals and semimetals of high molecular weight and density five times greater than the density of water, with potential toxicity to all forms of biota [3]. The toxic effects of toxic metals are associated with the production of free radicals in plants, animals and microbes, and carcinogenic action on humans [3]. At low concentrations, iron (Fe), zinc (Zn), copper (Cu), selenium (Se) and manganese (Mn) are essential for humans, animals, plants and microbe survival but can become toxic agents at higher concentrations. Arsenic (As), cadmium (Cd), lead (Pb), thallium (Tl), chromium (Cr), cadmium (Cd), nickel (Ni), aluminium (Al), bismuth (Bi), antimony (Sb), and mercury (Hg) serve no biological role [3]. Under specific concentrations and bioavailability, these metals induce toxicity by coordinating active sites (oxygen, nitrogen or sulfhydryl groups) of biological molecules, binding functional groups in proteins or replacing natural metal centres in different enzymes hindering their normal functioning in biochemical reactions. For instance, As blocks up to 200 enzymes, causing lipid peroxidation and defecting DNA synthesis [3,4,5]. Longstanding damage is associated with Cd, Hg and Pb accumulation in the body and the environment [4]. The acute and chronic harmfulness of toxic metals is influenced by their specific chemical properties and concentration, and the way of entry into the target organism [4,5]. They enter the plant along with water absorbed by root hairs or through stomata. Food and water ingestion, inhalation of air, permeation, and skin contact are the most common ways to come into humans and animals. In humans, some toxic metals are the cause of cancer, Alzheimer, Parkinson, bone mineralization, depression, vision and emotional disturbance, affecting DNA, RNA, reproductive system, and mental disorders in children [6]. Metals such as Cd, Pb, Ni, Co, Hg, Cr and Cu show severe toxicological effects on aquatic animals and microorganisms [7]. In plants, heavy metals interfere with photosynthesis reducing plant growth [8].
Toxic metals, associated with mining activities, soil smelters processing and manufacturing of metals and their sub-products or the use of sewage sludge in agriculture, affect microbiome in soil ecosystems [8,9,10]. Under opportune concentrations, toxic metals generate Reactive Oxygen Species, trigger lipid peroxidation, membrane and organelle damage and cell apoptosis [11] or affect mycorrhizal fungus functioning [12]. Metals like Cu, Cd, Mn, Pb, and Zn inhibit the growth of Aspergillus spp. and Sterigmatomyces halophillus [13], while Pb ions slows Aspergillus biennis growth rate [14], and Cd reduces Schizophyllum commune mycelial development [15]. In Neurospora crassa, Ni alters carbohydrate metabolism and induces membrane breaking [16].
The REEs consider the 15 lanthanides with atomic numbers 57–71, plus Scandium (Sc) and Yttrium (Y). These elements show similar chemical and physical properties related to an analogous external electronic configuration and are usually associated with the same deposits [17]. The impact of REEs on biota is not well defined. The REEs utilization as fertilizers, feed additives in fish and animal diets, and human medicine is well known and is further under development [18,19,20,21]. From soil, REEs are absorbed and bioaccumulated in plants [21,22,23] and can enter the food chain [19,20]. In humans, REEs exposure and accumulation induce toxicity [20,24,25,26,27,28,29], anti-angiogenic therapeutic activity [30], protective actions against bacteria, fungi and nematodes [31], hormonal and enzymatic activity stimulation, and immune system activation [20,28]. On various botanical species, REE exposure produces contradictory effects [32,33,34,35]. Isolates of Trichoderma atroviride, T. harzianum, Botrytis cinerea, Alternaria alternata, Fusarium solani, Rhizoctonia solani, and Sclerotinia sclerotiorum tolerated the presence of diverse REEs into the growing media [36]. Ganoderma lucidum and some Penicillium spp. were effective REE bio-sorbents [37,38]. Saccharomyces cerevisiae [37] and Kluyveromyces marxianus [39] accumulated neodymium (Nd). Pseudomonas aeruginosa built-up lanthanum (La), Erbium (Er), and Y, while Bacillus licheniformis, Bacillus subtilis, Brevibacterium helovolum and Rhodococcus erythropolis stored samarium (Sm) and other REEs [40]. The hormetic [41] behaviour of the toxic metals and REEs absorbed and accumulated in biological tissues is evident and well elucidated.
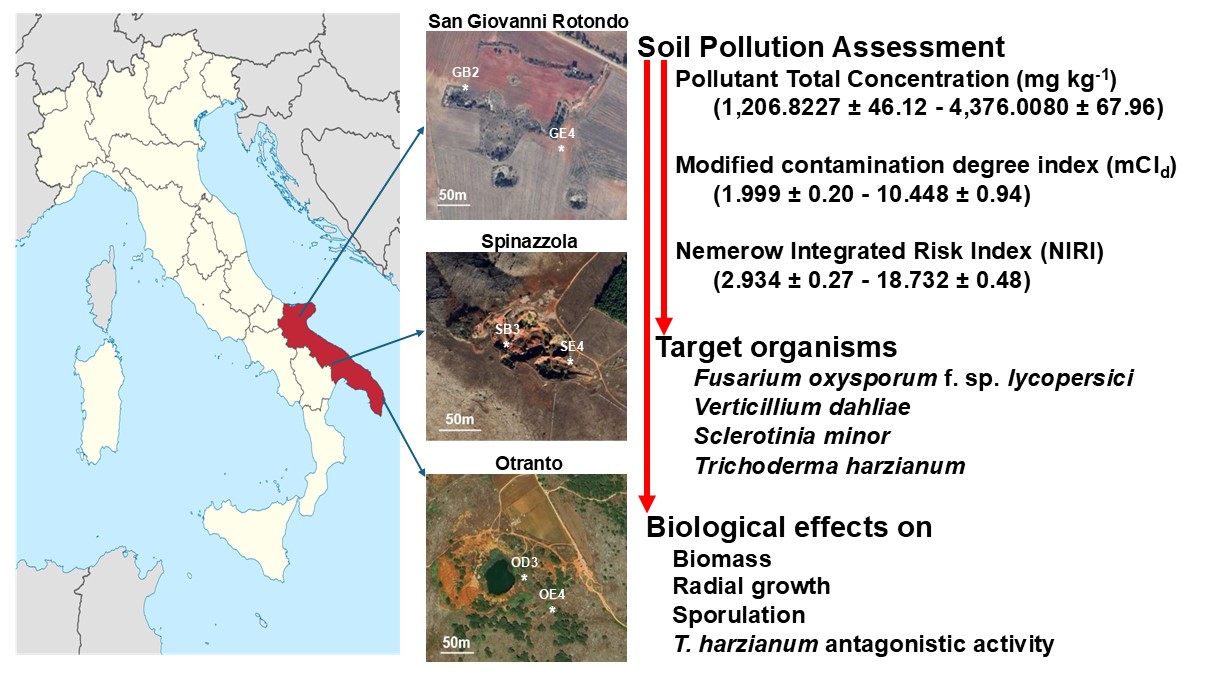
Most of the studies so far have been conducted on the effects triggered by REE and toxic metals exposures are based on experimental research under laboratory conditions with one element or their selected mixtures applying arbitrary concentrations, sometimes distinct from those found in natural systems [23,32,34,42]. The location around the source, precipitation, temperature, wind speed and direction, atmospheric relative humidity and composition affect biological effects and play a key role in exposure to environmental contaminants. After all, the ultimate harmful impact in bioassay shifts with the appropriateness of the target organism evaluated. These issues should be considered while monitoring and identifying the toxic effects of chemicals related to biota safety in various environments [43]. Based on these considerations, this work evaluated the impacts of soil containing heavy metals and REEs on three phytopathogenic fungi and one Biological Control Agent (BCA) of plant pathogens. Soil samples collected around three dismissed bauxite mines in Puglia, characterized in major and minor elements composition [44,45,46,47,48] were used in liquid and solid media experiments to evaluate the effects on growth, and dissemination structures production among the four tested fungal species. The antagonistic activity of the BCA strain mediated by secondary metabolites was also analysed. The biological impact of tested soils on selected fungi was modelled alongside the distance from the mine entrance and total pollutant concentration.
2. Materials and Methods
2.1. Strains, Media, and Growth Conditions
Sclerotinia minor strain DiSSPA 9S (SM), Fusarium oxysporum f. sp. lycopersici strain DiSSPA 259F (FOL) and Verticillium dahliae strain DiSSPA 23V (VD) were selected as phytopathogens, while Trichoderma harzianum specie-complex strain DiSSPA TH2 (TH) was considered as BCA. The strains SM, FOL, and VD were stored on sterile soil according to Toussoun and Nelson [49], whereas TH on Potato Dextrose Agar (Oxoid Ltd. Hampshire UK; PDA) slant tubes at 5±1 °C in the Plant Pathology culture collection at the Department of Soil, Plant and Food Sciences (Di.S.S.P.A.) - University of Bari Aldo Moro. All the strains were revitalized and routinely grown on PDA at 25±1 °C in the dark.
2.2. Soil Sampling and Risk Assessment
Soil samples (Table 1) were collected around three dismissed bauxite mines located in the Puglia region, Southern Italy, at San Giovanni Rotondo, Otranto and Spinazzola as reported by Brouziotis et al. [48]. For each location, soil samples were obtained near [GB2 (N 41.638893 E 15.714657), OD3 (N 40.132358 E 18.501168), SB3 (N 40.98953 E 16.179596)] and far [GE2 (N 41° 38.284020 E 15° 42.954480), OE4 (N 40.13133 E 18.501572), SE4 (N 40.98962 E 16.179776)] from each mine entrance at 20 and 50 m respectively.
The Pollutant Total Concentration (PTC) was calculated as the sum of each elemental concentration.
As a tool for pollution assessment, at each sampling point, the modified contamination degree index (mCId) was defined as mCId = [Σ(Cf)/n] where n indicates the total number of contaminants analysed; Cf corresponds to the contamination factors of each element considered and is calculated as Cf = Ci/Bi where Ci denotes the measured concentrations of heavy metals, Bi represents the geochemical background value of the metals [50,51]. Based on mCId, the level of soil pollution was classified [51,52] unpolluted (mCId < 1.5), low (1.5 ≤ mCId <2), moderate (2 ≤ mCId <4), high (4 ≤ mCId <8), very high (8 ≤ mCId <16), extremely high (16 ≤ mCId <32), ultra-high (mCId >32).
To evaluate the potential ecological risks of heavy metals, the Nemerow Integrated Risk Index (NIRI) was calculated as where is the ecological risk index evaluated as using the Ci and Bi defined previously; and indicates the toxicity coefficients of Cd, Cu, As, Ni, Cr, Pb, and Zn which are 30, 5, 10, 5, 2, 5 and 1 respectively [51,53]. The ecological risks of heavy metal pollution were considered [51,53] low (NIRI < 40), moderate (40≤ NIRI < 80), considerable (80 ≤ NIRI < 160), high (160 ≤ NIRI < 320) and extreme (NIRI ≥ 320).
Soil samples were dried (60 °C, 24 h), ground and sieved in 2-mm mesh steel nets, to run pH measurements and bioassays.
For soil pH measurement, 10 g were put into a 25 mL beaker, 10 mL of distilled water (0.2 μS m-1 at 25 °C, pH 5.7) was added, and the mixture was stirred for 30 min and allowed to stand for 30 min. The pH value of the suspension was determined with a corrected GLP22 pH meter (Crison Instrument S.p.A., Carpi, MO, I).

Table 1.
Chemical composition (mg kg-1 dry mass) of soil collected at 20 (GB2, OD3, SB3) and 50m (GE2, OE4, SE4) the mines entrance a.
Table 1.
Chemical composition (mg kg-1 dry mass) of soil collected at 20 (GB2, OD3, SB3) and 50m (GE2, OE4, SE4) the mines entrance a.
| Elements | San Giovanni Rotondo [45,47,48] | Otranto [45,46,47] | Spinazzola [44,46,47] | |||
| GB2 | GE2 | OD3 | OE4 | SB3 | SE4 | |
| Arsenic (As) | bdl b | bdl | 44.300 ± 14.228 | 44.300 ± 14.228 | 56.909 ± 14.758 | 56.909 ± 14.758 |
| Barium (Ba) | 31.400 ± 8.683 | 31.400 ± 8.683 | 16.201 ± 1.436 | 16.201 ± 1.436 | 54.136 ± 17.651 | 54.136 ± 17.651 |
| Cobalt (Co) | 24.533 ± 8.975 | 24.533 ± 8.975 | 51.150 ± 3.319 | 51.150 ± 3.319 | 47.636 ± 28.954 | 47.636 ± 28.954 |
| Chromium (Cr) | 468.667 ± 10.257 | 468.667 ± 10.257 | 999.501 ± 64.521 | 999.501 ± 64.521 | 754.091 ± 23.661 | 754.091 ± 23.661 |
| Caesium (Cs) | bdl | bdl | bdl | bdl | 6.600 ± 1.795 | 6.600 ± 1.795 |
| Copper (Cu) | bdl | bdl | 55.501 ± 7.591 | 55.501 ± 7.591 | bdl | bdl |
| Gallium (Ga) | 56.200 ± 12.718 | 56.200 ± 12.718 | 58.351 ± 5.244 | 58.351 ± 5.244 | 65.864 ± 9.131 | 65.864 ± 9.131 |
| Hafnium (Hf) | bdl | bdl | 12.147 ± 0.587 | 12.147 ± 0.587 | bdl | bdl |
| Molybdenum (Mo) | bdl | bdl | 7.051 ± 0.825 | 7.051 ± 0.825 | bdl | bdl |
| Niobium (Nb) | 81.800 ± 19.818 | 81.800 ± 19.818 | 75.101 ± 3.582 | 75.101 ± 3.582 | 112.727 ± 10.864 | 112.727 ± 10.864 |
| Nickel (Ni) | 197.333 ± 25.765 | 197.333 ± 25.765 | 190.501 ± 43.585 | 190.501 ± 43.585 | 458.182 ± 59.892 | 458.182 ± 59.892 |
| Rubidium (Rb) | bdl | bdl | bdl | bdl | 16.273 ± 7.953 | 16.273 ± 7.953 |
| Lead (Pb) | bdl | bdl | 94.151 ± 21.404 | 94.151 ± 21.404 | bdl | bdl |
| Strontium (Sr) | 27.600 ± 3.757 | 27.600 ± 3.757 | 54.301 ± 5.956 | 54.301 ± 5.956 | 14.682 ± 2.697 | 14.682 ± 2.697 |
| Thorium (Th) | bdl | bdl | 47.358 ± 9.756 | 47.358 ± 9.756 | bdl | bdl |
| Uranium (U) | bdl | bdl | 10.784 ± 10.777 | 10.784 ± 10.777 | bdl | bdl |
| Vanadium (V) | 346.800 ± 89.405 | 346.800 ± 89.405 | 206.351 ± 15.277 | 206.351 ± 15.277 | 482.000 ± 74.485 | 482.000 ± 74.485 |
| Zinc (Zn) | bdl | bdl | 265.509 ± 21.392 | 265.509 ± 21.392 | bdl | bdl |
| Zirconium (Zr) | bdl | bdl | 549.901 ± 18.049 | 549.901 ± 18.049 | 545.682 ± 41.891 | 545.682 ± 41.891 |
| ΣREE c | 944.77 ± 14.307 | 337.49± 10.703 | 687.75 ± 22.969 | 387.61 ± 29.369 | 1762.68 ± 22.174 | 826.51 ± 24.471 |
| Pollutant TotalConcentration | 1,814.1027 ± 88.08 | 1,206.8227 ± 46.12 | 3,426.2600 ± 19.49 | 3,126.1200 ± 32.31 | 4,376.0080 ± 67.96 | 3,439.8380 ± 98.23 |
a Values are the means ± sd; b bdl = below detection limit; c ΣREE include Ce, Dy, Er, Eu, Gd, Ho, La, Lu, Nd, Pr, Sm, Sc, Tb, Tm, Y, Yb.
2.3. In Vitro Assay on Target Organisms
The effects of soil on SM, FOL, VD, and TH were analysed using liquid and solid media.
Liquid cultures were achieved in 100 mL Erlenmeyer flasks containing 40 mL of half-strength Potato Dextrose Broth (Oxoid Ltd., ½PDB) amended with 1% fine soil from GB2, GE2, OD3, OE4, SB3 or SE4 samples. As a control, Erlenmeyer containing ½PDB were prepared. After sterilization (121 °C, 20 min), every flask was inoculated with 105 conidia of TH, FOL or VD or 105 fragments of SM mycelia. For each fungal species and soil, three flasks were prepared. The experiment was repeated twice. After eight days of incubation (25±1 °C, in the dark, 150 rpm on an orbital shaker), the average conidia concentrations in FOL, VD and TH, cultures and microsclerotia presence in SM were determined by a Thoma chamber under an optical microscope Zeiss standard mod 16 in control (Sc) and ½PDB amended with soil sample (Ss) cultures. The sporulation inhibition rate (SIR) was calculated as SIR=100×[(Sc-Ss)/Sc]. The mycelia were separated by filtration on cheesecloth and two layers of Miracloth (Calbiochem-Merck-Sigma-Aldrich, Milan, I) and centrifugation (8,000 rpm, 20 min, 4 °C, Thermo Scientific SL8R centrifuge, Thermo Fisher Scientific Inc., Osterode am Harz, D). The collected mycelia were washed four times each with 40 mL of sterile distilled water to remove adherent soil, frozen, lyophilized, and weighed to obtain dry weight (dwt) biomass. The Percentage of Growth Inhibition Rate (PGIR) was calculated as PGIR=100×[(Bc-Bs)/Bc] where Bc and Bs are the average dwt biomass in control or with soil sample, respectively. The pH of each culture filtrate (CF) was measured.
Petri dishes (diam 90 mm) containing PDA (24 g L-1) amended with 1% fine soil samples GB2, GE2, OD3, OE4, SB3 or SE4 were used in solid culture experiments. Sterilized (121 °C, 20 min) media were left to cool down to 50 °C and poured (10 ml per dish). Plates containing PDA were prepared as a control. Each plate was inoculated in the centre with a mycelium plug (diam 3 mm) obtained from FOL, VD, SM and TH fresh colony, sealed with Parafilm M and incubated at 25±1 °C, in the dark. For each fungal species and soil sample, three plates were prepared. The experiment was repeated twice. Every day for three weeks, colony growth was measured as two orthogonal diameters. The percentage of radial growth inhibition (PRGI) was calculated considering the average of colony diameters on PDA (Dc) and PDA amended with soil (Ds) in the formula PRGI=100×(Dc-Ds)/Dc. After 35 days of incubation, 10 plugs (diam 5 mm) were taken randomly from each FOL, VD and TH colony, transferred into 10 ml of sterile distilled water, vigorously vortexed and used for conidia enumeration by a Thoma chamber. At the same time, microsclerotia produced by SM were collected in 10 ml of sterile distilled water, counted under a stereoscope and expressed per plate. The SIR was calculated.
2.4. Effects on Trichoderma Harzianum Antagonistic Activity
Liquid cultures of TH were prepared in ½PDB (control) and ½PDB amended with 1% of GB2, GE2, OD3, OE4, SB3 or SE4 samples. Each flask (50 ml) was inoculated with 100 μL of conidia suspension (106 conidia per ml) collected from a colony of TH grown on PDA for 8 days at 25±1 °C, darkness. The inoculated flasks were incubated at 25±1 °C, in the dark on an orbital shaker (150 rpm). After 8 days, the CFs were collected by filtration on Miracloth and centrifugation (8000 rpm, 80 min, 4 °C, Thermo Scientific SL8R centrifuge) and sterilized through 0.22 μm Millipore membranes. These TH CFs were embedded in melted sterile PDA in the ratio 3:1 (v/v; TH CF : PDA), vigorously shaken, and poured into a 90 mm Petri dish (30 ml per plate), to obtain TH CF Control (TH½PDB), TH CF on ½PDB plus GB2 (TH½PDB-GB2), TH CF on ½PDB plus GE4 (TH½PDB-GE4), TH CF on ½PDB plus OD3 (TH½PDB-OD3), TH CF on ½PDB plus OE4 (TH½PDB-OE4), TH CF on ½PDB plus SB3 (TH½PDB-SB3), TH CF on ½PDB plus SE4 (TH½PDB-SD4). As a non-inoculate control, ½PDB (½PDB-NI) was prepared. Plugs (diam 8 mm) were taken with a cork bore and used to set up antagonism tests according to Figure 1.
Every organism (FOL, VD or SM) and TH CFs combinations were prepared in triplicate. The inoculated plates were sealed with Parafilm M and incubated (25±1 °C) in the dark. The advance of considered fungus and the colonization of the plugs were monitored daily for three weeks. The percentage of plug colonization (PPC) was calculated as PPC=100×(PCon-PTHCF)/PCon where PCon and PTHCF represent the number of colonized plugs containing ½PDB and TH CF, respectively. The experiment was repeated three times.
2.4. Statistical Analysis
For each parameter the mean and standard deviation (sd) were calculated. Data were subjected to Shapiro–Wilk’s and Levene’s tests to verify the distribution’s normality and the variances’ homoscedasticity, respectively. Subsequently, the variance analysis (ANOVA) with a 95% confidence level and Fisher’s least significant difference (LSD) were performed using SAS software version 9.0 for Windows. The percentage values were arcsine transformed before the analysis. The plots were assembled using Excel software. The Pearson correlation coefficient (r) was estimated among the PTC, mCId or NIRI values and each effect on tested fungi. The variables were considered [54]: uncorrelated (r = 0), positively correlated (r > 0), or negatively (r < 0) correlated. The correlation was considered weak (r < |0.3|), moderate (|0.31| < r < |0.7|), or strong (r > |0.71|).
3. Results
3.1. Soil Characterization
The fine soil collected around the three dismissed bauxite mines in Puglia have pH values (Table 2) ranging from 7.89±0.13 (sample OE4) to 8.42±0.12 (sample SE4). Based on mCId (Table 2), soils at the collection points recorded low (GE4), moderate (GB2), high (OE3, OE4, SE4) and very high (SB3) pollution. As shown in Table 2, low NIRI values were recorded across all sampling points.
3.2. Effects of Soil Samples on Target Organisms
The four target fungi recorded different responses depending on the culture performed and soil samples considered.
In the liquid culture (Figure 2), GB2, GE4, OD3, OE4 and SE4 soil samples slowed TH growth by an average of 63.16%, while SB3 induced a 7.34% increase. Samples OD3 and OE4 increased VD mycelial mass by 29.73 and 43.22%, respectively. The SM growth increased between 18.95 and 58.44% for OD3 and SE4, respectively. The FOL growth ranged from the 8.33% enhancement in the media containing GB2 to 8.58% inhibition in ½PDB amended with SB3. The presence of soil samples affected VD, TH and FOL conidia production (Figure 2). Microsclerotia were not produced by SM under all the tested media.
At the end of the growing time, different CF pHs were recorded (Table 3). Compared to the non-inoculated media (Table 3), TH, VD, and SM development decreased the pH, while FOL behaviour was conditioned by the soil sample added in the medium: pH increase for GB2, GE4, and SE4, and a slight reduction was recorded for OD3, OE4, and SB3.
On PDA amended with the fine soil from GB2, GE2, OD3, OE4, SB3 and SE4 samples, the PRGI (Figure 3) outlines a different behaviour for each tested fungal species. Compared with the control, the six soil samples inhibited VD (21.9-32.2%) and SM (4.3-65.2%) radial growth, TH was inhibited (22-25%) by the presence of GB2 and SB3, while growth enhances were recorded on PDA modified with OE4 (9.5%), OD3 (15.8%), GE4 (15.8%) and SB4 (17.1%). Growth improvement (65-86%) was recorded in FOL cultures.
All soil samples inhibited the production of FOL conidia (Figure 3): GB2, OD3 and SB3, collected near the mine entrances were the more efficient (61-64% SIR). Compared to the PDA control, VD increased conidia production in the media containing soil samples (27-36%) apart from SB3 which recorded a 12% reduction. All tested soil samples stimulated TH conidia production. The SM microsclerotia production did not undergo significant variations.
FOL, VD and SM showed different comportment towards the plugs containing the TH CFs (Figure 4). All the ½PDB-NI plugs containing uninoculated PDB were colonized bay FOL, SM and VD mycelia, while the plugs incorporating TH CFs revealed different PPC (Figure 4).
An antagonistic action was observed for the plugs containing the TH½PDB CF that inhibited the growth of the three target fungi. SM failed to colonize plugs containing TH CFs obtained in the presence of each tested soil sample. The TH½PDB-OD3 and TH½PDB-SE4 plugs were completely not colonized by FOL. VD did not colonize TH½PDB-GB2 plugs, while different PPCs were recorded for the other plugs. SM did not colonize TH½PDB plugs including the fine soil samples.
The Pearson correlation coefficient (Figure 5) shows different correlations levels among the PTC, mCId, or NIRI and the recorded parameters. The presence of GB2, GE4, OD3 and OE4 soil samples positively correlated PTC and mCId to pH, PGIR and SIR of liquid cultures, and PRGI and SIR collected from PDA experiments. The antagonistic effects of TH culture filtrates were uncorrelated to PTC and mCId. The NIRI values showed a positive weak correlation between GB2 and GE4 soil samples and the biological parameter recorded for the tested organisms. Samples OD3, OE4, SB3 and SE4 noted a weak negative correlation with pH, PGIR and SIR of liquid cultures, and PRGI and SIR related to solid media experiments. The PPCs explain a weak positive correlation with each tested soil sample.
4. Discussion
Living organisms are affected by the chemical composition of their environment. Particularly, metals when present under excessive concentrations affect biota activity and survival. At the same time, microorganisms remove metal ions, including toxic (heavy) metals and REEs, by adsorption on the surface of microbial cells, onto extracellular biopolymers (e.g., polyglutamic acid and polysaccharides), into microbial cells, by bio-minerals such as manganese oxide [10,55,56].
Soil contamination by toxic metals is the result of natural processes and human activities. Damage to soil microorganisms has often been reported as the final effect of metal toxicity. Severe ammonification and nitrification alteration were the first observations associated with Cu, Zn, Fe, Pb [57] or manganese [58] presence and concentrations. To avoid ecosystem disturbance and human exposure measures to limit the toxic metal loading rates were introduced in many European countries [10] and are constantly updated.
Contradictory are the conclusions carried out on REEs effects on biota: toxicity [19,24,25,26,27,28,32,33,34,35] or positive impacts [20,28,31]. The extensive information on the biota exposure to pollutants collected from laboratory studies shows disparity in critical toxic concentrations and related effects [10]. To avoid this limiting factor, soil samples collected around three dismissed bauxite mines in the Puglia region, at San Giovanni Rotondo, Otranto and Spinazzola were used as natural sources of pollutants, mainly toxic metals and REEs [48,49,50,51,52], against target organisms selected among soil-borne fungi. The elected mines played a key role in the “Italian” aluminium production. The mines in San Giovanni Rotondo and Otranto were utilized from 1937 to 1973 and 1940 to 1976, respectively; while the mine in Spinazzola, discovered in 1935, remained active from 1950 to 1978. The soil around these three dismissed bauxite mines is the reddish clay soil called Terra Rossa characterized by hematite, kaolinite, and goethite present on the surface of limestone [59].
All mining procedures, including bauxite extraction, cause drastic disturbances in landscape and soil properties. The soil around mines is characterized by poor soil structure, nutrient deficiency, low fertility, high concentrations of toxic metals (i.e., As, Cr, Cs, Cu, Mo, Pb) and REEs (e.g., Ce, Dy, Er, Eu, Gd, Ho, La, Lu, Nd, Pr, Sm, Sc, Tb, Tm, Y, Yb); loss of P, K, Ca, Al and Mg content, effective cation exchange capacity, soil organic carbon and nitrogen pool; increase soil pH, electrical conductivity, and bulk density, reduced soil microbiota communities and improved ecological risk assessment [2,44,45,46,47,48,60,61,62,63,64,65].
The presence of pollutants such as PAHs, C10-C40 hydrocarbons, metals and REEs has also been ascertained in dust and surface layers of soil in areas of high industrial activity of Taranto and Augusta Priolo, showing differences in topographic distribution among the soil sampling sites [66,67]. From the tree site explored in this study, soil samples taken in the area near (GB2, OD3, SB3) and in the zone far (GE2, OE4, SE4) from the mine entrance, confirm the presence of As, Ba, Co, Cr, Cs, Cu, Ga, Hf, Mo, Nb, Ni, Rb, Pb, Sr, Th, U, V, Zn, Zr, Y, La, Ce, Pr, Nd, Sm, Eu, Gd, Tb, Dy, Ho, Er, Tm, Yb and Lu. The average concentration of these elements reached 1510.4627 (GE2 = 1814.1027, GB2 = 1206.8227), 3276.19 (OE4 = 3426.2600, OD3 = 3126.1200) and 3907.9230 (SE4 = 4376.0080, SB3 3439.8380) mg kg-1 in San Giovanni Rotondo, Otranto and Spinazzola, respectively.
Soil pH impacts the solubility, bioavailability and overall mobility of heavy metals and REE. Higher pH values generally reduce the mobility and availability, while lower values, particularly below pH 5, can increase heavy metals solubility and toxicity [51,68]. Following these indications, heavy metals and REEs in the soils of the three here considered sites (pH values in the range 7.89±0.13 - 8.42±0.12) should have a low bioavailability. The pH of post-bauxite mining soils herein analysed is higher than the pH 4.96 recorded in Indonesia [69] and 5.1 in Brazil [60] but agrees with soil analysed in the Philippines [61].
The ranking of mCd values for the six soil samples was as follows: GE4 (1.999±0.20) < GB2 (3.891±0.52) < OE4 (4.555±0.07) < OE3 (5.324±0.21) < SE4 (6.703±0.46) < SB3 (10.448±0.94). According to the proposed classifications [51,52], collected soils showed low (GE4), moderate (GB2), high (OE3, OE4, SE4) and very high (SB3) pollution. The distribution of pollution levels is reduced as the distance from the mine entrance increases. It is speculated that the mining of the deposit is one of the reasons for the high mCd.
The calculation of NIRI eliminates the effect of the number of metals considered, avoids the overestimation caused by summing numerical values, combines the toxic response factors and allows an accurate valuation of metal’s contribution to the potential ecological risk. No differences were recorded between the samples taken near or far from the mine entrance. Distinctions are possible among the three dismissed mines. The mean NIRI values in the San Giovanni Rotondo, Otranto, and Spinazzola were 18.732±0.48, 15.533±0.45, and 2.934±0.27, respectively, with the San Giovanni Rotondo and Otranto being significantly higher than the Spinazzola area. Most sampling sites in the study areas that have high or very high pollution levels, shows low potential ecological risk.
The potential effects of soil samples were established on three strains of fungal plant pathogens and one BCA as target organisms selected based on their soil-borne habitus.
Fusarium oxysporum f. sp. lycopersici, widespread throughout the Mediterranean basin, is the significant pathogen of tomato vascular wilt. It lives as a saprophyte on tomato plant residues in the soil, differentiates one-celled microconidia (2.5-4 × 6-15 μm), 2-3 septate macroconidia (2.5-3.3 × 3.5-5.5 μm) and, formed in older mycelium, spherical chlamydospores (5.70-7.5 μm in diameter) that favour its preservation for up to 10 years. This fungus infects tomato seedlings close to transplanting and those in greenhouses and open-field cultivation. Yield losses due to disease can reach 45-55% and extend up to 70% under favourable conditions (27-30 °C). The typical symptoms of vascular wilt caused by F. oxysporum f. sp. lycopersici include yellowing of the basal leaves and epinasty (downward curvature of the leaf lamina). The intercellular colonization of the cortical tissues and xylem establishes a typical dark brown colour of the roots, collar and stems, leading to wilting and death of the entire plant or its sectors [70,71].
Sclerotinia minor is a polyphagous fungus that infects cucurbits, carrots, celery, fennel, lettuce, cabbage, tomato, artichoke, thistle, sunflower, and other botanical species during cultivation, and pears, lemons, and other fruits during post-harvest storage. Plants attacked by this pathogen show, at the base or along the stem, brownish lesions that evolve into rotting areas covered with cottony white efflorescence in which masses of blackish microsclerotia (8-10 mm) appear and allow the pathogen preservation in the soil for several years. On tomatoes, S. minor leads the plant to a rapid death. Conditions favourable to infection are 15-28 °C and some days of high humidity [71].
Verticillium dahliae is the agent of Verticillium wilt on vegetables (e.g., tomato, pepper, potato, eggplant, and artichoke), olives, vine and other three hundred botanical species. This fungus survives for many years in the soil as microsclerotia or hyphae and conidia in crop residues. The infection of the host plant starts from the roots, directly or through wounds, and colonizes the xylem vessels producing conidia that will be transported in an acropetal direction. As a defence mechanism, the infected plant produces gums and tyloses inside the tracheid which, together with mycelium and conidia of the fungus, cause the clogging of the xylem vessels, wilt and death of the plant. On tomatoes, the root system remains intact, wilting symptoms involve first on the basal oldest leaves, and then affect the younger ones, with a reduction in yield of 20-50% [71,72].
Species of the genus Trichoderma, including T. harzianum, are saprophytic, filamentous fungi ubiquitous in the soil and plant rhizosphere. They are strong opportunists, and producers of considerable amounts of conidia, antimicrobial substances, and lytic enzymes. These species are worldwide applied as BCAs to manage nematodes and plant disease agents such as Phytophthora, Pythium, Rhizopus, Athelia, Verticillium, Rhizoctonia, Fusarium, Sclerotinia and other phytopathogenic fungal species [73,74,75]. These fungi have been extensively utilized as biofertilizers to promote vegetative growth under different agricultural conditions by producing plant growth stimulants [74].
Starting from these considerations, the soil samples taken around the three dismissed bauxite mines in Puglia were tested in liquid and solid (agarized) media since the physical-chemical properties of the media can affect dynamics of fungal growth response, metabolisms and xenobiotic uptake, accumulation and efficacy. The liquid culture tests support a more extensive contact between soil samples and fungal-produced structures. Assessing the soil instead of individual elements or mixtures prepared in the laboratory is the singleness of the methodologic approach used in this work. The effects on biota caused by environmentally relevant exposures are usually related to mixtures and not only to one or a few representative substances.
Assayed on PDA all the tested soil samples decreased VD (22-32%) and SM (4-65%) or increased FOL (65-86%) radial development. The TH growth is soil-dependent: reduced by GB2 (25%) and SB3 (22%) samples, and stimulated by OE4 (9.5%), OD3 (15.8%), GE4 (15.8%) and SB4 (17.1%). Soils also affected the differentiation of dissemination or maintenance structures. Conidia production was reduced in FOL (37-78%) and increased in TH (5-109%) and VD (12-41%) while no effects were recorded on SM microsclerotia.
Media composition (e.g., nutrient sources, pH) and growth parameters (e.g., light/dark cycle, temperature, relative humidity, static/stirred conditions) influence fungal biomass production and sporulation. Structures useful for dissemination such as conidia and conservation structures such as microsclerotia represent, for the producers’ fungi, high ecological versatility, and security in perpetuation in time and space while waiting for the host [76]. Certainly, solid media, i.e., PDA, provide water and nutrients and allow the fungal hypha to grow and develop, whereas liquid cultures, both in static and stirred conditions, could slow down fungal growth, however they grant contact between mycelia and any xenobiotics to be tested. Media composition and growing conditions also affect fungal primary and secondary metabolisms and optimize the yields of specific active metabolite in medicinal fungi or drug-producing microbe [77,78,79]. The experiments conducted in the liquid PDB amended with the established soil samples indicate different performance in terms of growth and differentiation of dissemination or maintenance structures among the four fungal strains tested. The soil samples reduced the growth of TH (61.5-65%, except for sample SB3 which induced 7.3% biomass increment) and stimulated SM (8-58%) and FOL (0.5-10%, apart from sample SB3 which reduce SM and FOL biomasses of 2.5 and 8%, respectively). A different comportment was recorded for VD: growth reduction (3%) by the GB2 and GE4 samples, stimulation by the OD3 (30%) and OE4 (43%). Under the same growing conditions, the four fungal species also differentiated the production of conservation and diffusion structures.
Liquid cultures have some advantages: (i) rapid mycelium growth, (ii) uniform presence of the sample to be assessed, and (iii) easy sporulation verification. In addition, liquid cultures are usually used to stimulate the production of fungal secondary metabolites that are toxic to plants or other fungi. An inhibition in the conidia production was recorded in VD (85-90.8%) and TH (20-70.23%), while FOL increased (1.3-3.5-fold) conidia differentiation when compared with the controls in PDB. In all liquid cultures, SM did not produce microsclerotia.
Trichoderma species are components of soil microbiome and are widely promoted as biopesticides, biofertilizers and soil amendments, due to their ability to produce secondary metabolites with antimicrobial properties effective in vitro against several plant diseases agents [73] and work as defence substance activators in plants, stimulating pathogenesis-related proteins (PR-proteins) and enzymes, including peroxidase, chitinase, beta-1,3-glucanase and lipoxygenase [74]. Under the growth condition herein applied, the strain TH reduced antimicrobial effects against FOL, VD and SM depending on the soil sample and fungal strain considered. The presence of GB2, GE2, OD3, OE4, SB3 and SE4 soil samples in the liquid media also altered TH microbicidal action. The TH filtrates obtained in PDB, embedded in PDA inhibited the plugs colonization by FOL, VD and SM. The CFs obtained in ½PDB modified with the addition of 1% soil samples have, in all cases, avoided the growth of SM leaving its antagonistic activity unaltered. Soil-dependent behaviour was observed in FOL and VD: TH½PDB-OD3 and TH½PDB-SE4 blocked FOL growth; VD did not colonize TH½PDB-GB2 plugs and partially colonized the other.
Despite different target organisms, our results agree with previously published data regarding the soil of other industrial installations [66,67,71]. Fungal growth, sporulation and antagonistic activity are affected by the soil’s physical structure, chemical composition, elemental content and availability. However, hypotheses can be made to be supported by further evidence. It could be hypothesized that toxic metals and REEs may favour the growth and conidia production of Trichoderma in the soil, but reduce their antagonistic activity mediated by fungitoxic molecules. The employment of the results discussed here could mitigate the deleterious effects of V. dahliae and S. minor disease. In fact, by slowing down their saprophyte phase the requirement of a longer time to start the infectious process may be hypothesized. Stimulating the production of conidia in V. dahliae improves the possibility of dissemination. On the contrary, F. oxysporum f. sp. lycopersici could mitigate conidia production and the lower dispersion, increasing saprophyte mycelial growth.
The widespread presence of toxic metals and REEs in the soil, beyond the possible accumulation of undesired elements in crops, could influence the growth of T. harzianum, V. dahliae, S. minor, F. oxysporum f. sp. lycopersici, and, presumably, other fungal populations (e.g., microbiome, rhizosphere, symbionts, plant growth-promoting rhizobacteria, mycorrhizal fungi, endophytic bacteria, actinomycetes, nematodes, protozoans, soil-borne agent of plant disease, and BCAs), their development and dispersal, biological interactions with soil microbiome and plants, and disease/protection ability. In addition, T. harzianum strain TH, V. dahliae strain VD, S. minor strain SM and F. oxysporum f. sp. lycopersici strain FOL could be investigated in the REE and heavy metals soil bioremediation.
These data are preliminary; further tests are needed with a wider range of S. minor, F. oxysporum f. sp. lycopersici, V. dahliae, T. harzianum isolates and other soil-borne fungi to verify the universality of the obtained findings. In addition, the three sites here considered, along with many others dismissed mines represent important tourist attractions, cultural elements, included in geoparks, transformed into museums [1]; are visited by an ever-increasing number of people and subject to the pilgrimage and recreative activities of curious people of all ages. For this reason, it is crucial to study and monitor heavy metals and REEs as responsible for alterations in the chemical and biological composition of soils and to control and characterize some endogenous fungus strains.
Final speculation is necessary for Pearson correlation coefficient. The bio-toxic effects of the tested soil samples were positively weak/moderate related to PTC and mCId ascribing a dose-effect relationship with the elemental concentration of pollutants and their contamination factor. At the same, a weak positive or negative correlation is associated with NIRI values. It is evident that Cd, Cu, As, Ni, Cr, Pb, and Zn, included in the NIRI calculations, are partly associated with biological effects recorded on FOL, VD, SM and TH strains growth with the soil samples. Other toxic components could be considered valuable sources of toxicity. Considering the elemental composition of soil samples collected at the three dismissed bauxite mines, the REE content reached 52.08, 27.97, 20.07, 12.39, 40.28 and 24.03% in GB2, GE2, OD3, OE4, SB3, and SE4, respectively. With high probability, the effect on the four tested fungi here detected and discussed will be due to REEs. Moreover, the impact of other chemicals such as heavy metals (e.g., Zn, Pb, Cd, Hg) that could contribute to toxicity, should be considered.
5. Conclusions
The six soil samples collected from three dismissed bauxite mines in Puglia revealed consistent levels of toxic metals and REEs higher than those reported in other sites. These soil samples affected the four strains of target pathogens inducing different effects on radial growth and spore production on solid (agarized) PDA and liquid PDB. The experienced strains compensate for growth reduction by the greater production of reproductive structures, or, vice versa, enhance biomasses production to balance low conidia differentiation. Soil samples also affected the antimicrobial activity of the TH strain. Data collected, analysed and discussed here suggests the need to monitor toxic metals and, in particular, REEs impact on the biota in the environment and the soils, especially in locations of naturalistic, geological, and tourist interest.
Author Contributions
Conceptualization, G.L.B., and F.To.; methodology, G.L.B., Fa.M., S.P., F.To., F.Tu., I.G., G.F., Fo.M and A.A.B; validation, G.L.B., and F.T.; formal analysis, G.L.B., Fo.M., S.P., F.Tu., I.G., G.F., and A.A.B.; investigation, G.L.B., Fa.M., S.P., F.Tu. and A.A.B.; resources, G.L.B. and F.To.; data curation, G.L.B., Fa.M., Fo.M., S.P., F.To., I.G., G.F., F.Tu. and A.A.B.; writing—original draft preparation, G.L.B., S.P., Fo.M., F.Tu., I.G., A.A.B., Fa.M., G.F., G.L. and F.To.; writing—review and editing, G.L.B., S.P., Fo.M., F.Tu., I.G., A.A.B., Fa.M., G.F., G.L. and F.To.; supervision, G.L.B., and F.To.; project administration, F.To.; funding acquisition, F.To, G.L.B. and Fa.M.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the projects “TREDIMPOSTRA- LE TERRE RARE, ELEMENTI GEOCHIMICI DI IMPORTANZA STRATEGICA: PRESENZA NEL SUOLO E SOTTOSUOLO ED EFFETTI IN ORGANISMI MODELLO” (CUP H91I21001690006—Horizon Europe Seeds University of Bari D.R. 1940, 04/06/2021).
Data Availability Statement
The datasets presented in this study are included in the article, further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Varriale, R.; Aldighieri, B.; Genovese, L. Dismissed mines: from the past to the future. Heritage 2023, 6, 2152–2185. [CrossRef]
- Masinga, P.; Simbanegavi, T.T.; Makuvara, Z.; Marumure J.; Chaukura N.; Gwenzi W. Emerging organic contaminants in the soil–plant-receptor continuum: transport, fate, health risks, and removal mechanisms. Environ. Monit. Assess. 2024, 196, 367. [CrossRef]
- Järup, L. Hazards of heavy metal contamination. Br. Med. Bull. 2003, 68, 167–82. [CrossRef]
- Zoroddu, M.A.; Aaseth, J.; Crisponi, G.; Medici, S.; Peana, M.; Nurchi, V.M. The essential metals for humans: a brief overview. J. Inor. Biochem. 2019, 195, 120–129. [CrossRef]
- Peana, M.; Pelucelli, A.; Medici, S.; Cappai, R.; Nurchi, V.M.; Zoroddu, M.A. Metal toxicity and speciation: a review. Curr. Med. Chem. 2021, 28(35), 7190–7208. [CrossRef]
- Jan, A.T.; Azam, M.; Siddiqui, K.; Ali, A.; Choi, I.; Haq, Q.M. Heavy metals and human health: mechanistic insight into toxicity and counter defense system of antioxidants. Int. J. Mol. Sci. 2015, 16(12), 29592–29630. [CrossRef]
- Priyadarshini, E.; Priyadarshini, S.S.; Cousins, B.G.; Pradhan, N. Metal-fungus interaction: review on cellular processes underlying heavy metal detoxification and synthesis of metal nanoparticles. Chemosphere 2021, 274, 129976. [CrossRef]
- Dalvi, A.A.; Bhalerao, S.A. Response of plants towards heavy metal toxicity: an overview of avoidance, tolerance and uptake mechanism. Ann. Plant Sci. 2013, 2, 362–368.
- Mohamadhasani, F.; Rahimi, M. Growth response and mycoremediation of heavy metals by fungus Pleurotus sp. Sci. Rep. 2022, 12, 19947. [CrossRef]
- Giller K.E.; Witter E.; Mcgrath S.P. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: a review. Soil Biol. Biochem. 1998, 30(10–11), 1389-1414. [CrossRef]
- Igiri, B.E.; Okoduwa, S.I.R.; Idoko, G.O.; Akabuogu, E.P.; Adeyi, A.O.; Ejiogu, I.K. Toxicity and bioremediation of heavy metals contaminated ecosystem from tannery wastewater, a review. J. Toxicol. 2018, 2568038. [CrossRef]
- Del Val C.; Barea J.M.; Azcón-Aguilar C. Diversity of arbuscular mycorrhizal fungus populations in heavy-metal- contaminated soils. Appl. Environ. Microbiol. 1999, 65(2), 718-723. [CrossRef]
- Bano, A.; Hussai, J.; Akbarm, A.; Mehmood, K.; Anwar, M.; Hasni, M.S.; Ullah, S.; Sajid, S.; Ali, I. Biosorption of heavy metals by obligate halophilic fungi. Chemosphere 2018, 199, 218–222. [CrossRef]
- Graz, M.; Pawlikowska-Pawlega, B.; Jarosz-Wilkolazka, A. Growth inhibition and intracellular distribution of Pb ions by the white-rot fungus Abortiporus biennis. Int. Biodeterior. Biodegra. 2011, 65(1), 124–129. [CrossRef]
- Lilly, W.W.; Wallweber, G.J.; Lukefahr, T.A. Cadmium absorption and its effects on growth and mycelial morphology of the basidiomycete fungus, Schizophyllum commune. Microbios 1992, 72, 227–237.
- Mohan, P.M.; Pratap Rudra, M.P.; Sastry, K.S. Nickel transport in nickel–resistant strains of Neurospora crassa. Curr. Microbiol. 1984, 10, 125–128. [CrossRef]
- Haque, N.; Hughes, A.; Lim, S.; Vernon, C. Rare earth elements: overview of mining, mineralogy, uses, sustainability and environmental impact. Resources 2014, 3, 614–635. [CrossRef]
- Arciszewska, Ż.; Gama, S.; Leśniewska, B.; Malejko, J.; Nalewajko-Sieliwoniuk, E.; Zambrzycka-Szelewa, E.; Godlewska-Żyłkiewicz, B. The translocation pathways of rare earth elements from the environment to the food chain and their impact on human health. Process Saf. Environ. 2022, 168, 205–223. [CrossRef]
- Tommasi, F.; Thomas, P.J.; Pagano, G.; Perono, G.A.; Oral, R.; Lynos, D.M.; Toscanesi, M.; Trifuoggi, M. Review of rare earth elements as fertilizers and feed additives: a knowledge gap analysis. Arch. Environ. Contam. Toxicol. 2021, 81(4), 531–540. [CrossRef]
- Pagano, G.; Aliberti, F.; Guida, M.; Oral, R.; Sicialiano, A.; Trifuoggi, M.; Tommasi, F. Rare earth elements in human and animal health: state of art and research priorities. Environ. Research 2015, 142, 215–220. [CrossRef]
- Hu, Z.; Richter, H.; Sparovek, G.; Schnug, E. Physiological and biochemical effects of rare earth elements on plants and their agricultural significance: a review. J. Plant. Nutr. 2004, 27(1), 183–220. [CrossRef]
- Tao, Y.; Shen, L.; Feng, C.; Yang, R.; Qu, J.; Ju, H.; Zhang, Y. Distribution of rare earth elements (REEs) and their roles in plant growth: a review. Environ. Pollut. 2022, 298, 118540. [CrossRef]
- Tyler, G. Rare earth elements in soil and plant systems—a review. Plant Soil 2004, 267(1–2), 191–206. [CrossRef]
- McDonald, J.W.; Ghio, A.J.; Sheehan, C.E.; Bernhardt, P.F.; Roggli, V.L. Rare earth (Cerium oxide) pneumoconiosis: analytical scanning electron microscopy and literature review. Modern Pathol. 1995, 8, 859–865.
- Tong, S.L.; Zhu, W.Z.; Gao, Z.H.; Meng, Y.X.; Peng, R.L.; Lu, G.C. Distribution characteristics of rare earth elements in children’s scalp hair from a rare earths mining area in southern China. J. Environ. Sci. Heal. A 2004, 39(9), 2517–2532. [CrossRef]
- Bernstein, E.J.; Schmidt-Lauber, C.; Kay, J. Nephrogenic systemic fibrosis: a systemic fibrosing disease resulting from gadolinium exposure. Best Prac. Res. Cl. Rh. 2012, 26, 489–503. [CrossRef]
- Wei, B.; Li, Y.; Li, H.; Yu, J.; Ye, B.; Liang, T. Rare earth elements in human hair from a mining area of China. Ecotoxicol. Environ. Saf. 2013, 96, 118–123. [CrossRef]
- Pagano, G.; Guida, M.; Tommasi, F.; Oral, R. Health effects and toxicity mechanisms of rare earth elements-knowledge gaps and research prospects. Ecotoxicol. Environ. Saf. 2015, 115(5), 40–48. [CrossRef]
- Brouziotis, A.A.; Giarra, A.; Libralato, G.; Pagano, G.; Guida, M.; Trifuoggi, M. Toxicity of rare earth elements: an overview on human health impact. Front. Environ. Sci. 2022, 10, 948041. [CrossRef]
- Giri, S.; Karakoti, A.; Graham, R.P.; Maguire, J.L.; Reilly, C.M.; Seal, S.; Rattan, R.; Shridhar, V. Nanoceria: A rare-earth nanoparticle as a novel anti-angiogenic therapeutic agent in ovarian cancer. PLoS One 2013, 8(1), e54578. [CrossRef]
- Wakabayashi, T.; Ymamoto, A.; Kazaana, A.; Nakano, Y.; Nojiri, Y.; Kashiwazaki, M. Antibacterial, antifungal and nematocidal activities of rare earth ions. Biol. Trace Elem. Res. 2016, 174(2), 464e470. [CrossRef]
- Tommasi, F.; d’Aquino, L. Rare earth elements and plants. In Rare Earth Elements in Human and Environmental Health: At the Crossroad Between Toxicity and Safety; Pagano G. Ed.; Pan Stanford Publishing Pte. Ltd.: Singapore, 2017; pp. 107–125.
- Zicari, M.A.; d’Aquino, L.; Paradiso, A, Mastrolitti, S.; Tommasi, F. Effect of cerium on growth and antioxidant metabolism of Lemna minor L. Ecotoxicol. Environ. Saf. 2018, 163, 536–543. [CrossRef]
- Gjata, I.; Tommasi, F.; De Leonardis, S.; Dipierro, N.; Paciolla, C. Cytological alterations and oxidative stress induced by Cerium and Neodymium in lentil seedlings and onion bulbs. Front. Environ. Sci. 2022, 10, 969162. [CrossRef]
- Gjata, I.; van Drimmelen, C.K.E.; Tommasi, F. Paciolla, C.; Heise, S. Impact of rare earth elements in sediments on the growth and photosynthetic efficiency of the benthic plant Myriophyllum aquaticum. J. Soil. Sediment. 2024, 24, 3814–3823. [CrossRef]
- d’Aquino, L.; Morgana, M.; Carboni, M.A.; Staiano, M.; Vittori Antisari, M.; Re, M.; Lorito, M.; Vinale, F.; Abadi, K.M.; Woo, S.L. Effect of some rare earth elements on the growth and lanthanide accumulation in different Trichoderma strains. Soil Biol. Biochem. 2009, 41(12), 2406–2413. [CrossRef]
- Palmieri, M.C.; Garcia, O.; Melnikov, P. Neodymium biosorption from acidic solutions in batch system. Process Biochem. 2000, 36(5), 441–444. [CrossRef]
- Qu, Y.; Lian, B. Bioleaching of rare earth and radioactive elements from red mud using Penicillium tricolor RM-10. Bioresource Technol. 2013, 136, 16–23. [CrossRef]
- Vlachou, A.; Symeopoulos, B.; Bourikas, K.; Koutinas, A.A. A comparative study of neodymium uptake by yeast cells. Radiochim. Acta 2008, 97(8), 437–441.
- Tsuruta, T. Accumulation of rare earth elements in various microorganisms. J. Rare Earth. 2007, 25, 526–532. [CrossRef]
- Calabrese, B.J. Hormetic mechanisms. Crit. Rev. Toxicol. 2013, 43, 580–606. [CrossRef]
- d’Aquino, L.; Tommasi, F. Rare Earth Elements and Microorganism. In Rare Earth Elements in Human and Environmental Health: At the Crossroad Between Toxicity and Safety; Pagano G. Ed.; Pan Stanford Publishing Pte. Ltd.: Singapore, 2017; 127–141.
- Hassan, SH.A.; Van Ginkel, S.W.; Hussein, M.A.M.; Abskharon, R.; Oh, S-E. Toxicity assessment using different bioassays and microbial biosensors. Environ. Int. 2016, 92–93, 106–118. [CrossRef]
- Mongelli, G.; Boni, M.; Buccione, R.; Sinisi, R. Geochemistry of the Apulian karst bauxites (southern Italy): chemical fractionation and parental affinities. Ore Geol. Rev. 2014, 63, 9–21. [CrossRef]
- Mongelli, G.; Buccione, R.; Gueguen, E.; Langone, A.; Sinisi R. Geochemistry of the Apulian allochthonous karst bauxite, Southern Italy: distribution of critical elements and constraints on Late Cretaceous Peri-Tethyan palaeogeography. Ore Geol. Rev. 2016, 77, 246–259. [CrossRef]
- Mongelli, G.; Boni, M.; Oggiano, G.; Mameli, P.; Sinisi R.; Buccione, R.; Mondillo, N. Critical metals distribution in Tethyan karst bauxite: the cretaceous Italian ores. Ore Geol. Rev. 2017, 86, 526–536. [CrossRef]
- Sinisi, R. Cretaceous bauxite from San Giovanni Rotondo (Apulia, Southern Italy): a provenance tool. Minerals 2018, 8, 567. [CrossRef]
- Brouziotis, A.A.; Heise, S.; Saviano, L.; Zhang, K.; Giarra, A.; Bau, M.; Tommasi, F.; Guida, M.; Libralato, G.; Trifuoggi, M. Levels of rare earth elements on three abandoned mining sites of bauxite in southern Italy: a comparison between TXRF and ICP-MS. Talanta 2024, 275, 126093. [CrossRef]
- Toussoun, T.A.; Nelson, P.E. A pictorial guide to the identification of Fusarium species according to the taxonomic system of Snyder and Hansen. The Pennsylvania State University Press: London, UK, 1968.
- Sudarningsih, S.; Fahruddin, F.; Lailiyanto, M.; Noer, A.A.; Husain, S.; Siregar, S.S.; Wahyono, S.C.; Ridwan, I. Assessment of soil contamination by heavy metals: a case of vegetable production center in Banjarbaru Region, Indonesia. Pol. J. Environ. Stud. 2023, 32, 249–257. [CrossRef]
- Cao Z.; Qian H.; Gao Y.; Li K.; Liu Y.; Shi X, Li, S.; Zhao, W.; Yang, S.; Tian, P.; Wu, P.; Ma, Y. Ecological risk assessment and source identification of heavy metals in the sediments of the Danjiang River Basin: a quantitative method combining multivariate analysis and the APCS-MLR model. Ecol. Indic. 2025, 175, 113518. [CrossRef]
- Xiao, H.; Shahab, A.; Xi, B.; Chang, Q.; You, S.; Li, J.; Sun, X.; Huang, H.; Li, X. Heavy metal pollution, ecological risk, spatial distribution, and source identification in sediments of the Lijiang River. China. Environ. Pollut. 2021, 269, 116189. [CrossRef]
- Men, C.; Liu, R.; Xu, L.; Wang, Q.; Guo, L.; Miao, Y.; Shen, Z. Source-specific ecological risk analysis and critical source identification of heavy metals in road dust in Beijing. China. J. Hazard. Mater. 2020, 388, 121763. [CrossRef]
- Miot, H.A. Correlation analysis in clinical and experimental studies. J, Vasc. Bras. 2018, 17, 275–279. [CrossRef]
- Klaus-Joerger, T.; Joerger, R.; Olsson, E.; Granqvist, C-G. Bacteria as workers in the living factory: metal-accumulating bacteria and their potential for materials science. Trends Biotechnol. 2001, 19, 15–20. [CrossRef]
- Moriwaki, H.; Yamamoto, H. Interactions of microorganisms with rare earth ions and their utilization for separation and environmental technology. Appl. Microbiol. Biotechnol. 2013, 97(1), 1–8. Epub 2012 Nov 1. [CrossRef]
- Lipman, C.B.; Burgess, P.S. The effects of copper, zinc, iron and lead salts on ammonification and nitrification in soils. Univ. Calif. Publ. Agric. Sci. 1914, 1,127–139.
- Brown, P.E.; Minges, G.A. The effect of some manganese salts on ammonification and nitrification. Soil Science 1916, 2, 67-85.
- Micheletti, F.; Fornelli, A.; Spalluto, L.; Parise, M.; Gallicchio, S.; Tursi, F.; Festa, V. Petrographic and geochemical inferences for genesis of terra rossa: a case study from the Apulian Karst (Southern Italy). Minerals 2023, 13, 499. [CrossRef]
- Vilas Boas, H.F.; Almeida, L.F.J.; Teixeira, R.S.; Souza, I.F.; Silva, I.R. Soil organic carbon recovery and coffee bean yield following bauxite mining. Land Degrad. Dev. 2018, 29, 1565–1573. [CrossRef]
- Shrestha, R.K.; Lal, R. Changes in physical and chemical properties of soil after surface mining and reclamation. Geoderma 2011, 161(3-4), 168–176. [CrossRef]
- Ling, K.Y.; Zhu, X.Q.; Tang, H.S.; Du, S.J.; Gu, J. Geology and geochemistry of the Xiaoshanba bauxite deposit, Central Guizhou Province, SW China: implications for the behavior of trace and rare earth elements. J. Geochem. Explor. 2018, 190, 170–186. [CrossRef]
- Atibu, E.K.; Devarajan, N.; Laffite, A.; Giuliani, G.; Salumu, J.A.; Muteb, R.C. Mulaji, C.K.; Otamonga, J.-P.; Elongo, V.; Mpiana, P.T.; Poté, J. Assessment of trace metal and rare earth elements contamination in rivers around abandoned and active mine areas. The case of Lubumbashi River and Tshamilemba Canal, Katanga, Democratic Republic of Congo. Geochemistry 2016, 76(3), 353–362. [CrossRef]
- Nkrumah, P.N.; Erskine, P.D.; Erskine, J.D.; Van Der Ent, A. Rare earth elements (REE) in soils and plants of a uranium-REE mine site and exploration target in Central Queensland, Australia. Plant Soil 2021, 464(1–2), 375–389. [CrossRef]
- Reinhardt, N.; Proenza, J.A.; Villanova-de-Benavent, C.; Aiglsperger, T.; Bover-Arnal, T.; Torró, L.; Salas, R.; Dziggel, A. Geochemistry and mineralogy of rare earth elements (REE) in Bauxitic ores of the Catalan Coastal Range, NE Spain. Minerals 2018, 8(12), 562. [CrossRef]
- Trifuoggi, M.; Pagano, G.; Oral, R.; Gravina, M.; Toscanesi, M.; Mozzillo, M.; Siciliano, A.; Burić, P.; Lyons, D.M.; Palumbo, A.; Thomas, P.J.; D’Ambra, L.; Crisci, A.; Guida, M.; Tommasi, F. Topsoil and urban dust pollution and toxicity in Taranto (southern Italy) industrial area and in a residential district. Environ. Monit. Assess. 2019, 191(1), 43. [CrossRef]
- Tommasi, F.; Lyons, D.M, Pagano, G.; Kovaçic, I.; Trifuoggi, M. Geospatial pattern of topsoil pollution and multi-endpoint toxicity in the petrochemical area of Augusta-Priolo (eastern Sicily, Italy). Chemosphere 2023, 333, 138802. [CrossRef]
- Naz, M.; Dai, Z.; Hussain, S.; Tariq, M.; Danish, S.; Khan, I.U.; Qi, S.; Du, D. The soil pH and heavy metals revealed their impact on soil microbial community. J. Environ. Manage. 2022, 321, 115770. [CrossRef]
- Prematuri, R.; Turjaman, M.; Sato, T.; Tawaraya, K. Post Bauxite mining land soil characteristics and its effects on the growth of Falcataria moluccana (Miq.) Barneby & J. W. Grimes and Albizia saman (Jacq.) Merr. Appl. Envir. Soil Sci. 2020, 6764380. [CrossRef]
- Maurya, S.; Dubey, S.; Kumari, R.; Verma, R. Management tactics for fusarium wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici (Sacc.): a review. Int. J. Res. Pharm. Pharm. Sci. 2019, 4(5), 1–7.
- Olivier, R.P. Agrios’ Plant Pathology, 6th Ed. Academic Press: London, UK, 2024.
- Pegg, G.F.; Brady, B.L. Verticillium Wilts. CABI Publishing: Wallingford, UK, 2002.
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, L.E.; Marra, R.; Woo, L.S.; Lorito M. Trichoderma-plant-pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Ruocco, M.; Woo, S.; Lorito, M. Trichoderma secondary metabolites that affect plant metabolism. Nat. Prod. Commun. 2012, 7, 1545–1550. [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65(2), 168–178. [CrossRef]
- Cooke, R.C.; Rayner, A.D.M. Ecology of Saprotrophic Fungi. Longman Publishing: London, UK, 1984.
- Miao, L.; Kwong, T.F.N.; Qian, P-Y. Effect of culture conditions on mycelial growth, antibacterial activity, and metabolite profiles of the marine-derived fungus Arthrinium c.f. saccharicola. Appl. Microbiol. Biotechnol. 2006, 72,1063–1073. [CrossRef]
- Pu, X.; Qu, X.; Chen, F.; Bao, J.; Zhang, G.; Luo, Y. Camptothecin-producing endophytic fungus Trichoderma atroviride LY357: isolation, identification, and fermentation conditions optimization for camptothecin production. Appl. Microbiol. Biotechnol. 2013, 97(21), 9365–9375. [CrossRef]
- VanderMolen, K.M.; Raja, H.A.; El-Elimat, T.; Oberlies, N.H. Evaluation of culture media for the production of secondary metabolites in a natural products screening program. AMB Express 2013, 3(1), 71. [CrossRef]
Figure 1.
Scheme used for antagonistic activity plate assays between target fungal species (●) and biological control agent culture filtrate (●) plugs.
Figure 1.
Scheme used for antagonistic activity plate assays between target fungal species (●) and biological control agent culture filtrate (●) plugs.
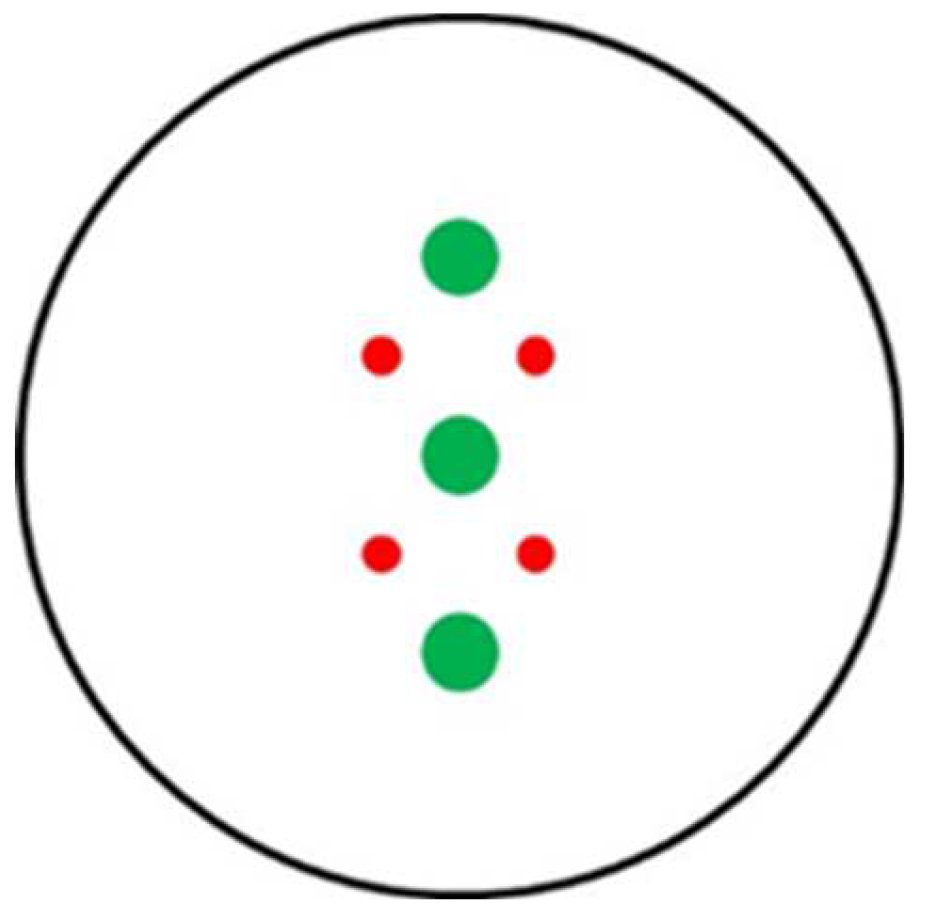
Figure 2.
Percentage of growth inhibition/enhancement of Fusarium oxysporum f. sp. lycopersici (■), Verticillium dahliae (■), Sclerotinia minor (■) and Trichoderma harzianum (■) mycelia in half strength Potato Dextrose Broth (½PDB) amended with fine soil (1%) of GB2 (½PDB+GB2), GE2 (½PDB+GE4), OD3 (½PDB+OD3), OE4 (½PDB+OE4), SB3 (½PDB+SB4) and SE4 (½PDB+SE4) samples. Each histogram represents the average of six replicates ± sd. The number in brackets indicates the sporulation inhibition rate. For each isolate or parameter, values marked with the same letter are not significant for Fischer’s LSD at P<0.05. Soil sample abbreviations and characteristics are in Table 1. NP = Not produced.
Figure 2.
Percentage of growth inhibition/enhancement of Fusarium oxysporum f. sp. lycopersici (■), Verticillium dahliae (■), Sclerotinia minor (■) and Trichoderma harzianum (■) mycelia in half strength Potato Dextrose Broth (½PDB) amended with fine soil (1%) of GB2 (½PDB+GB2), GE2 (½PDB+GE4), OD3 (½PDB+OD3), OE4 (½PDB+OE4), SB3 (½PDB+SB4) and SE4 (½PDB+SE4) samples. Each histogram represents the average of six replicates ± sd. The number in brackets indicates the sporulation inhibition rate. For each isolate or parameter, values marked with the same letter are not significant for Fischer’s LSD at P<0.05. Soil sample abbreviations and characteristics are in Table 1. NP = Not produced.
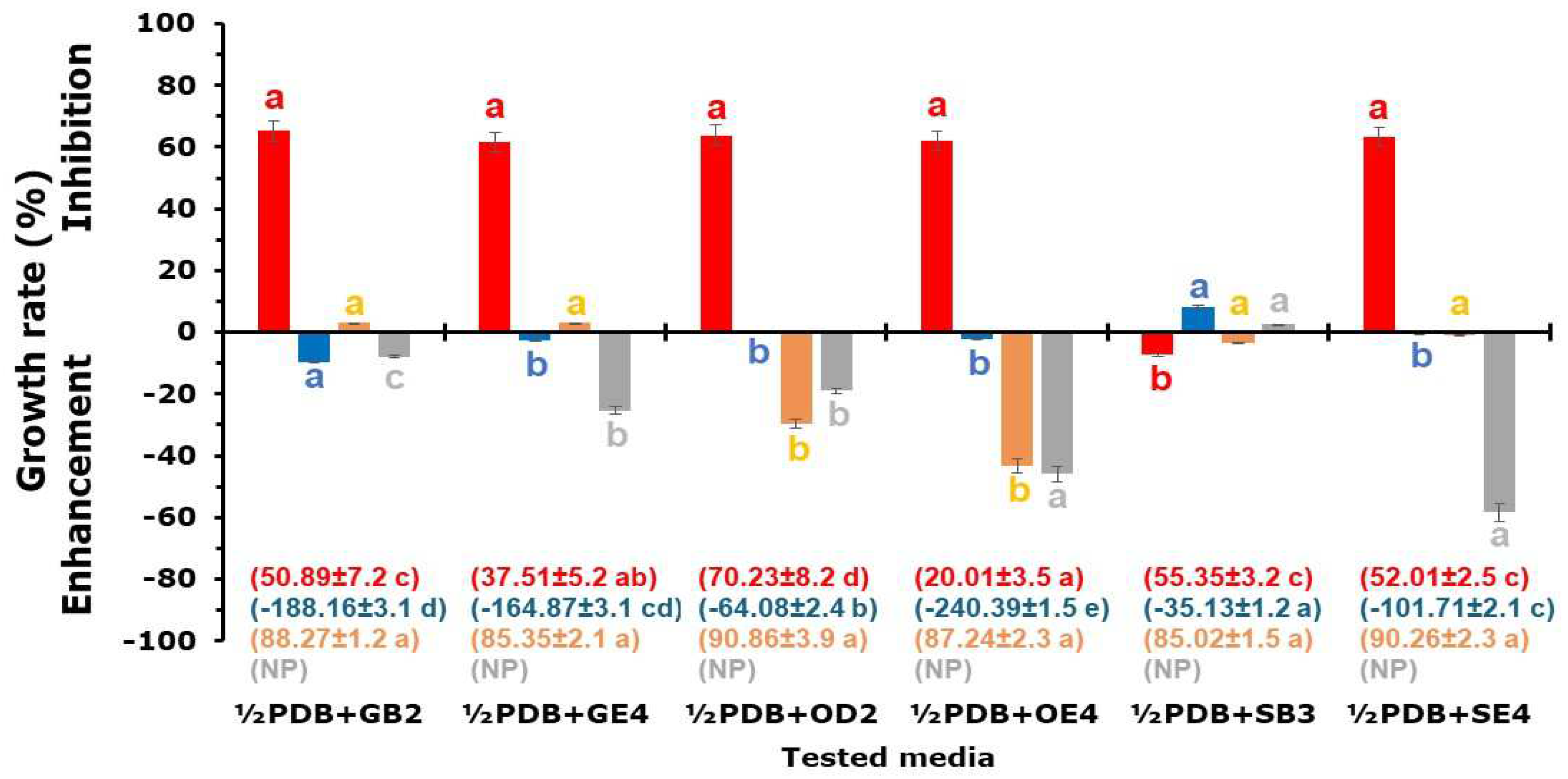
Figure 3.
Percentage of radial growth inhibition/enhancement of Fusarium oxysporum f. sp. lycopersici (■), Verticillium dahliae (■), Sclerotinia minor (■) and Trichoderma harzianum (■) on Potato Dextrose Agar (PDA) and PDA amended with GB2 (PDA+GB2), GE4 (PDA+GE4), OD3 (PDA+OD3), OE4 (PDA+0E4), SB3 (PDA+SB3) and SE4 (PDA+SE4) fine soil (1%) samples at 25±1°C, in the dark. Histograms represent the average of six replicates ± sd. The number in brackets indicates the sporulation inhibition rate. For each fungal isolate or parameter, values marked with the same letter are not significant for Fischer’s LSD at P<0.05. Soil sample abbreviations and characteristics are in Table 1.
Figure 3.
Percentage of radial growth inhibition/enhancement of Fusarium oxysporum f. sp. lycopersici (■), Verticillium dahliae (■), Sclerotinia minor (■) and Trichoderma harzianum (■) on Potato Dextrose Agar (PDA) and PDA amended with GB2 (PDA+GB2), GE4 (PDA+GE4), OD3 (PDA+OD3), OE4 (PDA+0E4), SB3 (PDA+SB3) and SE4 (PDA+SE4) fine soil (1%) samples at 25±1°C, in the dark. Histograms represent the average of six replicates ± sd. The number in brackets indicates the sporulation inhibition rate. For each fungal isolate or parameter, values marked with the same letter are not significant for Fischer’s LSD at P<0.05. Soil sample abbreviations and characteristics are in Table 1.
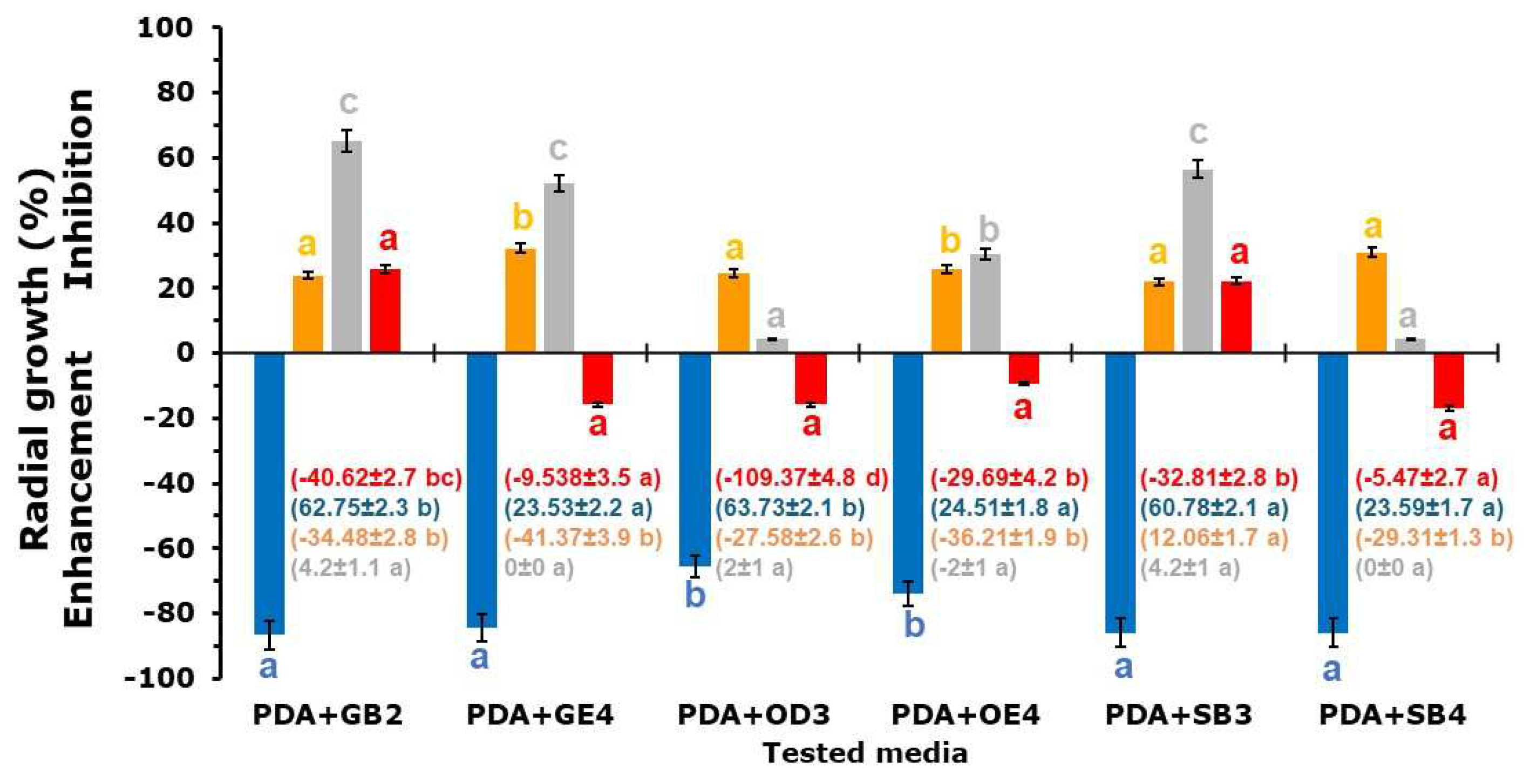
Figure 4.
Examples of the growth of Fusarium oxysporum f. sp. lycopersici (FOL), Verticillium dahliae (VD) and Sclerotinia minor (SM) in the presence of agar plugs containing the filtrate of Trichoderma harzianum (TH) obtained from liquid cultures (8 days, 25±1 °C, 100 rpm) in half strength Potato Dextrose Broth (½PDB; TH½PDB) and ½PDB modified with the addition of fine soil (1%) of samples GB2 (TH½PDB-GB2), GE2 (TH½PDB-GE2), OD3 (TH½PDB-OD3), OE4 (TH½PDB-OE4), SB3 (TH½PDB-SB3) and SE4 (TH½PDB-SB4). ½PDB-NI = control ½PDB not inoculated. For soil sample abbreviations and characteristics see Table 1. The red numbers indicate the percentage of plug colonization (PPC) calculated as PPC=100×(PCon-PTHCF)/PCon where PCon and PTHCF represent the number of colonized plugs containing PDB and TH CF, respectively.
Figure 4.
Examples of the growth of Fusarium oxysporum f. sp. lycopersici (FOL), Verticillium dahliae (VD) and Sclerotinia minor (SM) in the presence of agar plugs containing the filtrate of Trichoderma harzianum (TH) obtained from liquid cultures (8 days, 25±1 °C, 100 rpm) in half strength Potato Dextrose Broth (½PDB; TH½PDB) and ½PDB modified with the addition of fine soil (1%) of samples GB2 (TH½PDB-GB2), GE2 (TH½PDB-GE2), OD3 (TH½PDB-OD3), OE4 (TH½PDB-OE4), SB3 (TH½PDB-SB3) and SE4 (TH½PDB-SB4). ½PDB-NI = control ½PDB not inoculated. For soil sample abbreviations and characteristics see Table 1. The red numbers indicate the percentage of plug colonization (PPC) calculated as PPC=100×(PCon-PTHCF)/PCon where PCon and PTHCF represent the number of colonized plugs containing PDB and TH CF, respectively.
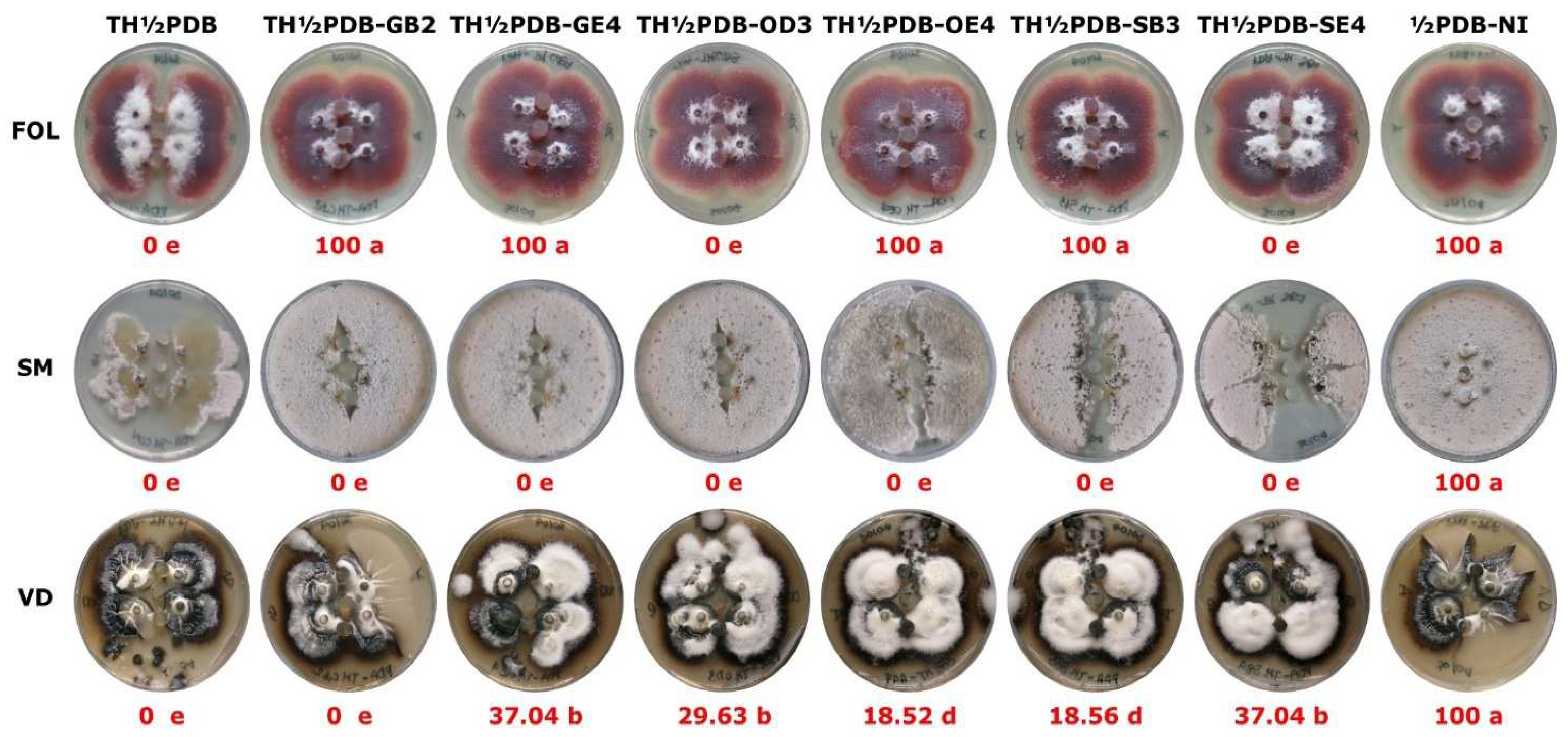
Figure 5.
Pearson correlation coefficient among the pollutant total concentration (A), modified contamination degree index (B) or Nemerow integrated risk index (C) and pH culture filtrate, percentage of growth inhibition rate (PGIR), or sporulation inhibition rate in liquid (SIR-L) and agarized (SIR-A) cultures of Fusarium oxysporum f. sp. lycopersici (FOL), Verticillium dahliae (VD), Trichoderma harzianum (TH) and Sclerotinia minor (SM) grown in half-strength Potato Dextrose Broth or Potato Dextrose Agar amended with 1% fine soil from GB2, GE2, OD3, OE4, SB3 or SE4 soil samples. Interpretation of the reference colour (D). “*” and “**” indicate statistical significance at P≤0.05 and P≤0.01, respectively. a MNP = Microsclerotia not produced. Soil sample abbreviations and characteristics are in Table 1.
Figure 5.
Pearson correlation coefficient among the pollutant total concentration (A), modified contamination degree index (B) or Nemerow integrated risk index (C) and pH culture filtrate, percentage of growth inhibition rate (PGIR), or sporulation inhibition rate in liquid (SIR-L) and agarized (SIR-A) cultures of Fusarium oxysporum f. sp. lycopersici (FOL), Verticillium dahliae (VD), Trichoderma harzianum (TH) and Sclerotinia minor (SM) grown in half-strength Potato Dextrose Broth or Potato Dextrose Agar amended with 1% fine soil from GB2, GE2, OD3, OE4, SB3 or SE4 soil samples. Interpretation of the reference colour (D). “*” and “**” indicate statistical significance at P≤0.05 and P≤0.01, respectively. a MNP = Microsclerotia not produced. Soil sample abbreviations and characteristics are in Table 1.
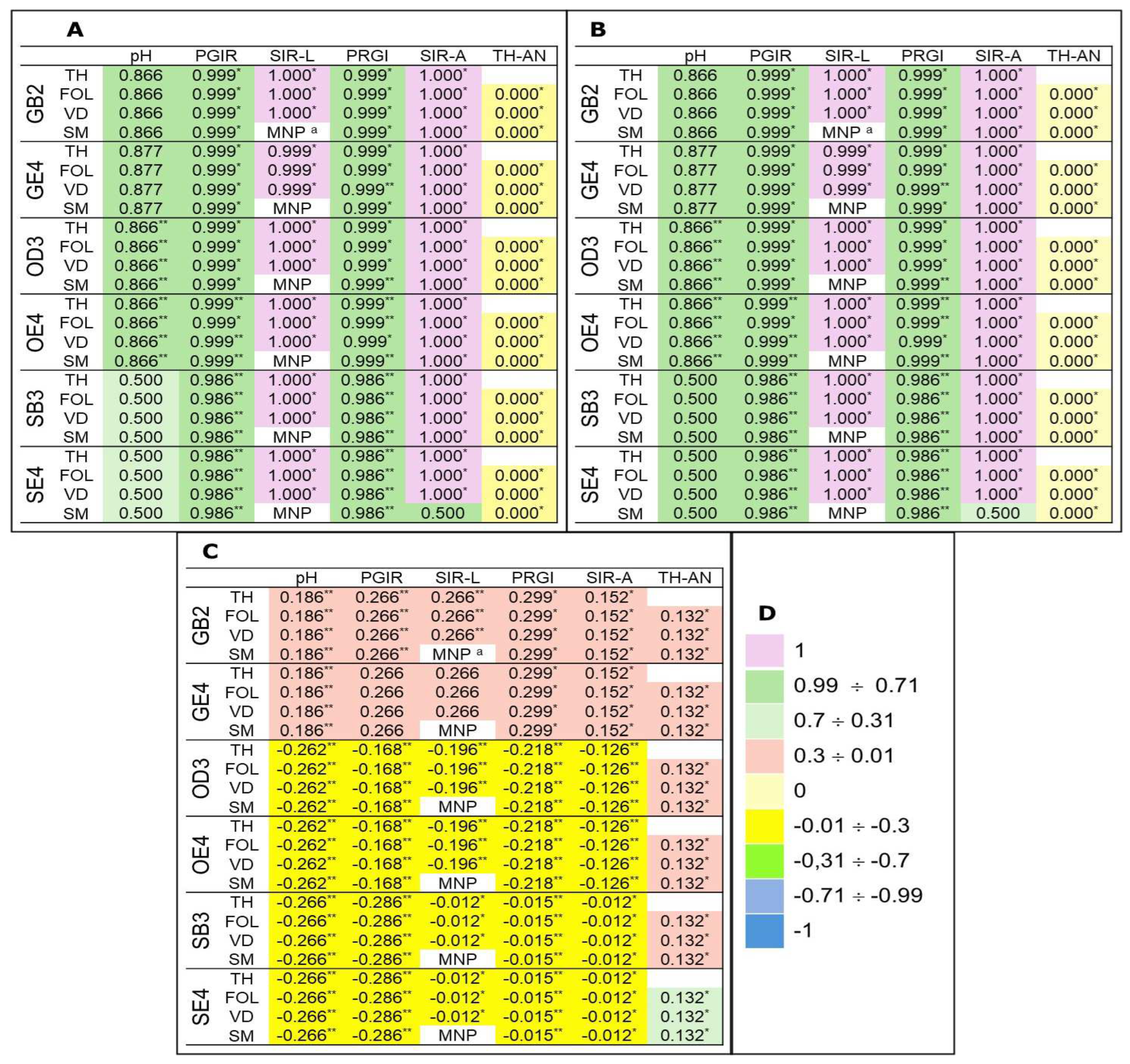
Table 2.
Values of pH, modified contamination degree index (mCId) and Nemerow Integrated Risk Index (NIRI) of soil samples collected from dismissed bauxite mines in San Giovanni Rotondo (GB2, GE4), Otranto (OD3, OE4), and Spinazzola (SB3, SE4).
Table 2.
Values of pH, modified contamination degree index (mCId) and Nemerow Integrated Risk Index (NIRI) of soil samples collected from dismissed bauxite mines in San Giovanni Rotondo (GB2, GE4), Otranto (OD3, OE4), and Spinazzola (SB3, SE4).
| Parameter | Soil samples a,b | |||||
| GB2 | GE4 | OD3 | OE4 | SB3 | SE4 | |
| pH | 8.34±0.12a | 8.24±0.15a | 8.01±0.11ab | 7.89±0.13b | 8.04±0.14ab | 8.42±0.12a |
| mCId | 3.891±0.52d | 1.999±0.20e | 5.324±0.21b | 4.555±0.07c | 10.448±0.94a | 6.703±0.46b |
| NIRI | 2.934±0.27c | 2.934±0.27c | 15.533±0.45b | 15.533±0.49b | 18.732±0.48a | 18.732±0.48a |
a For each parameter, values marked with the same letter are not significant for Fischer’s LSD at P<0.05. b Soil sample abbreviations and characteristics are in Table 1.
Table 1.
Final pH of Fusarium oxysporum f. sp. lycopersici (FOL), Verticillium dahliae (VD), Trichoderma harzianum (TH) and Sclerotinia minor (SM) cultures in half strength Potato Dextrose Broth (½PDB) or ½PDB amended with GB2, GE2, OD3, OE4, SB3 and SE4 soil samples a,b,c.
Table 1.
Final pH of Fusarium oxysporum f. sp. lycopersici (FOL), Verticillium dahliae (VD), Trichoderma harzianum (TH) and Sclerotinia minor (SM) cultures in half strength Potato Dextrose Broth (½PDB) or ½PDB amended with GB2, GE2, OD3, OE4, SB3 and SE4 soil samples a,b,c.
| Media | ½PDB NId | FOL | VD | TH | SM |
| ½PDB | 5.61±0.1 a | 5.13±0.1 a | 4.54±0.1 a | 3.82±0.1 a | 3.82±0.1 a |
| ½PDB+GB2 | 6.02±0.1 b | 6.18±0.1 b | 5.06±0.1 ab | 4.21±0.1 ab | 4.22±0.1 ab |
| ½PDB+GE4 | 5.99 ±0.1 b | 6.11±0.1 b | 5.11±0.1 b | 3.93±0.1 a | 3.91±0.1 a |
| ½PDB+OD3 | 6.08±0.1 b | 5.18±0.1 a | 4.59±0.1 a | 4.21±0.1 ab | 4.21±0.1 ab |
| ½PDB+OE4 | 6.02±0.1 b | 5.08±0.1 a | 4.94±0.1 a | 3.85±0.1 a | 3.85±0.1 a |
| ½PDB+SB3 | 6.06±0.1 b | 5.12±0.1 a | 5.01±0.1 ab | 4.61±0.1 b | 4.63±0.1 b |
| ½PDB+SE4 | 6.02±0.1 b | 6.11±0.1 b | 4.85±0.1 a | 3.76±0.1 ab | 3.73±0.1 a |
a Each value represents the average of six repetitions ± sd; b Soil sample abbreviations and characteristics are in Table 1; c For each column, values with the same letter are not significant for Fischer’s LSD at P<0.05; c NI = Not Inoculated.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



