Submitted:
28 May 2025
Posted:
29 May 2025
You are already at the latest version
Abstract
Introduction: Kala-azar is a protracted disease caused by the zoonotic protozoa Leishmania infantum and by the anthroponotic L. donovani, transmitted by sandflies. Fever, anemia, and hepatosplenomegaly may be followed by hemorrhages and bacterial infections. The pathogenesis is related to exaggerated cytokine release that may provoke death. Patients and methods: Seventy-two hospitalized patients were studied. Clinical data and mortality were analyzed regarding the blood and bone marrow L. infantum load, and plasma IL-1, IL6, IL-8, IL10, IL-12, TNF-, and TGF- concentrations. Results: Cytokines were high. Blood and bone marrow L. infantum kDNA were correlated and increased with the time of disease. Blood and bone marrow load were higher in men, adults, and HIV-infected patients, and were strongly associated with mortality. IL-6 was independently associated with sepsis. In the multivariate analysis, IL-12 was the only one associated with blood L. infantum load, and the correlation was negative. Conclusions: Since L. infantum loads were associated with disease severity, but plasma cytokine levels were not, other mechanisms must be considered to explain disease progression leading to death. Plasma IL-12 appears to reduce blood parasite load in L. infantum kala-azar, suggesting the presence of a persistent, albeit weak, acquired immune response that is ultimately overwhelmed by a progressive, inflammatory, and inefficient innate systemic response.
Keywords:
kala-azar
; visceral leishmaniasis
; Leishmania infantum
; pathogenesis
; cytokines
; immunity
; parasitism
; mortality
1. Introduction
Kala-azar, or visceral leishmaniasis, is a lethal parasitic disease with protracted symptoms. Most patients report low-grade fever. Patients also report inappetence, diarrhea, and weight loss and appear malnourished and anemic, with enlarged spleen and liver [1]. Jaundice, vomiting, and emaciation, signs of bleeding and, together with bacterial infections, are frequent and may lead to death [2,3]. The disease is commoner and is more lethal at the extremes of age [4,5]. Immunosuppressed patients, are also at higher risk [6,7].
Two species of protozoa are the cause of disease. Leishmania donovani is the agent in East Africa and South Asia and is restricted to humans. Currently, L. donovani kala-azar has been put under control in South Asia through a set of well-coordinated control measures [8]. L. infantum causes the disease among humans and other mammals in Central and Western Asia, the Mediterranean Basin, and the Americas. Differently of L. donovani kala-azar, the incidence of L. infantum kala-azar remains untouched by control measures [9]. Transmission occurs mainly by the bite of several species of sandflies. The flagellar promastigotes develop in the insects and in culture media. After the infective bite, promastigotes are phagocytized by neutrophils and monocytes. They lose the flagella and survive as amastigotes in macrophages distributed in the spleen, in the liver Kupffer cells, bone marrow, lymph nodes [10,11,12]. They circulate in the blood inside monocytes and neutrophils in very low concentration [13,14].
There are no known Leishmania virulence factors that can directly harm mammal host cells or tissues and cause disease [3]. Instead, they lead to the sickness through host response as long as they surpass the host’s innate and acquired defenses and progressively multiply, generating the typical signs and symptoms and, eventually, death [15]. As it happens with other infectious diseases, systemic inflammation is triggered by pro-inflammatory cytokines while regulatory cytokines limit inflammation and presumably immunity [11,16]. This wave of cytokines is associated with hemorrhagic manifestations, and bacterial infections [17].
How precisely the parasites and the host are intertwined to develop the lethal disease phenotype has been investigated but is still a matter of conjecture [18]. Evaluations of the role of the host showed that cytokines such as IL-6, IL-8, and INF-g, sCD14 might be involved in more life-threatening disease [17,19]. Similarly, the larger the L. infantum load, the more severe the disease is [20,21]. Interestingly, the L. infantum genome explains around 80% of the mortality of human patients, suggesting a complex parasite interaction with host response [22]. However, it is not known how the interplay of parasites and host molecules is for generating the outcome of death even after prompt diagnosis and treatment. Mortality remains 10% over the years and, it is increasing in some parts of the world [23]. Therefore, this study was developed to shed light on the connections between L. infantum burden, a set of host molecules, life-threatening kala-azar, and immunity, in order to better understand the pathogenesis of severe kala-azar.
2. Materials and Methods
2.1. Patients
All kala-azar patients were treated at the “Natan Portella” Tropical Disease Institute in Teresina, Brazil, and confirmed kala-azar cases by the presence of L. infantum amastigotes on bone marrow smears or the presence of promastigotes in culture were included in this study. The study was performed on a sample of 72 patients, sequentially admitted with available clinical information taken by a single physician and with cryopreserved plasma, blood, and bone marrow samples obtained before treatment.
2.2. Medical Data
The following parameters of VL patients were analyzed: age, sex, history of vomiting, somnolence, inappetence, diarrhea, abdominal pain, cough, shortness of breath, oliguria, and presence of fever > 39 °C, pallor, irritability, edema, bleeding, jaundice, dyspnea, lung X-ray abnormalities, urinary tract infection, renal failure, pneumonia, sepsis, HIV-1 infection, spleen and liver size, and complete blood count. Fever was present in all but one patient and, therefore, was not analyzed. The registered bleeding variables included any history of hemorrhagic manifestations on admission or during hospital stay, such as gingival or gastrointestinal bleeding, hematoma at venoclysis site, petechiae, and bruise. The variables relative to hemorrhages were collapsed into two variables: “reported bleeding” and “detected bleeding”. The first was informed by the patients and the second was observed by the medical team. A patient was taken as having pneumonia when typical symptoms and signs of pneumonia were present in the X-ray. Bacterial cultures were routinely requested, but many patients were already receiving antibiotic therapy. Hyperventilation, tissue hypoperfusion, venous saturation, oliguria, and altered consciousness defined sepsis. Infections were divided into two categories: “sepsis” and “any bacterial infection”. “Sepsis” refers to those patients who fulfilled the criteria described above, while “any bacterial infection” described the diagnosis of any type of bacterial infection.
Since the number of deaths was too small to detect statistically significant clinical associations with severe disease, the score system software Kala-Cal® was also used as a proxy of disease gravity. The system uses data such as edema, jaundice, dyspnea, HIV-coinfection, vomiting, bacterial infections, and hemorrhages, and, indeed, it was significantly associated with mortality. The software has been published elsewhere [5] and can be easily reached at https://www.sbmt.org.br/kalacal/. In synthesis, disease severity was evaluated by six variables: “death”, “chance of death > 10% by using Kala-Cal®”, “reported bleeding”, “detected bleeding”, “sepsis”, and “any bacterial infection”.
2.3. DNA Isolation, Purity, and Standardization
Bone marrow and blood samples were collected both in heparinized and in citrated tubes. Isolated parasites were stored in liquid nitrogen. DNA isolation was performed by the QIAmp DNA Blood Mini Kit (Qiagen, Germany) according to the manufacturer’s instructions with 200.0 μL of bone marrow or blood. The purity and DNA concentration was evaluated in a spectrophotometer (NanoDrop ND-1000 spectrophotometer; Thermo Fisher Scientific, Waltham, MA). DNA samples were normalized to a concentration of 5 ng/μL, concentrated or diluted with distilled water when necessary.
2.4. Quantitative PCR
Quantitative PCR was based on the TaqMan probe to quantify L. infantum on blood and bone marrow accurately. The target sequence for L. infantum detection consisted of FAM – TTT TGA ACG GGA TTT CTG – MGB-NFQ (GenBank AF169140). Specific primers based on kinetoplast DNA consisted of 5’ – GGC GTT CTG CAA AAT CGG AAA A – 3’ (forward) and 5’ - CCG ATT TTT GGC ATT TTT GGT CGA T – 3’ (reverse), (Applied Biosystems, Foster City, CA, USA) were used (Zacarias et al.,2017). A standard curve was constructed using 10-fold serially diluted L. infantum DNA corresponding to 104 to 1 parasite per reaction.
Albumin was selected as the housekeeping gene to equalize the parasite count in the bone marrow. The number of parasites was expressed as a ratio to the number of human nucleated cells. The primers were 5’ - GCT GTC ATC TCT TGT GGG CTG T – 3’(forward) and 5’ – ACT CAT GGG AGC TGC TGG TTC – 3’ (reverse). The probe was VIC- GG AGA GAT TTG TGT GGG CAT GAC A –TAMRA (GenBank NG009291) (Verma et al., 2010). The standard curve was constructed using 10-fold serially diluted human cells DNA corresponding to 2 x 104 to 2 human nucleated cells per reaction.
Amplification and detection were performed in StepOne™ Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). Cycling parameters were 50oC for 2 minutes, 95oC for 10 minutes, and 40 cycles at 95oC for 15 s and 60oC for 1 minute. Standards, samples, and negative controls were analyzed in duplicates. The threshold cycle (Ct) value was plotted by the standard curve. The cut-off between reactions was 20%, and no deviation proportions were considered.
2.5. Plasma Cytokines
The plasma specimens were stored at - 20oC. Serum IL-1β, IL-6, IL-8, IL-10, TNF-α, IL-12 were measured using a high-sensitivity multiplex inflammatory cytokine panel by cytometric bead array (CBA), plus cytokines IL-17 and TGF-β, which were measured independently by CBA-flex set (BD Biosciences, San Jose, CA, USA), on the BDFAcs Array (BD Biosciences, San Jose, CA, USA), following the manufacturer’s instructions. Measurements of each sample were performed in duplicate, and the average of the two measurements was used. Standard curves were derived from the cytokine standards supplied with the kit 10-fold dilution. The lower limits of detection for specific analyses ranged from 0.40 pg/mL for IL-8, 0.04 pg/mL for IL-1β, 0.04 pg/mL for IL-6, 0.97 pg/mL for IL-10, 0.03 pg/mL for TNF-α, 0.06 pg/mL for IL-12, and 0.05 pg/mL for TGF-β, based on standards curve dilution.
2.6. Statistical Analysis
Proportions and 95% confidence intervals were calculated for the clinical and demographic variables. The median and interquartile intervals, as well as the means, were calculated for kDNA loads. The Kolmogorov–Smirnov test compared the plasma cytokines with the standard values taken from the literature [24]. The Wilcoxon rank-sum test compared blood and bone marrow kDNA load according to demographic and clinical data. Similarly, the test was used to analyze the concentration of cytokines. Pearson’s correlation test evaluated the correlation between death, “risk of death > 10% by Kala-Cal®”, “reported bleeding”, “detected bleeding” “sepsis”, and “any bacterial infection”. The Spearman’s correlation test was used to test the correlation between the natural logarithm of blood and bone marrow kDNA burdens and their correlation with the time of disease, as indicated by the time with fever. Finally, simple and multiple linear and quantile regression were applied to test whether the cytokines predicted the blood and bone marrow kDNA load. The statistical package Stata/IC 15.1 (College Station, TX) was used to analyze the data.
3. Results
3.1. Study Population
Table 1 shows the characteristics of the study population. Forty-two patients were male (58.3%) and 30 female (41.7%). The median age was 7.5 and the mean was 15.2 years. Seventeen participants (23.6%) were under two, 22 (30.6%) were under four, and 61 were children under 15. Six were older than 40 (8.3%). Among the 70 who were tested for HIV, 13 were positive (18.6%). Four (5.6%) patients died. Male sex was associated with HIV infection (p = 0.032).
3.2. Clinical Findings
Table 1 also shows the clinical characteristics of the study population. The mean Kala-Cal® chance of death was 12.1%, and the median was 6.5. Twenty-five patients had a chance of death of at least 10% (34.7%). More than 40% had hemorrhages or infections. Fifteen patients had “reported bleeding” manifestation (20.8%). Ten patients filled the criteria for sepsis (14.1%). Four patients had some sort of bleeding at hospital stay e.g., “detected bleeding” (5.6%). Any bacterial infection was detected in 23 (31.9%) patients. Mortality was associated with “detected bleeding” (p = 0.014) and with sepsis syndrome (p = 0.008), as well as with any bacterial of infection (p = 0.052). The six markers of disease severity were significantly and positively correlated, except “death” with “reported bleeding” and “probability of death > 10%” with “sepsis” or “any bacterial infection”, which were positively but not statistically correlated. The supplemental table informs the correlation matrix between these variables.
The chance of death calculated by Kala-Cal® was well correlated with the occurrence of death (r = 0.40, p < 0.001). As expected at admission, the estimated chance of death among those who survived was 8.9%, much less than chance of those who actually lately died (65.3%).
3.3. Quantity of Parasite Load and Cytokines
Table 2 depicts the blood and bone marrow L. infantum load, plasma cytokine concentrations, and reference values. The median blood kDNA concentration was 856.7 kDNA amastigote-equivalents/mL (AEq/mL), and the mean was 3,515.4 AEq/mL. The median bone marrow kDNA concentration was 55.7 AEq/106 DNA equivalents of human cells (HCEq), with IQ 3.7 – 400.7 AEq/106HCEq. The standard deviation was 1.7 larger than the mean blood kDNA, while the same relationship was 3.2 for bone marrow kDNA, demonstrating the higher variability of the bone marrow count. The standard values were assumed to be zero, although some asymptomatic patients may harbor minimal blood kDNA load [25,26,27,28]. Regarding the cytokines, IL-6, IL-8, IL-10, and TGF-b had medians and means much above the values of a healthy population, while IL-1b and IL-12 had a more modest increase. Despite the high plasma concentration, cytokines were not detected in a relevant proportion of patients: 28% for IL-12, 19% for IL-1b, 9% for TNF-a, and 6% for IL-6.
1Parasite-equivalents. 2AEq/106 equivalents of human cells. 3Kildey et al, 2014. 4Grainger et al, 2000. 5Not available. 6Assumed to be zero in persons without visceral leishmaniasis.
3.4. Blood and Bone Marrow L. infantum Load
The Spearman’s correlation test revealed a moderate, but statistically significant correlation between the natural logarithm of blood and the natural logarithm of bone marrow parasite load (r = 0.48, p < 0.001) (Figure 1). Bone marrow kDNA load without including human cell counts as the denominator in the parameter was poorly correlated with the parasite blood load (r = 0.21, p = 0.077).
3.5. Time of Disease, Blood and Bone Marrow L. infantum Load, Plasma Cytokines, and Severity
The blood and bone marrow L. infantum kDNA load increased with the time of disease, as estimated by the duration of fever (Figure 2). The Spearman’s correlation coefficient for blood kDNA with time was 0.33 (p < 0.005), and for the bone marrow, it was 0.38 (p = 0.001). There was no correlation of any cytokine with time. At the univariate linear regression analysis of the relationship of the natural logarithm of time with fever with the natural logarithm of blood kDNA load, the probability of rejecting the model was < 0.005, and the p-value of the coefficient was 0.005. For predicting the natural logarithm of the bone marrow load, the probability of rejecting the model was < 0.001, and the p-value of the coefficient was < 0.001. There was no association of any cytokine with the time of disease. Higher chance of death and “detected bleeding” were slightly correlated with the time of fever (r = 0.23, p = 0.056 and r = 0.24, p = 0.044, respectively).
3.6. Blood and Bone Marrow L. infantum Load, Age, Sex, HIV Infection, and Kala-Azar Severity
Table 3 shows the relationships between the L. infantum load with age, sex, HIV infection, and markers of kala-azar severity. Individuals with 15 years of age or older had higher blood median load (p = 0.022), and patients 40 years old or older had the highest blood load (median = 9,129.5 AEq/mL) (data not shown). The same happened in the bone marrow (p = 0.043). The blood and bone marrow load were higher in men, p = 0.012 and p = 0.073, respectively. HIV-infected patients had eight times higher blood load than those non-infected (p = 0.006). On the other hand, in bone marrow, although higher, this difference was non-significant. At the multivariate quantile regression, HIV status did not stand out, and only sex and age were associated with blood kDNA load.
The four patients who died had kDNA in the blood with a median 14.2 times higher than the survivors but non-significantly associated to death (p = 0.169), likely due to the small number of deceased patients. The bone marrow L. infantum load was also higher in the deceased in those who survived, but not statistically significant. The concentration of kDNA in the blood was 11.4 times higher in those with a chance of death estimated above 10% (p < 0.001). In the bone marrow, it was 14.5 times higher in those with a higher than 10% chance of death (p = 0.001).
The association of the two variables relative to hemorrhagic phenomena, “reported bleeding” and “detected bleeding”, were not concordant regarding L. infantum load in the blood and in the bone marrow. Blood kDNA was ten times higher in those four patients with “detected bleeding”, than in those with “reported bleeding”. The opposite happened in the bone marrow: L. infantum load was not that high in those with “detected bleeding” but was 10 times higher in those patients with “reported bleeding” (p = 0.12).
3.7. Plasma Cytokines, Age, Sex, HIV Infection, and Markers of Kala-Azar Severity
Table 4 shows the associations between age, sex, HIV infection, and the clinical manifestations of the severity of kala-azar with the seven measured plasma cytokines. There was a paucity of statistically solid associations between plasma cytokines and demographic and clinical data. IL-10 was almost two times higher in children (p = 0.02). IL-10 was also much higher in women than in men (p = 0.06). Similarly, IL-10 was the only cytokine associated with HIV infection, being significantly lower in those with HIV (p = 0.04). Possibly, due to the small number of deaths in the study population (four), no cytokine had any statistically significant association with death. However, the median of IL-6 was noticeably more than two times higher in those who died. The used proxy of death and disease severity, e.g., the chance of death according to the application of the Kala-Cal® software, showed no significant association with any cytokines. Only three patients with measured cytokines had “detected bleeding”. However, in these three patients, IL-6 was almost four times higher (p = 0.03). Only IL-8 was associated with “reported bleeding” (p = 0.04). Sepsis syndrome had some significant associations with cytokines. IL-12 was higher in those with the syndrome (p = 0.05), as well as nearly significant with IL-1b (p = 0.06), IL-6 (p = 0.06), and IL-10 (p = 0.07). By the end of these results, the variable “any bacterial infection” was statistically associated with IL-1b (p = 0.01), IL-6 (p = 0.01), IL-8 (p = 0.03), IL-10 (p = 0.04) and TNF-a (p = 0.01). However, no statistically significant association would exist if the Bonferroni analysis for multiple comparisons is applied. In the multivariate linear regression, IL-6 was the only predictor of sepsis (p = 0.04), confirmed in a multiple quantile regression. No cytokine was found predicting the chance of death or any other clinical outcome at the p-value <0.05 (data of regressions not shown).
3.8. Regression Analysis Between L. infantum Load and Plasma Cytokines
In the univariate linear regression analysis, IL-12 negatively and significantly predicted blood kDNA levels (p = 0.002). Similarly, TNF-α also showed a negative and significant prediction (p = 0.018). However, when controlling for IL-12, the effect of TNF-α was no longer significant. In the multivariate linear regression, only IL-12 remained a significant predictor of blood kDNA load (Figure 3); no other cytokines were associated. Finally, no cytokine predicted kDNA levels in the bone marrow. The models adjusted R2 and pseudo R2, respectively, were low, indicating that the studied cytokines were poor predictors of L. infantum load.
4. Discussion
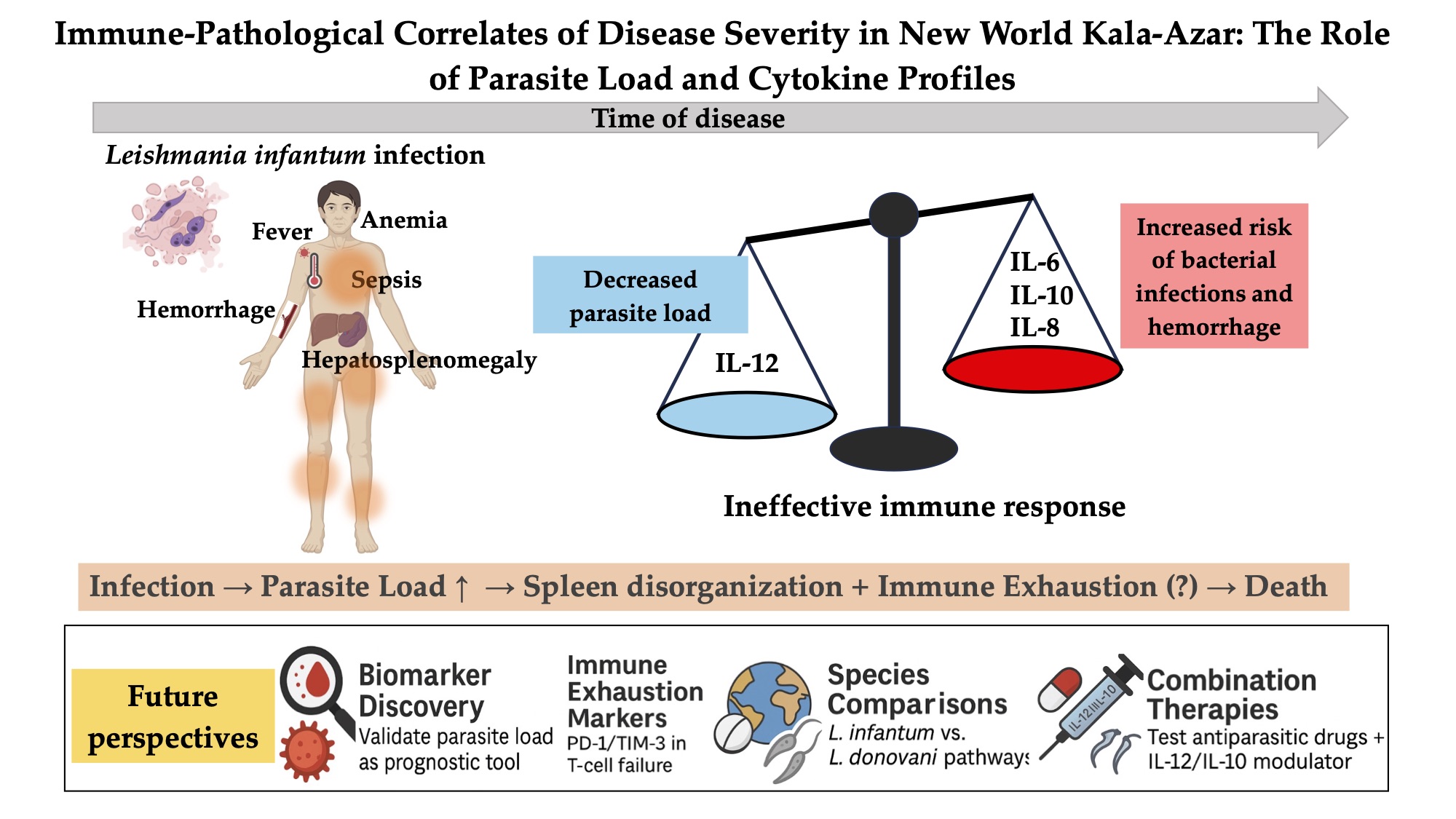
This study found that patients with kala-azar have a high inflammatory status and have signs of substantial risk of death. On the other hand, the data also shows that children, women, and persons living with HIV have a more prominent regulatory immune response. Markedly, however, IL-1b, IL-6, IL-8, IL-10, IL-12, TNF-a, and TGF-b were not, or were poorly associated with the risk of death. Moreover, while the studied cytokines did not increase with time of disease, blood and bone marrow L. infantum load expanded progressively. Additionally, blood and bone marrow L. infantum load were well correlated, and the parasite load was higher among men, adults, and HIV-infected patients. Finally, higher blood and bone marrow kDNA loads were associated with increased severity. Most plasma cytokines were not related to blood and bone marrow L. infantum load, but higher plasma IL-12 was independently associated with lower L. infantum load.
The study’s weakest point was its cross-sectional design, which did not allow the identification of causal relationships, except with death. Other key cytokines and molecules, such as INF-g, were not studied. Additionally, bone marrow cytokines were not measured, and the spleen size was not regularly registered, hiding the organ’s importance to the observed values. However, the findings are of interest for helping to understand the pathogenesis of life-threatening kala-azar and to highlight the progressive immunological failure of patients with kala-azar.
This study population describes the well-known male and children predominance in kala-azar caused by L. infantum [29]. HIV coinfection was higher than usual for the region and the country [30,31]. Although mortality was lower in this sample, more than one-third of the patients had a chance of death estimated over 10% and hemorrhages and bacterial infections. These findings demonstrate how severe the disease can be and stress the importance of understanding the pathogenetic mechanisms of these complications. When compared with blood donors [24], plasma cytokine concentrations where very high, confirming the role of inflammation. Indeed, previous works have already shown very high values plasma cytokines in patients in East-Africa, Brazil and India for INF-g, IL-8, TGF-b, L-10, and IL-6 [32,33,34,35].
Children had higher concentrations of most cytokines than older patients, but only IL-10 reached statistical significance. This finding might support a previous observation and suggests a more immunotolerant state of children, independently of kala-azar [36]. Regarding sex, IL-10 was higher in females. Although another study in healthy adult individuals could not observe this difference, the present findings suggest that a sex-dependent IL-10 response in patients with kala-azar may actually exist [37].
Another previous work has shown that HIV-infected patients under antiretroviral therapy (ART) had lower IL-10 than patients, both in those patients not on ART and in those long-term non-progressors [38,39]. In the present study, IL-10 was lower in patients coinfected with HIV than in those non-coinfected, suggesting that HIV infection attenuates the regulatory innate response in kala-azar.
Based on observational data, it has been proposed that plasma IL-6 and IL-8, INF-g, IL-27, and soluble CD14 are the major mediators of the pathogenicity of kala-azar [3,17,19]. Due to its overlapping actions with characteristics of kala-azar, such as hemorrhages, anemia, hypoalbuminemia, and hyperglobulinemia, IL-6 was proposed to be the best explanatory cytokine for the complications of kala-azar [40]. However, in the present study, neither the chance of death at hospital admission nor the occurrence of death, revealed associations of plasma cytokines with signs of severity strong enough. Indeed, when the p-values were corrected by the Bonferroni method for multiple comparisons, no association between clinical presentation and any of the seven studied cytokines was found. Additionally, at the multivariate linear regression, no cytokine was found predicting the chance of death. This set of data on the role of cytokines detected in the blood suggests that the core of the pathogenic phenomena that lead to severe kala-azar or death may not rely primarily on the direct effect of pro- or anti-inflammatory cytokines as previously proposed [17,19].
This study observed that the cytokine response does not change with time after the disease starts, and this finding may influence the comprehension of the framework for the pathogenesis of kala-azar. Although cytokines did not change, L. infantum load and disease severity increased with time. The phenomenon does not look to be determined by progressive spleen enlargement since parasite load in the bone marrow also increased similarly.
The median blood and bone marrow amounts of L. infantum kDNA in this sample were in the range of the concentrations previously found in Teresina and elsewhere using the same protocol [20,21,41]. As expected, blood and bone marrow kDNA concentrations correlated well, indicating that bone marrow is balanced with systemic parasitism. Men and adults had higher blood and bone marrow loads, likely due to the modulatory effect of testosterone and dihydrotestosterone on men, who are the majority of adult patients with kala-azar [42]. Interestingly, HIV infection was not independently associated with a higher load. One possible explanation is that the patients with HIV had a relapsing course of kala-azar and, therefore, were under secondary prophylaxis with liposomal amphotericin B for L. infantum to prevent kala-azar relapses.
The disease severity associated with L. infantum load is a relevant finding and deserves further discussion. Unfortunately, the study design cannot assess the direction of causality: if L. infantum load worsens the disease via a linkage with a specific, unknown, factor or if a broad ongoing multifactorial lymphoid disruption leads to a non-specific, progressive and generalized failure of immunity and then to higher parasite load.
The direct cytokines’ contributions to the manifestations of severe have already been studied [17,19], but here, blood cytokines were also compared with blood and bone marrow L. infantum kDNA load, and with clinical presentation, and risk of death. IL-12 and TNF-a were found to be associated with blood L. infantum load, but IL-12 was the only one to be an independent predictor of blood kDNA load, and, importantly, this action was to decrease the L. infantum load in a dose-dependent manner. Indeed, this finding is consistent with the canonical role of innate-immunity-derived IL-12, which promotes antigen-specific Th1 response by T-cell activation, proliferation, and differentiation through secretion of INF-g [12,43,44,45,46,47,48]. Unfortunately, this finding had not been forecasted, and INF-g was not measured in the present study. However, previous two articles analyzed the association of plasma cytokines with the blood load of L. donovani kala-azar in India and Africa [49,50]. They found a positive correlation only with IL-10, TGF-b, and IL-17 but not with IFN-γ, TNF-α, IL-6, IL-4, IL-2, IL-12, and IL-22, suggesting that immunoregulatory cytokines are the primary controllers of blood parasite load in L. donovani kala-azar. However, although Teles et al [51], in Brazil, and van Dijk et al[35], in Uganda, also found a positive correlation with IL-10, they identified a negative correlation with IFN-γ. The difference of the data from India with the findings in Brazil and Uganda suggests that L. infantum and Indian L. donovani differ on the host control of kala-azar: while for the disease caused by L. infantum and by Ugandan parasites it seems to persist a more prominent role of sustained, acquired, type Th1 response, in Indian L. donovani innate regulatory cytokines “deal the cards” and play an absolute permissive role on parasite load.
IL-12/IL-10 interaction is the line of balance in kala-azar: at the infection site, intracellular amastigotes’ molecules drive the infected macrophages for a predominant IL-12 or IL-10 synthesis [10,15]. In the majority of infections, Th1 response prevails, and stimulates T-cells to synthesize INF-g, which activate macrophages to produce free radicals that kill the amastigotes, and the infection becomes controlled [11,12,52,53]. In a small proportion of immunocompetent humans, IL-10 prevails and disrupts the macrophage IL-12 signaling to CD4+ and CD8+, blocking the secretion of INF-g and preventing macrophage activation for intracellular defense [47,53,54,55,56,57]. All these events may happen entirely at the innate immunity level. However, with time, acquired immunity develops. If there is a Th1-type response with T-cells secreting INF-g, memory T-lymphocytes generate Leishmania-specific clones, and the host becomes immune, as seen by the high proportion of persons with cellular immunity to Leishmania who had never developed symptoms and lived in endemic areas [25,58]. However, if a regulatory profile is maintained, parasite load increases, disease and complications appear, and the host eventually dies. This study shows that despite the dominance of IL-10 effects on L. donovani kala-azar, acquired immunity persistently influences parasitism in L. infantum and in Ugandan kala-azar.
Therefore, the IL-12 response in L. infantum kala-azar reported here and ING-g elsewhere suggest that an overwhelmed but enduring acquired immunity persists for a while during disease. Another reason for this hypothesis is that IL-12 is maintained for an extended period after cure, compared to the cytokines secreted after stimulation of the innate response, such as the rapid fall – one to two weeks – of the cytokines mentioned above, as previously published [35,59]. Therefore, IL-12 seems to be part of acquired immunity, not of innate immunity, since it is long-lasting, due to the memory T-cells developed after earlier antigenic priming of T-cells, while IL-1, IL-6, IL-8, IL-10, and TNF-a, albeit at higher concentrations, last only during the antigenic stimulus, as characteristically occurs with innate immunity. Nevertheless IL-10 has been described associated with T-regulatory cytokines as part of the acquired immunity, it typically falls to very low levels after the cure of kala-azar [33,34,35,60].
Similarly, the early fall of INF-g in the study of LIMA et al [59] and van DIJK et al [35] suggests that most plasma INF-g is originated from cells belonging to the innate immunity such as neutrophils, eosinophils, NK cells, or even T-cells in an antigen-independent process [33,61,62], not by TCR antigen-specific CD-4+ and CD8+ T-cells. Therefore, this study suggests that in kala-azar, innate and acquired immunity coexists with disease. With treatment, the hidden cellular immunity is established but relapses indicate that the parasite persists even after immunity develops and patients become asymptomatic, e.g., non-sterile immunes. This equilibrium may have advantages for both: long-lasting immunity at the cost of some chance of parasite remission and transmission.
While L. infantum kala-azar remains with signs of some effective defense, L. donovani kala-azar not. The two parasites cause remarkable phenotypic differences, nevertheless being relatively genetically similar. L. infantum kala-azar seems to derive from the older L. donovani kala-azar[63]. While L. donovani kala-azar is mostly transmitted among humans, L. infantum kala-azar is zoonotic with a breadth of mammal hosts [1]. L. donovani kala-azar develops in older individuals, while L. infantum kala-azar hits younger immunocompetent persons, primarily children, signaling its higher force of infection [29]. L. donovani kala-azar leads to post-kala-azar dermal leishmaniasis, which is rare with L. infantum [64]. There are other clinical and epidemiological differences between the two species, but comparative, well-controlled, head-to-head studies between the two species and places still need to be done, from genomic analysis to innate and acquired immunity and pathogenesis.
Another open question from this study is why bone marrow L. infantum load did not show a similar association with IL-12 or TNF-a as blood load did. One explanation is that bone marrow control of Leishmania parasitism has in situ peculiarities not captured by the measured plasma cytokines. An insightful study investigated the association between bone marrow cytokines and local L. infantum burden. The findings revealed that IFN-γ was associated with a reduction in bone marrow parasite load, whereas interleukin-10 (IL-10) correlated with an increase in parasite burden. Interestingly, a strong and statistically significant positive correlation between IFN-γ and IL-10 levels was also observed. However, IL-12 or TNF-a were not found associated with parasite burden [51]. These finding illustrates the strong opposing effects of concomitant, protective versus permissive, cytokines each one downregulating (or upregulating) the other, in the bone marrow. It was not clear if the local findings in bone marrow can be generalized for the blood.
Since cytokines are pleiotropic and redundant, with synergistic actions, it is difficult to understand their isolated role in diseases [56]. Here, the risk of death and complications were not firmly associated with specific cytokines, except the association of IL-6 with sepsis in the multivariate regression analysis. However, since sepsis is the main cause of cytokine storms [56,65], it is not valid to infer that the rise of IL-6 was due to L. infantum instead of the opportunistic bacteria lipopolysaccharide LPS. Remarkably, mortality and the chance of death were not associated with any plasma cytokines. Although INF- was not measured in this study, it does not seem to be a candidate for kala-azar severity since its side effects are mild to the human host and do not match with the symptoms of complicated kala-azar [66]
Therefore, the question is how a microorganism without virulence factors that do not directly harm human cells or tissues would lead to death [15,18]. The progressive rise of parasite load indicates a failing acquired immunity, suggesting that a higher parasite load is a consequence and not a cause of immune failure. Two distinct mechanisms without the direct and specific action of cytokines may be pointed out to explain the global defense failure. One is T-cell exhaustion, and the other is spleen disorganization. T-cell exhaustion is a dysfunction of T-cells, mostly CD8+ cells, occurring naturally and during chronic infections and cancer [67]. The other seemingly alternative is the disorganization of lymph node and the spleen architecture, a phenomenon that disrupts the white pulp structure [68]. Both mechanisms are exacerbated in kala-azar and may hypothetically result in increasing amastigote load and in a progressive acquired immune failure and, thereafter, augmented probability of complications and death [69,70,71]. These two mechanisms of immune deterioration occur simultaneously in kala-azar but a clear molecular link between them has not been identified yet [72]. They are natural processes of apparently accelerated in kala-azar by an elusive systemic factor, likely linked to the prolonged and intense inflammatory status.
In summary, L. infantum load was associated with life-threatening kala-azar, but it is unknown how. In contrast, circulating cytokines were poorly associated with phenotypes of severe disease. As expected, Il-12 has an enduring, strong, negative effect on L. infantum proliferation during kala-azar. Parasite load worsened with time but not cytokines, suggesting a cytokine-independent immunological failure process that results in progressively severe disease and death. However, there are still no key host factors leading to complications and death by kala-azar. Consequently, it may only be conjectured that the immunological long-term consequences of sustained infection and inflammation, such as persistent immune activation, may lead to immune exhaustion and overall immunological disorganization.
Author Contributions
Conceptualization, IS; and CC; Methodology, IS, DA, JA; Formal Analysis, IS., CC.; Investigation, DC, IS, DA, JA.; Resources, IS, CC.; Data Curation, IS, CC.; Writing – Original Draft Preparation, IS, CC.; Writing – Review & Editing, IS, CC, DC; Visualization, GF; Supervision, CC.; Project Administration, CC.; Funding Acquisition, CC.
Funding
This research was funded by the Conselho Nacional de Pesquisas - CNPq – number 475102/2011-8.
Informed Consent Statement
The project was approved by the Research Ethics Committee of the Federal University of Piauí (CAE: 44037015.3.0000.5314), by the precepts of the Resolution of the National Health Council-CNS 466/12, which deals with the Guidelines and Norms for Research Involving Human Subjects. All patients included in the study, or their guardians, signed an informed consent form.
Data Availability Statement
Data is available through permission given directly by the authors IS and CC.
Acknowledgments
The authors acknowledge the Instituto de Doenças Tropicais Natan Portella for the full support of the study, including patient evaluation, laboratory data use, and the infra-structure of the Laboratory of Leishmaniasis.
Conflicts of Interest
None do declare.
References
- Burza S, Croft SL, Boelaert M. Leishmaniasis. Lancet. 2018 Sep;392(10151):951–70.
- Andrade TM, Carvalho EM, Rocha H. Bacterial Infections in Patients with Visceral Leishmaniasis. J Infect Dis. 1990 Dec 1;162(6):1354–9.
- Costa CHN, Werneck GL, Costa DL, Holanda TA, Aguiar GB, Carvalho AS, et al. Is severe visceral leishmaniasis a systemic inflammatory response syndrome? A case control study. Rev Soc Bras Med Trop. 2010 Aug;43(4):386–92. 86822010000400010&lng=en&tlng=en.
- Sampaio MJA de Q, Cavalcanti NV, Alves JGB, Fernandes Filho MJC, Correia JB. Risk Factors for Death in Children with Visceral Leishmaniasis. Franco-Paredes C, editor. PLoS Negl Trop Dis. 2010 Nov 2;4(11):e877.
- Costa DL, Rocha RL, Chaves E de BF, Batista VG de V, Costa HL, Costa CHN. Predicting death from kala-azar: construction, development, and validation of a score set and accompanying software. Rev Soc Bras Med Trop. 2016 Dec;49(6):728–40.
- Akuffo H, Costa C, van Griensven J, Burza S, Moreno J, Herrero M. New insights into leishmaniasis in the immunosuppressed. Rafati S, editor. PLoS Negl Trop Dis . 2018 May 10;12(5):e0006375.
- Kurizky PS, Marianelli FF, Cesetti MV, Damiani G, Sampaio RNR, Gonçalves LMT, et al. A comprehensive systematic review of leishmaniasis in patients undergoing drug-induced immunosuppression for the treatment of dermatological, rheumatological and gastroenterological diseases. Rev Inst Med Trop Sao Paulo. 2020;62.
- Rahim S, Karim MM. The Elimination Status of Visceral Leishmaniasis in Southeast Asia Region. Acta Parasitol. 2024 Sep 20;69(3):1704–16.
- da Rocha ICM, dos Santos LHM, Coura-Vital W, da Cunha GMR, Magalhães F do C, da Silva TAM, et al. Effectiveness of the Brazilian Visceral Leishmaniasis Surveillance and Control Programme in reducing the prevalence and incidence of Leishmania infantum infection. Parasit Vectors. 2018 Dec 12;11(1):586.
- McCall LI, Zhang WW, Matlashewski G. Determinants for the Development of Visceral Leishmaniasis Disease. Chitnis CE, editor. PLoS Pathog. 2013 Jan 3;9(1):e1003053.
- Volpedo G, Pacheco-Fernandez T, Bhattacharya P, Oljuskin T, Dey R, Gannavaram S, et al. Determinants of Innate Immunity in Visceral Leishmaniasis and Their Implication in Vaccine Development. Front Immunol. 2021 Oct 12;12.
- Bogdan C, Islam NAK, Barinberg D, Soulat D, Schleicher U, Rai B. The immunomicrotope of Leishmania control and persistence. Trends Parasitol. 2024 Sep;40(9):788–804.
- Silva JC, Zacarias DA, Silva VC, Rolão N, Costa DL, Costa CH. Comparison of optical microscopy and quantitative polymerase chain reaction for estimating parasitaemia in patients with kala-azar and modelling infectiousness to the vector Lutzomyia longipalpis. Mem Inst Oswaldo Cruz. 2016 Jul 18;111(8):517–22.
- Ferreira GR, Santos-Oliveira JR, Silva-Freitas ML, Honda M, Costa DL, Da-Cruz AM, et al. Biomarkers of disease severity in patients with visceral leishmaniasis co-infected with HIV. Cytokine. 2022 Jan;149:155747.
- Al-Khalaifah, HS. Major Molecular Factors Related to Leishmania Pathogenicity. Front Immunol. 2022 Jun 13;13.
- Samant M, Sahu U, Pandey SC, Khare P. Role of Cytokines in Experimental and Human Visceral Leishmaniasis. Front Cell Infect Microbiol. 2021 Feb 18;11.
- Costa DL, Rocha RL, Carvalho RMA, Lima-Neto AS, Harhay MO, Costa CHN, et al. Serum cytokines associated with severity and complications of kala-azar. Pathog Glob Health. 2013 Mar 12;107(2):78–87.
- Costa CHN, Chang KP, Costa DL, Cunha FVM. From Infection to Death: An Overview of the Pathogenesis of Visceral Leishmaniasis. Pathogens. 2023 Jul 24;12(7):969.
- dos Santos PL, de Oliveira FA, Santos MLB, Cunha LCS, Lino MTB, de Oliveira MFS, et al. The Severity of Visceral Leishmaniasis Correlates with Elevated Levels of Serum IL-6, IL-27 and sCD14. Oliveira SC, editor. PLoS Negl Trop Dis. 2016 Jan 27;10(1):e0004375.
- Silva JM, Zacarias DA, de Figueirêdo LC, Soares MRA, Ishikawa EAY, Costa DL, et al. Bone Marrow Parasite Burden among Patients with New World Kala-Azar is Associated with Disease Severity. Am Soc Trop Med Hyg. 2014 Apr 2;90(4):621–6.
- Zacarias DA, Rolão N, de Pinho FA, Sene I, Silva JC, Pereira TC, et al. Causes and consequences of higher Leishmania infantum burden in patients with kala-azar: a study of 625 patients. Trop Med Int Heal. 2017 Jun 2;22(6):679–87.
- Grace CA, Sousa Carvalho KS, Sousa Lima MI, Costa Silva V, Reis-Cunha JL, Brune MJ, et al. Parasite Genotype Is a Major Predictor of Mortality from Visceral Leishmaniasis. Weiss LM, editor. MBio. 2022 Dec 20;13(6).
- Cota G, Erber AC, Schernhammer E, Simões TC. Inequalities of visceral leishmaniasis case-fatality in Brazil: A multilevel modeling considering space, time, individual and contextual factors. Ramos AN, editor. PLoS Negl Trop Dis. 2021 Jul 1;15(7):e0009567.
- Kildey K, Rooks K, Weier S, Flower RL, Dean MM. Effect of age, gender and mannose-binding lectin (MBL) status on the inflammatory profile in peripheral blood plasma of Australian blood donors. Hum Immunol. 2014 Sep;75(9):973–9.
- Porcino GN, Carvalho KSS, Braz DC, Costa Silva V, Costa CHN, de Miranda Santos IKF. Evaluation of methods for detection of asymptomatic individuals infected with Leishmania infantum in the state of Piauí, Brazil. Rafati S, editor. PLoS Negl Trop Dis. 2019 Jul 1;13(7):e0007493.
- Das VNR, Bimal S, Siddiqui NA, Kumar A, Pandey K, Sinha SK, et al. Conversion of asymptomatic infection to symptomatic visceral leishmaniasis: A study of possible immunological markers. Brodskyn CI, editor. PLoS Negl Trop Dis. 2020 Jun 18;14(6):e0008272.
- Chakravarty J, Hasker E, Kansal S, Singh OP, Malaviya P, Singh AK, et al. Determinants for progression from asymptomatic infection to symptomatic visceral leishmaniasis: A cohort study. Franco-Paredes C, editor. PLoS Negl Trop Dis. 2019 Mar 27;13(3):e0007216.
- Virginia Batista Vieira A, Farias PCS, Silva Nunes Bezerra G, Xavier AT, Sebastião Da Costa Lima Júnior M, Silva ED Da, et al. Evaluation of molecular techniques to visceral leishmaniasis detection in asymptomatic patients: a systematic review. Expert Rev Mol Diagn . 2021 May 4;21(5):493–504.
- Harhay MO, Olliaro PL, Vaillant M, Chappuis F, Lima MA, Ritmeijer K, et al. Who Is a Typical Patient with Visceral Leishmaniasis? Characterizing the Demographic and Nutritional Profile of Patients in Brazil, East Africa, and South Asia. Am Soc Trop Med Hyg. 2011 Apr 5;84(4):543–50.
- Leite de Sousa-Gomes M, Romero GAS, Werneck GL. Visceral leishmaniasis and HIV/AIDS in Brazil: Are we aware enough? Gradoni L, editor. PLoS Negl Trop Dis. 2017 Sep 25;11(9):e0005772.
- Machado CAL, Sevá A da P, Silva AAFA e, Horta MC. Epidemiological profile and lethality of visceral leishmaniasis/human immunodeficiency virus co-infection in an endemic area in Northeast Brazil. Rev Soc Bras Med Trop . 2021;54. 86822021000100316&tlng=en.
- Hailu A, van der Poll T, Berhe N, Kager PA. Elevated plasma levels of interferon (IFN)-gamma, IFN-gamma inducing cytokines, and IFN-gamma inducible CXC chemokines in visceral leishmaniasis. Am J Trop Med Hyg. 2004 Nov;71(5):561–7.
- Peruhype-Magalhães V, Martins-Filho OA, Prata A, Silva LDA, Rabello A, Teixeira-Carvalho A, et al. Mixed inflammatory/regulatory cytokine profile marked by simultaneous raise of interferon-γ and interleukin-10 and low frequency of tumour necrosis factor-α+ monocytes are hallmarks of active human visceral Leishmaniasis due to Leishmania chagasi infectio. Clin Exp Immunol. 2006 Aug 25;146(1):124–32.
- Ansari NA, Saluja S, Salotra P. Elevated levels of interferon-γ, interleukin-10, and interleukin-6 during active disease in Indian kala azar. Clin Immunol. 2006 Jun;119(3):339–45.
- van Dijk NJ, Carter J, Kiptanui D, Mens PF, Schallig HDFH. A case–control study on risk factors for visceral leishmaniasis in West Pokot County, Kenya. Trop Med Int Heal. 2024 Oct 4;29(10):904–12.
- Decker ML, Grobusch MP, Ritz N. Influence of Age and Other Factors on Cytokine Expression Profiles in Healthy Children—A Systematic Review. Front Pediatr. 2017 Dec 14;5.
- Ringleb M, Javelle F, Haunhorst S, Bloch W, Fennen L, Baumgart S, et al. Beyond muscles: Investigating immunoregulatory myokines in acute resistance exercise – A systematic review and meta-analysis. FASEB J. 2024 Apr 15;38(7).
- Tasca KI, Correa CR, Caleffi JT, Mendes MB, Gatto M, Manfio VM, et al. Asymptomatic HIV People Present Different Profiles of sCD14, sRAGE, DNA Damage, and Vitamins, according to the Use of cART and CD4 + T Cell Restoration. J Immunol Res. 2018;2018:1–11.
- Guedes DL, Silva ED da, Castro MCAB, Júnior WLB, Ibarra-Meneses AV, Tsoumanis A, et al. Comparison of serum cytokine levels in symptomatic and asymptomatic HIV-Leishmania coinfected individuals from a Brazilian visceral leishmaniasis endemic area. Guizani I, editor. PLoS Negl Trop Dis. 2022 Jun 17;16(6):e0010542.
- Hunter CA, Jones SA. IL-6 as a keystone cytokine in health and disease. Nat Immunol. 2015 May;16(5):448–57.
- Mary C, Faraut F, Lascombe L, Dumon H. Quantification of Leishmania infantum DNA by a Real-Time PCR Assay with High Sensitivity. J Clin Microbiol. 2004 Nov;42(11):5249–55.
- de Araújo Albuquerque LP, da Silva AM, de Araújo Batista FM, de Souza Sene I, Costa DL, Costa CHN. Influence of sex hormones on the immune response to leishmaniasis. Parasite Immunol. 2021 Oct 2;43(10–11).
- Murray HW, Rubin BY, Rothermel CD. Killing of intracellular Leishmania donovani by lymphokine-stimulated human mononuclear phagocytes. Evidence that interferon-gamma is the activating lymphokine. J Clin Invest. 1983 Oct 1;72(4):1506–10.
- Reiner NE, Ng W, Wilson CB, McMaster WR, Burchett SK. Modulation of in vitro monocyte cytokine responses to Leishmania donovani. Interferon-gamma prevents parasite-induced inhibition of interleukin 1 production and primes monocytes to respond to Leishmania by producing both tumor necrosis factor-alpha and in. J Clin Invest. 1990 Jun 1;85(6):1914–24.
- Heinzel FP, Schoenhaut DS, Rerko RM, Rosser LE, Gately MK. Recombinant interleukin 12 cures mice infected with Leishmania major. J Exp Med . 1993 May 1;177(5):1505–9.
- Murray HW, Hariprashad J. Interleukin 12 is effective treatment for an established systemic intracellular infection: experimental visceral leishmaniasis. J Exp Med. 1995 Jan 1;181(1):387–91.
- Ghalib HW, Piuvezam MR, Skeiky YA, Siddig M, Hashim FA, El-Hassan AM, et al. Interleukin 10 production correlates with pathology in human Leishmania donovani infections. J Clin Invest. 1993 Jul 1;92(1):324–9.
- Bacellar O, Brodskyn C, Guerreiro J, Barral-Netto M, Costa CH, Coffman RL, et al. Interleukin-12 Restores Interferon- Production and Cytotoxic Responses in Visceral Leishmaniasis. J Infect Dis. 1996 Jun 1;173(6):1515–8.
- Verma S, Kumar R, Katara GK, Singh LC, Negi NS, Ramesh V, et al. Quantification of Parasite Load in Clinical Samples of Leishmaniasis Patients: IL-10 Level Correlates with Parasite Load in Visceral Leishmaniasis. Rodrigues MM, editor. PLoS One. 2010 Apr 9;5(4):e10107.
- Bhattacharya P, Ghosh S, Ejazi SA, Rahaman M, Pandey K, Ravi Das VN, et al. Induction of IL-10 and TGFβ from CD4+CD25+FoxP3+ T Cells Correlates with Parasite Load in Indian Kala-azar Patients Infected with Leishmania donovani. Bates PA, editor. PLoS Negl Trop Dis. 2016 Feb 1;10(2):e0004422.
- Teles L de F, Viana AG, Cardoso MS, Pinheiro GRG, Bento GA, Lula JF, et al. Evaluation of medullary cytokine expression and clinical and laboratory aspects in severe human visceral leishmaniasis. Parasite Immunol. 2021 Dec 11;43(12).
- Carvalho EM, Badaró R, Reed SG, Jones TC, Johnson WD. Absence of gamma interferon and interleukin 2 production during active visceral leishmaniasis. J Clin Invest. 1985 Dec 1;76(6):2066–9.
- Carvalho EM, Bacellar O, Brownell C, Regis T, Coffman RL, Reed SG. Restoration of IFN-gamma production and lymphocyte proliferation in visceral leishmaniasis. J Immunol. 1994 Jun 15;152(12):5949–56.
- Karp CL, El-Safi SH, Wynn TA, Satti MM, Kordofani AM, Hashim FA, et al. In vivo cytokine profiles in patients with kala-azar. Marked elevation of both interleukin-10 and interferon-gamma. J Clin Invest. 1993 Apr 1;91(4):1644–8.
- Nylén S, Sacks D. Interleukin-10 and the pathogenesis of human visceral leishmaniasis. Trends Immunol. 2007 Sep;28(9):378–84.
- Kany S, Vollrath JT, Relja B. Cytokines in Inflammatory Disease. Int J Mol Sci. 2019 Nov 28;20(23):6008.
- Fabri A, Kandara K, Coudereau R, Gossez M, Abraham P, Monard C, et al. Characterization of Circulating IL-10-Producing Cells in Septic Shock Patients: A Proof of Concept Study. Front Immunol. 2021 Feb 4;11.
- D’Oliveira Júnior A, Costa SRM, Bispo Barbosa A, Orge Orge M de LG, Carvalho EM. Asymptomatic Leishmania chagasi Infection in Relatives and Neighbors of Patients with Visceral Leishmaniasis. Mem Inst Oswaldo Cruz. 1997 Jan;92(1):15–20.
- Lima S, Braz D, Silva V, Farias T, Zacarias DADAADA, Silva JC, et al. Biomarkers of the early response to treatment of visceral leishmaniasis: A prospective cohort study. Parasite Immunol. 2021 Jan 20;43(1).
- Saraiva M, Vieira P, O’Garra A. Biology and therapeutic potential of interleukin-10. J Exp Med. 2020 Jan 6;217(1).
- Munk RB, Sugiyama K, Ghosh P, Sasaki CY, Rezanka L, Banerjee K, et al. Antigen-Independent IFN-γ Production by Human Naïve CD4+ T Cells Activated by IL-12 Plus IL-18. Klinman D, editor. PLoS One . 2011 May 10;6(5):e18553.
- Lee HG, Cho MJ, Choi JM. Author Correction: Bystander CD4+ T cells: crossroads between innate and adaptive immunity. Exp Mol Med. 2023 Jun 8;55(6):1275–1275.
- Reis-Cunha JL, Grace CA, Ahmed S, Harnqvist SE, Lynch CM, Boité MC, et al. The global dispersal of visceral leishmaniasis occurred within human history . 2024.
- Zijlstra, EE. The immunology of post-kala-azar dermal leishmaniasis (PKDL). Parasit Vectors. 2016 Dec 23;9(1):464.
- Jarczak D, Nierhaus A. Cytokine Storm—Definition, Causes, and Implications. Int J Mol Sci. 2022 Oct 3;23(19):11740.
- Miller CHT, Maher SG, Young HA. Clinical Use of Interferon-γ. Ann N Y Acad Sci. 2009 Dec 14;1182(1):69–79.
- Baessler A, Vignali DAA. T Cell Exhaustion. Annu Rev Immunol. 2024 Jun 28;42(1):179–206.
- Sonar SA, Watanabe M, Nikolich JŽ. Disorganization of secondary lymphoid organs and dyscoordination of chemokine secretion as key contributors to immune aging. Semin Immunol. 2023 Nov;70:101835.
- Gautam S, Kumar R, Singh N, Singh AK, Rai M, Sacks D, et al. CD8 T Cell Exhaustion in Human Visceral Leishmaniasis. J Infect Dis. 2014 Jan 15;209(2):290–9.
- Silva-O’Hare J, de Oliveira IS, Klevorn T, Almeida VA, Oliveira GGS, Atta AM, et al. Disruption of Splenic Lymphoid Tissue and Plasmacytosis in Canine Visceral Leishmaniasis: Changes in Homing and Survival of Plasma Cells. Khan WN, editor. PLoS One . 2016 May 31;11(5):e0156733.
- Hermida M d’El R, de Melo CVB, Lima I dos S, Oliveira GG de S, Dos-Santos WLC. Histological Disorganization of Spleen Compartments and Severe Visceral Leishmaniasis. Front Cell Infect Microbiol. 2018 Nov 13;8.
- de Souza TL, da Silva AVA, Pereira L de OR, Figueiredo FB, Mendes Junior AAV, Menezes RC, et al. Pro-Cellular Exhaustion Markers are Associated with Splenic Microarchitecture Disorganization and Parasite Load in Dogs with Visceral Leishmaniasis. Sci Rep. 2019 Sep 10;9(1):12962.
Figure 1.
Correlation between plasma and bone marrow parasite load (r = 0.48, p-value < 0.001). Legend: *Natural logarithm of kDNA amastigote-equivalents/mL of plasma (AEq/mL). ** *Natural logarithm of AEq/106 DNA equivalents of human cells.
Figure 1.
Correlation between plasma and bone marrow parasite load (r = 0.48, p-value < 0.001). Legend: *Natural logarithm of kDNA amastigote-equivalents/mL of plasma (AEq/mL). ** *Natural logarithm of AEq/106 DNA equivalents of human cells.
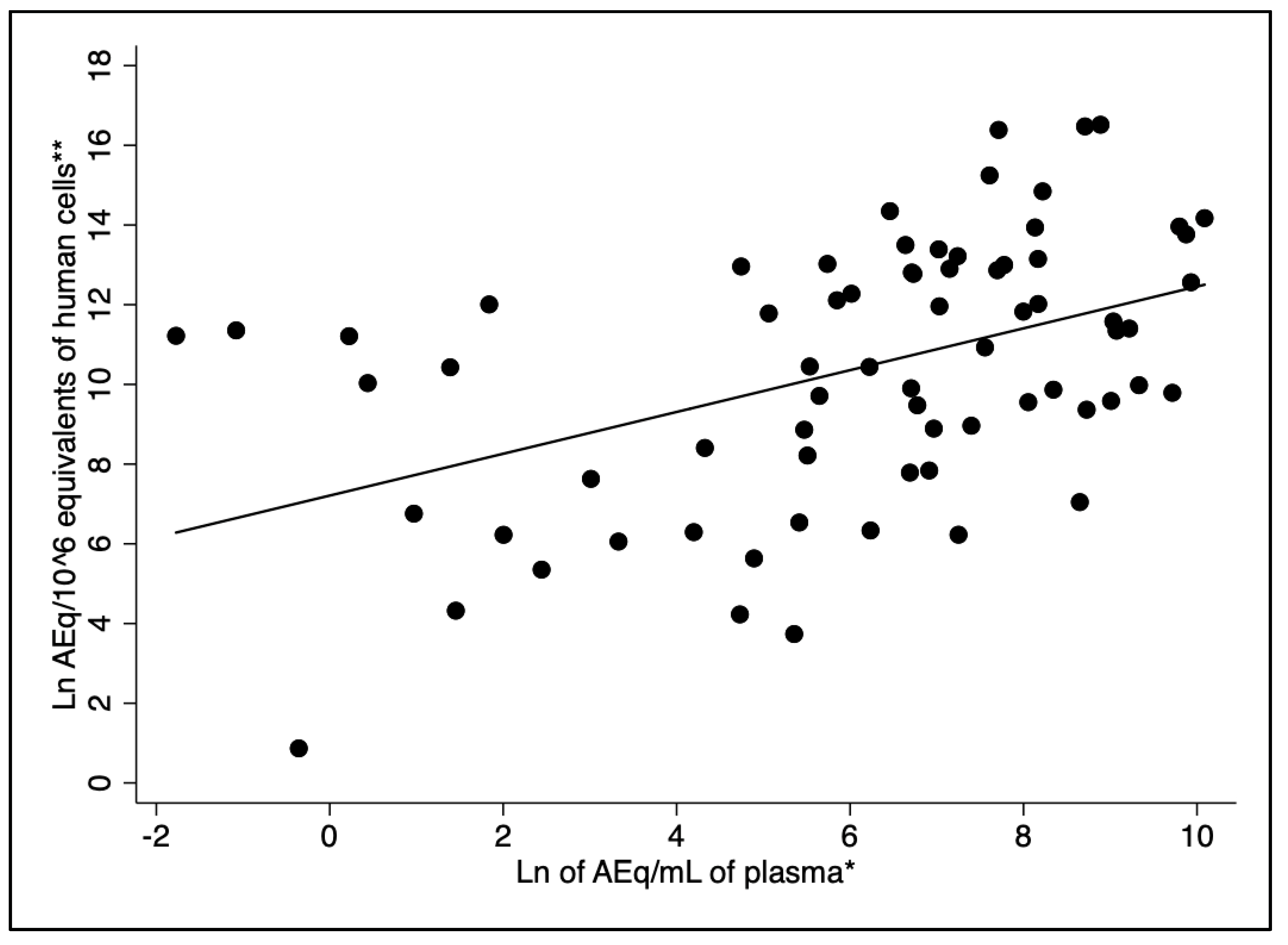
Figure 2.
Increasing parasite load with time of fever. A: plasma load (r = 0.33, p-value < 0.005); B: bone marrow load (r = 0.38, p-value < 0.001).
Figure 2.
Increasing parasite load with time of fever. A: plasma load (r = 0.33, p-value < 0.005); B: bone marrow load (r = 0.38, p-value < 0.001).
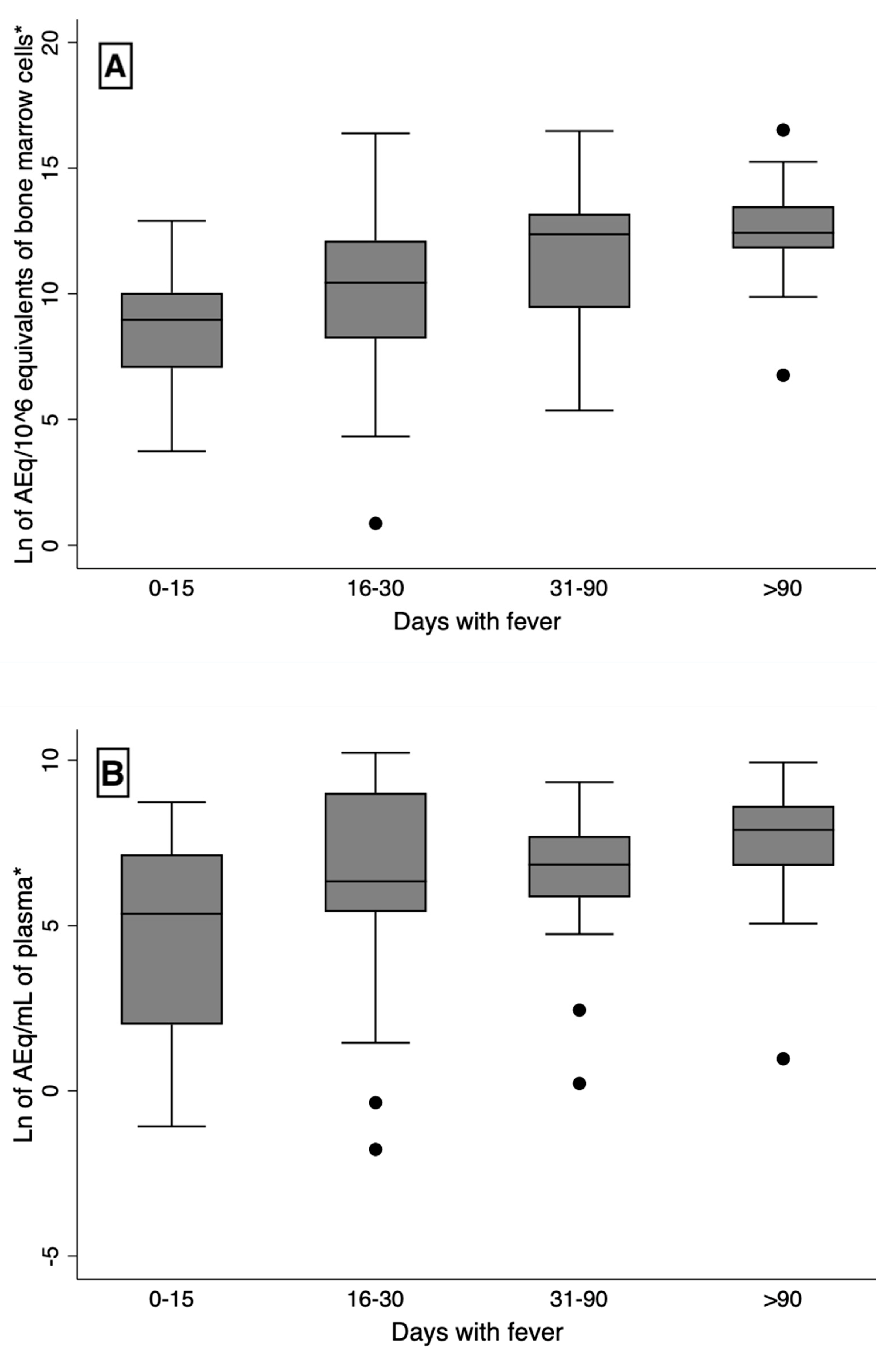
Figure 3.
Above: output of the multivariate linear regression analysis for prediction of of plasma parasite kDNA load by plasma cytokines. Bellow: output of the multivariate quantile regression analysis for prediction of plasma parasite kDNA load by plasma cytokines.
Figure 3.
Above: output of the multivariate linear regression analysis for prediction of of plasma parasite kDNA load by plasma cytokines. Bellow: output of the multivariate quantile regression analysis for prediction of plasma parasite kDNA load by plasma cytokines.
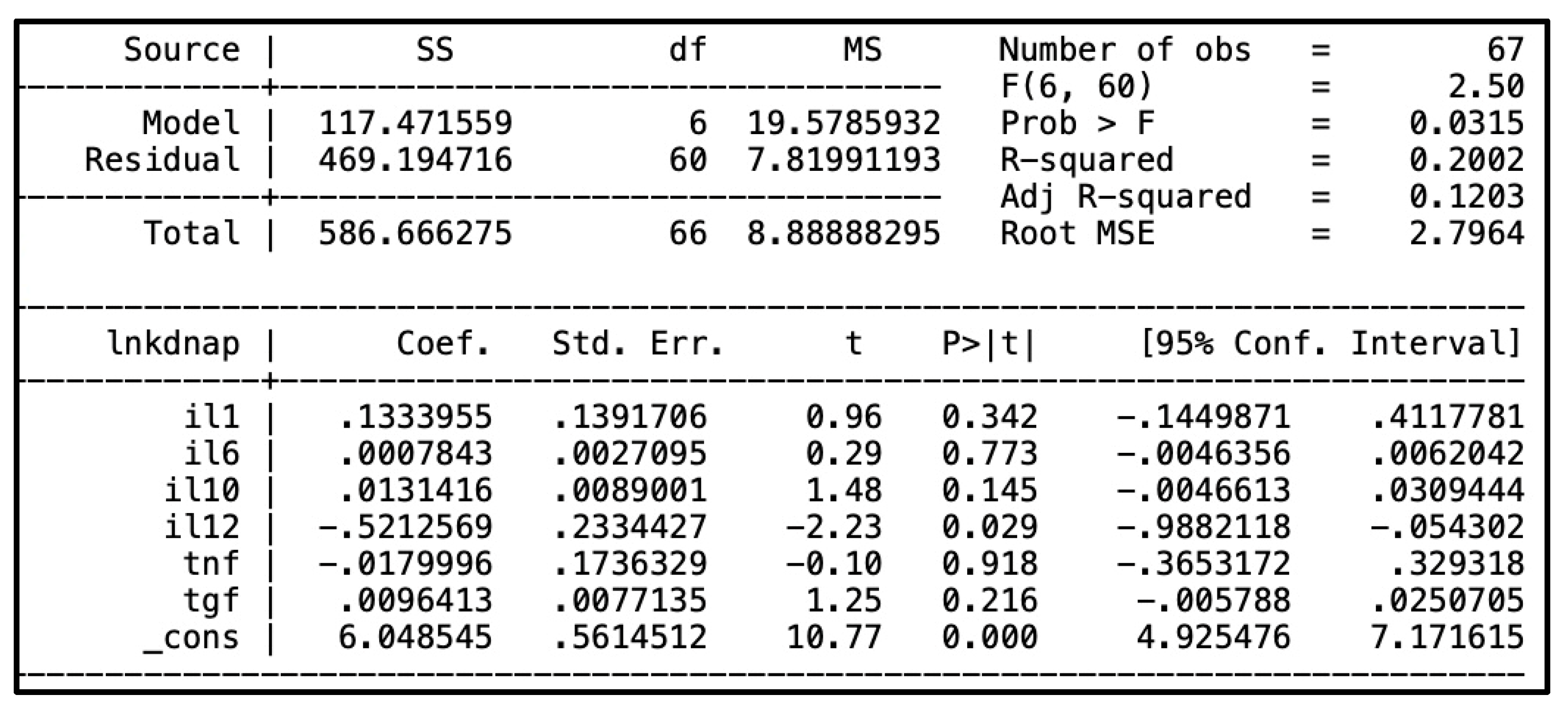
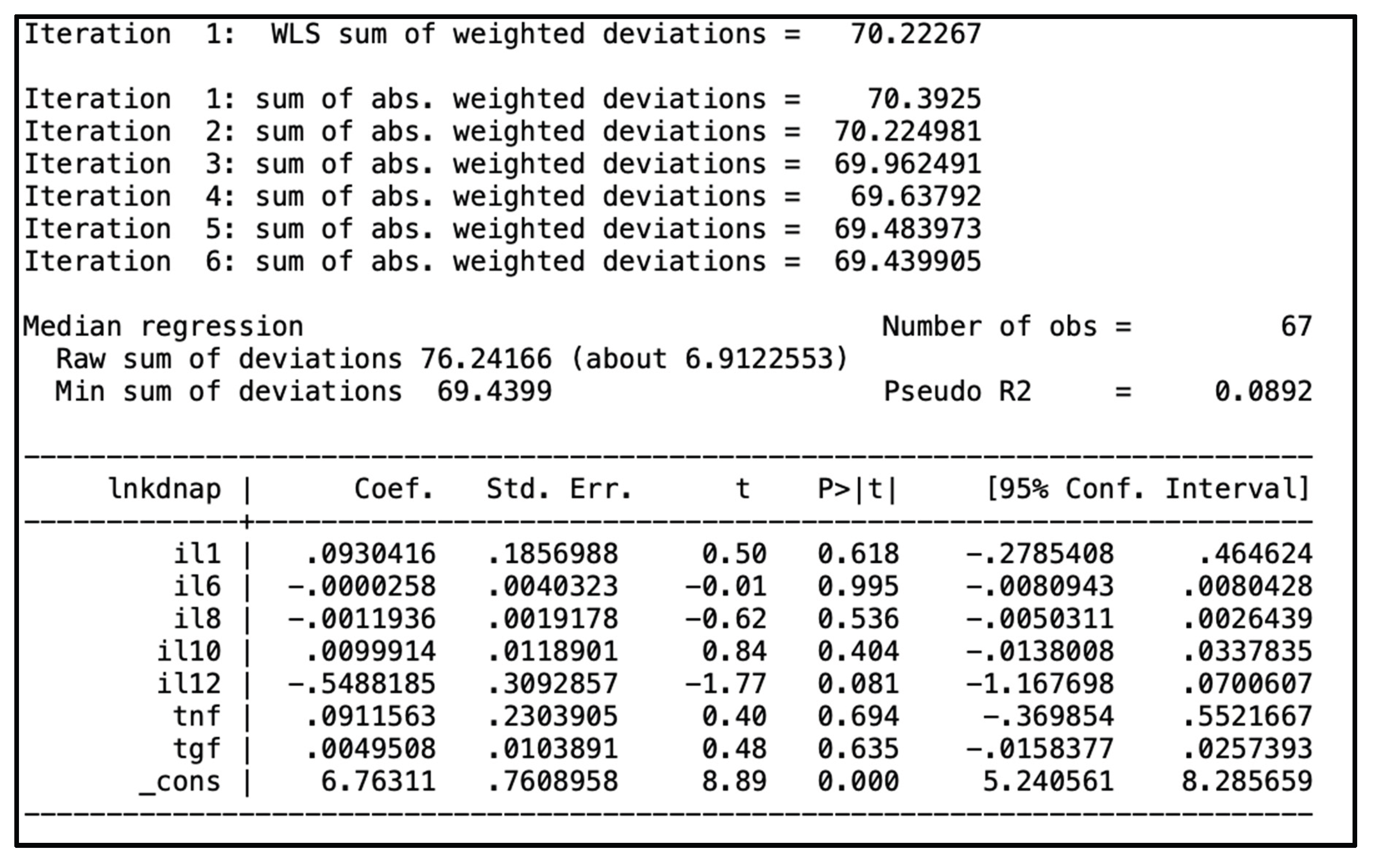

Table 1.
Characteristics of the study population.
| Characteristic | Number (%) | 95% CI1 |
| Sex | ||
| Male | 42 (58.3) | 46.1; 69.85 |
| Female | 30 (41.7) | 30.2; 53.89 |
| Age groups (Years) | ||
| <2 | 17 (23.6) | 14.0; 35.0 |
| 2<4 | 5 (6.9) | 2.2; 15.4 |
| 4<15 | 22 (30.6) | 20.2; 42.5 |
| 15<40 | 22 (30.6) | 20.2; 42.5 |
| 40+ | 6 (8.3) | 3.1; 17.3 |
| HIV2 (number, %) | 13 (18.6) | 10.3; 30.0 |
| Deaths (number, %) | 4 (5.6) | 1.5; 13.61 |
| Chance of death > 10% by Kala-Cal® | 25 (34.7) | 23.9; 46.9 |
| Hemorrhages or infections | 31 (43.7) | 31.9; 56.0 |
| Reported bleeding | 4 (5.6%) | 1.5; 13.6 |
| Detected bleeding | 15 (20.8) | 12.2; 32.0 |
| Sepsis | 10 (14.1) | 7.0; 24.4 |
| Any bacterial infection | 23 (31.9) | 21.4; 44.0 |
195% confidence interval. 2Only 70 patients tested for HIV infection.
Table 2.
Concentration of kDNA and cytokines in the plasma of patients with kala-azar.
| Variables | Median | Interquartile intervals | Mean | Reference values (median) | Kolmogorov–Smirnov test (p-value) |
| Plasma kDNA (AEq1 /mL) | 856.7 | 145.5-3,527.9 | 3,515.4 | 06 | 0.000 |
| Bone marrow kDNA (AEq/106HCEq2) | 55.7 | 3.6 – 4,008 | 889.9 | 06 | 0.000 |
| IL-1b pg/mL | 0.9 | 0.2 – 2.1 | 2.0 | 0.18 (0–3.66)3 | 0.000 |
| IL-6 pg/mL | 9.5 | 2.4 – 28.0 | 41.7 | 0 (0–0)3 | 0.000 |
| IL-8 pg/mL | 26.2 | 9.8 – 145.5 | 146.9 | 0 (0–0)3 | 0.000 |
| IL-10 pg/mL | 18.4 | 8.7 – 35 | 30.2 | 0 (0–0)3 | 0.000 |
| IL-12 pg/mL | 1.2 | 0.0 – 2.5 | 1.8 | 0 (0–0)3 | 0.000 |
| TNF-a pg/mL | 1.0 | 0.3 – 3.0 | 2.2 | 0 (0–0)3 | 0.000 |
| TGF-b ng/mL | 23.6 | 11.2 – 42.4 | 39.6 | NA4,5 | - |
Table 3.
Table 2. Concentration of Leishmania infantum amastigotes estimated by kDNA in the blood of patients with kala-azar.
Table 3.
Table 2. Concentration of Leishmania infantum amastigotes estimated by kDNA in the blood of patients with kala-azar.
| Markers of severe disease (number of patients) |
Blood AEq1 median, (mean) |
p-value2 |
Bone marrow AEq/109HCEq median, (mean) |
p-value3 |
| Age (years) | ||||
| <15 (27) | 508 (2,513) | 27 (519) | ||
| 15+ (44) | 2,679 (5,091) | 0.022 | 137 (1,494) | 0.043 |
| Sex | ||||
| Female (30) | 329 (1,580) | 28 (631) | ||
| Male (41) | 1,452 (4,898) | 0.012 | 90 (1,079) | 0.073 |
| HIV | ||||
| Yes (13) | 4,207 (7,016) | 56 (2,867) | ||
| No (57) | 511 (2,768) | 0.006 | 35 (500) | 0.168 |
| Hospital outcome | ||||
| Death (4) | 11,826 (11,925) | 487 (605) | ||
| Survival (67) | 830 (3,020) | 0.169 | 56 (907) | 0.273 |
| Chance of death > 10% by Kala-Cal® | ||||
| > 10% (25) | 3,532 (6,617) | 290 (2,280) | ||
| <10% (46) | 311 (1,866) | <0.001 | 20 (350) | 0.001 |
| Reported bleeding | ||||
| Yes (4) | 8,855 (10,440) | 89 (406) | ||
| No (67) | 830 (3,108) | 0.350 | 56 (919) | 0.517 |
| Detected bleeding | ||||
| Yes (15) | 823 (4,427) | 214 (1,406) | ||
| No (56) | 876 (3,276) | 0.873 | 21 (752) | 0.012 |
| Sepsis | ||||
| Yes (10) | 888 (5,619) | 247 (375) | ||
| No (60) | 1,130 (3,210) | 0.856 | 34 (988) | 0.374 |
| Any bacterial infection | ||||
| Yes (23) | 837 (3,679) | 425 (466) | ||
| No (48) | 1,059 (3,439) | 0.758 | 65 (1,093) | 0.722 |
1Amastigote-equivalents/mL 2Wilcoxon ranksum test. 3AEq/106 equivalents of human cells.
Table 4.
Plasma concentration of cytokines according to demographic data, and HIV-infection, and markers of kala-azar severity.
Table 4.
Plasma concentration of cytokines according to demographic data, and HIV-infection, and markers of kala-azar severity.
| Variables (number of patients) |
IL-1b median, (mean) |
p-value1 |
IL-6 median, (mean) |
p-value |
IL-8 median, (mean) |
p-value |
IL-10 median, (mean) |
p-value |
IL-12 median, (mean |
p-value |
TNF-a median, (mean) |
p-value |
TGF-b median, (mean) |
p-value |
| Age (years) | ||||||||||||||
| <15 (27) | 1.1 (2.1) | 14.0 (49.3) | 28.8 (130.7) | 23.4 (36.4) | 1.4 (2.1) | 1.2 (2.5) | 23.1 (40.5) | |||||||
| 15+ (44) | 0.4 (1.9) | 0.18 | 7.8 (28.9) | 0.16 | 23.3 (174.2) | 0.95 | 12.8 (19.8) | 0.02 | 0.6 (1.3) | 0.18 | 0.9 (1.6) | 0.52 | 24.5 (38.0) | 0.95 |
| Sex | ||||||||||||||
| Female (30) | 0.9 (1.9) | 8.4 (55.9) | 25.6 (118.7) | 23.1 (32.6) | 1.0 (2.2) | 2.1 (3.1) | 13.9 (40.6) | |||||||
| Male (41) | 1.0 (2.0) | 0.95 | 13.6 (32.6) | 0.79 | 26.2 (164.8) | 0.99 | 13.6 (28.7) | 0.06 | 1.3 (1.6) | 0.92 | 0.9 (1.7) | 0.54 | 28.5 (38.9) | 0.25 |
| HIV-infection | ||||||||||||||
| Yes (12) | 1.3 (2.7) | 8.1 (13.0) | 46.7 (188.1) | 12.4 (14.3) | 1.2 (1.7) | 1.6 (1.9) | 16.5 (35.9) | |||||||
| No (57) | 0.9 (1.9) | 0.46 | 13.0 (48.8) | 0.50 | 25.4 (140.5) | 0.50 | 21.8 (34.2) | 0.04 | 1.3 (1.9) | 0.87 | 1.0 (2.3) | 0.70 | 24.05 (40.1) | 0.65 |
| Hospital outcome | ||||||||||||||
| Death (4) | 1.3 (1.3) | 24.7 (44.4) | 48.1 (173.9) | 12.0 (31.5) | 1.4 (1.8) | 1.2 (1.8) | 18.6 (20.4) | |||||||
| Survival (62) | 0.9 (2.1) | 0.45 | 9.1 (42.2) | 0.44 | 26.4 (147.5) | 0.96 | 18.7 (30.5) | 0.44 | 1.2 (1.8) | 0.61 | 1.0 (2.3) | 0.84 | 24.1 (20.4) | 0.58 |
| Chance of death2 | ||||||||||||||
| > 10% (23) | 1.0 (2.5) | 15.4 (44.3) | 65.2 (206.5) | 16.2 (27.8) | 1.1 (1.6) | 1.0 (2.3) | 24.5 (45.4) | |||||||
| <10% (43) | 0.9 (1.8) | 0.33 | 9.5 (44.3) | 0.45 | 24.6 (118.5) | 0.10 | 19.1 (32.0) | 0.59 | 1.3 (1.9) | 0.68 | 0.9 (2.2) | 0.36 | 22.3 (36.1) | 0.53 |
| Reported bleeding | ||||||||||||||
| Yes (3) | 1.6 (1.8) | 34.1 (63.7) | 78.6 (246.1) | 34.7 (51.3) | 2.0 (2.1) | 0.4 (1.8) | 15.7 (19.8) | |||||||
| No (63) | 0.9 (2.0) | 0.14 | 8.6 (41.2) | 0.03 | 24.9 (144.5) | 0.16 | 16.6 (29.5) | 0.15 | 1.2 (1.8) | 0.44 | 1.0 (2.3) | 0.83 | 23.6 (40.3) | 0.51 |
| Detected bleeding | ||||||||||||||
| Yes (13) | 0.9 (2.8) | 17.2 (24.9) | 88.5 (242.7) | 16.1 (24.1) | 2.0 (2.5) | 2.1 (2.2) | 42.1 (59.8) | |||||||
| No (53) | 1.0 (1.9) | 0.50 | 8.6 (46.6) | 0.42 | 24.6 (126.2) | 0.04 | 21.8 (32.1) | 0.57 | 1.2 (1.7) | 0.13 | 1.0 (2.3) | 0.66 | 20.9 (34.3) | 0.09 |
| Sepsis | ||||||||||||||
| Yes (10) | 1.6 (1.9) | 23.0 (137.3) | 110.2 (229.9) | 36.9 (37.3) | 1.9 (2.5) | 2.4 (2.5) | 18.3 (31.9) | |||||||
| No (56) | 0.8 (2.0) | 0.06 | 8.1 (25.3) | 0.06 | 24.7 (134.7) | 0.21 | 16.1 (29.3) | 0.07 | 0.9 (1.7) | 0.04 | 0.9 (2.2) | 0.17 | 25.7 (40.7) | 0.41 |
| Any bacterial infection | ||||||||||||||
| Yes (22) | 1.6 (2.4) | 25.2 (83.1) | 67.0 (203.9) | 30.5 (40.6) | 1.9 (2.1) | 2.4 (2.8) | 18.9 (33.0) | |||||||
| No (44) | 0.7 (1.8) | 0.01 | 7.2 (21.9) | 0.01 | 22.6 (121.8) | 0.03 | 16.0 (25.5) | 0.04 | 0.9 (1.7) | 0.11 | 0.8 (2.0) | 0.01 | 29.5 (42.5) | 0.24 |
1Wilcoxon ranksum test. 2Kala-Cal®.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



