Submitted:
26 May 2025
Posted:
26 May 2025
You are already at the latest version
Abstract
Farmed fish encounter persistent challenges stemming from a variety of environmental challenges and human activities that induce both acute and chronic stress, resulting in increased vulnerability to diseases and mortality. Therefore, it is essential to comprehend the stressors and corresponding stress responses in fish to recognize and mitigate harmful stress during aquaculture practices. In this review, we provide an overview of the various stressors present in aquaculture, along with the resultant alterations in neuroendocrine responses, including the BSC axis, HPI axis, and CNSS, as well as neurotransmitter levels within the nervous system, experienced by fish under different types of stress. Fish possess well-developed endogenous nervous and endocrine systems that respond to stress, with complex communication networks among these systems demonstrating distributed collaboration. An understanding of the neuroendocrine and neurotransmitter responses to stress may enhance our comprehension of fish stress mechanisms and facilitate the development of an integrated dietary supplementation strategy and improve their resilience against the diverse stresses encountered in aquaculture.
Keywords:
Stress
; Aquaculture
; Neural responses
; Neuroendocrine responses
; Fish
1. Introduction
Over the past few decades, the growing requirement for fish protein and the change in dietary habits have caused the explosive development of aquaculture [1,2]. However, the growth of aquaculture is limited by the various environmental and husbandry-related stress which results in the decline of disease resistance and subsequent death in fish. While the definition of stress remains a topic of debate, the most recent literature, acknowledged by the majority of researchers, suggests that the colloquial use of the term "stress" should be restricted to situations in which an environmental demand exceeds an organism’s natural regulatory capacity and adaptive resources in a maladaptive manner [3,4]. It has been demonstrated that, in response to stress, fish exhibit a series of neural and neuroendocrine reactions aimed at participating in stressor recognition or alleviating the adverse effects of stressors and maintaining homeostasis [5,6]. Depending on the intensity and duration of the stressor, acute stress often shows beneficial effects, while chronic stress normally stands on the opposite side, that increases the fish vulnerability to fatal diseases [7]. The terms “acute” and “chronic” are context-dependent, which should be based on the duration of its consequences on the physiology of the animal [8]. For instance, being caught in a net or escaping from a predator would typically be classified as “acute stress,” whereas being overcrowded in a tank or occupying a low position in the social hierarchy would generally be categorized as “chronic stress”. Therefore, it is crucial to address the understanding of the mechanisms of the neural and neuroendocrine responses when fish cope with stress. This article presents a comprehensive overview of the prevalent stressors in aquaculture, as well as the intricate interplay between neural and neuroendocrine responses in fish during periods of stress. It aims to facilitate a deeper understanding of fish stress and to illustrate an integrated adaptation strategy in fish that enhances their resilience against the multifaceted challenges they encounter.
2. Stressors in Aquaculture
Common stressors in aquafarming activities, such as unsuitable temperature (high or low temperature), pH (overacid or overalkali), dissolved oxygen (hypoxia or hyperxia), microorganisms (pathogenic parasite, bacteria or viruses), and overcrowding, etc., can be classified as environmental and anthropogenic stressors according to the stress sources, which can be further divided into physical, chemical, biological, and procedural stressors (Table 1). Changes in these stressors during the culture period can adversely affect the behavior, growth, immunity, and reproduction of fish, leading to economic loss to aquafarmers. Oxygen delivery limitations and cardiac failure have been known as the critical element in sublethal thermal stress [9]; however, stressor usually does not appear alone. For instance, increases in water temperature will not only elevate the fish oxygen demand and reduce the dissolved oxygen in the water, but also increase the toxicity of harmful substances (such as unionized ammonia, toxic microalgae, etc. [10,11]), and these will accelerate the homeostasis imbalance in fish. Meanwhile, unlike livestock, fish live in water, which make it hard for aquafarmers to detect anomalies in time. Therefore, the stress in aquaculture should not be overlooked, environmental monitoring and control are required the intensive and sustainable aquaculture. However, it is impossible to maintain an absolute stable condition without any stressors during farming processes. In addition, fish have a well-developed and complex endogenic neural and neuroendocrine regulation, which can induce a series of signaling cascades to deal with threatened homeostasis caused by stress. Understanding the cellular and molecular mechanisms of the neural and neuroendocrine responses and their network during stress, may help to reduce the negative effects of stress, improve disease resistance and develop strategies to mitigate the harmful effects of stress by regulating the neural and neuroendocrine system in fish.
3. Neurotransmitters and Neuroendocrine Systems Respond to Stress in Fish
When fish are stressed, the primary stress responses are the neural and neuroendocrine responses. A perceived threat triggers the activation of the neural circuit in the brain, leading to the activation of downstream endocrine systems and other responsive organs [3,12]. The nervous system utilizes a variety of neurotransmitters in response to stress (Figure 1). In fish, three major neuroendocrine systems respond to stress: the brain-sympathetic-chromaffin cells (BSC) axis, the hypothalamus-pituitary-interrenal (HPI) axis, and the caudal neurosecretory system (CNSS) (Figure 2, Table 2).
3.1. Neurotransmitters During Stress
3.1.1. Acetylcholine (ACh)
ACh is a neurotransmitter released by cholinergic neurons and is found in both the peripheral nervous system (PNS) and central nervous system (CNS) of fish [13]. Cholinergic systems in the brain play a regulatory role in numerous physiological processes, including muscle contraction, autonomic regulation, cognitive function, and stress responses. Cholinergic neurons are distinguished by the presence of choline acetyltransferase (ChAT), which facilitates ACh synthesis, and the vesicular acetylcholine transporter (VAChT), which transports ACh into vesicles. The presynaptic release of ACh from vesicles into the synaptic cleft triggers the activation of nicotinic and muscarinic ACh receptors (nAChR and mAChR, respectively) before undergoing degradation by acetylcholinesterase (AChE) into acetate and choline. The latter is then transported back into the presynaptic neurons via the high-affinity choline transporter (HACT) [14].
It is widely acknowledged that the majority of stress leads to a rapid, transiently enhanced release of ACh in the cholinergic system [15,16]. The release of ACh may modulate the activation of stress-related signaling pathways. However, depending on the source and intensity of the stressors, the level of ACh may decrease following stress under certain conditions [17]. Due to the low concentrations and rapid turnover of ACh, measuring its levels in the fish brain has been challenging. Therefore, it is common practice to assess the activity of ACh-related enzymes and the expression of ACh receptors in order to evaluate the responses of the cholinergic system during stress. Positive correlations have been observed between the ACh content and the ChAT activity in various fish, including carp (Cyprinus carpio), rainbow trout (Oncorhynchus mykiss) and eel (Anguilla anguilla) [18], while inhibition in the expression and activity of AChE has been noted during different stress conditions, such as heavy mental load [19], temperature changes in carp (C. carpio)[20], pesticide exposure in tilapia (Oreochromis mossambicus) [21,22] , parasite infestation in killfish (Fundulus parvipinnis) [23], and exposure to fired underwater currents in rainbow trout (O. mykiss) [24]. In addition, AChE is widely recognized as the primary neurotoxic biomarker that reflects cholinergic responses during various stressors in fish [25]. Reduced AChE activity implies a decline in cholinergic transmission and subsequent accumulation of ACh. A study by Jifa et al. (2006) reported no significant inhibition of AChE in the brain of Japanese seabass (Lateolabrax japonicus) exposed to benzo[a]pyrene (BaP) at concentrations of 2 and 20 µg L⁻¹ for 6, 12, and 18 days. This suggests that these concentrations had no appreciable effect on AChE activity [26]. Additionally, studies had shown that fish exposed to polycyclic aromatic hydrocarbons (PAHs) with a low molecular weight did not induce changes or cause stimulation of AChE activity [27]. It has also been reported that stress can induce the up-regulation of AChR in fish [28]. The activation of nAChR can induce anxiolytic like effects in zebrafish [29], while the antagonist of mAChR can disrupt the stress responses caused by organophosphorus exposure [30].
3.1.2. Glutamate (Glu) and Gamma-Aminobutyric Acid (GABA)
Glu and GABA are the principal neurotransmitters in the central nervous system. Glu and GABA are synthesized not only in the brain, but also in peripheral tissues and organs [31,32]. Glu primarily functions as an excitatory neurotransmitter, whereas GABA serves as an inhibitory neurotransmitter. These two substances exhibit opposing yet complementary physiological roles, collectively regulating the normal functioning of the nervous system [33]. Glutamate is synthesized in neurons from intermediates of the glucose-derived tricarboxylic acid cycle and branched-chain amino acids, a process catalyzed by glutaminase utilizing glutamine (Gln) as a precursor. Within neurons, cytoplasmic glutamate is encapsulated into vesicles by vesicular glutamate transporters for subsequent extracellular release. Upon neuronal depolarization and the initiation of signal transduction, these glutamate-containing vesicles are released into the synaptic cleft, where they bind to three types of ionotropic glutamate receptors (iGluRs), N-methyl-D-aspartic acid receptor (NMDA), A-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid receptor (AMPA), and Kainate receptors [34] — as well as metabotropic glutamate receptors (mGluRs), before being removed and converted into Gln through the action of glutamine synthetase [35]. The synthesis of GABA is facilitated by the enzymatic action of glutamic acid decarboxylase (GAD), which catalyzes the α-decarboxylation reaction using Glu as a precursor. GABA exerts its effects through GABAA and GABAB receptors, which are classified into two primary categories: ionotropic receptors and metabotropic receptors, respectively [36].
When subjected to stress, maintaining the balance of Glu and GABA is crucial for the physiological adaptation of fish. An imbalance between Glu and GABA can disrupt their homeostasis, leading to increased excitatory toxicity, which ultimately results in cellular swelling, irreversible neuronal damage, and even cell death [37,38]. It has been reported that glutamic acid effectively alleviates the adverse effects of saline-alkali environments on aquatic organisms such as grass carp and tilapia [39,40]. In response to heat stress challenges, rainbow trout activate the glutamate-glutamine metabolic pathway to mitigate oxidative damage caused by elevated temperatures [41]. Notably, under saline-alkali environmental stress conditions, disruption or blockage of the glutamate metabolic pathway directly affects ammonia excretion processes in crucian carp, resulting in nitrogenous waste accumulation within the body that may ultimately trigger an inflammatory response [42]. In aquatic animals, GABA plays a crucial role in regulating glucose homeostasis during fasting stress [43] , alleviating symptoms of hyperglycemia, and enhancing the resilience of animals subjected to heat stress [44]. Further research has demonstrated that incorporating GABA into animal feed can significantly increase serum GABA levels while concurrently reducing serum concentrations of glucose and corticosterone hormones in stressful environments, thereby effectively mitigating the stress response [45,46]. In summary, Glu and GABA system plays an indispensable regulatory role under various environmental stresses.
3.1.3. Catecholamine (CA)
CAs are synthesized in the brain, in the adrenal medulla (head kidney in fish), and by certain sympathetic nerve fibers. These CAs, which include dopamine (DA), norepinephrine (NE), and epinephrine (E), are biosynthesized from tyrosine (Tyr). The process begins with hydroxylation catalyzed by tyrosine hydroxylase (TH), converting Tyr into dihydroxyphenylalanine (DOPA). Following this, DOPA undergoes decarboxylation mediated by aromatic amino acid decarboxylase (AADC) to yield DA. In certain neurons, DA functions directly as a neurotransmitter, whereas in others, DA may be further transported into chromaffin granules for additional processing. Within these granules, DA is further hydroxylated by dopamine-β-hydroxylase (DbH) to produce NE, which is then methylated through a reaction catalyzed by phenylethanol-N-methyl transferase (PNMT) to form E [47]. The activities of CAs are mediated by their binding to various receptors. In detail, the dopamine receptors are classified in two different families, D1-like (encompassing D1 and D5 subtypes) and D2-like (including D2, D3, and D4 subtypes), while adrenergic receptors are categorized into three different families, ADRα1 (α1A, α1B, α1D), ADRα2 (α2A, α2B, α2C) and ADRβ (β1, β2, and β3), based on their molecular structure and receptor pharmacological properties [48,49]. CAs play a pivotal role intricate processes like behavior, movement, emotion, and memory [50,51,52].
When fish and other vertebrates experience stress, they rapidly release CAs to elevate plasma glucose levels, thereby providing essential energy support for the instinctive ’fight-or-flight’ response [53]. In marine organisms, environmental factors significantly influence their stress responses. For example, in response to fluctuations in seawater temperature and salinity, Harpagifer antarcticus exhibits significant alterations in various neurotransmitters and DA and NE levels within its system, further emphasizing the critical role of stress responses in the adaptation of marine organisms to environmental changes [54]. Zebrafish subjected to acute osmotic pressure changes and hypoxia exhibit significant reduction in NE and E levels within their brains, which subsequently affect their reproductive signaling processes, thereby revealing the potential impact of stress responses on fish reproductive health [55]. The hypoxia-tolerant tropical fish Pygocentrus nattereri experiences notable effects of endogenous myocardial NE and E on cardiac function when exposed to hypoxic conditions, facilitating its ability to sustain vital activities in low-oxygen environments [56]. Furthermore, E play a crucial role in regulating energy balance in the livers of chondrichthyan fish, particularly under stressful conditions [57]. In aquaculture practices, the farming environment exerts a profound influence on fish stress responses. Gilthead seabream larvae, when subjected to adverse conditions such as high-density farming and fasting, exhibit significant alterations in their DA and NE levels and brain monoamines to adapt to these challenges [58]. Conversely, following exposure to low farming densities, O. mykiss experiences disruptions in the levels of brain CAs, leading to a series of physiological abnormalities such as reduced survival rates, impaired growth and feed efficiency, and dysregulation of immune and inflammatory systems [59]. Additionally, the CA levels in rainbow trout are also influenced by farming density, further affirming the significance of the farming environment on fish stress responses [59]. In addition to farming density, various environmental factors can also influence fish stress responses. For instance, exposure to AlCl3 results in decreased levels of GABA, DA, and NE in zebrafish, while glutamate levels are elevated [60]. Similarly, in adult zebrafish exposed to methamphetamine (METH), their brain levels of DA and NE decrease in a time- and concentration-dependent manner, accompanied by an increase in anxiety-like behavior [61].
3.1.4. Serotonin
The monoamine serotonin, also known as 5-hydroxytryptamine (5-HT), is a major neurotransmitter in the CNS and PNS of fish. It plays a crucial role in vasoconstriction [62], hemostasis, intestinal motility [63], and stress responses [64]. 5-HT is synthesized from dietary L-tryptophan in a two-step process: first, it undergoes hydroxylation by tryptophan hydroxylase (Tph) to form 5-hydroxytryptophan (5-HTP), and then it is decarboxylated by aromatic amino acid decarboxylase (Aadc) to form 5-HT. 5-HT is further metabolized to 5-hydroxyindole acetic acid (5-HIAA) through a combination of monoamine oxidase (Mao) and aldehyde dehydrogenase (Ad) [65]. In addition, serotonergic neurons can also be identified based on 5-HT transporter (Sert) and 5-HT receptors.
Many studies have demonstrated that stress can induce hyperactivity of serotonin metabolism in fish [66,67]. In tilapia and zebrafish, social stress and acute isolation stress could lead to a reduction in the level of 5-HT and an increase in its metabolites 5-HIAA [68,69,70]. The tryptophan hydroxylase 1a (tph1a) gene was significantly upregulated in the gill of zebrafish after long-term exposure of microplastics and heavy metal [71]. Meanwhile, researches have also shown that after repeated heat stress and acute ammonia exposure, there is a significant reduction in the levels of 5-HT and the expression of tph1 and tph2 in the brain [72,73]. Acute social stress has been reported to downregulate the transcription of Mao A gene expression and enhance 5-HT metabolism in the brain of tilapia [70]. Additionally, a study on rainbow trout demonstrated that MAO activity decreased in the brain stem and telencephalon but increased in the optic tectum and hypothalamus during confinement stress [74]. Serotonin exerts its functions by binding to specific serotonin receptors. In fish, seven families of 5-HT receptors with at least 39 genes have been identified [75]. Many of the receptors were reported to response various stressors [75,76]. Inhibition of 5-HT receptors has been shown to attenuate stress responses in flatfish [77], salmon [78], tilapia [68], and zebrafish [79], while the agonists of 5-HT receptors can lead to enhanced stress responses in the non-stressed fish [80]. In addition, some of the agonists of 5-HT receptors (i.e. buspirone and MK-212) decreased stress-induced behavior in fish [78,79,81]. Emerging evidence suggests individual variability in sensitivity to serotonin fluctuations within the same species. Fish exhibiting divergent stress-coping styles, such as proactive versus reactive coping, display differences in 5-HT-related functions. These differences were reflected in a generally higher brainstem 5-HT concentration and a lower telencephalic 5-HT activity, i.e. the ratio of 5-HIAA to 5-HT, in proactive fish compared to reactive one [82].
3.2. Neuroendocrine Systems During Stress
Brain-Sympathetic-Chromaffin Cells (BSC) Axis
The autonomic nervous system provides the most immediate ‘fight-or-flight’ responses to stressor (in seconds) through its two divisions, the sympathetic nervous system (SNS) as well as the parasympathetic nervous system (PNS). During stress, the sympathetic neurons in the spinal cord can release ACh and further activates acetylcholine receptors on chromaffin cells in the head kidney of the fish triggering depolarization and Ca²⁺ influx, which facilitates CAs release into the blood circulation and finally act on the peripheral target tissues [83,84]. In the PNS, efferent parasympathetic nerve signals are transduced from the central nervous system to their targets by preganglionic and postganglionic neurons. ACh is a major neurotransmitter in the PNS, and is activated when the stressful state is alleviated [85]. In general, the two systems usually regulate important functions in an antagonistic manner to achieve homeostasis during stress [84].
Glutamate is found to activate NMDA/AMPA receptors in brain regions (paraventricular neucleus, PVN) to increase sympathetic outflow [86], while the activation of mGluRs in PVN shows effectively reduce sympathetic outflow [87]. In addition, GABAergic inhibition in the PVN can reduce sympathetic activation by its GABAA and GABAB receptors [88]. Studies have demonstrated that chromaffin cells express various iGluRs and mGluRs, which exert distinct effects on the regulation of basal and glutamate-induced CA secretion through the nitric oxide/3’,5’-cyclic guanosine monophosphate (NO/cGMP) pathway. Additionally, glutamate-mediated activation of neuronal nitric oxide synthase (nNOS) in chromaffin cells highlights the involvement of both PKA and PKC signaling pathways in the apoptotic effects induced by glutamate [89]. In chromaffin cells, GABA is synthesized, stored in granules, and released upon nicotinic ACh receptor activation. Exogenous GABA stimulates GABAA receptors, inducing CA secretion via action potentials while inhibiting trans-synaptically evoked excitation [32]. GABA facilitates secretion when synaptic excitation is low but inhibits it when excitation is high [90]. Although chromaffin cells do not synthesize 5-HT, they accumulate small amounts through SERT-mediated uptake [91]. Recent studies have demonstrated that 5-HT1A receptors inhibit catecholamine (CA) secretion and exocytosis from adrenal chromaffin cells via an atypical mechanism that does not involve the modulation of cellular excitability, voltage-gated Ca2+ channels, potassium channels, or intracellular calcium levels [92].
The BSC axis can rapidly increase blood pressure and heart rate during acute stress, and quickly subside after the stress due to reflex parasympathetic activation [93]. However, chronic stress continues to activate the SNS, leading to increased synthesis, storage, and basal levels of CAs, as well as decreased levels of ACh [85]. In fish, it has been recognized that both acute and chronic stressors such as air exposure [94], handling stress [76], and hypoosmotic shock [95], can induce CA biosynthesis and elevate plasma CA concentrations. In coral trout, acute stress can trigger rapid changes in body color through the activation of catecholaminergic signaling. In vitro incubation with NE induces aggregation of chromatosomes while intraperitoneal injection leads to lightening of body color [96]. In gilthead sea bream, the elevation of plasma CAs was observed to be induced by chronic high stocking density and food deprivation, suggesting an activation of the fish’s BSC axis during chronic stress [58].
3.3. Hypothalamus-Pituitary-Interrenal (HPI) Axis
In the HPI axis, stress activates the hypophysiotropic neurons in the paraventricular nucleus (PVN) of the hypothalamus secreting arginine vasotocin (AVT) and the corticotropin-releasing hormone (CRH). These releasing hormones stimulate the anterior pituitary to generate pro-opiomelanocortin (POMC) which is processed into the adrenocoticotropic hormone (ACTH), also known as corticotropin, into the blood circulatory system. When ACTH reaches the head kidney, it binds with the melanocortin 2 receptor (MC2R) on cortisol-producing cells, resulting in the activation of steroidogenic acute regulatory protein (STAR) for the generation of corticosteroids, a class of steroid hormones [97]. Cortisol is a major corticosteroid in fish, which functions primarily to increase blood flow and heart rate, mobilize energy stores to muscle during stress by binding with corticosteroid receptors, namely the glucocorticoid receptor (GR) and mineralocorticoid receptor (MR), on the target cells [98]. Cortisol is the end-product of HPI axis activation and can also induce negative feedback on HPI axis, regulating the hypothalamic CRH and pituitary ACTH synthesis and secretion.
In fish, various stressors, such as temperature [99], crowding [100], hypoxia [101], light-dark cycles, and fasting [102], have been shown to activate the HPI axis, with cortisol levels widely recognized as indicators of the stress response. In rainbow trout and lake whitefish, the cortisol response to stressors immediately after hatching is absent when siblings experience stress during the embryonic stage [101,103]. The activation of the HPI axis typically occurs in response to acute and high-intensity stress, depending on both intensity and duration. Conversely, under chronic stress conditions, the capacity of the HPI axis to mount an appropriate stress response is diminished, particularly within the pituitary gland [104]. Notably, chronic heavy mental exposure has been found to alter the HPI axis and inhibit cortisol responses to acute stressors [105,106]. However, in Atlantic salmon, chronic stress was also observed to prolong HPI axis activation during eight weeks of daily crowding [100]. Additionally, the response of the HPI axis to stress has been shown to be influenced by maternal conditions. For example, maternal cortisol exposure or sequestration during embryogenesis can mediate the development and sensitivity of the HPI axis in their offspring [107,108].
Moreover, the neural and neuroendocrine system have largely complementary actions during stress. For instance, noradrenergic and adrenergic projections to the hypophysiotrophic zone of the PVN can participate in HPI activation [109]. ACh was known to stimulates cortisol secretion through mAChR in adrenal cells [110,111]. Meanwhile, cortisol was also reported to induce ACh accumulation and result in tissue damage and impairs regeneration [112]. CRH-releasing neurons have projections to the locus coeruleus which can directly send projection to the sympathetic and parasympathetic preganglionic neurons for enhancing the sympathetic activity and reducing the parasympathetic activity [85].
Strong evidence indicates that glutamate drives hypothalamic-pituitary-adrenal (HPA) axis stress responses through excitatory signaling mediated by iGluRs activity [113]. Meanwhile, other studies have shown that Glu may participate in the feedback loop of the HPA axis via kainate receptors and mGluRs, which inhibit HPA activation [114]. Additionally, cortisol secretion during acute stress induces the release of glutamate in limbic and cortical regions [115], while inhibition of glutamate receptors results in the stimulation of adrenocorticotropic hormone (ACTH) release during stress [116]. Taken together, these findings suggest that glutamate plays a complex role in the excitation of corticotropin-releasing hormone (CRH) neurons, acting at multiple levels to both drive HPA axis responses and limit over-activation. The role of GABA in HPA axis has been well established. CRH neurons have been shown to be regulated by both phasic and tonic GABAergic inhibition, which are distinct types of GABAergic inhibition mediated by specific subtypes of GABAA receptors [117]. Meanwhile, the cortisol also increase GABAergic signaling through changes in the expression of postsynaptic GABAARs and tonic GABAergic inhibition, contributing to the inhibition of CRH neurons [118]. These results provide novel negative feedback onto the HPA axis.
NE and E are known to stimulate hypothalamic CRH secretion via alpha 1 and alpha 2 receptors, while CA may also contribute to the feedback loop of HPA axis. Study investigated corticosterone’s long-term regulation of Crh and Avp gene expression in the PVH using a saporin-based immunocytotoxin targeting catecholaminergic neurons. In adrenalectomized animals with corticosterone replacement, corticosterone’s suppression of Crh expression required intact catecholaminergic innervation. Avp expression elevation post-adrenalectomy was similarly suppressed in lesioned animals. These findings demonstrate that corticosterone-catecholamine interactions critically modulate hypothalamic Crh and Avp expression [119]. In addition, another CA, DA is also found to regulate the activation of HPI axis during stress. By contacting with their receptors on CRH-containing neurons in the PVN, DA can regulate the HPI axis during stress [120]. Dopaminergic neurons are the primary source of DA, which also seems to regulate HPA axis [121]. Meanwhile, several dopaminergic regions in the brain express glucocorticoid receptors and administration of glucocorticoids can increase the level of extracellular DA [121].
Study has shown that midbrain serotonin neurons directly innervate CRH cells in the hypothalamic PVN to stimulate CRH and POMC release. This subsequently leads to ACTH secretion and activation of the hypothalamic-pituitary-adrenal (HPA) axis, mediated by 5-HT1A, 5-HT2A, 5-HT2C, and possibly also 5-HT1B receptors [122,123]. In human colonic enterochromaffin (EC) cells, EC cell-specific CRH1 expression and their activation pathways driving serotonin release/synthesis [124]. Meanwhile, chronic corticosterone exposure elevates TPH2 protein expression and mediate hypersensitivity of stress/anxiety circuits via opposing CRH receptor actions on serotonin synthesis [125].
3.4. Caudal Neurosecretory System (CNSS)
In addition to the common systems, fish possess a distinct neuroendocrine system located at the caudal end of the spinal cord, referred to as the CNSS. This system includes a neurohemal organ known as the urophysis and magnocellular peptide-synthesizing neuroendocrine neurons, termed Dahlgren cells, which were first described in 1914 [126]. CNSS plays the important roles in osmoregulation [127], thermoregulation [128], and other stress responses (such as ammonia, hypoxia, isolation and subordination) [129]. Studies have shown that the activities of the CNSS are innervated by the midbrain and medulla [130], then release neuropeptides into the circulation from tail vein. Recent studies demonstrated that some neurotransmitters, including DA [131], adrenaline [132], serotonin [133], taurine [134], nitric oxide [127], Glu [135] and GABA [136], were found to regulate the firing activities of Dahlgren cells through their receptors in CNSS, reflecting the crosstalk between nervous system and CNSS. The major synthesized neuropeptides in the CNSS are AVT, Isotocin (IT) [137], CRH, Urotensin I (UI, uts1) [138] and Urotensin II (UII, uts2) [139], parathyroid hormone related protein (PTHrP) [140]. It has been proved that the CNSS is the major circulating source of CRH, UI, UII, and PTHrP in fish [140,141,142].
Research has demonstrated that various stressors, including handling, osmotic pressure, thermal conditions, hyperammonemia, hypoxia, isolation, and subordination, can influence the expression of crh, uts1, and uts2 synthesized in the CNSS [127,129,143,144,145]. The expression changes of these genes in CNSS varied depending on the type of stressor: handling stress resulted in an increased expression of all three genes; acute seawater/freshwater transfer experiments elevated the expression of uts1 while decreasing that of uts2; acute hypothermal stress, but not hyperthermal stress, enhanced the expression of all three genes; subordination had no significant effect; isolation led to a reduction in the expression of crh and uts1, whereas hyperammonemia and hypoxia increased the expression levels of crh and/or uts1. In addition, both in vivo and in vitro studies showed that administration of UI and UII cause the increase in cortisol secretion [146]. In comparison to the individual administration of ACTH, UI, or UII, the enhanced release of cortisol following perifusion with a combination of ACTH and either UI or UII is significantly greater, indicating that interrenal UI and UII potentiate the steroidogenic effects of ACTH [147]. Given that fish possess relatively low blood pressure and slow fluid circulation velocity, the CNSS may play a role in stress responses by providing complementary regulation through the control of circulating levels of stress-related hormones and modulating cortisol production in relation to specific stressors. Current studies indicate that the contributions of the brain and CNSS to stress responses appear to be dependent on factors such as the type of stressor, duration, intensity, and specific regions within fish [148].
4. Dietary Supplements on Stress Mitigation Through Neuroendocrine and Neurotransmitters Regulations
Traditional pharmaceutical therapy in aquaculture normally focuses on the substance in the water that induces the infection after the onset of the disease, which is often palliative and induces secondary environmental pollution, for instance, using antibiotics and malachite green to treat bacterial and fungal infections. However, most of the infections occur after the failure of stress regulations. Using dietary supplements to preserve the beneficial effects and reduce the harmful effects of stress is more environment friendly and seems to be a viable and sustainable strategy to develop immunocompetence and disease resistance in aquatic animals. According to their components, they are divided into nutritional and non-nutritional supplements.
4.1. Nutritional Supplements
Recently, dietary intervention of amino acids, such as phenylalanine, has been considered to reduce the stress-induced physiological injury in fish through neuroendocrine and neurotransmitters regulations [149].
Phenylalanine (Phe) and tyrosine (Tyr) is the precursor of the neurotransmitters, catecholamines (NE, E and DA) [150,151,152]. Oral administration of the Phe-enriched diet is found to induce a high level of DA and minimize the effect of repeated handling stress in juvenile soles (Solea senegalensis) [153]. Dietary supplementation with 5% of Phe supplementation diet, shows a reduction on several stress markers (glucose, cortisol, NE, E, and DA) in meagres (Argyrosomus regius), but not in gilthead seabreams (Sparus aurata), during netting/chasing stress [154]. However, gilthead seabreams (S. aurata) fed with diets supplemented with 5% of Phe or Tyr showed a reduction in various stress markers during chonic confinement and netting/chasing stress [155] In addition, phenylalanine could also eliminate the reactive oxygen species (ROS) and mitigate the oxidative damage during stress [155,156]. However, it is still unclear that whether the stress mitigation is caused by amino acids themselves or their metabolites.
Tryptophan (Trp) is an essential amino acid that plays critical roles in the regulating the stress response. It serves as a precursor for serotonin and melatonin synthesis. Studies had demonstrated that dietary Trp at 0.72 % level supplementation could reduce serum cortisol levels and oxidative stress responses during thermal stress in rohu (Labeo rohita) [157]. Additionally, dietary Trp at 5g kg−1 levels had been shown to enhance intestinal antioxidant capacity and modulated inflammatory responses under high-stocking density conditions in rainbow trout (Oncorhynchus mykiss) [158]. Tryptophan supplementation had also been associated with decreased levels of POMC b-like peptides, which were involved in adrenocorticotropic hormone and cortisol secretion. When stressed fish (Dicentrarchus labrax) fed a Trp-supplemented diet were subjected to an inflammatory stimulus, plasma cortisol levels decreased and the expression of genes involved in the neuroendocrine response was altered, which seemed to be mediated by altered patterns in serotonergic activity [159].
Except the amino acids, some fatty acids and vitamins are also found to regulate cortisol release during stress. For instance, prostaglandin (PGE2) has been reported to be involved in modulating the HPI axis and affect the cortisol release in fish [160]. Vitamins are being used to mitigate various husbandry and physical stressors in aquaculture. For instance, dietary administration of vitamin C and E can protect the fish from death or other negative effects induced by various stressors, such as heat, hypoxia and handling [161,162,163]. More importantly, vitamin C and E seem to mitigate the stress by modulating the neuroendocrine and immune reactions. High dose vitamin C and E can block steroidogenesis by preventing the conversion of unsaturated lipids into cholesterol esters, and hindering the formation of cortisol [163,164,165].
4.2. Non-Nutritional Supplements
Except the nutritional additives, some non-nutritional compounds are also known to modulate immune function and enhance stress tolerance in fish. Levamisole, a synthesized phenyl imidazothiazole, has been reported to improve the stress tolerance [165,166]. Even with low supplementation of levamisole (0.1%), rainbow trout fingerlings show a higher tolerance during the crowded stress [167]. After 15-days oral administration of levamisole, it has been found that levamisole can modulate the plasma cortisol and enhance the innate immune responses against the bacterial infection in air-exposure-stressed pacu (Piaractus mesopotamicus) [168]. Clinical results in human showed that levamisole was helpful in reducing or stopping steroids and affecting the glucocorticosteroids level [169]. Another in vitro experiment showed that levamisole could significantly suppress the elevation of ROS, triggered by glutamate through stabilizing mitochondrial membrane potential, and protect the cell death [170]. However, levamisole mechanism of action in fish is poorly understood.
Recently, application of some biological derivatives, such as prebiotics and probiotics, also show a mitigation of stress via increasing the level of neurotransmitters in brain and modifying host-microbial community in fish intestine [171,172]. The application of mixed water probiotic is reported to reduce cortisol levels, improve tolerance to various husbandry stressors in fish [173,174].
In addition, some herbal medicines are also reported to demonstrate anti-stress effects in aquatic animals. The emodin, as the active component of anthraquinone extract, can protect the Wuchang bream (Megalobrama amblycephala) from the crowding stress not only by reducing the serum cortisol [163], which is also proved in common carp (Cyprinus carpio) [175]. By targeting the NF-κB signaling, the emodin improves the antioxidant capacity [176]. The in vitro experiment shows that the emodin can induce antioxidant defenses in peripheral blood leukocytes by Nrf2-Keap1 signaling [177]. Moreover, dietary supplementation with 1% turmeric can significantly reduce the plasma cortisol and enhance the lysozyme levels during transportation, which is used as an excellent feed additive to ameliorate the adverse effects of transportation in common carp (C. carpio) [178].
5. Conclusions and Perspectives
The steadily increasing demand for fish-derived protein necessitates the expansion and intensification of aquaculture practices, which in turn has heightened various environmental and husbandry-related stressors affecting fish. Fish possess well-developed endogenous nervous and endocrine systems that respond to stress, with complex communication networks among these systems demonstrating distributed collaboration. The nervous system plays a crucial role in detecting challenges, facilitating neural regulation, and innervating the endocrine system. Concurrently, the endocrine system subsequently releases a variety of stress-related hormones to reorganize energy resources, modify metabolic processes, and regulate immune responses. Future research on fish neural and neuroendocrine stress responses should focus on: 1) Investigating the specific mechanisms and differences in neuroendocrine stress responses across different fish species; 2) Understanding the coordinated regulatory mechanisms of the neuroendocrine stress system as fish respond to multiple stressors; 3) Enhancing and elucidating the interactions between the nervous system and neuroendocrine system in fish; 4) Exploring strategies to enhance aquaculture efficiency and improve survival rates by modulating neuroendocrine stress responses.
Author Contributions
Mingzhe Yuan: Conceptualization, Software, Investigation, Resources, Visualization, Writing - original draft, Writing - review & editing. Qian Fang: Investigation, Writing -original draft. Weiqun Lu: Conceptualization, Supervision. Xubo Wang: Supervision, Writing - review & editing. Cheong-Meng Chong: Conceptualization, Funding acquisition, Supervision, Writing - original draft, Writing - review & editing. Shan Chen: Funding acquisition.
Data availability statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
This work was supported by Ningbo Natural Science Foundation (Youth Foundation, 2024J449), Research Start-up Funding Project at Ningbo University (ZX2024000042 and ZX2024000043), and the Research Fund of University of Macau (File no. SRG2021-00013-ICMS and MYRG2020-00004-ICMS).
Conflicts of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- C. E. Boyd, A.A. McNevin, R.P. Davis, The contribution of fisheries and aquaculture to the global protein supply. Food Secur 2022, 14, 805–827. [Google Scholar] [CrossRef] [PubMed]
- R. L. Naylor, R.W. Hardy, A.H. Buschmann, S.R. Bush, L. Cao, D.H. Klinger, D.C. Little, J. Lubchenco, S.E. Shumway, M. Troell, A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar]
- Agorastos, G.P. Chrousos, The neuroendocrinology of stress: the stress-related continuum of chronic disease development. Mol Psychiatry 2022, 27, 502–513. [Google Scholar] [CrossRef]
- J. M. Koolhaas, A. Bartolomucci, B. Buwalda, S.F. de Boer, G. Flugge, S.M. Korte, P. Meerlo, R. Murison, B. Olivier, P. Palanza, G. Richter-Levin, A. Sgoifo, T. Steimer, O. Stiedl, G. van Dijk, M. Wohr, E. Fuchs, Stress revisited: a critical evaluation of the stress concept. Neurosci Biobehav Rev 2011, 35, 1291–1301. [Google Scholar]
- E.C. Urbinati, F.S. E.C. Urbinati, F.S. Zanuzzo, J.D. Biller, Stress and immune system in fish, Biology and physiology of freshwater neotropical fish, Elsevier2020, pp. 93-114.
- G. Nardocci, C. Navarro, P.P. Cortes, M. Imarai, M. Montoya, B. Valenzuela, P. Jara, C. Acuna-Castillo, R. Fernandez, Neuroendocrine mechanisms for immune system regulation during stress in fish. Fish & shellfish immunology 2014, 40, 531–538. [Google Scholar]
- F. S. Dhabhar, The short-term stress response - Mother nature’s mechanism for enhancing protection and performance under conditions of threat, challenge, and opportunity. Front Neuroendocrinol 2018, 49, 175–192. [Google Scholar] [CrossRef]
- R. Boonstra, C. Fox, Reality as the leading cause of stress: rethinking the impact of chronic stress in nature. Functional Ecology 2012, 27, 11–23. [Google Scholar]
- D.H. Evans, J.B. D.H. Evans, J.B. Claiborne, S. Currie, The physiology of fishes: Fourth edition, CRC Press, New York, 2013.
- R. Raman, C. Prakash, M. Makesh, N. Pawar, Environmental stress mediated diseases of fish: an overview. Adv Fish Res 2013, 5, 141–158. [Google Scholar]
- S. S.U.H. Kazmi, Y.Y.L. Wang, Y.-E. Cai, Z. Wang, Temperature effects in single or combined with chemicals to the aquatic organisms: An overview of thermo-chemical stress. Ecological Indicators 2022, 143, 109354. [Google Scholar] [CrossRef]
- M. L. Pfau, S.J. Russo, Peripheral and Central Mechanisms of Stress Resilience. Neurobiol Stress 2015, 1, 66–79. [Google Scholar] [CrossRef]
- M. Rima, Y. Lattouf, M. Abi Younes, E. Bullier, P. Legendre, J.M. Mangin, E. Hong, Dynamic regulation of the cholinergic system in the spinal central nervous system. Sci Rep 2020, 10, 15338. [Google Scholar] [CrossRef] [PubMed]
- X. Gu, X. Wang, An overview of recent analysis and detection of acetylcholine. Anal Biochem 2021, 632, 114381. [Google Scholar]
- Y. S. Mineur, T.N. Mose, L. Vanopdenbosch, I.M. Etherington, C. Ogbejesi, A. Islam, C.M. Pineda, R.B. Crouse, W. Zhou, D.C. Thompson, M.P. Bentham, M.R. Picciotto, Hippocampal acetylcholine modulates stress-related behaviors independent of specific cholinergic inputs. Mol Psychiatry 2022, 27, 1829–1838. [Google Scholar]
- S. Yang, T. Yan, L. Zhao, H. Wu, Z. Du, T. Yan, Q. Xiao, Effects of temperature on activities of antioxidant enzymes and Na(+)/K(+)-ATPase, and hormone levels in Schizothorax prenanti. J Therm Biol 2018, 72, 155–160. [Google Scholar] [CrossRef]
- S. Sarasamma, G. S. Sarasamma, G. Audira, S. Juniardi, B.P. Sampurna, S.-T. Liang, E. Hao, Y.-H. Lai, C.-D. Hsiao, Zinc Chloride Exposure Inhibits Brain Acetylcholine Levels, Produces Neurotoxic Signatures, and Diminishes Memory and Motor Activities in Adult Zebrafish. International Journal of Molecular Sciences, 2018.
- Szabo, J. Nemcsok, P. Kasa, D. Budai, Comparative study of acetylcholine synthesis in organs of freshwater teleosts. Fish Physiol Biochem 1991, 9, 93–99. [Google Scholar] [CrossRef]
- S. Bejaoui, I. Chetoui, F. Ghribi, D. Belhassen, B.B. Abdallah, C.B. Fayala, S. Boubaker, S. Mili, N. Soudani, Exposure to different cobalt chloride levels produces oxidative stress and lipidomic changes and affects the liver structure of Cyprinus carpio juveniles. Environ Sci Pollut Res Int 2024, 31, 51658–51672. [Google Scholar]
- S. J. Woo, J.K. Chung, Effects of trichlorfon on oxidative stress, neurotoxicity, and cortisol levels in common carp, Cyprinus carpio L., at different temperatures. Comp Biochem Physiol C Toxicol Pharmacol 2020, 229, 108698. [Google Scholar]
- S. Ghosh, R. Bhattacharya, S. Pal, N.C. Saha, Benzalkonium chloride induced acute toxicity and its multifaceted implications on growth, hematological metrics, biochemical profiles, and stress-responsive biomarkers in tilapia (Oreochromis mossambicus). Environ Sci Pollut Res Int 2024, 31, 52147–52170. [Google Scholar] [CrossRef]
- N. Deb, S. Das, Acetylcholine esterase and antioxidant responses in freshwater teleost, Channa punctata exposed to chlorpyrifos and urea. Comp Biochem Physiol C Toxicol Pharmacol 2021, 240, 108912. [Google Scholar]
- V. C. Renick, K. Weinersmith, D.E. Vidal-Dorsch, T.W. Anderson, Effects of a pesticide and a parasite on neurological, endocrine, and behavioral responses of an estuarine fish. Aquat Toxicol 2016, 170, 335–343. [Google Scholar] [CrossRef]
- A. M. Villalba, A. De la Llave-Propin, J. De la Fuente, C. Perez, E.G. de Chavarri, M.T. Diaz, A. Cabezas, R. Gonzalez-Garoz, F. Torrent, M. Villarroel, R. Bermejo-Poza, Using underwater currents as an occupational enrichment method to improve the stress status in rainbow trout. Fish Physiol Biochem 2024, 50, 463–475. [Google Scholar]
- S. Kar, B. Senthilkumaran, Recent advances in understanding neurotoxicity, behavior and neurodegeneration in siluriformes. Aquaculture and Fisheries 2024, 9, 404–410. [Google Scholar] [CrossRef]
- W. Jifa, Z. Yu, S. Xiuxian, W. You, Response of integrated biomarkers of fish (Lateolabrax japonicus) exposed to benzo[a]pyrene and sodium dodecylbenzene sulfonate. Ecotoxicol Environ Saf 2006, 65, 230–236. [Google Scholar] [CrossRef]
- H.F. Olivares-Rubio, J.J. H.F. Olivares-Rubio, J.J. Espinosa-Aguirre, Acetylcholinesterase activity in fish species exposed to crude oil hydrocarbons: A review and new perspectives. Chemosphere, 2021. [Google Scholar]
- T. Zhao, L. Zheng, Q. Zhang, S. Wang, Q. Zhao, G. Su, M. Zhao, Stability towards the gastrointestinal simulated digestion and bioactivity of PAYCS and its digestive product PAY with cognitive improving properties. Food Funct 2019, 10, 2439–2449. [Google Scholar] [CrossRef]
- Alzualde, O. Jaka, D. Latino, O. Alijevic, I. Iturria, J.H. de Mendoza, P. Pospisil, S. Frentzel, M.C. Peitsch, J. Hoeng, K. Koshibu, Effects of nicotinic acetylcholine receptor-activating alkaloids on anxiety-like behavior in zebrafish. J Nat Med 2021, 75, 926–941. [Google Scholar] [CrossRef]
- J. G. Santos da Rosa, H.H. Alcantara Barcellos, M. Fagundes, C. Variani, M. Rossini, F. Kalichak, G. Koakoski, T. Acosta Oliveira, R. Idalencio, R. Frandoloso, A.L. Piato, L. Jose Gil Barcellos, Muscarinic receptors mediate the endocrine-disrupting effects of an organophosphorus insecticide in zebrafish. Environ Toxicol 2017, 32, 1964–1972. [Google Scholar]
- J. Du, X.H. Li, Y.J. Li, Glutamate in peripheral organs: Biology and pharmacology. Eur J Pharmacol 2016, 784, 42–48. [Google Scholar] [CrossRef]
- K. Harada, H. Matsuoka, H. Fujihara, Y. Ueta, Y. Yanagawa, M. Inoue, GABA Signaling and Neuroactive Steroids in Adrenal Medullary Chromaffin Cells. Front Cell Neurosci 2016, 10, 100. [Google Scholar]
- D. Zhang, Z. Hua, Z. Li, The role of glutamate and glutamine metabolism and related transporters in nerve cells. CNS Neurosci Ther 2024, 30, e14617. [Google Scholar] [CrossRef]
- R. Sprengel, A. R. Sprengel, A. Eltokhi, Ionotropic Glutamate Receptors (and Their Role in Health and Disease), in: D.W. Pfaff, N.D. Volkow, J.L. Rubenstein (Eds.), Neuroscience in the 21st Century, Springer International Publishing, Cham, 2022, pp. 57-86.
- K. T. Lee, H.S. Liao, M.H. Hsieh, Glutamine Metabolism, Sensing and Signaling in Plants. Plant Cell Physiol 2023, 64, 1466–1481. [Google Scholar]
- Z. Heli, C. Hongyu, B. Dapeng, T. Yee Shin, Z. Yejun, Z. Xi, W. Yingying, Recent advances of gamma-aminobutyric acid: Physiological and immunity function, enrichment, and metabolic pathway. Front Nutr 2022, 9, 1076223. [Google Scholar] [CrossRef] [PubMed]
- N. Hossein-Javaheri, L.T. Buck, GABA receptor inhibition and severe hypoxia induce a paroxysmal depolarization shift in goldfish neurons. J Neurophysiol 2021, 125, 321–330. [Google Scholar] [CrossRef] [PubMed]
- N. J. Allen, D. Attwell, The effect of simulated ischaemia on spontaneous GABA release in area CA1 of the juvenile rat hippocampus. J Physiol 2004, 561, 485–498. [Google Scholar] [CrossRef] [PubMed]
- J. Jiang, X.Y. Wu, X.Q. Zhou, L. Feng, Y. Liu, W.D. Jiang, P. Wu, Y. Zhao, Glutamate ameliorates copper-induced oxidative injury by regulating antioxidant defences in fish intestine. Br J Nutr 2016, 116, 70–79. [Google Scholar] [CrossRef]
- M. Wang, E. Li, Y. Huang, W. Liu, S. Wang, W. Li, L. Chen, X. Wang, Dietary supplementation with glutamate enhanced antioxidant capacity, ammonia detoxification and ion regulation ability in Nile tilapia (Oreochromis niloticus) exposed to acute alkalinity stress. Aquaculture 2025, 594, 741360. [Google Scholar]
- L. Li, Z. Liu, J. Quan, J. Lu, G. Zhao, J. Sun, Metabonomics analysis reveals the protective effect of nano-selenium against heat stress of rainbow trout (Oncorhynchus mykiss). J Proteomics 2022, 259, 104545. [Google Scholar] [CrossRef]
- Y. Sun, C. Y. Sun, C. Geng, W. Liu, Y. Liu, L. Ding, P. Wang, Investigating the Impact of Disrupting the Glutamine Metabolism Pathway on Ammonia Excretion in Crucian Carp (Carassius auratus) under Carbonate Alkaline Stress Using Metabolomics Techniques, Antioxidants (Basel) 13(2) (2024).
- K. El-Naggar, S. El-Kassas, S.E. Abdo, A.A.K. Kirrella, R.A. Al Wakeel, Role of gamma-aminobutyric acid in regulating feed intake in commercial broilers reared under normal and heat stress conditions. J Therm Biol 2019, 84, 164–175. [Google Scholar] [CrossRef]
- S. Sohrabipour, M.R. Sharifi, A. Talebi, M. Sharifi, N. Soltani, GABA dramatically improves glucose tolerance in streptozotocin-induced diabetic rats fed with high-fat diet. Eur J Pharmacol 2018, 826, 75–84. [Google Scholar] [CrossRef]
- C. Zhang, X. Wang, C. Wang, Y. Song, J. Pan, Q. Shi, J. Qin, L. Chen, Gamma-aminobutyric acid regulates glucose homeostasis and enhances the hepatopancreas health of juvenile Chinese mitten crab (Eriocheir sinensis) under fasting stress. Gen Comp Endocrinol 2021, 303, 113704. [Google Scholar] [CrossRef]
- C. M. Ncho, A. Goel, V. Gupta, C.M. Jeong, Y.H. Choi, Embryonic manipulations modulate differential expressions of heat shock protein, fatty acid metabolism, and antioxidant-related genes in the liver of heat-stressed broilers. PLoS One 2022, 17, e0269748. [Google Scholar]
- J. Motiejunaite, L. Amar, E. Vidal-Petiot, Adrenergic receptors and cardiovascular effects of catecholamines. Ann Endocrinol 2021, 82, 193–197. [Google Scholar] [CrossRef] [PubMed]
- V. G. Barsagade, Dopamine system in the fish brain: A review on current knowledge. J Entomol Zool Stud 2020, 8, 2549–2555. [Google Scholar] [CrossRef]
- W. Joyce, J. Warwicker, H.A. Shiels, S.F. Perry, Evolution and divergence of teleost adrenergic receptors: why sometimes ’the drugs don’t work’ in fish. J Exp Biol 2023, 226, jeb245859. [Google Scholar]
- C. C. Lapish, S. Ahn, L.M. Evangelista, K. So, J.K. Seamans, A.G. Phillips, Tolcapone enhances food-evoked dopamine efflux and executive memory processes mediated by the rat prefrontal cortex. Psychopharmacology 2009, 202, 521–530. [Google Scholar] [CrossRef]
- T. C. Jhou, Dopamine and anti-dopamine systems: polar opposite roles in behavior. The FASEB Journal 2013, 27, 80.2–802. [Google Scholar]
- K. Domschke, B. Winter, A. Gajewska, S. Unterecker, B. Warrings, A. Dlugos, S. Notzon, K. Nienhaus, F. Markulin, A. Gieselmann, C. Jacob, M.J. Herrmann, V. Arolt, A. Muhlberger, A. Reif, P. Pauli, J. Deckert, P. Zwanzger, Multilevel impact of the dopamine system on the emotion-potentiated startle reflex. Psychopharmacology (Berl) 2015, 232, 1983–1993. [Google Scholar]
- J. Aerts, Quantification of a Glucocorticoid Profile in Non-pooled Samples Is Pivotal in Stress Research Across Vertebrates. Front Endocrinol (Lausanne) 2018, 9, 635. [Google Scholar] [CrossRef]
- L. Vargas-Chacoff, J.L.P. Muñoz, D. Ocampo, K. Paschke, J.M. Navarro, The effect of alterations in salinity and temperature on neuroendocrine responses of the Antarctic fish Harpagifer antarcticus. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 2019, 235, 131–137. [Google Scholar]
- S. Sreelekshmi, K. Manish, M.C. Subhash Peter, R. Moses Inbaraj, Analysis of neuroendocrine factors in response to conditional stress in zebrafish Danio rerio (Cypriniformes: Cyprinidae). Comp Biochem Physiol C Toxicol Pharmacol 2022, 252, 109242. [Google Scholar]
- W. Joyce, C.J.A. Williams, S. Iversen, P.G. Henriksen, M. Bayley, T. Wang, The effects of endogenous and exogenous catecholamines on hypoxic cardiac performance in red-bellied piranhas. J Exp Zool A Ecol Integr Physiol 2019, 331, 27–37. [Google Scholar] [CrossRef]
- A. N. Schoen, A.M. Weinrauch, I.A. Bouyoucos, J.R. Treberg, W. Gary Anderson, Hormonal effects on glucose and ketone metabolism in a perfused liver of an elasmobranch, the North Pacific spiny dogfish, Squalus suckleyi. Gen Comp Endocrinol 2024, 352, 114514. [Google Scholar] [CrossRef]
- M.A. Lopez-Patino, A.K. M.A. Lopez-Patino, A.K. Skrzynska, F. Naderi, J.M. Mancera, J.M. Miguez, J.A. Martos-Sitcha, High Stocking Density and Food Deprivation Increase Brain Monoaminergic Activity in Gilthead Sea Bream (Sparus aurata). Animals (Basel) 11(6) (2021).
- J. Roy, F. J. Roy, F. Terrier, M. Marchand, A. Herman, C. Heraud, A. Surget, A. Lanuque, F. Sandres, L. Marandel, Effects of Low Stocking Densities on Zootechnical Parameters and Physiological Responses of Rainbow Trout (Oncorhynchus mykiss) Juveniles, Biology (Basel) 10(10) (2021).
- K. Kaur, R.K. Narang, S. Singh, AlCl(3) induced learning and memory deficit in zebrafish. Neurotoxicology 2022, 92, 67–76. [Google Scholar] [CrossRef] [PubMed]
- J. Bedrossiantz, M. Bellot, P. Dominguez-Garcia, M. Faria, E. Prats, C. Gomez-Canela, R. Lopez-Arnau, E. Escubedo, D. Raldua, A Zebrafish Model of Neurotoxicity by Binge-Like Methamphetamine Exposure. Front Pharmacol 2021, 12, 770319. [Google Scholar]
- M. H.B. Amador, M.D. McDonald, Is serotonin uptake by peripheral tissues sensitive to hypoxia exposure?. Fish Physiol Biochem 2022, 48, 617–630.
- Mardones, R. Oyarzun-Salazar, B.S. Labbe, J.M. Miguez, L. Vargas-Chacoff, J.L.P. Munoz, Intestinal variation of serotonin, melatonin, and digestive enzymes activities along food passage time through GIT in Salmo salar fed with supplemented diets with tryptophan and melatonin. Comp Biochem Physiol A Mol Integr Physiol 2022, 266, 111159. [Google Scholar]
- E. Hoglund, O. Overli, S. Winberg, Tryptophan Metabolic Pathways and Brain Serotonergic Activity: A Comparative Review. Front Endocrinol (Lausanne) 2019, 10, 158. [Google Scholar] [CrossRef]
- C. Lillesaar, The serotonergic system in fish. J Chem Neuroanat 2011, 41, 294–308. [Google Scholar] [CrossRef]
- T. Backstrom, S. Winberg, Serotonin Coordinates Responses to Social Stress-What We Can Learn from Fish. Front Neurosci 2017, 11, 595. [Google Scholar] [CrossRef]
- N. Khan, P. Deschaux, Role of serotonin in fish immunomodulation. J Exp Biol 1997, 200, 1833–1838. [Google Scholar] [CrossRef]
- C. H. Lim, T. Soga, I.S. Parhar, Social stress-induced serotonin dysfunction activates spexin in male Nile tilapia (Oreochromis Niloticus). Proc Natl Acad Sci U S A 2023, 120, e2117547120. [Google Scholar] [CrossRef]
- S. Shams, D. Seguin, A. Facciol, D. Chatterjee, R. Gerlai, Effect of social isolation on anxiety-related behaviors, cortisol, and monoamines in adult zebrafish. Behav Neurosci 2017, 131, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Y. Higuchi, T. Soga, I.S. Parhar, Social Defeat Stress Decreases mRNA for Monoamine Oxidase A and Increases 5-HT Turnover in the Brain of Male Nile Tilapia (Oreochromis niloticus). Front Pharmacol 2018, 9, 1549. [Google Scholar]
- D. Santos, A. Luzio, L. Felix, J. Bellas, S.M. Monteiro, Oxidative stress, apoptosis and serotonergic system changes in zebrafish (Danio rerio) gills after long-term exposure to microplastics and copper. Comp Biochem Physiol C Toxicol Pharmacol 2022, 258, 109363. [Google Scholar]
- X. Gao, X. Wang, X. Wang, Y. Fang, S. Cao, B. Huang, H. Chen, R. Xing, B. Liu, Toxicity in Takifugu rubripes exposed to acute ammonia: Effects on immune responses, brain neurotransmitter levels, and thyroid endocrine hormones. Ecotoxicol Environ Saf 2022, 244, 114050. [Google Scholar] [CrossRef]
- Y. Shimomura, M. Inahata, M. Komori, N. Kagawa, Reduction of Tryptophan Hydroxylase Expression in the Brain of Medaka Fish After Repeated Heat Stress. Zoolog Sci 2019, 36, 223–230. [Google Scholar] [CrossRef]
- J. Schjolden, K.G. Pulman, T.G. Pottinger, O. Tottmar, S. Winberg, Serotonergic characteristics of rainbow trout divergent in stress responsiveness. Physiol Behav 2006, 87, 938–947. [Google Scholar] [CrossRef]
- Z. S. Hou, M.Q. Liu, H.S. Wen, Q.F. Gao, Z. Li, X.D. Yang, K.W. Xiang, Q. Yang, X. Hu, M.Z. Qian, J.F. Li, Identification, characterization, and transcription of serotonin receptors in rainbow trout (Oncorhynchus mykiss) in response to bacterial infection and salinity changes. Int J Biol Macromol 2023, 249, 125930. [Google Scholar]
- J. Martorell-Ribera, M.T. Venuto, W. Otten, R.M. Brunner, T. Goldammer, A. Rebl, U. Gimsa, Time-Dependent Effects of Acute Handling on the Brain Monoamine System of the Salmonid Coregonus maraena. Front Neurosci 2020, 14, 591738. [Google Scholar] [CrossRef]
- M. Shi, E.J. Rupia, P. Jiang, W. Lu, Switch from fight-flight to freeze-hide: The impacts of severe stress and brain serotonin on behavioral adaptations in flatfish. Fish Physiol Biochem 2024, 50, 891–909. [Google Scholar] [CrossRef]
- S. Shapouri, A. Sharifi, O. Folkedal, T.W.K. Fraser, M.A. Vindas, Behavioral and neurophysiological effects of buspirone in healthy and depression-like state juvenile salmon. Front Behav Neurosci 2024, 18, 1285413. [Google Scholar] [CrossRef]
- R. X. do Carmo Silva, B.G. do Nascimento, G.C.V. Gomes, N.A.H. da Silva, J.S. Pinheiro, S.N. da Silva Chaves, A.F.N. Pimentel, B.P.D. Costa, A.M. Herculano, M. Lima-Maximino, C. Maximino, 5-HT2C agonists and antagonists block different components of behavioral responses to potential, distal, and proximal threat in zebrafish. Pharmacol Biochem Behav 2021, 210, 173276. [Google Scholar]
- L. R. Medeiros, M.C. Cartolano, M.D. McDonald, Crowding stress inhibits serotonin 1A receptor-mediated increases in corticotropin-releasing factor mRNA expression and adrenocorticotropin hormone secretion in the Gulf toadfish. J Comp Physiol B 2014, 184, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Z. K. Varga, D. Pejtsik, L. Biro, A. Zsigmond, M. Varga, B. Toth, V. Salamon, T. Annus, E. Mikics, M. Aliczki, Conserved Serotonergic Background of Experience-Dependent Behavioral Responsiveness in Zebrafish (Danio rerio). J Neurosci 2020, 40, 4551–4564. [Google Scholar] [CrossRef] [PubMed]
- E. Hoglund, M. Moltesen, M.F. Castanheira, P.O. Thornqvist, P.I.M. Silva, O. Overli, C. Martins, S. Winberg, Contrasting neurochemical and behavioral profiles reflects stress coping styles but not stress responsiveness in farmed gilthead seabream (Sparus aurata). Physiol Behav 2020, 214, 112759. [Google Scholar]
- Y. M. Ulrich-Lai, J.P. Herman, Neural regulation of endocrine and autonomic stress responses. Nat Rev Neurosci 2009, 10, 397–409. [Google Scholar] [CrossRef]
- E. Scott-Solomon, E. Boehm, R. Kuruvilla, The sympathetic nervous system in development and disease. Nat Rev Neurosci 2021, 22, 685–702. [Google Scholar] [CrossRef]
- E. Won, Y.K. Kim, Stress, the Autonomic Nervous System, and the Immune-kynurenine Pathway in the Etiology of Depression. Curr Neuropharmacol 2016, 14, 665–673. [Google Scholar] [CrossRef]
- S. S. Kulkarni, N.A. Mischel, P.J. Mueller, Revisiting differential control of sympathetic outflow by the rostral ventrolateral medulla. Front Physiol 2022, 13, 1099513. [Google Scholar]
- J. J. Zhou, J. Pachuau, D.P. Li, S.R. Chen, H.L. Pan, Group III metabotropic glutamate receptors regulate hypothalamic presympathetic neurons through opposing presynaptic and postsynaptic actions in hypertension. Neuropharmacology 2020, 174, 108159. [Google Scholar] [CrossRef]
- R. J. Wang, Q.H. Zeng, W.Z. Wang, W. Wang, GABA(A) and GABA(B) receptor-mediated inhibition of sympathetic outflow in the paraventricular nucleus is blunted in chronic heart failure. Clin Exp Pharmacol Physiol 2009, 36, 516–522. [Google Scholar] [CrossRef]
- R. Perez-Rodriguez, A.M. Olivan, C. Roncero, J. Moron-Oset, M.P. Gonzalez, M.J. Oset-Gasque, Glutamate triggers neurosecretion and apoptosis in bovine chromaffin cells through a mechanism involving NO production by neuronal NO synthase activation. Free Radic Biol Med 2014, 69, 390–402. [Google Scholar] [CrossRef] [PubMed]
- M. Inoue, K. Harada, H. Matsuoka, A. Warashina, Paracrine role of GABA in adrenal chromaffin cells. Cell Mol Neurobiol 2010, 30, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- R. L. Brindley, M.B. Bauer, R.D. Blakely, K.P.M. Currie, Serotonin and Serotonin Transporters in the Adrenal Medulla: A Potential Hub for Modulation of the Sympathetic Stress Response. ACS Chem Neurosci 2017, 8, 943–954. [Google Scholar] [CrossRef]
- M. B. Bauer, R.L. Brindley, K.P. Currie, Serotonergic regulation of catecholamine exocytosis from adrenal chromaffin cells involves two mechanistically and temporally distinct pathways. Biophysical Journal 2024, 123, 382a. [Google Scholar]
- H. Kalamarz-Kubiak, Cortisol in Correlation to Other Indicators of Fish Welfare, Corticosteroids, IntechOpen, London, UK, 2018, pp. 155-183.
- A. K. Skrzynska, E. Maiorano, M. Bastaroli, F. Naderi, J.M. Miguez, G. Martinez-Rodriguez, J.M. Mancera, J.A. Martos-Sitcha, Impact of Air Exposure on Vasotocinergic and Isotocinergic Systems in Gilthead Sea Bream (Sparus aurata): New Insights on Fish Stress Response. Front Physiol 2018, 9, 96. [Google Scholar]
- E. S. Chelebieva, E.S. Kladchenko, I.V. Mindukshev, S. Gambaryan, A.Y. Andreyeva, ROS formation, mitochondrial potential and osmotic stability of the lamprey red blood cells: effect of adrenergic stimulation and hypoosmotic stress. Fish Physiol Biochem 2024, 50, 1341–1352. [Google Scholar] [CrossRef]
- N. Zhao, K. Jiang, X. Ge, J. Huang, C. Wu, S.X. Chen, Neurotransmitter norepinephrine regulates chromatosomes aggregation and the formation of blotches in coral trout Plectropomus leopardus. Fish Physiol Biochem 2024, 50, 705–719. [Google Scholar] [CrossRef]
- C. A. Shaughnessy, V.D. Myhre, D.J. Hall, S.D. McCormick, R.M. Dores, Hypothalamus-pituitary-interrenal (HPI) axis signaling in Atlantic sturgeon (Acipenser oxyrinchus) and sterlet (Acipenser ruthenus). Gen Comp Endocrinol 2023, 339, 114290. [Google Scholar] [CrossRef]
- E. Faught, M.J.M. Schaaf, Molecular mechanisms of the stress-induced regulation of the inflammatory response in fish. Gen Comp Endocrinol 2024, 345, 114387. [Google Scholar] [CrossRef]
- Torres-Martinez, R.S. Hattori, J.I. Fernandino, G.M. Somoza, S.D. Hung, Y. Masuda, Y. Yamamoto, C.A. Strussmann, Temperature- and genotype-dependent stress response and activation of the hypothalamus-pituitary-interrenal axis during temperature-induced sex reversal in pejerrey Odontesthes bonariensis, a species with genotypic and environmental sex determination. Mol Cell Endocrinol 2024, 582, 112114. [Google Scholar]
- M. I. Virtanen, M.H. Iversen, D.M. Patel, M.F. Brinchmann, Daily crowding stress has limited, yet detectable effects on skin and head kidney gene expression in surgically tagged atlantic salmon (Salmo salar). Fish & shellfish immunology 2024, 152, 109794. [Google Scholar]
- L. M. Whitehouse, E. Faught, M.M. Vijayan, R.G. Manzon, Hypoxia affects the ontogeny of the hypothalamus-pituitary-interrenal axis functioning in the lake whitefish (Coregonus clupeaformis). Gen Comp Endocrinol 2020, 295, 113524. [Google Scholar] [CrossRef] [PubMed]
- N. Saiz, M. N. Saiz, M. Gomez-Boronat, N. De Pedro, M.J. Delgado, E. Isorna, The Lack of Light-Dark and Feeding-Fasting Cycles Alters Temporal Events in the Goldfish (Carassius auratus) Stress Axis, Animals (Basel) 11(3) (2021).
- G. Ghaedi, B. Falahatkar, V. Yavari, M.T. Sheibani, G.N. Broujeni, The onset of stress response in rainbow trout Oncorhynchus mykiss embryos subjected to density and handling. Fish Physiol Biochem 2015, 41, 485–493. [Google Scholar] [CrossRef]
- Madaro, R.E. Olsen, T.S. Kristiansen, L.O. Ebbesson, T.O. Nilsen, G. Flik, M. Gorissen, Stress in Atlantic salmon: response to unpredictable chronic stress. J Exp Biol 2015, 218, 2538–2550. [Google Scholar]
- Z. Arab-Bafrani, E. Zabihi, S.M. Hoseini, H. Sepehri, M. Khalili, Silver nanoparticles modify the hypothalamic-pituitary-interrenal axis and block cortisol response to an acute stress in zebrafish, Danio rerio. Toxicol Ind Health 2022, 38, 201–209. [Google Scholar] [CrossRef]
- M. S. Tellis, D. Alsop, C.M. Wood, Effects of copper on the acute cortisol response and associated physiology in rainbow trout. Comp Biochem Physiol C Toxicol Pharmacol 2012, 155, 281–289. [Google Scholar] [CrossRef]
- M. Q. Zhuo, X. Chen, L. Gao, H.T. Zhang, Q.L. Zhu, J.L. Zheng, Y. Liu, Early life stage exposure to cadmium and zinc within hour affected GH/IGF axis, Nrf2 signaling and HPI axis in unexposed offspring of marine medaka Oryzias melastigma. Aquat Toxicol 2023, 261, 106628. [Google Scholar] [CrossRef]
- D. Nesan, M.M. Vijayan, Maternal Cortisol Mediates Hypothalamus-Pituitary-Interrenal Axis Development in Zebrafish. Sci Rep 2016, 6, 22582. [Google Scholar] [CrossRef]
- P. M. Plotsky, E.T. Cunningham, Jr., E.P. Widmaier, Catecholaminergic modulation of corticotropin-releasing factor and adrenocorticotropin secretion. Endocr Rev 1989, 10, 437–458. [Google Scholar] [CrossRef]
- S. W. Walker, M.W. Strachan, E.R. Lightly, B.C. Williams, I.M. Bird, Acetylcholine stimulates cortisol secretion through the M3 muscarinic receptor linked to a polyphosphoinositide-specific phospholipase C in bovine adrenal fasciculata/reticularis cells. Mol Cell Endocrinol 1990, 72, 227–238. [Google Scholar] [CrossRef]
- Y.M. Ulrich-Lai, W.C. Y.M. Ulrich-Lai, W.C. Engeland, Sympatho-adrenal activity and hypothalamic–pituitary–adrenal axis regulation, in: T. Steckler, N.H. Kalin, J.M.H.M. Reul (Eds.), Handbook of Stress and the Brain - Part 1: The Neurobiology of Stress, Elsevier2005, pp. 419-435.
- E.B. Ormaechea, M.E. E.B. Ormaechea, M.E. Cornide-Petronio, E. Negrete-Sánchez, C.G.Á. De León, A.I. Álvarez-Mercado, J. Gulfo, M.B. Jiménez Castro, J. Gracia-Sancho, C. Peralta, Effects of Cortisol-Induced Acetylcholine Accumulation on Tissue Damage and Regeneration in Steatotic Livers in the Context of Partial Hepatectomy Under Vascular Occlusion, Transplantation 102 (2018).
- Y. M. Ulrich-Lai, K.R. Jones, D.R. Ziegler, W.E. Cullinan, J.P. Herman, Forebrain origins of glutamatergic innervation to the rat paraventricular nucleus of the hypothalamus: differential inputs to the anterior versus posterior subregions. J Comp Neurol 2011, 519, 1301–1319. [Google Scholar] [CrossRef] [PubMed]
- N. K. Evanson, J.P. Herman, Role of Paraventricular Nucleus Glutamate Signaling in Regulation of HPA Axis Stress Responses. Interdiscip Inf Sci 2015, 21, 253–260. [Google Scholar]
- Sewanu Stephen Godonu, N. Francis-Lyons, Role Of Cortisol in The Synthesis of Glutamate During Oxidative Stress. Int. J. Sci. R. Tech. 2025, 2, 100–104. [Google Scholar]
- D. Jezova, E. Jurankova, M. Vigas, [Glutamate neurotransmission, stress and hormone secretion]. Bratisl Lek Listy 1995, 96, 588–596.
- J. Maguire, The relationship between GABA and stress: ’it’s complicated’. J Physiol 2018, 596, 1781–1782.
- P. L.W. Colmers, J.S. Bains, Balancing tonic and phasic inhibition in hypothalamic corticotropin-releasing hormone neurons. J Physiol 2018, 596, 1919–1929. [Google Scholar] [CrossRef]
- K. L. Kaminski, A.G. Watts, Intact catecholamine inputs to the forebrain are required for appropriate regulation of corticotrophin-releasing hormone and vasopressin gene expression by corticosterone in the rat paraventricular nucleus. J Neuroendocrinol 2012, 24, 1517–1526. [Google Scholar] [CrossRef]
- E. H. Douma, E.R. de Kloet, Stress-induced plasticity and functioning of ventral tegmental dopamine neurons. Neurosci Biobehav Rev 2020, 108, 48–77. [Google Scholar] [CrossRef]
- G.D. Stanwood, Dopamine and Stress, in: G. Fink (Ed.), Stress: Physiology, Biochemistry, and Pathology, Academic Press2019, pp. 105-114.
- N. R. Hanley, L.D. Van de Kar, Serotonin and the neuroendocrine regulation of the hypothalamic--pituitary-adrenal axis in health and disease. Vitam Horm 2003, 66, 189–255. [Google Scholar]
- H. Jorgensen, U. Knigge, A. Kjaer, M. Moller, J. Warberg, Serotonergic stimulation of corticotropin-releasing hormone and pro-opiomelanocortin gene expression. J Neuroendocrinol 2002, 14, 788–795. [Google Scholar] [CrossRef]
- S. V. Wu, P.Q. Yuan, J. Lai, K. Wong, M.C. Chen, G.V. Ohning, Y. Tache, Activation of Type 1 CRH receptor isoforms induces serotonin release from human carcinoid BON-1N cells: an enterochromaffin cell model. Endocrinology 2011, 152, 126–137. [Google Scholar] [CrossRef] [PubMed]
- N. C. Donner, P.H. Siebler, D.T. Johnson, M.D. Villarreal, S. Mani, A.J. Matti, C.A. Lowry, Serotonergic systems in the balance: CRHR1 and CRHR2 differentially control stress-induced serotonin synthesis. Psychoneuroendocrinology 2016, 63, 178–190. [Google Scholar] [CrossRef] [PubMed]
- U. Dahlgren, THE ELECTRIC MOTOR NERVE CENTERS IN THE SKATES (RAJIDAe). Science 1914, 40, 862–863.
- C. Cioni, E. C. Cioni, E. Angiulli, M. Toni, Nitric Oxide and the Neuroendocrine Control of the Osmotic Stress Response in Teleosts, Int J Mol Sci 20(3) (2019).
- M. Yuan, X. Li, W. Lu, The caudal neurosecretory system: A novel thermosensitive tissue and its signal pathway in olive flounder (Paralichthys olivaceus). J Neuroendocrinol 2020, 32, e12876. [Google Scholar] [CrossRef]
- N. J. Bernier, S.L. Alderman, E.N. Bristow, Heads or tails? Stressor-specific expression of corticotropin-releasing factor and urotensin I in the preoptic area and caudal neurosecretory system of rainbow trout. J Endocrinol 2008, 196, 637–648. [Google Scholar]
- J. P. O’Brien, R.M. Kriebel, Brain stem innervation of the caudal neurosecretory system. Cell Tissue Res 1982, 227, 153–160. [Google Scholar]
- Z. Lan, W. Zhang, J. Xu, M. Zhou, Y. Chen, H. Zou, W. Lu, Modulatory effect of dopamine receptor 5 on the neurosecretory Dahlgren cells of the olive flounder, Paralichthys olivaceus. Gen Comp Endocrinol 2018, 266, 67–77. [Google Scholar] [CrossRef]
- M. Shi, C. Liu, Y. Qin, L. Yv, W. Lu, alpha1 and beta3 adrenergic receptor-mediated excitatory effects of adrenaline on the caudal neurosecretory system (CNSS) in olive flounder, Paralichthys olivaceus. Gen Comp Endocrinol 2024, 349, 114468. [Google Scholar] [CrossRef]
- P. Jiang, S. Fang, N. Huang, W. Lu, The excitatory effect of 5-HT(1A) and 5-HT(2B) receptors on the caudal neurosecretory system Dahlgren cells in olive flounder, Paralichthys olivaceus. Comp Biochem Physiol A Mol Integr Physiol 2023, 283, 111457. [Google Scholar]
- W. Zhang, Z. Lan, K. Li, C. Liu, P. Jiang, W. Lu, Inhibitory role of taurine in the caudal neurosecretory Dahlgren cells of the olive flounder, Paralichthys olivaceus. Gen Comp Endocrinol 2020, 299, 113613. [Google Scholar] [CrossRef]
- Z. Lan, J. Xu, Y. Wang, W. Lu, Modulatory effect of glutamate GluR2 receptor on the caudal neurosecretory Dahlgren cells of the olive flounder, Paralichthys olivaceus. Gen Comp Endocrinol 2018, 261, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Z. Lan, W. Zhang, J. Xu, W. Lu, GABA(A) receptor-mediated inhibition of Dahlgren cells electrical activity in the olive flounder, Paralichthys olivaceus. Gen Comp Endocrinol 2021, 306, 113753. [Google Scholar] [CrossRef] [PubMed]
- M. Gozdowska, M. Slebioda, E. Kulczykowska, Neuropeptides isotocin and arginine vasotocin in urophysis of three fish species. Fish Physiol Biochem 2013, 39, 863–869. [Google Scholar] [CrossRef]
- H. Zhou, C. Ge, A. Chen, W. Lu, Dynamic Expression and Regulation of Urotensin I and Corticotropin-Releasing Hormone Receptors in Ovary of Olive Flounder Paralichthys olivaceus. Front Physiol 2019, 10, 1045. [Google Scholar] [CrossRef]
- X. Li, H. Zhou, C. Ge, K. Li, A. Chen, W. Lu, Dynamic changes of urotensin II and its receptor during ovarian development of olive flounder Paralichthys olivaceus. Comp Biochem Physiol B Biochem Mol Biol 2023, 263, 110782. [Google Scholar]
- W. Lu, Y. Jin, J. Xu, M.P. Greenwood, R.J. Balment, Molecular characterisation and expression of parathyroid hormone-related protein in the caudal neurosecretory system of the euryhaline flounder, Platichthys flesus. Gen Comp Endocrinol 2017, 249, 24–31. [Google Scholar] [CrossRef]
- W. Lu, L. Dow, S. Gumusgoz, M.J. Brierley, J.M. Warne, C.R. McCrohan, R.J. Balment, D. Riccardi, Coexpression of corticotropin-releasing hormone and urotensin i precursor genes in the caudal neurosecretory system of the euryhaline flounder (Platichthys flesus): a possible shared role in peripheral regulation. Endocrinology 2004, 145, 5786–5797. [Google Scholar]
- W. Lu, M. Greenwood, L. Dow, J. Yuill, J. Worthington, M.J. Brierley, C.R. McCrohan, D. Riccardi, R.J. Balment, Molecular characterization and expression of urotensin II and its receptor in the flounder (Platichthys flesus): a hormone system supporting body fluid homeostasis in euryhaline fish. Endocrinology 2006, 147, 3692–3708. [Google Scholar]
- Y. Qin, M. Shi, Y. Wei, W. Lu, The role of NMDA receptors in fish stress response: Assessments based on physiology of the caudal neurosecretory system and defensive behavior. J Neuroendocrinol 2024, 36, e13448. [Google Scholar] [CrossRef]
- M. Yuan, X. Li, T. Long, Y. Chen, W. Lu, Dynamic Responses of the Caudal Neurosecretory System (CNSS) Under Thermal Stress in Olive Flounder (Paralichthys olivaceus). Front Physiol 2019, 10, 1560. [Google Scholar]
- W. Lu, G. Zhu, A. Chen, X. Li, C.R. McCrohan, R. Balment, Gene expression and hormone secretion profile of urotensin I associated with osmotic challenge in caudal neurosecretory system of the euryhaline flounder, Platichthys flesus. Gen Comp Endocrinol 2019, 277, 49–55. [Google Scholar] [CrossRef] [PubMed]
- C. J. Kelsall, R.J. Balment, Native urotensins influence cortisol secretion and plasma cortisol concentration in the euryhaline flounder, platichthys flesus. Gen Comp Endocrinol 1998, 112, 210–219. [Google Scholar] [CrossRef]
- D. E. Arnold-Reed, R.J. Balment, Peptide hormones influence in vitro interrenal secretion of cortisol in the trout, Oncorhynchus mykiss. Gen Comp Endocrinol 1994, 96, 85–91. [Google Scholar] [CrossRef] [PubMed]
- K. Rousseau, F. Girardot, C. Parmentier, H. Tostivint, The Caudal Neurosecretory System: A Still Enigmatic Second Neuroendocrine Complex in Fish. Neuroendocrinology 2025, 115, 154–194. [Google Scholar]
- M. Herrera, J.M. Mancera, B. Costas, The Use of Dietary Additives in Fish Stress Mitigation: Comparative Endocrine and Physiological Responses. Front Endocrinol (Lausanne) 2019, 10, 447. [Google Scholar] [CrossRef]
- M. Herrera, M.A. Herves, I. Giraldez, K. Skar, H. Mogren, A. Mortensen, V. Puvanendran, Effects of amino acid supplementations on metabolic and physiological parameters in Atlantic cod (Gadus morhua) under stress. Fish Physiol Biochem 2017, 43, 591–602. [Google Scholar] [CrossRef]
- S. M. Hoseini, M. Ahmad Khan, M. Yousefi, B. Costas, Roles of arginine in fish nutrition and health: insights for future researches. Reviews in Aquaculture 2020, 12, 2091–2108. [Google Scholar] [CrossRef]
- N. Salamanca, I. N. Salamanca, I. Giráldez, E. Morales, I. de La Rosa, M. Herrera, Phenylalanine and Tyrosine as Feed Additives for Reducing Stress and Enhancing Welfare in Gilthead Seabream and Meagre, Animals, 2021.
- B. Costas, C. Aragao, J.L. Soengas, J.M. Miguez, P. Rema, J. Dias, A. Afonso, L.E. Conceicao, Effects of dietary amino acids and repeated handling on stress response and brain monoaminergic neurotransmitters in Senegalese sole (Solea senegalensis) juveniles. Comp Biochem Physiol A Mol Integr Physiol 2012, 161, 18–26. [Google Scholar] [CrossRef]
- N. Salamanca, I. Giráldez, E. Morales, I. de La Rosa, M. Herrera, Phenylalanine and Tyrosine as Feed Additives for Reducing Stress and Enhancing Welfare in Gilthead Seabream and Meagre. Animals-Basel 2021, 11, 45. [Google Scholar]
- N. Salamanca, O. Moreno, I. Giraldez, E. Morales, I. de la Rosa, M. Herrera, Effects of Dietary Phenylalanine and Tyrosine Supplements on the Chronic Stress Response in the Seabream (Sparus aurata). Front Physiol 2021, 12, 775771. [Google Scholar]
- W. Li, L. Feng, Y. Liu, W.D. Jiang, S.Y. Kuang, J. Jiang, S.H. Li, L. Tang, X.Q. Zhou, Effects of dietary phenylalanine on growth, digestive and brush border enzyme activities and antioxidant capacity in the hepatopancreas and intestine of young grass carp (Ctenopharyngodon idella). Aquaculture Nutrition 2015, 21, 913–925. [Google Scholar]
- P. Kumar, S. Saurabh, A.K. Pal, N.P. Sahu, A.R. Arasu, Stress mitigating and growth enhancing effect of dietary tryptophan in rohu (Labeo rohita, Hamilton, 1822) fingerlings. Fish Physiol Biochem 2014, 40, 1325–1338. [Google Scholar] [CrossRef] [PubMed]
- S. M. Hoseini, M. Yousefi, A. Taheri Mirghaed, B.A. Paray, S.H. Hoseinifar, H. Van Doan, Effects of rearing density and dietary tryptophan supplementation on intestinal immune and antioxidant responses in rainbow trout (Oncorhynchus mykiss). Aquaculture 2020, 528, 735537. [Google Scholar] [CrossRef]
- D. Peixoto, I. Carvalho, M. Machado, C. Aragao, B. Costas, R. Azeredo, Dietary tryptophan intervention counteracts stress-induced transcriptional changes in a teleost fish HPI axis during inflammation. Sci Rep 2024, 14, 7354. [Google Scholar] [CrossRef]
- Gupta, B. Lahlou, J. Botella, J. Porthe-Nibelle, In vivo and in vitro studies on the release of cortisol from interrenal tissue in trout. I. Effects of ACTH and prostaglandins. Exp Biol 1985, 43, 201–212. [Google Scholar]
- J. Niu, L.X. Tian, Y.J. Liu, K.S. Mai, H.J. Yang, C.X. Ye, W. Gao, Nutrient values of dietary ascorbic acid (l-ascorbyl-2-polyphosphate) on growth, survival and stress tolerance of larval shrimp,Litopenaeus vannamei. Aquaculture Nutrition 2009, 15, 194–201. [Google Scholar] [CrossRef]
- J. Ming, J. Xie, P. Xu, X. Ge, W. Liu, J. Ye, Effects of emodin and vitamin C on growth performance, biochemical parameters and two HSP70s mRNA expression of Wuchang bream (Megalobrama amblycephala Yih) under high temperature stress. Fish & shellfish immunology 2012, 32, 651–661. [Google Scholar]
- B. Liu, P. Xu, J. Xie, X. Ge, S. Xia, C. Song, Q. Zhou, L. Miao, M. Ren, L. Pan, R. Chen, Effects of emodin and vitamin E on the growth and crowding stress of Wuchang bream (Megalobrama amblycephala). Fish & shellfish immunology 2014, 40, 595–602. [Google Scholar]
- M. A.O. Dawood, S. Koshio, Vitamin C supplementation to optimize growth, health and stress resistance in aquatic animals. Reviews in Aquaculture 2016, 10, 334–350. [Google Scholar]
- Ciji, M.S. Akhtar, Stress management in aquaculture: a review of dietary interventions. Reviews in Aquaculture 2021, 13, 2190–2247. [Google Scholar] [CrossRef]
- A. H. Sherif, M.E. Mahfouz, Immune status of Oreochromis niloticus experimentally infected with Aeromonas hydrophila following feeding with 1, 3 β-glucan and levamisole immunostimulants. Aquaculture 2019, 509, 40–46. [Google Scholar] [CrossRef]
- S. Meshkini, N. Delirezh, A.A. Tafi, Effects of levamisole on immune responses and resistance against density stress in rainbow trout fingerling (Oncorhynchus mykiss). Journal of Veterinary Research 2017, 72, 129–136. [Google Scholar]
- E. Pahor-Filho, A.S.C. Castillo, N.L. Pereira, F. Pilarski, E.C. Urbinati, Levamisole enhances the innate immune response and prevents increased cortisol levels in stressed pacu (Piaractus mesopotamicus). Fish & shellfish immunology 2017, 65, 96–102. [Google Scholar]
- L. Jiang, I. Dasgupta, J.A. Hurcombe, H.F. Colyer, P.W. Mathieson, G.I. Welsh, Levamisole in steroid-sensitive nephrotic syndrome: usefulness in adult patients and laboratory insights into mechanisms of action via direct action on the kidney podocyte. Clin Sci (Lond) 2015, 128, 883–893. [Google Scholar] [CrossRef]
- M. Shukry, T. Kamal, R. Ali, F. Farrag, E. Almadaly, A.A. Saleh, M. Abu El-Magd, Pinacidil and levamisole prevent glutamate-induced death of hippocampal neuronal cells through reducing ROS production. Neurol Res 2015, 37, 916–923. [Google Scholar] [CrossRef]
- M. A.O. Dawood, N.M. Eweedah, E.M. Moustafa, E.M. Farahat, Probiotic effects of Aspergillus oryzae on the oxidative status, heat shock protein, and immune related gene expression of Nile tilapia (Oreochromis niloticus) under hypoxia challenge. Aquaculture 2020, 520, 734669. [Google Scholar] [CrossRef]
- Chudzik, A. Orzylowska, R. Rola, G.J. Stanisz, Probiotics, Prebiotics and Postbiotics on Mitigation of Depression Symptoms: Modulation of the Brain-Gut-Microbiome Axis. Biomolecules 2021, 11, 1000. [Google Scholar] [CrossRef]
- L. C. Gomes, R.P. Brinn, J.L. Marcon, L.A. Dantas, F.R. Brandão, J.S. de Abreu, P.E.M. Lemos, D.M. McComb, B. Baldisserotto, Benefits of using the probiotic Efinol®L during transportation of cardinal tetra,Paracheirodon axelrodi(Schultz), in the Amazon. Aquaculture Research 2009, 40, 157–165. [Google Scholar] [CrossRef]
- N. Eissa, H.P. Wang, H. Yao, E. Abou-ElGheit, Mixed Bacillus Species Enhance the Innate Immune Response and Stress Tolerance in Yellow Perch Subjected to Hypoxia and Air-Exposure Stress. Sci Rep 2018, 8, 6891. [Google Scholar] [CrossRef]
- J. Xie, B. Liu, Q. Zhou, Y. Su, Y. He, L. Pan, X. Ge, P. Xu, Effects of anthraquinone extract from rhubarb Rheum officinale Bail on the crowding stress response and growth of common carp Cyprinus carpio var. Jian. Aquaculture 2008, 281, 5–11. [Google Scholar] [CrossRef]
- C. Song, B. Liu, J. Xie, X. Ge, Z. Zhao, Y. Zhang, H. Zhang, M. Ren, Q. Zhou, L. Miao, P. Xu, Y. Lin, Comparative proteomic analysis of liver antioxidant mechanisms in Megalobrama amblycephala stimulated with dietary emodin. Sci Rep 2017, 7, 40356. [Google Scholar] [CrossRef] [PubMed]
- D. A. Tadese, C. Song, C. Sun, B. Liu, B. Liu, Q. Zhou, P. Xu, X. Ge, M. Liu, X. Xu, M. Tamiru, Z. Zhou, A. Lakew, N.T. Kevin, The role of currently used medicinal plants in aquaculture and their action mechanisms: A review. Reviews in Aquaculture 2021, 14, 816–847. [Google Scholar]
- S. M. Hoseini, S.K. Gupta, M. Yousefi, E.V. Kulikov, S.G. Drukovsky, A.K. Petrov, A. Taheri Mirghaed, S.H. Hoseinifar, H. Van Doan, Mitigation of transportation stress in common carp, Cyprinus carpio, by dietary administration of turmeric. Aquaculture 2022, 546, 737380. [Google Scholar] [CrossRef]
- C.-H. Chang, Y.-C. C.-H. Chang, Y.-C. Wang, T.-H. Lee, Hypothermal stress-induced salinity-dependent oxidative stress and apoptosis in the livers of euryhaline milkfish, Chanos chanos, Aquaculture 534 (2021).
- C. Hassenruck, H. C. Hassenruck, H. Reinwald, A. Kunzmann, I. Tiedemann, A. Gardes, Effects of Thermal Stress on the Gut Microbiome of Juvenile Milkfish (Chanos chanos), Microorganisms 9(1) (2020).
- Topal, S. Ozdemir, H. Arslan, S. Comakli, How does elevated water temperature affect fish brain? (A neurophysiological and experimental study: Assessment of brain derived neurotrophic factor, cFOS, apoptotic genes, heat shock genes, ER-stress genes and oxidative stress genes). Fish & shellfish immunology 2021, 115, 198–204. [Google Scholar]
- X. Li, P. X. Li, P. Wei, S. Liu, Y. Tian, H. Ma, Y. Liu, Photoperiods affect growth, food intake and physiological metabolism of juvenile European Sea Bass (Dicentrachus labrax L.). Aquacult Rep 20 (2021).
- P. Konkal, C.B. Ganesh, Continuous Exposure to Light Suppresses the Testicular Activity in Mozambique Tilapia Oreochromis mossambicus (Cichlidae). J Ichthyol+ 2020, 60, 660–667.
- S. Suzuki, E. S. Suzuki, E. Takahashi, T.O. Nilsen, N. Kaneko, H. Urabe, Y. Ugachi, E. Yamaha, M. Shimizu, Physiological changes in off-season smolts induced by photoperiod manipulation in masu salmon (Oncorhynchus masou), Aquaculture 526 (2020).
- Y. P. Sapozhnikova, A.G. Koroleva, V.M. Yakhnenko, M.L. Tyagun, O.Y. Glyzina, A.B. Coffin, M.M. Makarov, A.N. Shagun, V.A. Kulikov, P.V. Gasarov, S.V. Kirilchik, I.V. Klimenkov, N.P. Sudakov, P.N. Anoshko, N.A. Kurashova, L.V. Sukhanova, Molecular and cellular responses to long-term sound exposure in peled (Coregonus peled). J Acoust Soc Am 2020, 148, 895. [Google Scholar]
- H. Kusku, S. Ergun, S. Yilmaz, B. Guroy, M. Yigit, Impacts of Urban Noise and Musical Stimuli on Growth Performance and Feed Utilization of Koi fish (Cyprinus carpio) in Recirculating Water Conditions. Turk J Fish Aquat Sc 2019, 19, 513–523. [Google Scholar]
- H. Kusku, Ü. Yigit, S. Yilmaz, M. Yigit, S. Ergün, Acoustic effects of underwater drilling and piling noise on growth and physiological response of Nile tilapia (Oreochromis niloticus). Aquaculture Research 2020, 51, 3166–3174. [Google Scholar] [CrossRef]
- B. Falahatkar, A.S. Amlashi, M. Kabir, Ventilation frequency in juvenile beluga sturgeon Huso huso L. exposed to clay turbidity. J Exp Mar Biol Ecol 2019, 513, 10–12. [Google Scholar] [CrossRef]
- T. C.T. Phan, A.V. Manuel, N. Tsutsui, T. Yoshimatsu, Impacts of short-term salinity and turbidity stress on the embryonic stage of red sea bream Pagrus major. Fisheries Sci 2019, 86, 119–125. [Google Scholar]
- M. Hasenbein, N.A. Fangue, J. Geist, L.M. Komoroske, J. Truong, R. McPherson, R.E. Connon, Assessments at multiple levels of biological organization allow for an integrative determination of physiological tolerances to turbidity in an endangered fish species. Conserv Physiol 2016, 4, cow004. [Google Scholar] [CrossRef] [PubMed]
- M. D.D. Carneiro, L.C. Maltez, R.V. Rodrigues, M. Planas, L.A. Sampaio, Does acidification lead to impairments on oxidative status and survival of orange clownfish Amphiprion percula juveniles?. Fish Physiol Biochem 2021, 47, 841–848.
- M. D.D. Carneiro, S. Garcia-Mesa, L.A. Sampaio, M. Planas, Primary, secondary, and tertiary stress responses of juvenile seahorse Hippocampus reidi exposed to acute acid stress in brackish and seawater. Comp Biochem Physiol B Biochem Mol Biol 2021, 255, 110592. [Google Scholar]
- L. Pellegrin, L.F. L. Pellegrin, L.F. Nitz, L.C. Maltez, C.E. Copatti, L. Garcia, Alkaline water improves the growth and antioxidant responses of pacu juveniles (Piaractus mesopotamicus), Aquaculture 519 (2020).
- Bal, S.G. Pati, F. Panda, L. Mohanty, B. Paital, Low salinity induced challenges in the hardy fish Heteropneustes fossilis; future prospective of aquaculture in near coastal zones, Aquaculture 543 (2021).
- M. A.O. Dawood, A.E. Noreldin, H. Sewilam, Long term salinity disrupts the hepatic function, intestinal health, and gills antioxidative status in Nile tilapia stressed with hypoxia. Ecotoxicol Environ Saf 2021, 220, 112412. [Google Scholar] [CrossRef]
- N.A. Mohamed, M.F. N.A. Mohamed, M.F. Saad, M. Shukry, A.M.S. El-Keredy, O. Nasif, H. Van Doan, M.A.O. Dawood, Physiological and ion changes of Nile tilapia (Oreochromis niloticus) under the effect of salinity stress, Aquacult Rep 19 (2021).
- J. Zeng, N.A. Herbert, W. Lu, Differential Coping Strategies in Response to Salinity Challenge in Olive Flounder. Front Physiol 2019, 10, 1378. [Google Scholar] [CrossRef]
- J.-s. Huang, Z.-x. J.-s. Huang, Z.-x. Guo, J.-d. Zhang, W.-z. Wang, Z.-l. Wang, E. Amenyogbe, G. Chen, Effects of hypoxia-reoxygenation conditions on serum chemistry indicators and gill and liver tissues of cobia (Rachycentron canadum), Aquacult Rep 20 (2021).
- N. Schafer, J. N. Schafer, J. Matousek, A. Rebl, V. Stejskal, R.M. Brunner, T. Goldammer, M. Verleih, T. Korytar, Effects of Chronic Hypoxia on the Immune Status of Pikeperch (Sander lucioperca Linnaeus, 1758), Biology (Basel) 10(7) (2021).
- X. Zheng, D. X. Zheng, D. Fu, J. Cheng, R. Tang, M. Chu, P. Chu, T. Wang, S. Yin, Effects of hypoxic stress and recovery on oxidative stress, apoptosis, and intestinal microorganisms in Pelteobagrus vachelli, Aquaculture 543 (2021).
- H. Su, D. Ma, H. Zhu, Z. Liu, F. Gao, Transcriptomic response to three osmotic stresses in gills of hybrid tilapia (Oreochromis mossambicus female x O. urolepis hornorum male). Bmc Genomics 2020, 21, 110. [Google Scholar]
- Y. Zhao, C. Zhang, H. Zhou, L. Song, J. Wang, J. Zhao, Transcriptome changes for Nile tilapia (Oreochromis niloticus) in response to alkalinity stress. Comp Biochem Physiol Part D Genomics Proteomics 2020, 33, 100651. [Google Scholar]
- N. M. Noor, M. De, Z.C. Cob, S.K. Das, Welfare of scaleless fish, Sagor catfish ( Hexanematichthys sagor) juveniles under different carbon dioxide concentrations. Aquaculture Research 2021, 52, 2980–2987. [Google Scholar] [CrossRef]
- V.C. Mota, T.O. V.C. Mota, T.O. Nilsen, J. Gerwins, M. Gallo, J. Kolarevic, A. Krasnov, B.F. Terjesen, Molecular and physiological responses to long-term carbon dioxide exposure in Atlantic salmon (Salmo salar), Aquaculture 519 (2020).
- M. Machado, F. Arenas, J.C. Svendsen, R. Azeredo, L.J. Pfeifer, J.M. Wilson, B. Costas, Effects of Water Acidification on Senegalese Sole Solea senegalensis Health Status and Metabolic Rate: Implications for Immune Responses and Energy Use. Front Physiol 2020, 11, 26. [Google Scholar] [CrossRef]
- Y. Zhang, X.-F. Y. Zhang, X.-F. Liang, S. He, L. Li, Effects of long-term low-concentration nitrite exposure and detoxification on growth performance, antioxidant capacities, and immune responses in Chinese perch (Siniperca chuatsi), Aquaculture 533 (2021).
- J. Yu, Y. Xiao, Y. Wang, S. Xu, L. Zhou, J. Li, X. Li, Chronic nitrate exposure cause alteration of blood physiological parameters, redox status and apoptosis of juvenile turbot (Scophthalmus maximus). Environ Pollut 2021, 283, 117103. [Google Scholar] [CrossRef]
- X. Q. Gao, F. Fei, H.H. Huo, B. Huang, X.S. Meng, T. Zhang, B.L. Liu, Impact of nitrite exposure on plasma biochemical parameters and immune-related responses in Takifugu rubripes. Aquat Toxicol 2020, 218, 105362. [Google Scholar] [CrossRef]
- S. Xue, J. Lin, Y. Han, Y. Han, Ammonia stress–induced apoptosis by p53–BAX/BCL-2 signal pathway in hepatopancreas of common carp (Cyprinus carpio). Aquacult Int 2021, 29, 1895–1907. [Google Scholar] [CrossRef]
- S. Xue, J. S. Xue, J. Lin, Q. Zhou, H. Wang, Y. Han, Effect of ammonia stress on transcriptome and endoplasmic reticulum stress pathway for common carp (Cyprinus carpio) hepatopancreas, Aquacult Rep 20 (2021).
- S. Cao, D. S. Cao, D. Zhao, R. Huang, Y. Xiao, W. Xu, X. Liu, Y. Gui, S. Li, J. Xu, J. Tang, F. Qu, Z. Liu, S. Liu, The influence of acute ammonia stress on intestinal oxidative stress, histology, digestive enzymatic activities and PepT1 activity of grass carp (Ctenopharyngodon idella), Aquacult Rep 20 (2021).
- M. C.B. Kiemer, K.D. Black, D. Lussot, A.M. Bullock, I. Ezzi, The effects of chronic and acute exposure to hydrogen sulphide on Atlantic salmon (Salmo salar L.). Aquaculture 1995, 135, 311–327.
- T. Bagarinao, I. Lantin-Olaguer, The sulfide tolerance of milkfish and tilapia in relation to fish kills in farms and natural waters in the Philippines, Hydrobiologia 1998, 382, 137–150.
- V. Valenzuela-Muñoz, D. V. Valenzuela-Muñoz, D. Valenzuela-Miranda, A.T. Gonçalves, B. Novoa, A. Figueras, C. Gallardo-Escárate, Induced-iron overdose modulate the immune response in Atlantic salmon increasing the susceptibility to Piscirickettsia salmonis infection, Aquaculture 521 (2020).
- M. Singh, A.S. Barman, A.L. Devi, A.G. Devi, P.K. Pandey, Iron mediated hematological, oxidative and histological alterations in freshwater fish Labeo rohita. Ecotoxicol Environ Saf 2019, 170, 87–97. [Google Scholar] [CrossRef]
- S. Asif, M. Javed, S. Abbas, F. Ambreen, S. Iqbal, Growth responses of carnivorous fish species under the chronic stress of water-borne copper. Iran J Fish Sci 2021, 20, 773–788. [Google Scholar]
- J. S. Paul, B.C. Small, Chronic exposure to environmental cadmium affects growth and survival, cellular stress, and glucose metabolism in juvenile channel catfish (Ictalurus punctatus). Aquat Toxicol 2021, 230, 105705. [Google Scholar] [CrossRef]
- Kakade, E.S. Salama, F. Pengya, P. Liu, X. Li, Long-term exposure of high concentration heavy metals induced toxicity, fatality, and gut microbial dysbiosis in common carp, Cyprinus carpio. Environ Pollut 2020, 266, 115293. [Google Scholar] [CrossRef]
- Y. Kong, M. Li, X. Shan, G. Wang, G. Han, Effects of deltamethrin subacute exposure in snakehead fish, Channa argus: Biochemicals, antioxidants and immune responses. Ecotoxicol Environ Saf 2021, 209, 111821. [Google Scholar] [CrossRef]
- L. Zhao, G. Tang, C. Xiong, S. Han, C. Yang, K. He, Q. Liu, J. Luo, W. Luo, Y. Wang, Z. Li, S. Yang, Chronic chlorpyrifos exposure induces oxidative stress, apoptosis and immune dysfunction in largemouth bass (Micropterus salmoides). Environ Pollut 2021, 282, 117010. [Google Scholar] [CrossRef]
- M. D. Baldissera, C.F. Souza, R. Zanella, O.D. Prestes, A.D. Meinhart, A.S. Da Silva, B. Baldisserotto, Behavioral impairment and neurotoxic responses of silver catfish Rhamdia quelen exposed to organophosphate pesticide trichlorfon: Protective effects of diet containing rutin. Comp Biochem Physiol C Toxicol Pharmacol 2021, 239, 108871. [Google Scholar]
- P. V. Zimba, L. Khoo, P.S. Gaunt, S. Brittain, W.W. Carmichael, Confirmation of catfish, Ictalurus punctatus (Rafinesque), mortality from Microcystis toxins. J Fish Dis 2008, 24, 41–47. [Google Scholar]
- M. J. Bakke, T.E. Horsberg, Effects of algal-produced neurotoxins on metabolic activity in telencephalon, optic tectum and cerebellum of Atlantic salmon (Salmo salar). Aquat Toxicol 2007, 85, 96–103. [Google Scholar] [CrossRef]
- M. A.O. Dawood, E.M. Moustafa, M.S. Gewaily, S.E. Abdo, M.F. AbdEl-Kader, M.S. SaadAllah, A.H. Hamouda, Ameliorative effects of Lactobacillus plantarum L-137 on Nile tilapia (Oreochromis niloticus) exposed to deltamethrin toxicity in rearing water. Aquat Toxicol 2020, 219, 105377. [Google Scholar] [CrossRef]
- X. Chen, H. X. Chen, H. Yi, S. Liu, Y. Zhang, Y. Su, X. Liu, S. Bi, H. Lai, Z. Zeng, G. Li, Promotion of pellet-feed feeding in mandarin fish (Siniperca chuatsi) by Bdellovibrio bacteriovorus is influenced by immune and intestinal flora, Aquaculture 542 (2021).
- A. I. Zaineldin, S. Hegazi, S. Koshio, M. Ishikawa, M.A.O. Dawood, S. Dossou, Z. Yukun, K. Mzengereza, Singular effects of Bacillus subtilis C-3102 or Saccharomyces cerevisiae type 1 on the growth, gut morphology, immunity, and stress resistance of red sea bream (Pagrus major). Ann Anim Sci 2021, 21, 589–608. [Google Scholar]
- L. Kolek, J. L. Kolek, J. Szczygieł, Ł. Napora-Rutkowski, I. Irnazarow, Effect of Trypanoplasma borreli infection on sperm quality and reproductive success of common carp (Cyprinus carpio L.) males, Aquaculture 539 (2021).
- X. Xie, J. Kong, J. Huang, L. Zhou, Y. Jiang, R. Miao, F. Yin, Integration of metabolomic and transcriptomic analyses to characterize the influence of the gill metabolism of Nibea albiflora on the response to Cryptocaryon irritans infection. Vet Parasitol 2021, 298, 109533. [Google Scholar] [CrossRef]
- M.P. Gonzalez, S.L. M.P. Gonzalez, S.L. Marin, M. Mancilla, H. Canon-Jones, L. Vargas-Chacoff, Fin Erosion of Salmo salar (Linnaeus 1758) Infested with the Parasite Caligus rogercresseyi (Boxshall & Bravo 2000), Animals (Basel) 10(7) (2020).
- K. Filipsson, E. Bergman, L. Greenberg, M. Osterling, J. Watz, A. Erlandsson, Temperature and predator-mediated regulation of plasma cortisol and brain gene expression in juvenile brown trout (Salmo trutta). Front Zool 2020, 17, 25. [Google Scholar] [CrossRef]
- R. Kortet, M.V.M. Laakkonen, J. Tikkanen, A. Vainikka, H. Hirvonen, Size-dependent stress response in juvenile Arctic charr (Salvelinus alpinus) under prolonged predator conditioning. Aquaculture Research 2019, 50, 1482–1490. [Google Scholar] [CrossRef]
- H. Ling, S.J. Fu, L.Q. Zeng, Predator stress decreases standard metabolic rate and growth in juvenile crucian carp under changing food availability. Comp Biochem Physiol A Mol Integr Physiol 2019, 231, 149–157. [Google Scholar] [CrossRef]
- T. N. Barreto, C.N.P. Boscolo, E. Gonçalves-de-Freitas, Homogeneously sized groups increase aggressive interaction and affect social stress in Thai strain Nile tilapia (Oreochromis niloticus). Mar Freshw Behav Phy 2015, 48, 309–318. [Google Scholar] [CrossRef]
- L. M. Gomez-Laplaza, E. Morgan, The influence of social rank in the angelfish, Pterophyllum scalare, on locomotor and feeding activities in a novel environment. Lab Anim 2003, 37, 108–120. [Google Scholar] [CrossRef]
- K. A. Sloman, K.M. Gilmour, A.C. Taylor, N.B. Metcalfe, Physiological effects of dominance hierarchies within groups of brown trout, Salmo trutta, held under simulated natural conditions. Fish Physiology and Biochemistry 2000, 22, 11–20. [Google Scholar] [CrossRef]
- Z. Zhang, Y. Z. Zhang, Y. Fu, H. Guo, X. Zhang, Effect of environmental enrichment on the stress response of juvenile black rockfish Sebastes schlegelii, Aquaculture 533 (2021).
- Z. Zhang, Y. Z. Zhang, Y. Fu, Z. Zhang, X. Zhang, S. Chen, A Comparative Study on Two Territorial Fishes: The Influence of Physical Enrichment on Aggressive Behavior, Animals (Basel) 11(7) (2021).
- Z. Zhang, X. Z. Zhang, X. Xu, Y. Wang, X. Zhang, Effects of environmental enrichment on growth performance, aggressive behavior and stress-induced changes in cortisol release and neurogenesis of black rockfish Sebastes schlegelii, Aquaculture 528 (2020).
- L. Li, Y. L. Li, Y. Shen, W. Yang, X. Xu, J. Li, Effect of different stocking densities on fish growth performance: A meta-analysis, Aquaculture 544 (2021).
- C. Poltronieri, R. Laura, D. Bertotto, E. Negrato, C. Simontacchi, M.C. Guerrera, G. Radaelli, Effects of exposure to overcrowding on rodlet cells of the teleost fish Dicentrarchus labrax (L.). Vet Res Commun 2009, 33, 619–629.
- C. M.A. Caipang, M.F. Brinchmann, I. Berg, M. Iversen, R. Eliassen, V. Kiron, Changes in selected stress and immune-related genes in Atlantic cod,Gadus morhua, following overcrowding. Aquaculture Research 2008, 39, 1533–1540. [Google Scholar] [CrossRef]
- M.D. Baldissera, C. M.D. Baldissera, C. de Freitas Souza, A.L. Val, B. Baldisserotto, Involvement of purinergic signaling in the Amazon fish Pterygoplichthys pardalis subjected to handling stress: Relationship with immune response, Aquaculture 514 (2020).
- P. Sun, F. Yin, B. Tang, Effects of Acute Handling Stress on Expression of Growth-Related Genes in Pampus argenteus. J World Aquacult Soc 2016, 48, 166–179. [Google Scholar]
- J. Zhao, Y. Zhu, D. Yang, J. Chen, Y. He, X. Li, X. Feng, B. Xiong, Cage-cultured Largemouth Bronze Gudgeon, Coreius guichenoti: Biochemical Profile of Plasma and Physiological Response to Acute Handling Stress. J World Aquacult Soc 2013, 44, 628–640. [Google Scholar] [CrossRef]
- C. Raposo de Magalhães, D. C. Raposo de Magalhães, D. Schrama, C. Nakharuthai, S. Boonanuntanasarn, D. Revets, S. Planchon, A. Kuehn, M. Cerqueira, R. Carrilho, A.P. Farinha, P.M. Rodrigues, Metabolic Plasticity of Gilthead Seabream Under Different Stressors: Analysis of the Stress Responsive Hepatic Proteome and Gene Expression, Front Mar Sci 8 (2021).
- K. Takahashi, R. Masuda, Net-chasing training improves the behavioral characteristics of hatchery-reared red sea bream (Pagrus major) juveniles. Can J Fish Aquat Sci 2018, 75, 861–867. [Google Scholar] [CrossRef]
- Ziegelbecker, K.M. Sefc, Growth, body condition and contest performance after early-life food restriction in a long-lived tropical fish. Ecol Evol 2021, 11, 10904–10916. [Google Scholar] [CrossRef]
- L. Su, S. Luo, N. Qiu, X. Xiong, J. Wang, The Reproductive Strategy of the Rare Minnow (Gobiocypris rarus) in Response to Starvation Stress. Zool Stud 2020, 59, e1. [Google Scholar]
- M. Varju-Katona, T. Muller, Z. Bokor, K. Balogh, M. Mezes, Effects of various lengths of starvation on body parameters and meat composition in intensively reared pikeperch (Sander lucioperca L.). Iran J Fish Sci 2020, 19, 2062–2076.
- M. Bortoletti, L. M. Bortoletti, L. Maccatrozzo, G. Radaelli, S. Caberlotto, D. Bertotto, Muscle Cortisol Levels, Expression of Glucocorticoid Receptor and Oxidative Stress Markers in the Teleost Fish Argyrosomus regius Exposed to Transport Stress, Animals (Basel) 11(4) (2021).
- M. Vanderzwalmen, J. M. Vanderzwalmen, J. McNeill, D. Delieuvin, S. Senes, D. Sanchez-Lacalle, C. Mullen, I. McLellan, P. Carey, D. Snellgrove, A. Foggo, M.E. Alexander, F.L. Henriquez, K.A. Sloman, Monitoring water quality changes and ornamental fish behaviour during commercial transport, Aquaculture 531 (2021).
- E. E. Grausgruber, M.J. Weber, Effects of Stocking Transport Duration on Age-0 Walleye. J Fish Wildl Manag 2021, 12, 70–82. [Google Scholar] [CrossRef]
Figure 1.
Graphical description of the synthesis and deliver of neurotransmitters during stress in fish.
Figure 1.
Graphical description of the synthesis and deliver of neurotransmitters during stress in fish.
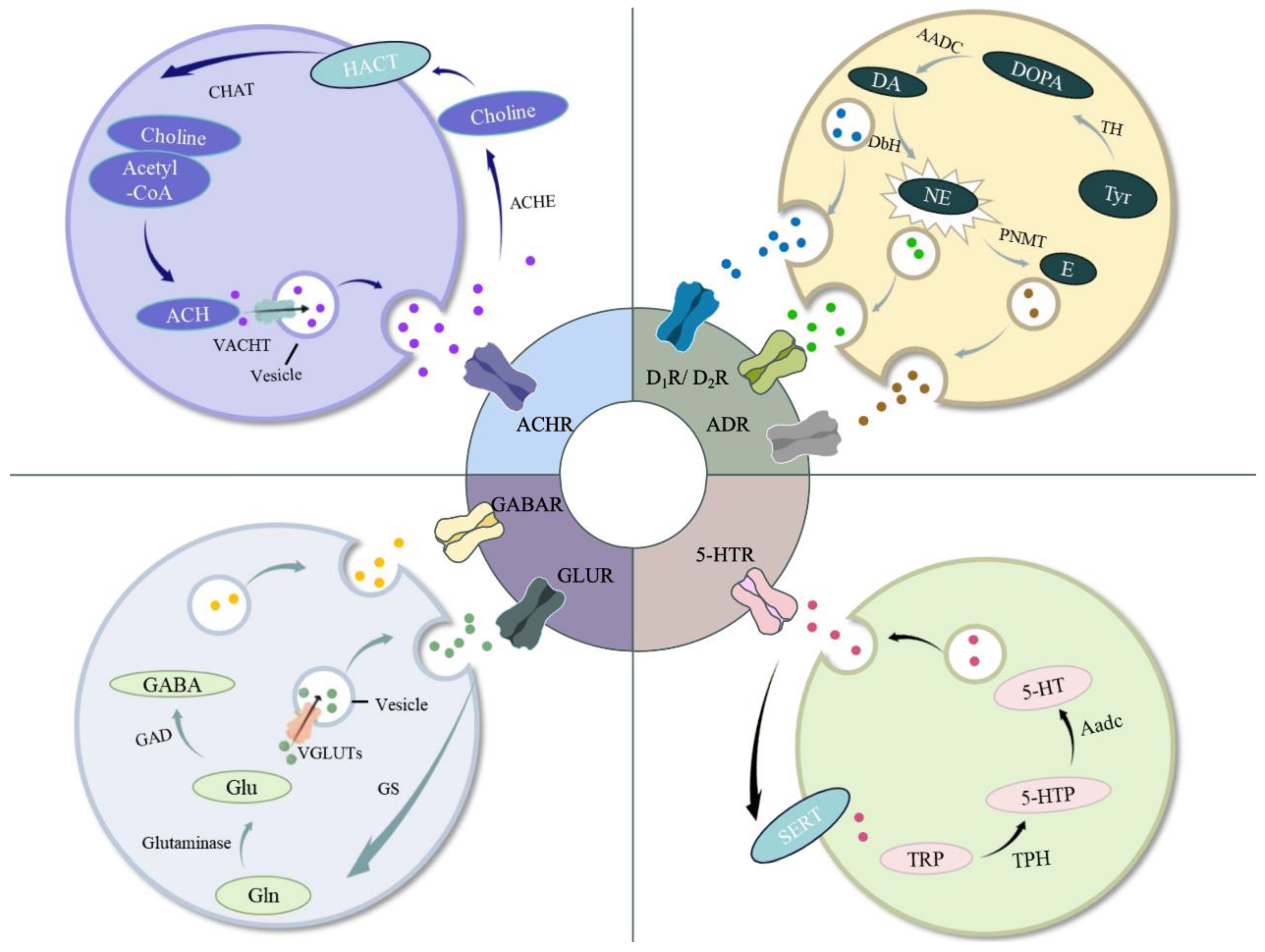
Figure 2.
The schematic diagram shows the neuroendocrine systems in fish. When stress signal is perceived, the neurotransmitter, acetylcholine (ACh), will be released from sympathetic nerve fibers to (i) stimulate the catecholamine (CA) synthesis in the chromaffin cells; (ii) participate in activating the hypothalamus releasing corticotropin-releasing hormone (CRH) and stimulate the pituitary gland releasing adrenocoticotropic hormone (ACTH), which can induce the cortisol secretion in the interrenal cells. (iii) The stress signal will also activate the neurons in midbrain regulating the caudal neurosecretory system (CNSS). CRH, urotensin I (UI) and urotensin II (UII) are synthesized in the Dahlgren cells, stored in the urophysis and released into the circulation from tail vein.
Figure 2.
The schematic diagram shows the neuroendocrine systems in fish. When stress signal is perceived, the neurotransmitter, acetylcholine (ACh), will be released from sympathetic nerve fibers to (i) stimulate the catecholamine (CA) synthesis in the chromaffin cells; (ii) participate in activating the hypothalamus releasing corticotropin-releasing hormone (CRH) and stimulate the pituitary gland releasing adrenocoticotropic hormone (ACTH), which can induce the cortisol secretion in the interrenal cells. (iii) The stress signal will also activate the neurons in midbrain regulating the caudal neurosecretory system (CNSS). CRH, urotensin I (UI) and urotensin II (UII) are synthesized in the Dahlgren cells, stored in the urophysis and released into the circulation from tail vein.
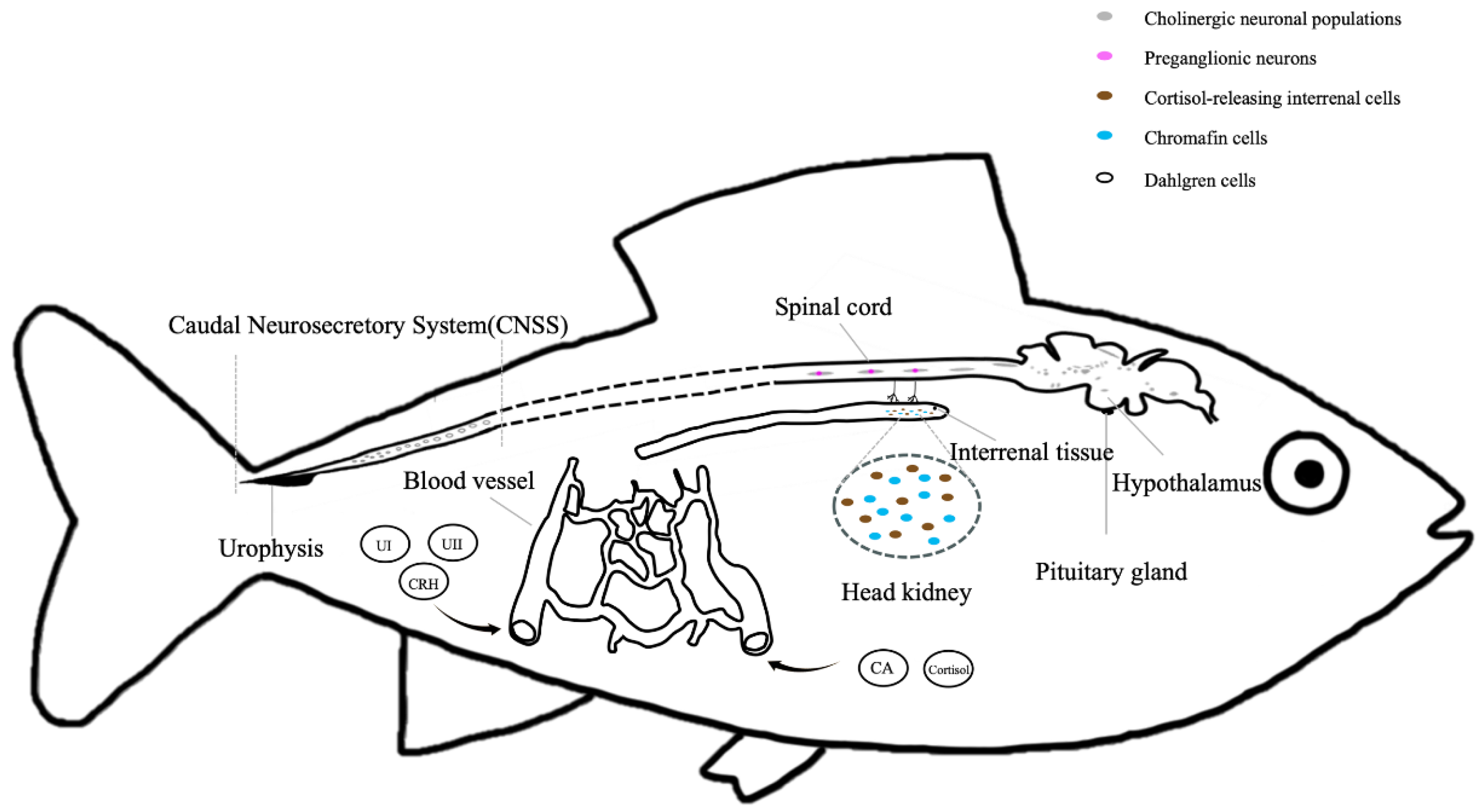

Table 1.
Common stressors in aquaculture.
| Type of stressors | Examples | Reference |
|---|---|---|
| Physical | Water temperature; Photoperiod; Sound; Turbidity … |
[179,180,181] [182,183,184] [185,186,187] [188,189,190] |
| Chemical | pH; Salinity; Dissolved Oxygen; Alkalinity; Carbon dioxide complexities; Nitrite; Ammonia; Hydrogen sulfide; Iron; Heavy metal; Pesticides … |
[191,192,193] [194,195,196,197] [198,199,200] [201,202] [203,204,205] [206,207,208] [209,210,211] [212,213] [214,215] [216,217,218] [219,220,221] |
| Biological | Algal toxicosis; Microorganisms; Parasites; Predators; Social rank; Environmental enrichment … |
[222,223] [224,225,226] [227,228,229] [230,231,232] [233,234,235] [236,237,238] |
| Procedural | Overcrowding; Handling; Netting Feeding; Transportation … |
[239,240,241] [242,243,244] [245,246] [247,248,249] [250,251,252] |
Table 2.
Comparison of neuroendocrine systems in fish.
| System | Primary Components | Response Timing | Magnitude/Duration |
|---|---|---|---|
| Brain-Sympathetic-Chromaffin Cells (BSC) Axis | Brain (sympathetic neurons); Sympathetic nerves; Chromaffin cells (in head kidney; catecholamines) |
Immediate (<1 min to hours) | Rapid onset; Short duration (minutes to hours); High peak magnitude |
| Hypothalamus-Pituitary-Interrenal (HPI) Axis | Hypothalamus (CRH/ACTH-releasing factors); Pituitary (ACTH); Interrenal tissue (cortisol/corticosterone) |
Delayed (hours to days) | Slower onset; Long-lasting (days to weeks); Moderate to high magnitude (sustained) |
| Caudal Neurosecretory System (CNSS) | Dahlgren cell (urotensins, isotocin, CRH); Urophysis; |
Variable (minutes to hours, depending on stimulus) | Moderate onset; Moderate duration (hours); Target-specific magnitude |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



