
Submitted:
13 May 2025
Posted:
13 May 2025
You are already at the latest version
Abstract
Heavy metal contamination in soil, especially cadmium (Cd) and lead (Pb), poses serious environmental and health risks, particularly in mining regions. While this contamination affects most organisms present in such areas, some filamentous fungi proliferate and immobilize metals in contaminated areas. In this work, six filamentous fungi tolerant to high concentrations of these metals were identified by macroscopic and microscopic morphological characteristics, as well as molecularly, through conserved regions of internal transcribed spacers (ITS). Tolerance to Cd and Pb was evaluated in solid and liquid culture media, and half the maximum inhibitory concentration (IC50) was assessed. Pb tolerance was obtained in Penicillium simplicissimum, Paecilomyces lilacinus, and Rhizopus microsporus (IC50: 3874, 1176, and 211.80 mg/L). Cd tolerance was also detected in Paecilomyces lilacinus, Fusarium oxysporum, Rhizopus microsporus, and Cunninghamella sp. (IC50: 311, 223, 29.25, and 25.18 mg/L). The results indicate that these fungi have evolved effective survival strategies in contaminated environments and highlight their potential for future applications in the bioremediation of multi-metal contaminated soils. This study lays the groundwork for exploring tolerance mechanisms and evaluating the efficacy of native fungal isolates in mitigating heavy metal contamination.
Keywords:
Cadmium
; Lead
; fungus
; tolerance
; contamination
; metals
1. Introduction
Mining is one of the most important activities in terms of economic impact in a vast number of countries, nonetheless, large amounts of residues are generated from gold and silver extraction with mercury and cyanide. Mining residues are accumulated in open spaces and its environmental impact is significant due to the highly toxic concentration of heavy metals, which are disseminated by various means and contaminate anthropic and natural spaces, damaging diverse organisms and biogeochemical cycles [1,2,3].
In Mexico, mining industries are of a great tradition since pre-Hispanic times, located principally in the north and center of the country. In the state of Zacatecas, Mexico, specifically in Concepción del Oro, the main economic activity is the mining of lead, copper, zinc, silver and gold, besides marble, onyx and quartz. In mining residues, known as mining tailings, Cd and Pb concentrations that surpass maximum permissible limits (MPL) have been found (37 and 400 mg/Kg, respectively) [4,5].
Heavy metal contamination is considerably extended throughout nature, and toxicity may affect different organisms and biogeochemical cycles [1]. The main toxicity mechanisms of metals at a molecular level include: 1) Blockage of biomolecules essential functional groups, due to metallic cations affinity to sulfhydryl groups in proteins, denaturing them. 2) Cation displacement in important enzymes like Rubisco, that loses its function when divalent cations such as Co2+, Ni2+, Zn2+, Cd2+ and Pb2+ replace Mg2+. 3) Reactive Oxygen Species (ROS) generation due to Fe2+ or Cu+ auto oxidation, which results in H2O2 and OH radicals that cause irreversible damage to carbohydrates, DNA, proteins and lipids [6].
Cadmium is present in concentrations of 0.1-0.5 mg/Kg in soil, mainly in minerals, and in copper, lead and zinc residues. In soil, Cd is immobilized in organic matter, but its bioavailability remains, especially in conditions of acid pH. Its water mobility occurs as Cd2+ or like soluble complexes with anions and organic matter. Cd toxicity is a result of its capability for damaging DNA and cell membranes, binding to proteins sulfhydryl groups, and protein denaturing. Some microorganisms are resistant to Cd presence through biomineral precipitation such as phosphates, carbonates and sulfides, and few can synthesize CdS nanoparticles [7,8].
Lead contamination is one of the most dangerous and common due to activities like mining and battery manufacturing, being highly toxic and persistent in the environment. Pb2+ can replace Ca2+ in cells, damaging DNA, proteins and cell membranes, in addition to protein synthesis inhibition and ROS generation. Tolerance to Pb in microorganisms has been developed because of ATPase efflux pumps activity and Pb nanoparticle synthesis, as well as immobilizing Pb in the soil by mineral precipitation of pyrophyllite and lead oxalate [8,9].
A great diversity of microorganisms is harbored in the soil, which are crucial for its fertility and biogeochemical cycles; of these microorganisms, 50% are fungi [10,11,12]. Within the most found fungi genera in soil are Aspergillus, Penicillium, Rhizopus and Trichoderma [13]. Fungal communities that flourish in contaminated areas are constantly exposed to high concentrations of xenobiotics, so they have developed a superior tolerance to metals in comparison to bacteria and actinomycetes. It has been shown that some fungi possess the ability to immobilize and degrade toxic compounds to more stable forms through biotransformation, biosorption, biolixiviation, biomineralization, enzyme-catalyzed transformation and toxic elements storage through intracellular accumulation [14,15,16]. Consequently, soil microbiomes play a fundamental role in heavy metal toxicity mitigation in the environment [4,17,18].
It has been suggested that fungi that thrive in contaminated soils have a potential to immobilize metals and be applied for soil remediation [19,20,21,22]. For this reason, the aim of the study was to isolate and identify morphological and molecular filamentous fungi from mining contaminated soils, to evaluate their tolerance to increasing concentrations of cadmium and lead with morphological changes of fungal colonies and to estimate the half-maximum inhibitory concentration of the tolerant isolates.
2. Materials and Methods
Obtention and morphological identification of fungal isolates
Samples of heavy metal contaminated soil were obtained from a community in Concepción del Oro, Zacatecas, México (24°42′N 101°25′W); 57 fungal isolates were obtained and conserved via standard methods (Os-1, Os-2, Os-3, …, Os-56 y Os-57) in an internal collection at Environmental Studies Laboratory, Universidad Autónoma de Aguascalientes, México [4]. The isolates were incubated in PDA at 28 °C in darkness for 7 days, were prepared with lactophenol cotton blue staining and observed through microscope (40x) for morphological identification [23].
Cd and Pb tolerance evaluation
Tolerance to heavy metals of 57 fungal isolates was analyzed by means of mycelium diameter and changes in its morphological characteristics when exposed to increasing concentrations of Cd or Pb. Metal stock solutions of CdCl2 or Pb(NO3)2 were prepared according to Văcar et al. [24] and sterilized under UV light for 30 min. Different metal concentrations were adjusted by adding metal stock solutions to sterile culture media. Concentrations increasing from 1000 to 12,000 mg/L were used for Pb tolerant isolates and from 50 to 1050 mg/L for Cd tolerant isolates. To evaluate isolates growth, PDA media was inoculated with 5 x 105 spores/mL [25] and were incubated for 7 days at 28 °C in the dark. Mycelium diameters were measured and compared against control (PDA without metal). By means of this test, the 6 fungal isolates with the highest tolerance to cadmium and lead were selected for the study.
To determine isolates growth in different metal concentrations mycelium dry weight was measured and half maximal inhibitory concentration (IC50) was calculated for 6 isolates tolerant to high concentrations of Pb or Cd. Inoculum preparation was done according to methodology proposed by Janicki, et al. [25], with modifications. 5 x 105 spores/mL of tolerant fungus were inoculated in 30 mL of PDB and incubated for 24 h at 32 °C in continuous agitation. For each assay, 15% v/v of homogenized inoculum was added to flasks with different metal concentrations and incubated for 24 h at 32 °C in continuous agitation. Mycelia were washed twice with distilled water, filtered through Ahlstrom 54 filter paper and dried for 5 h at 60 °C. Filter paper was weighed and IC50 value was calculated [26].
Standardized PDA (BD Bioxon) and PDB (BD Difco) media, which have a constant pH of 5.6 to 5.8, were used in the experiments. Although the pH was not adjusted after sterilization or during incubation, the same conditions were applied to all media, and all experiments were performed in parallel with controls, minimizing variability due to physicochemical conditions. The metal stock solutions were sterilized with UV light (254 nm) to prevent any alteration in the medium composition.
Molecular identification of tolerant fungi
Monosporic cultures of the 6 tolerant fungi were obtained following methodology proposed by Rangel-Muñoz et al. [27]. Spores were cultured in PDB at 28 °C for 24 h in the dark. Genomic DNA was extracted according to Aljanabi & Martinez [28] with modifications. DNA electrophoresis was performed in 1% agarose and quantified in NanoDrop 2000 (ThermoFisher Scientific) with GeneSnap (SynGene). PCR products were ligated to pJET 1.2 plasmid (ampicillin resistance) and Escherichia coli DH5α strains were transformed via heat shock [29] to increase the number of copies. Transformed colonies were subjected to plasmid extraction (Plasmid Mini-Prep Kit - Column Kit, Jena Bioscence). Internal Transcribed Spacer (ITS) regions were amplified using ITS 4 (5’TCCTCCGCTTATTGATATGC3’) and ITS 5 (5’GGAAGTAAAAGTCGTAACAAGG3’) primers. In order to confirm ITS amplified fragment, a restriction enzyme reaction was performed, and plasmids were sequenced at Biotechnology Institute in Universidad Nacional Autónoma de Mexico. The sequences obtained were assembled with the Seqman program; they were also aligned and compared with the sequences present in the NCBI database using the Basic Local Alignment Search Tool (BLAST).
Statistical analysis
Mean value and standard deviation for 3 replicates of mycelial growth diameter in different concentrations of Cd or Pb and for growth inhibition records were calculated. To compare average growth diminution for each isolate ANOVA and Tukey HSD tests were applied (p<0.05). For IC50 determination, a non-linear regression between mycelial growth and Cd or Pb concentrations was calculated. All statistical analyses were performed with GraphPad Prism 9.0.0.
3. Results
Isolates identification
Macroscopic characteristics of front and back of the colonies of the six fungal isolates tolerant to Cd or Pb, and microscopic features at 40X magnification of the fruiting body and spores were observed for identification at genera level (Figure 1). The isolate identified as Penicillium sp. presented green velvety mycelium at the front of the colony and brush-like fruiting body. Paecilomyces sp. had a light pink and powdery colony front, and its fruiting body was composed of verticillated conidiophores. Rhizopus sp. colonies presented fuzzy aerial mycelium, characteristic given due to the millimetric structures of the fruiting bodies. The isolate identified as Fusarium sp. presented a creamy lilac colony and cylindrical fruiting bodies organized in rafts. Cuninghamella sp. isolate presented dense white aerial mycelium and specific straight sporangiophores with visible terminal vesicles.
To identify the genera of fungi tolerant to Cd or Pb through distinctive characteristics, morphology techniques were applied (Table 1). To assure a precise identification of fungi, ITS 4 and 5 were amplified (Figure 2). For all tolerant Cd or Pb isolates bands in the range of 500-700 bp were observed (corresponding to ITS 4 and 5); they were purified and sequenced. The retrieved sequences had more than 80% coincidence with fungal species registered at NCBI database when aligned.
Tolerance to Cd or Pb
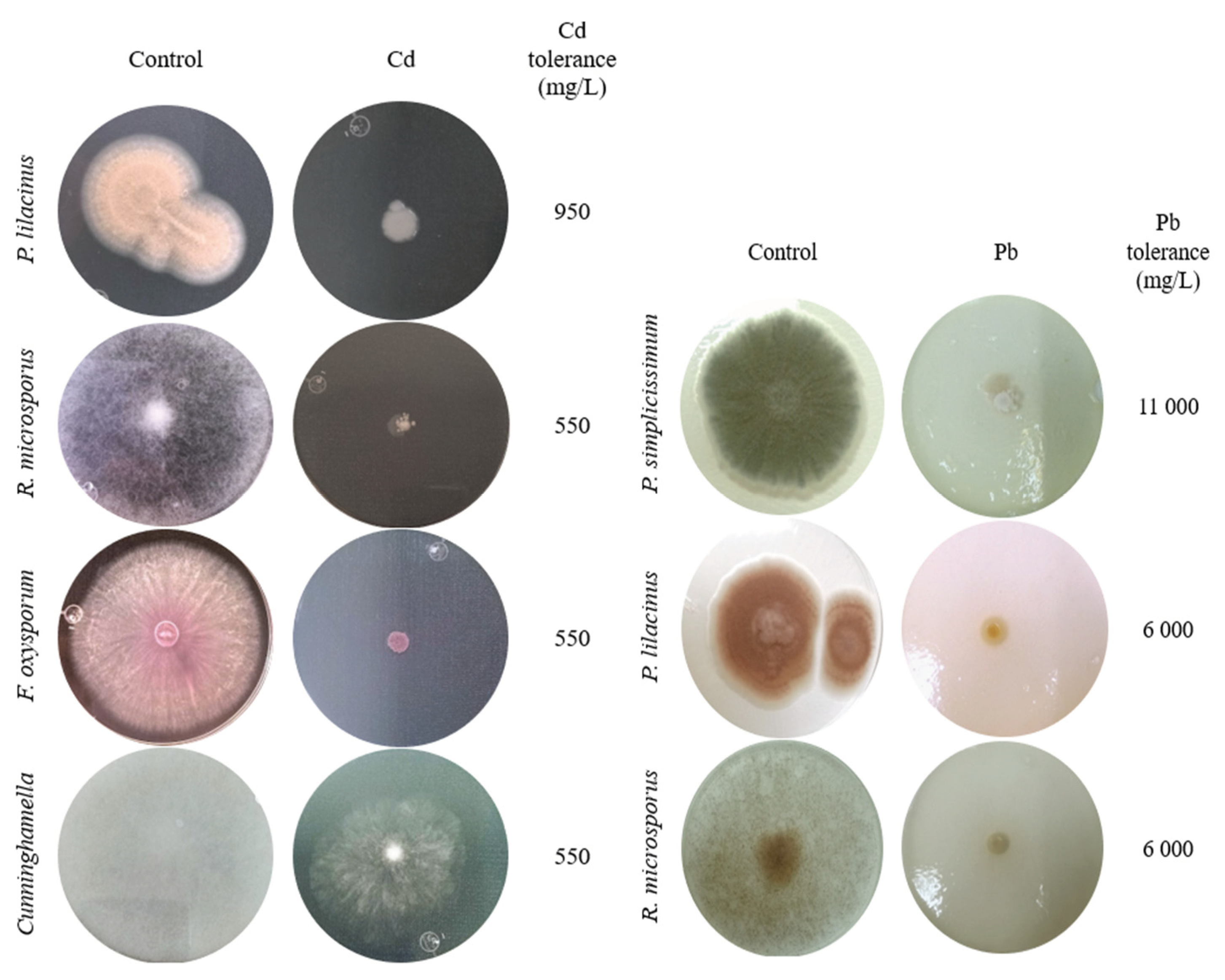
All Cd resistant isolates showed significant differences in comparison to control in terms of diameter length (p<0.05) in every concentration tested (Figure 3). Specifically, P. lilacinus showed growth in a Cd concentration of 950 mg/L (Figure 3A), presenting only diameter reduction, while F. oxysporum and R. microsporus also had mycelium reduction, without adverse effects on mycelium coloration (Figure 5), in the metal concentration of 550 mg/L (Figure 3B,C). Cuninghamella sp. showed colony color change from grayish white to translucent white (Figure 5), along with considerable aerial mycelium reduction in a concentration of 550 mg/L of Cd (Figure 3D).
P. simplicissimum exhibited growth up to 11,000 mg/L of Pb (Figure 4A). Nonetheless, morphological changes were detected such as irregular edges, bulges and color change from green to yellowish white, alongside mycelium diameter reduction as metal concentration in culture media increased (Figure 5). P. lilacinus grew up to a Pb concentration of 6,000 mg/L (Figure 4B), showing diameter reduction and colony coloration changes from lilac to yellowish white; meanwhile, R. microsporus had an aerial mycelium and diameter reduction as Pb concentration increased up to 6,000 mg/L (Figure 4C), from which growth was not observed anymore (Figure 5).
Figure 3.
Fungal colony diameter in different concentrations of Cd (n=3). A) Paecilomyces lilacinus, B) Fusarium oxysporum, C) Rhizopus microsporus, D) Cuninghamella sp.; * Significant difference (P < 0.05) between mean fungal growth in media culture without Cd (control) and growth in different Cd concentrations.
Figure 3.
Fungal colony diameter in different concentrations of Cd (n=3). A) Paecilomyces lilacinus, B) Fusarium oxysporum, C) Rhizopus microsporus, D) Cuninghamella sp.; * Significant difference (P < 0.05) between mean fungal growth in media culture without Cd (control) and growth in different Cd concentrations.
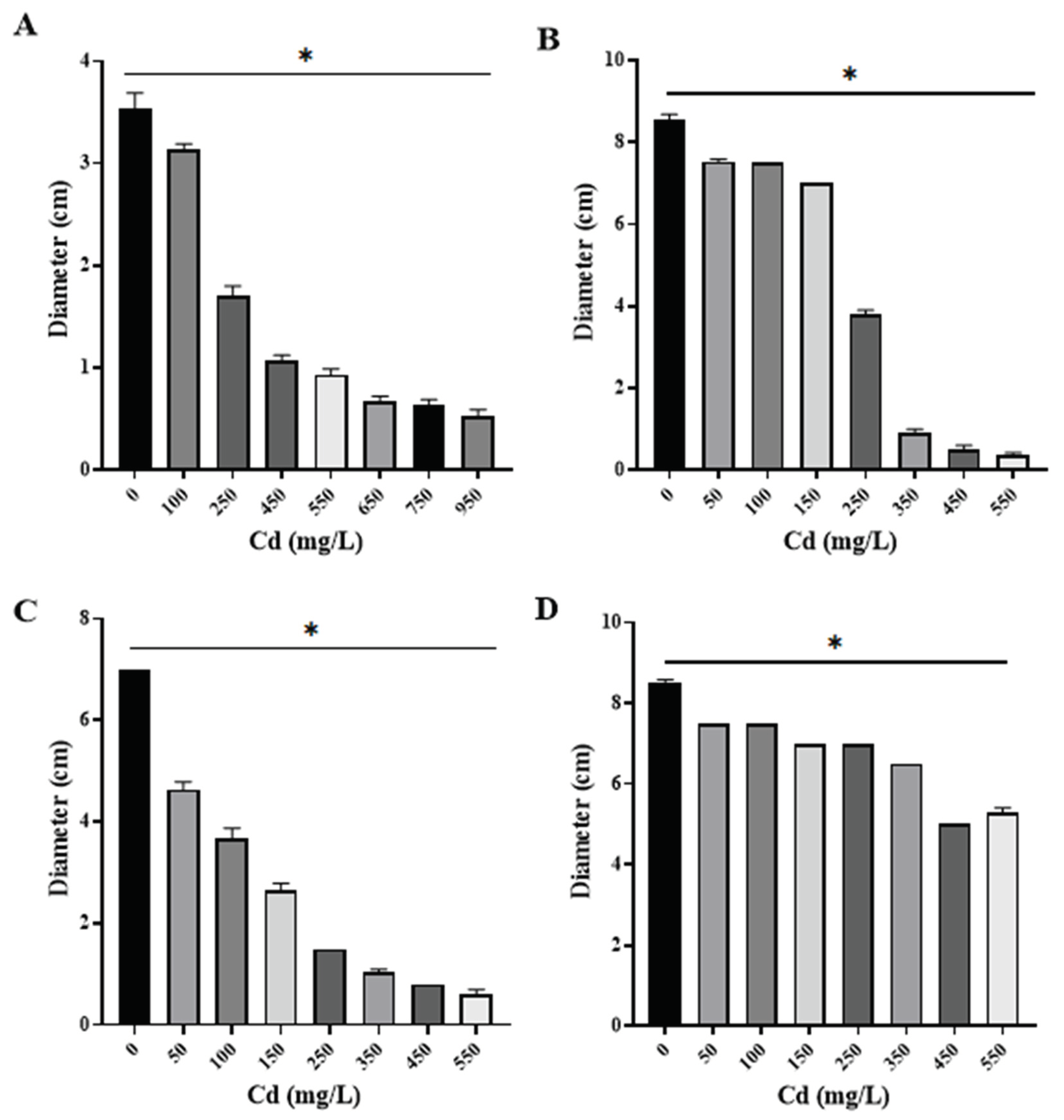
Figure 4.
Fungal colony diameters in different concentrations of Pb (n=3). A) Os1 Penicillium simplicissimum. B) Os6 Paecilomyces lilacinus. C) Os7 Rhizopus microsporus. * Significant difference (P < 0.05) between control (mean fungal growth in media culture without Pb) and growth in different Pb concentrations. ns, no significant.
Figure 4.
Fungal colony diameters in different concentrations of Pb (n=3). A) Os1 Penicillium simplicissimum. B) Os6 Paecilomyces lilacinus. C) Os7 Rhizopus microsporus. * Significant difference (P < 0.05) between control (mean fungal growth in media culture without Pb) and growth in different Pb concentrations. ns, no significant.
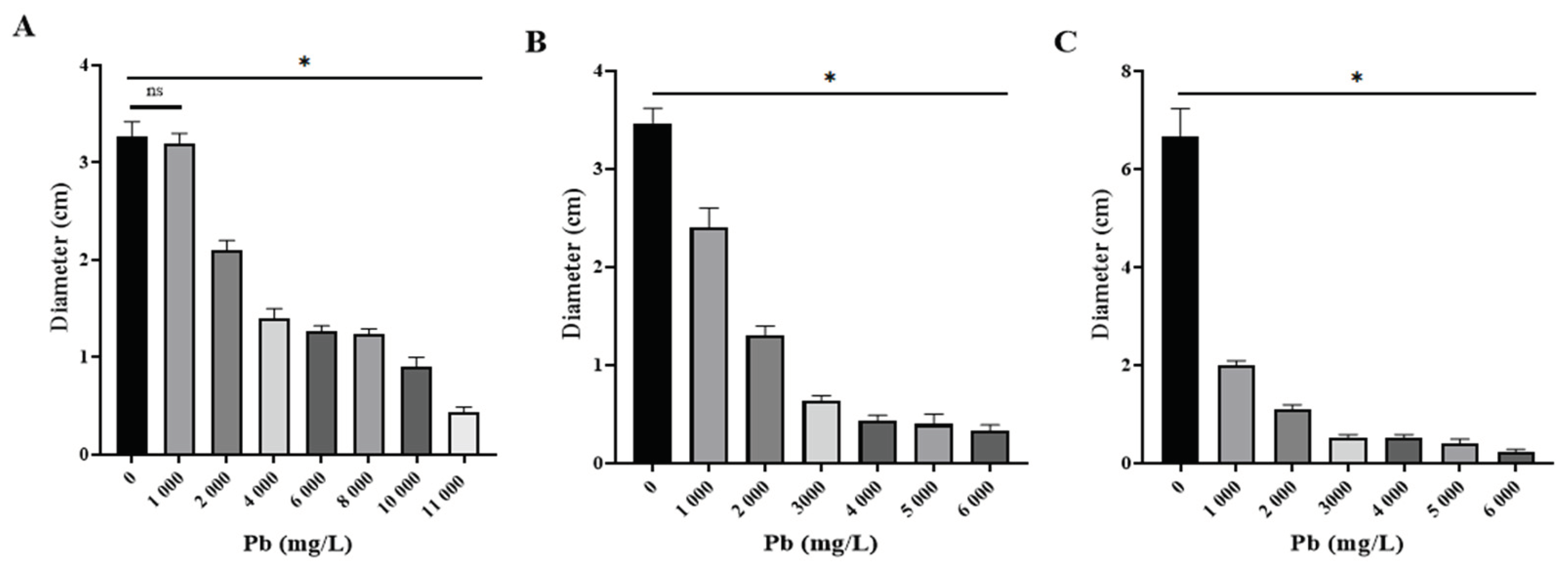
Figure 5.
Tolerance and morphological changes of fungal colonies in PDA with and without Cd or Pb.
Tolerance levels to cadmium and lead of the isolates were also estimated based on the metal concentration to inhibit 50% of fungal growth (IC50), based on biomass production in different concentrations of Cd or Pb in culture media (Figure 6 and Figure 7). Fungal isolates growth varied depending on metal concentration, and all displayed a different IC50 value.
Regarding Cd exposure, Paecilomyces lilacinus and Fusarium oxysporum exhibited a gradual growth diminishment, as metal concentration increased (Figure 6). Their IC50 values were 311 mg/L and 223 mg/L, respectively (Figure 6A,B). In contrast, R. microsporus and Cuninghamella sp had a significant growth reduction of 48.7% and 41.8%, with IC50 values of 29.3 mg/L and 25.2 mg/L (Figure 6C,D), respectively, regarding control. With reference to Pb exposure, P. simplicissimum, had a visible growth reduction in 2,000 mg/L and then displayed gradual reduction as metal concentration increased, reaching an IC50 value of 3,874 mg/L (Figure 7A), in contrast, P. lilacinus growth was diminished in 28.51% as the first metal concentration (500 mg/L) was added, with an IC50 of 1,176 mg/L (Figure 7B), whereas R. microsporus showed growth reduction of 27.9% in 50 mg/L and an IC50 of 212 mg/L (Figure 7C).
4. Discussion
In this study, high tolerance to Pb and to Cd was detected in three and four native fungal species, respectively, all isolated from Mexican contaminated soils with mining tailings. These findings suggest that some fungi can survive because they have developed the capacity to immobilize heavy metals that are present in the soil, which may be relevant in mitigation of mining pollution and thus diminish the risk of exposure and damage of human and animal neighboring populations.
As a first step to exploring native fungi in regions affected by metal pollution, metal-tolerant fungi were identified. Researches indicates that Beauveria bassiana can tolerance elevated concentrations of Pb(II), although this processes results in a gradual decline of fungal biomass [30]. Similarly, Trichoderma viride exhibits adaptive responses of its growth upon exposure to higher Pb2+ concentrations, suggesting a mechanism of adaptation that depends on concentration [31]. Furthermore, research conducted by Chen et al. [32] has revealed that fungi like S. chinense, T. asperellum and Coriolopsis sp. can utiliza both surface binding to the metal and intracellular sequestration as tolerance strategies. White rot fungi (Phellinus spp., Phlebia spp., Pleurotus spp., etc.), also show the ability to adsorb and accumulate metals, making them promising candidates for selective sorption of heavy metal ions contaminating polluted waters [33]. These authors’ results highlight the importance of identifying and characterizing metal-tolerant fungi to enhance our understanding of their mechanisms of tolerance and their potential applications in environmental remediation efforts.
The tolerant fungi were morphologically identified (Figure 1), and their identity was verified with the results obtained from molecular tests by sequencing fungal highly conserved ITS fragments and their corresponding alignments to recorded sequences in the NCBI database (Table 1). Recent research on fungal communities in mine contaminates soils has employed ITS based sequencing to successfully characterize taxonomic composition and species diversity [34]. Similarly, studies isolating metal tolerant fungi from contaminate environments, such as Mucor sp. CBRF59, have based on ITS sequences in combination with morphological data for fungal identification [35]. ITS-based methods have also been used to identify dominant taxa involved in metal tolerance, uptake and accumulation in arbuscular mycorrhizal fungi [36].
In addition to ITS, analysis of other genetic materials or whole genome approaches have also been used to obtain higher taxonomic resolution; however, the internal transcribed spacer region remains a widely used and accepted molecular marker for molecular identification of fungi, especially in ecological studies and settings such as the present study. Therefore, the use of ITS in our study is consistent with current methodologies for fungal identification of environmental interest, making it sufficient for the selection of strains with potential for biotechnological application.
The outcomes of this study are aligning earlier findings that emphasize the benefits of native microorganisms in mitigating heavy metal stress. Sagar et al. [37] point out that Enterobacter sp. PR14 was demonstrates plasmid-mediated tolerance to multiple heavy metal contamination, highlighting the genetic basis of resistance. In addition, recent research has revealed that some fungi can synthesize nanovesicles and extracellular polymeric substances (EPS) to capture and immobilize heavy metal ions in soil [38]., which offers new perspectives for the use of metal-tolerant fungi under conditions of intense contamination, as occurs in soils contaminated by mining tailings. In agriculture, the usefulness of microbial-assisted bioremediation strategies has also been shown; an example is the use of Mesorhizobium RC3 for growth enhancement and nodulation of chickpea under chromium stress [39]. In summary, these authors’ reports reinforce the idea that the fungal strains used in our study could have the potential to be valuable resources for the bioremediation of sites contaminated with multiple metals.
The isolates exhibited tolerance to different concentrations of Cd or Pb. About Pb exposure, it was observed that one isolate, P. simplicissimum, showed tolerance up to 11,000 mg/L, whereas P. lilacinus. and R. microsporus isolates showed tolerance to 6,000 mg/L. In reference to Cd exposure, P. lilacinus was tolerant to 950 mg/L, while Cunninghamella sp., R. microsporus y F. oxysporum displayed tolerance to a limit of 550 mg/L. All isolates experienced growth reduction under the influence of these two heavy metals, since mycelial radial growth was shorter in comparison to controls (Figure 3, Figure 4 and Figure 5). This radial reduction has been previously reported in other fungi. Urquhart el al. [40], described that Paecilomyces variotii growth was inhibited at 1,000 mg/L of Pb after three days of incubation. Other studies reported inhibitory concentrations of 1,000 mg/L of Pb for Penicillium sp. and of 843 mg/L of Cd for Paecilomyces sp. [9,41]. Zeng et al. [42], reported growth of high-density white mycelium with yellow bottom for P. lilacinus in Cd concentrations as high as 8,950 mg/L; this study was carried out with an isolate from a cadmium smelting plant. The results of these authors together with the findings of this work reveal the importance of thoroughly investigating the tolerance of fungi presents in soils contaminated with heavy metals.
It has also been reported that native saprotrophic micro fungi exhibit high tolerance levels to different pollutants, metals included. Within the tolerant fungi we can find Aspergillus sp., Trichoderma sp., Penicillium sp., Geotrichum sp. and Cladosporium sp [43,44,45,46]. However, our results report levels of tolerance (Figure 5) to Cd or Pb that have not been previously reported for fungi such as P. lilacinus and Cuninghamella sp. And in fungi such as Penicillium, Rhizopus and Fusarium, higher tolerances are reported (Figure 5) than those found in the works of the aforementioned authors.
Tolerance to heavy metals by filamentous fungi is not restricted to cadmium or lead. In the work by Chun, et al. [47], Fusarium sp. and Trichoderma sp. isolated from abandoned mines showed tolerance to Cu. Aspergillus sp., Penicillium sp. y Rhizopus sp. have also shown tolerance to Cd and Pb, with morphological changes in colonies, and, additionally, to Cr, Cu and Zn [48,49]. Fungi tend to react with colony morphological changes such as size reduction and color change. Such changes were visible in this study when high concentrations of Pb were added to P. simplicissimum cultures, while the presence of Cd in cultures resulted in inhibition of colony growth without color changes or changes in colony shape compared to the control (Figure 5). Oladipo et al. [50], analyzed the response to heavy metals in terms of growth and tolerance in filamentous fungi isolated from gold and precious stones mining sites. These researchers recorded that Rhizopus microsporus tolerated a total concentration of 250 mg/Kg of Pb; this tolerance was 24 times lower than that exhibited by the R. microsporus isolate in this study (Figure 5); this suggests that each fungal species exhibits a particular response depending on the area where it was isolated, and that even the tolerance mechanism would be different depending on the metal it is exposed to.
A study by Văcar et al. [24], recorded that IC50 Pb concentration reached for Fusarium oxysporum was 1,568 mg/L, although the colony had a considerable reduction in size. For Paecilomyces spp isolated from Cd contaminated soils, a tolerance of 10 mg/L to Cd and 1,243 mg/L to Pb [40]; authors describe an irregular growth pattern attributable to Pb, and growth inhibition with 10 mg/L of Cd. Regarding other fungi, as Mucor sp., biomass considerably decreased in presence of small concentrations of Cd and Pb [35]. The same inhibition pattern is visible in the results shown with R. microsporus in the presence of Cd or Pb, as well as in Cuninghamella sp in the presence of Cd (Figure 6 and Figure 7). These results suggest that fungi have tolerance to heavy metals, and specific growth patterns as shown in the six isolates found in this study, hence it is important to continue researching the specific tolerance of native fungi and to test their efficacy to remediate soils from contaminants they are competent for [51], which is intrinsically related to the ambient conditions that these fungi were isolated from.
These findings mark one of the first reports of filamentous fungi isolated from soils impacted by mining in Concepcion del Oro, Zacatecas, Mexico, an area that has not been extensively studied in terms of microbial ecology. Additionally, it is significant that the elevated IC50 values observed for Penicillium simplicissimum (Pb) and Paecilomyces lilacinus (Cd and Pb) surpass those reported in previous studies of the same fungi, indicating exceptional tolerance that could reflect long term adaptation to extreme metal concentrations. The identification of dual metal tolerance in P. lilacinus further underscores its potential as a suitable option for bioremediation efforts in sites affected by multiple metal contaminants. While these experiments were performed under controlled in vitro conditions, the results provide a strong basis for future investigations in more applicable environmental contexts involving highly tolerant organisms isolated from the contaminated areas.
5. Conclusions
This research highlights the significant tolerance to cadmium and lead in filamentous fungi isolated from soils contaminated by mining tailings in a region with scarce microbial ecological characterization. The isolates showed tolerance to elevated concentrations of Pb and Cd, with Penicillium simplicissimum and Paecilomyces lilacinus showing particularly high IC50 values, higher than previously reported, suggesting a long-term adaptation to severe pollution. The dual metal tolerance detected in P. lilacinus further supports the potential for bioremediation applications in multi-metal contaminated environments. Although experiments were conducted under controlled in vitro conditions, the results provide a relevant basis for future studies in more complex environmental conditions. Finally, these findings emphasize the importance of identifying native metal-tolerant fungi as an initial approach to comprehending their tolerance mechanisms and assess their potential contribution to the remediation of polluted soils.
Author Contributions
We show aprpreciation to all contributions made by the authors of this work as follows: Data curation, Oscar Flores Amaro and Laura Yamamoto Flores; Formal analysis, Denisse Ramos Suárez; Investigation, Denisse Ramos Suárez; Methodology, J. Felix Gutiérrez Corona and Juan Bautista Bautista; Project administration, Alma Guerro Barrera and Francisco Avelar Gonzalez; Software, J. Felix Gutiérrez Corona and Juan Bautista Bautista; Supervision, Arturo Valdivia Flores and Francisco Avelar Gonzalez; Writing – original draft, Denisse Ramos Suárez; Writing – review & editing, Arturo Valdivia Flores and Francisco Avelar Gonzalez.
Funding
The research leading to these results received funding from CONAHCYT under Grant Agreement No 811213.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed at the corresponding author(s).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ITS | Internal Transcribed Spacers |
| Cd | Cadmium |
| Pb | Lead |
| MLP | Maximum Permissible Limits |
| IC50 | Half maximal Inhibitory Concentration |
| bp | Base Pair |
References
- M. H. Wong, “Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils,” Chemosphere, vol. 50, no. 6, pp. 775–780, 2003. [CrossRef]
- S. A. Covarrubias, J. A. García Berumen, and J. J. Peña Cabriales, “Microorganisms role in the bioremediation of contaminated soils with heavy metals,” Acta Univ., vol. 25, no. NE-3, pp. 40–45, 2015. [CrossRef]
- F. García Aguirre, “La minería en México,” Espac. para el Cap. a cielo abierto, INEGI, pp. 128–136, 2012, Available: http://www.redalyc.org/pdf/124/12426062013.pdf.
- O. A. Flores-Amaro et al., “Characterization and evaluation of the bioremediation potential of Rhizopus microsporus Os4 isolated from arsenic-contaminated soil,” Water. Air. Soil Pollut., vol. 235, no. 8, 2024. [CrossRef]
- DOF, NORMA OFICIAL MEXICANA NOM-147-SEMARNAT/SSA1-2004, Que establece criterios para determinar las concentraciones de remediación de suelos contaminados por arsénico, bario, cadmio, cromo hexavalente, mercurio, níquel, plata, selenio, talio y/o vanadio. 2007, pp. 35–96. Available: http://www2.inecc.gob.mx/publicaciones/libros/402/cuencas.html.
- S. A. Covarrubias and J. J. Peña Cabriales, “Contaminación ambiental por metales pesados en México: Problemática y estrategias de fitorremediación,” Rev. Int. Contam. Ambient., vol. 33, pp. 7–21, 2017. [CrossRef]
- Agency for Toxic Substances and Disease Registry Sep, “Toxicological Profile for Cadmium,” ATSDR’s Toxicol. Profiles, no. September, 2002. [CrossRef]
- L. Newsome and C. Falagán, “The Microbiology of Metal Mine Waste: Bioremediation Applications and Implications for Planetary Health,” GeoHealth, vol. 5, no. 10, pp. 1–53, 2021. [CrossRef]
- D. Tian et al., “A new insight into lead (II) tolerance of environmental fungi based on a study of Aspergillus niger and Penicillium oxalicum,” Environ. Microbiol., vol. 21, no. 1, pp. 471–479, 2019. [CrossRef]
- FAO & ITPS, Status of the World’s Soil Resources (SWSR) – Technical Summary. Food and Agriculture Organization of the United Nation and Intergovernmental echnical Panel on Soils, Rome, Italy. 2015. Available: http://www.fao.org/3/i5126e/i5126e.pdf link accessed on 28/04/2020.
- I. E. García, “Microorganismos del suelo y sustentabilidad de los agroecocistemas,” Rev. Argent. Microbiol., vol. 43, no. 1, pp. 1–3, 2011, Available: https://www.redalyc.org/pdf/2130/213019226001.pdf.
- J. A. Samaniego-Gaxiola and Y. Chew-Madinaveitia, “Diversidad de géneros de hongos del suelo en tres campos con diferente condición agrícola en La Laguna, México,” Rev. Mex. Biodivers., vol. 78, no. 2, pp. 383–390, 2007.
- M. Delgado, “Los microorganismos del suelo en la nutrición vegetal,” Investig. ORIUS Biotecnol., pp. 1–9, 2008, Available: https://portalcamaronero.com/wp-content/uploads/2016/10/Los-microorganismos-del-suelo-en-la-nutrición-vegetal.pdf.
- E. P. Burford, M. Fomina, and G. M. Gadd, “Fungal involvement in bioweathering and biotransformation of rocks and minerals,” Mineral. Mag., vol. 67, no. 6, pp. 1127–1155, 2003. [CrossRef]
- X. Yu and Q. Zhan, “Phosphate-Mineralization Microbe Repairs Heavy Metal Ions That Formed Nanomaterials in Soil and Water,” Nanomater. - Toxicity, Hum. Heal. Environ., pp. 3–9, 2020. [CrossRef]
- X. Zhang et al., “Microbial controls on heavy metals and nutrients simultaneous release in a seasonally stratified reservoir,” Environ. Sci. Pollut. Res., vol. 29, no. 2, pp. 1937–1948, 2022. [CrossRef]
- A. S. Ayangbenro and O. O. Babalola, “A new strategy for heavy metal polluted environments: A review of microbial biosorbents,” Int. J. Environ. Res. Public Health, vol. 14, no. 1, 2017. [CrossRef]
- J. Qiu et al., “Experimental and modeling studies of competitive Pb (II) and Cd (II) bioaccumulation by Aspergillus niger,” Appl. Microbiol. Biotechnol., vol. 105, no. 16–17, pp. 6477–6488, 2021. Available: https://doi.org/10.1007/s00253-021-11497-3. [CrossRef]
- A. Kapoor, T. Viraraghavan, and D. R. Cullimore, “Removal of heavy metals using the fungus Aspergillus niger,” Bioresour. Technol., vol. 70, pp. 95–104, 1999.
- S. E. Hassan, M. Hijri, and M. St-Arnaud, “Effect of arbuscular mycorrhizal fungi on trace metal uptake by sunflower plants grown on cadmium contaminated soil,” N. Biotechnol., vol. 30, no. 6, pp. 780–787, 2013. Available: http://dx.doi.org/10.1016/j.nbt.2013.07.002. [CrossRef]
- Y. Xie, H. Luo, Z. Du, L. Hu, and J. Fu, “Identification of cadmium-resistant fungi related to Cd transportation in bermudagrass [Cynodon dactylon (L.) Pers.],” Chemosphere, vol. 117, no. 1, pp. 786–792, 2014. Available: http://dx.doi.org/10.1016/j.chemosphere.2014.10.037. [CrossRef]
- M. Refaey, A. M. Abdel-Azeem, H. H. Abo Nahas, M. A. Abdel-Azeem, and A. A. El-Saharty, Role of Fungi in Bioremediation of Soil Contaminated with Heavy Metals, no. June. 2021. [CrossRef]
- A. Bonifaz Trujillo, Micología Médica Básica, Cuarta. Ciudad de Mexico: Mc Graw-Hill, 2012. Available: https://mega.nz/folder/UR0D0YwL#eRPcO3H4VRhMrkJeVBtUoA.
- C. L. Văcar et al., “Heavy metal-resistant filamentous fungi as potential mercury bioremediators,” J. Fungi, vol. 7, no. 5, 2021. [CrossRef]
- T. Janicki, J. Długoński, and M. Krupiński, “Detoxification and simultaneous removal of phenolic xenobiotics and heavy metals with endocrine-disrupting activity by the non-ligninolytic fungus Umbelopsis isabellina,” J. Hazard. Mater., vol. 360, pp. 661–669, 2018. Available: https://doi.org/10.1016/j.jhazmat.2018.08.047. [CrossRef]
- M. Le Berre, J. Q. Gerlach, I. Dziembała, and M. Kilcoyne, “Calculating Half Maximal Inhibitory Concentration (IC50) Values from Glycomics Microarray Data Using GraphPad Prism,” Methods Mol. Biol., vol. 2460, pp. 89–111, 2022. [CrossRef]
- E. J. Rangel-Muñoz et al., “Assessment of the Potential of a Native Non-Aflatoxigenic Aspergillus flavus Isolate to Reduce Aflatoxin Contamination in Dairy Feed,” Toxins (Basel)., vol. 14, no. 7, pp. 1–14, 2022. [CrossRef]
- S. M. Aljanabi and I. Martinez, “Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques,” Nucleic Acids Res., vol. 25, pp. 4692–4693, 1997.
- J. Sambrook and D. . Russell, Molecular cloning: a laboratory manual, 3rd ed., vol. 2. New York: Cold Sring Harbor Labratory, 2001. [CrossRef]
- D. Gola, A. Malik, M. Namburath, and S. Z. Ahammad, “Removal of industrial dyes and heavy metals by Beauveria bassiana: FTIR, SEM, TEM and AFM investigations with Pb(II),” Environ. Sci. Pollut. Res., vol. 25, no. 21, pp. 20486–20496, 2018. [CrossRef]
- D. Luo, S. Qiang, R. Geng, L. Shi, J. Song, and Q. Fan, “Mechanistic study for mutual interactions of Pb2+ and Trichoderma viride,” Ecotoxicol. Environ. Saf., vol. 233, Mar. 2022. [CrossRef]
- S. H. Chen, S. L. Ng, Y. L. Cheow, and A. S. Y. Ting, “A novel study based on adaptive metal tolerance behavior in fungi and SEM-EDX analysis,” J. Hazard. Mater., vol. 334, pp. 132–141, 2017. [CrossRef]
- H. R. Noormohamadi, M. R. Fat’hi, M. Ghaedi, and G. R. Ghezelbash, “Potentiality of white-rot fungi in biosorption of nickel and cadmium: Modeling optimization and kinetics study,” Chemosphere, vol. 216, pp. 124–130, 2019. Available: https://doi.org/10.1016/j.chemosphere.2018.10.113. [CrossRef]
- F. Ye, D. Gong, C. Pang, J. Luo, X. Zeng, and C. Shang, “Analysis of Fungal Composition in Mine-Contaminated Soils in Hechi City,” Curr. Microbiol., vol. 77, no. 10, pp. 2685–2693, 2020. Available: https://doi.org/10.1007/s00284-020-02044-w. [CrossRef]
- Z. Deng et al., “Characterization of Cd- and Pb-resistant fungal endophyte Mucor sp. CBRF59 isolated from rapes (Brassica chinensis) in a metal-contaminated soil,” J. Hazard. Mater., vol. 185, no. 2–3, pp. 717–724, 2011. Available: http://dx.doi.org/10.1016/j.jhazmat.2010.09.078. [CrossRef]
- M. J. Salazar et al., “Pb accumulation in spores of arbuscular mycorrhizal fungi,” Sci. Total Environ., vol. 643, pp. 238–246, 2018. Available: https://doi.org/10.1016/j.scitotenv.2018.06.199. [CrossRef]
- A. Sagar, R. Riyazuddin, P. K. Shukla, P. W. Ramteke, and R. Z. Sayyed, “Heavy metal stress tolerance in Enterobacter sp. PR14 is mediated by plasmid,” Indian J. Exp. Biol., vol. 58, no. 2, pp. 115–121, 2020. [CrossRef]
- V. Budamagunta et al., “Nanovesicle and extracellular polymeric substance synthesis from the remediation of heavy metal ions from soil,” Environ. Res., vol. 219, p. 114997, Feb. 2023. Available: https://www.sciencedirect.com/science/article/abs/pii/S0013935122023246. [Accessed: Apr. 24, 2025]. [CrossRef]
- H. Naz et al., “Mesorhizobium improves chickpea growth under chromium stress and alleviates chromium contamination of soil,” J. Environ. Manage., vol. 338, p. 117779, Jul. 2023. Available: https://www.sciencedirect.com/science/article/abs/pii/S0301479723005674?via%3Dihub. [Accessed: Apr. 24, 2025]. [CrossRef]
- A. S. Urquhart, N. F. Chong, Y. Yang, and A. Idnurm, “A large transposable element mediates metal resistance in the fungus Paecilomyces variotii,” Curr. Biol., vol. 32, no. 5, pp. 937-950.e5, 2022. Available: https://doi.org/10.1016/j.cub.2021.12.048. [CrossRef]
- M. Słaba, E. Gajewska, P. Bernat, M. Fornalska, and J. Długoński, “Adaptive alterations in the fatty acids composition under induced oxidative stress in heavy metal-tolerant filamentous fungus Paecilomyces marquandii cultured in ascorbic acid presence,” Environ. Sci. Pollut. Res., vol. 20, no. 5, pp. 3423–3434, 2013. [CrossRef]
- X. Zeng, J. Tang, H. Yin, X. Liu, P. Jiang, and H. Liu, “Isolation, identification and cadmium adsorption of a high cadmium-resistant Paecilomyces lilacinus,” African J. Biotechnol., vol. 9, no. 39, pp. 6525–6533, 2010.
- E. Alori and O. Fawole, “Phytoremediation of Soils Contaminated with Aluminium and Manganese by Two Arbuscular Mycorrhizal Fungi,” J. Agric. Sci., vol. 4, no. 8, pp. 246–252, 2012. [CrossRef]
- A. Hassan, A. Pariatamby, I. C. Ossai, and F. S. Hamid, “Bioaugmentation assisted mycoremediation of heavy metal and/metalloid landfill contaminated soil using consortia of filamentous fungi,” Biochem. Eng. J., vol. 157, no. February, p. 107550, 2020. Available: https://doi.org/10.1016/j.bej.2020.107550. [CrossRef]
- P. K. Joshi, A. Swarup, S. Maheshwari, R. Kumar, and N. Singh, “Bioremediation of Heavy Metals in Liquid Media Through Fungi Isolated from Contaminated Sources,” Indian J. Microbiol., vol. 51, no. 4, pp. 482–487, 2011. [CrossRef]
- E. Sey and E. J. D. Belford, “Heavy Metals Tolerance Potential of Fungi Species Isolated from Gold Mine Tailings in Ghana,” J. Environ. Heal. Sustain. Dev., vol. 6, no. 1, pp. 1231–1242, 2021. [CrossRef]
- S. J. Chun, Y. J. Kim, Y. Cui, and K. H. Nam, “Ecological network analysis reveals distinctive microbial modules associated with heavy metal contamination of abandoned mine soils in Korea,” Environ. Pollut., vol. 289, no. January, p. 117851, 2021. Available: https://doi.org/10.1016/j.envpol.2021.117851. [CrossRef]
- F. Liaquat et al., “Evaluation of metal tolerance of fungal strains isolated from contaminated mining soil of Nanjing, China,” Biology (Basel)., vol. 9, no. 12, pp. 1–12, 2020. [CrossRef]
- M. Zotti, S. Di Piazza, E. Roccotiello, G. Lucchetti, M. G. Mariotti, and P. Marescotti, “Microfungi in highly copper-contaminated soils from an abandoned Fe-Cu sulphide mine: Growth responses, tolerance and bioaccumulation,” Chemosphere, vol. 117, no. 1, pp. 471–476, 2014. Available: http://dx.doi.org/10.1016/j.chemosphere.2014.08.057. [CrossRef]
- O. G. Oladipo, O. O. Awotoye, A. Olayinka, C. C. Bezuidenhout, and M. S. Maboeta, “Heavy metal tolerance traits of filamentous fungi isolated from gold and gemstone mining sites,” Brazilian J. Microbiol., vol. 49, no. 1, pp. 29–37, 2018. Available: http://dx.doi.org/10.1016/j.bjm.2017.06.003. [CrossRef]
- A. Singh and A. Roy, In Fungal communities for the remediation of environmental pollutants, A. N. Yada., vol. 1. Springer International Publishing, 2021. [CrossRef]
Figure 1.
Morphological identification of fungi tolerant to Cd or Pb. Macroscopic and microscopic characteristics of the colonies.
Figure 1.
Morphological identification of fungi tolerant to Cd or Pb. Macroscopic and microscopic characteristics of the colonies.
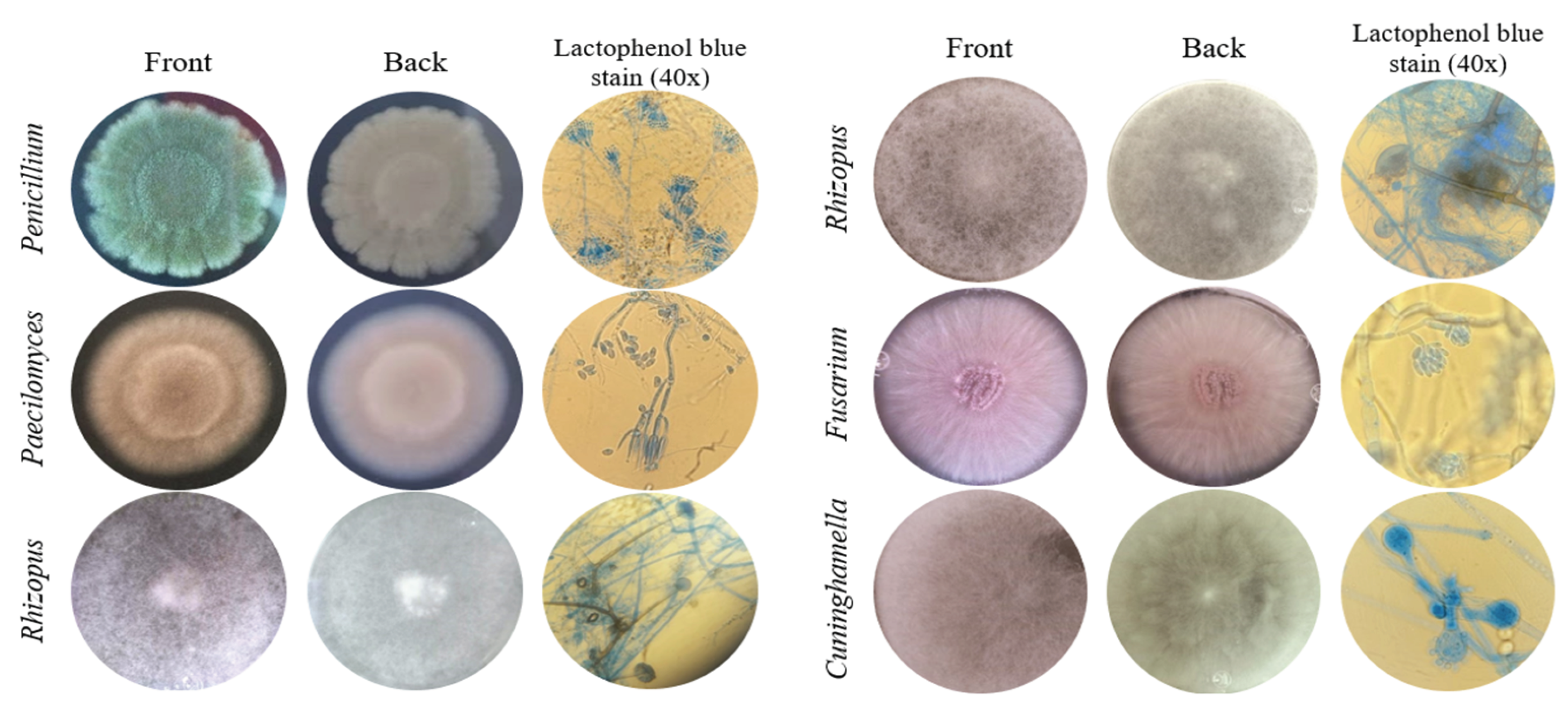
Figure 2.
Amplification of Internal Transcribed Spacers (ITS) 4 and 5 of isolated fungi.
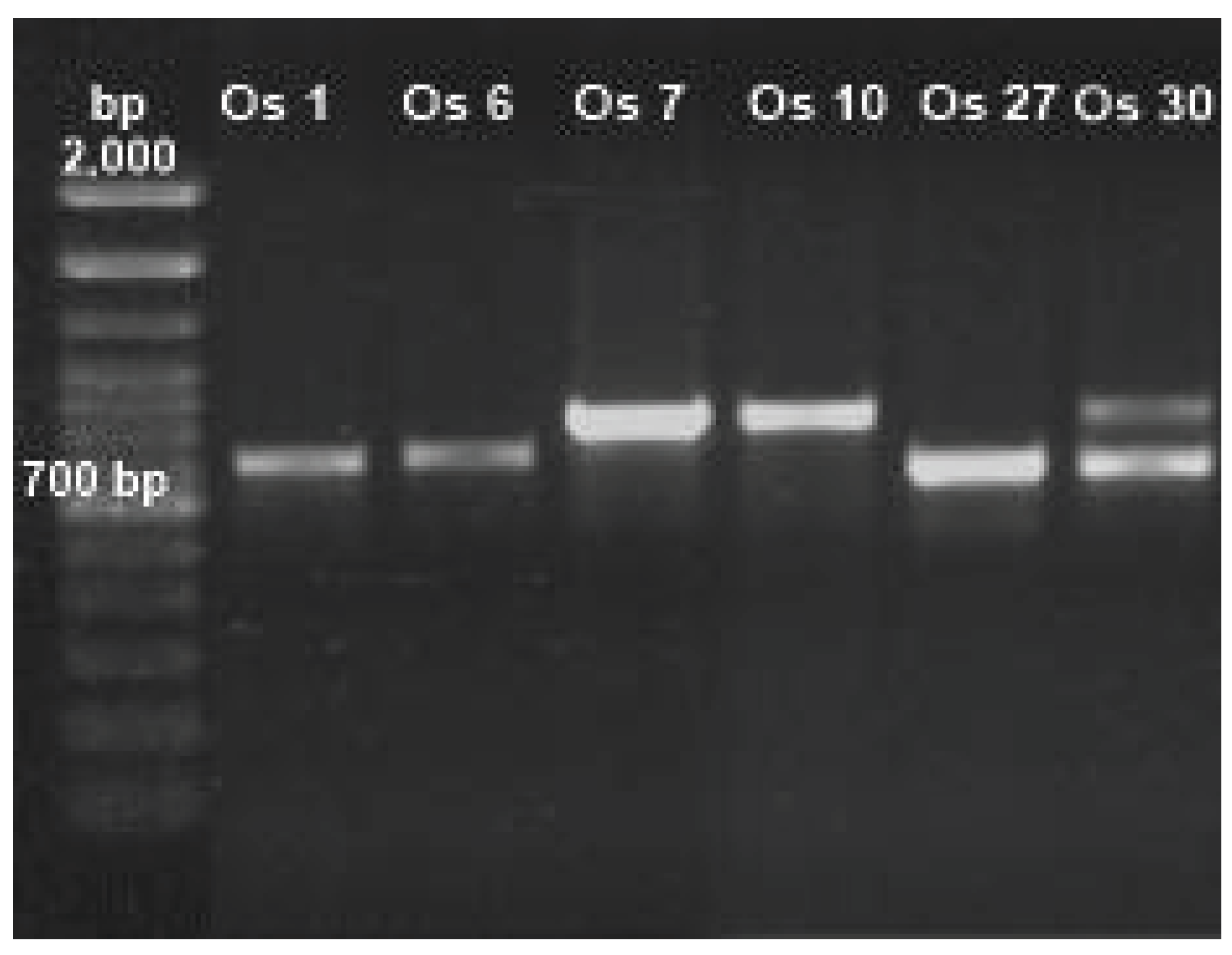
Figure 6.
Half maximal inhibitory concentration (IC50) of isolated fungi in different concentrations of Cd. The dotted line corresponds to IC50 values. A) Paecilomyces lilacinus, 311.3 mg/L; B) Fusarium oxysporum, 223 mg/L; C) Rhizopus microsporus, 29.25 mg/L; D) Cuninghamella sp, 25.18 mg/L. R2 = determination coefficient. Sx.y = Standard deviation Cd*Dry biomass.
Figure 6.
Half maximal inhibitory concentration (IC50) of isolated fungi in different concentrations of Cd. The dotted line corresponds to IC50 values. A) Paecilomyces lilacinus, 311.3 mg/L; B) Fusarium oxysporum, 223 mg/L; C) Rhizopus microsporus, 29.25 mg/L; D) Cuninghamella sp, 25.18 mg/L. R2 = determination coefficient. Sx.y = Standard deviation Cd*Dry biomass.
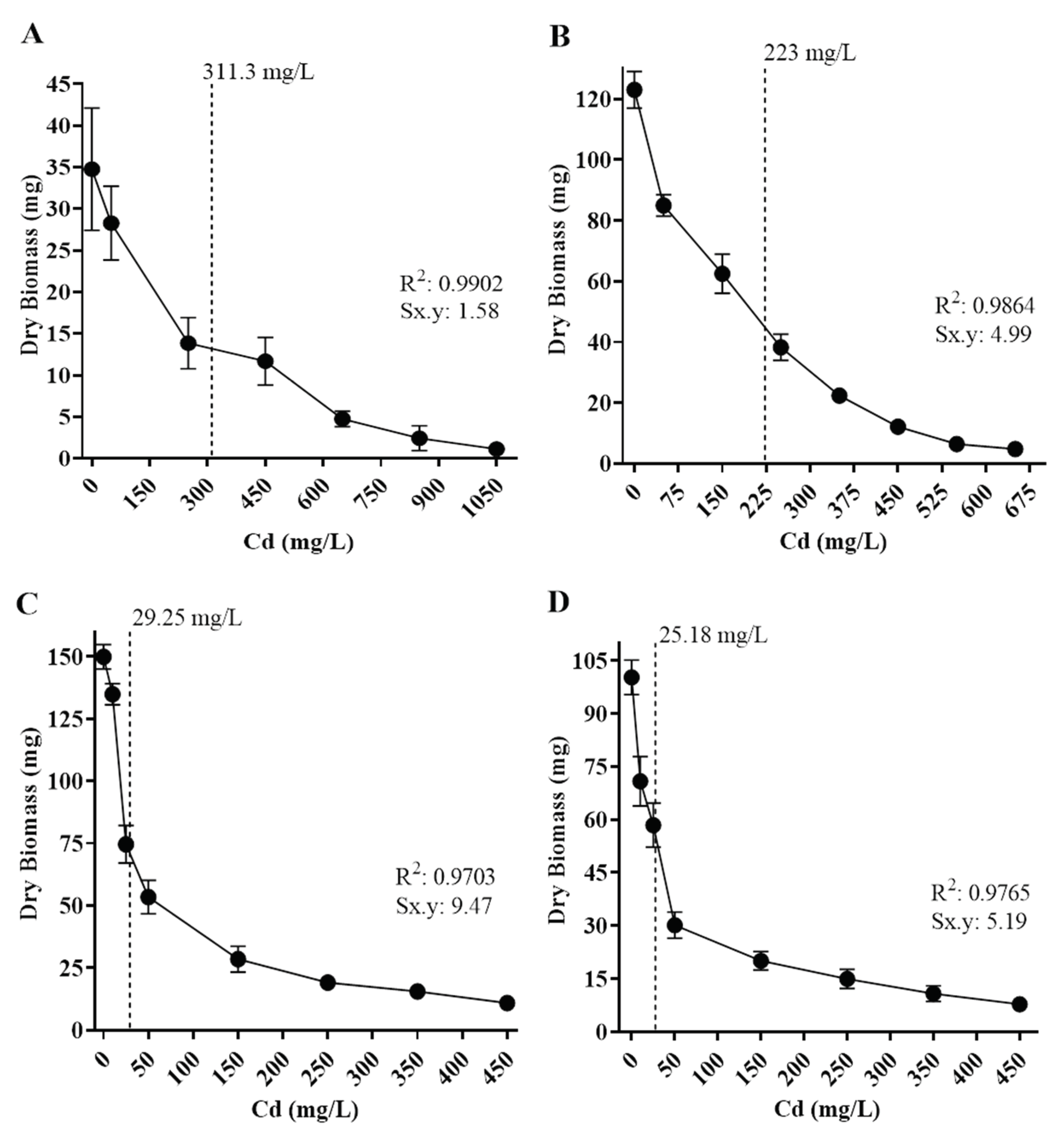
Figure 7.
Half maximal inhibitory concentration (IC50) of isolated fungi in different concentrations of Pb. The dotted line corresponds to IC50 values. A) Penicillium simplicissimum, 3,874 mg/L; B) Paecilomyces lilacinus, 1,176 mg/L; C) Rhizopus microsporus, 211.8 mg/L. Sx.y: Standard deviation dry biomass*Pb.
Figure 7.
Half maximal inhibitory concentration (IC50) of isolated fungi in different concentrations of Pb. The dotted line corresponds to IC50 values. A) Penicillium simplicissimum, 3,874 mg/L; B) Paecilomyces lilacinus, 1,176 mg/L; C) Rhizopus microsporus, 211.8 mg/L. Sx.y: Standard deviation dry biomass*Pb.
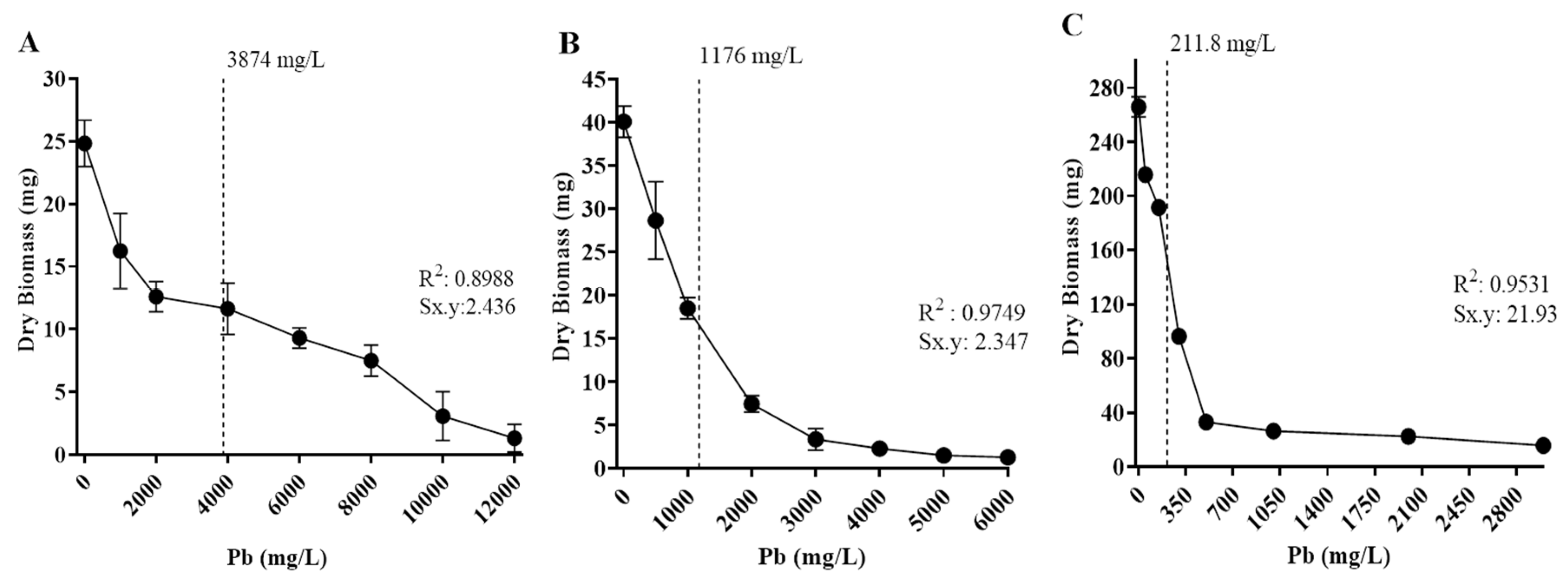

Table 1.
Morphological and molecular identification of fungi tolerant to Cd or Pb obtained from soils contaminated with mining tailings.
Table 1.
Morphological and molecular identification of fungi tolerant to Cd or Pb obtained from soils contaminated with mining tailings.
| Isolate ID | Morphological identification | Size (bp) | Molecular identification | Coincidence (%) | Access |
|---|---|---|---|---|---|
|
Os 1 Os 6 Os 7 |
Penicillium sp. | 552 | P. simplicissimum | 99.4 | MW485753.1 |
| Paecilomyces sp. | 641 | P. lilacinus | 99.8 | MT453285.1 | |
| Rhizopus sp. | 664 | R. microsporus | 100 | MH473977.1 | |
|
Os 10 Os 27 |
Rhizopus sp. | 666 | R. microsporus | 100 | MH473977.1 |
| Fusarium sp. | 513 | F. oxysporum | 99.6 | KX655587.1 | |
|
Os 30 Os 1 |
Cuninghamella sp. | 715 | Cuninghamella sp. | 87.5 | OR096349.1 |
| Penicillium sp. | 552 | P. simplicissimum | 99.4 | MW485753.1 |
* NCBI (National Center for Biotechnology Information): https://blast.ncbi.nlm.nih.gov/. * bp: base pairs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



