
Submitted:
25 May 2025
Posted:
26 May 2025
You are already at the latest version
Abstract
Background/Objectives: The insula plays a role in various medical conditions, including eating disorders, addiction, and chronic pain. Repetitive transcranial magnetic stimulation (rTMS) has emerged as a promising therapeutic avenue, yet few studies have investigated its modulation effects on the insula. Moreover, direct evidence of target engagement remains scarce. This study aimed to stimulate the insula with rTMS and assess BOLD signal modulation through concurrent functional magnetic resonance imaging (fMRI). Methods: Ten participants were recruited and six underwent a single session of 5 Hz high-frequency rTMS over the right insular cortex inside the MRI scanner, using a compatible MRI-B91 TMS coil. Stimulation consisted of 10 trains of 10 seconds, with 50-second interval between trains. Frameless stereotactic neuronavigation ensured precise targeting. Paired t-tests were used to compare the mean BOLD signal obtained between stimulation trains with resting-state fMRI acquired before the rTMS stimulation session (significant cluster threshold of 10 voxels; False Discovery Rate at q < 0.01). Results: Increased activity was observed in the anterior, middle, and middle-inferior insula, while deactivations occurred in the ventral anterior and posterior insula. Two participants reported dysgeusia, providing further evidence of insular modulation. Conclusions: This study provides neuroimaging evidence for rTMS-induced insular modulation. Our results are highly relevant for future clinical applications, with potential therapeutic avenues in individuals with conditions where insular dysfunction plays a key role.
Keywords:
Insular cortex
; rTMS
; fMRI
; Neuromodulation
; Neurostimulation
; High-frequency
; Dysgeusia
; BOLD signal
; Neuronavigation
; Concurrent rTMS-fMRI.
1. Introduction
The insula is a deep structure located behind the frontal, parietal, and temporal opercula. Anatomically, it is divided into two portions by the central insular sulcus: the anterior insula (aI) and the posterior insula (pI) [1,2]. Functionally, the insula is divided into three subregions: the mid-posterior insula, and the ventral and dorsal aI. The insula has been implicated in a variety of functions, including autonomic and vestibular functions, interoception, sensory, cognitive, and affective processes [3,4,5,6,7,8]. The mid-posterior insula is connected to primary and secondary somatosensory cortices, whereas the dorsal aI is connected to the dorsal anterior cingulate cortex (dACC) and prefrontal cortex (PFC). The ventral aI shows functional connectivity with the inferior frontal gyrus and the temporal lobe including the amygdala [9].
Research suggests that the insula plays a key role in the regulation of appetite and eating behavior [3,10], as well as interoceptive awareness, including the perception of both hunger and satiety [11]. Functional magnetic resonance imaging (fMRI) studies have shown altered activity in the insula in obese patients [12,13,14]. Studies using electrocortical stimulation in epileptic patients undergoing neurosurgery showed that stimulation of the mid-insula induced gustatory hallucinations, such as acid and metallic tastes [15,16,17]. Furthermore, the insula is involved in the modulation of pain [18,19,20,21,22,23,24,25,26,27]. In individuals with chronic pain conditions, fMRI studies have shown altered activity in the insular cortex [28,29,30,31]. These changes may reflect maladaptive neuroplasticity in which the brain's response to pain is exaggerated or misregulated, contributing to persistent pain [32]. Two studies using intracranial neuronal recordings showed that the earliest response to painful laser stimulation occurs in the pI [33,34]. Low-frequency electrocortical stimulation of the pI elicited pain symptoms [21], whereas subthreshold high-frequency electrocortical stimulation transiently elevated temperature pain thresholds [35,36].
Repetitive transcranial magnetic stimulation (rTMS) is a non-invasive neurostimulation technique that appears to be promising for the treatment of several medical conditions. Technically, rTMS modulates neuronal activity by inducing electrical currents through time-shifting magnetic field pulses [37]. High-frequency rTMS has been widely used in the treatment of depression, mainly targeting the left dorsolateral PFC (DLPFC) [38,39,40,41]. It has also been shown to be safe and potentially effective in the modulation of craving, alcohol and cigarette consumption, and decision-making in addiction when applied to the DLPFC, medial PFC, and dACC [42,43,44,45,46,47,48,49,50,51,52,53]. A recent review reported significant analgesic effects of motor cortex rTMS in neuropathic pain, supported by 14 randomized placebo-controlled trials involving approximately 750 patients [54]. The pI is a promising target, but current evidence remains limited. Finally, a systematic review found that high-frequency rTMS targeting the bilateral DLPFC and the insula was associated with the greatest reduction in body mass index [55].
To date, only a few studies have examined the insula as a therapeutic target, but results are promising. Spagnolo and colleagues [56] reported that low-frequency deep rTMS at an intensity of 120% of the individual’s motor threshold (MT) over the right aI, using an H8 coil, was safe but did not affect performance on behavioral tasks, highlighting that high-frequency stimulation may be required to effectively activate the insula. In 2017, a clinical trial explored the potential of targeting the insula in the treatment of addiction. The study aimed to measure changes in dopamine levels using Positron Emission Tomography (PET) with [11C]-(+)-propyl-hexahydro-naphtho-oxazin. Participants underwent three PET scans after different rTMS sessions (sham, 1 Hz, or 10 Hz). The results showed that low-frequency rTMS targeting the insula significantly decreased dopamine levels in the substantia nigra, sensorimotor striatum, and associative striatum [57]. Dinur-Klein and colleagues [58] showed that high-frequency deep rTMS of the PFC and insula bilaterally reduced cigarette consumption and nicotine dependence, in contrast to low-frequency or sham treatments, and appears to be a promising treatment strategy for addiction. These findings are consistent with Ibrahim and colleagues, who reported the usefulness of combining deep insula rTMS with medication to improve smoking abstinence rates [59]. In addition, deep continuous theta burst stimulation of the right operculo-insular cortex selectively impaired the perception of thermonociceptive input from Aδ-fibre thermonociceptors, without affecting the perception of innocuous warm, cold, or vibrotactile sensations [60]. Another study reported subjective changes in cold perception following rTMS over the posterior-superior insula at a frequency of 10 Hz with an intensity at 80% of MT [61]. In addition, deep posterior-superior insula rTMS was associated with a significant reduction in pain intensity in refractory peripheral neuropathic pain [62]. Finally, a pilot study suggested that deep rTMS over the insula was safe, effective, and well-tolerated in patients with anorexia nervosa [63]. While these study results are promising, a common limitation is the uncertainty regarding whether the stimulation effectively reaches the insula considering its deep location in the Sylvian fissure.
Our study aimed to assess the feasibility of modulating insular activity using high-frequency rTMS and to determine the optimal rTMS parameters required to effectively modulate insular activity. rTMS was applied directly within the MRI scanner, providing a direct measure of neuronal modulation. This simultaneous rTMS-fMRI integration allowed characterization of changes in local cerebral blood flow and oxygenation, which in turn modulated the blood-oxygenation-level-dependent (BOLD) imaging signal [64].
2. Materials and Methods
Participants: Ten healthy participants (five women), all right-handed, were recruited in this study. Inclusion criteria required that participants had no significant health problems, no diagnosis of neurological and/or psychiatric disorders, and no condition that would make MRI and rTMS unsafe (e.g., metallic implants, pacemakers). Prior to rTMS modulation, each participant underwent an individual T1-weighted MRI scan on a separate day to rule out any medical conditions that might affect participation. One participant was excluded due to structural brain abnormalities (SBJ8). Another participant initially agreed to participate in the rTMS but later declined (SBJ7). A technical issue with the coil prevented us from performing stimulation in one participant (SBJ10). We had to interrupt the stimulation in another participant because of severe pain in the right temple caused by the pulses administration from the TMS coil (SBJ5). Thus, a total of six participants received rTMS modulation over the right insular cortex with simultaneous fMRI recording. The study was approved by the CHUM ethics committee (2019-7917, 18.122). All participants gave informed consent prior to all study procedures.
Identification of the target: To target the insula using frameless stereotactic neuronavigation, the previously acquired anatomical MRI was integrated into the Brainsight system, which generates a 3D model of the brain (Brainsight system®, Polaris system, Rogue Research). Four parts of the insula were identified (i.e., the limen insula, the superior border/temporal operculum, the anterior border/frontal operculum, and the posterior border/parietal) (inspired by Ciampi de Andrade and colleagues [61]). Real-time tracking was used to monitor the position of the TMS coil over the participant's head while they lay on the MRI table. At this stage, the MRI table was not yet positioned inside the scanner. The coil was attached to the TMS support, which can be inserted into the MRI scanner during rTMS modulation (see Figure 1). We used three tracking tools: the subject tracker, positioned on the participant's forehead; the coil tracker, attached directly to the TMS coil to guide the coil handle relative to the magnetic field hotspot; and the pointer tool. The TMS coil was positioned on the right side for all participants. However, the exact placement of the coil was individualized for each participant, considering the unique anatomical landmarks of each subject and the limited angulation of the coil in the MRI. Once the coil was positioned over the insula, the subject was moved into the MRI scanner with the TMS coil attached to the support. Table 1 and Table 2 show the individual motor threshold (MT) and the targeted insular subregions.
rTMS procedure: We used the MRI-B91 coil with compressed air cooling (Magventure®, MagProX100, Denmark). This coil generates a magnetic field of 3 to 6 kT/s for deep stimulation (3-5 cm) at 100% stimulator intensity and is compatible with MRI. Individual MT was determined by stimulating the left primary motor cortex and identifying the minimum intensity required to activate the right abductor pollicis brevis muscle in at least 6 out of 10 trials. High-frequency stimulation was performed with 10 trains of 10 seconds each at 5 Hz, with a 50-second interval between trains. Each stimulation train consisted of 50 pulses, for a total of 500 pulses. Stimulation intensity was delivered between 60% and 75% of the individual MT, depending on the participant's tolerance for discomfort during repetitive pulses.
fMRI image acquisition: All participants underwent fMRI acquisition simultaneously with rTMS. Prior to this, a resting-state functional MRI (RS-fMRI) was performed without rTMS. A 3 Tesla whole-body scanner (Skyra, Siemens) with a 16-channel head antenna was used to acquire fMRI data. We used a T2*-weighted gradient echo planar imaging sequence (i.e., slices = 49, TR = 3000 ms, TE = 30 ms, flip angle = 90°; FOV phase = 100%, FOV read = 230 mm, matrix = 64 x 64 matrix, voxel resolution = 2.4 x 2.4 x 3 mm, slice thickness = 3 mm). Participants were instructed to keep their eyes open during the rTMS-fMRI acquisition.
fMRI data processing: The alignment between fMRI slices and rTMS was ensured by synchronizing the onset of rTMS with the fMRI acquisition. We precisely calculated the timing of each stimulation event relative to the fMRI recording. Several parameters were used to determine the onset of rTMS relative to the MRI scans, including the total number of slices, the repetition time (i.e., TR), the echo time (i.e., TE), and the duration of the stimulation trains. This approach allowed us to align rTMS events with specific fMRI slices. We took great care to minimize artifacts from the simultaneous use of TMS and MRI. However, our fMRI data were noisy due to artifacts when TMS pulses occurred. Therefore, we decided to restrict analysis to MRI slices obtained in between stimulations and exclude the MRI slices acquired during stimulations from further analysis. We used Statistical Parametric Mapping software (SPM12) on MATLAB (MATLAB software R2022b) for fMRI data processing. Motion correction was performed using SPM's Realign function, which registered all functional images to a reference image (i.e., the average of all images). The corrected images were visually inspected to confirm the effectiveness of the motion correction. Slice timing correction was then performed. The anatomical MRI was then normalized to the Montreal Neurological Institute (MNI) brain template. Coregistration was performed to align the functional images with the high-resolution anatomical MRI images of each individual subject. Finally, the coregistered functional images were normalized to the MNI standard brain template.
Statistical analysis: Statistical analyses were performed using SPM12 software and xjView toolbox (https://www.alivelearn.net/xjview/). Analyses focused on the fMRI images obtained between rTMS trains. Specifically, paired t-tests were performed to compare the BOLD signal between the average of all fMRI images obtained between stimulation trains to the RS-fMRI images (i.e., RS-fMRI images acquired without the rTMS procedure) using a p-value of 0.01. The interval between stimulation trains was defined as the period from 20 to 50 seconds after the onset of the rTMS trains. This was considered the optimal choice based on increased statistical power. We then used the xjView toolbox to show insular activation and deactivation patterns. The initial voxel-wise threshold was set at p < 0.001 uncorrected. Significant clusters were then identified using the False Discovery Rate (FDR) correction at q < 0.01, with a minimum cluster size threshold of 10 voxels. Due to the insufficient number of fMRI images per rTMS train, it was not possible to make comparisons between different stimulation trains or to visualize the time course of BOLD changes after rTMS. For better understanding, the results are presented on the MNI brain template.
3. Results
One subject's data was excluded from analysis due to significant noise contamination that compromised data quality (SBJ9). For the remaining five participants, we identified between one and two significant activation clusters per subject that survived FDR correction. These activations were characterized by both increases and decreases in BOLD signal, indicating neural responses to stimulation. Specifically, we observed activation in the left (L) middle/aI (SBJ1), L middle insula (SBJ1, SBJ3), L mid-inferior insula (SBJ4), L pI (SBJ6), and right (R) mid-inferior insula (SBJ3). Deactivation was observed in the L ventral aI/aI (SBJ2, SBJ4), and the R and L pI (SBJ4) (see Table 2, Figure 2). In addition, two participants experienced a metallic taste immediately after rTMS. Significant activations were also observed in other brain regions, including the frontal (L and R inferior frontal gyrus, L middle frontal gyrus), temporal (L superior temporal gyrus, L temporal pole), parietal (R supramarginal gyrus, R post-central gyrus), R posterior cingulate, L and R putamen, L parahippocampal gyrus, and L and R globus pallidus regions (see Supplementary Table A1).
Due to the variability in stimulation targets across participants, group analyses were not performed. Heterogeneity in target locations would have introduced significant variability, complicating group-level inferences and potentially masking meaningful patterns of activation.
4. Discussion
In this study, we investigated the feasibility of modulating insular activity using rTMS with concurrent fMRI recording. A single high-frequency 5 Hz rTMS session was administered to six healthy participants over the R insula. Concurrent fMRI revealed significant BOLD signal modulations characterized by increased activity (i.e., activation, positive BOLD signal) in the L middle insula, L and R mid-inferior insula, and L pI subregions. Significant decreased activity (i.e., deactivation, negative BOLD signal) was observed in the L ventral aI, and L and R pI. Two participants reported dysgeusia (i.e., metallic taste). High-frequency stimulation with the MRI-B91 coil over the insular subregions was generally well tolerated despite the limited sample size of this preliminary study.
Our findings provide neuroimaging evidence that rTMS can modulate the insula despite its relatively deep-seated location. Of course, we cannot exclude indirect neural activation and/or deactivation of the insula by rTMS given the dense connectivity of the insula with numerous cerebral regions, including the overlying opercula. For instance, intermittent theta-burst stimulation over frontal regions has been shown to indirectly suppress insula activation by modulating fronto-insular connectivity [65]. Addicott and al. [66] also reported an increase in resting-state functional connectivity between the right postcentral gyrus (PCG) and the left insula following both 1 Hz and 10 Hz rTMS over the right PCG. However, the fact that dysgeusia was reported by two participants suggests that the insula was directly stimulated [67]. The symptom of dysgeusia is consistent with previous electrocortical stimulation studies that have reported gustatory hallucinations, such as metallic taste, following middle insula stimulation [15,16,17]. The two participants who experienced gustatory hallucinations showed an increased BOLD signal in the middle insula (see Table 2). This finding supports the role of this insular subregion in the integration of taste-related sensory inputs [67,68,69,70]. The concordance between increased BOLD activity in the mid-insula and the occurrence of gustatory sensations suggests that the activation threshold of this region may be modulated by rTMS, potentially leading to transient changes in sensory perception, as observed in our study.
In our study, both BOLD signal activations and/or deactivations were observed among our participants. This may possibly be explained by differences in the targeted insular subregions. Furthermore, while low-frequency rTMS is widely recognized for its deactivation/inhibitory effects on neural excitation, the mechanism of action of high-frequency stimulation (i.e., ≥ 5 Hz) remains debated [57]. Indeed, this paradoxical inhibition may result from the activation of inhibitory interneurons, particularly fast-spiking GABAergic circuits, as well as homeostatic plasticity mechanisms that regulate cortical excitability. In addition, stimulation-induced modulation of functional connectivity may lead to downstream inhibitory effects in interconnected neural networks. Furthermore, these activations and/or deactivations in neural responses appear to be influenced by the length and intensity of the stimulation train, with longer and more intense trains potentially enhancing inhibitory effects through sustained interneuron activation and synaptic plasticity [71,72]. In addition, insular response could differ between individuals depending on baseline excitability [73]. Ko and colleagues [74] suggested that the inconsistent findings in the literature of activation or deactivation across studies may be due to the study populations (i.e., healthy vs. clinical), which may reflect differences in neurochemistry, structural integrity, and connectivity. Finally, it may be possible that 5 Hz rTMS falls into a "grey zone" between excitatory and inhibitory effects. Studies suggests that rTMS responses are frequency dependent, and certain frequencies, such as 5 Hz, may produce both excitatory and inhibitory results depending on the context, stimulation parameters, and neural circuits [72,75].
The fact that rTMS over the right insula resulted in the modulation of BOLD activity in both hemispheres is not necessarily surprising considering the connectivity between both insulae and the widespread connectivity of the insula to surrounding lobes [9,76]. In our study, the activated areas are known to be structurally and functionally connected to the insula (see Supplementary Table A1). It is well established that rTMS has widespread effects beyond the target region. Previous neuroimaging studies have reported that rTMS induces cortical activation in both the stimulated and non-stimulated hemispheres [77,78,79]. Negative BOLD responses in the contralateral hemisphere have been observed after both high-frequency [77] and low-frequency rTMS [80]. Future research should explore the neurotransmitter systems underlying these effects. In addition, it would be valuable to investigate the variability of these responses across different rTMS frequencies and intensities, as well as in different populations.
Choosing the optimal coil is critical for effective deep structure stimulation. A study conducted by Lu and Ueno [81] demonstrated that double-cone, H-coil, and HCA coils show significantly deeper field penetration compared to the conventional “figure of eight” (Fo8) coil, albeit at the expense of inducing higher and more widespread magnetic fields in superficial cortical regions. A double-cone and an HCA coil show a superior ability to stimulate deep brain regions compared to an H-coil. At a depth of 40-60 mm, the volume of brain stimulated above threshold by the H-coil is greater than that stimulated by the double-cone coil, suggesting that the stimulation focus of the double-cone coil is superior at this depth. Conversely, at a depth of 60-80 mm, the volume of brain stimulated above threshold by the H-coil decreases rapidly, while that stimulated by the double-cone coil increases. Currently, there is no consensus on the optimal coil for targeting the insular cortex, as some studies have used a double-cone coil (e.g., D-B80 butterfly coil) [61], an H-coil [56,57,58,59,63,82,83], or a Fo8 coil [66,84,85]. The stimulation intensity varied widely across these studies. Spagnolo et al. [56] reported that a superficial cortical intensity of 145% of MT was required to reach a depth of 4 cm below the scalp. However, such high intensity exceeds safety guidelines for insular stimulation [37,86]. Although some studies suggest that higher stimulation thresholds yield better results [57,85], we were unable to apply higher intensities due to the moderate pain induced by pulses at the temple in three participants. We had no choice but to use an MRI-compatible coil for the purpose of these experiments, and thus the intensity of the magnetic field was reduced compared to standard coils due to the thicker casing [87,88].
Several limitations need to be taken into consideration. First, the pulses induced by the coil caused a slight movement of the head during stimulation, making it impossible to obtain clear fMRI images simultaneously with rTMS. Consequently, our results are based on fMRI data acquired immediately post- and between trains of stimulation. High-frequency rTMS has been shown to modulate neuronal activity for up to one hour after stimulation. However, it remains unclear whether this post-stimulation activity mirrors the immediate effects observed during stimulation [89]. Second, we performed only a single high-frequency rTMS session and did not include low-frequency stimulation or a sham condition. Thus, we report the acute effects of rTMS over the insular cortex but cannot infer long-term impacts. Third, the simultaneous rTMS-fMRI recording had inherent challenges, including noise, limited coil positioning on the scalp, restricted movement, and limitations in the choice of coil and rTMS parameters. Even with these limitations, we are confident that our target was effectively stimulated, as evidenced by fMRI images and dysgeusia symptoms specifically associated with the insular cortex.
5. Conclusions
In conclusion, this preliminary work demonstrates that high-frequency rTMS can be applied to the insular cortex without significant adverse effects, and that it is possible to reach the insula with TMS using an MRI-B91 coil guided by frameless stereotactic neuronavigation. Our findings are highly relevant for future clinical applications, where insular modulation could potentially be used for therapeutic avenues such as obesity and chronic pain. Despite the small sample size, this study contributes to the growing body of literature on the effects of neuromodulation with rTMS. Further research needs to validate and extend upon these preliminary observations, including a larger sample size and objective assessments of changes in somatosensory, emotional, and cognitive functions.
Author Contributions
Conceptualization, D.C., O.B. and D.K.N.; methodology, D.C., O.B., L.D.B and D.K.N.; data acquisition, D.C., C.P. and M.R.; formal analysis, D.C., M.R. and K.P.; writing—original draft preparation, D.C., O.B., A.A. and D.K.N.; writing—review and editing, D.C., O.B., L.D.B., K.P. and D.K.N; supervision, O.B. and D.K.N.; funding acquisition, O.B. and D.K.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Quebec Bio-Imaging Network, grant number 51561, the Canada Research Chair Program, CRC-2022-00358, the NSERC, RGPIN-2022-04255, and the CIHR, PJT 148563.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of the Centre de Recherche du Centre Hospitalier de l’Université de Montréal (CER-CRCHUM (18-122; on August 30, 2018).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Dataset available on request from the authors. The data are not publicly available due to ethical reasons.
Acknowledgments
The authors would like to thank all participants for their time and patience during the experiments, as well as the staff of the Neuroimaging platform at the Centre de Recherche du Centre Hospitalier de l’Université de Montréal.
Conflicts of Interest
The authors declare that they have no relevant conflict of interest.
Appendix A
Appendix A.1

Table A1.
Regions showing significant changes in BOLD signal after rTMS trains over the insula.
| Subjects | Side | Anatomic regions | MNI Coordinates | T-score | Cluster size (# of voxel) | ||
| x | y | z | |||||
| SBJ1 | L | Inferior frontal gyrus | -30 | 28 | -7 | 3.84 | 170 |
| L | Middle frontal gyrus | -38 | 40 | -9 | 4.89 | 154 | |
| L | BA 47 | -33 | 31 | -6 | 3.28 | 53 | |
| L | Insula | -34 | 11 | -3 | 3.17 | 23 | |
| L | Putamen | -23 | -3 | -1 | 3.93 | 22 | |
| L | Lateral globus pallidus | -19 | -3 | -6 | 3.52 | 13 | |
| L | Insula | -41 | -1 | 1 | 3.35 | 10 | |
| SBJ2 | L | Insula/BA 13 | -34 | 13 | -9 | 3.27 | 67 |
| L | Temporal pole | -46 | 8 | -8 | 3.32 | 50 | |
| L | Superior temporal gyrus | -43 | 13 | -9 | 3.32 | 45 | |
| SBJ3 | L | Midbrain | -1 | -31 | -15 | 4.44 | 340 |
| L | Insula (middle) | -39 | -1 | -2 | 3.32 | 34 | |
| R | Insula (middle-inferior) | 39 | -12 | -3 | 2.73 | 47 | |
| L | Parahippocampal gyrus | -15 | -40 | -5 | 3.06 | 212 | |
| R | Putamen | 27 | -17 | 1 | 2.72 | 56 | |
| L | Putamen | -28 | -15 | -2 | 2.76 | 57 | |
| R | Globus pallidus | 22 | -17 | 1 | 2.7 | 67 | |
| R | Supramarginal gyrus | 37 | -48 | 29 | 3.31 | 33 | |
| R | Post-central gyrus | 53 | -17 | 28 | 2.92 | 26 | |
| SBJ4 | L | Superior temporal gyrus * | -47 | -11 | -1 | 3.47 | 27 |
| L | Insula (middle-inferior) * | -35 | -4 | -7 | 2.36 | 20 | |
| R | Inferior frontal gyrus | -43 | 20 | -15 | 2.90 | 534 | |
| R | BA 41 | 47 | -36 | 13 | 2.95 | 86 | |
| L | Superior temporal gyrus | -38 | -40 | 5 | 3.02 | 358 | |
| R | Insula (pI) | 39 | -25 | 4 | 2.18 | 131 | |
| L | Insula (pI) | -36 | -18 | 6 | 1.88 | 134 | |
| L | Insula (aI) | -34 | 11 | -14 | 2.56 | 189 | |
| R | Globus pallidus | 25 | -12 | 1 | 1.91 | 81 | |
| SBJ6 | L | Insula (pI) | -43 | -25 | 16 | 3.22 | 54 |
| R | Posterior cingulate | 5 | -51 | 23 | 4.57 | 173 | |
| L | Superior temporal gyrus | -37 | -45 | 15 | 2.81 | 24 | |
Clusters ≥ 10 voxels, FDR-corrected at q < 0.01 / q < 0.001 / q = 0.05 * (trend toward significance). pI: posterior insula; aI: anterior insula; L: left; R: right.
References
- Kurth, F.; Zilles, K.; Fox, P.T.; Laird, A.R.; Eickhoff, S.B. , A link between the systems: functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct Funct 2010, 214, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Ture, U.; Yasargil, D.C.; Al-Mefty, O.; Yasargil, M.G. , Topographic anatomy of the insular region. J Neurosurg 1999, 90, 720–733. [Google Scholar] [CrossRef] [PubMed]
- Avery, J.A.; Kerr, K.L.; Ingeholm, J.E.; Burrows, K.; Bodurka, J.; Simmons, W.K. , A common gustatory and interoceptive representation in the human mid-insula. Hum Brain Mapp 2015, 36, 2996–3006. [Google Scholar] [CrossRef]
- Craig, A. , How do you feel—now? The anterior insula and human awareness. Nature reviews neuroscience 2009, 10, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D. , Interoception: the sense of the physiological condition of the body. Curr Opin Neurobiol 2003, 13, 500–505. [Google Scholar] [CrossRef]
- Craig, A.D. , Emotional moments across time: a possible neural basis for time perception in the anterior insula. Philos Trans R Soc Lond B Biol Sci 2009, 364, 1933–1942. [Google Scholar] [CrossRef]
- Gogolla, N. , The insular cortex. Curr Biol 2017, 27, R580–R586. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Nomi, J.S.; Hebert-Seropian, B.; Ghaziri, J.; Boucher, O. , Structure and Function of the Human Insula. J Clin Neurophysiol 2017, 34, 300–306. [Google Scholar] [CrossRef]
- Ghaziri, J.; Tucholka, A.; Girard, G.; Houde, J.C.; Boucher, O.; Gilbert, G.; Descoteaux, M.; Lippe, S.; Rainville, P.; Nguyen, D.K. , The Corticocortical Structural Connectivity of the Human Insula. Cereb Cortex 2017, 27, 1216–1228. [Google Scholar] [CrossRef]
- Frank, S.; Kullmann, S.; Veit, R. , Food related processes in the insular cortex. Front Hum Neurosci 2013, 7, 499. [Google Scholar] [CrossRef]
- Simmons, W.K.; DeVille, D.C. , Interoceptive contributions to healthy eating and obesity. Curr Opin Psychol 2017, 17, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Lee, S.; Preissl, H.; Schultes, B.; Birbaumer, N.; Veit, R. , The obese brain athlete: self-regulation of the anterior insula in adiposity. PLoS One 2012, 7, e42570. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, I.; Jurado, M.A.; Garolera, M.; Segura, B.; Sala-Llonch, R.; Marques-Iturria, I.; Pueyo, R.; Sender-Palacios, M.J.; Vernet-Vernet, M.; Narberhaus, A.; Ariza, M.; Junque, C. , Alterations of the salience network in obesity: a resting-state fMRI study. Hum Brain Mapp 2013, 34, 2786–2797. [Google Scholar] [CrossRef]
- Makaronidis, J.M.; Batterham, R.L. , Obesity, body weight regulation and the brain: insights from fMRI. Br J Radiol 2018, 91, 20170910. [Google Scholar] [CrossRef]
- Mazzola, L.; Mauguiere, F.; Isnard, J. , Electrical Stimulations of the Human Insula: Their Contribution to the Ictal Semiology of Insular Seizures. J Clin Neurophysiol 2017, 34, 307–314. [Google Scholar] [CrossRef]
- Pugnaghi, M.; Meletti, S.; Castana, L.; Francione, S.; Nobili, L.; Mai, R.; Tassi, L. , Features of somatosensory manifestations induced by intracranial electrical stimulations of the human insula. Clin Neurophysiol 2011, 122, 2049–2058. [Google Scholar] [CrossRef] [PubMed]
- Stephani, C.; Fernandez-Baca Vaca, G.; Maciunas, R.; Koubeissi, M.; Luders, H.O. , Functional neuroanatomy of the insular lobe. Brain Struct Funct 2011, 216, 137–149. [Google Scholar] [CrossRef]
- Geuter, S.; Boll, S.; Eippert, F.; Buchel, C. , Functional dissociation of stimulus intensity encoding and predictive coding of pain in the insula. Elife 2017, 6, e24770. [Google Scholar] [CrossRef]
- Kong, J.; White, N.S.; Kwong, K.K.; Vangel, M.G.; Rosman, I.S.; Gracely, R.H.; Gollub, R.L. , Using fMRI to dissociate sensory encoding from cognitive evaluation of heat pain intensity. Hum Brain Mapp 2006, 27, 715–721. [Google Scholar] [CrossRef]
- Lu, C.; Yang, T.; Zhao, H.; Zhang, M.; Meng, F.; Fu, H.; Xie, Y.; Xu, H. , Insular cortex is critical for the perception, modulation, and chronification of pain. J Neuroscience bulletin 2016, 32, 191–201. [Google Scholar] [CrossRef]
- Mazzola, L.; Faillenot, I.; Barral, F.G.; Mauguiere, F.; Peyron, R. , Spatial segregation of somato-sensory and pain activations in the human operculo-insular cortex. Neuroimage 2012, 60, 409–418. [Google Scholar] [CrossRef]
- Moayedi, M.; Weissman-Fogel, I. , Is the insula the "how much" intensity coder? J Neurophysiol 2009, 102, 1345–1347. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, Y.; Quevedo, A.S.; McHaffie, J.G.; Kraft, R.A.; Coghill, R.C. , Brain mechanisms supporting discrimination of sensory features of pain: a new model. J Neurosci 2009, 29, 14924–14931. [Google Scholar] [CrossRef]
- Roy, M.; Piche, M.; Chen, J.I.; Peretz, I.; Rainville, P. , Cerebral and spinal modulation of pain by emotions. Proc Natl Acad Sci U S A 2009, 106, 20900–20905. [Google Scholar] [CrossRef]
- Schulz, E.; Stankewitz, A.; Witkovsky, V.; Winkler, A.M.; Tracey, I. , Strategy-dependent modulation of cortical pain circuits for the attenuation of pain. Cortex 2019, 113, 255–266. [Google Scholar] [CrossRef]
- Singer, T.; Seymour, B.; O'Doherty, J.; Kaube, H.; Dolan, R.J.; Frith, C.D. , Empathy for pain involves the affective but not sensory components of pain. Science 2004, 303, 1157–1162. [Google Scholar] [CrossRef] [PubMed]
- Starr, C.J.; Sawaki, L.; Wittenberg, G.F.; Burdette, J.H.; Oshiro, Y.; Quevedo, A.S.; Coghill, R.C. , Roles of the insular cortex in the modulation of pain: insights from brain lesions. J Neurosci 2009, 29, 2684–2694. [Google Scholar] [CrossRef]
- Apkarian, A.V.; Bushnell, M.C.; Treede, R.D.; Zubieta, J.K. , Human brain mechanisms of pain perception and regulation in health and disease. Eur J Pain 2005, 9, 463–484. [Google Scholar] [CrossRef] [PubMed]
- Baliki, M.N.; Geha, P.Y.; Jabakhanji, R.; Harden, N.; Schnitzer, T.J.; Apkarian, A.V. , A preliminary fMRI study of analgesic treatment in chronic back pain and knee osteoarthritis. Mol Pain 2008, 4, 47. [Google Scholar] [CrossRef]
- Cauda, F.; Palermo, S.; Costa, T.; Torta, R.; Duca, S.; Vercelli, U.; Geminiani, G.; Torta, D.M. , Gray matter alterations in chronic pain: A network-oriented meta-analytic approach. Neuroimage Clin 2014, 4, 676–686. [Google Scholar] [CrossRef]
- Mandloi, S.; Syed, M.; Shoraka, O.; Ailes, I.; Kang, K.C.; Sathe, A.; Heller, J.; Thalheimer, S.; Mohamed, F.B.; Sharan, A.; Harrop, J.; Krisa, L.; Matias, C.; Alizadeh, M. , The role of the insula in chronic pain following spinal cord injury: A resting-state fMRI study. J Neuroimaging 2023, 33, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Labrakakis, C. , The Role of the Insular Cortex in Pain. Int J Mol Sci 2023, 24, 5736. [Google Scholar] [CrossRef] [PubMed]
- Frot, M.; Magnin, M.; Mauguiere, F.; Garcia-Larrea, L. , Human SII and posterior insula differently encode thermal laser stimuli. Cereb Cortex 2007, 17, 610–620. [Google Scholar] [CrossRef]
- Frot, M.; Mauguiere, F. , Dual representation of pain in the operculo-insular cortex in humans. Brain 2003, 126 Pt 2, 438–450. [Google Scholar] [CrossRef]
- Denis, D.J.; Marouf, R.; Rainville, P.; Bouthillier, A.; Nguyen, D.K. , Effects of insular stimulation on thermal nociception. Eur J Pain 2016, 20, 800–810. [Google Scholar] [CrossRef]
- Bergeron, D.; Obaid, S.; Fournier-Gosselin, M.P.; Bouthillier, A.; Nguyen, D.K. , Deep Brain Stimulation of the Posterior Insula in Chronic Pain: A Theoretical Framework. Brain Sci 2021, 11, 639. [Google Scholar] [CrossRef]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Safety of, T.M.S.C.G. , Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin Neurophysiol 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [PubMed]
- Kaster, T.S.; Daskalakis, Z.J.; Noda, Y.; Knyahnytska, Y.; Downar, J.; Rajji, T.K.; Levkovitz, Y.; Zangen, A.; Butters, M.A.; Mulsant, B.H.; Blumberger, D.M. , Efficacy, tolerability, and cognitive effects of deep transcranial magnetic stimulation for late-life depression: a prospective randomized controlled trial. Neuropsychopharmacology 2018, 43, 2231–2238. [Google Scholar] [CrossRef]
- Levkovitz, Y.; Isserles, M.; Padberg, F.; Lisanby, S.H.; Bystritsky, A.; Xia, G.; Tendler, A.; Daskalakis, Z.J.; Winston, J.L.; Dannon, P.; Hafez, H.M.; Reti, I.M.; Morales, O.G.; Schlaepfer, T.E.; Hollander, E.; Berman, J.A.; Husain, M.M.; Sofer, U.; Stein, A.; Adler, S.; Deutsch, L.; Deutsch, F.; Roth, Y.; George, M.S.; Zangen, A. , Efficacy and safety of deep transcranial magnetic stimulation for major depression: a prospective multicenter randomized controlled trial. World Psychiatry 2015, 14, 64–73. [Google Scholar] [CrossRef]
- Modirrousta, M.; Meek, B.P.; Wikstrom, S.L. , Efficacy of twice-daily vs once-daily sessions of repetitive transcranial magnetic stimulation in the treatment of major depressive disorder: a retrospective study. Neuropsychiatr Dis Treat 2018, 14, 309–316. [Google Scholar] [CrossRef]
- Tavares, D.F.; Myczkowski, M.L.; Alberto, R.L.; Valiengo, L.; Rios, R.M.; Gordon, P.; de Sampaio-Junior, B.; Klein, I.; Mansur, C.G.; Marcolin, M.A.; Lafer, B.; Moreno, R.A.; Gattaz, W.; Daskalakis, Z.J.; Brunoni, A.R. , Treatment of Bipolar Depression with Deep TMS: Results from a Double-Blind, Randomized, Parallel Group, Sham-Controlled Clinical Trial. Neuropsychopharmacology 2017, 42, 2593–2601. [Google Scholar] [CrossRef]
- De Ridder, D.; Vanneste, S.; Kovacs, S.; Sunaert, S.; Dom, G. , Transient alcohol craving suppression by rTMS of dorsal anterior cingulate: an fMRI and LORETA EEG study. Neurosci Lett 2011, 496, 5–10. [Google Scholar] [CrossRef]
- Ekhtiari, H.; Tavakoli, H.; Addolorato, G.; Baeken, C.; Bonci, A.; Campanella, S.; Castelo-Branco, L.; Challet-Bouju, G.; Clark, V.P.; Claus, E.J.N. , Transcranial electrical and magnetic stimulation (tES and TMS) for addiction medicine: a consensus paper on the present state of the science and the road ahead. Neurosci Biobehav Rev 2019, 104, 118–140. [Google Scholar] [CrossRef]
- Hoppner, J.; Broese, T.; Wendler, L.; Berger, C.; Thome, J. , Repetitive transcranial magnetic stimulation (rTMS) for treatment of alcohol dependence. World J Biol Psychiatry 2011, 12 Suppl 1 (sup1), 57–62. [Google Scholar] [CrossRef]
- Makani, R.; Pradhan, B.; Shah, U.; Parikh, T. , Role of Repetitive Transcranial Magnetic Stimulation (rTMS) in Treatment of Addiction and Related Disorders: A Systematic Review. Curr Drug Abuse Rev 2017, 10, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.; Urban, N.; Grassetti, A.; Chang, D.; Hu, M.-C.; Zangen, A.; Levin, F.R.; Foltin, R.; Nunes, E.V.J. , Transcranial magnetic stimulation of medial prefrontal and cingulate cortices reduces cocaine self-administration: a pilot study. Front Psychiatry 2018, 9, 80. [Google Scholar] [CrossRef]
- Mishra, B.R.; Nizamie, S.H.; Das, B.; Praharaj, S.K. , Efficacy of repetitive transcranial magnetic stimulation in alcohol dependence: a sham-controlled study. Addiction 2009, 105, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Politi, E.; Fauci, E.; Santoro, A.; Smeraldi, E. , Daily sessions of transcranial magnetic stimulation to the left prefrontal cortex gradually reduce cocaine craving. Am J Addict 2008, 17, 345–346. [Google Scholar] [CrossRef] [PubMed]
- Rapinesi, C.; Del Casale, A.; Di Pietro, S.; Ferri, V.R.; Piacentino, D.; Sani, G.; Raccah, R.N.; Zangen, A.; Ferracuti, S.; Vento, A.E.; Angeletti, G.; Brugnoli, R.; Kotzalidis, G.D.; Girardi, P. , Add-on high frequency deep transcranial magnetic stimulation (dTMS) to bilateral prefrontal cortex reduces cocaine craving in patients with cocaine use disorder. Neurosci Lett 2016, 629, 43–47. [Google Scholar] [CrossRef]
- Su, H.; Zhong, N.; Gan, H.; Wang, J.; Han, H.; Chen, T.; Li, X.; Ruan, X.; Zhu, Y.; Jiang, H.; Zhao, M. , High frequency repetitive transcranial magnetic stimulation of the left dorsolateral prefrontal cortex for methamphetamine use disorders: A randomised clinical trial. Drug Alcohol Depend 2017, 175, 84–91. [Google Scholar] [CrossRef]
- Terraneo, A.; Leggio, L.; Saladini, M.; Ermani, M.; Bonci, A.; Gallimberti, L. , Transcranial magnetic stimulation of dorsolateral prefrontal cortex reduces cocaine use: A pilot study. Eur Neuropsychopharmacol 2016, 26, 37–44. [Google Scholar] [CrossRef]
- Wing, V.C.; Barr, M.S.; Wass, C.E.; Lipsman, N.; Lozano, A.M.; Daskalakis, Z.J.; George, T.P. , Brain stimulation methods to treat tobacco addiction. Brain Stimul 2013, 6, 221–230. [Google Scholar] [CrossRef]
- Zangen, A.; Moshe, H.; Martinez, D.; Barnea-Ygael, N.; Vapnik, T.; Bystritsky, A.; Duffy, W.; Toder, D.; Casuto, L.; Grosz, M.L. , Repetitive transcranial magnetic stimulation for smoking cessation: a pivotal multicenter double-blind randomized controlled trial. J World Psychiatry 2021, 20, 397–404. [Google Scholar] [CrossRef]
- Garcia-Larrea, L. , Non-invasive cortical stimulation for drug-resistant pain. Curr Opin Support Palliat Care 2023, 17, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.Y.; Zeng, B.S.; Chen, Y.W.; Hung, C.M.; Sun, C.K.; Cheng, Y.S.; Stubbs, B.; Carvalho, A.F.; Brunoni, A.R.; Su, K.P.; Tu, Y.K.; Wu, Y.C.; Chen, T.Y.; Lin, P.Y.; Liang, C.S.; Hsu, C.W.; Tseng, P.T.; Li, C.T. , Efficacy and acceptability of noninvasive brain stimulation interventions for weight reduction in obesity: a pilot network meta-analysis. Int J Obes (Lond) 2021, 45, 1705–1716. [Google Scholar] [CrossRef]
- Spagnolo, P.A.; Wang, H.; Srivanitchapoom, P.; Schwandt, M.; Heilig, M.; Hallett, M. , Lack of Target Engagement Following Low-Frequency Deep Transcranial Magnetic Stimulation of the Anterior Insula. Neuromodulation 2019, 22, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Jacobs, M.; Cho, S.S.; Boileau, I.; Blumberger, D.; Heilig, M.; Wilson, A.; Daskalakis, Z.J.; Strafella, A.P.; Zangen, A.; Le Foll, B. , Deep TMS of the insula using the H-coil modulates dopamine release: a crossover [(11)C] PHNO-PET pilot trial in healthy humans. Brain Imaging Behav 2018, 12, 1306–1317. [Google Scholar] [CrossRef] [PubMed]
- Dinur-Klein, L.; Dannon, P.; Hadar, A.; Rosenberg, O.; Roth, Y.; Kotler, M.; Zangen, A. , Smoking cessation induced by deep repetitive transcranial magnetic stimulation of the prefrontal and insular cortices: a prospective, randomized controlled trial. Biol Psychiatry 2014, 76, 742–749. [Google Scholar] [CrossRef]
- Ibrahim, C.; Tang, V.M.; Blumberger, D.M.; Malik, S.; Tyndale, R.F.; Trevizol, A.P.; Barr, M.S.; Daskalakis, Z.J.; Zangen, A.; Le Foll, B. , Efficacy of insula deep repetitive transcranial magnetic stimulation combined with varenicline for smoking cessation: A randomized, double-blind, sham controlled trial. Brain Stimul 2023, 16, 1501–1509. [Google Scholar] [CrossRef]
- Lenoir, C.; Algoet, M.; Mouraux, A. , Deep continuous theta burst stimulation of the operculo-insular cortex selectively affects Adelta-fibre heat pain. J Physiol 2018, 596, 4767–4787. [Google Scholar] [CrossRef]
- Ciampi de Andrade, D.; Galhardoni, R.; Pinto, L.F.; Lancelotti, R.; Rosi, J., Jr.; Marcolin, M.A.; Teixeira, M.J. , Into the island: a new technique of non-invasive cortical stimulation of the insula. Neurophysiol Clin 2012, 42, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Dongyang, L.; Fernandes, A.M.; da Cunha, P.H.M.; Tibes, R.; Sato, J.; Listik, C.; Dale, C.; Kubota, G.T.; Galhardoni, R.; Teixeira, M.J.; Aparecida da Silva, V.; Rosi, J.; Ciampi de Andrade, D. , Posterior-superior insular deep transcranial magnetic stimulation alleviates peripheral neuropathic pain - A pilot double-blind, randomized cross-over study. Neurophysiol Clin 2021, 51, 291–302. [Google Scholar] [CrossRef]
- Knyahnytska, Y.O.; Blumberger, D.M.; Daskalakis, Z.J.; Zomorrodi, R.; Kaplan, A.S. , Insula H-coil deep transcranial magnetic stimulation in severe and enduring anorexia nervosa (SE-AN): a pilot study. Neuropsychiatr Dis Treat 2019, 15, 2247–2256. [Google Scholar] [CrossRef]
- Bestmann, S.; Ruff, C.C.; Blankenburg, F.; Weiskopf, N.; Driver, J.; Rothwell, J.C. , Mapping causal interregional influences with concurrent TMS-fMRI. Exp Brain Res 2008, 191, 383–402. [Google Scholar] [CrossRef]
- Iwabuchi, S.J.; Raschke, F.; Auer, D.P.; Liddle, P.F.; Lankappa, S.T.; Palaniyappan, L. , Targeted transcranial theta-burst stimulation alters fronto-insular network and prefrontal GABA. Neuroimage 2017, 146, 395–403. [Google Scholar] [CrossRef]
- Addicott, M.A.; Luber, B.; Nguyen, D.; Palmer, H.; Lisanby, S.H.; Appelbaum, L.G. , Low-and high-frequency repetitive transcranial magnetic stimulation effects on resting-state functional connectivity between the postcentral gyrus and the insula. J Brain connectivity 2019, 9, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Small, D.M. , Taste representation in the human insula. Brain Struct Funct 2010, 214, (5–6). [Google Scholar] [CrossRef] [PubMed]
- Avery, J.A.; Gotts, S.J.; Kerr, K.L.; Burrows, K.; Ingeholm, J.E.; Bodurka, J.; Martin, A.; Kyle Simmons, W. , Convergent gustatory and viscerosensory processing in the human dorsal mid-insula. Hum Brain Mapp 2017, 38, 2150–2164. [Google Scholar] [CrossRef]
- Rolls, E.T. , Functions of the anterior insula in taste, autonomic, and related functions. Brain Cogn 2016, 110, 4–19. [Google Scholar] [CrossRef]
- Small, D.M.; Gregory, M.D.; Mak, Y.E.; Gitelman, D.; Mesulam, M.M.; Parrish, T. , Dissociation of neural representation of intensity and affective valuation in human gustation. Neuron 2003, 39, 701–711. [Google Scholar] [CrossRef]
- de Jesus, D.R.; de Souza Favalli, G.P.; Hoppenbrouwers, S.S.; Barr, M.S.; Chen, R.; Fitzgerald, P.B.; Daskalakis, Z.J. , Determining optimal rTMS parameters through changes in cortical inhibition. J Clinical Neurophysiology 2014, 125, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, P.B.; Fountain, S.; Daskalakis, Z.J. , A comprehensive review of the effects of rTMS on motor cortical excitability and inhibition. Clin Neurophysiol 2006, 117, 2584–2596. [Google Scholar] [CrossRef]
- Maeda, F.; Keenan, J.P.; Tormos, J.M.; Topka, H.; Pascual-Leone, A. , Interindividual variability of the modulatory effects of repetitive transcranial magnetic stimulation on cortical excitability. Exp Brain Res 2000, 133, 425–430. [Google Scholar] [CrossRef]
- Ko, J.H.; Tang, C.C.; Eidelberg, D. , Brain stimulation and functional imaging with fMRI and PET. Handb Clin Neurol 2013, 116, 77–95. [Google Scholar] [PubMed]
- Pascual-Leone, A.; Valls-Sole, J.; Wassermann, E.M.; Hallett, M. , Responses to rapid-rate transcranial magnetic stimulation of the human motor cortex. Brain 1994, 117 Pt 4, 847–858. [Google Scholar] [CrossRef]
- Ghaziri, J.; Fei, P.; Tucholka, A.; Obaid, S.; Boucher, O.; Rouleau, I.; Nguyen, D.K. , Resting-State Functional Connectivity Profile of Insular Subregions. Brain Sci 2024, 14, 742. [Google Scholar] [CrossRef]
- Shimomura, T.; Fujiki, M.; Ohba, H.; Kochiyma, T.; Sugita, K.; Matsuta, H.; Kawasaki, Y.; Oonishi, K.; Fudaba, H.; Kamida, T.J.J.N.N., Contralateral negative bold responses in the motor network during subthreshold high-frequency interleaved TMS-fMRI over the human primary motor cortex. 2018, 9, 1–8.
- Tian, D.; Izumi, S.I. , Interhemispheric Facilitatory Effect of High-Frequency rTMS: Perspective from Intracortical Facilitation and Inhibition. Brain Sci 2022, 12, 970. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Hanajima, R.; Shirota, Y.; Ohminami, S.; Tsutsumi, R.; Terao, Y.; Ugawa, Y.; Hirose, S.; Miyashita, Y.; Konishi, S.; Kunimatsu, A.; Ohtomo, K. , Bidirectional effects on interhemispheric resting-state functional connectivity induced by excitatory and inhibitory repetitive transcranial magnetic stimulation. Hum Brain Mapp 2014, 35, 1896–1905. [Google Scholar] [CrossRef]
- Plow, E.B.; Cattaneo, Z.; Carlson, T.A.; Alvarez, G.A.; Pascual-Leone, A.; Battelli, L. , The compensatory dynamic of inter-hemispheric interactions in visuospatial attention revealed using rTMS and fMRI. Front Hum Neurosci 2014, 8, 226. [Google Scholar] [CrossRef]
- Lu, M.; Ueno, S. , Comparison of the induced fields using different coil configurations during deep transcranial magnetic stimulation. PLoS One 2017, 12, e0178422. [Google Scholar] [CrossRef]
- Lee, M.R.; Caparelli, E.C.; Leff, M.; Steele, V.R.; Maxwell, A.M.; McCullough, K.; Salmeron, B.J.J.N.T. a. t. N. I., Repetitive transcranial magnetic stimulation delivered with an H-coil to the right insula reduces functional connectivity between insula and medial prefrontal cortex. 2020, 23, 384–392.
- Perini, I.; Kampe, R.; Arlestig, T.; Karlsson, H.; Lofberg, A.; Pietrzak, M.; Zangen, A.; Heilig, M. , Repetitive transcranial magnetic stimulation targeting the insular cortex for reduction of heavy drinking in treatment-seeking alcohol-dependent subjects: a randomized controlled trial. Neuropsychopharmacology 2020, 45, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Hodkinson, D.J.; Bungert, A.; Bowtell, R.; Jackson, S.R.; Jung, J. , Operculo-insular and anterior cingulate plasticity induced by transcranial magnetic stimulation in the human motor cortex: a dynamic casual modeling study. J Neurophysiol 2021, 125, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Moeller, S.J.; Gil, R.; Weinstein, J.J.; Baumvoll, T.; Wengler, K.; Fallon, N.; Van Snellenberg, J.X.; Abeykoon, S.; Perlman, G.; Williams, J.; Manu, L.; Slifstein, M.; Cassidy, C.M.; Martinez, D.M.; Abi-Dargham, A., Deep rTMS of the insula and prefrontal cortex in smokers with schizophrenia: Proof-of-concept study. Schizophrenia (Heidelb) 2022, 8, 6.
- Rossi, S.; Antal, A.; Bestmann, S.; Bikson, M.; Brewer, C.; Brockmoller, J.; Carpenter, L.L.; Cincotta, M.; Chen, R.; Daskalakis, J.D.; Di Lazzaro, V.; Fox, M.D.; George, M.S.; Gilbert, D.; Kimiskidis, V.K.; Koch, G.; Ilmoniemi, R.J.; Lefaucheur, J.P.; Leocani, L.; Lisanby, S.H.; Miniussi, C.; Padberg, F.; Pascual-Leone, A.; Paulus, W.; Peterchev, A.V.; Quartarone, A.; Rotenberg, A.; Rothwell, J.; Rossini, P.M.; Santarnecchi, E.; Shafi, M.M.; Siebner, H.R.; Ugawa, Y.; Wassermann, E.M.; Zangen, A.; Ziemann, U.; Hallett, M.; basis of this article began with a Consensus Statement from the Ifcn Workshop on "Present, F. o. T. M. S. S. E. G. S. O. u. t. A., Safety and recommendations for TMS use in healthy subjects and patient populations, with updates on training, ethical and regulatory issues: Expert Guidelines. Clin Neurophysiol 2021, 132, 269–306.
- Koponen, L.M.; Nieminen, J.O.; Ilmoniemi, R.J. , Multi-locus transcranial magnetic stimulation-theory and implementation. Brain Stimul 2018, 11, 849–855. [Google Scholar] [CrossRef]
- Nieminen, J.O.; Koponen, L.M.; Ilmoniemi, R.J. , Experimental Characterization of the Electric Field Distribution Induced by TMS Devices. Brain Stimul 2015, 8, 582–589. [Google Scholar] [CrossRef]
- Siebner, H.R.; Bergmann, T.O.; Bestmann, S.; Massimini, M.; Johansen-Berg, H.; Mochizuki, H.; Bohning, D.E.; Boorman, E.D.; Groppa, S.; Miniussi, C.; Pascual-Leone, A.; Huber, R.; Taylor, P.C.; Ilmoniemi, R.J.; De Gennaro, L.; Strafella, A.P.; Kahkonen, S.; Kloppel, S.; Frisoni, G.B.; George, M.S.; Hallett, M.; Brandt, S.A.; Rushworth, M.F.; Ziemann, U.; Rothwell, J.C.; Ward, N.; Cohen, L.G.; Baudewig, J.; Paus, T.; Ugawa, Y.; Rossini, P.M., Consensus paper: combining transcranial stimulation with neuroimaging. Brain Stimul 2009, 2, 58–80.
Figure 1.
The MRI-B91 coil is attached to an MRI-compatible TMS support while the subject lies on the MRI table, with the coil positioned on the scalp (A). A 16-channel head antenna is placed over the participant's head to optimize signal reception during the fMRI acquisition (B and C).
Figure 1.
The MRI-B91 coil is attached to an MRI-compatible TMS support while the subject lies on the MRI table, with the coil positioned on the scalp (A). A 16-channel head antenna is placed over the participant's head to optimize signal reception during the fMRI acquisition (B and C).
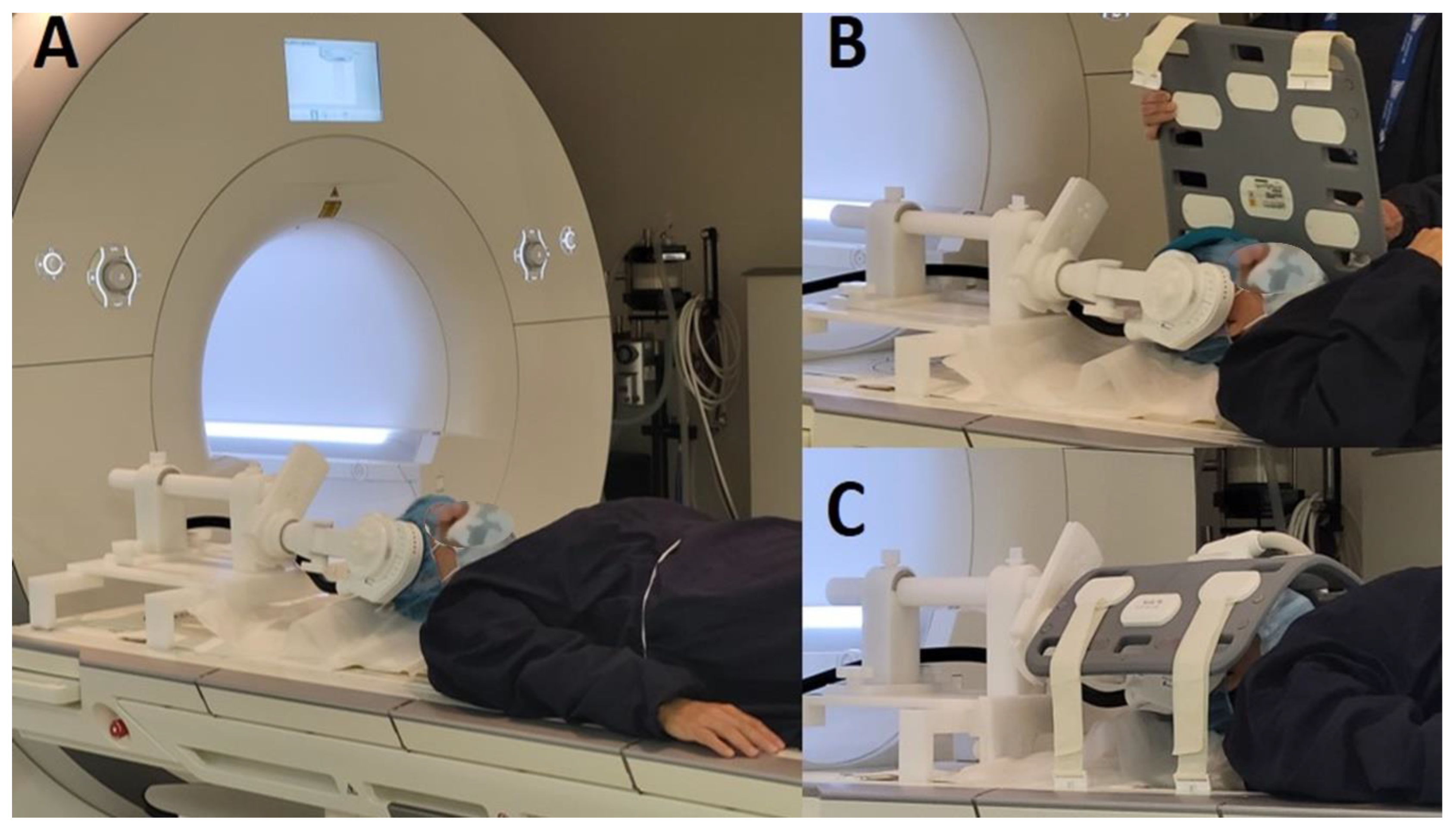
Figure 2.
Change in BOLD activity during post-trains intervals compared to the resting-state fMRI. Results are displayed at q < 0.01 or q < 0.001, FDR-corrected. The left side represents BOLD signal activation/excitation (SBJ1, SBJ3, and SBJ4) and the right side represents BOLD signal deactivation/inhibition (SBJ2, SBJ4, and SBJ6). From top to bottom, a sagittal view (right and left) followed by a coronal view.
Figure 2.
Change in BOLD activity during post-trains intervals compared to the resting-state fMRI. Results are displayed at q < 0.01 or q < 0.001, FDR-corrected. The left side represents BOLD signal activation/excitation (SBJ1, SBJ3, and SBJ4) and the right side represents BOLD signal deactivation/inhibition (SBJ2, SBJ4, and SBJ6). From top to bottom, a sagittal view (right and left) followed by a coronal view.
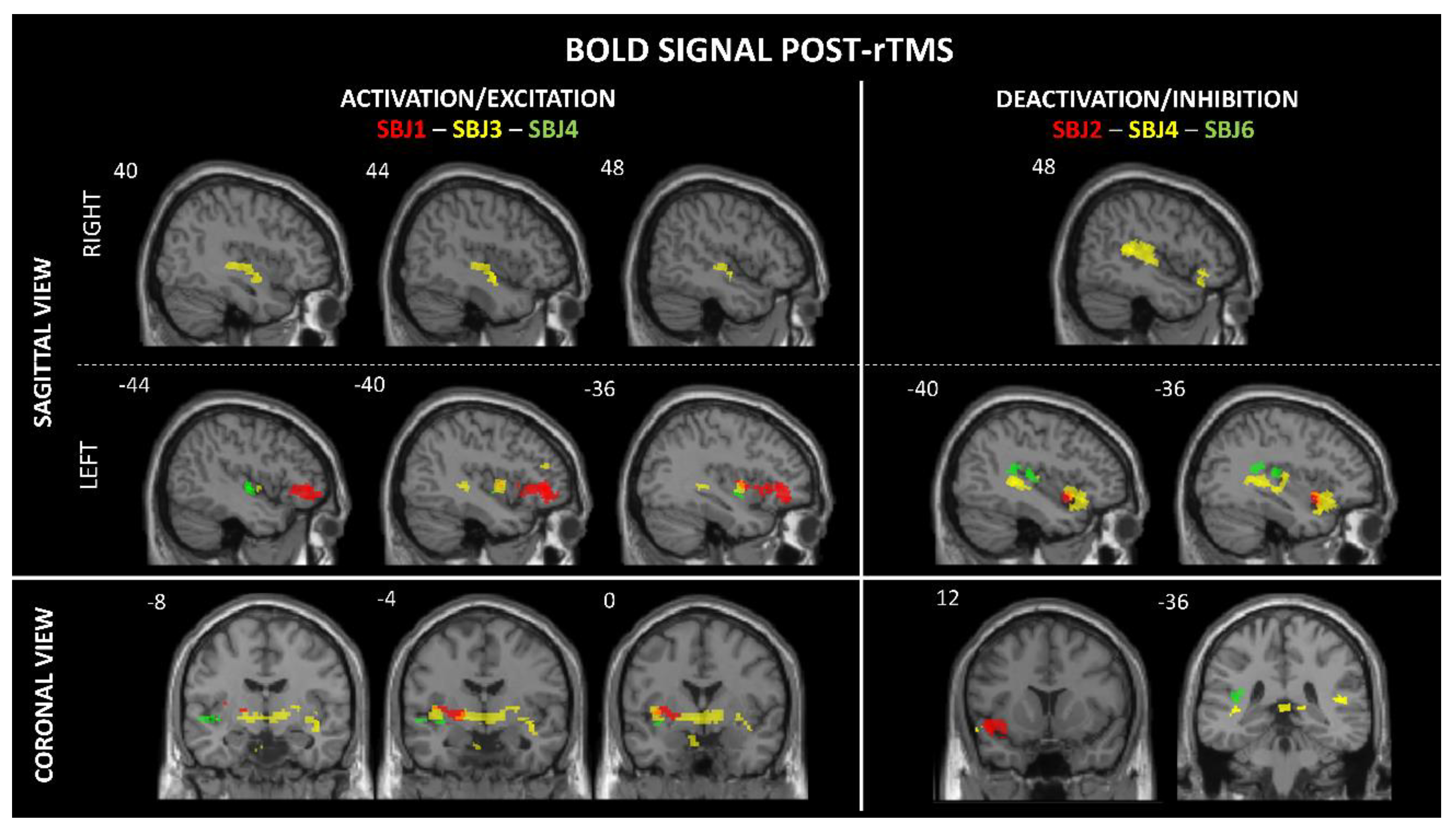
Table 1.
Descriptive characteristics of participants, motor threshold parameters, and excluded subjects.
Table 1.
Descriptive characteristics of participants, motor threshold parameters, and excluded subjects.
| Subjects | Sex | Motor threshold parameters | Reason of exclusion | ||
| Stimulation threshold | Stimulation intensity | Stimulation amplitude | |||
| SBJ1 | F | 69 | 75% | 52 | - |
| SBJ2 | F | 69 | 75% | 52 | - |
| SBJ3 | M | 77 | 60% | 50 | - |
| SBJ4 | F | 63 | 70% | 45 | - |
| SBJ5 | M | 60 | 60% | 36 | Stimulation interrupted: Pain at right temple |
| SBJ6 | M | 55 | 70% | 39 | - |
| SBJ7 | F | - | - | - | Participation declined |
| SBJ8 | M | - | - | - | Exclusion: Contraindication for rTMS-MRI |
| SBJ9 | F | 69 | 75% | 52 | - |
| SBJ10 | M | - | - | - | Technical issue: rTMS coil malfunction |
F = female; M = male. Participants included (n=6) and excluded (n=4).
Table 2.
BOLD responses in the insular cortex.
| Brainsight parameters | MNI coordinates | BOLD signal activity |
Side | T-Value | Cluster size | FDR- correction |
Sensory responses induced by rTMS |
||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Targeted insular subregions | Significant BOLD changes in the insular subregions |
x | y | z | |||||||
| SBJ1 | mid-pI | middle/aI | -34 | 11 | -3 | A | L | 3.17 | 23 | < 0.001 | Metallic taste |
| middle | -41 | -1 | 1 | A | L | 3.35 | 10 | < 0.001 | |||
| SBJ2 | ventral aI | ventral aI | -34 | 12 | -15 | D | L | 3.27 | 67 | < 0.001 | - |
| SBJ3 | aI | middle | -39 | -1 | -2 | A | L | 3.32 | 34 | < 0.001 | - |
| mid-inferior | 39 | -12 | -3 | A | R | 2.73 | 47 | < 0.001 | |||
| SBJ4 | Inferior pI | mid-inferior | -35 | -4 | -7 | A | L | 2.36 | 20 | 0.05 * | Metallic taste |
| pI | 39 | -25 | 4 | D | R | 2.18 | 131 | < 0.01 | |||
| pI | -36 | -18 | 6 | D | L | 1.88 | 134 | < 0.01 | |||
| aI | -34 | 11 | -14 | D | L | 2.56 | 189 | < 0.01 | |||
| SBJ6 | Superior pI | pI | -43 | -25 | 16 | A | L | 3.22 | 54 | < 0.001 | - |
| SBJ9 | Inferior pI | Excluded from analysis due to excessive noise contamination | |||||||||
Clusters ≥ 10 voxels, FDR-corrected at q < 0.01 / q < 0.001 / q = 0.05 * (trend toward significance). pI: posterior insula; aI: anterior insula; L: left; R: right; A: activation; D: deactivation. Participant # SBJ9 excluded from analysis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



