
Submitted:
10 May 2025
Posted:
12 May 2025
You are already at the latest version
Abstract
Immunization is a procedure that trains and strengthens host immunity against harmful foreign agents from damaging and taking the host life. Immunization, also termed vaccination, has opened the era of modern medicine, not only contributed to the independence of the USA but also the health of the citizens and the prosperity of the nation. Resonating with the USA growth, vaccinology as a scientific discipline studying vaccine and immunization needs to take a leap forward. Generating the next generation vaccine and eliciting newly defined host immunity are imperative to boost the immunity of we the people and the strength of the nation.
Keywords:
immunization
; epigenetic immunity
; chromatin vaccine (cVacc)
; p21Cip1Waf1Sdi1 (p21)
; recombination-activating gene 2 protein (RAG2)
; CD4 T-cells
; effective HIV vaccine
Introduction
It appears to be the time for Vaccinology to take a leap, since a mandatory smallpox variolation bolstered the troops of Washington and won the War of Independence. Nobel laureate Theiler and his team at the Rockefeller Foundation provided safe and affordable protection against yellow fever by a single dose offering long-term immunity. Then, polio vaccines not only have had a profound, positive impact on societies, drastically reduced polio cases and prevented paralysis and death, but also have led to the establishment of robust health systems and infrastructure for disease surveillance, prevention, treatment and cure.
Footsteps of Trailblazers in Immunization
Immunization and its impact on national independence and societal prosperity can be traced back to the American Revolution. George Washington mandated inoculation (a precursor to vaccination) for his troops to combat the deadly threat of smallpox, a decision that significantly boosted health and the fighting strength of the Continental Army during the War [1,2]. This deed, too, paved a way for Edward Jenner’s discovery of the smallpox vaccine and its usage: vaccination or immunization. Jenner studied applying cowpox to prevent smallpox, revolutionized medicine and laid a cornerstone for immunology, vaccinology, and a combination of both, vaccination or immunization [3,4].
The research programs aiming to protect national strength and health, specifically development of vaccines, have made significant contributions to medicine throughout the history of the USA. In particular, military research programs on immunization have not only benefited the health and strength of the US army but also that of the general public.
Yellow fever was a serious problem for US troops in 1898 [2]. In response, the army created a Yellow Fever Commission, headed by physician Walter Reed. The Commission members traveled abroad and, after extensive experiments and observation, validated a theory presented by Cuban physician Carlos Finlay two decades earlier: mosquitoes were responsible for the spread of the disease. The researchers showed that a virus was the cause of yellow fever, transmitted from one person to another using mosquitoes as vectors.
The American physician William Gorgas then organized collaborative programs, which allowed for the completion of the Panama Canal by the Americans after French efforts were abandoned due to the difficulties of both yellow fever and malaria. Gorgas then joined the Rockefeller Foundation’s Yellow Fever Commission as director of its work on the eradication of yellow fever. Years later, Harvard instructor Max Theiler also joined the Commission and developed the 17D vaccine against yellow fever. Theiler, a virologist, eventually won the Nobel Prize for his efforts, and the 17D vaccine remains in use today [5].
The majority of acute respiratory disease cases among US military trainees were the result of adenovirus infections after World War II, so an adenovirus vaccine was created at Walter Reed Army Institute of Research (WRAIR). After extensive military studies, adenovirus vaccines were given to new military trainees “within hours after their arrival” at basic training beginning in 1971.
The vaccine’s manufacturer, however, ended the vaccine production in 1994, and all stocks were depleted by 1999 as well. Outbreaks of acute respiratory disease caused by adenoviruses rose among military trainees following discontinuation of the vaccination program. So in 2001, the Army provided funds to re-establish an adenovirus vaccine, and the government contracted with a manufacturer to restore a production line for adenovirus type 4 and type 7 vaccine tablets. The vaccine was licensed in 2011, and the US military deployed it to training facilities in the same year. Surveillance of adenovirus illness since then shows a marked decrease in incidence of all serotypes of adenovirus after re-introduction of the vaccine [6].
Today, military researchers remain heavily involved in efforts to develop treatments and vaccines for human immunodeficiency virus (HIV) infection and malaria. The US Military HIV Research Program (MHRP) at the Walter Reed Army Institute of Research studies not only care and treatment of HIV but prevention as well. MHRP scientists, collaborating with the National Institutes of Allergy and Infectious Diseases (NIAID), developed a HIV vaccine candidate that has reached human clinical testing in combination with another vaccine product. Separately, US military researchers have also contributed to the testing of the leading vaccine candidate for malaria.
Vaccinology Today
Since the War of Independence, our knowledge on vaccinology – vaccine and immunization, has developed with the Nation. Vaccines now consist of preventive and therapeutic vaccines, to prevent and treat both acute infections and chronic diseases. The immunization has proceeded into immunotherapy to accommodate the vaccines of different types with multiple functions. For example, infusion of antibodies to prevent HIV infection is an immunotherapy derived from passive immunization. Applying stem cells to treat type I diabetes is an immunotherapy to correct an immune disorder and allow insulin-producing beta cells to grow and be functional.
Although the advancement of immunotherapy is impressive, developing effective vaccines against current diseases remains a challenge, for example, in developing effective HIV, malaria, COVID [7,8,9,10,11,12], or cancer vaccines. If there are no effective vaccines, there will be no effective immunizations nor immunotherapies. In this context, immunogens and vaccines are equivalent and with the same meaning.
After the War of Independence to now the era of precision personalized medicine, vaccinology is on the cusp of a leap, derived from our efforts in exploring the basic principles of vaccine manufacture and host intrinsic immunity. This is the foundation to generate new antimicrobial drugs and novel immunotherapies.
We have proposed to apply a chromatin vaccine (cVacc) to elicit the immunity of our DNA – the epigenetic immunity for production of effective HIV vaccines. The concept and process can be tuned to develop other vaccines or immunogens to generate new antimicrobial drugs and novel immunotherapies. Here, we outline how to apply a cVacc to elicit the host epigenetic immunity.
A Leap Forward in Immunization by cVacc Elicited Epigenetic Immunity
Nothing can stop an idea whose time has come. In an era of modern medicine, we maneuver cutting edge science to treat diseases. Prophylactic and therapeutic vaccines and vaccinations are among them. Vaccination comprises two parts: immunogen and immunity. An immunogen needs to target the immunity to produce results. Knowing the immunogen and its targeted immunity lies at the heart of a successful vaccination.
We will apply a cVacc to target the epigenetic immunity – the immunity of our DNA. We will elicit the power of our genome’s immunity to control the HIV infection by targeting its lifecycle. We aim to force HIV to be the latest member of the ancient human endogenous retroviruses (HERVs), and succumb HIV DNA into a new enhancer or promoter or transposon in our genome, enriching our DNA’s function. We will do this like our body has done to the HERVs.
This study will provide three deliverables, embodying how our DNA’s immunity – epigenetic immunity can be enhanced, which may not only benefit our generation but also generations to come. Furthermore, we hope the knowledge and techniques applied and developed in this research will facilitate advancement of virology, immunology, vaccinology, and also other disciplines.
The NIH human genome project and the epigenomics roadmap consortia have provided a large amount of data that revolutionized our understanding of disease and treatment. Specifically, 8% of the human genome consists of the sequences of HERVs. The epigenetic silencing and memory are the main sources to control gene expressions, specifically HERVs [13,14,15,16,17,18,19,20].
Empirical studies further unveil that our genome has an intrinsic immunity that protects the integrity of our DNA from pathogen invasion and insult, which is underpinned by epigenetic regulation. We have named the immunity of our DNA as the epigenetic immunity [7,21,22,23]. Studies further show that the immune surveillance is built on the epigenetic silencing and memory. Viruses, or other pathogens, hijack host epigenetic machinery and impair such immune responses [24,25,26,27,28,29,30,31,32,33,34,35,36,37].
Epigenetic immunity occurs in every cell with DNA but not in the cell without it, such as erythrocytes (red blood cells). It protects our DNA against pathogen infection via DNA methylation, histone modification, and noncoding RNA (ncRNA) function. Note that the same molecular mechanism of DNA immunity confers an immune phenomenon that is studied in innate immune cells, firstly in myeloid cells and is termed as trained immunity. The immune phenomena, however, are revealed and studied further beyond the innate immune cells to lymphocytes, and up to the hematopoietic stem and progenitor cells (HSPCs) [38,39,40,41,42,43,44,45,46,47,48,49,50,51].
We aim to study the same molecular mechanism to improve our understanding of human immunity. We propose to define the biomarkers, a relevant immune signaling pathway, and immune effector differentiation that embody the function of epigenetic immunity to prevent, control, and treat viral infections by three deliverables. We focus on HIV as an example due to the historical and scientific reasons [52,53,54,55,56,57,58,59,60].
The first deliverable of this research is the generation of chromatin vaccines (cVacc) (Figure 1). cVacc is a functional gene transcription unit with an enhancer in a nucleosome format that resists nuclease degradation while mediating epigenetic silencing of viral RNA by ncRNA function and the enhancer decommissioning process [7,21,22,23].
- Processing 2-/1-LTR circles by ChIP-seq, etc.
- Exposing the 2-/1-LTR circles to cell cultures
- Colony formation assay for CD4 T-cells
- Testing CD4 T-cells antigen specific function and memory in vitro and in vivo
HIV infects naive CD4 T-cells that have the property of stem cells for self-renewal and multipotency to differentiate into lineage effector cells. The naive CD4 T-cells mimic stem cell asymmetric division upon an antigen stimulation and differentiate into memory and effector immune cells. Enhancer decommissioning processes occur routinely in hematopoietic stem and progenitor cells (HSPCs) when differentiating into linage cells. The same genomic DNA of a naïve CD4 T-cell that is in a progenitor status and after enhancer decommissioning, asymmetrically, differentiates into daughter immune memory and effector cells. A process mimics that of a HSPC.
cVacc functions as an immunogen and acts to prime the naive CD4 T-cell enhancer by triggering signaling transduction pathways of Toll like receptors (TLR), DNA/RNA sensors, interferon, and NFkB to load transcription factors (TF)/RNA polymerase II (RNAPII) to the CD4 T-cell enhancer [61,62,63,64,65,66,67,68,69,70,71,72]. This primed cell enhancer allows CD4 T-cells to rapidly differentiate into memory and effector cells upon encountering HIV.
In other words, the vaccinated CD4 T-cells, specifically the CD4 T memory cells, counteract the HIV proviral enhancer not only by competitively loading TF/RNAPII on their enhancers, but by enhancer RNA (eRNA) and other ncRNA function as well. Both converge to silence the HIV RNA expression while allowing CD4 T-cells to differentiate into a spawn of new daughter anti-HIV memory and effector cells to establish a long lasting anti-HIV immunity. Such a mechanism bestows durability on an anti-HIV vaccine.
Molecularly, taking away the TF and RNAPII, the transcription complex/machinery, from a promoter or enhancer is named decommissioning. Priming the CD4 T-cell enhancer with the rationed transcriptional complex/machinery in a cell and competitively taking away from that of the HIV promoter if the virus comes later, underpins the molecular mechanism of cVacc to prevent HIV infection and reconstitute patient immunity. Cellularly, such a vaccine strategy regenerates the normal function of CD4 T-cells that are destined to execute in human immunity being a commander-in-chief, allowing CD4 T-cells to launch effective immunities protecting host from numerous infections of viruses, bacteria, fungi, etc., while keeping host in an immune homeostasis state.
Moreover, cVacc is made from the aborted, unintegrated 1-LTR and 2-LTR circles that are wrapped with histones and resist the nuclease degradation [73,74,75,76]. This feature, differing from an mRNA vaccine that needs a cold-chain to maintain its function, allows cVacc to be managed and administered wider via ordinary methods to prevent and treat viral infections. cVacc can be applied to the routes of viral entry up to the target cells without jabbing, and aiming to elicit IgA, IgM, IgG, and innate, adaptive, as well as epigenetic immunity in a systemic manner. Such a strategy not only stimulates local, mucosa, or tissue specific immunity but the entire host immunity as well.
These features, specifically silencing the viral mRNA expression and cutting off downstream mutagenesis, allow cVacc to be the next generation vaccine compared to COVID mRNA vaccines. Additionally in COVID, mRNA vaccines work better than the adenovirus-vector vaccines, and are reported to have engaged in the epigenetic responses in inhibition of the viral infections [7,8,9,10,11,12]. The stimulation of epigenetic immune responses, however, differs from eliciting the epigenetic immunity. cVacc elicits the epigenetic immunity [7,21,22,23].
To summarize, the strength of cVacc stems from its formats and components in priming the TF/RNAPII transcriptional complex to bind to the CD4 T-cell super-enhancers. Such immunization strategy not only decommissions the HIV promoter and silences the HIV RNA production, but also develops antigen specific CD4 T-cells in multi-lineages to reconstitute patient immunity.
Sequentially, immunization by cVacc generates the 2nd deliverable of the study, which are the recombination-activating gene 2 protein (RAG2) regulated antigen specific CD4 T-cells. In mammals, RAG2 is a far more evolved immune machinery than the CRISPR-Cas in bacteria, since RAG2 directs the specific antigen binding receptor formation of both T and B cells after an antigen or immunogen stimulation [77,78,79,80,81,82,83].
The essence of vaccination is to generate antigen specific CD4 T-cells, including memory CD4 T- cells. The primed naïve CD4 T-cells differentiate into antigen specific CD4 T-cells with their specific antigen-binding receptors, i.e., T cell receptors (TCRs). Based on the known and emerging knowledge of T and B lymphocytes, a class switch recombination process occurs in the maturation of both cells, specifically in formation of specific antigen binding receptors [77,78,79,80,81,82,83].
Studies show that RAG2 plays a pivotal role in the T and B lymphocyte maturation and their antigen binding receptor formations after stimulation by immunogens. The function of RAG2, however, is cell cycle dependent [77,78,79,80,81,82,83]. Ours and others experimental results have shown that p21Cip1Waf1Sdi1 (p21), a single cell checkpoint protein, not only restricts HIV infection in HSPCs, CD4 T-cells, and macrophages, but controls immune cell cycle status as well [53,84,85,86,87,88,89,90,91,92,93]. Through regulating the p21-signalosome pathway, we can regulate the function of RAG2, generate antigen specific CD4 T-cells for immunotherapy, and reconstitute patient immunity (Figure 2). This differs from CAR T cells that are TCR indirectly specific and with significant side effects [94,95,96,97,98].
With the same molecular mechanisms and a technological tweaking, immunization via cVacc produces a third deliverable – generation of RAG2 regulated antigen specific CD4 T follicular helper cells (Tfh) and B cells. B lymphocytes generate antibodies and play important roles in host antiviral immunities. From the HIV/AIDS model, we understand that the CD4 (Tfh) cell is indispensable for B cell function, specifically in producing antibodies [84,85,86,87,88,99,100,101,102,103]. This CD4 (Tfh) cell helped B cell function is, however, governed by the p21-signalsome-RAG2 pathway also (Figure 2) [54,84,85,86,87,88,89,90,91,92,93,99,100,101,102,103]. In addition to generating the antigen specific CD4 (Tfh) cells and using tuned techniques, we will generate antigen specific B cells that exert humoral immunity against the viral infection.
Note that RAG2, along with RAG1, is a key player in B cell antigen receptor (BCR) formation by initiating V(D)J recombination, a process that reshuffles the immunoglobulin gene segments to generate a diverse range of BCRs in responding to pathogens [83,104,105,106,107]. The function of RAG2 in formation of BCRs that bind to pathogenic antigen or immunogen differs from other B cell surface receptor formation or the antibody maturation regulated by activation-induced cytidine deaminase (AID) [77,78,79,80,81,82,83,104,105,106,107].
In the above study, we will apply technologies including but not limited to Chromatin Immunoprecipi-tation Sequencing (ChIP-seq), ChIP and lambda exonuclease digestion (exo) followed by high-throughput sequencing (ChIP-exo), Multi-omics analyses, Methylation-specific PCR, Cell colony formation assay, Flow cytometry, Stem cell culture, Organoid culture, Humanized mouse model, and Artificial intelligence (AI) simulated clinical testing of effective HIV vaccines. We aim to follow in the footsteps of pioneers, continue on a national journey, to make a scientific progress and fulfill a duty that the era has bestowed on our generation.
Discussion, Conclusion, and Future Direction
The vaccinology needs a leap forward based on the knowledge from the NIH human genome project and the epigenomics road map consortium. Notably, the achievements on the human genome’s research occur at the same time as that of HIV/AIDS research [13,14,15,16,17,18,19,20]. The research on HIV/AIDS, a naturally occurring module for loss-of-function in human immunity, has greatly facilitated studies on human immunodeficiency at a scale of DNA sequencing to epigenetic immunity that may lead to an HIV cure.
Specifically, cVacc elicits a kinetic replenishment of antigen specific immune cells, and sets a foundation of vaccine durability that serves as a remedy against “vaccine breakthrough”. Vaccine breakthrough is a recent emerging phenomenon occurring to vaccination/immunization [7,8,9,10,11,12]. Using a cVacc to elicit the immunity of our DNA – the epigenetic immunity, not only generates an effective HIV vaccine but other vaccines preventing and treating a variety of diseases to benefit generations yet to come.
References
- Glynn, I. , Glynn, J. The Life and Death of Smallpox. New York: Cambridge University Press, 2004.
- U.S. Military and Vaccine History. HistoryOfVaccines.org. Available online: https://historyofvaccines.org › how-are-vaccines-made (accessed on 2025 Mar 30).
- Joklik, Willett, Amos, Wilfer. Zinsser Microbiology. 19th Edition. Appleton & Lange, 1988.
- Plotkin, SA. Vaccines: past, present and future. Nat Med. 2005 Apr;11(4 Suppl):S5-11.
- Norrby, E. Yellow fever and Max Theiler: the only Nobel Prize for a virus vaccine. J. Exp. Med. 2007, 204, 2779–2784. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.C. Adenovirus 4 and 7 Vaccine: New Body Armor for U.S. Marine Corps Officer Trainees. J. Infect. Dis. 2019, 221, 685–686. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Askenase, P.; Jaenisch, R.; Crumpacker, C.S. Approaches to pandemic prevention – the chromatin vaccine. Front. Immunol. 2023, 14, 1324084. [Google Scholar] [CrossRef]
- Talotta, R. COVID-19 mRNA vaccines as hypothetical epigenetic players: Results from an in silico analysis, considerations and perspectives. Vaccine 2023, 41, 5182–5194. [Google Scholar] [CrossRef] [PubMed]
- Maurya R, Shamim U, Mishra P, Swaminathan A, Raina A, Tarai B, Budhiraja S, Pandey R. Intertwined Dysregulation of Ribosomal Proteins and Immune Response Delineates SARS-CoV-2 Vaccination Breakthroughs. Microbiol Spectr. 2023 Jun 15;11(3):e0429222.
- Mortari, E.P.; Ferrucci, F.; Zografaki, I.; Carsetti, R.; Pacelli, L. T and B cell responses in different immunization scenarios for COVID-19: a narrative review. Front. Immunol. 2025, 16, 1535014. [Google Scholar] [CrossRef]
- Samaan, P.; Korosec, C.S.; Budylowski, P.; Chau, S.L.L.; Pasculescu, A.; Qi, F.; Delgado-Brand, M.; Tursun, T.R.; Mailhot, G.; Dayam, R.M.; et al. mRNA vaccine-induced SARS-CoV-2 spike-specific IFN-γ and IL-2 T-cell responses are predictive of serological neutralization and are transiently enhanced by pre-existing cross-reactive immunity. J. Virol. 2025, 99, e0168524. [Google Scholar] [CrossRef]
- Simonis, A.; Theobald, S.J.; E Koch, A.; Mummadavarapu, R.; Mudler, J.M.; Pouikli, A.; Göbel, U.; Acton, R.; Winter, S.; Albus, A.; et al. Persistent epigenetic memory of SARS-CoV-2 mRNA vaccination in monocyte-derived macrophages. Mol. Syst. Biol. 2025, 21, 341–360. [Google Scholar] [CrossRef]
- Green, E.D.; Watson, J.D.; Collins, F.S. Human Genome Project: Twenty-five years of big biology. Nature 2015, 526, 29–31. [Google Scholar] [CrossRef]
- Stein, R.A.; DePaola, R.V. Human endogenous retroviruses: our genomic fossils and companions. Physiol. Genom. 2023, 55, 249–258. [Google Scholar] [CrossRef]
- Roadmap Epigenomics Consortium; Kundaje, A. ; Meuleman, W.; Ernst, J.; Bilenky, M.; Yen, A.; Heravi-Moussavi, A.; Kheradpour, P.; Zhang, Z.; et al. Integrative analysis of 111 reference human epigenomes. Nature 2015, 518, 317–330. [Google Scholar] [CrossRef]
- The ENCODE Project Consortium; Moore, J. E.; Purcaro, M.J.; Pratt, H.E.; Epstein, C.B.; Shoresh, N.; Adrian, J.; Kawli, T.; Davis, C.A.; Dobin, A.; et al. Expanded encyclopaedias of DNA elements in the human and mouse genomes. Nature 2020, 583, 699–710. [Google Scholar] [CrossRef]
- Jou, J.; Gabdank, I.; Luo, Y.; Lin, K.; Sud, P.; Myers, Z.; Hilton, J.A.; Kagda, M.S.; Lam, B.; O'Neill, E.; et al. The ENCODE Portal as an Epigenomics Resource. Curr. Protoc. Bioinform. 2019, 68, e89–e89. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.A.; Hitz, B.C.; Sloan, C.A.; Chan, E.T.; Davidson, J.M.; Gabdank, I.; Hilton, J.A.; Jain, K.; Baymuradov, U.K.; Narayanan, A.K.; et al. The Encyclopedia of DNA elements (ENCODE): data portal update. Nucleic Acids Res. 2018, 46, D794–D801. [Google Scholar] [CrossRef]
- Pehrsson, E.C.; Choudhary, M.N.K.; Sundaram, V.; Wang, T. The epigenomic landscape of transposable elements across normal human development and anatomy. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Amin, V.; Harris, R.A.; Onuchic, V.; Jackson, A.R.; Charnecki, T.; Paithankar, S.; Subramanian, S.L.; Riehle, K.; Coarfa, C.; Milosavljevic, A. Epigenomic footprints across 111 reference epigenomes reveal tissue-specific epigenetic regulation of lincRNAs. Nat. Commun. 2015, 6, 6370–6370. [Google Scholar] [CrossRef]
- Zhang JL, Crumpacker CS. Towards a Cure, does host immunity play a role? mSphere. 2017; 2:e00138-17 (and references therein).
- Zhang JL, Crumpacker CS. An integrative immunobiology and inflammation study on cytomegalovirus. Integrative Immunobiology and Inflammation 2016; [S.l.] v. 1, n. 1. [Available online to 2018].
- Zhang J, Crumpacker C. HIV UTR, LTR, and Epigenetic Immunity. Viruses. 2022;18;14(5): 1084.
- Cullen, H.; Schorn, A.J. Endogenous Retroviruses Walk a Fine Line between Priming and Silencing. Viruses 2020, 12, 792. [Google Scholar] [CrossRef]
- Jakobsson, J.; Vincendeau, M. SnapShot: Human endogenous retroviruses. Cell 2022, 185, 400–400.e1. [Google Scholar] [CrossRef]
- Alcazer, V.; Bonaventura, P.; Tonon, L.; Michel, E.; Mutez, V.; Fabres, C.; Chuvin, N.; Boulos, R.; Estornes, Y.; Maguer-Satta, V.; et al. HERVs characterize normal and leukemia stem cells and represent a source of shared epitopes for cancer immunotherapy. Am. J. Hematol. 2022, 97, 1200–1214. [Google Scholar] [CrossRef]
- Mantovani, F.; Kitsou, K.; Magiorkinis, G. HERVs: Expression Control Mechanisms and Interactions in Diseases and Human Immunodeficiency Virus Infection. Genes 2024, 15, 192. [Google Scholar] [CrossRef]
- Hurst, T.P.; Magiorkinis, G. Epigenetic Control of Human Endogenous Retrovirus Expression: Focus on Regulation of Long-Terminal Repeats (LTRs). Viruses 2017, 9, 130. [Google Scholar] [CrossRef]
- Shah, A.H.; Gilbert, M.; E Ivan, M.; Komotar, R.J.; Heiss, J.; Nath, A. The role of human endogenous retroviruses in gliomas: from etiological perspectives and therapeutic implications. Neuro-Oncology 2021, 23, 1647–1655. [Google Scholar] [CrossRef]
- Torre, D.; Fstkchyan, Y.S.; Ho, J.S.Y.; Cheon, Y.; Patel, R.S.; Degrace, E.J.; Mzoughi, S.; Schwarz, M.; Mohammed, K.; Seo, J.-S.; et al. Nuclear RNA catabolism controls endogenous retroviruses, gene expression asymmetry, and dedifferentiation. Mol. Cell 2023, 83, 4255–4271.e9. [Google Scholar] [CrossRef] [PubMed]
- Tchasovnikarova, I.A.; Timms, R.T.; Matheson, N.J.; Wals, K.; Antrobus, R.; Göttgens, B.; Dougan, G.; Dawson, M.A.; Lehner, P.J. Epigenetic silencing by the HUSH complex mediates position-effect variegation in human cells. Science 2015, 348, 1481–1485. [Google Scholar] [CrossRef]
- Meyer TJ, Rosenkrantz JL, Carbone L, Chavez SL. Endogenous Retroviruses: With Us and against Us. Front Chem. 2017 Apr 7;5:23.
- Reiss, D.; Mager, D.L. Stochastic epigenetic silencing of retrotransposons: Does stability come with age? Gene 2007, 390, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, M.; Faure-Dupuy, S. Virus hijacking of host epigenetic machinery to impair immune response. J. Virol. 2023, 97, e0065823. [Google Scholar] [CrossRef]
- Capparelli R, Iannelli D. Epigenetics and Helicobacter pylori. Int J Mol Sci. 2022 Feb 3;23(3):1759.
- Kim, E.-J.; Ma, X.; Cerutti, H. Gene silencing in microalgae: Mechanisms and biological roles. Bioresour. Technol. 2015, 184, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Adhya, D.; Basu, A. Epigenetic modulation of host: new insights into immune evasion by viruses. J. Biosci. 2010, 35, 647–663. [Google Scholar] [CrossRef]
- Sun JC, Lopez-Verges S, Kim CC, DeRisi JL, Lanier LL. NK cells and immune „memory”. J Immunol. 2011 Feb 15;186(4):1891-7.
- Netea, M.G.; Quintin, J.; van der Meer, J.W. Trained Immunity: A Memory for Innate Host Defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.B.; Ifrim, D.C.; Saeed, S.; Jacobs, C.; van Loenhout, J.; de Jong, D.; Stunnenberg, H.G.; et al. Bacille Calmette-Guérin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc. Natl. Acad. Sci. 2012, 109, 17537–17542. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.; Jacobs, C.; Xavier, R.J.; van der Meer, J.W.; van Crevel, R.; Netea, M.G. BCG-induced trained immunity in NK cells: Role for non-specific protection to infection. Clin. Immunol. 2014, 155, 213–219. [Google Scholar] [CrossRef]
- Stevens, W.B.; Netea, M.G.; Kater, A.P.; van der Velden, W.J. 'Trained immunity: consequences for lymphoid malignancies. Haematologica 2016, 101, 1460–1468. [Google Scholar] [CrossRef] [PubMed]
- Gourbal, B.; Pinaud, S.; Beckers, G.J.M.; Van Der Meer, J.W.M.; Conrath, U.; Netea, M.G. Innate immune memory: An evolutionary perspective. Immunol. Rev. 2018, 283, 21–40. [Google Scholar] [CrossRef]
- Groh, L.; Netea, M.G.; Riksen, N.P.; Keating, S.T. Getting to the Marrow of Trained Immunity. Epigenomics 2018, 10, 1151–1154. [Google Scholar] [CrossRef]
- van der Heijden CDCC, Noz MP, Joosten LAB, Netea MG, Riksen NP, Keating ST. Epigenetics and Trained Immunity. Antioxid Redox Signal. 2018 Oct 10;29(11):1023-1040.
- Fok, E.T.; Davignon, L.; Fanucchi, S.; Mhlanga, M.M. The lncRNA Connection Between Cellular Metabolism and Epigenetics in Trained Immunity. Front. Immunol. 2019, 9, 3184. [Google Scholar] [CrossRef]
- Domínguez-Andrés, J.; AB Joosten, L.; Netea, M.G. Induction of innate immune memory: the role of cellular metabolism. Curr. Opin. Immunol. 2019, 56, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.M.; Mills, K.H.G.; Basdeo, S.A. The Effects of Trained Innate Immunity on T Cell Responses; Clinical Implications and Knowledge Gaps for Future Research. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, K.T.M.; Mills, K.H. Trained Innate Immunity in Hematopoietic Stem Cell and Solid Organ Transplantation. Transplantation 2021, 105, 1666–1676. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, P.; Jayaseelan, V.P. Implication of epitranscriptomics in trained innate immunity and COVID-19. Epigenomics 2021, 13, 1077–1080. [Google Scholar] [CrossRef]
- Kain BN, Tran BT, Luna PN, Cao R, Le DT, Florez MA, Maneix L, Toups JD, Morales-Mantilla DE, Koh S, Han H, Jaksik R, Huang Y, Catic A, Shaw CA, King KY. Hematopoietic stem and progenitor cells confer cross-protective trained immunity in mouse models. iScience. 2023 Aug 9;26(9):107596.
- Stier, S.; Cheng, T.; Forkert, R.; Lutz, C.; Dombkowski, D.M.; Zhang, J.L.; Scadden, D.T. Ex vivo targeting of p21Cip1/Waf1 permits relative expansion of human hematopoietic stem cells. Blood 2003, 102, 1260–1266. [Google Scholar] [CrossRef]
- Zhang, J.; Attar, E.; Cohen, K.; Crumpacker, C.; Scadden, D. Silencing p21Waf1/Cip1/Sdi1 expression increases gene transduction efficiency in primitive human hematopoietic cells. Gene Ther. 2005, 12, 1444–1452. [Google Scholar] [CrossRef]
- Zhang, J.; Scadden, D.T.; Crumpacker, C.S. Primitive hematopoietic cells resist HIV-1 infection via p21Waf1/Cip1/Sdi1. J. Clin. Investig. 2007, 117, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Cheng J, Ke Q, Jin Z, Wang H, Kocher O, Morgan J, Crumpacker C, Zhang JL. Cytomegalo-virus Infection Causes an Increase of Arterial Blood Pressure. PLoS Pathog 2009; 5(5)e: 1000427.
- Zhang, J.; Crumpacker, C.S. Hematopoietic stem and progenitor cells in HIV/AIDS and immune reconstitution. Cell Res. 2010, 20, 745–747. [Google Scholar] [CrossRef] [PubMed]
- Zhang JL, Poznansky M, Crumpacker CS. Naïve and memory CD4+ T-cells in HIV eradication and immunization. J Infect Dis 2012; 206(4): 617-8.
- Zhang JL, Crumpacker CS. Eradication of HIV and Cure of AIDS, Now and How? Front Immunol 2013; 4:337.
- Zhang, J.; Crumpacker, C. Hematopoietic Stem and Immune Cells in Chronic HIV Infection. Stem Cells Int. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Askenase, P.; Crumpacker, C.S. Systems Vaccinology in HIV Vaccine Development. Vaccines 2022, 10, 1624. [Google Scholar] [CrossRef]
- Hirahara, K.; Vahedi, G.; Ghoreschi, K.; Yang, X.-P.; Nakayamada, S.; Kanno, Y.; O’shea, J.J.; Laurence, A. Helper T-cell differentiation and plasticity: insights from epigenetics. Immunology 2011, 134, 235–245. [Google Scholar] [CrossRef]
- Nakayamada, S.; Kanno, Y.; Takahashi, H.; Jankovic, D.; Lu, K.T.; Johnson, T.A.; Sun, H.-W.; Vahedi, G.; Hakim, O.; Handon, R.; et al. Early Th1 Cell Differentiation Is Marked by a Tfh Cell-like Transition. Immunity 2011, 35, 919–931. [Google Scholar] [CrossRef]
- Nakayamada, S.; Takahashi, H.; Kanno, Y.; O'Shea, J.J. Helper T cell diversity and plasticity. Curr. Opin. Immunol. 2012, 24, 297–302. [Google Scholar] [CrossRef]
- Hirahara, K.; Poholek, A.; Vahedi, G.; Laurence, A.; Kanno, Y.; Milner, J.D.; O’shea, J.J. Mechanisms underlying helper T-cell plasticity: Implications for immune-mediated disease. J. Allergy Clin. Immunol. 2013, 131, 1276–1287. [Google Scholar] [CrossRef]
- Wei, G. , Wei L., Zhu J., Zang C., Hu-Li J., Yao Z., Cui K., Kanno Y., Roh T.Y., Watford W.T., et al. Global mapping of H3K4me3 and H3K27me3 reveals specificity and plasticity in lineage fate determination of differentiating CD4+ T cells. Immunity. 2009;30:155–167.
- Vahedi, G.; Kanno, Y.; Sartorelli, V.; O'Shea, J.J. Transcription factors and CD4 T cells seeking identity: masters, minions, setters and spikers. Immunology 2013, 139, 294–298. [Google Scholar] [CrossRef]
- Johnson, J.L.; Georgakilas, G.; Petrovic, J.; Kurachi, M.; Cai, S.; Harly, C.; Pear, W.S.; Bhandoola, A.; Wherry, E.J.; Vahedi, G. Lineage-Determining Transcription Factor TCF-1 Initiates the Epigenetic Identity of T Cells. Immunity 2018, 48, 243–257. [Google Scholar] [CrossRef]
- Durek, P.; Nordström, K.; Gasparoni, G.; Salhab, A.; Kressler, C.; de Almeida, M.; Bassler, K.; Ulas, T.; Schmidt, F.; Xiong, J.; et al. Epigenomic Profiling of Human CD4+ T Cells Supports a Linear Differentiation Model and Highlights Molecular Regulators of Memory Development. Immunity 2016, 45, 1148–1161. [Google Scholar] [CrossRef]
- Issuree, P.D. , Day K., Au C., Raviram R., Zappile P., Skok J.A., Xue H.H., Myers R.M., Littman D.R. Stage-specific epigenetic regulation of CD4 expression by coordinated enhancer elements during T cell development. Nat. Commun. 2018;9:3594.
- Maqbool, M.A.; Pioger, L.; El Aabidine, A.Z.; Karasu, N.; Molitor, A.M.; Dao, L.T.; Charbonnier, G.; van Laethem, F.; Fenouil, R.; Koch, F.; et al. Alternative Enhancer Usage and Targeted Polycomb Marking Hallmark Promoter Choice during T Cell Differentiation. Cell Rep. 2020, 32, 108048. [Google Scholar] [CrossRef] [PubMed]
- Paiano, J.J.; Johnson, J.L.; Vahedi, G. Enhancing our understanding of enhancers in T-helper cells. Eur. J. Immunol. 2015, 45, 2998–3001. [Google Scholar] [CrossRef] [PubMed]
- Witte, S.; O'Shea, J.J.; Vahedi, G. Super-enhancers: Asset management in immune cell genomes. Trends Immunol. 2015, 36, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Bukrinsky, M.; Sharova, N.; Stevenson, M. Human immunodeficiency virus type 1 2-LTR circles reside in a nucleoprotein complex which is different from the preintegration complex. J. Virol. 1993, 67, 6863–6865. [Google Scholar] [CrossRef]
- Cara, A.; Vargas, J.; Keller, M.; Jones, S.; Mosoian, A.; Gurtman, A.; Cohen, A.; Parkas, V.; Wallach, F.; Chusid, E.; et al. Circular Viral DNA and Anomalous Junction Sequence in PBMC of HIV-Infected Individuals with No Detectable Plasma HIV RNA. Virology 2002, 292, 1–5. [Google Scholar] [CrossRef]
- Irwan, I.D.; Karnowski, H.L.; Bogerd, H.P.; Tsai, K.; Cullen, B.R. Reversal of Epigenetic Silencing Allows Robust HIV-1 Replication in the Absence of Integrase Function. mBio 2020, 11. [Google Scholar] [CrossRef]
- Geis, F.K.; Kelenis, D.P.; Goff, S.P. Two lymphoid cell lines potently silence unintegrated HIV-1 DNAs. Retrovirology 2022, 19, 1–10. [Google Scholar] [CrossRef]
- Jiang, H.; Ross, A.E.; Desiderio, S. Cell Cycle-dependent Accumulation in Vivo of Transposition-competent Complexes between Recombination Signal Ends and Full-length RAG Proteins. 279, 8478. [Google Scholar] [CrossRef]
- Li, Z.; Dordai, D.I.; Lee, J.; Desiderio, S. A Conserved Degradation Signal Regulates RAG-2 Accumulation during Cell Division and Links V(D)J Recombination to the Cell Cycle. Immunity 1996, 5, 575–589. [Google Scholar] [CrossRef]
- Lin, W.C.; Desiderio, S. Cell cycle regulation of V(D)J recombination-activating protein RAG-2. Proc. Natl. Acad. Sci. 1994, 91, 2733–2737. [Google Scholar] [CrossRef]
- Matthews AG, Oettinger MA. Regulation of RAG transposition. Adv Exp Med Biol. 2009;650:16-31.
- Teng G, Schatz DG. Regulation and Evolution of the RAG Recombinase. Adv Immunol. 2015;128:1-39.
- Jiang, H.; Chang, F.-C.; Ross, A.E.; Lee, J.; Nakayama, K.; Desiderio, S. Ubiquitylation of RAG-2 by Skp2-SCF Links Destruction of the V(D)J Recombinase to the Cell Cycle. Mol. Cell 2005, 18, 699–709. [Google Scholar] [CrossRef]
- Tanner A, Hallam SJ, Nielsen SJ, Cuadra GI, Berges BK. Development of human B cells and antibodies following human hematopoietic stem cell transplantation to Rag2(-/-)γc(-/-) mice. Transpl Immunol. 2015 Jun;32(3):144-50.
- Fuente, C.; Maddukuri, A.; Kehn, K.; Baylor, S.Y.; Deng, L.; Pumfery, A.; Kashanchi, F. Pharmacological Cyclin-Dependent Kinase Inhibitors as HIV-1 Antiviral Therapeutics. Curr. HIV Res. 2003, 1, 131–152. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Tungaturthi, P.K.; Ayyavoo, V.; Ghafouri, M.; Ariga, H.; Khalili, K.; Srinivasan, A.; Amini, S.; E Sawaya, B. The Role of Vpr in the Regulation of HIV-1 Gene Expression. Cell Cycle 2006, 5, 2626–2638. [Google Scholar] [CrossRef]
- Chen, H.; Li, C.; Huang, J.; Cung, T.; Seiss, K.; Beamon, J.; Carrington, M.F.; Porter, L.C.; Burke, P.S.; Yang, Y.; et al. CD4+ T cells from elite controllers resist HIV-1 infection by selective upregulation of p21. J. Clin. Investig. 2011, 121, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Yu XG, Lichterfeld M. Elite control of HIV: p21 (waf-1/cip-1) at its best. Cell Cycle. 3: 2011;10(19), 2011.
- Saez-Cirion, A.; Hamimi, C.; Bergamaschi, A.; David, A.; Versmisse, P.; Melard, A.; Boufassa, F.; Barre-Sinoussi, F.; Lambotte, O.; Rouzioux, C.; et al. Restriction of HIV-1 replication in macrophages and CD4+ T cells from HIV controllers. Blood 2011, 118, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chao, W.; Saini, M.; Potash, M.J. A Common Path to Innate Immunity to HIV-1 Induced by Toll-Like Receptor Ligands in Primary Human Macrophages. PLOS ONE 2011, 6, e24193. [Google Scholar] [CrossRef]
- Valle-Casuso, J.C.; Allouch, A.; David, A.; Lenzi, G.M.; Studdard, L.; Barré-Sinoussi, F.; Müller-Trutwin, M.; Kim, B.; Pancino, G.; Sáez-Cirión, A. p21 Restricts HIV-1 in Monocyte-Derived Dendritic Cells through the Reduction of Deoxynucleoside Triphosphate Biosynthesis and Regulation of SAMHD1 Antiviral Activity. J. Virol. 2017, 91, e01324–17. [Google Scholar] [CrossRef]
- Zhang J, Crumpacker C. Personal communication & unpublished studies, 2018.
- Osei Kuffour E, Schott K, Jaguva Vasudevan AA, Holler J, Schulz WA, Lang PA, Lang KS, Kim B, Häussinger D, König R, Münk C. USP18 (UBP43) Abrogates p21-Mediated Inhibition of HIV-1. J Virol. 2018 Sep 26;92(20):e00592-18.
- Shi, B.; Sharifi, H.J.; DiGrigoli, S.; Kinnetz, M.; Mellon, K.; Hu, W.; de Noronha, C.M.C. Inhibition of HIV early replication by the p53 and its downstream gene p21. Virol. J. 2018, 15, 53. [Google Scholar] [CrossRef]
- Chohan, K.L.; Siegler, E.L.; Kenderian, S.S. CAR-T Cell Therapy: the Efficacy and Toxicity Balance. Curr. Hematol. Malign- Rep. 2023, 18, 9–18. [Google Scholar] [CrossRef]
- D’Agostino, M.; Raje, N. Anti-BCMA CAR T-cell therapy in multiple myeloma: can we do better? Leukemia 2020, 34, 21–34. [Google Scholar] [CrossRef]
- Majzner, R.G.; Mackall, C.L. Tumor Antigen Escape from CAR T-cell Therapy. Cancer Discov. 2018, 8, 1219–1226. [Google Scholar] [CrossRef]
- Curran, K.J.; Margossian, S.P.; Kernan, N.A.; Silverman, L.B.; Williams, D.A.; Shukla, N.; Kobos, R.; Forlenza, C.J.; Steinherz, P.; Prockop, S.; et al. Toxicity and response after CD19-specific CAR T-cell therapy in pediatric/young adult relapsed/refractory B-ALL. Blood 2019, 134, 2361–2368. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, A.V.; Turtle, C.J. Toxicities of CD19 CAR-T cell immunotherapy. Am. J. Hematol. 2019, 94, S42–S49. [Google Scholar] [CrossRef] [PubMed]
- Brenna, E.; Davydov, A.N.; Ladell, K.; McLaren, J.E.; Bonaiuti, P.; Metsger, M.; Ramsden, J.D.; Gilbert, S.C.; Lambe, T.; Price, D.A.; et al. CD4+ T Follicular Helper Cells in Human Tonsils and Blood Are Clonally Convergent but Divergent from Non-Tfh CD4+ Cells. Cell Rep. 2020, 30, 137–152.e5. [Google Scholar] [CrossRef]
- Mintz, M.A.; Cyster, J.G. T follicular helper cells in germinal center B cell selection and lymphomagenesis. Immunol. Rev. 2020, 296, 48–61. [Google Scholar] [CrossRef]
- Song, W.; Craft, J. T follicular helper cell heterogeneity: Time, space, and function. Immunol. Rev. 2019, 288, 85–96. [Google Scholar] [CrossRef]
- Crotty, S. Follicular helper CD4 T cells (TFH). Annu Rev Immunol. 2011;29:621-63.
- Crotty, S. T Follicular Helper Cell Biology: A Decade of Discovery and Diseases. Immunity 2019, 50, 1132–1148. [Google Scholar] [CrossRef]
- Wesemann, D.R.; Portuguese, A.J.; Meyers, R.M.; Gallagher, M.P.; Cluff-Jones, K.; Magee, J.M.; Panchakshari, R.A.; Rodig, S.J.; Kepler, T.B.; Alt, F.W. Microbial colonization influences early B-lineage development in the gut lamina propria. Nature 2013, 501, 112–115. [Google Scholar] [CrossRef]
- Braams, M.; Pike-Overzet, K.; Staal, F.J.T. The recombinase activating genes: architects of immune diversity during lymphocyte development. Front. Immunol. 2023, 14, 1210818. [Google Scholar] [CrossRef]
- Sandel, P.C.; Monroe, J.G. Negative Selection of Immature B Cells by Receptor Editing or Deletion Is Determined by Site of Antigen Encounter. Immunity 1999, 10, 289–299. [Google Scholar] [CrossRef]
- Maës, J.; Caspi, Y.; Rougeon, F.; Haimovich, J.; Goodhardt, M. Secondary V(D)J Rearrangements and B Cell Receptor-Mediated Down-Regulation of Recombination Activating Gene-2 Expression in a Murine B Cell Line. J. Immunol. 2000, 165, 703–709. [Google Scholar] [CrossRef]
Figure 1.
How to make cVacc.
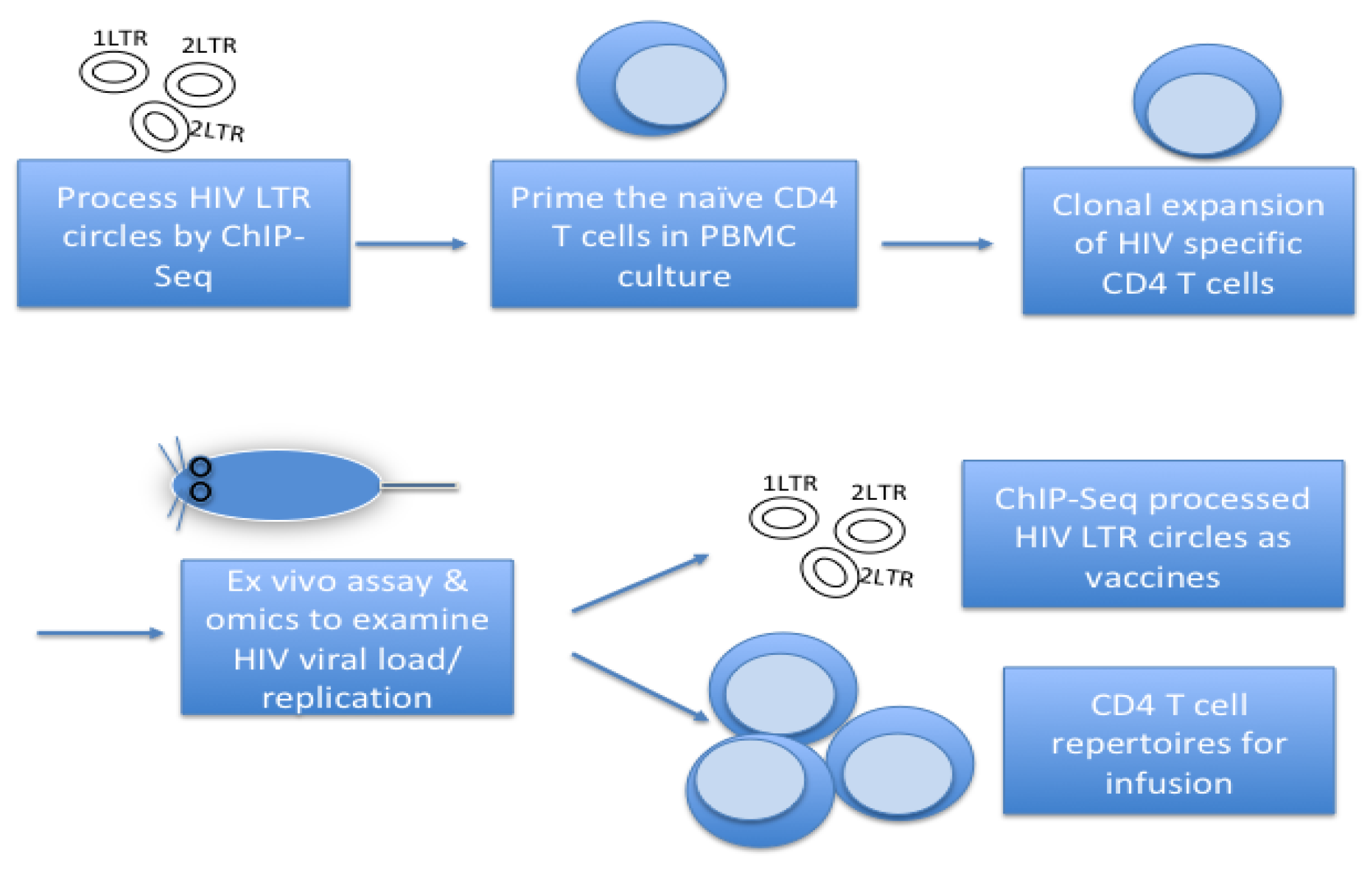
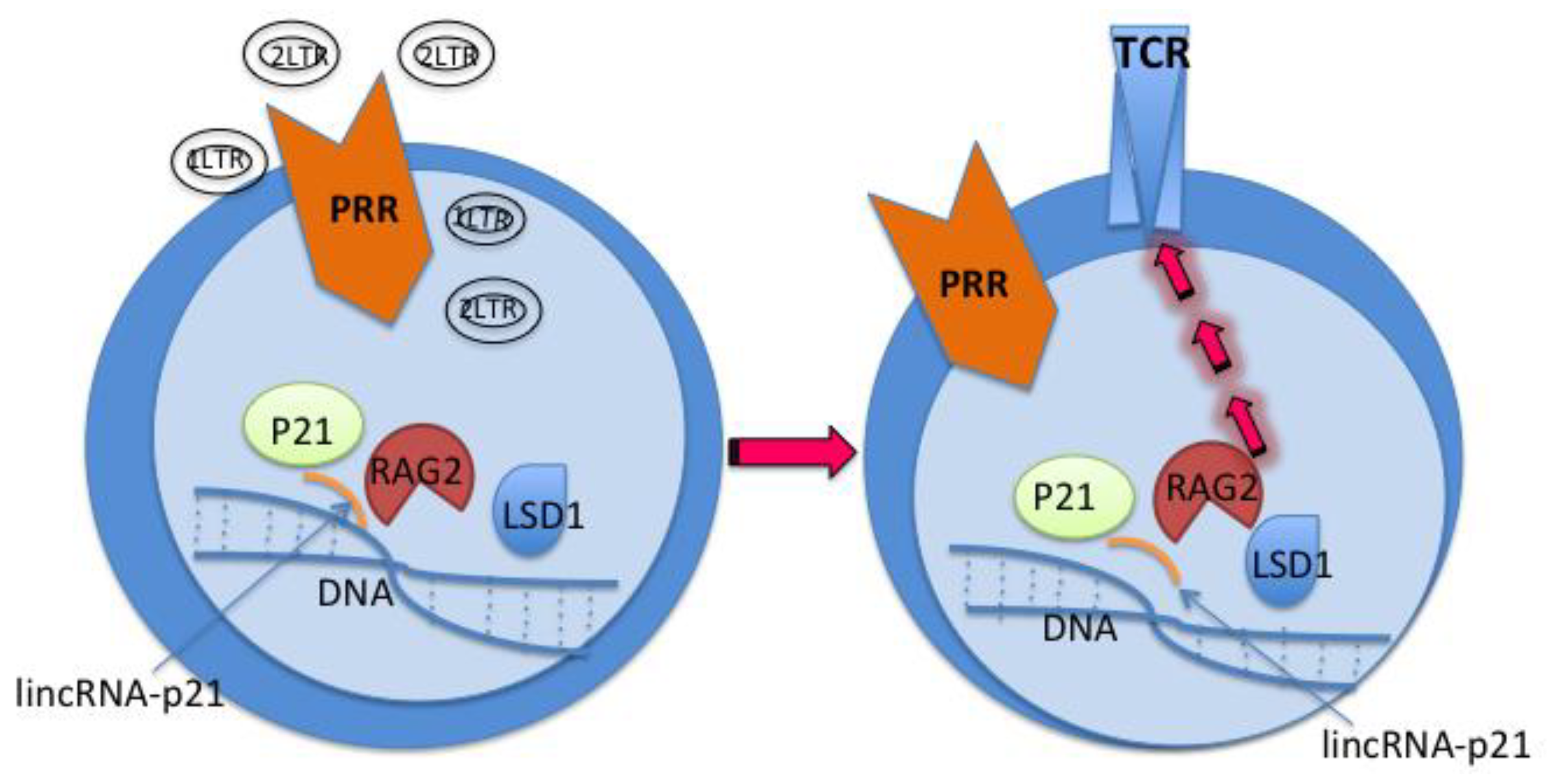
Figure 2.
Developing HIV specific T-cells & B-cells via the p21 signalosome pathway*.PRR: pattern recognition receptors consisted of sensor like receptors - Toll like receptors, DNA/RNA sensors, etc. TCR: T cell antigen receptor. RAG2: recombination activating gene 2 protein. LSD1: lysine specific demethylase 1. *The posited pathway elicited by cVacc is derived from our preliminary, unpublished studies. These can be further confirmed with new lab resources.
Figure 2.
Developing HIV specific T-cells & B-cells via the p21 signalosome pathway*.PRR: pattern recognition receptors consisted of sensor like receptors - Toll like receptors, DNA/RNA sensors, etc. TCR: T cell antigen receptor. RAG2: recombination activating gene 2 protein. LSD1: lysine specific demethylase 1. *The posited pathway elicited by cVacc is derived from our preliminary, unpublished studies. These can be further confirmed with new lab resources.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



