
Submitted:
13 May 2025
Posted:
15 May 2025
You are already at the latest version
Abstract
Dimethyl trisulfide (DMTS) is a naturally occurring organic polysulfide associated with protective functions such as antioxidant and neuroprotective effects. DMTS is a lipophilic transient receptor potential ankyrin 1 (TRPA1) ligand that reaches the central nervous system (CNS). Its role in the CNS, particularly regarding depression‐like behaviour, has yet to be explored. This study investigates the influence of DMTS on stress responses and whether this effect is mediated through the TRPA1 ion channel, known for its role in stress adaptation. Using a mouse model exposed to chronic unpredictable mild stress (CUMS), we examined the impact of DMTS on depression‐like behaviour and anxiety, and identified the involved brain regions. Our methods involved testing both Trpa1 wild‐type and gene‐knockout mice under CUMS conditions and DMTS treatment. Various behavioural assessments—including the open field, marble burying, tail suspension, forced swim, and sucrose preference tests—were performed to evaluate anxiety and depression‐like behaviour. Additionally, we measured body weight changes and the relative weights of the thymus and adrenal glands, while serum levels of corticosterone and adrenocorticotropic hormone were quantified via ELISA. FOSB immunohistochemistry was utilized to assess chronic neuronal activation in stress‐ relevant brain areas. Results showed that CUMS induces depression‐like behaviour in a TRPA1‐ dependent manner and that DMTS treatment significantly reduced these effects when TRPA1 channels were functional. DMTS also mitigated thymus involution due to hypothalamic‐pituitary‐ adrenal (HPA) axis dysregulation. Overall, DMTS appears to relieve depressive and anxiety symptoms through TRPA1‐mediated pathways, suggesting its potential as a dietary supplement or adjunct therapy for depression and anxiety.
Keywords:
dimethyl trisulfide
; mechanism
; TRPA1
; ion channels
; depression
; anxiety
; chronic stress
1. Introduction
The expression of the transient receptor potential ankyrin 1 (TRPA1) ion channel is well known in peripheral nociceptive nerves, but there is increasing evidence of its central nervous system (CNS) presence [1]. Trpa1 mRNA expression was confirmed in certain stress-related limbic brain areas, including the olfactory bulb, piriform cortex [20], and hypothalamus [21]; however, the highest level of Trpa1 mRNA was detected in the urocortinergic cells of the centrally projecting Edinger-Westphal nucleus (EWcp), both in mice and human [2]. In addition, the functional activity of TRPA1 was demonstrated using a specific and potent agonist (JT010) by patch clamp electrophysiology in acute EWcp slices [3]. Researchers from our institution confirmed that Trpa1 was downregulated in the EWcp in chronic unpredictable mild stress (CUMS) model in mice and human suicide victims [2]. Moreover, altered stress adaptation ability was observed in Trpa1 gene-deficient mice in a single prolonged stress model of posttraumatic stress disorder (PTSD) [4]. This suggests that TRPA1 in EWcp neurons might contribute to regulating depression-like behaviour and stress adaptation response in mice [2].
In addition to electrophilic and non-electrophilic activators, naturally occurring compounds can also be included as activators of TRPA1 ion channels. These include wasabi, cinnamaldehyde, eugenol, phenylpropenes of cloves, piperine in black pepper, curcumin or gingerol. It is fascinating the activation of TRPA1 by compounds produced by Brassica (mustard) and Allium (onions and garlic) plants, such as pungent isothiocyanates and allicin, respectively [5].
Among these agonists, lipophilic dimethyl trisulphide (DMTS) has been proven to reach the brain. DMTS is a sulfur-based molecule found in garlic, onion, broccoli, and similar plants. DMTS is readily available in high purity, is naturally occurring, stable, and can be reasonably priced. DMTS has an elimination half-life of 36 minutes and accumulates mainly in red blood cells [6]. It readily crosses the blood-brain barrier and can be detected in brain tissue, including stress-relevant brain areas, such as EWcp [7,8].
Literature suggests that sulfide attenuates depression-like behaviour induced by CUMS [9,10]. Some suggest that the effects of sulfides are mediated by their oxidized forms, called polysulfides [11,12,13], which have been detected in the CNS of rodents. DMTS is an organic polysulfide, the main differences between organic and inorganic polysulfides are reflected in half-life, purity and availability. Organic polysulfides have more favourable pharmacokinetic properties. Although the role of sulfide in the CNS is being actively investigated, little is known about its effects on anxiety and depression [14]. In our previous experiments, DMTS inhibited spontaneous motor activity and respiration in mice [15]. Given that CNS inhibitors/depressants (e.g. benzodiazepines and barbiturates) tend to reduce levels of anxiety, we hypothesized that an appropriately chosen dose of DMTS may have an anxiolytic and antidepressant effect without interfering with movement and breathing. Our previous findings suggest that one of the main targets of DMTS effects is the TRPA1 cation channel [16]. The TRPA1 ion channel was activated by sodium polysulfide in the CNS astrocytes [17]. Our results imply that TRPA1 expressed on Chinese hamster ovary (CHO) cells is activated by DMTS and sodium polysulfide (POLY) [15,18]. Inhibition of motor activity of mice by DMTS was mediated by the TRPA1 channel [15].
It raises the importance of research on the topic that the detailed pathomechanism of these disorders remains unclear due to the lack of a universally accepted animal model in basic research, and that the effective treatment of anxiety and depression is also unmet with current treatment strategies [19].
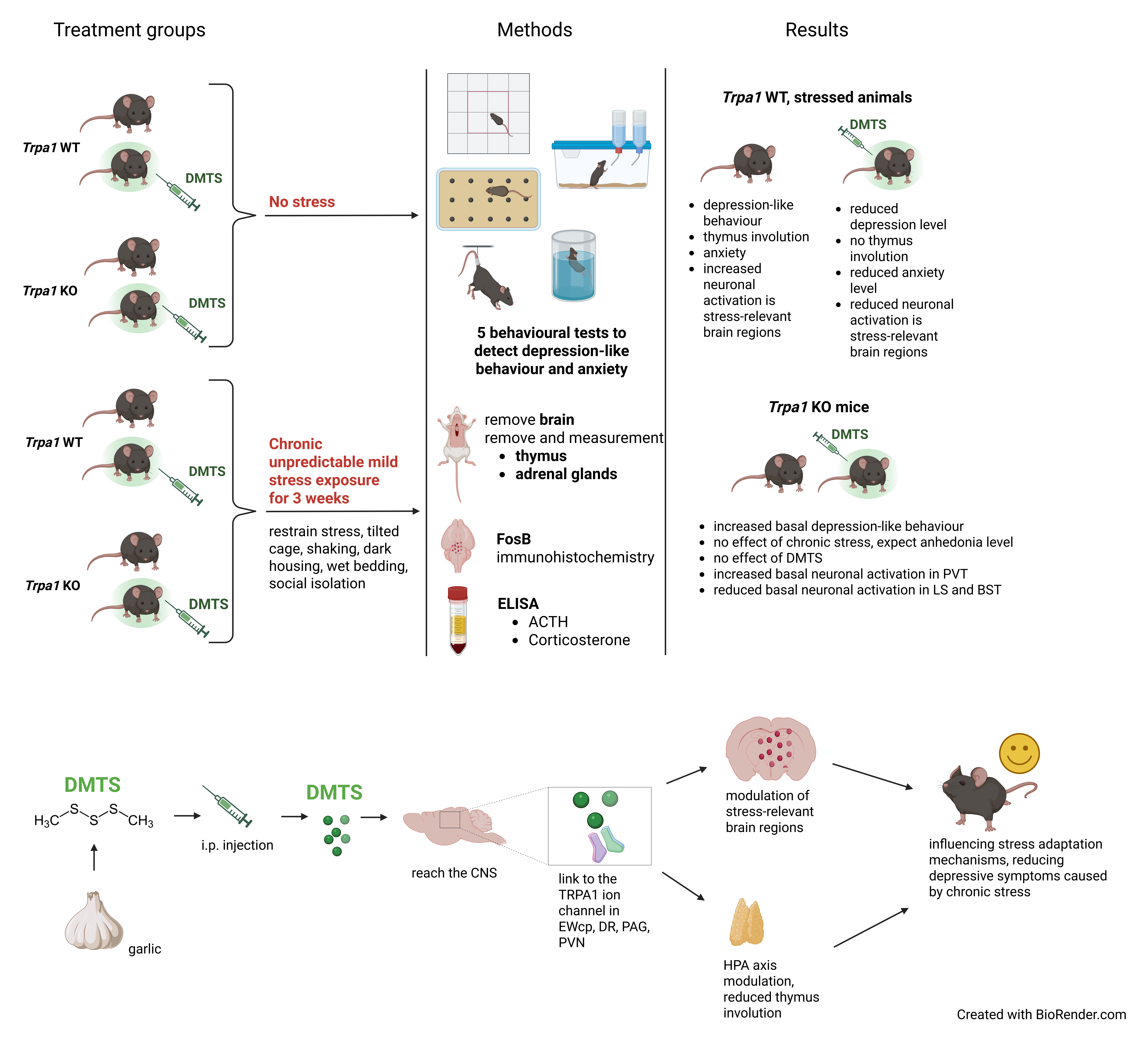
In the present study, we aimed to investigate whether the TRPA1 agonist DMTS modulates stress-related brain processes, such as anxiety or depression. We tested the impact of DMTS on a mouse model of depression with special focus on the involvement of the TRPA1 channel. We carried out behavioural studies using Trpa1 wild-type (WT) and knockout (KO) mice in CUMS model to detect anxiety- and depression-like behaviour. We tested physical and endocrine parameters to assess the stress adaptation responses. FOSB immunohistochemical analysis was utilized as a marker of chronic neuronal activation and enzyme-linked immunosorbent assay (ELISA) was performed to measure the plasma corticosterone (CORT) and adrenocorticotropic hormone (ACTH) levels.
2. Materials and Methods
2.1. Animals
Experiments were conducted on genetically modified male mice (adult, 25-30 g) lacking functional Trpa1 and their WT counterparts. Age-matched animals were used. The animals were housed in a temperature- and humidity-controlled environment with a 12-hour light-dark cycle (lights were turned on at 6:00 a.m.). Mice were housed in standard polycarbonate cages (365 mm × 207 mm × 144 mm) in groups of 4-6 mice per cage at the animal facility of the Department of Pharmacology and Pharmacotherapy, University of Pécs. They were provided standard rodent food and tap water ad libitum, ensuring their well-being. All experimental procedures were carried out according to the European Communities Council Directive of 2010/63/EU. The studies were approved by the Animal Welfare Committee of the University of Pécs and the National Scientific Ethics Committee for Animal Experiments in Hungary (permission number: BA/73/0476-9/2022). As previously described, Trpa1 KO mice were bred on a C57BI/6 background [18].
2.2. Chemicals
DMTS (Sigma-Aldrich, Budapest, Hungary) solutions were prepared in physiological saline containing polysorbate 80 (Sigma-Aldrich, Budapest, Hungary). A solution of 3% m/v was prepared by dissolving polysorbate 80 in physiological saline. The DMTS stock solution was obtained by dissolving DMTS at 10 mg/ml. Solutions were diluted further with physiological saline to obtain working solutions. Solutions were administered intraperitoneally (i.p.) to the animals (10 ml/kg). The vehicle used in the experiments contained 1.5% m/v polysorbate 80.
2.3. Experimental Design
2.3.1. Dose Determination and Treatment Groups
The appropriate intraperitoneal dose of DMTS was determined using OFT (see chapter 2.5.1.). The test was used to monitor the general locomotor activity of the Trpa1 WT animals. Evaluation of behavioural tests for chronic stress can be compromised by altered motor activity. The distance travelled by the mice and the time spent moving were evaluated. For dose testing, animals were divided into 4 groups. Doses of 30 mg/kg and 40 mg/kg of DMTS and respective doses of vehicle were tested. In addition, there were naïve and vehicle treated groups with 8 animals in each group.
After dose determination, in the second experiment, we divided the animals into 12 treatment groups. Trpa1 KO mice were utilized to explore the mechanism of action and to validate the role of the TRPA1 receptor. We used stressed and non-stressed groups within the two mouse genotypes. Naïve, vehicle-treated, and DMTS-treated subgroups were included within WT and KO groups (8-16 mice per group; Table 1).
2.3.2. Experimental Schedule
The total experimental period lasted for 24 days. The CUMS paradigm was applied from day 0 to day 21. Treatments were applied on days 16 and 20 (Figure 1A). There was an interval of 3 days between the two treatment days to allow for DMTS elimination from the body. DMTS and its vehicle were administered i.p. with a 30G needle in the lateral lower abdomen of the conscious animal. Injections were given seven times, once every hour at a dose of 30 mg/kg. Behavioural tests were carried out 24 hours after the last injection. The animals were terminated on day 23, 48 hours after the last stressor was applied, and their organs were collected. Control animals were measured in behavioural tests on days 7 and 11, and were treated with DMTS or the corresponding vehicle on the preceding days, in the same way as stressed animals (Figure 1B).
2.4. CUMS Paradigm
We used the CUMS model of depression, because the validity of this model was repeatedly confirmed [9,20,21]. The 3-week chronic stress paradigm consisted of 4 various mid-day stressors (restraint stress, tilted cage, shaker stress, dark room), applied between 10 a.m. and 2 p.m., as well as 3 types of overnight stressors (wet bedding, social isolation, group holding) (Table 2). In the case of „restraint stress”, the mice were closed into a plastic tube equipped with a perforated conical tip and several additional ventilation holes for 60 mins. For “tilted cage”, their cages were fixed at a 45° angle for 3 hours. For the “shaker stress”, mice were placed on a laboratory shaker set to 60 rounds per minute. For “dark room”, animals were kept in the dark for 3 hours during their daytime light hours. The overnight stress exposure period started at 6 p.m. and lasted until 6 a.m., except on “group holding” days, where animals were left undisturbed. For “wet bedding”, the sawdust was moisturized with 250 mL of tap water. The next morning mice were placed on fresh, dry bedding. For “social isolation”, mice were individually housed overnight. The next morning the original groups, previously housed in the same cage, were reunited. Stressors were applied randomly twice daily, day and night, for 3 weeks (Table 2.). All groups of animals received the stressors in the same order and no stressors were applied during the last 48 hours. Control groups were not stressed. The body weights of all mice were measured on days 1 and 15. The last body weight measurement took place before the anaesthetic injection on day 24, immediately before perfusion or decapitation.
2.5. Behavioural Tests
Five well-established behavioural tests were used to verify the anxiety (open field test (OFT) and marble burying test (MBT)), the anhedonia (sucrose preference test (SPT)) and the depression-like behaviour (forced swim test (FST), and tail suspension test (TST)), as well as to test the impact of DMTS.
2.5.1. Open Field Test
The OFT was conducted before performing the CUMS protocol to determine the optimal dosage of DMTS that does not impede the spontaneous movements of the animals. Animals were placed in the middle of a 60x60 cm box surrounded by 60 cm high walls. The behaviour of the animals was filmed for 7 minutes with a digital camera, and the last 5 minutes of the video recordings were evaluated. The evaluation began 30 seconds after the appearance of the animal in the open arena. Doses of 30 mg/kg and 40 mg/kg of DMTS and respective doses of vehicle were tested. The assessment continued for 5 minutes. The time spent moving and the total distance covered by the animal were recorded by the Noldus EthoVision XT 15 (Noldus) software.
Later in the setting of the CUMS protocol, the OFT was used to assess the anxiety level and exploratory behaviour of the mice on day 21. In this case, we evaluated the time spent in the centre of the opened arena. Anxious animals spend less time in the open central part of the arena. The time spent in the peripheral area alongside the walls is proportional to the level of anxiety [22].
2.5.2. Marble Burying Test
Mice were subjected to MBT for 30 min on day 17 of the CUMS protocol. Animals were placed one by one into a separate cage, where 24 coloured glass marbles of about 1 cm diameter had been placed. After thirty minutes, the number of marbles buried by the animals in the box was recorded. Marbles buried up to at least two-thirds of the diameter were counted as buried. More buried marbles indicate higher level of anxiety [23,24].
2.5.3. Sucrose Preference Test
On day 21 of the CUMS protocol, mice were subjected to SPT for 12 hours. Anhedonia is an important sign of depression characterised by a lack of interest in activities that normally bring pleasure. In animals, this can be assessed through the SPT. The test involves habituating the mice to two drinking bottles, one containing 1% m/v sucrose solution and the other containing tap water. The two bottles were offered randomly over four nights without separating the animals. On the second day, the animals were deprived of water for 12 hours before the dark phase of their circadian rhythm. Mice were placed individually into cages lined with clean litter and were randomly offered two drinking bottles, one with tap water and the other with sucrose solution at the beginning of the dark cycle. Consumption of tap water and sucrose solution was measured by weighing the bottles before and after the experiment. The sucrose preference was calculated as [consumption of sucrose solution/(consumption of water + consumption of sucrose solution) ]*100.
2.5.4. Forced Swim Test and Tail Suspension Test
On day 17 of the CUMS protocol, mice were subjected to FST. Animals were placed in a clear plastic container partially filled with water at 23 °C for 6 minutes. This test was conducted avoiding danger, but also escape of the mice from the container. Animals were recorded and the last 4 minutes of the test were evaluated. Water was drained from the fur of mice and they were returned to their cages when dry.
On day 21 of the CUMS protocol, mice were subjected to TST for 5 min. The TST involved holding the animals by their tails without causing any pain and securing them in a suspended position using adhesive tape. The suspension surface was 50 cm above the ground. Behaviour of the mice was recorded using a video camera. Care was taken not to injure the animals.
In both tests, EthoVision XT (Noldus) software was used to evaluate the time the animals spent moving and immobile, as well as the frequency of high activity periods.
Experiments were conducted in a secluded room to ensure the animals did not witness each other swimming or hanging. Bottles used in the experiments were thoroughly cleaned after each use. Mice were allowed to acclimate to the laboratory for 60 minutes each day for the three days before the experiment.
2.6. Termination
2.6.1. Perfusion and Tissue Collection
48 hours after any manipulation, half of each experimental group was anaesthetised by urethane (i.p., 2.4 g/kg). Animals were weighed, and their tails were clipped. Mice were then perfused transcardially with 20 mL of ice-cold 0.1 M phosphate-buffered saline (PBS; pH 7.4), followed by 150 mL of 4% paraformaldehyde solution in Millonig buffer (pH 7.4) for 15 minutes. Adrenal glands and thymus were removed and weighed using a Sartorius analytic scale (Sartorius AG). Data were corrected for body weight.
Brain samples were dissected and post-fixed in the same fixative solution for 72 hours at 4°C. We collected 5 series of 30 μm sections (Leica VT1000S vibratome; Leica Biosystems) and stored them in PBS containing 0.01% sodium azide at 4°C. Immunohistochemistry was performed to detect the expression of FOSB protein in the EWcp (Bregma -2.92 mm to -4.04 mm), dorsal raphe nucleus (DR; Bregma: -4.04 to -4.16 mm), basolateral amygdala (BLA; Bregma: -0.58 to -1.46 mm), lateral septum (LS; Bregma: 0.26 to -0.10 mm), paraventricular nucleus of the thalamus (PVT; Bregma: -0.22 to -0.70 mm), paraventricular nucleus of the hypothalamus (PVN; Bregma: 0.26 mm) and bed nucleus of the stria terminalis (BST; Bregma: 0.62 to -0.22 mm) according to Paxinos and Franklin, 2001.
2.6.2. Decapitation and Blood Sample Collection
The other half of each experimental group was terminated by cervical dislocation and blood was collected (500 μL) by cardiac puncture into a syringe containing 50 μL of 7.5% m/m EDTA solution (Sigma). Plasma samples were stored at -80 °C until ACTH and CORT content was determined by ELISA.
2.7. FOSB Immunohistology
Immunohistochemistry was carried out as described previously by Kormos et al. [20]. After washing sections were treated by 1% hydrogen-peroxide (H2O2) in PBS to inhibit the endogenous peroxidase activity. Permeabilization was carried out with 0.5% Triton X-100 solution for 30 min. Normal horse serum (2%; NHS, Jackson Immunoresearch, Europe Ltd., Suffolk, UK) in PBS for 30 min was applied to block the non-specific binding sites. Subsequently, sections were transferred into a solution of mouse anti-FOSB antibodies (Santa Cruz, sc-48 Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) in 1:500 dilution in PBS and 2% NHS in PBS overnight, at room temperature. After 2x15 min of washing in PBS, sections were treated with 1:200 diluted biotinylated horse anti-mouse antibodies for 60 min (Vectastain ABC Elite Kit, Vector Lbs., Burlingame, CA, USA) followed by 2 x15 min rinses in PBS. Preparations were then treated with peroxidase-conjugated avidin-biotin complex (Vectastain ABC Elite Kit) according to the supplier’s protocol for 60 min. After rinsing in PBS, the immunoreaction was developed in Tris buffer (pH 7.4) with 0.02% 3,3’ diamino-benzidine (DAB) (Sigma) and 0.03% v/v H2O2. The chemical reaction was accomplished under visual control using a microscope to optimise the signal/background ratio and was stopped after 7 min with PBS. Afterwards, the preparations were rinsed with PBS and mounted on slides coated with gelatin. After drying, slides were transferred into ascending ethanol solutions (70% (10 min), 96% (10 min), absolute (10 min)), then into xylene for 2x20 min and coverslipped using Depex (Fluka, Heidelberg, Germany) mounting medium.
2.8. Microscopy and Digital Imaging
DAB-labelled sections were examined and digitalised using a Nikon Microphot FXA microscope with a Spot RT camera (Nikon, Tokyo, Japan). Five serial sections were photographed at 90 lm luminous flux for each animal. The areas examined were EWcp, DR, BLA, LS, PVT, PVN, and BST.
2.9. Enzyme-Linked Immunosorbent Assay
Serum ACTH (VTSZ: 38220000) and CORT (VTSZ: 38220000) concentrations were measured using a commercially available ELISA set (Abcam). Blood was collected from the animals, centrifuged (3000 rpm, 5 min at 4 °C), and the supernatant was isolated and stored at -80°C until assayed.
When determining CORT, 25 μl of sample and standard solutions were added to the pre-coated antibody plate and incubated for 2 h at 37 °C followed by washing with 200 μl wash buffer. Then 50 μl of SP Conjugate reagent was added and incubated for 30 min with shaking. After another wash, 50 μl of chromogen substrate was added to each well and incubated until the optimal blue colour density developed. The reaction was stopped by adding 50 μl of stop solution, and absorbance was read at 450 nm using a microplate reader (Multiskan EX, Thermoscientific, USA).
During ACTH determination 50 μl of standard or sample and 50 μL of antibody cocktail were added to the wells. After incubation for 1 hour at room temperature, 350 μl of wash buffer per well was added, followed by 100 μl of 3,3’,5,5’ tetramethylbenzidine solution. The reaction was terminated with 100 μl stop solution per well after incubation for 20 minutes. Optical density (OD) values were read at 450 nm.
Serum CORT concentration was measured in ng/ml and ACTH concentration in pg/ml. In both cases, standard curves were generated and the unknown sample concentrations were calculated.
2.7. Evaluation Methods
2.7.1. Noldus EthoVision XT 15
The Noldus EthoVision XT system is designed to monitor animal movements in a laboratory setting. It uses an overhead camera and tracks animals based on black-and-white contrast, providing a reliable evaluation method compared to subjective techniques. The system generates a wide range of data and can be consistently applied across different laboratories [25].
In the case of OFT evaluation starts 30 seconds after the animal appears in the arena and lasts for 5 minutes. The time spent moving and the distance travelled during this time are assessed. In the case of FST, the last 4 minutes of the 6-minute recording were evaluated. We measured how long the animal was inactive and how often it entered a higher activity state based on the changes of the outline of the image of the mouse. Similarly, in the case of TST, the program assessed the time spent immobile and the number of active periods.
2.7.2. ImageJ
ImageJ software (Version 1.54d) and the IHC Toolbox was used for the morphometry. By outlining the brain area to be examined and setting the appropriate thresholds, the program separates the cells from the background based on density, counts the number of cells within the area, and calculates the size of the outlined area. The IHC Toolbox extension also includes a model explicitly optimised for DAB staining. The program allows manual or automatic cell counting. In our case, manual counting was impractical, given the large number of cells, so we used automatic counting. We converted the RGB image to greyscale (8-bit) then selected the area of interest. We highlighted all the structures to be counted using the Threshold command then, the program calculated the number of structures highlighted relative to the selected area.
2.8. Statistics
Data are presented as the mean and standard error of the mean. Three-way analysis of variance (ANOVA) was used to evaluate immunohistochemistry data. Normality of the data distribution was checked by the Shapiro-Wilk test and the homogeneity of variance by Bartlett's Chi-squared test [26]. Behavioural data were evaluated by one- (OFT for dose determination) and three-way ANOVA. All post hoc analyses were performed using the Tukey test based on the ANOVA's first- or second-order effects. Statistical evaluation was performed using GraphPad Prism 8.
3. Results
3.1. Finding the Suitable Dose of DMTS Using Open Field Test
Distance travelled was measured in centimetres (Figure 2A) and time spent moving was measured in seconds (Figure 2B). DMTS at 40 mg/kg significantly reduced both the time spent moving and the distance travelled in the observed time interval compared to the untreated group. No inhibition was observed at 30 mg/kg DMTS treatment. This dose was used in the following experiments using CNS models of depression-like behaviour. Neither the vehicle of 40 mg/kg, nor that of 30 mg/kg DMTS affected spontaneous movement of the animals. The vehicle of 30 mg/kg DMTS was used in further studies.
3.2. Validity of the Model
3.2.1. Endocrine and Physical Parameters
3.2.1.1. Changes in body Weights
In order to test the effects of CUMS, body weight change of the animals was recorded (Figure 3A). Wild-type, non-stressed mice tended to gain weight over the three weeks. CUMS effectively reduced body weight gain in non-injected, vehicle-treated and DMTS-treated Trpa1 WT mice (p < 0.001) compared to non-stressed mice. In contrast, KO mice did not lose weight significantly. The main effects of stress (p < 0.0001) and genotype (p < 0.0002) were significant, while the treatment effect was below the statistical threshold. The stress x genotype (p < 0.0004) interaction was significant (Table 3.). DMTS treatment did not reverse the effect of CUMS on body weight change in WT mice in comparison with naïve or vehicle-treated stress-exposed WTs. CUMS did not induce body weight change in KO mice, as revealed by post hoc comparisons in non-injected, vehicle-treated and DMTS-treated pairs of KO groups. Interestingly, non-stressed KO mice did not gain weight in any treatment groups, unlike non-stressed WT mice that gained 2-3 g (main genotype effect; p > 0.001). A comparison of body weight data (Figure 3B) before CUMS exposure revealed statistical differences between Trpa1 WT and KO mice (main genotype effect; p = 0.004) and between the Trpa1 WT naïve or vehicle-treated and DMTS-treated mice (p < 0.001). Post hoc tests confirmed that CUMS-exposed Trpa1 WT mice had lower body weight than control ones (main stress effect; p < 0.0001). Absolute body weight in stressed WT and Trpa1 KO mice did not differ significantly. Both the stress x genotype (p < 0.0001), stress x treatment (p = 0.0280), treatment x genotype (p = 0.0045) and treatment x stress x genotype (p = 0.0364) interactions were significant. Body weight data collectively prove that the stressors were effective in WT mice; interestingly, KO mice react differently to the same stress effect (Table 3).
3.2.1.2. Adrenal Gland and Thymus Weight
There was no difference in basal relative adrenal weight between the two genotypes. Relative adrenal weight was higher after exposure to CUMS (main effect of stress; p > 0.001) both in Trpa1 WT and KO mice in all treatment groups, and after treatment in Trpa1 WT and KO, non-stressed mice (main effect of treatment; p < 0.0001) in comparison to the naïve and vehicle-treated groups, without a difference in genotypes (Figure 3C). There was no interaction (Table 3). There was no difference in basal relative thymus weight between the two genotypes. CUMS exposure significantly reduced thymus weight (main effect of stress; p = 0.0028) in WT but not in the Trpa1 KO mice (main effect of genotype; p > 0.0001) in the naïve and vehicle-treated groups (Figure 3D). In WT mice that received DMTS treatment in addition to chronic stress, there was a lack of reduction in thymus weight (p < 0.0078) compared to the naïve and vehicle-treated groups. There was interaction between stress and genotype (p = 0.0017).
3.2.1.3. CORT and ACTH Serum Concentration
There was no difference in basal CORT levels between the two genotypes. CUMS significantly enhanced the CORT concentrations (main effect of stress; p = 0.0357) only in WT mice, with a difference in genotypes (Figure 3E). DMTS treatment significantly boosted the CORT concentration in non-stressed animals without a difference in genotypes (main effect of treatment; p = 0.0493). There was no interaction between the three factors.
There was a significant difference in basal ACTH levels between the two genotypes (Figure 2F). The baseline ACTH level in Trpa1 KO mice was slightly but significantly higher than in WT mice (p < 0.001). ANOVA revealed that CUMS influenced ACTH levels differently in the two genotypes (genotype x stress interaction; p < 0.0001). Post hoc tests showed that CUMS effectively increased the serum ACTH concentration in WT mice (main effect of stress; p < 0.0001); however, it failed to induce any further increase in KO mice. DMTS treatment significantly increased the serum ACTH concentrations (p = 0.0040), without a main effect, only in WT mice (treatment x genotype interaction; p = 0.0323; Table 3).
3.2.2. Behavioural Tests
3.2.2.1. Tests to Detect Anxiety Level
Time spent on the periphery (Figure 4B) was one of the main anxiety parameters of the OFT, and there were no differences in baseline between the two genotypes. Surprisingly, the chronic stress effect in any genotype did not affect this value.
However, DMTS treatment had different effects in the two genotypes (treatment x genotype interaction; p = 0.0118). DMTS treatment increased the time spent in the periphery in WT, non-stressed mice, while it had no effect in Trpa1 KO, non-stressed animals. Nevertheless, DMTS treatment had no effect in WT, stressed animals. However, it reduced the time spent in the periphery zone in Trpa1 KO, stress-exposed animals (treatment x stress interaction; p = 0.0002). We also examined the distance travelled at the periphery compared to the central zone (Figure 4B). There was also no difference in baseline, stress and treatment-free values between the two genotypes. Trpa1 WT and KO mice moved more in the periphery zone after CUMS exposure (main effect of stress; p < 0.0001), and this was also observed in non-stressed WT DMTS-treated animals (main effect of treatment; p = 0.0333). In contrast, in KOs (treatment x genotype interaction; p < 0.0001), the impact of DMTS was observed in the stressed animals (treatment x stress interaction; p < 0.0001), with less movement in the periphery zone after treatment (Table 3).
Regarding the marble burying test, we observed a baseline difference in anxiety levels between the two genotypes. Trpa1 KO mice buried significantly more marbles than WTs (main effect of genotype; p < 0.0001). Chronic stress-exposed WT animals buried significantly more marbles compared to the non-stressed animals in the MBT (main effect of stress; p > 0.0001; Figure 4C), but not the Trpa1 KO groups (stress x genotype interaction; p > 0.0001). Concerning DMTS treatment, we observed that the effect of the substance resulted in an increase in the number of buried marbles in WT non-stressed animals but a decrease in stressed animals compared to naïve or vehicle-treated groups (treatment x stress interaction; p = 0.0003). Furthermore, treatment x genotype x stress interaction was significant (p = 0.0047; Table 3).
3.2.2.2. Tests to Detect Anhedonia and Depression-like Behaviour
The preference of WT animals for sweetened water was significantly higher than that of Trpa1 KO animals in all treatment groups in the SPT (main effect of genotype; p > 0.0001; Figure 4D). CUMS exposure affected the sucrose preference in WT and Trpa1 KO animals (main stress effect; p > 0.0001). DMTS treatment increased the sucrose preference – i.e. reduced anhedonia – in stressed WT animals (main effect of treatment; p = 0.0474), with no impact in KO animals (stress x treatment y genotype interaction; p = 0.0359; Table 3).
FST and TST assessed the immobility time representing depression-like behaviour.
Regarding the forced swimming test, we found a baseline difference in depression levels between the two genotypes. Non-stressed Trpa1 KO mice had higher immobility scores than their WT counterparts (main effect of genotype; p > 0.0001). CUMS affected active duration (the main effect of stress; p < 0.0001) differently in the two genotypes in the FST (stress x genotype interaction; p > 0.0001; Figure 4E). Trpa1 KO mice showed no change in activity after chronic stress. This augmented depression-like behaviour was not altered by CUMS exposure in Trpa1 KO mice. DMTS treatment significantly enhanced the activity of WT, chronic stress-exposed animals (main effect of treatment; p = 0.0233). The treatment x genotype x stress interaction was also significant (p = 0.0070; Table 3).
In all treatment groups, Trpa1 KO animals showed significantly lower activity than the WTs in the TST (main effect of genotype; p > 0.0001; Figure 4F). As a result of CUMS, WT mice had significantly lower activity (p > 0.001) in naïve and vehicle-treated groups. DMTS-treated, stressed animals produced a higher level of activity (main effect of treatment; p = 0.0045). The impact of CUMS was not observed in Trpa1 KO mice (stress x genotype interaction; p = 0.0323), taking into account all treatment groups. The treatment x genotype x stress interaction was also significant (p = 0.0222; Table 3).
3.3. Pattern of FOSB Neuronal Activation in Stress-Related Brain Areas
The neuronal activation was assessed by FOSB immunocytochemistry in 8 stress-related brain areas.
3.3.1. Centrally Projecting Edinger-Westphal Nucleus
We found no difference in basal FOSB levels between the two genotypes in EWcp. FOSB reactivity was higher in WT naïve and vehicle-treated groups following CUMS exposure (main effect of stress; p = 0.0008). ANOVA found the main effects of genotype (p = 0.0014) and stress x genotype (p < 0.0001) interactions significant (Figure 5). The DMTS-treated group showed significantly lower neuronal activation after CUMS exposure compared to the naïve or vehicle-treated groups (p < 0.0001), but only in the WT mice. The treatment x stress genotype interaction was significant (p = 0.0448; Table 4).
3.3.2. Dorsal Raphe Nucleus
We found no difference in basal FOSB levels between the two genotypes in DR. In naïve and vehicle-treated WT mice, CUMS induced more than threefold FOSB positivity (p = 0.0002). The effect of stress (p = 0.0002) and genotype (p = 0.0338), in addition, stress x treatment (p = 0.0276) and treatment x stress x genotype (p < 0.0057) interactions had a statistical interaction (Table 4). In contrast, if WT mice were DMTS treated, the FOSB elevation was significantly reduced (p = 0.004). CUMS and DMTS treatment did not induce the rise of FOSB expression in the DR of KO mice (Figure 6).
3.3.3. Periaqueductal Gray Matter
Basal FOSB reactivity did not differ between the two genotypes in the PAG brain area. CUMS influenced FOSB expression in the PAG (main effect of stress; p < 0.0001 Figure 7). Stressed WT mice showed almost three times higher neuronal activity than non-stressed animals. CUMS did not increase FOSB immunoreactivity in KO mice (stress x genotype interaction; p = 0.0006). DMTS treatment effectively reduced FOSB immunoreactivity in the PAG in WT stressed animals (main effect of treatment; p = 0.0311), but it had no effect in Trpa1 KO ones (treatment x genotype interaction; p = 0.0128) or non-stressed WT animals (treatment x stress interaction; p = 0.0005). Treatment x stress x genotype interaction was not significant (p = 0.0555; Table 4).
3.3.4. Paraventricular Nucleus of the Thalamus
We found a difference in basal FOSB reactivity between the two genotypes in the PVT brain area. KO, non-stressed mice showed higher FOSB positive cell counts in the PVT than their WT counterparts (p = 0.0041). CUMS exposure significantly increased FOSB immunoreactivity in WT mice in the PVT (p = 0.0468; Figure 8) without the main effect of treatment but had no effect in Trpa1 KO mice (stress x genotype interaction; p = 0.0001). DMTS treatment enhanced FOSB immunoreactivity in WT non-stressed animals and it reduced FOSB positive cell counts in WT stressed animals. Still, in both cases, the effect was only a tendency and not statistically significant. DMTS treatment did not affect KO mice. The treatment x stress x genotype interaction was significant (p = 0.0397; Table 4).
3.3.5. Lateral Septal Nucleus
We found a difference in basal FOSB reactivity between the two genotypes in the LS brain area. Trpa1 KO non-stressed animals exhibited significantly lower FOSB positive cell counts than their WT counterparts (main effect of genotype; p > 0.0001). ANOVA revealed the main effect of stress in the LS (p = 0.0004; Figure 9). In WT mice CUMS increased the FOSB positive cell counts, but KO mice did not show CUMS-induced FOSB elevation, but only a tendency. DMTS treatment significantly reduced the number of FOSB positive cells in WT stressed animals compared to the naïve or vehicle-treated groups (p = 0.0010), without the main effect of treatment. DMTS treatment did not impact Trpa1 KO mice (treatment x genotype interaction; p = 0.0255). Treatment x stress x genotype interaction was also significant (p = 0.0296; Table 4).
3.3.6. Paraventricular Nucleus of the Hypothalamus
In basal FOSB reactivity, no difference was found between the two genotypes in the PVN brain area. The assessment of FOSB immunoreactivity in the PVN of treated mice (Figure 10) revealed the main effects of stress (p < 0.0001) and genotype (p < 0.0005) to be highly significant. ANOVA found an interaction between treatment x stress (p = 0.0077) and treatment x genotype (p = 0.0257). Post hoc comparisons revealed that CUMS caused about twice the FOSB increase in naïve and vehicle-treated WT and KO animals (p < 0.01) without the main genotype effect. Interestingly, DMTS treatment caused an effective decrease in the number of FOSB positive cells in WT chronic stress-exposed mice (p = 0.0001), but not in the KO stressed mice (treatment x genotype interaction; p = 0.0257). DMTS treatment did not affect non-stressed WT and KO mice (treatment x stress interaction; p = 0.0077; Table 4).
3.3.7. Bed Nucleus of the Stria Terminalis
We found a difference in basal FOSB reactivity between the two genotypes in the BST (Figure 11). FOSB immunoreactivity was significantly higher in the WT mice compared to the non-stressed groups in every treatment group as a result of CUMS (main effect of stress; p < 0.0001), but chronic stress exposure did not affect Trpa1 KO mice (stress x genotype interaction; p = 0.0158). In addition, FOSB cell count was significantly higher in Trpa1 KO mice in comparison to WT animals (main effect of genotype; p < 0.0001). There was no main effect of treatment or any other interaction (Table 4).
3.3.8. Basolateral Amygdala
4. Discussion
We tested the effects of a naturally occurring polysulfide found in garlic – DMTS – on chronic stress-induced anxiety and depression-like behaviour. We hypothesize that the TRPA1 ion channel might be involved as a possible mediator of DMTS effects. To the best of our knowledge, the anti-anxiety and anti-depressant effects of DMTS have not been studied before. However, a recent study by our research team has demonstrated the role of DMTS in acute stress [27].
4.1. Validity of the Model
We decided to utilise the CUMS model because it is a widely recognised and well-studied model in mice [28]. Exposure to CUMS resulted in typical physiological [29,30,31], hormonal (ACTH, CORT) and behavioural (anxiety, anhedonia, and depression-like) changes indicating heightened activity within the HPA axis [20,32]. The stress induced by the anaesthetic injection administered before perfusion may have resulted in acute elevations in ACTH and CORT levels. Despite this potential confounding factor, our observations of increased adrenal weight, diminished thymus mass, and body weight reduction strongly indicate prolonged constitutive activity of the HPA axis. These findings confirm the validity and reliability of our model. An interesting observation is that in the absence of the Trpa1 gene, animals respond differently to the effects of chronic stress, suggesting a stress adaptation disorder in the lack of this gene [2].
4.2. Normal Stress Response Requires TRPA1 Ion Channel, But the Impact of DMTS Is Independent of the TRPA1 on Physical and Endocrine Parameters
We found TRPA1-related basal alterations in ACTH levels, body weight changes, and absolute body weights, but not in the relative weights of the thymus and adrenal glands or CORT levels. Non-stressed Trpa1 KO mice showed reduced body weight gain compared to WT ones, and their absolute body weight was lower, too. This could be explained by potential anorexigenic effect of TRPA1. Some studies suggest that the lack of TRPA1 leads to an upregulation of urocortin 1 (UCN1) [2]. UCN1 belongs to the corticotrophin-releasing hormone (CRH) family, with an anorexigenic effect [2,33]. Basal levels of ACTH were elevated in KO mice and could not increase further upon CUMS. No basal difference in CORT levels was found, but the CORT level response to chronic stress differs between the two genotypes. A statistically insignificant elevation of CORT levels was observed in response to stress in KO animals. This suggests that TRPA1 modulates the HPA axis response at the anterior pituitary or adrenal cortex levels. There was no basal difference in thymus weights between the two genotypes, but there was a difference in response to chronic stress. No stress-induced alteration of the thymus weight was observed in gene KO animals. This correlates with increased CORT levels under chronic stress, as glucocorticoids exert an immunosuppressive effect associated with a decrease in the thymus weight. TRPA1 may modulate lymphocyte differentiation during chronic stress, too. The TRPA1 protein has a dual role in immunity. It acts as a sensor for cellular stress, tissue injury, and harmful external stimuli and triggers defensive responses. However, when not properly regulated, it can also contribute to the impaired function of the immune system [34,35]. TRPA1 activity may play a role in developing immune cells of primary and secondary immune organs [35,36,37].
Overall, chronic stress affected all parameters in Trpa1 WT animals. Most parameters were not affected by stress in KO mice, except the weight of the adrenal glands. The HPA axis function is altered in the absence of the TRPA1 ion channel and the channel may be required for normal stress response.
DMTS treatment resulted in increased resting CORT and ACTH levels, as well as adrenal gland weight. This was not observed regarding ACTH levels in KO animals. DMTS affected these values differently in stressed animals. In the early 2000s, Russo and colleagues proved that H2S could modulate the hypothalamic-pituitary system [38]. The effects of DMTS on CORT levels and adrenal weight are indeed not mediated through the TRPA1 ion channel. Elevated ACTH levels may have occurred due to the involvement of the TRPA1 ion channel. DMTS probably affected the pituitary gland via the TRPA1 ion channel and the adrenal gland through other routes. The reduction of thymus weight under chronic stress was prevented by DMTS treatment, suggesting that it somehow diminished the immunosuppressant effect of glucocorticoids. TRPA1 ion channel might be implicated, given the difference in KO animals. The protective effect of sulfides has been described in several papers explaining the positive effect of DMTS on the immune system under oxidative stress [39] and chronic endoplasmic reticulum stress [40]. Although there was no significant effect on body weight change and absolute body weight, there was a trend towards a slight reduction of body weight gain in WT animals. The TRPA1 ion channel might have anorexigenic effect [41].
Overall, DMTS treatment elevates CORT levels in both genotypes of control animals. DMTS treatment did not decrease thymus size in wild-type mice and it did not prevent thymus weight reduction in KO ones because the weight did not increase in response to stress. Our results suggest that the effect of DMTS in control is not TRPA1-dependent. DMTS has no impact on physical and endocrine parameters in stressed mice. It is important to mention that at termination, DMTS may no longer maintain plasma levels as high as at the time of behavioural testing, which may explain the absence of impact on parameters.
4.3. Lack of TRPA1 Abolished the Inhibitory Effect of DMTS on CUMS-Induced Depression-like Behaviour of Mice
Gene-knockout mice showed basally elevated anxiety levels in the MBT but not in the OFT. KO animals presented increased depression-like behaviour in the SPT, FST and TST. Basal anxiety levels are not different between the two genotypes, according to OFT data. Chronic stress caused a similar increase in anxiety levels in KO animals and their WT counterparts. The absence of any basal genotype-related difference in the OFT test results supports the idea that effects could be specific to the test [42].
Under chronic stress increased anxiety levels, anhedonia and reduced activity were detected in wild-type mice indicating depression-like behaviour. Basal depression-like phenotype observed in KO mice might be explained by increased UCN1 and CRH2 receptor signalling in the DR [43,44,45].
The two genotypes differed in their response to stress. KO animals appeared to be unaffected by stress, probably because they already exhibited depression-like behaviour.
CUMS induced depression- and anxiety-like phenomena in FST, TST, SPT, MBT and OFT in the naïve stressed group compared to the control group in WT animals. Stress-induced anxiety was prevented by the lack of Trpa1.
Trpa1 KO animals showed no effect of CUMS on depression-like behaviour both in FST and TST. However, anhedonia developed upon chronic stress in KO animals in SPT, which is a key symptom of depression [46]. Our results align with those of Kormos et al. [2]. The lack of TRPA1 also prevented the stress-related increase of anxiety levels in the MBT but not in the OFT. The changes observed may be due to increased basal depression-like behaviour in Trpa1 KO mice, which prevented further increases in depressive parameters.
Conflicting findings were obtained regarding the impact of DMTS on anxiety levels. DMTS treatment can increase baseline anxiety levels of mice via TRPA1 ion channel activation in the OFT. DMTS treatment reduced the anxiety levels of stressed KO animals. The treatment increased baseline anxiety levels in the MBT partly via TRPA1, it seemed to reduce anxiety levels in stressed, anxious animals in a TRPA1-dependent way as well. The fact that DMTS had a TRPA1-mediated differential effect on anxiety and depression-like behaviour in animals proposes that anxiety may be differentially regulated from depression. The TRPA1 ion channel also plays a role in this differential regulation [2]. Previous studies revealed an inhibitory action of H2S on anxiety-like behaviour mediated by mitigation of oxidative stress [47] or regulation of neuroplasticity [48].
Our findings regarding the effect of DMTS on depression-like behaviour were more consistent. DMTS-treated WT mice exhibited reduced anhedonia and depression-like behaviour when exposed to chronic stress. On the other hand, there was no effect of DMTS on depression-like behaviour in KO animals. The treatment did not change their baseline depression-like behaviour. DMTS did not prevent the development of anhedonia in KO mice. The protective effect of DMTS treatment against anhedonia is mediated through the TRPA1 ion channel. Anhedonia extends beyond major depression into the field of anxiety disorders. Anhedonia may be present in generalised anxiety disorder [49]. Our results obtained in the OFT, a test assessing anxiety, as well as in the relative mass of the adrenal glands in KO animals corroborate these findings. This is important because extant psychological and pharmacological therapies are relatively ineffective against anhedonia. There is an unmet therapeutic need for this high-risk symptom. Current psychological and medicinal treatments of anxiety and depression focus largely on reducing negative mood rather than improving positive mood [46].
Overall, the CUMS paradigm resulted in anxiety and depression-like phenotype and the response to chronic stress seems to be TRPA1-dependent. DMTS treatment alleviates depression-like behaviour and anxiety. In most cases this is a TRPA1-independent process. Except for sucrose preference, TRPA1 is required for DMTS to mitigate anhedonia. One possible underlying mechanism for this is that H2S has TRPA1-independent antagonistic effect on depression-like behaviour mediated by facilitation of hippocampal long-term potentiation and augmentation of synaptic neurotransmission involved in the regulation of synaptic plasticity [22]. In addition to cognitive impairment, several studies have linked depression and anxiety to alterations in synaptic plasticity [50,51,52].
4.4. Effect of DMTS Treatment on Neuronal Activation Depends on Stress Exposure and the Presence of Functional Trpa1
ΔFOSB accumulates after repeated stimulation. Growing evidence suggests that the accumulation is a homeostatic response to chronic stress. Overexpression of ΔFOSB in the brain has been found to enhance resilience to stress, while inhibition of its activity has been associated with increased susceptibility to stress [53]. The FOSB antibody used in the present study recognises all splice variants of the full-length FOSB and ΔFOSB proteins. The latter isoform is involved in chronic stress adaptation [54,55,56,57].
In our previous study, we performed RNAscope ISH to detect Trpa1 expression in the lateral septum (LS) and paraventricular nucleus of the thalamus (PVT). RNAscope is more appropriate than immunohistochemistry due to the lack of sufficiently specific commercially available antibodies [58,59,60]. Our findings suggest that the LS and PVT areas do not express Trpa1 [27].
The basal neuronal activation of the two genotypes was not different in EWcp, DR, PAG, PVN and BLA brain areas. However, our findings revealed a fundamental difference between Trpa1 WT and KO mice. FOSB immunoreactivity in KO non-stressed mice was higher in the PVT but lower in the LS and BST areas compared to their WT counterparts.
The results of our study indicate that exposure to chronic stress significantly increased neuronal activation in the tested brain areas in Trpa1 WT animals except the BLA. Interestingly, the impact of chronic stress on FOSB expression was observed in the PVN brain area of KO mice, although to a lesser extent than in WT animals. This suggests that the elevated neuronal activation response to chronic stress in PVN is TRPA1-dependent. TRPA1-dependent activation can occur directly or indirectly, depending on whether the ion channel is expressed in the brain area.
In the EWcp, DR, and PAG, we obtained similar results for FOSB immunoreactivity in response to chronic stress regarding the differences observed between genotypes. DMTS treatment reduced the elevated neuronal activity after chronic stress exposure which is in line with the findings from behavioural tests of depression-like behaviour. Neuronal activation in these brain areas was not significantly higher after chronic stress in DMTS-treated animals. Since no increase in neuronal activation after chronic stress was observed in KO mice, the effect of DMTS cannot be regarded as TRPA1-dependent. The neurons that produce 5-HT in the DR and the cells that produce UCN1 in the EWcp both play a role in the development of mood disorders and anxiety related to stress [61]. When the neurons in the DR are activated, anxiety-like behaviour arises [62]. The Trpa1 ion channel was detected in certain cells of the DR but not in serotonergic cells (Milicic et al. under publication), suggesting different ways of mediating the effects of DMTS in this area. EWcp urocortinergic neurons interact with DR serotoninergic neurons, forming a back-and-forth connection between the two nuclei. The DR has CRH receptors. The projection is affected by UCN1 and modulates 5-HT release, affecting mood and anxiety [63,64]. The urocortinergic neurons from the EWcp nucleus project primarily to the DR and PVN areas, which are involved in the stress response [63,65,66]. It is thought that the projections from the DR cause the suppression of PAG cells. Changes in the levels of extracellular 5-HT in the DR have been shown to decrease the panic responses triggered by PAG stimulation [67].
Although the PVT does not express the ion channel [27], we noticed a difference in basal neuronal activation between the two genotypes, which has not changed in response to chronic stress. It is important to note that some of the efferents from the EWcp expressing Trpa1 innervate the PVT. This neuronal input may explain the difference in basal neuronal FOSB immunoreactivity in KO animals [68]. The PVT is heavily influenced by input from the hypothalamus, which includes a dense network of neuropeptide-containing neurons [69]. Prefrontal cortical areas, such as the infralimbic, prelimbic, and insular cortices, play a crucial role in providing input to the PVT. The dorsomedial nucleus of the hypothalamus, the PAG, and the lateral parabrachial nucleus also contribute to the input received by the PVT. Additionally, the PVT receives projections from various neurons located throughout the brainstem and hypothalamus. These widespread projections to the PVT demonstrate a remarkable diversity in neurochemical content, including fibrous projections involving monoamines (such as dopamine, noradrenaline, adrenaline, and serotonin (5-HT)) as well as an extensive array of neuropeptides [70]. The neuronal activity-enhancing effect of chronic stress was observed in the PVT, too. Neuronal activation did not increase further in response to chronic stress in KO mice, maybe because it was already elevated. The effect of DMTS was not statistically significant in the PVT area. There was a tendency for DMTS to reduce neuronal activation in wild-type mice exposed to chronic stress .
The LS has been shown to play a key role in emotional processes and responses to stress. Some studies suggest that the LS promotes coping behaviour during active stress and is involved in an HPA axis inhibitory mechanism mediated at least in part by septal 5-HT1A receptors and does not undergo glucocorticoid-mediated feedback [71]. Even though Trpa1 is not present in the LS area [27], we still noted a difference in the baseline neuronal activation of Trpa1 KO and WT mice in that brain region following chronic stress and DMTS treatment. TRPA1 may be involved in facilitating the activation-boosting effect of DMTS through an indirect mechanism. The LS is reached by a highly intense urocortinergic fibre network from the EWcp [72]. It is known that UCN1 cells express the highest amount of Trpa1 in the mouse brain [2]. The absence of TRPA1 in the EWcp nucleus in KO animals may explain the findings in the LS region. DMTS treatment did not decrease neuronal activation in stressed KO animals and chronic stress did not increase it.
No basal difference in neuronal activation was observed between the two genotypes in the PVN area. Chronic stress strongly increased neuronal activation in WT and KO animals despite the PVN expressing Trpa1 (Milicic et al., under publication). Other processes are probably involved in adapting the PVN to chronic stress besides TRPA1. The PVN is a region that is essential for the initiation of stress responses. Chronic stress induces changes in the expression of peptides, neurotransmitter receptors and neuronal excitability in the PVN [73,74,75]. PVN neurons are hyperreactive and hyperresponsive to stimuli following repeated stress, contributing to the acute hormone release-promoting effect of chronic stress. Chronic stress increases the number of presynaptic excitatory neurotransmitter boutons (glutamate and norepinephrine) targeting CRH cell bodies and dendrites [76]. We noticed a reduction in neuronal activity in WT animals following the application of DMTS, indicating that the TRPA1 ion channel is at least partly involved in mediating the effects of DMTS in the PVN region. It is important to note that changes in the SPT after DMTS application and chronic stress exposure are consistent with changes in the PVN. Considering that the PVN expresses the TRPA1 ion channel and the SPT test showed no effect of DMTS in the KO animals, the positive action of DMTS on anhedonia levels may be due to the activation of TRPA1 ion channels.
There is a significant difference in neuronal activation between the two genotypes in the BST region, with lower activation in the gene knockout mice. Neuronal activation was increased in response to chronic stress. Similarly to the LS and PVT areas, we found a difference in basal and post-chronic-stress neuronal activity in the BST area between WT and KO animals. DMTS treatment had no effect on neuronal activation in this brain region in either group. The BST is believed to be a hub connecting limbic cognitive centres and nuclei involved in processing reward, stress, and anxiety. It also receives incoming information about systemic stressors like hypertension and haemorrhage [77]. As a result, the BST appears to have an important role in coordinating physiological and behavioural responses, linking structures such as the amygdala, hippocampus and medial prefrontal cortex (MPFC) to hypothalamic and brainstem regions involved in autonomic and neuroendocrine functions [78]. Some peptides present in the BST, such as pituitary adenylate cyclase-activating polypeptide (PACAP), can directly activate CRH neurons. There may be potential excitatory peptidergic projections to the PVN [79]. PACAP is up-regulated in the BST by chronic stress, leading to hyperactivity of the HPA axis [72]. The impact of the BST on how the body responds to stress depends on how long the exposure to stress lasts. For instance, when the ventral part of the BST is damaged, it reduces the response to acute stress, but it significantly increases the stress response of the body after prolonged exposure to unpredictable stress. Different groups of cells may be activated by sudden stress compared to prolonged one. Damage to the dorsal part of the BST heightens the stress response after prolonged exposure to unpredictable stress, indicating its role in keeping the stress system from becoming too sensitive after prolonged stress [80].
There was no difference in neuronal activation between the two genotypes in the BLA. Stress had no effect in either group, nor did DMTS treatment affect neuronal activation in this brain region. The BLA is a critical brain region involved in fear and anxiety [81]. Recent studies suggest that the medial amygdala (MeA), BLA, and central amygdala (CeA) do not exhibit increased FOSB/ΔFOSB expression in response to acute restraint stress or chronic variable/repeated stress [82].
It is also worth mentioning that the magnitude of neuronal activation was specific to the given brain area. We observed a nearly fourfold increase in FOSB expression in the EWcp and DR. It was approximately threefold in the PAG and PVN areas. We observed a 2-2.5-fold increase in the PVT and BST and only a 1.5-fold increase in the LS.
Summarising the effects of DMTS treatment, we observed that it did not impact FOSB expression in KO mice. However, FOSB immunoreactivity was significantly lower in WT DMTS-treated animals than in their stressed but untreated counterparts in all brain areas that we examined, except in the BST and BLA. Notably, the magnitude of the impact of DMTS treatment was also brain area specific. Brain areas known to express TRPA1 (EWcp, PAG, DR) exhibited the largest effect upon DMTS administration. Brain areas that do not express the TRPA1 (LS, PVT) ion channel also responded to DMTS, but the effect was 2-3x less. In brain areas that do not express Trpa1, indirect activation from an area expressing TRPA1 may be the explanation [68]. TRPA1 is only partially involved in mediating the effects of DMTS in stress adaptation among other processes.
The protective effect of sulfides, specifically that of H2S, in the CNS have been previously described. They have been studied in various neurodegenerative diseases, focusing on protein S-persulfidation [83], the role of sulfide in neuroprotection through the TRPA1 ion channel [84] or its participation in neuromodulation through the reduction of endoplasmic reticulum (ER) stress [85]. H2S used in previous studies cannot directly modify cysteines by persulfidation. Polysulfides, such as alkyl trisulfides like DMTS, can directly participate in the reaction [86]. Chemically, alkyl trisulfides form tri- and disulfide metabolites with thiol groups of cysteine amino acids, while inorganic polysulfides lead to persulfidation of proteins. For this reason, the two groups of compounds (organic and inorganic polysulfides) may have different biological effects. DMTS has an elimination half-life of 36 minutes and accumulates mainly in red blood cells. It readily crosses the blood-brain barrier and can be detected in brain tissue. It is also patented in the United States of America as an intramuscular antidote for cyanide poisoning [87,88,89,90,91,92].
The changes in physical/endocrine, behavioural and neuronal activation parameters in response to chronic stress and DMTS treatment can be related in some instances. In Trpa1 wild-type mice, anxiety and depression-like behaviour under chronic stress were measured. DMTS attenuated all these effects.
We observed correlations between relative adrenal weights, SPT findings, OFT results and PVN brain area activation changes after chronic stress in gene knockout mice. Relative adrenal weight, depression and anxiety parameters in gene-knockout animals were not affected by DMTS. The chronic stress-induced anhedonia assessed in the SPT is not a TRPA1-dependent process but that TRPA1 must partially mediate the reduction in anhedonia by DMTS treatment. In addition, based on the neuronal activation pattern in the PVN, this brain area mediates the antidepressant effect of DMTS on anhedonia in a TRPA1-dependent manner.
Antidepressant and anxiolytic effects of DMTS may be mediated through modulation of the HPA axis and may be mediated by the EWcp, DR, PAG, LS and PVN either directly or indirectly through the TRPA1 ion channel.
A brief summary of our main findings (Figure 13):
- Trpa1 deficiency affects/modifies the response to chronic stress, both at the level of behaviour and neuronal activation.
- CUMS leads to anxiety and depression-like behaviour, dysregulation of the HPA axis and activation of brain areas important in stress adaptation in a TRPA1-dependent manner.
- DMTS alleviates depression-like behaviour in Trpa1 WT mice.
- Lack of TRPA1 reverses the antidepressant-like action of DMTS in chronic stress-exposed mice according to the SPT test.
- DMTS alleviates thymus involution caused by dysregulation of the HPA axis due to chronic stress.
- The action of DMTS on anxiety and depression-like behaviour may be differentially and independently regulated.
- The absence of TRPA1 ion channel mitigates the inhibitory effect of DMTS on chronic stress-induced neuronal activation in PVN.
5. Conclusions
We confirmed the role of the TRPA1 ion channel in stress adaptation processes using a CUMS mouse model. We identified the potential anti-anxiety and anti-depressant effects of the naturally occurring organic polysulfide DMTS and the partial role of the TRPA1 ion channel in mediating these effects. We identified the brain areas mediating the effects of DMTS on stress adaptation processes and the extent of the impact in each brain area.
Our findings provide a novel insight into the mechanism of the ameliorating effect of DMTS on depression and anxiety and suggest that TRPA1 is a promising therapeutic target for mood disorders, which exploits new paths for the prevention and cure of depression under chronic stress.
Based on all this, DMTS is an ideal candidate for further investigation as a dietary supplement or complementary therapeutic in the treatment of anxiety and depression.
6. Limitations
It is necessary to point out the shortcomings of our experiments. The behavioural phenotype of our global KO mouse strain may be due to developmental compensation. We cannot exclude the possibility that other peripheral or central mechanisms, which we did not study here, may contribute to the behavioural phenotype of the KO mice, as the functional receptor was deleted both peripherally and centrally [2,93,94]. In this study, female mice and tissue samples were not examined. The oestrous cycle may influence the responses in female mice, as some of the brain areas that we examined express the oestrogen receptor ß [95,96,97,98,99,100,101]. Another limitation is related to the substance used in our research study. DMTS has not been investigated in behavioural tests before except our recently published paper [27], so there is not enough data on the possible side effects of DMTS on the CNS and animal behaviour.
Author Contributions
Conceptualization, G.P.; methodology, G.P., K.G., V.K.; validation G.P.; formal analysis, G.P.; writing—original draft preparation, K.G.; writing—review and editing, G.P. and V.K.; visualization, G.P., K.G., and V.K.; supervision, G.P.; funding acquisition, G.P. and E.P. E.P. provided the C57BL6J, Trpa1 KO, and WT mice. All the authors have read and agreed to the published version of the manuscript. All the authors have read and agreed to the published version of the manu-script.
Funding
This work was supported by OTKA FK 132454 from the National Research, Development, and Innovation Office, Hungary. VK was supported by the János Bolyai Research Scholarship of the Hungarian Academy of Sciences (BO/00750/22/5) and TKP2021-EGA-16.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, the 1998/XXVIII Act of the Hungarian Parliament on Animal Protection and Consideration Decree of Scientific Procedures of Animal Experiments (243/1998), the European Communities Council Directive of 2010/63/EU, and the requirements of the International Association for the Study of Pain (IASP) and was approved by the National Scientific Ethical Committee on Animal Experimentation (license number BA02/2000-31/2022) approved on 30 May 2022).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We wish to thank Ágnes Dombi and Csenge Sánta for their expert technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 5-HT | Serotonin |
| ACTH | Adrenocorticotropic hormone |
| ANOVA | Analysis of variance |
| BLA | Basolateral amygdala |
| BST | Bed nucleus of the stria terminalis |
| CeA | Central amygdala |
| CHO | Chinese hamster ovary |
| CORT | Corticosterone |
| CNS | Central nervous system |
| CRF | Corticotropin-releasing factor |
| CUMS | Chronic unpredictable mild stress |
| DAB | 3,3′-Diaminobenzidine |
| DARK | Dark room |
| DMTS | Dimethyl trisulfide |
| DR | Dorsal raphe nucleus |
| ELISA | Enzyme-linked immunosorbent assay |
| ER | Endoplasmic reticulum |
| EWcp | Centrally projecting Edinger-Westphal nucleus |
| FTS | Forced swim test |
| GH | Group holding |
| H2O2 | Hydrogen peroxide |
| H2S | Hydrogen sulfide |
| HPA | Hypothalamic-pituitary-adrenal |
| IHC | Immunohistochemistry |
| i.p. | Intraperitoneal |
| ISOL | Social isolation |
| KO | Knockout |
| LS | Lateral septum |
| MBT | Marble burying test |
| MeA | Medial amygdala |
| mPFC | Medial prefrontal cortex |
| OFT | Open field test |
| PACAP | Pituitary adenylate cyclase-activating polypeptide |
| PAG | Periaqueductal gray matter |
| PBS | Phosphate-buffered saline |
| POLY | Inorganic polysulfide |
| PTSD | Post-traumatic stress disorder |
| PVN | Paraventricular nucleus of the hypothalamus |
| PVT | Paraventricular nucleus of the thalamus |
| REST | Restraint stress |
| SHAKE | Shaker stress |
| SPT | Sucrose preference test |
| TILT | Tilted cage |
| TRPA1 | Transient receptor potential ankyrin 1 |
| TST | Tail suspension test |
| UCN1 | Urocortin 1 |
| WET | Wet bedding |
| WM | Body weight measurements |
| WT | Wild type |
References
- Shi, L.; Lin, Y.; Jiao, Y.; Herr, S.A.; Tang, J.; Rogers, E.; Chen, Z.; Shi, R. Acrolein scavenger dimercaprol offers neuroprotection in an animal model of Parkinson’s disease: implication of acrolein and TRPA1. Transl. Neurodegener. 2021, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kormos, V.; Kecskés, A.; Farkas, J.; Gaszner, T.; Csernus, V.; Alomari, A.; Hegedüs, D.; Renner, É.; Palkovits, M.; Zelena, D.; et al. Peptidergic neurons of the Edinger–Westphal nucleus express TRPA1 ion channel that is downregulated both upon chronic variable mild stress in male mice and in humans who died by suicide. J. Psychiatry Neurosci. 2022, 47, E162–E175. [Google Scholar] [CrossRef] [PubMed]
- Al-Omari, A.; Kecskés, M.; Gaszner, B.; Biró-Sütő, T.; Fazekas, B.; Berta, G.; Kuzma, M.; Pintér, E.; Kormos, V. Functionally active TRPA1 ion channel is downregulated in peptidergic neurons of the Edinger-Westphal nucleus upon acute alcohol exposure. Front. Cell Dev. Biol. 2023, 10, 1046559. [Google Scholar] [CrossRef] [PubMed]
- Konkoly, J.; Kormos, V.; Gaszner, B.; Correia, P.; Berta, G.; Biró-Sütő, T.; Zelena, D.; Pintér, E. Transient receptor potential ankyrin 1 ion channel expressed by the Edinger-Westphal nucleus contributes to stress adaptation in murine model of posttraumatic stress disorder. Front. Cell Dev. Biol. 2022, 10, 1059073. [Google Scholar] [CrossRef]
- Talavera, K.; Startek, J.B.; Alvarez-Collazo, J.; Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Naert, R.; Nilius, B. Mammalian Transient Receptor Potential TRPA1 Channels: From Structure to Disease. Physiol. Rev. 2020, 100, 725–803. [Google Scholar] [CrossRef]
- De Silva, D.; Lee, S.; Duke, A.; Angalakurthi, S.; Chou, C.-E.; Ebrahimpour, A.; Thompson, D.E.; Petrikovics, I. Intravascular Residence Time Determination for the Cyanide Antidote Dimethyl Trisulfide in Rat by Using Liquid-Liquid Extraction Coupled with High Performance Liquid Chromatography. J. Anal. Methods Chem. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Kiss, L.; Bocsik, A.; Walter, F.R.; Ross, J.; Brown, D.; A Mendenhall, B.; Crews, S.R.; Lowry, J.; Coronado, V.; E Thompson, D.; et al. From the Cover: In Vitro and In Vivo Blood-Brain Barrier Penetration Studies with the Novel Cyanide Antidote Candidate Dimethyl Trisulfide in Mice. Toxicol. Sci. 2017, 160, 398–407. [Google Scholar] [CrossRef]
- Kiss, L.; Holmes, S.; Chou, C.-E.; Dong, X.; Ross, J.; Brown, D.; Mendenhall, B.; Coronado, V.; De Silva, D.; Rockwood, G.A.; et al. Method development for detecting the novel cyanide antidote dimethyl trisulfide from blood and brain, and its interaction with blood. J. Chromatogr. B 2017, 1044-1045, 149–157. [Google Scholar] [CrossRef]
- Liu, S.-Y.; Li, D.; Zeng, H.-Y.; Kan, L.-Y.; Zou, W.; Zhang, P.; Gu, H.-F.; Tang, X.-Q. Hydrogen Sulfide Inhibits Chronic Unpredictable Mild Stress-Induced Depressive-Like Behavior by Upregulation of Sirt-1: Involvement in Suppression of Hippocampal Endoplasmic Reticulum Stress. Int. J. Neuropsychopharmacol. 2017, 20, 867–876. [Google Scholar] [CrossRef]
- Wei, L.; Kan, L.-Y.; Zeng, H.-Y.; Tang, Y.-Y.; Huang, H.-L.; Xie, M.; Zou, W.; Wang, C.-Y.; Zhang, P.; Tang, X.-Q. BDNF/TrkB Pathway Mediates the Antidepressant-Like Role of H2S in CUMS-Exposed Rats by Inhibition of Hippocampal ER Stress. NeuroMolecular Med. 2018, 20, 252–261. [Google Scholar] [CrossRef]
- Cortese-Krott, M.M.; Kuhnle, G.G.C.; Dyson, A.; Fernandez, B.O.; Grman, M.; Dumond, J.F.; Barrow, M.P.; McLeod, G.; Nakagawa, H.; Ondrias, K.; et al. Key bioactive reaction products of the NO/H2S interaction are S/N-hybrid species, polysulfides, and nitroxyl. Proc. Natl. Acad. Sci. USA 2015, 112, E4651–E4660. [Google Scholar] [CrossRef] [PubMed]
- Cortese-Krott, M.M.; Pullmann, D.; Feelisch, M. Nitrosopersulfide (SSNO − ) targets the Keap-1/Nrf2 redox system. Pharmacol. Res. 2016, 113, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Greiner, R.; Pálinkás, Z.; Bäsell, K.; Becher, D.; Antelmann, H.; Nagy, P.; Dick, T.P. Polysulfides Link H2S to Protein Thiol Oxidation. Antioxidants Redox Signal. 2013, 19, 1749–1765. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wu, L. Trend in H2S Biology and Medicine Research—A Bibliometric Analysis. Molecules 2017, 22, 2087. [Google Scholar] [CrossRef]
- Pozsgai, G.; Payrits, M.; Sághy, É.; Sebestyén-Bátai, R.; Steen, E.; Szőke, É.; Sándor, Z.; Solymár, M.; Garami, A.; Orvos, P.; et al. Analgesic effect of dimethyl trisulfide in mice is mediated by TRPA1 and sst 4 receptors. Nitric Oxide 2017, 65, 10–21. [Google Scholar] [CrossRef]
- Dombi, Á.; Sánta, C.; Bátai, I.Z.; Kormos, V.; Kecskés, A.; Tékus, V.; Pohóczky, K.; Bölcskei, K.; Pintér, E.; Pozsgai, G. Dimethyl Trisulfide Diminishes Traumatic Neuropathic Pain Acting on TRPA1 Receptors in Mice. Int. J. Mol. Sci. 2021, 22, 3363. [Google Scholar] [CrossRef]
- Kimura, Y.; Mikami, Y.; Osumi, K.; Tsugane, M.; Oka, J.; Kimura, H. Polysulfides are possible H 2 S-derived signaling molecules in rat brain. FASEB J. 2013, 27, 2451–2457. [Google Scholar] [CrossRef]
- Bátai, I.Z.; Horváth, Á.; Pintér, E.; Helyes, Z.; Pozsgai, G. Role of Transient Receptor Potential Ankyrin 1 Ion Channel and Somatostatin sst4 Receptor in the Antinociceptive and Anti-inflammatory Effects of Sodium Polysulfide and Dimethyl Trisulfide. Front. Endocrinol. 2018, 9, 55. [Google Scholar] [CrossRef]
- Voineskos, D.; Daskalakis, Z.J.; Blumberger, D.M. Management of Treatment-Resistant Depression: Challenges and Strategies. Neuropsychiatr. Dis. Treat. 2020, ume 16, 221–234. [Google Scholar] [CrossRef]
- Kormos, V.; Gáspár, L.; Kovács, L.Á.; Farkas, J.; Gaszner, T.; Csernus, V.; Balogh, A.; Hashimoto, H.; Reglődi, D.; Helyes, Z.; et al. Reduced response to chronic mild stress in PACAP mutant mice is associated with blunted FosB expression in limbic forebrain and brainstem centers. Neuroscience 2016, 330, 335–358. [Google Scholar] [CrossRef]
- Ruilian, L.; Honglin, Q.; Jun, X.; Jianxin, L.; Qingyun, B.; Yilin, C.; Haifeng, M. H2S-mediated aerobic exercise antagonizes the hippocampal inflammatory response in CUMS-depressed mice. J. Affect. Disord. 2021, 283, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-L.; Xie, B.; Zhang, C.; Xu, K.-L.; Niu, Y.-Y.; Tang, X.-Q.; Zhang, P.; Zou, W.; Hu, B.; Tian, Y. Antidepressant-like and anxiolytic-like effects of hydrogen sulfide in behavioral models of depression and anxiety. Behav. Pharmacol. 2013, 24, 590–597. [Google Scholar] [CrossRef]
- Kedia, S.; Chattarji, S. Marble burying as a test of the delayed anxiogenic effects of acute immobilisation stress in mice. J. Neurosci. Methods 2014, 233, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Njung'E, K.; Handley, S.L. Evaluation of marble-burying behavior as a model of anxiety. Pharmacol. Biochem. Behav. 1991, 38, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.L.; Gardner, J.M.; Karmel, B.Z.; Kim, S.-Y. Rating scale measures are associated with Noldus EthoVision-XT video tracking of behaviors of children on the autism spectrum. Mol. Autism 2014, 5, 15–15. [Google Scholar] [CrossRef]
- S.S. Shapiro, M.B. Wilk, An Analysis of Variance Test for Normality (Complete Samples), (2024).
- Göntér, K.; Dombi, Á.; Kormos, V.; Pintér, E.; Pozsgai, G. Examination of the Effect of Dimethyl Trisulfide in Acute Stress Mouse Model with the Potential Involvement of the TRPA1 Ion Channel. Int. J. Mol. Sci. 2024, 25, 7701. [Google Scholar] [CrossRef]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef]
- Boleij, H.; Willems, J.; Leijten, M.; Klooster, J.V.; Lesscher, H.; Kirchhoff, S.; Lavrijsen, M.; Arndt, S.S.; Ohl, F. Chronic social stress does not affect behavioural habituation in male CD1 mice. Behav. Brain Res. 2014, 273, 34–44. [Google Scholar] [CrossRef]
- Harris, R.B.S. Chronic and acute effects of stress on energy balance: are there appropriate animal models? Am. J. Physiol. Integr. Comp. Physiol. 2015, 308, R250–R265. [Google Scholar] [CrossRef]
- Rabasa, C.; Pastor-Ciurana, J.; Delgado-Morales, R.; Gómez-Román, A.; Carrasco, J.; Gagliano, H.; García-Gutiérrez, M.S.; Manzanares, J.; Armario, A. Evidence against a critical role of CB1 receptors in adaptation of the hypothalamic–pituitary–adrenal axis and other consequences of daily repeated stress. Eur. Neuropsychopharmacol. 2015, 25, 1248–1259. [Google Scholar] [CrossRef]
- Farkas, J.; Kovács, L.Á.; Gáspár, L.; Nafz, A.; Gaszner, T.; Ujvári, B.; Kormos, V.; Csernus, V.; Hashimoto, H.; Reglődi, D.; et al. Construct and face validity of a new model for the three-hit theory of depression using PACAP mutant mice on CD1 background. Neuroscience 2017, 354, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Spina, M.; Merlo-Pich, E.; Chan, R.K.W.; Basso, A.M.; Rivier, J.; Vale, W.; Koob, G.F. Appetite-Suppressing Effects of Urocortin, a CRF-Related Neuropeptide. Science 1996, 273, 1561–1564. [Google Scholar] [CrossRef] [PubMed]
- Landini, L.; de Araujo, D.S.M.; Titiz, M.; Geppetti, P.; Nassini, R.; De Logu, F. TRPA1 Role in Inflammatory Disorders: What Is Known So Far? Int. J. Mol. Sci. 2022, 23, 4529. [Google Scholar] [CrossRef] [PubMed]
- Naert, R.; López-Requena, A.; Talavera, K. TRPA1 Expression and Pathophysiology in Immune Cells. Int. J. Mol. Sci. 2021, 22, 11460. [Google Scholar] [CrossRef]
- Szabó, K.; Kemény, Á.; Balázs, N.; Khanfar, E.; Sándor, Z.; Boldizsár, F.; Gyulai, R.; Najbauer, J.; Pintér, E.; Berki, T. Presence of TRPA1 Modifies CD4+/CD8+ T Lymphocyte Ratio and Activation. Pharmaceuticals 2022, 15, 57. [Google Scholar] [CrossRef]
- Szabó, K.; Makkai, G.; Konkoly, J.; Kormos, V.; Gaszner, B.; Berki, T.; Pintér, E. TRPA1 Covalent Ligand JT010 Modifies T Lymphocyte Activation. Biomolecules 2024, 14, 632. [Google Scholar] [CrossRef]
- Russo, D.; Tringali, G.; Ragazzoni, E.; Maggiano, N.; Menini, E.; Vairano, M.; Preziosi, P.; Navarra, P. Evidence That Hydrogen Sulphide Can Modulate Hypothalamo-Pituitary-Adrenal Axis Function: In Vitro and In Vivo Studies in the Rat. J. Neuroendocr. 2000, 12, 225–233. [Google Scholar] [CrossRef]
- Hassan, E.; Kahilo, K.; Kamal, T.; El-Neweshy, M.; Hassan, M. Protective effect of diallyl sulfide against lead-mediated oxidative damage, apoptosis and down-regulation of CYP19 gene expression in rat testes. Life Sci. 2019, 226, 193–201. [Google Scholar] [CrossRef]
- Wang, H.; Shi, X.; Qiu, M.; Lv, S.; Liu, H. Hydrogen Sulfide Plays an Important Protective Role through Influencing Endoplasmic Reticulum Stress in Diseases. Int. J. Biol. Sci. 2020, 16, 264–271. [Google Scholar] [CrossRef]
- Sodhi, R.K.; Singh, R.; Bansal, Y.; Bishnoi, M.; Parhar, I.; Kuhad, A.; Soga, T. Intersections in Neuropsychiatric and Metabolic Disorders: Possible Role of TRPA1 Channels. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef]
- Renard, C.E.; Dailly, E.; David, D.J.; Hascoet, M.; Bourin, M. Monoamine metabolism changes following the mouse forced swimming test but not the tail suspension test. Fundam. Clin. Pharmacol. 2003, 17, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Júnior, E.D.D.S.; Da Silva, A.V.; Da Silva, K.R.; Haemmerle, C.A.; Batagello, D.S.; Da Silva, J.M.; Lima, L.B.; Da Silva, R.J.; Diniz, G.B.; Sita, L.V.; et al. The centrally projecting Edinger–Westphal nucleus—I: Efferents in the rat brain. J. Chem. Neuroanat. 2015, 68, 22–38. [Google Scholar] [CrossRef] [PubMed]
- van der Doelen, R.H.; Robroch, B.; Arnoldussen, I.A.; Schulpen, M.; Homberg, J.R.; Kozicz, T. Serotonin and urocortin 1 in the dorsal raphe and Edinger–Westphal nuclei after early life stress in serotonin transporter knockout rats. Neuroscience 2017, 340, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Waselus, M.; Nazzaro, C.; Valentino, R.J.; Van Bockstaele, E.J. Stress-Induced Redistribution of Corticotropin-Releasing Factor Receptor Subtypes in the Dorsal Raphe Nucleus. Biol. Psychiatry 2009, 66, 76–83. [Google Scholar] [CrossRef]
- Craske, M.G.; Meuret, A.E.; Ritz, T.; Treanor, M.; Dour, H.J. Treatment for Anhedonia: A Neuroscience Driven Approach. Depression Anxiety 2016, 33, 927–938. [Google Scholar] [CrossRef]
- Yakovleva, O.; Bogatova, K.; Mukhtarova, R.; Yakovlev, A.; Shakhmatova, V.; Gerasimova, E.; Ziyatdinova, G.; Hermann, A.; Sitdikova, G. Hydrogen Sulfide Alleviates Anxiety, Motor, and Cognitive Dysfunctions in Rats with Maternal Hyperhomocysteinemia via Mitigation of Oxidative Stress. Biomolecules 2020, 10, 995. [Google Scholar] [CrossRef]
- Chen, M.; Pritchard, C.; Fortune, D.; Kodi, P.; Grados, M. Hydrogen sulfide: a target to modulate oxidative stress and neuroplasticity for the treatment of pathological anxiety. Expert Rev. Neurother. 2019, 20, 109–121. [Google Scholar] [CrossRef]
- Prenoveau, J.M.; Zinbarg, R.E.; Craske, M.G.; Mineka, S.; Griffith, J.W.; Epstein, A.M. Testing a hierarchical model of anxiety and depression in adolescents: A tri-level model. J. Anxiety Disord. 2010, 24, 334–344. [Google Scholar] [CrossRef]
- Vose, L.R.; Stanton, P.K. Synaptic Plasticity, Metaplasticity and Depression. Curr. Neuropharmacol. 2017, 15, 71–86. [Google Scholar] [CrossRef]
- Sha, Z.; Xu, J.; Li, N.; Li, O. Regulatory Molecules of Synaptic Plasticity in Anxiety Disorder. Int. J. Gen. Med. 2023, ume 16, 2877–2886. [Google Scholar] [CrossRef]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: new insights from stress and rapid-acting antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. ∆FosB: A transcriptional regulator of stress and antidepressant responses. Eur. J. Pharmacol. 2015, 753, 66–72. [Google Scholar] [CrossRef] [PubMed]
- E.J. Nestler, M.B. Kelz, J. Chen, DFosB: a molecular mediator of long-term neural and behavioral plasticity, (1999).
- Perrotti, L.I.; Hadeishi, Y.; Ulery, P.G.; Barrot, M.; Monteggia, L.; Duman, R.S.; Nestler, E.J. Induction of ΔFosB in Reward-Related Brain Structures after Chronic Stress. J. Neurosci. 2004, 24, 10594–10602. [Google Scholar] [CrossRef] [PubMed]
- Sterrenburg, L.; Gaszner, B.; Boerrigter, J.; Santbergen, L.; Bramini, M.; Elliott, E.; Chen, A.; Peeters, B.W.M.M.; Roubos, E.W.; Kozicz, T. Chronic Stress Induces Sex-Specific Alterations in Methylation and Expression of Corticotropin-Releasing Factor Gene in the Rat. PLOS ONE 2011, 6, e28128. [Google Scholar] [CrossRef]
- Vialou, V.; Thibault, M.; Kaska, S.; Cooper, S.; Gajewski, P.; Eagle, A.; Mazei-Robison, M.; Nestler, E.J.; Robison, A. Differential induction of FosB isoforms throughout the brain by fluoxetine and chronic stress. Neuropharmacology 2015, 99, 28–37. [Google Scholar] [CrossRef]
- Virk, H.S.; Rekas, M.Z.; Biddle, M.S.; Wright, A.K.A.; Sousa, J.; Weston, C.A.; Chachi, L.; Roach, K.M.; Bradding, P. Validation of antibodies for the specific detection of human TRPA1. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Patil, M.; Jeske, N.; Akopian, A. Transient receptor potential V1 regulates activation and modulation of transient receptor potential A1 by Ca2+. Neuroscience 2010, 171, 1109–1119. [Google Scholar] [CrossRef]
- Rojas-Galvan, N.S.; Ciotu, C.I.; Heber, S.; Fischer, M.J. Correlation of TRPA1 RNAscope and Agonist Responses. J. Histochem. Cytochem. 2024, 72, 275–287. [Google Scholar] [CrossRef]
- Kozicz, T.; Tilburg-Ouwens, D.; Faludi, G.; Palkovits, M.; Roubos, E. Gender-related urocortin 1 and brain-derived neurotrophic factor expression in the adult human midbrain of suicide victims with major depression. Neuroscience 2008, 152, 1015–1023. [Google Scholar] [CrossRef]
- Bouwknecht, J.A.; Spiga, F.; Staub, D.R.; Hale, M.W.; Shekhar, A.; Lowry, C.A. Differential effects of exposure to low-light or high-light open-field on anxiety-related behaviors: Relationship to c-Fos expression in serotonergic and non-serotonergic neurons in the dorsal raphe nucleus. Brain Res. Bull. 2007, 72, 32–43. [Google Scholar] [CrossRef]
- Kozicz, T. The missing link; the significance of urocortin 1/urocortin 2 in the modulation of the dorsal raphe serotoninergic system. Mol. Psychiatry 2009, 15, 340–341. [Google Scholar] [CrossRef] [PubMed]
- Neufeld-Cohen, A.; Tsoory, M.M.; Evans, A.K.; Getselter, D.; Gil, S.; Lowry, C.A.; Vale, W.W.; Chen, A. A triple urocortin knockout mouse model reveals an essential role for urocortins in stress recovery. Proc. Natl. Acad. Sci. 2010, 107, 19020–19025. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt, J.C.; Vaughan, J.; Arias, C.; Rissman, R.A.; Vale, W.W.; Sawchenko, P.E. Urocortin Expression in Rat Brain: Evidence Against a Pervasive Relationship of Urocortin-Containing Projections With Targets Bearing Type 2 CRF Receptors. J. Comp. Neurol. 1999, 415, 285–312. [Google Scholar] [CrossRef]
- Kozicz, T. On the role of urocortin 1 in the non-preganglionic Edinger–Westphal nucleus in stress adaptation. Gen. Comp. Endocrinol. 2007, 153, 235–240. [Google Scholar] [CrossRef]
- de Bortoli, V.C.; Yamashita, P.S.d.M.; Zangrossi, H. 5-HT1A and 5-HT2A receptor control of a panic-like defensive response in the rat dorsomedial hypothalamic nucleus. J. Psychopharmacol. 2013, 27, 1116–1123. [Google Scholar] [CrossRef]
- Zuniga, A.; E Ryabinin, A. Involvement of Centrally Projecting Edinger–Westphal Nucleus Neuropeptides in Actions of Addictive Drugs. Brain Sci. 2020, 10, 67. [Google Scholar] [CrossRef]
- Kirouac, G.J. Placing the paraventricular nucleus of the thalamus within the brain circuits that control behavior. Neurosci. Biobehav. Rev. 2015, 56, 315–329. [Google Scholar] [CrossRef]
- Ma, J.; du Hoffmann, J.; Kindel, M.; Beas, B.S.; Chudasama, Y.; Penzo, M.A. Divergent projections of the paraventricular nucleus of the thalamus mediate the selection of passive and active defensive behaviors. Nat. Neurosci. 2021, 24, 1429–1440. [Google Scholar] [CrossRef]
- Singewald, G.M.; Rjabokon, A.; Singewald, N.; Ebner, K. The Modulatory Role of the Lateral Septum on Neuroendocrine and Behavioral Stress Responses. Neuropsychopharmacology 2010, 36, 793–804. [Google Scholar] [CrossRef]
- Hammack, S.E.; Cheung, J.; Rhodes, K.M.; Schutz, K.C.; Falls, W.A.; Braas, K.M.; May, V. Chronic stress increases pituitary adenylate cyclase-activating peptide (PACAP) and brain-derived neurotrophic factor (BDNF) mRNA expression in the bed nucleus of the stria terminalis (BNST): Roles for PACAP in anxiety-like behavior. Psychoneuroendocrinology 2009, 34, 833–843. [Google Scholar] [CrossRef]
- Kiss, A.; Aguilera, G. Regulation of the hypothalamic pituitary adrenal axis during chronic stress: responses to repeated intraperitoneal hypertonic saline injection. Brain Res. 1993, 630, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Verkuyl, J.M.; Hemby, S.E.; Joëls, M. Chronic stress attenuates GABAergic inhibition and alters gene expression of parvocellular neurons in rat hypothalamus. Eur. J. Neurosci. 2004, 20, 1665–1673. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.R.; Cullinan, W.E.; Herman, J.P. Organization and regulation of paraventricular nucleus glutamate signaling systems: N-methyl-D-aspartate receptors. J. Comp. Neurol. 2005, 484, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Flak, J.N.; Ostrander, M.M.; Tasker, J.G.; Herman, J.P. Chronic stress-induced neurotransmitter plasticity in the PVN. J. Comp. Neurol. 2009, 517, 156–165. [Google Scholar] [CrossRef]
- Crestani, C.C.; Alves, F.H.; Gomes, F.V.; Resstel, L.B.; Correa, F.M.; Herman, J.P. Mechanisms in the Bed Nucleus of the Stria Terminalis Involved in Control of Autonomic and Neuroendocrine Functions: A Review. Curr. Neuropharmacol. 2013, 11, 141–159. [Google Scholar] [CrossRef]
- Herman, J.P.; Figueiredo, H.; Mueller, N.K.; Ulrich-Lai, Y.; Ostrander, M.M.; Choi, D.C.; Cullinan, W.E. Central mechanisms of stress integration: hierarchical circuitry controlling hypothalamo–pituitary–adrenocortical responsiveness. Front. Neuroendocr. 2003, 24, 151–180. [Google Scholar] [CrossRef]
- V. Grinevich, A. Fournier, G. Pelletier, Effects of pituitary adenylate cyclase-activating polypeptide Ž PACAP. on corticotropin-releasing hormone Ž CRH. gene expression in the rat hypothalamic paraventricular nucleus, (1997).
- Choi, D.C.; Evanson, N.K.; Furay, A.R.; Ulrich-Lai, Y.M.; Ostrander, M.M.; Herman, J.P. The Anteroventral Bed Nucleus of the Stria Terminalis Differentially Regulates Hypothalamic-Pituitary-Adrenocortical Axis Responses to Acute and Chronic Stress. Endocrinology 2007, 149, 818–826. [Google Scholar] [CrossRef]
- Deji, C.; Yan, P.; Ji, Y.; Yan, X.; Feng, Y.; Liu, J.; Liu, Y.; Wei, S.; Zhu, Y.; Lai, J. The Basolateral Amygdala to Ventral Hippocampus Circuit Controls Anxiety-Like Behaviors Induced by Morphine Withdrawal. Front. Cell. Neurosci. 2022, 16, 894886. [Google Scholar] [CrossRef]
- Kovács, L.Á.; Füredi, N.; Ujvári, B.; Golgol, A.; Gaszner, B. Age-Dependent FOSB/ΔFOSB Response to Acute and Chronic Stress in the Extended Amygdala, Hypothalamic Paraventricular, Habenular, Centrally-Projecting Edinger-Westphal, and Dorsal Raphe Nuclei in Male Rats. Front. Aging Neurosci. 2022, 14, 862098. [Google Scholar] [CrossRef]
- Sun, H.-J.; Wu, Z.-Y.; Nie, X.-W.; Bian, J.-S. Role of Hydrogen Sulfide and Polysulfides in Neurological Diseases: Focus on Protein S-Persulfidation. Curr. Neuropharmacol. 2021, 19, 868–884. [Google Scholar] [CrossRef]
- Kimura, H. Physiological role of hydrogen sulfide and polysulfide in the central nervous system. Neurochem. Int. 2013, 63, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bian, J.-S. Hydrogen Sulfide: A Neuromodulator and Neuroprotectant in the Central Nervous System. ACS Chem. Neurosci. 2014, 5, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Hydrogen Sulfide (H2S) and Polysulfide (H2Sn) Signaling: The First 25 Years. Biomolecules 2021, 11, 896. [Google Scholar] [CrossRef] [PubMed]
- De Silva, D.; Lee, S.; Duke, A.; Angalakurthi, S.; Chou, C.-E.; Ebrahimpour, A.; Thompson, D.E.; Petrikovics, I. Intravascular Residence Time Determination for the Cyanide Antidote Dimethyl Trisulfide in Rat by Using Liquid-Liquid Extraction Coupled with High Performance Liquid Chromatography. J. Anal. Methods Chem. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Dong, X.; Kiss, L.; Petrikovics, I.; Thompson, D.E. Reaction of Dimethyl Trisulfide with Hemoglobin. Chem. Res. Toxicol. 2017, 30, 1661–1663. [Google Scholar] [CrossRef]
- Manandhar, E.; Maslamani, N.; Petrikovics, I.; Rockwood, G.A.; Logue, B.A. Determination of dimethyl trisulfide in rabbit blood using stir bar sorptive extraction gas chromatography-mass spectrometry. J. Chromatogr. A 2016, 1461, 10–17. [Google Scholar] [CrossRef]
- Petrikovics, I.; Kiss, L.; Chou, C.-E.; Ebrahimpour, A.; Kovács, K.; Kiss, M.; Logue, B.; Chan, A.; Manage, A.B.W.; Budai, M.; et al. Antidotal efficacies of the cyanide antidote candidate dimethyl trisulfide alone and in combination with cobinamide derivatives. Toxicol. Mech. Methods 2019, 29, 438–444. [Google Scholar] [CrossRef]
- A Rockwood, G.; E Thompson, D.; Petrikovics, I. Dimethyl trisulfide. Toxicol. Ind. Heal. 2016, 32, 2009–2016. [Google Scholar] [CrossRef]
- Warnakula, I.K.; Ebrahimpour, A.; Li, S.Y.; Ralalage, R.D.G.; Hewa-Rahinduwage, C.C.; Kiss, M.; Rios, C.T.; Kelley, K.D.; Whiteman, A.C.; Thompson, D.E.; et al. Evaluation of the Long-Term Storage Stability of the Cyanide Antidote: Dimethyl Trisulfide and Degradation Product Identification. ACS Omega 2020, 5, 27171–27179. [Google Scholar] [CrossRef]
- Konkoly, J.; Kormos, V.; Gaszner, B.; Sándor, Z.; Kecskés, A.; Alomari, A.; Szilágyi, A.; Szilágyi, B.; Zelena, D.; Pintér, E. The Role of TRPA1 Channels in the Central Processing of Odours Contributing to the Behavioural Responses of Mice. Pharmaceuticals 2021, 14, 1336. [Google Scholar] [CrossRef]
- Olah, E.; Rumbus, Z.; Kormos, V.; Tekus, V.; Pakai, E.; Wilson, H.V.; Fekete, K.; Solymar, M.; Kelava, L.; Keringer, P.; et al. The Hypothermic Effect of Hydrogen Sulfide Is Mediated by the Transient Receptor Potential Ankyrin-1 Channel in Mice. Pharmaceuticals 2021, 14, 992. [Google Scholar] [CrossRef] [PubMed]
- Cano, G.; Hernan, S.L.; Sved, A.F. Centrally Projecting Edinger-Westphal Nucleus in the Control of Sympathetic Outflow and Energy Homeostasis. Brain Sci. 2021, 11, 1005. [Google Scholar] [CrossRef] [PubMed]
- Donner, N.; Handa, R. Estrogen receptor beta regulates the expression of tryptophan-hydroxylase 2 mRNA within serotonergic neurons of the rat dorsal raphe nuclei. Neuroscience 2009, 163, 705–718. [Google Scholar] [CrossRef]
- Leite, C.; Madeira, M.D.; Sá, S.I. Effects of sex steroids and estrogen receptor agonists on the expression of estrogen receptor alpha in the principal division of the bed nucleus of the stria terminalis of female rats. Brain Res. 2014, 1582, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Süß, T.; Oliveira, V.E.d.M.; Neumann, I.D.; Bludau, A. Neurobiology of the lateral septum: regulation of social behavior. Trends Neurosci. 2022, 45, 27–40. [Google Scholar] [CrossRef]
- Sheng, Z.; Kawano, J.; Yanai, A.; Fujinaga, R.; Tanaka, M.; Watanabe, Y.; Shinoda, K. Expression of estrogen receptors (α, β) and androgen receptor in serotonin neurons of the rat and mouse dorsal raphe nuclei; sex and species differences. Neurosci. Res. 2004, 49, 185–196. [Google Scholar] [CrossRef]
- Stern, J.; Zhang, W. Preautonomic neurons in the paraventricular nucleus of the hypothalamus contain estrogen receptor β. Brain Res. 2003, 975, 99–109. [Google Scholar] [CrossRef]
- Tian, Z.; Wang, Y.; Zhang, N.; Guo, Y.-Y.; Feng, B.; Liu, S.-B.; Zhao, M.-G. Estrogen receptor GPR30 exerts anxiolytic effects by maintaining the balance between GABAergic and glutamatergic transmission in the basolateral amygdala of ovariectomized mice after stress. Psychoneuroendocrinology 2013, 38, 2218–2233. [Google Scholar] [CrossRef]
Figure 1.
Experimental timeline of control (A) and chronic stress-exposed (B) animals. IHC: immunohistochemistry; ACTH: adrenocorticotropic hormone; CORT: corticosterone; CUMS: chronic unpredictable mild stress; DMTS: dimethyl trisulfide; ELISA: enzyme-linked immunosorbent assay.
Figure 1.
Experimental timeline of control (A) and chronic stress-exposed (B) animals. IHC: immunohistochemistry; ACTH: adrenocorticotropic hormone; CORT: corticosterone; CUMS: chronic unpredictable mild stress; DMTS: dimethyl trisulfide; ELISA: enzyme-linked immunosorbent assay.
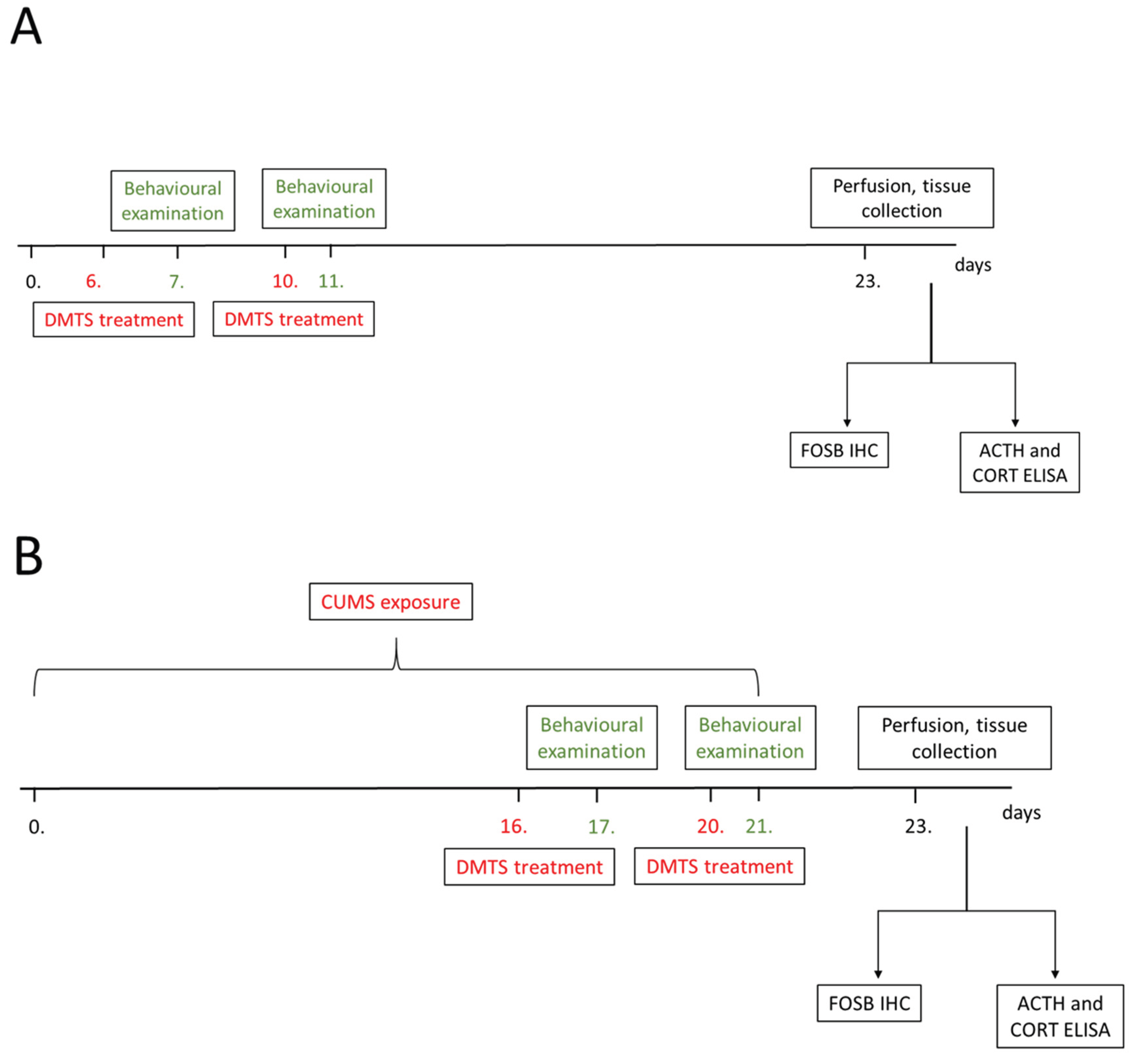
Figure 2.
Establishing the dose of dimethyl trisulfide (DMTS) via open field test. Symbols represent from left to right untreated (naïve), vehicle-treated, 30 mg/kg (DMTS30) and 40 mg/kg DMTS-treated (DMTS40) Trpa1 WT mice. Distance travelled (A) and mobility time (B) are plotted on the left y axes (One-way analysis of variance; **p < 0.01; n = 8-10 per group).
Figure 2.
Establishing the dose of dimethyl trisulfide (DMTS) via open field test. Symbols represent from left to right untreated (naïve), vehicle-treated, 30 mg/kg (DMTS30) and 40 mg/kg DMTS-treated (DMTS40) Trpa1 WT mice. Distance travelled (A) and mobility time (B) are plotted on the left y axes (One-way analysis of variance; **p < 0.01; n = 8-10 per group).
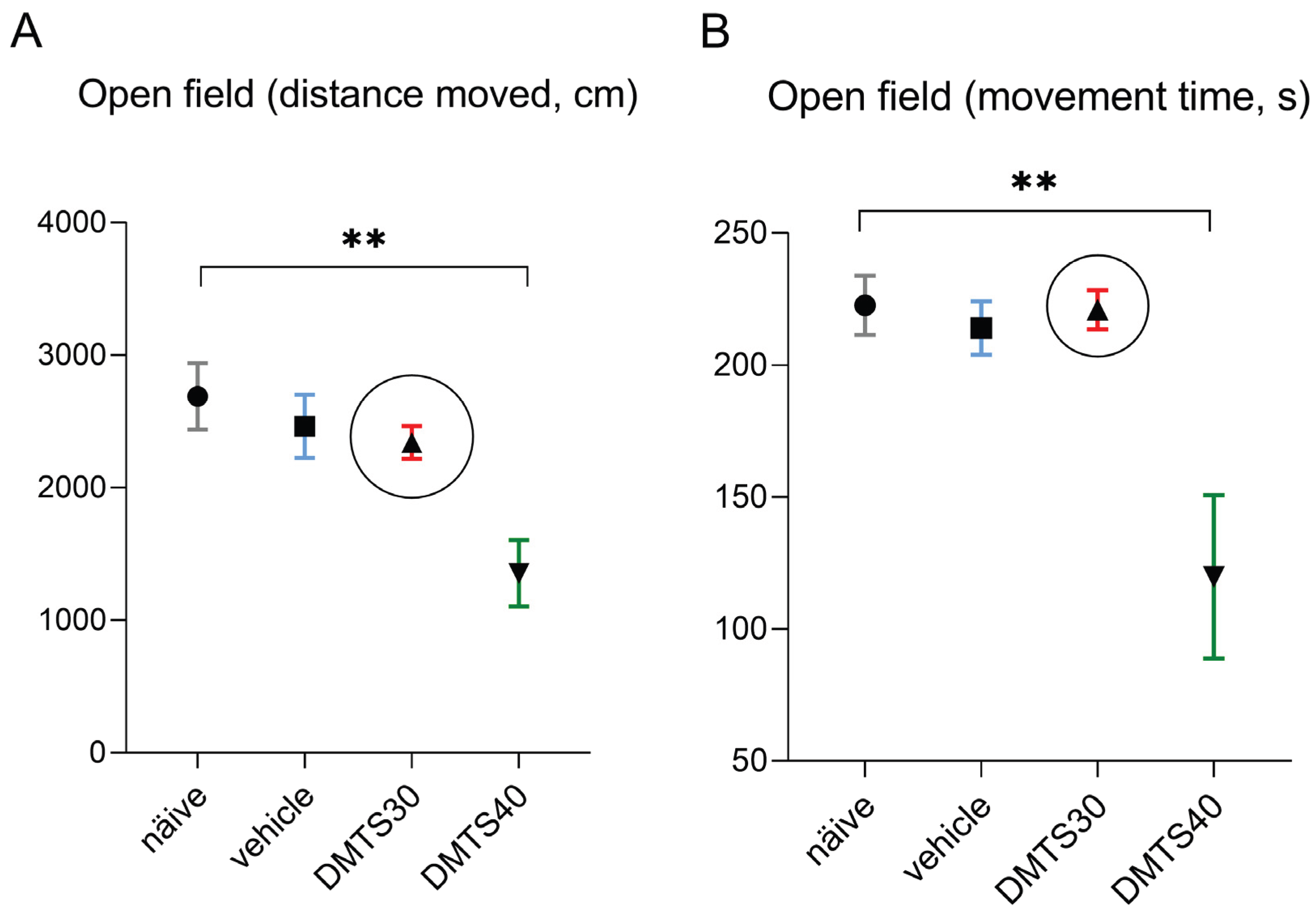
Figure 3.
The efficacy of chronic unpredictable mild stress (CUMS) exposure and dimethyl trisulfide (DMTS) treatment. We found that body weight change (A), relative adrenal weight (C) and relative thymus weight (D) mirrored somatic changes induced by chronic stress. Panel (B) presents absolute body weight of mice at the end of the in vivo experiment. We assessed the hypothalamic-pituitary-adrenal axis activity by determining serum corticosterone (E) and adrenocorticotropic hormone (ACTH) concentration (F) (Three-way analysis of variance followed by a Tukey’s post hoc test; *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001; n = 8-16 per group). Only the relevant significance levels have been indicated to facilitate clarity.
Figure 3.
The efficacy of chronic unpredictable mild stress (CUMS) exposure and dimethyl trisulfide (DMTS) treatment. We found that body weight change (A), relative adrenal weight (C) and relative thymus weight (D) mirrored somatic changes induced by chronic stress. Panel (B) presents absolute body weight of mice at the end of the in vivo experiment. We assessed the hypothalamic-pituitary-adrenal axis activity by determining serum corticosterone (E) and adrenocorticotropic hormone (ACTH) concentration (F) (Three-way analysis of variance followed by a Tukey’s post hoc test; *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001; n = 8-16 per group). Only the relevant significance levels have been indicated to facilitate clarity.
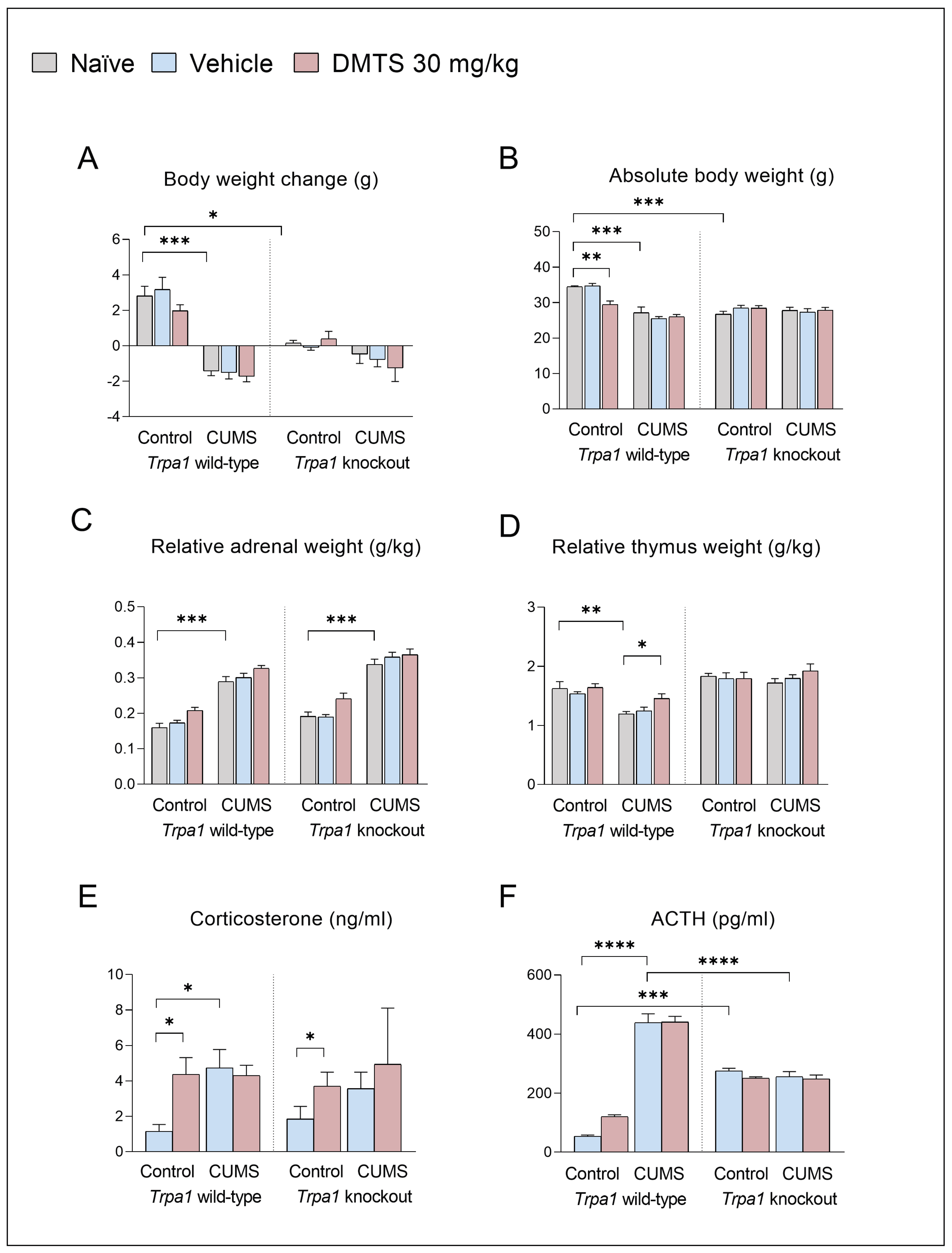
Figure 4.
Effect of chronic unpredictable mild stress (CUMS) and dimethyl trisulfide (DMTS) treatment on anxiety and depression-related behaviour. In order to assess the level of anxiety, we evaluated the time spent (A) and the distance travelled (B) in the periphery part of the arena in the open field test, and we counted the number of hidden marbles in the marble buried test (C). To detect depression-like behaviour, we determined the presence of anhedonia with the sucrose preference test (D), moreover, we assessed inactive duration in the forced swim test (E) and tail suspension test (F) (Three-way analysis of variance followed by a Tukey’s post hoc test; *p < 0.05; **p < 0.01; ***p < 0.001; n = 8-16 per group). Only the relevant significance levels have been indicated to facilitate clarity.
Figure 4.
Effect of chronic unpredictable mild stress (CUMS) and dimethyl trisulfide (DMTS) treatment on anxiety and depression-related behaviour. In order to assess the level of anxiety, we evaluated the time spent (A) and the distance travelled (B) in the periphery part of the arena in the open field test, and we counted the number of hidden marbles in the marble buried test (C). To detect depression-like behaviour, we determined the presence of anhedonia with the sucrose preference test (D), moreover, we assessed inactive duration in the forced swim test (E) and tail suspension test (F) (Three-way analysis of variance followed by a Tukey’s post hoc test; *p < 0.05; **p < 0.01; ***p < 0.001; n = 8-16 per group). Only the relevant significance levels have been indicated to facilitate clarity.
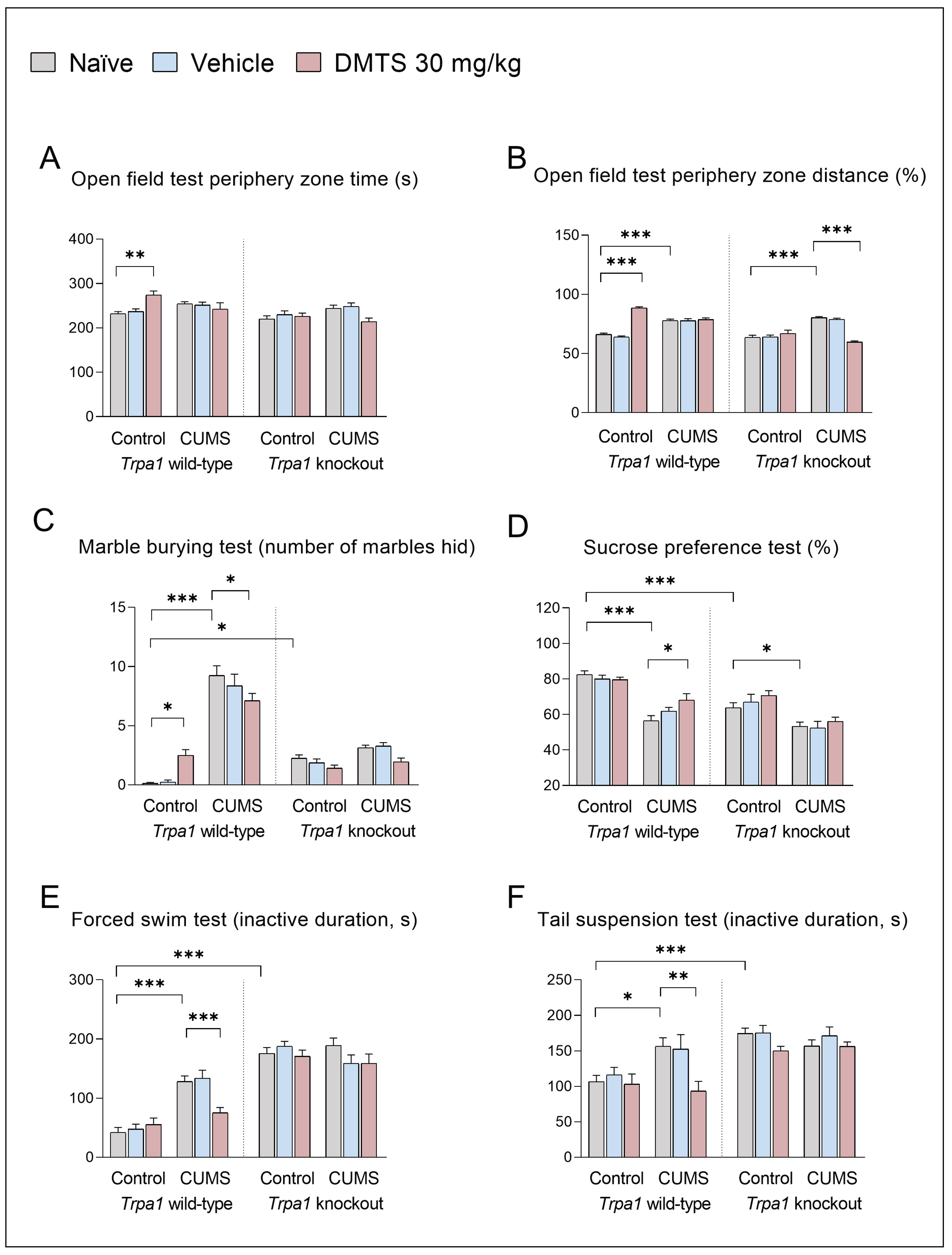
Figure 5.
FOSB immunohistochemistry in centrally projecting Edinger-Westphal nucleus (EWcp), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05; **p < 0.01; n = 5-6 per group. Black line: scale bar 200 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
Figure 5.
FOSB immunohistochemistry in centrally projecting Edinger-Westphal nucleus (EWcp), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05; **p < 0.01; n = 5-6 per group. Black line: scale bar 200 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.

Figure 6.
FOSB immunohistochemistry in dorsal raphe nucleus (DR), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05; **p < 0.01; n = 5-6 per group. Black line: scale bar 200 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
Figure 6.
FOSB immunohistochemistry in dorsal raphe nucleus (DR), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05; **p < 0.01; n = 5-6 per group. Black line: scale bar 200 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
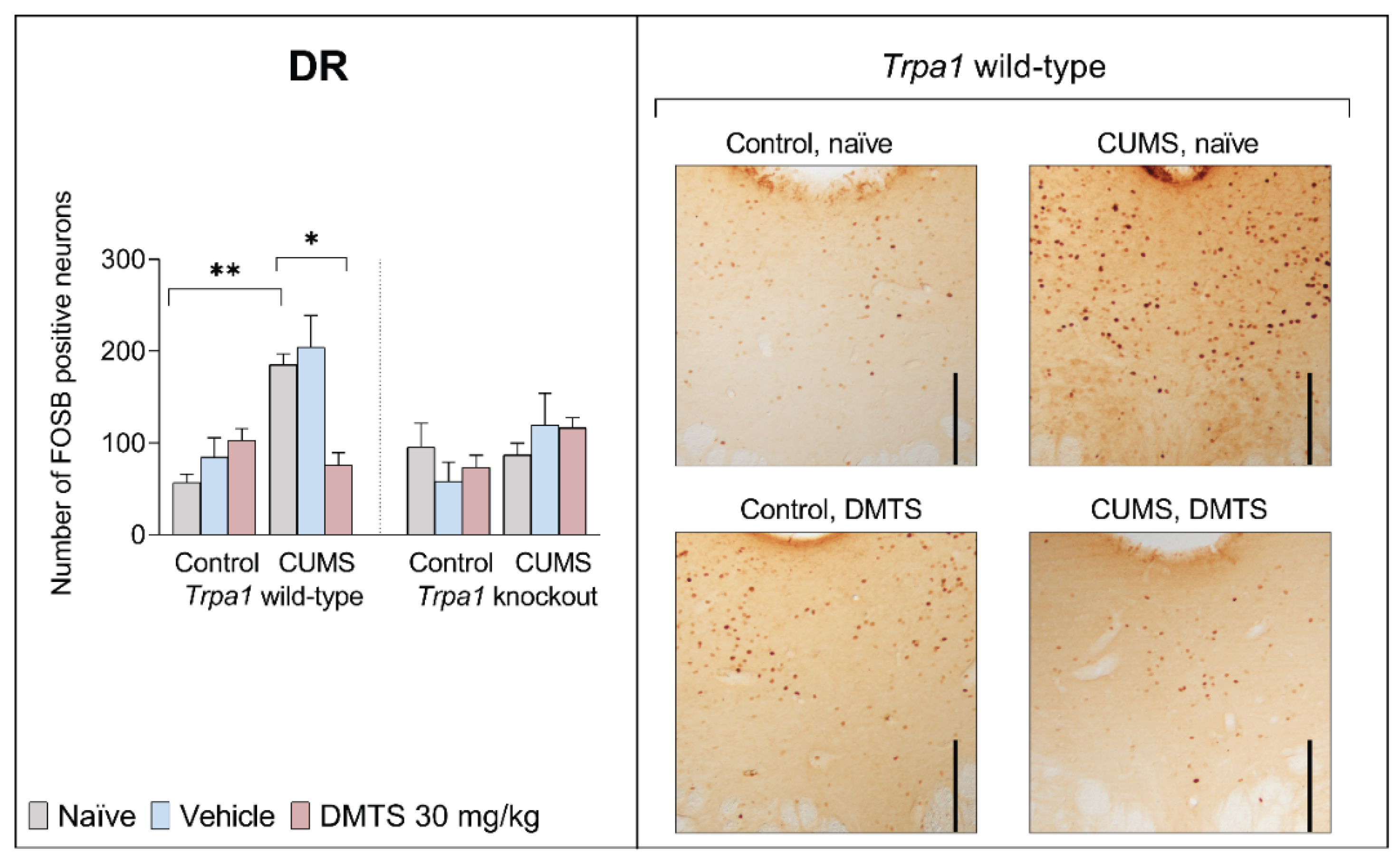
Figure 7.
FOSB immunohistochemistry in periaqueductal gray matter (PAG), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: ***p < 0.001; n = 5-6 per group. Black line: scale bar 500 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
Figure 7.
FOSB immunohistochemistry in periaqueductal gray matter (PAG), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: ***p < 0.001; n = 5-6 per group. Black line: scale bar 500 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
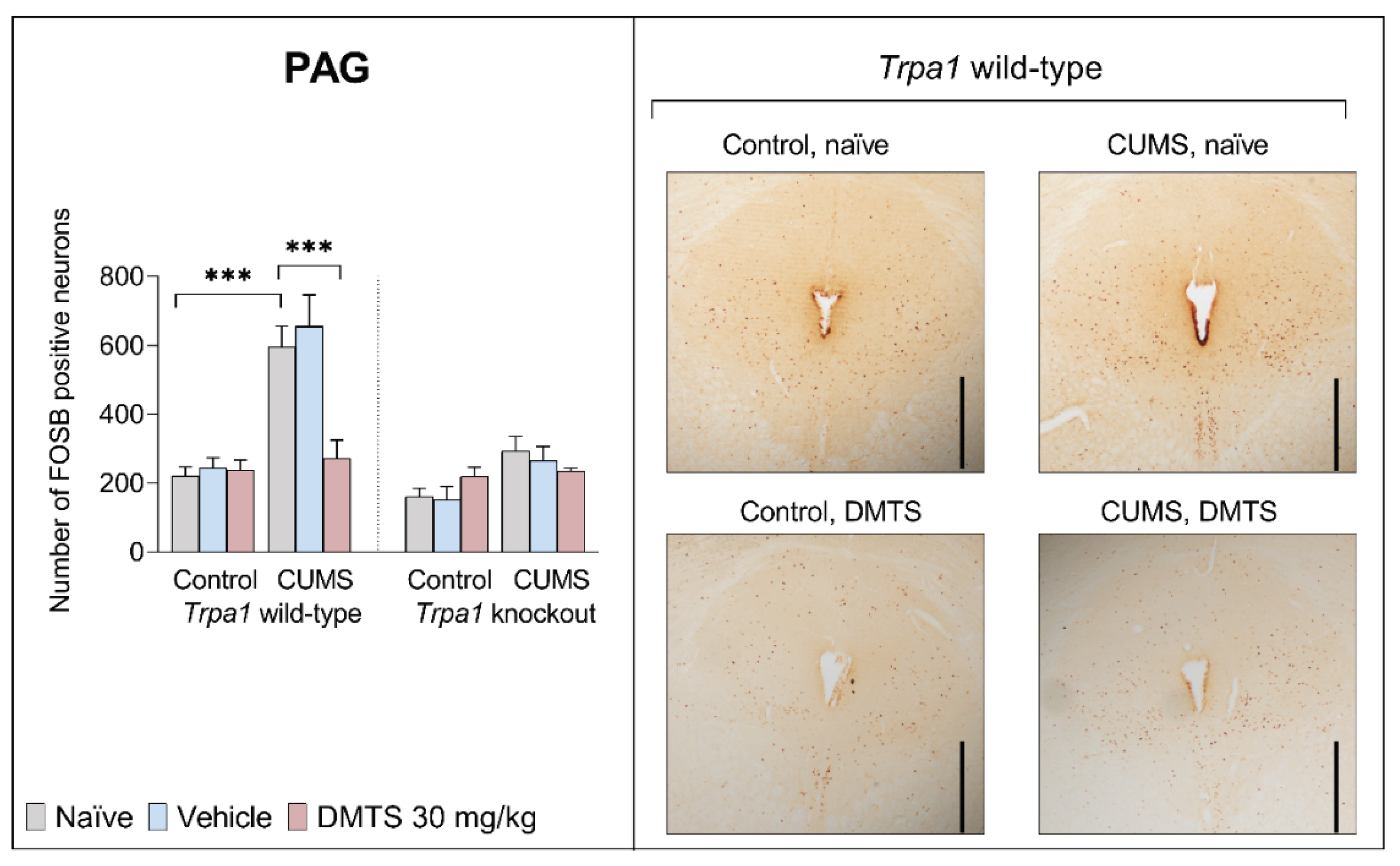
Figure 8.
FOSB immunohistochemistry in paraventricular nucleus of the thalamus (PVT), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05; n = 5-6 per group. Black line: scale bar 200 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
Figure 8.
FOSB immunohistochemistry in paraventricular nucleus of the thalamus (PVT), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05; n = 5-6 per group. Black line: scale bar 200 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
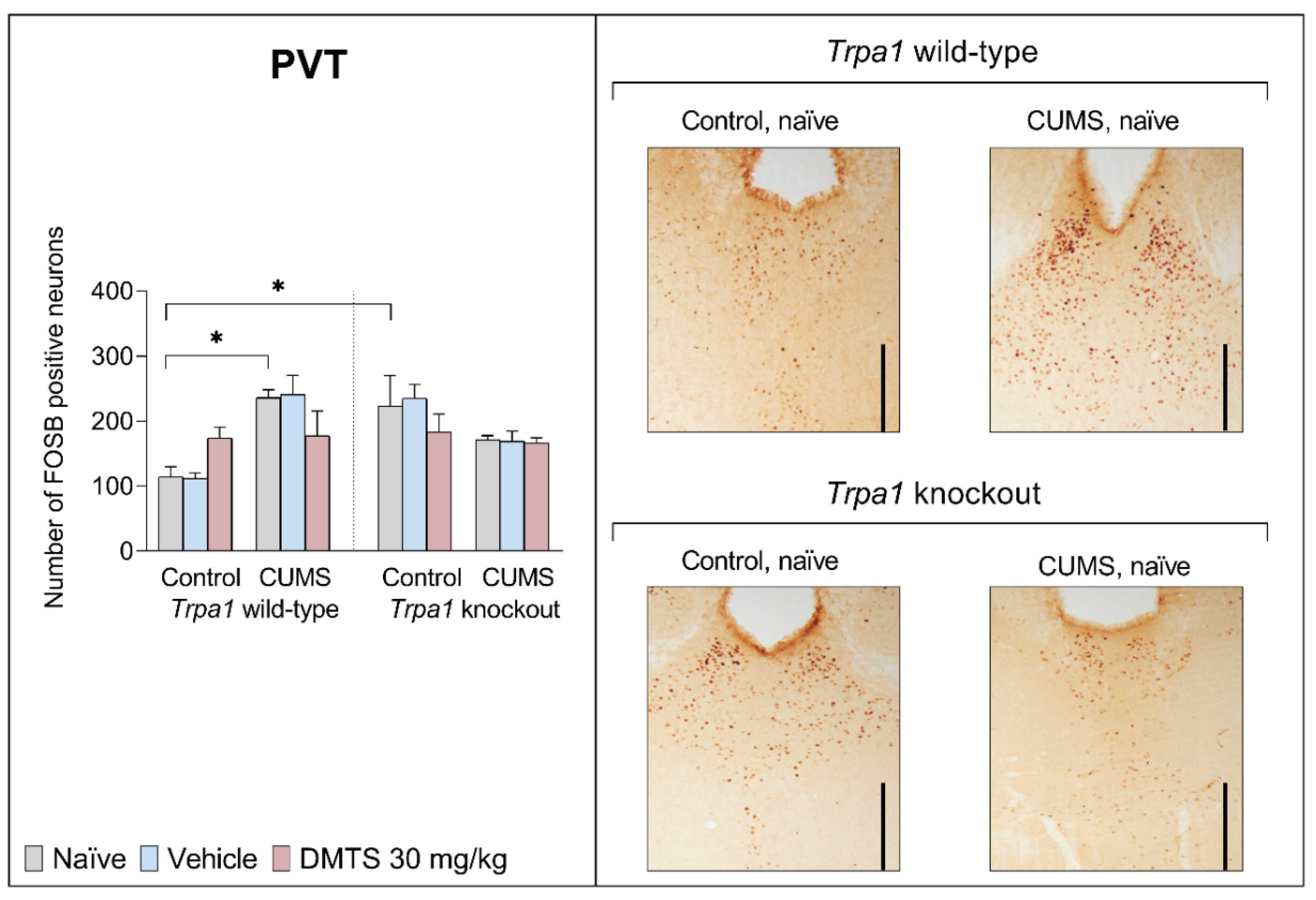
Figure 9.
FOSB immunohistochemistry in lateral septal nucleus (LS), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05, **p < 0.01; n = 5-6 per group. Black line: scale bar 500 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
Figure 9.
FOSB immunohistochemistry in lateral septal nucleus (LS), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05, **p < 0.01; n = 5-6 per group. Black line: scale bar 500 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
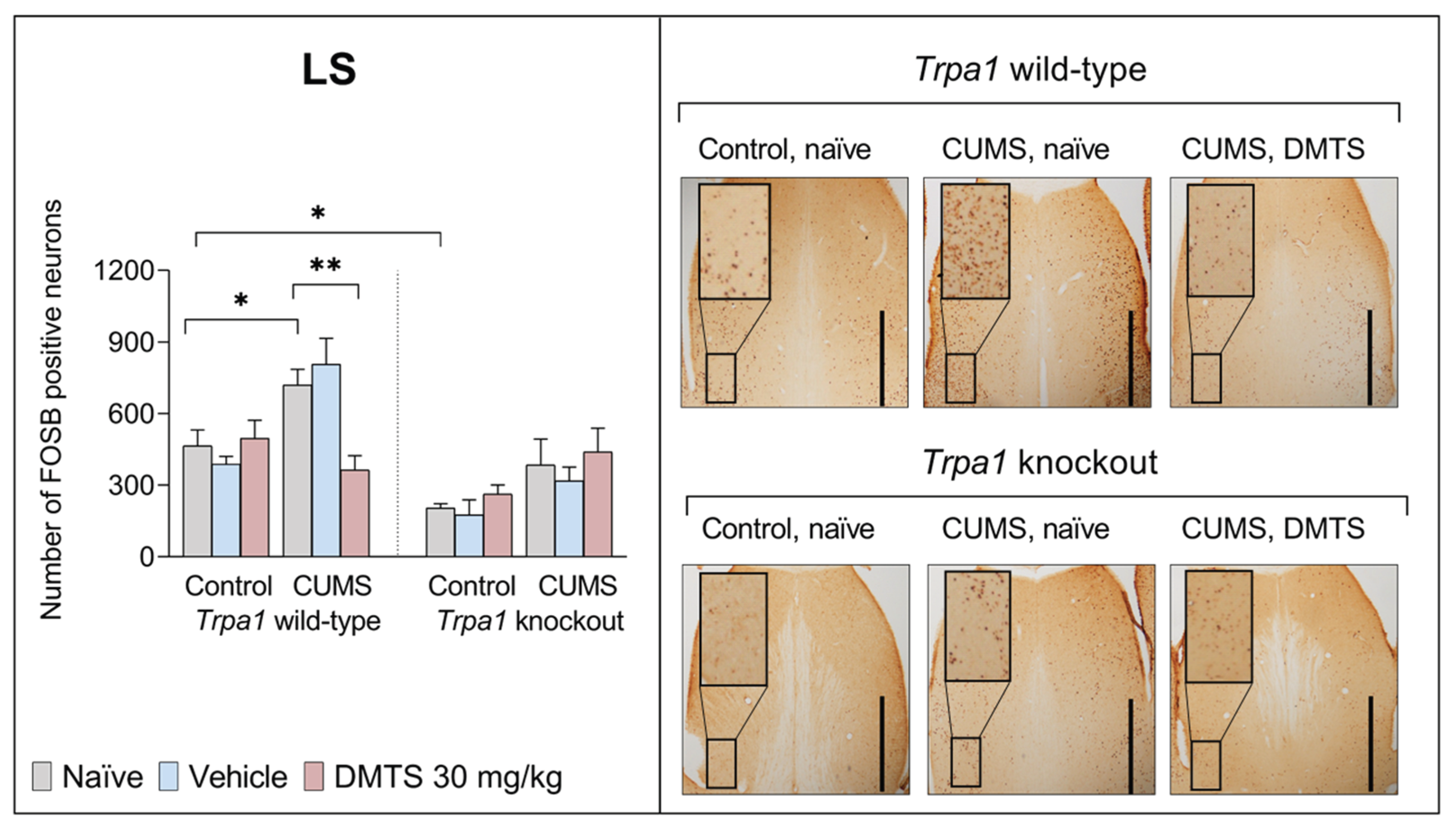
Figure 10.
FOSB immunohistochemistry in paraventricular nucleus of the hypothalamus (PVN), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: **p < 0.01, ***p < 0.001; n = 5-6 per group. Black line: scale bar 200 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
Figure 10.
FOSB immunohistochemistry in paraventricular nucleus of the hypothalamus (PVN), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: **p < 0.01, ***p < 0.001; n = 5-6 per group. Black line: scale bar 200 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
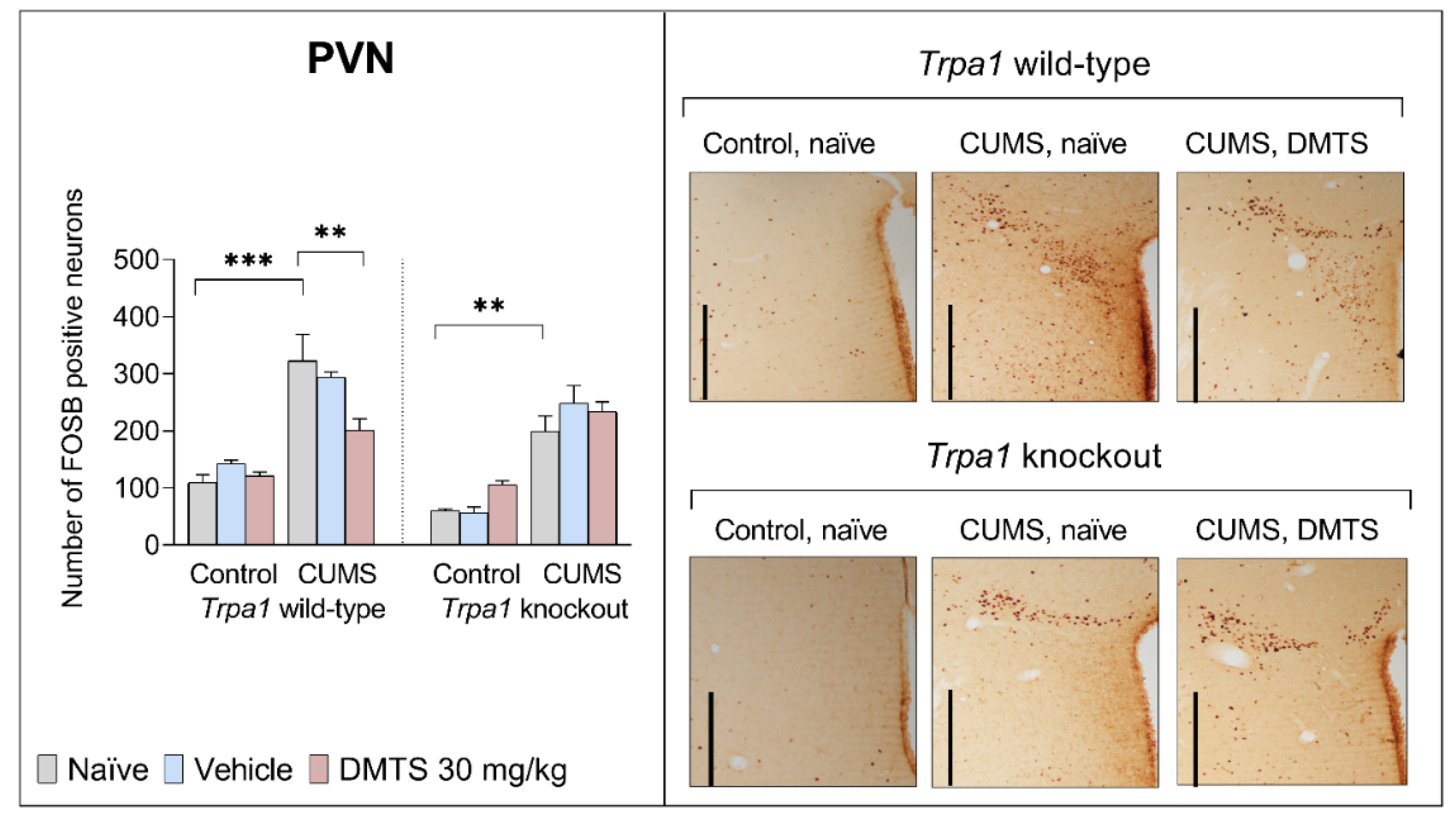
Figure 11.
FOSB immunohistochemistry in bed nucleus of the stria terminalis (BST), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05, **p < 0.01; n = 5-6 per group. Black line: scale bar 500 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
Figure 11.
FOSB immunohistochemistry in bed nucleus of the stria terminalis (BST), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: *p < 0.05, **p < 0.01; n = 5-6 per group. Black line: scale bar 500 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
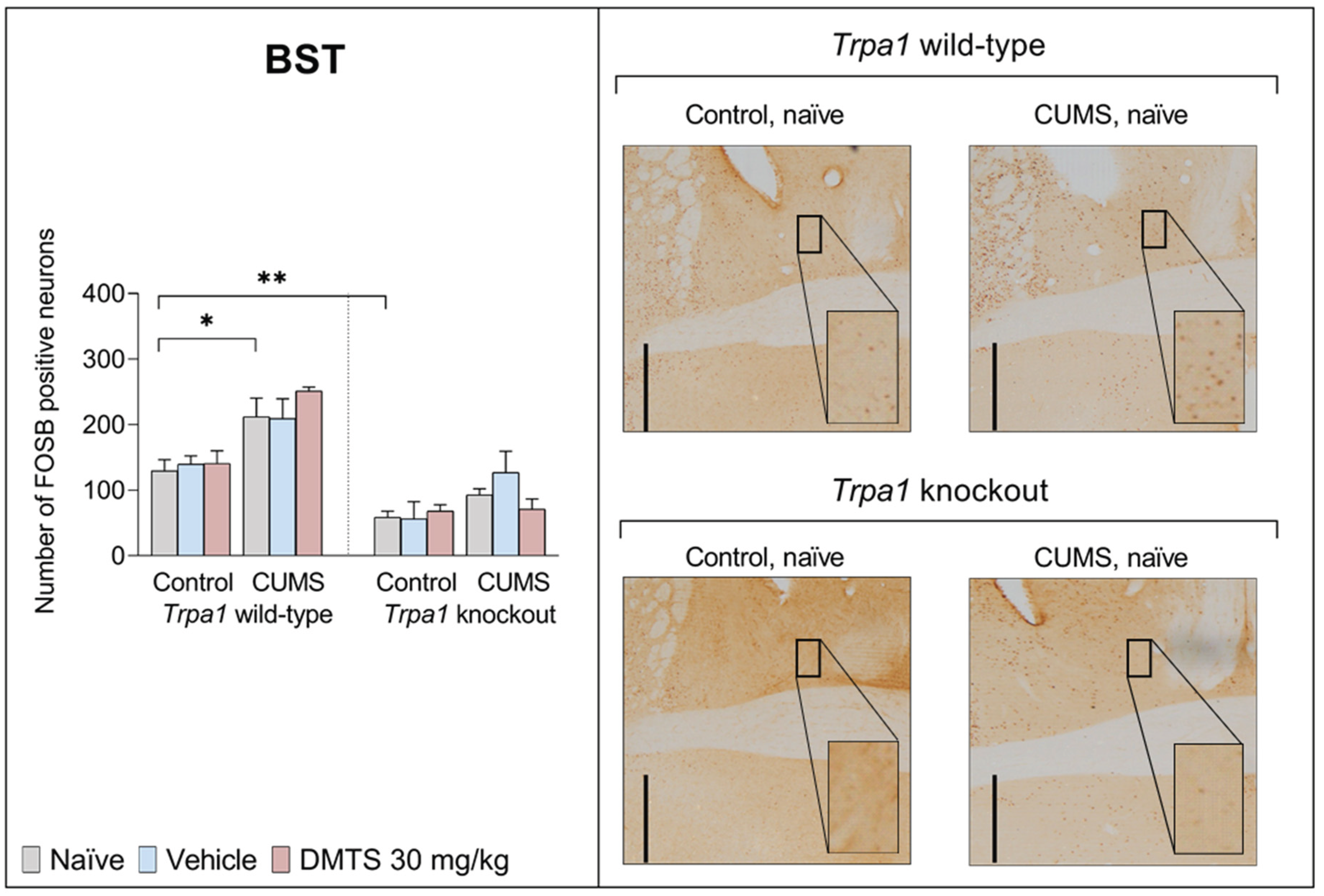
Figure 12.
FOSB immunohistochemistry in basolateral amygdala (BLA), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: n = 5-6 per group. Black line: scale bar 500 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
Figure 12.
FOSB immunohistochemistry in basolateral amygdala (BLA), representative micrographs. Bar graphs show the number of FOSB immunoreactive neurons in each group. Three-way analysis of variance followed by a Tukey’s post hoc test: n = 5-6 per group. Black line: scale bar 500 m. Control = non-stressed; CUMS = chronic unpredictable mild stress; DMTS = dimethyl trisulfide. Only the relevant significance levels have been indicated to facilitate clarity.
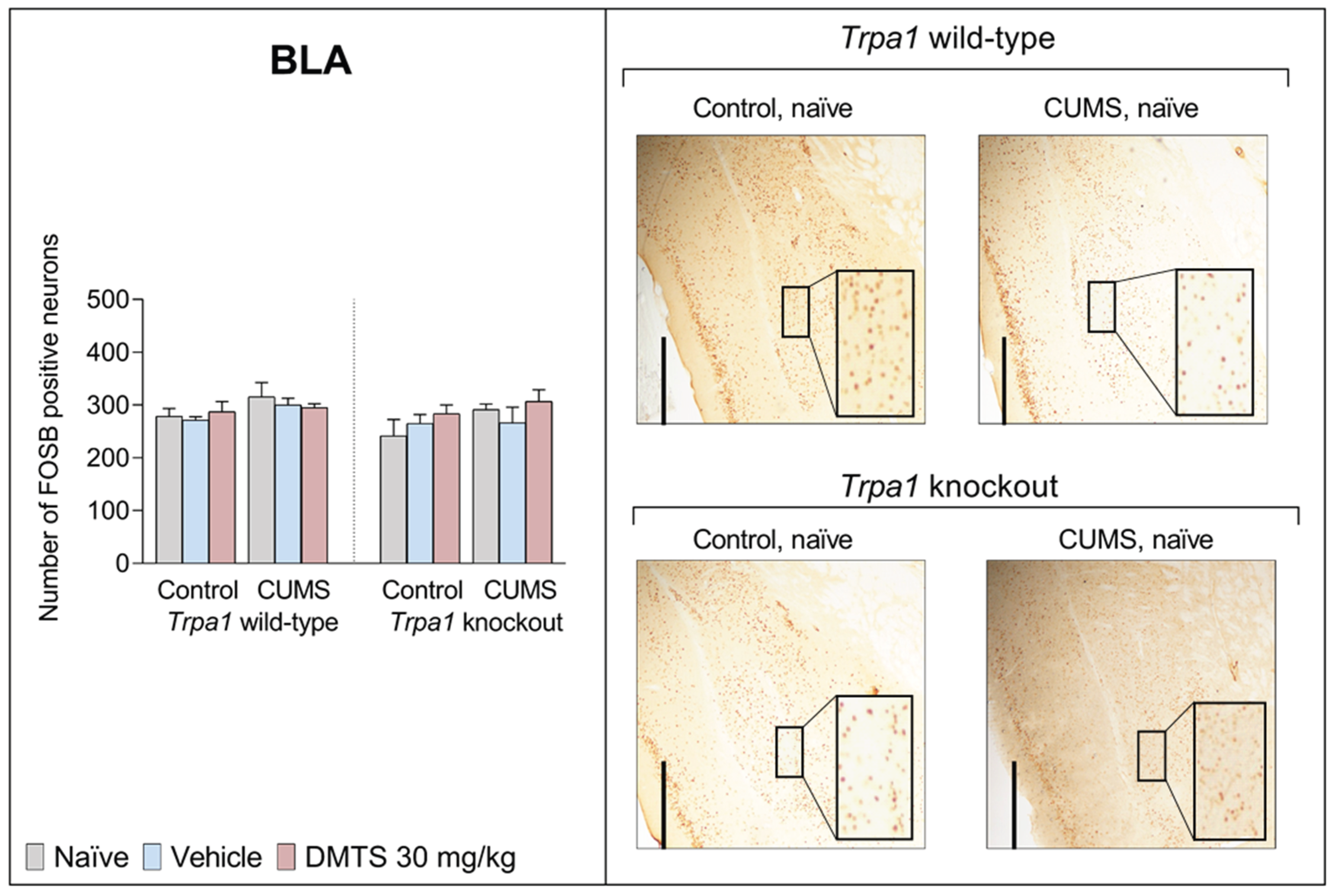
Figure 13.
Summary of possible mechanisms underlying the effects of chronic stress and DMTS treatment on depression-like behaviour and anxiety. Red arrows = inhibition; green arrows = stimulation; blue arrow = modulation; HPA = hypothalamic-pituitary-adrenal axis; TRPA1 = transient receptor potential ankyrin 1; ACTH = adrenocorticotropic hormone; CRH = corticotropin-releasing hormone; DMTS = dimethyl trisulfide; CRF = corticotropin-releasing factor; 5-HT1A = serotonin 1A; UCN1 = urocortin 1; PAG = periaqueductal gray matter; DR = dorsal raphe nucleus; EWcp = centrally projecting Edinger-Westphal nucleus; BST = bed nucleus of the stria terminalis; BLA = basolateral amygdala; LS = lateral septum; PVT = paraventricular nucleus of the thalamus; PVN = paraventricular nucleus of the hypothalamus.
Figure 13.
Summary of possible mechanisms underlying the effects of chronic stress and DMTS treatment on depression-like behaviour and anxiety. Red arrows = inhibition; green arrows = stimulation; blue arrow = modulation; HPA = hypothalamic-pituitary-adrenal axis; TRPA1 = transient receptor potential ankyrin 1; ACTH = adrenocorticotropic hormone; CRH = corticotropin-releasing hormone; DMTS = dimethyl trisulfide; CRF = corticotropin-releasing factor; 5-HT1A = serotonin 1A; UCN1 = urocortin 1; PAG = periaqueductal gray matter; DR = dorsal raphe nucleus; EWcp = centrally projecting Edinger-Westphal nucleus; BST = bed nucleus of the stria terminalis; BLA = basolateral amygdala; LS = lateral septum; PVT = paraventricular nucleus of the thalamus; PVN = paraventricular nucleus of the hypothalamus.
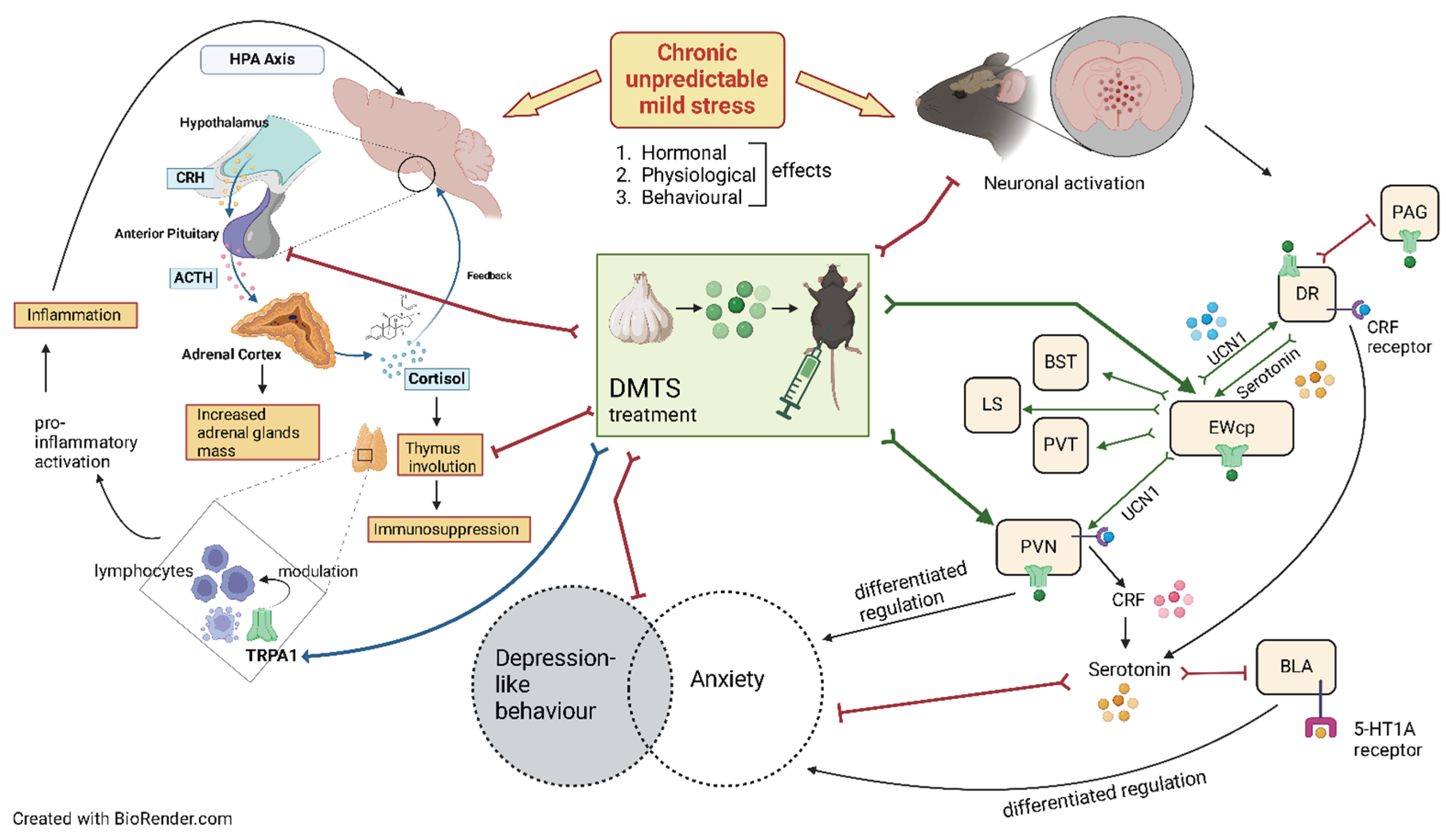

Table 1.
The table summarises the treatment groups and the element numbers. Trpa1: transient receptor potential ankyrin 1; WT: wild type; KO: knockout; CUMS: chronic unpredictable mild stress; DMTS30: dimethyl trisulfide 30 mg/kg dose; naïve: non-treated.
Table 1.
The table summarises the treatment groups and the element numbers. Trpa1: transient receptor potential ankyrin 1; WT: wild type; KO: knockout; CUMS: chronic unpredictable mild stress; DMTS30: dimethyl trisulfide 30 mg/kg dose; naïve: non-treated.
| Trpa1 WT | |||||
| Non-stressed | CUMS | ||||
| naïve | vehicle-treated | DMTS30-treated | naïve | vehicle-treated | DMTS30-treated |
| 8 | 16 | 16 | 8 | 16 | 16 |
| Trpa1 KO | |||||
| Non-stressed | CUMS | ||||
| naïve | vehicle-treated | DMTS30-treated | naïve | vehicle-treated | DMTS30-treated |
| 8 | 16 | 16 | 8 | 16 | 16 |
Table 2.
Timeline of chronic unpredictable mild stress protocol with mid-day and overnight stressors. DARK = dark room; GH = group holding; REST = restraint stress; SHAKE = shaker stress; ISOL = social isolation; TILT = tilted cage; WM: body weight measurement; WET = wet bedding.
Table 2.
Timeline of chronic unpredictable mild stress protocol with mid-day and overnight stressors. DARK = dark room; GH = group holding; REST = restraint stress; SHAKE = shaker stress; ISOL = social isolation; TILT = tilted cage; WM: body weight measurement; WET = wet bedding.
| No. of days | Mid-day stressors | Overnight stressors |
| 1. | WM, REST | WET |
| 2. | DARK | ISOL |
| 3. | TILT | GH |
| 4. | DARK | WET |
| 5. | SHAKE | GH |
| 6. | TILT | ISOL |
| 7. | REST | GH |
| 8. | TS, TILT | WET |
| 9. | SHAKE | ISOL |
| 10. | REST | WET |
| 11. | TILT | GH |
| 12. | SHAKE | WET |
| 13. | DARK | GH |
| 14. | TILT | WET |
| 15. | WM, REST | GH |
| 16. | TILT | WET |
| 17. | REST | GH |
| 18. | DARK | WET |
| 19. | TILT | GH |
| 20. | DARK | WET |
| 21. | REST | ISOL |
Table 3.
Summary of the statistics of physical parameters, endocrine and behavioural data by three-way analysis of variance (ANOVA). ACTH = adrenocorticotropic hormone; CUMS = chronic unpredictable mild stress. Significant values are highlighted in bold.
Table 3.
Summary of the statistics of physical parameters, endocrine and behavioural data by three-way analysis of variance (ANOVA). ACTH = adrenocorticotropic hormone; CUMS = chronic unpredictable mild stress. Significant values are highlighted in bold.
| Main effects | Interactions | |||||||
| Variable | Treatment | Genotype | CUMS | Treatment x genotype | CUMS x treatment | CUMS x genotype | CUMS x treatment x genotype | |
| MBT, number of marbles hid | F2,149 | 0.9901 | 57.66 | 186.3 | 2.325 | 8.423 | 110.6 | 5.556 |
| p | 0.374 | <0.0001 | <0.0001 | 0.1013 | 0.0003 | <0.0001 | 0.0047 | |
| OFT, periphery zone time | F2,72 | 0.1927 | 13.57 | 1.654 | 4.723 | 45583 | 0.7645 | 0.3862 |
| p | 0.8252 | 0.0004 | 0.204 | 0.0118 | 0.0002 | 0.3859 | 0.6815 | |
| OFT, periphery zone distance | F2,72 | 3.657 | 30498 | 46.64 | 93.32 | 61.24 | 2.189 | 0.3012 |
| p | 0.0333 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.1455 | 0.7413 | |
| FST, activity | F2,89 | 3.923 | 148.1 | 18.73 | 0.7528 | 1.389 | 53.56 | 5.322 |
| p | 0.0233 | <0.0001 | <0.0001 | 0.474 | 0.2559 | <0.0001 | 0.007 | |
| TST, activity | F2,73 | 5.814 | 45322 | 1.963 | 0.9288 | 0.8436 | 4.864 | 4.135 |
| p | 0.0045 | <0.0001 | 0.1678 | 0.3997 | 0.4366 | 0.0323 | 0.0222 | |
| SPT, sucrose preference | F2,78 | 3.172 | 13516 | 31079 | 0.02048 | 45304 | 2.555 | 3.496 |
| p | 0.0474 | <0.0001 | <0.0001 | 0.9797 | 0.3292 | 0.1146 | 0.0359 | |
| Serum ACTH | F 1,16 | 0.7709 | 0.3015 | 243.3 | 5.494 | 1.222 | 275.1 | 3.534 |
| p | 0.3929 | 0.5905 | <0.0001 | 0.0323 | 0.2854 | <0.0001 | 0.0785 | |
| Serum corticosterone | F1,30 | 4.197 | 0.03008 | 4.84 | 0.02317 | 1.975 | 0.03587 | 1.174 |
| p | 0.0493 | 0.8635 | 0.0357 | 0.88 | 0.1702 | 0.8511 | 0.2872 | |
| Relative adrenal weight | F1,56 | 10.87 | 25.35 | 415.3 | 0.009887 | 1.105 | 2.79 | 0.5117 |
| p | <0.0001 | <0.0001 | <0.0001 | 0.9902 | 0.3383 | 0.1004 | 0.6023 | |
| Relative thymus weight | F2,121 | 2.248 | 55.79 | 9.346 | 0.3015 | 2.179 | 10.35 | 0.01058 |
| p | 0.11 | <0.0001 | 0.0028 | 0.7402 | 0.1176 | 0.0017 | 0.9895 | |
| Change in body weight | F2,107 | 15.02 | 14.43 | 45.22 | 2.162 | 0.969 | 13.44 | 0.7456 |
| p | 0.1215 | 0.0002 | <0.0001 | 0.1201 | 0.3833 | 0.0004 | 0.4773 | |
| Body weight | F2,77 | 2.326 | 13.49 | 50.72 | 5.797 | 3.747 | 43.66 | 3.461 |
| p | 0.1045 | 0.0004 | <0.0001 | 0.0045 | 0.028 | <0.0001 | 0.0364 | |
Table 4.
Summary of statistical analyses of FOSB immunoreactivity in various stress-related brain areas. Analysis was performed by three-way analysis of variance (ANOVA). EWcp = centrally projecting Edinger-Westphal nucleus; LS = lateral septal nucleus; PAG = periaqueductal gray matter; DR = dorsal raphe nucleus; PVN = paraventricular nucleus of the hypothalamus; PVT = paraventricular nucleus of the thalamus; BLA = basolateral amygdala; BST = bed nucleus of the stria terminalis; CUMS = chronic unpredictable mild stress. Significant values are highlighted in bold.
Table 4.
Summary of statistical analyses of FOSB immunoreactivity in various stress-related brain areas. Analysis was performed by three-way analysis of variance (ANOVA). EWcp = centrally projecting Edinger-Westphal nucleus; LS = lateral septal nucleus; PAG = periaqueductal gray matter; DR = dorsal raphe nucleus; PVN = paraventricular nucleus of the hypothalamus; PVT = paraventricular nucleus of the thalamus; BLA = basolateral amygdala; BST = bed nucleus of the stria terminalis; CUMS = chronic unpredictable mild stress. Significant values are highlighted in bold.
| Main effects | Interactions | ||||||||
| Variable | Treatment | Genotype | CUMS | Treatment x genotype | CUMS x treatment | CUMS x genotype | CUMS x treatment x genotype | ||
| EWcp | F2,27 | 2.708 | 14.12 | 12.71 | 11.09 | 1.611 | 22.19 | 3.491 | |
| p | 0.0848 | 0.0008 | 0.0014 | 0.0003 | 0.2182 | <0.0001 | 0.0448 | ||
| LS | F2,27 | 0.5911 | 16.68 | 36.4 | 3.381 | 4.289 | 0.02364 | 4.091 | |
| p | 0.5616 | 0.0004 | <0.0001 | 0.0509 | 0.0255 | 0.8791 | 0.0296 | ||
| PAG | F2,27 | 4.024 | 58.24 | 29.59 | 10.78 | 5.261 | 15.65 | 45378 | |
| p | 0.0311 | <0.0001 | <0.0001 | 0.0005 | 0.0128 | 0.0006 | 0.0555 | ||
| DR | F2,27 | 1.434 | 20.61 | 5.002 | 4.278 | 2.267 | 3.268 | 6.678 | |
| p | 0.2559 | 0.0002 | 0.0338 | 0.0276 | 0.123 | 0.085 | 0.0057 | ||
| PVN | F2,27 | 0.2366 | 166 | 16.27 | 5.995 | 4.281 | 0.03658 | 3.206 | |
| p | 0.7911 | <0.0001 | 0.0005 | 0.0077 | 0.0257 | 0.8499 | 0.0583 | ||
| PVT | F2,27 | 0.4047 | 1.945 | 1.356 | 0.8528 | 0.4163 | 20.27 | 3.702 | |
| p | 0.6717 | 0.1759 | 0.2557 | 0.4387 | 0.6642 | 0.0001 | 0.0397 | ||
| BLA | F2,27 | 1.146 | 3.03 | 2.553 | 0.7379 | 1.137 | 0.04659 | 0.4684 | |
| p | 0.3347 | 0.0945 | 0.1232 | 0.4887 | 0.3375 | 0.8309 | 0.6316 | ||
| BST | F2,27 | 0.2597 | 39.09 | 62.11 | 0.1842 | 1.029 | 6.838 | 2.529 | |
| p | 0.7733 | <0.0001 | <0.0001 | 0.833 | 0.3715 | 0.0158 | 0.1026 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



