
Submitted:
08 May 2025
Posted:
09 May 2025
You are already at the latest version
Abstract
Coastal wetlands play important environmental roles, but their hydrogeomorphological dynamics remain poorly understood under extreme climatic events. This study aimed to characterize the temporal and spatial variability of hydrogeomorphological attributes (vegetation, water, and soil) in the wetlands of Lagoa do Peixe National Park, Brazil. The methodology involved applying Principal Component Analysis in both temporal (T) and spatial (S) modes, and correlating decomposed spectral indices (NDVI, MNDWI, BI2) for each attribute to identify variability patterns. The results show that PCA highlighted the sensitivity of the Lagoon Terraces and the Lagoon Fringe, where the low slope amplifies hydrological variations. The soil exhibited greater complexity, reflecting its textural composition and distinct temporal response to the water balance. Vegetation and water showed strong correlation, with seasonal dynamics influenced by ENSO events (El Niño/La Niña). In the time series (2019–2024), precipitation and evapotranspiration exhibited distinct seasonal patterns, influenced by an extended La Niña period (2020–2023), characterized by water deficit. The temporal variability in response to climate extremes has implications for the conservation of these ecosystems, while spatial variability is more closely related to hydrogeomorphological characteristics.
Keywords:
hydrogeomorphological attributes
; remote sensing
; seasonal variability
; coastal wetlands
; Principal Component Analysis
; spectral indices
1. Introduction
Coastal wetlands play a fundamental role in providing habitats, mitigating floods [1,2,3], and in carbon sequestration and storage [4,5,6], contributing approximately 12% to the global carbon stock [7]. Moreover, they hold high economic value [8], making them essential ecosystems for both environmental and economic sustainability.
Despite their recognized environmental importance, studies indicate a 30% loss of global wetlands [9,10,11]. Between 1700 and 2020, wetland losses ranged from 21% to 35% [12]. From 1980 to 2014, 47% of Ramsar sites experienced suppression of associated wetlands [13]. Estimates of global wetland area range from 0.54 to 21.26 million km², but the specific spatial consistency of the wetland class is less than 1% [14]. However, discrepancies between estimates may partly reflect differences in wetland definitions. International treaties, such as the Ramsar Convention and the Kyoto Protocol, highlight the need for wetland conservation [9]. Moreover, preserving these ecosystems is essential for achieving the Sustainable Development Goals (SDGs) [15].
Climate change compromises the functioning of coastal wetlands [16], leading to alterations in hydrogeomorphology across multiple dimensions, such as changes in hydrological regimes, sea level rise, freshwater inflow, and the input of sediments and nutrients [17], these changes affect biodiversity [4], the biogeochemistry of these environments, and their capacity for carbon storage [16]. Characterizing the water balance can serve as a key parameter for analyzing wetland variability, while also providing valuable insights into the impacts of climate change on these hydrogeomorphological systems [18].
Qualitative and quantitative indicators of the hydrogeomorphological characteristics of wetlands, derived from optical sensors—such as spectral indices—enable the analysis of variability patterns and change trends in hydrogeomorphological attributes that are essential to the ecological health of these ecosystems [19].
Wetland variability patterns can be analyzed using multitemporal image transformations, such as Principal Component Analysis (PCA). PCA has been employed to detect changes in the wetlands of Zambia [20]; to identify spectral bands for differentiating wetland vegetation species in the Great Lakes region, United States [21]; to assess degradation in the Göksu Delta, Mediterranean coast of Turkey [22]; to identify spatiotemporal changes and monitor wetlands in the Yellow River Delta, China [23,24]; in Nigeria, to evaluate impacts on vegetation [24]; in spectral mixture models to quantify vegetation and water loss in the Andrade Mesa wetlands, on the U.S.–Mexico border [25]; and in Brazil, to analyze the spatiotemporal variability of vegetation cover in the Pantanal [26]; s well as to characterize the Taquari Megafan in the Pantanal [27].
Other authors have used PCA Orientation in T and S modes to identify recurring temporal and spatial patterns over time. This approach was applied in the study of the Marismas Nacionales coastal ecosystem (northwest Mexico) to identify patterns of variation in mangrove canopies [19]; and in Poyang Lake, China, it was used to delineate dynamic cover types [28].
However, PCA applications have been nearly nonexistent in understanding the spatial and temporal dynamics of hydrogeomorphological attributes in subtropical coastal wetlands. Based on these considerations, the aim of this study is to characterize the temporal and spatial variability of hydrogeomorphological attributes (vegetation, water, and soils) in the coastal wetlands of the Lagoa do Peixe National Park, a significant Ramsar site in the southernmost region of Brazil.
2. Materials and Methods
2.1. Study Area
In Brazil, coastal wetlands are distributed along the entire coastline, spanning both tropical and subtropical zones. They encompass a wide variety of ecosystems, including dunes, restingas, sandy beaches, rocky shores, lagoons, estuaries, salt marshes, mangroves, and coral reefs [29]. Due to its morphogenesis, the Coastal Plain of Rio Grande do Sul hosts the highest concentration of coastal wetlands [30]. However, trends of ecosystem loss have been identified throughout the period from 1985 to 2021 [31].
The Coastal Plain of Rio Grande do Sul records four episodes of transgression and regression during the Quaternary period, which gave rise to four depositional systems known as the Lagoon-Barrier System. Three of these are of Pleistocene age (I, II, and III), and one is of Holocene age (IV) [32,33,34].
The Lagoa do Peixe National Park is located in the central segment of the Coastal Plain of Rio Grande do Sul, between the Lagoa dos Patos and the Atlantic Ocean, within the municipalities of Tavares and Mostardas (Figure 1). It is a strictly protected Conservation Unit, established in 1986. Due to its importance for the conservation of coastal ecosystems and migratory bird habitats, it was designated as a Ramsar Site in 1993. Currently, it is part of the Atlantic Forest Biosphere Reserve and the Western Hemisphere Shorebird Reserve Network [35,36]. Coastal wetlands within Lagoa do Peixe National Park occupy 47% of the park’s area, and Lagoa do Peixe—the main water body—periodically connects with the Atlantic Ocean [37].
Within the park, five geological-geomorphological features are present: the Paleocliff of Barrier III; Colluvial Deposits of Barrier III; Lagoon Terrace I; Lagoon Terrace II; and Aeolian Dunes of Barrier IV [38]. The predominant soil type is Melanic Gleysol, a hydromorphic soil developed from recent, unconsolidated Holocene sediments, with clayey, clayey-sandy, and sandy textures [39]. Nine hydrogeomorphological compartments have been defined: Eolic Dunes, Lagoon Fringe, Lacustrine Fringe, Lacustrine, Lagoon Estuarine, Lacustrine Terrace, Lagoon Terrace, Depression, and Slope [40]. These areas are susceptible to flood pulses associated with extreme precipitation events.
Annual rainfall in the region ranges from 1,200 to 1,500 mm, with higher totals during winter and spring, occasionally exceeding the historical average. Precipitation is primarily driven by frontal systems resulting from the interaction between tropical maritime (mT) and polar maritime (mP) air masses, as well as by anomalies associated with phenomena such as the El Niño–Southern Oscillation (ENSO) and the Southern Annular Mode (SAM) [39,40]. El Niño events lead to above-average rainfall, while La Niña causes negative anomalies, significantly reducing water levels in Lagoa do Peixe [42,43,44,45].
The vegetation cover consists of Pioneer Formations or Restinga [34], influenced by marine and lacustrine conditions. Wet grasslands, swamp forests, marshes, tidal flats (marismas), and lagoonal water bodies are also present. Areas of Pinus spp. silviculture—an invasive species—have spread over the park’s sandy terrains [46], impacting the natural stability of dunes and contributing to the siltation of wetlands [47].
2.2. Data and Procedures
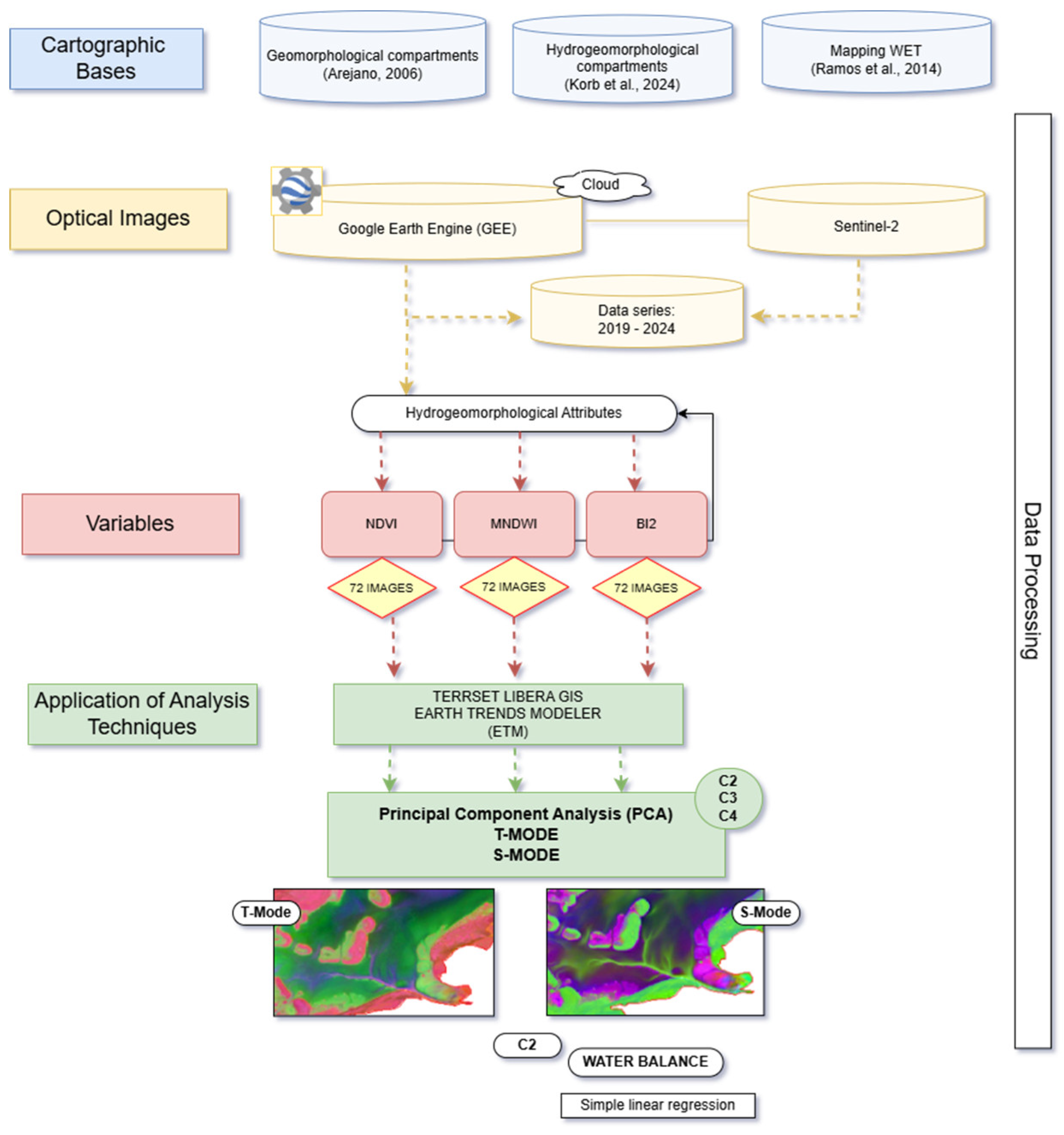
This study adopted the following cartographic bases: (a) geomorphological compartments [38]; (b) hydrogeomorphological compartments [40]; (c) wetland mapping by the Zoobotanical Foundation of Rio Grande do Sul [30]; (d) precipitation data from the Climate Hazards Group InfraRed Precipitation with Stations (CHIRPS) product, using Java Script based code structures in the Google Earth Engine (GEE) Code Editor platform; and, (e) evapotranspiration data from the Terra Moderate Resolution Imaging Spectroradiometer (MODIS) product, MOD16A2GF, accessed via the GEE platform.
Multispectral images from the Sentinel-2 satellite (MSI – Multispectral Instrument) were used, covering the period from 2019 to 2024, with a spatial resolution of 10 m, obtained through Google Earth Engine. One image per month was selected for each year of the time series, encompassing the four climatic seasons and different phases of the El Niño–Southern Oscillation (ENSO) variability (El Niño, La Niña, and Neutral).
Spectral indices related to hydrogeomorphological attributes were calculated on the GEE platform (Table 1): the Normalized Difference Vegetation Index (NDVI) for vegetation, the Modified Normalized Difference Water Index (MNDWI) for water, and the Second Brightness Index (BI2) for soil. The time series consisted of 72 images for each index.
2.3. Principal Component Analysis (PCA)
The characterization of the spatial and temporal patterns of hydrogeomorphological attributes was performed through the decomposition of the spectral index time series (2019–2024) using Principal Component Analysis (PCA), normalized and non-centered, with orientations in the T-mode (time) and S-mode (space).
In T-mode, the variables are temporal samples, allowing the identification of spatial patterns that recur over time. In S-mode, the variables are spatial samples, enabling the identification of temporal patterns that recur across space [27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51].
PCA was performed using the TerrSet Libera GIS software [54], Earth Trends Modeler (ETM) Module, generating images of the principal components, variance, eigenvalues, and loadings for each component in T-mode; in S-mode, the variance, loading images, and score profiles were obtained. Four principal components were extracted: C1, C2, C3, and C4.
In order to understand and highlight the variability of hydrogeomorphological attributes (vegetation, water, and soil), we chose to analyze only Principal Components 2, 3, and 4 of the PCA, since Principal Component 1 (C1) represents the predominant pattern. The temporal and spatial variations of interest in this study are better represented by the secondary components, which capture contrasts and heterogeneities not explained by C1.
To assess the dependency relationship between hydrogeomorphological attributes, Simple Linear Regression was applied to the loading values of Component 2 (C2) in T-mode and to the scores in S-mode for each spectral index. The model construction involved correlation analysis using the Pearson Correlation Coefficient. Data normality was assessed using the Kolmogorov-Smirnov test, with p-values > 0.05 indicating the null hypothesis, meaning the data follow a normal distribution.
Acceptance limits for the linear regression results were defined with a 95% confidence interval. The estimated coefficients were obtained using the least squares method, aiming to minimize the model residuals. The F-test was applied to assess statistical significance, with p > 0.05 established as the relevance criterion. All statistical analyses were performed using PASW Statistics 18 software.
Figure 2.
Methodological framework of the study.
3. Results
3.1. Temporal and Spatial Variability
The PCA results in T-mode highlight spatial patterns of hydrogeomorphological attributes of Coastal Wetlands recurring over time. The percentage of variance and eigenvalues (Figure 3) show that the first component (C1) is dominant across all indices, contributing 91.79%, 89.25%, and 62.56% of the variation in NDVI, MNDWI, and BI2, respectively, with high loading values (Figure 4). NDVI and MNDWI exhibit a variance structure concentrated in C1, while BI2 shows a broader distribution with C2, and the loading profile is distributed between C1 and C2 (Figure 4), indicating a temporal dynamics with greater variability.
The loading profiles of NDVI (Figures 4a, 4b, and 4c), T-Mode, show variations in the time series (2019–2024) and highlight distinct behaviors of vegetation cover, hydrological regime, and soil characteristics. C2 stands out with positive peaks in winter (June to August) and negative peaks in summer (December to March). C3 and C4 exhibit smaller amplitudes, with loading values ranging between -0,2 and 0,2.
The loading profile of MNDWI (Figure 4b) highlights hydrological variations. Although C2 represents less than 10% of the variance, it registers significant oscillations between -0,65 (February 2023) and 0,47 (June 2020). Positive peaks predominate in winter and spring (June to September), while negative peaks occur in summer and autumn. C3 and C4 exhibit more discrete variations, indicating intermediate hydrological patterns in areas with lower flooding frequency, with a water balance regulated by processes such as infiltration, evapotranspiration, and subsurface storage.
The loading profile of BI2 (Figure 4c) highlights patterns in the interaction between soil reflectance, moisture, land cover, and the hydrogeomorphological dynamics of the Coastal wetland. In T-mode, C1 records negative values for most of the time series. C2 presents positive loading values ranging from 0,81 (January 2019) to 0,03 (April 2023). Associated with higher positive loads, C2 is linked to areas of exposed soil and lower surface moisture, suggesting the capture of phenological change events in vegetation cover or drought periods. C3 and C4, with subtler fluctuations, represent intermediate patterns of moisture and soil exposure, associated with compartments with lower seasonal fluctuations. BI2 proves sensitive to soil moisture dynamics and substrate exposure, differentiating areas of exposed soil from areas with greater water variation.
The color composite images of components C2, C3, and C4 (Figure 5) show recurring spatial patterns over time. In NDVI, C2 highlights areas of greater variability in vegetation, with distinct patterns in the wetlands of the Lacustrine Fringe and the Lagoon Fringe, indicating the differentiated response of vegetation to water influence. C3 and C4 emphasize secondary variations associated with seasonal patterns, with a focus on transition areas between wet fields and lacustrine environments.
In MNDWI, C2 shows greater variability in areas of lagoons and marshes, which are wetlands more sensitive to hydrological variations. In BI2, C2 emphasizes regions with higher soil reflectance, while C3 and C4 display hybrid patterns that highlight the complex spectral response of areas susceptible to temporary flooding and seasonal soil exposure.
The results of the PCA in S-mode highlight temporal patterns of the hydrogeomorphological attributes of the Coastal wetland recurring in space. The percentage of variance and eigenvalues show that C1 represents the dominant spatial pattern in all attributes, contributing 84,70%, 84,98%, and 73.41% of the variation in NDVI, MNDWI, and BI2, respectively, with proportionally high scores (Figure 6).
In NDVI and MNDWI, C2, C3, and C4 capture smaller variations. C2 stands out as the second largest variability, especially in BI2 (16,01%), contrasting with vegetation (NDVI 8,34%) and water (MNDWI 7,82%). The high score value of C2 in BI2 (302.584.66) reinforces its relevance in discriminating secondary temporal patterns recurring in space. Components C3 and C4 represent less than 2% variance in all attributes.
The scores (Figure 7) in S-Mode indicate that C1 exhibits a predominant and stable trend throughout the temporal series for the three indices, reflecting the dominant spatial structure and being less susceptible to seasonal variations. C3 and C4 show variations of smaller amplitude compared to C1 and C2. The scores of these secondary components suggest short-term variations in vegetation cover and less intense spectral response in vegetation.
C2 of NDVI (Figure 7a) shows significant oscillations, with notable positive peaks in the winter months (June to August), with the highest recorded value being 1190.41 (August 2021), corresponding to areas of higher vegetative vigor. In autumn and summer, recurrent negative scores indicate that C2 captures disturbances in vegetation that are not apparent in C1. C3 and C4 show less pronounced variations, reflecting short-duration processes and local changes, with opposing fluctuations in the historical series.
The C2 of the MNDWI (Figure 7b) displays score oscillations reflecting the influence of hydrological events such as floods and droughts. The highest C2 scores occur during winter in all years, while the negative peaks appear in summer, in autumn (2020, 2021, and 2022), and in spring (2022).
The BI2 score profile stood out for presenting a distinct pattern, characterized by oscillations in C1, seasonal variations in C2, stability in C3, and negative values in C4, suggesting a differentiated soil dynamic. The C1 of BI2 (Figure 7c) shows positive score values in 60% of the time series, differing from the T-Mode, which showed negative values. Negative peaks occur throughout the years, mainly concentrated in certain months of summer, autumn, and spring. C2 displays less intense oscillations, with a maximum peak of 1063.18 (June 2020) and the lowest recorded value of -522.32 (January 2023). C3 shows loading values close to zero and predominantly positive; while C4 is mostly negative.
The recurrent temporal patterns of hydrogeomorphological attributes of the coastal wetlands, in the false-color composites C2, C3, and C4 in S-Mode (Figure 8), reveal differences between hydrogeomorphological compartments and wetland typologies. In the NDVI, C2 highlights areas of greater contrast corresponding to wet fields and marshes in the Lagoon Terrace and Lagoon Fringe, as well as palustrine vegetation in the Slope compartment—typologies with higher scores throughout the time series.
The false-color composite of the MNDWI highlights flooded areas and water bodies within the Estuarine Lagoon and Lacustrine compartments. C2 emphasizes the contrast between permanently flooded areas and those that are temporarily saturated. C3 and C4 reveal the influence of seasonal processes, such as water expansion and retraction, as well as the interference of suspended material, particularly in Lagoa do Peixe.
BI2, an indicator of soil moisture and exposed soil presence, shows distinct sensitivity to hydrological and geomorphological variations, with contrasts observed in the Lagoon Fringe, Depression, and parts of the Lagoon Terrace and their corresponding wetlands. The C2 of BI2 emphasizes transitions between wetlands and dry areas, capturing fluctuations in water availability. C3 reflects patterns associated with depositional processes related to flood events and water level retraction.
3.2. Relationship Between Hydrogeomorphological Attributes
The Pearson correlation coefficients of the loading values of C2, T-Mode, and the scores, S-Mode, demonstrate statistically significant associations (p < 0,001) between all analyzed combinations (Table 2 and Table 3).
The correlation between NDVI and MNDWI in T-Mode (p = 0.862) and S-Mode (p = 0.880) was strong and positive, indicating an association between vegetation and water surface variability in the compartments. The correlations between NDVI and BI2 (p = 0,550; 0,655) and between MNDWI and BI2 (p = 0,497; 0,642) were moderate and positive, suggesting that soil brightness (BI2) also shares part of the temporal and spatial variability.
The regression lines displayed positive linear trends between the analyzed variables (Figure 9, Figure 10). Figure 9a shows that 74,2% of the NDVI variability can be explained by MNDWI variations in T-Mode, and 77,4% in S-Mode (Figure 10a). In Figure 9b, in the regression between BI2 and MNDWI, only 24,7% of the BI2 variability is explained by MNDWI changes in T-Mode, and 41,2% in S-Mode (Figure 10b). Between NDVI and BI2 (Figure 9c), 30,3% of the temporal variability of NDVI can be explained by the corresponding variations of BI2 in T-Mode, and 42,9% in S-Mode (Figure 10c).
3.3. Hydrogeomorphological Attributes and Water Balance
In the time series (2019-2024), precipitation and evapotranspiration exhibited distinct seasonal patterns (Figures 11a, 11f), influenced by an extended La Niña period (2020-2023), characterized by water deficit. While precipitation showed greater variations associated with the influence of La Niña and El Niño, evapotranspiration remained relatively stable, with moderate fluctuations. An inverse relationship is observed, with stabilization or reduction of evapotranspiration during periods of higher precipitation, and reduction or constancy during periods of lower precipitation. This dynamic reflects the influence of the water balance in regulating water fluxes in the analyzed system.
The positive and negative peaks of the loading weights (T-Mode) and the scores of C2 (S-Mode) for each attribute follow the variations of the hydrological regime (Figures 11a, 11f). The highest values occur during periods of higher precipitation and positive water balance, while negative values are related to phases of water deficit. It is noteworthy that the oscillatory behavior of C2 of BI2 (Figures 11c, 11f) differs from the others, showing a delay in response to the water balance. C2 appears to directly reflect water variations, suggesting a dynamic interaction between vegetation, water availability, and soil structural conditions.
4. Discussion
The characterization of the temporal and spatial variability of hydrogeomorphological attributes (vegetation, water, and soils) in the wetlands of study area allowed for the identification of persistent patterns over time, spatial differentiation, as well as seasonal or episodic fluctuations. Understanding temporal and spatial behavior is essential for comprehending hydrogeomorphological dynamics and guiding conservation strategies aimed at maintaining the environmental health of these systems. The wetlands of the Park occupy interdunal areas between Barriers III and IV [38,40]. This compartment is characterized by low altimetric variation [55] exposed, along with the wetlands, to coastal dynamics, extreme climatic events [37], and anthropogenic impacts.
In this configuration, variability is predominantly determined by biophysical factors. Thus, the method adopted in this analysis was aimed at characterizing and understanding the mechanisms that regulate this variability.
Although PCA has been used in the investigation of inland wetlands [19,20,21,22,25,26,27,51,56], its application in interpreting the variability of coastal wetlands is still underexplored. Moreover, PCA has been predominantly applied in identifying dominant landscape patterns, detecting degradation, and supporting image classification. It uses spectral bands as input variables [22,51], spectral mixture [25], spectral indices such as NDVI and EVI related to vegetation’s biophysical atributes [27,37,57], and MNDWI for water [27].
The analysis of vegetation and its degradation in wetlands using conventional PCA [22], based on spectral bands as variables, showed limitations in distinguishing classes with similar spectral signatures, such as marsh vegetation and agricultural crops. However, other studies using conventional PCA and spectral indices (NDVI, MNDWI) proved suitable for analyzing changes in the Taquari Fan wetland in Brazil [27]; and on the coast of Mexico, the use of a vegetation index combined with PCA [19], was effective in identifying variation patterns in mangrove vegetation.
Our study employed multitemporal decomposition in both T (temporal) and S (spatial) modes, along with the use of spectral indices. In component formation, PCA in T-mode emphasized those components that stood out over time, while S-mode prioritized dominant spatial patterns. These two approaches provided complementary insights into the temporal and spatial variability present in the time series [52]. Radiometric indices of vegetation, water, and soil serve as predictor variables for the association of these attributes and the variability of coastal wetlands [58]. Applying PCA in both T and S modes, along with vegetation radiometric indices in the Pantanal [26], proved effective in investigating its temporal and spatial variability.
4.1. Temporal and Spatial Variability
Coastal wetlands exhibit high temporal and spatial complexity in their biophysical attributes [59]. The interactions among topography, soils, water flow, and vegetation differ across each hydrogeomorphological compartment [40], defining distinct functioning within each wetland typology [60]. Seasonal and interannual variations in coastal wetlands influence the connectivity processes of these environments [61,62]. Although these processes are essential for ecological exchanges within these ecosystems, when considering the increase in emergent plants and water levels, the relatively high soil moisture during extreme precipitation events can cause physiological stress in plants, limiting their photosynthesis and autotrophic respiration [63,64].
The hydrological dynamics, morphology, surface cover, and vegetation interact interdependently, modulating the temporal and spatial variability of hydrogeomorphological atributes [60]. In the study area, hydrological variability is primarily driven by flood pulses associated with extreme El Niño–Southern Oscillation events and the Southern Hemisphere Annular Mode [65].
Climatological variables have been identified as drivers of environmental change in coastal mangroves in Mexico [62], and in the phenology of the Pantanal in Brazil In the Pantanal, a strong relationship was observed between precipitation variability and vegetation patterns [67], with the seasonal hydrological pulse being the main determinant of plant productivity in flood-prone compartments [68]. In addition to precipitation, flood responses vary according to geomorphology and soil type, with sandy soils showing greater susceptibility to drought [26].
In both T-mode and S-mode analyses, vegetation, water, and soil in the Park wetlands showed positive linear associations, with high and moderate correlations. The evidence of temporal variability (T-mode) across space is linked to hydrological events such as flooding (El Niño) and droughts (La Niña), which directly affect the vigor and distribution of vegetation cover.
In S-mode, differences were identified among hydrogeomorphological compartments and wetland typologies, particularly highlighting the contrasts among sectors of the Lagoon Terrace, the Lagoon Fringe, and areas of marsh vegetation (in the Slope compartment).
4.2. Hydrogeomorphological Interactions
The strong linear association observed between NDVI and MNDWI reinforces the functional interdependence between vegetation and the presence of surface water in the Lagoa do Peixe National Park wetlands. This pattern highlights that variations in moisture levels directly affect the vigor and distribution of vegetation cover, both temporally and spatially.
On the other hand, the moderate correlations involving BI2 suggest that, although soil brightness is related to hydroecological processes, its behavior is influenced by other environmental factors beyond water availability or vegetation dynamics. The lower predictive capacity of the regressions involving BI2 indicates a more complex and heterogeneous soil response to hydrological processes.
In the interdune area of study area, corresponding to Lagoon Terraces 1 and 2, hydromorphic sandy-clayey soils predominate [40]. Under natural conditions, these hydromorphic soils remain saturated with water either continuously or during certain periods of the year. They have limited drainage and characteristics resulting from the influence of excessive moisture, whether permanent or temporary, caused by the proximity of the water table to the surface during specific seasons [69].
The relationship between the water balance, loading values, and the scores of hydrogeomorphological attributes revealed well-defined seasonal patterns between precipitation and evapotranspiration, modulated by climatic events associated with the prolonged La Niña period (2020–2022). In contrast, the year 2019, under El Niño influence, was marked by regular rainfall, which contributed to the stability of evapotranspiration observed during the following dry period.
Evapotranspiration in wetlands can remain stable even during water deficit periods due to a combination of ecohydrological factors, including physiological adaptations of vegetation, groundwater storage, or soil moisture [70,71]. Coastal dynamics elements, such as winds and tides, also modulate evapotranspiration behavior, as observed in the coastal wetlands of Chongming Island, China [72], where coastal wind and tidal dynamics introduced complex patterns into evapotranspiration behavior.
The oscillation of loading values (T-mode) and scores (S-mode) found in our study is consistent with research that highlights the direct influence of water availability on wetland dynamics [26,28,60,64,65,66,67,73], and the modulation of hydrogeomorphological processes [19,74,75,76,77,78].
In our study area, this oscillation is clearly expressed in the hydrogeomorphological compartments of Lagoon Terraces 1 and 2, where the interaction between seasonal hydrological dynamics and flat geomorphology intensifies the response of hydrogeomorphological attributes. Characterized by hydromorphic soils and low slope, the Lagoon Terraces show high sensitivity to fluctuations in the water table and to flood pulses, which explains the high loading values of Component 2 (C2) in T-mode, associated with extreme events, as well as the contrasting S-mode scores that distinguish permanently flooded areas from temporarily flooded ones.
This spatial compartmentalization highlights the role of geological characteristics in modulating wetland resilience, with the Lagoon Terraces acting as critical transition zones between lacustrine and palustrine environments. Small hydrological variations in these systems result in significant impacts on vegetation and soil exposure, as indicated by BI2 loadings and scores.
The distinct temporal and spatial response of BI2 suggests the existence of resilience mechanisms or delayed vegetation responses, possibly associated with factors such as rooting depth, soil water storage capacity, or physiological adaptations [70,71,76].
Additionally, the influence of soil structural conditions may explain variations in temporal response, as soil texture and porosity affect water retention and redistribution, thus modifying vegetation responses to precipitation events [26,79,80].
The spectral behavior of sandy, fine-textured soils, as found in the study area, is another factor explaining the temporal and spatial variability of this attribute, as detected by BI2. Sandy soils with finer texture tend to retain more moisture during precipitation events, increasing radiant energy absorption and reducing surface reflectance. This pattern is particularly evident in sandy soils with sparse vegetation cover, as observed in Lagoon Terraces 1 and 2 of the Lagoa do Peixe National Park.
Vegetation variability was influenced by wetland geological characteristics in a study conducted in the Pantanal [26], which identified early senescence of vegetation in areas dominated by sandy soils. According to the authors, this pattern emphasizes the direct influence of soil composition on vegetation dynamics, as sandy soils have lower water and nutrient retention capacity [26]. These characteristics may intensify the impacts of extreme climatic events, such as drought periods, directly affecting the ecological resilience of the ecosystem [79].
5. Conclusions
This study enabled the characterization of the temporal and spatial variability of hydrogeomorphological attributes (vegetation, water, and soil) in the coastal wetlands of Lagoa do Peixe National Park, contributing to the understanding of the dynamics and variability of this ecosystem, a Ramsar site of global relevance. The methodological approach, based on multitemporal analysis of spectral indices (NDVI, MNDWI, and BI2) and Principal Component Analysis (PCA) in both T-mode (temporal) and S-mode (spatial), proved effective in identifying recurring patterns and seasonal fluctuations, highlighting contrasts among hydrogeomorphological compartments.
The results showed that vegetation (NDVI) and water (MNDWI) exhibit a strong positive correlation, reinforcing the functional interdependence between these attributes. Vegetation cover dynamics responded directly to hydrological variations, with peaks in vegetative vigor associated with periods of higher precipitation and favorable water balance. In contrast, the response of soil (BI2) revealed greater complexity, indicating an influence not only of moisture but also of factors such as texture, surface exposure, and depositional processes. This heterogeneity underscores the importance of integrated analyses to understand the resilience of these ecosystems to climatic extremes.
The influence of El Niño and La Niña events stood out in modulating hydrogeomorphological variability, with water deficit periods (La Niña) accentuating temporal and spatial contrasts. Nevertheless, evapotranspiration remained relatively stable, suggesting adaptive vegetation mechanisms and soil water retention. This behavior reinforces the self-regulating capacity of wetlands, although more intense climate changes may challenge their long-term resilience.
From a spatial perspective, the Lagoon Fringe, Lagoon Terrace, and Hillslope areas showed the greatest variations, reflecting geomorphological differences and hydrological connectivity. These sectors are particularly sensitive to changes in precipitation regimes and extreme coastal events, requiring targeted conservation strategies.
Implications for Conservation and Future Research
The findings of this study highlight the need for:
- Continuous monitoring of coastal wetlands, integrating remote sensing and hydrological data to anticipate responses to climate change;
- Differentiated protection by compartment, considering the higher sensitivity of areas such as the Lagoon Fringe to hydrological variations;
- Investigation of subsurface processes, such as water storage and exchanges with the water table, to better understand soil and vegetation resilience.
In summary, the identified variability reflects the complexity and interdependence of hydrogeomorphological attributes in the Lagoa do Peixe National Park, reinforcing its role as a dynamic and sensitive ecosystem. The approach employed can be adapted to other Ramsar sites, contributing to the sustainable management of coastal wetlands facing increasing climatic and anthropogenic pressures.
Author Contributions
C.C.K.: conception, research, methodology, mapping, validation, data preparation, investigation, analysis, and writing of the article; L. A. G.: supervision, research, validation, review, analysis, and writing of the article; H.H.: review, supervision in the TerrSet Libera GIS software; T. F.B.: review, analysis, and writing of the article; C. S. C.: review.
Funding
Research funded by the Coordination for the Improvement of Higher Education Personnel (CAPES) through doctoral scholarships, process numbers 88887.645451/2021-00, 88887.488339/2020-00, and 88887.801261/2023-00; CNPq PQ scholarship - process number 301822/2022-0; Foundation for Research Support of the State of Rio Grande do Sul (FAPERGS) - Gaúcho Researcher Program, FAPERGS Public Notice 07/2021.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article. More detailed data can be provided upon request to the corresponding author.
Acknowledgments
To the Postgraduate Program in Remote Sensing of the State Center for Research in Remote Sensing and Meteorology/UFRGS; to CAPES and FAPERGS for their financial support. To ICMBio for the authorization for activities for scientific purposes.
Conflicts of Interest
The authors declare not having any conflict of interest.
References
- Mitsch, W.J.; Bernal, B.; Nahlik, A.M.; Mander, Ü.; Zhang, L.; Anderson, C.J.; Jørgensen, S.E.; Brix, H. Wetlands, Carbon, and Climate Change. Landscape Ecol 2013, 28, 583–597. [Google Scholar] [CrossRef]
- Simioni, J.P.D. Métodos de Classificação de Imagens de Satélite Para Delineamento de Banhados. Tese de Doutorado, Programa de Pós-Graduação em Sensoriamento Remoto, Universidade Federal do Rio Grande do Sul: Porto Alegre, 2021.
- Aitali, R.; Snoussi, M.; Kolker, A.S.; Oujidi, B.; Mhammdi, N. Effects of Land Use/Land Cover Changes on Carbon Storage in North African Coastal Wetlands. Journal of Marine Science and Engineering 2022, 10, 364. [Google Scholar] [CrossRef]
- Erwin, K.L. Wetlands and Global Climate Change: The Role of Wetland Restoration in a Changing World. Wetlands Ecol Manage 2009, 17, 71–84. [Google Scholar] [CrossRef]
- Ilyas, S.; Xu, X.; Jia, G.; Zhang, A. Interannual Variability of Global Wetlands in Response to El Niño Southern Oscillations (ENSO) and Land-Use. Front. Earth Sci. 2019, 7. [Google Scholar] [CrossRef]
- Zhang, Z.; Jiang, W.; Peng, K.; Wu, Z.; Ling, Z.; Li, Z. Assessment of the Impact of Wetland Changes on Carbon Storage in Coastal Urban Agglomerations from 1990 to 2035 in Support of SDG15.1. Science of The Total Environment 2023, 877, 162824. [Google Scholar] [CrossRef]
- Saunois, M.; Jackson, R.B.; Bousquet, P.; Poulter, B.; Canadell, J.G. The Growing Role of Methane in Anthropogenic Climate Change. Environ. Res. Lett. 2016, 11, 120207. [Google Scholar] [CrossRef]
- IPBE The IPBES Regional Assessment Report on Biodiversity and Ecosystem Services for the Americas; Zenodo. 2018.
- Kuenzer, C.; Bluemel, A.; Gebhardt, S.; Quoc, T.V.; Dech, S. Remote Sensing of Mangrove Ecosystems: A Review. Remote Sensing 2011, 3, 878–928. [Google Scholar] [CrossRef]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A Blueprint for Blue Carbon: Toward an Improved Understanding of the Role of Vegetated Coastal Habitats in Sequestering CO2. Frontiers in Ecology and the Environment 2011, 9, 552–560. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Megonigal, J.P. Tidal Wetland Stability in the Face of Human Impacts and Sea-Level Rise. Nature 2013, 504, 53–60. [Google Scholar] [CrossRef]
- Fluet-Chouinard, E.; Stocker, B.D.; Zhang, Z.; Malhotra, A.; Melton, J.R.; Poulter, B.; Kaplan, J.O.; Goldewijk, K.K.; Siebert, S.; Minayeva, T.; et al. Extensive Global Wetland Loss over the Past Three Centuries. Nature 2023, 614, 281–286. [Google Scholar] [CrossRef]
- Xi, Y.; Peng, S.; Ciais, P.; Chen, Y. Nature Climate Change. 2021, pp. 45–51p.
- Hu, S.; Niu, Z.; Chen, Y. Global Wetland Datasets: A Review. Wetlands 2017, 37, 807–817. [Google Scholar] [CrossRef]
- Yu, H.; Li, S.; Liang, Z.; Xu, S.; Yang, X.; Li, X. Multi-Source Remote Sensing Data for Wetland Information Extraction: A Case Study of the Nanweng River National Wetland Reserve. Sensors 2024, 24, 6664. [Google Scholar] [CrossRef] [PubMed]
- Salimi, S.; Almuktar, S.A.A.A.N.; Scholz, M. Impact of Climate Change on Wetland Ecosystems: A Critical Review of Experimental Wetlands. Journal of Environmental Management 2021, 286, 112160. [Google Scholar] [CrossRef]
- Day, J.W.; Christian, R.R.; Boesch, D.M.; Yáñez-Arancibia, A.; Morris, J.; Twilley, R.R.; Naylor, L.; Schaffner, L.; Stevenson, C. Consequences of Climate Change on the Ecogeomorphology of Coastal Wetlands. Estuaries and Coasts 2008, 31, 477–491. [Google Scholar] [CrossRef]
- Venegas-Cordero, N.; Marcinkowski, P.; Stachowicz, M.; Grygoruk, M. On the Role of Water Balance as a Prerequisite for Aquatic and Wetland Ecosystems Management: A Case Study of the Biebrza Catchment, Poland. Ecohydrology & Hydrobiology 2024, 24, 808–819. [Google Scholar] [CrossRef]
- Berlanga-Robles, C.A. Trends in Mangrove Canopy and Cover in the Teacapan-Agua Brava Lagoon System (Marismas Nacionales) in Mexico: An Approach Using Open-Access Geospatial Data. Wetlands 2024, 45, 1. [Google Scholar] [CrossRef]
- Munyati, C. Use of Principal Component Analysis (PCA) of Remote Sensing Images in Wetland Change Detection on the Kafue Flats, Zambia. Geocarto International 2004, 19, 11–22. [Google Scholar] [CrossRef]
- Torbick, N.; Becker, B. Evaluating Principal Components Analysis for Identifying Optimal Bands Using Wetland Hyperspectral Measurements From the Great Lakes, USA. Remote Sensing 2009, 1, 408–417. [Google Scholar] [CrossRef]
- Karabulut, M.; Küçükönder, M. An Examination of Temporal Changes in Göksu Delta (Turkey) Using Principle Component Analysis. SSRN Journal 2018. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, G.; Xue, B.; Zhang, M.; Tan, Z. Dynamic Landscapes and the Driving Forces in the Yellow River Delta Wetland Region in the Past Four Decades. Science of The Total Environment 2021, 787, 147644. [Google Scholar] [CrossRef]
- Yelwa, S.A.; Sanda, A.B.; Usman, U. Spatio-Temporal Assessment of Vegetation Resource Dynamics in Nigeria from SPOT Satellite Imageries. American Journal of Climate Change 2019, 8, 502–519. [Google Scholar] [CrossRef]
- Mattson, M.; Sousa, D.; Quandt, A.; Ganster, P.; Biggs, T. Mapping Multi-Decadal Wetland Loss: Comparative Analysis of Linear and Nonlinear Spatiotemporal Characterization. Remote Sensing of Environment 2024, 302, 113969. [Google Scholar] [CrossRef]
- de Almeida, T.I.R.; Penatti, N.C.; Ferreira, L.G.; Arantes, A.E.; do Amaral, C.H. Principal Component Analysis Applied to a Time Series of MODIS Images: The Spatio-Temporal Variability of the Pantanal Wetland, Brazil. Wetlands Ecol Manage 2015, 23, 737–748. [Google Scholar] [CrossRef]
- Pereira, L.E.; Noveli, R.A.P.; Filho, A.C.P. USO DE ANÁLISE DE COMPONENTES PRINCIPAIS (ACP) PARA CARACTERIZAÇÃO DAS SUB-REGIÕES DO MEGALEQUE DO TAQUARI – PANTANAL. Revista GeoPantanal 2023, 18, 114–126. [Google Scholar] [CrossRef]
- Dronova, I.; Gong, P.; Wang, L.; Zhong, L. Mapping Dynamic Cover Types in a Large Seasonally Flooded Wetland Using Extended Principal Component Analysis and Object-Based Classification. Remote Sensing of Environment 2015, 158, 193–206. [Google Scholar] [CrossRef]
- Junk, W.J. Definição, Delineamento e Classificação Brasileira Das Áreas Úmidas. In Inventário das áreas úmidas brasileiras: Distribuição, ecologia, manejo, ameaças e lacunas de conhecimento; Carlini & Caniato Editorial: Cuiabá - MT, 2024; 42p, ISBN 978-85-8009-353-7. [Google Scholar]
- Ramos, R.A.; Pasqualeto, A.I.; Balbueno, R.A.; Das Neves, D.D.; De Quadros, E.L.L. Mapeamento e Diagnóstico de Áreas Úmidas No Rio Grande Do Sul, Com o Uso de Ferramentas de Geoprocessamento. In Proceedings of the Anais; 2014; pp. 17–21. [Google Scholar]
- Belloli, T.F.; Guasselli, L.A.; Cunha, C.S.; Korb, C.C. Mudanças e Transição da Cobertura e Usoda Terra nas Áreas Úmidas da Região Geomorfológica Planície Costeira - RS. Geo UERJ 2024. [Google Scholar] [CrossRef]
- Villwock, J.A.; Tomazelli, L.J.; Loss, E.L.; Dehnhardt, E.A.; Bachi, F.A.; Dehnhardt, B.A. Quaternary of South America and Antarctic Peninsula. Geology of the Rio Grande Do Sul Coastal Province 1986, 4, 79–97. [Google Scholar]
- Tomazelli, L.J.; Villwock, J.A. O Cenozóico Do Rio Grande Do Sul: Geologia Da Planície Costeira. In Geologia do Rio Grande do Sul; Holz, F. & DeRos, L.F., 2000; p. 444.
- Schäffer, A.; Lanzer, R.; Scur, L. 2013.
- Moraes, V.L. Uso Do Solo e Conservação Ambiental No Parque Nacional daLagoa Do Peixe e Entorno (RS). Dissertação de Mestrado, Programa de Pós-Graduação em Geografia, Universidade Federal do Rio Grande do Sul: Porto Alegre, 2009.
- Portz, L.C.; Guasselli, L.A.; Corrêa, I.C.S. Variação Espacial e Temporal de NDVI Na Lagoa Do Peixe, RS. Revista Brasileira de Geografia Física 2011, 5, 897–908. [Google Scholar]
- Schossler, V. Influência Das Mudanças Climáticas Em Geoindicadores Na Costa Sul Do Brasil. Tese de Doutorado, Programa de Pós-Graduação em Geociências, Universidade Federal do Rio Grande do Sul: Porto Alegre, 2016.
- Arejano, T.B. Geologia e Evolução Holocênica Do Sistema Lagunar Da Lagoa Do Peixe, Litoral Médio Do Rio Grande Do Sul, Brasil. Tese de Doutorado, Universidade Federal do Rio Grande do Sul: Programa de Pós-Graduação em Geociências, 2006.
- Streck, E.V.; Kämpf, N.; Dalmolin, R.S.D.; Klamt, E.; Nascimento, P.C.; Scheneider, P.; Giasson, E.; Pinto,, L.F.S. (Eds.) Solos Do Rio Grande Do Sul, 2nd ed. Emater-Ascar: Porto Alegre, 2008.
- Korb, C.C.; Guasselli, L.A.; Belloli, T.F.; Cunha, C.S.; Bauer, A.L.; Brückmann, C. dos S. Hydrogeomorphological compartmentalization in coastal wetlands - Lagoa do Peixe National Park, Brazil. Revista Brasileira de Geomorfologia 2024, 25. [Google Scholar] [CrossRef]
- Reboita, M.S.; Ambrizzi, T.; Rocha, R.P. da Relationship between the Southern Annular Mode and Southern Hemisphere Atmospheric Systems Relação Entre o Modo Anular Sul e Os Sistemas Atmosféricos No Hemisfério Sul. Revista Brasileira de Meteorologia 2009, 24, 48–55. [Google Scholar] [CrossRef]
- Fontana, D.C.; Berlato, M.A. Influência do El Niño Oscilação Sul sobre a precipitação do Estado do Rio Grande do Sul. Revista Brasileira de Agrometeorologia 1997, 5, 127–132. [Google Scholar]
- Britto, F.P.; Barletta, R.; Mendonça, M. Variabilidade Espacial e Temporal da Precipitação Pluvial no Rio Grande Do Sul: influência do fenômeno El Niño Oscilação Sul. Revista Brasileira de Climatologia 2008, 3, 37–48. [Google Scholar] [CrossRef]
- Rodrigues, B.D. Comportamento Dos Sistemas Frontais No Estado Do Rio Grande Do Sul Durante Os Episódios ENOS. Dissertação de Mestrado, Programa de Pós-Graduação em Sensoriamento Remoto, Universidade Federal do Rio Grande do Sul: Porto Alegre, 2015.
- Sbruzzi, J.B. Análise Da Dinâmica Da Embocadura Da Lagoa Do Peixe - RS Utilizando Dados de Sensoriamento Remoto Orbital. Dissertação de Mestrado, Programa de Pós-Graduação em Sensoriamento Remoto, Universidade Federal do Rio Grande do Sul: Porto Alegre, 2015.
- Signori, L.M. Mapeamento Por Sensoriamento Remoto de Área de Pinus Spp No Parque Nacional Da Lagoa Do Peixe. Dissertação de Mestrado, Universidade Federal do Rio Grande do Sul: Programa de Pós-Graduação em Sensoriamento Remoto, 2018.
- Portz, L.; Manzolli, R.P.; Alcántara-Carrió, J.; Rockett, G.C.; Barboza, E.G. Degradation of a Transgressive Coastal Dunefield by Pines Plantation and Strategies for Recuperation (Lagoa Do Peixe National Park, Southern Brazil). Estuarine, Coastal and Shelf Science 2021, 259, 107483. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS (Earth Resources Technology Satellite).; Greenbelt, 1973; pp. 309–317.
- Xu, H. Modification of Normalised Difference Water Index (NDWI) to Enhance Open Water Features in Remotely Sensed Imagery. International Journal of Remote Sensing 2006, 27, 3025–3033. [Google Scholar] [CrossRef]
- Escadafal, R. Remote Sensing of Arid Soil Surface Color with Landsat Thematic Mapper. Advances in Space Research 1989, 9, 159–163. [Google Scholar] [CrossRef]
- Nepita-Villanueva, M.R.; Berlanga-Robles, C.A.; Ruiz-Luna, A.; Morales Barcenas, J.H. Spatio-Temporal Mangrove Canopy Variation (2001–2016) Assessed Using the MODIS Enhanced Vegetation Index (EVI). J Coast Conserv 2019, 23, 589–597. [Google Scholar] [CrossRef]
- Machado-Machado, E.A.; Neeti, N.; Eastman, J.R.; Chen, H. Implications of Space-Time Orientation for Principal Components Analysis of Earth Observation Image Time Series. Earth Sci Inform 2011, 4, 117–124. [Google Scholar] [CrossRef]
- Neeti, N.; Ronald Eastman, J. Novel Approaches in Extended Principal Component Analysis to Compare Spatio-Temporal Patterns among Multiple Image Time Series. Remote Sensing of Environment 2014, 148, 84–96. [Google Scholar] [CrossRef]
- Eastman, J.R. TerrSet Libera GIS. Eospatial Monitoring and Modeling System. Tutorial. 2024. [Google Scholar]
- Tagliani, C.R.; Hartmann, C.; Calliari, L. J.; Griep, G.H. Geologia e Geomorfologia Da Porção Sul Do Parque Nacional Da Lagoa Do Peixe, RS, Brasil. In Proceedings of the Anais; São Paulo; 1992; pp. 292–294. [Google Scholar]
- Wang, Y.; Jin, S.; Dardanelli, G. Vegetation Classification and Evaluation of Yancheng Coastal Wetlands Based on Random Forest Algorithm from Sentinel-2 Images. Remote Sensing 2024, 16, 1124. [Google Scholar] [CrossRef]
- Schossler, V. Morfodinâmica Da Embocadura Da Lagoa Do Peixe e Da Linha de Praia Adjacente. Dissertação de Mestrado, Programa de Pós-Graduação em Geociências, Universidade Federal do Rio Grande do Sul: Porto Alegre, 2011.
- Ramsar Convention Secretariat Wetland Inventory: A Ramsar Framework for Wetland Inventory and Ecological Character Description.; 4th ed.; Switzerland, 2010; Vol. 15;
- Doughty, C.L.; Ambrose, R.F.; Okin, G.S.; Cavanaugh, K.C. Characterizing Spatial Variability in Coastal Wetland Biomass across Multiple Scales Using UAV and Satellite Imagery. Remote Sensing in Ecology and Conservation 2021, 7, 411–429. [Google Scholar] [CrossRef]
- Brinson, M.M. A Hydrogeomorphic Classification for Wetlands 1993.
- Simioni, J.P.D.; Guasselli, L.A.; Etchelar, C.B. Connectivity among Wetlands of EPA of Banhado Grande, RS. RBRH 2017, 22. [Google Scholar] [CrossRef]
- Li, Z.; Sun, W.; Chen, H.; Xue, B.; Yu, J.; Tian, Z. Interannual and Seasonal Variations of Hydrological Connectivity in a Large Shallow Wetland of North China Estimated from Landsat 8 Images. Remote Sensing 2021, 13, 1214. [Google Scholar] [CrossRef]
- Islam, M.A.; Macdonald, S.E. Ecophysiological Adaptations of Black Spruce (Picea Mariana) and Tamarack (Larix Laricina) Seedlings to Flooding. Trees 2004, 18, 35–42. [Google Scholar] [CrossRef]
- Wei, S.; Han, G.; Chu, X.; Sun, B.; Song, W.; He, W.; Wang, X.; Li, P.; Yu, D. Impactos Prolongados de Eventos Extremos de Precipitação Enfraqueceram a Força Anual Do Sumidouro de CO2 Do Ecossistema Em Uma Área Úmida Costeira. Agricultural and Forest Meteorology 2021, 310, 108655. [Google Scholar] [CrossRef]
- Korb, C.C.; Guasselli, L.A.; Belloli, T.F.; Cunha, C.S. Dinâmica espaço-temporal de pulsos de inundações nas Áreas Úmidas do Parque Nacional da Lagoa do Peixe, sul do Brasil. Investigaciones Geográficas: Una mirada desde el sur. [CrossRef]
- Rioja-Nieto, R.; Barrera-Falcón, E.; Torres-Irineo, E.; Mendoza-González, G.; Cuervo-Robayo, A.P. Environmental Drivers of Decadal Change of a Mangrove Forest in the North Coast of the Yucatan Peninsula, Mexico. J Coast Conserv 2017, 21, 167–175. [Google Scholar] [CrossRef]
- Penatti, N.C.; Almeida, T.I.R. de; Ferreira, L.G.; Arantes, A.E.; Coe, M.T. Satellite-Based Hydrological Dynamics of the World’s Largest Continuous Wetland. Remote Sensing of Environment 2015, 170, 1–13. [Google Scholar] [CrossRef]
- Ivory, S.J.; McGlue, M.M.; Spera, S.; Silva, A.; Bergier, I. Vegetation, Rainfall, and Pulsing Hydrology in the Pantanal, the World’s Largest Tropical Wetland. Environ. Res. Lett. 2019, 14, 124017. [Google Scholar] [CrossRef]
- Empresa Brasileira de Pesquisa Agropecuária Sistema Brasileiro de Classificação de Solos; 2nd ed. 2006.
- Giardina, F.; Gentine, P.; Konings, A.G.; Seneviratne, S.I.; Stocker, B.D. Diagnosing Evapotranspiration Responses to Water Deficit across Biomes Using Deep Learning. New Phytologist 2023, 240, 968–983. [Google Scholar] [CrossRef]
- Stocker, B.D.; Tumber-Dávila, S.J.; Konings, A.G.; Anderson, M.C.; Hain, C.; Jackson, R.B. Global Patterns of Water Storage in the Rooting Zones of Vegetation. Nat. Geosci. 2023, 16, 250–256. [Google Scholar] [CrossRef]
- Huang, Y.; Guo, H.; Chen, X.; Chen, Z.; van der Tol, C.; Zhou, Y.; Tang, J. Meteorological Controls on Evapotranspiration over a Coastal Salt Marsh Ecosystem under Tidal Influence. Agricultural and Forest Meteorology 2019, 279, 107755. [Google Scholar] [CrossRef]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. Flood Pulse Concept in River- Floodplain-Systems.; 1989; Vol. 106, pp. 110–127.
- Wei, S.; Han, G.; Chu, X.; Sun, B.; Song, W.; He, W.; Wang, X.; Li, P.; Yu, D. Prolonged Impacts of Extreme Precipitation Events Weakened Annual Ecosystem CO2 Sink Strength in a Coastal Wetland. Agricultural and Forest Meteorology 2021, 310, 108655. [Google Scholar] [CrossRef]
- Ablat, X.; Liu, G.; Liu, Q.; Huang, C. Using MODIS-NDVI Time Series to Quantify the Vegetation Responses to River Hydro-Geomorphology in the Wandering River Floodplain in an Arid Region. Water 2021, 13, 2269. [Google Scholar] [CrossRef]
- Daly, E.; Porporato, A. Impact of Hydroclimatic Fluctuations on the Soil Water Balance. Water Resources Research 2006, 42. [Google Scholar] [CrossRef]
- Singh, M.; Sinha, R. Hydrogeomorphic Indicators of Wetland Health Inferred from Multi-Temporal Remote Sensing Data for a New Ramsar Site (Kaabar Tal), India. Ecological Indicators 2021, 127, 107739. [Google Scholar] [CrossRef]
- Lee, O.; Kim, H.S.; Kim, S. Hydrological Simple Water Balance Modeling for Increasing Geographically Isolated Doline Wetland Functions and Its Application to Climate Change. Ecological Engineering 2020, 149, 105812. [Google Scholar] [CrossRef]
- Penatti, N.C.; de Almeida, T.I.R. Subdvision of Pantanal Quaternary Wetlands: MODIS NDVI Time Series In The Indirect Detection Of Sediments Granulometry. The International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences, B8. [CrossRef]
- Narváez-Salcedo, S.; Hoyos, N.; Aldana-Domínguez, J. Spatial and Temporal Trend of EVI in Mangrove Forests of Coastal Lagoons in the Colombian Caribbean. 2022.
Figure 1.
Study area location.
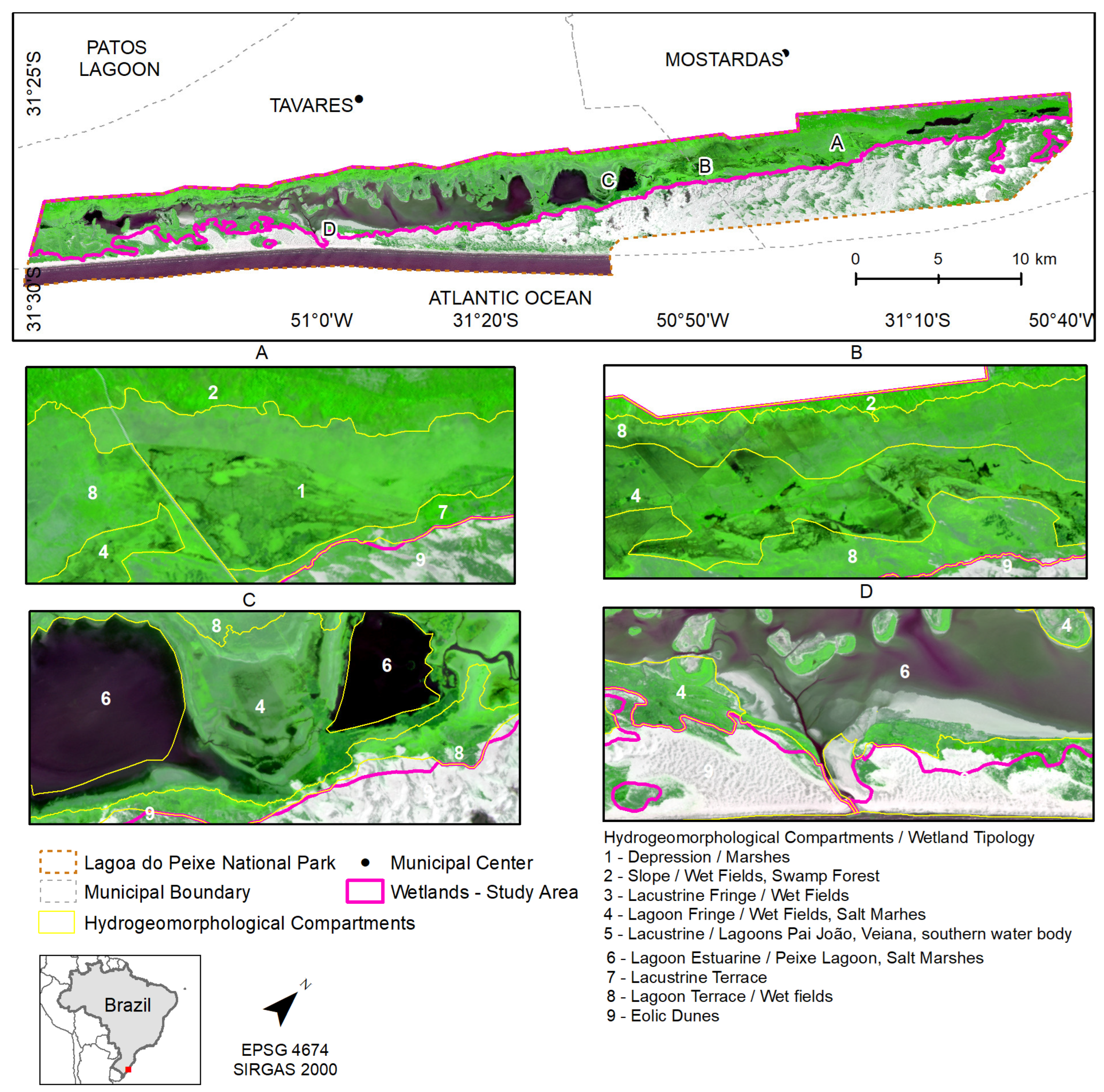
Figure 3.
Percentage of variance and eigenvalues associated with each Principal Component and Hydrogeomorphological Attribute, T-Mode.
Figure 3.
Percentage of variance and eigenvalues associated with each Principal Component and Hydrogeomorphological Attribute, T-Mode.
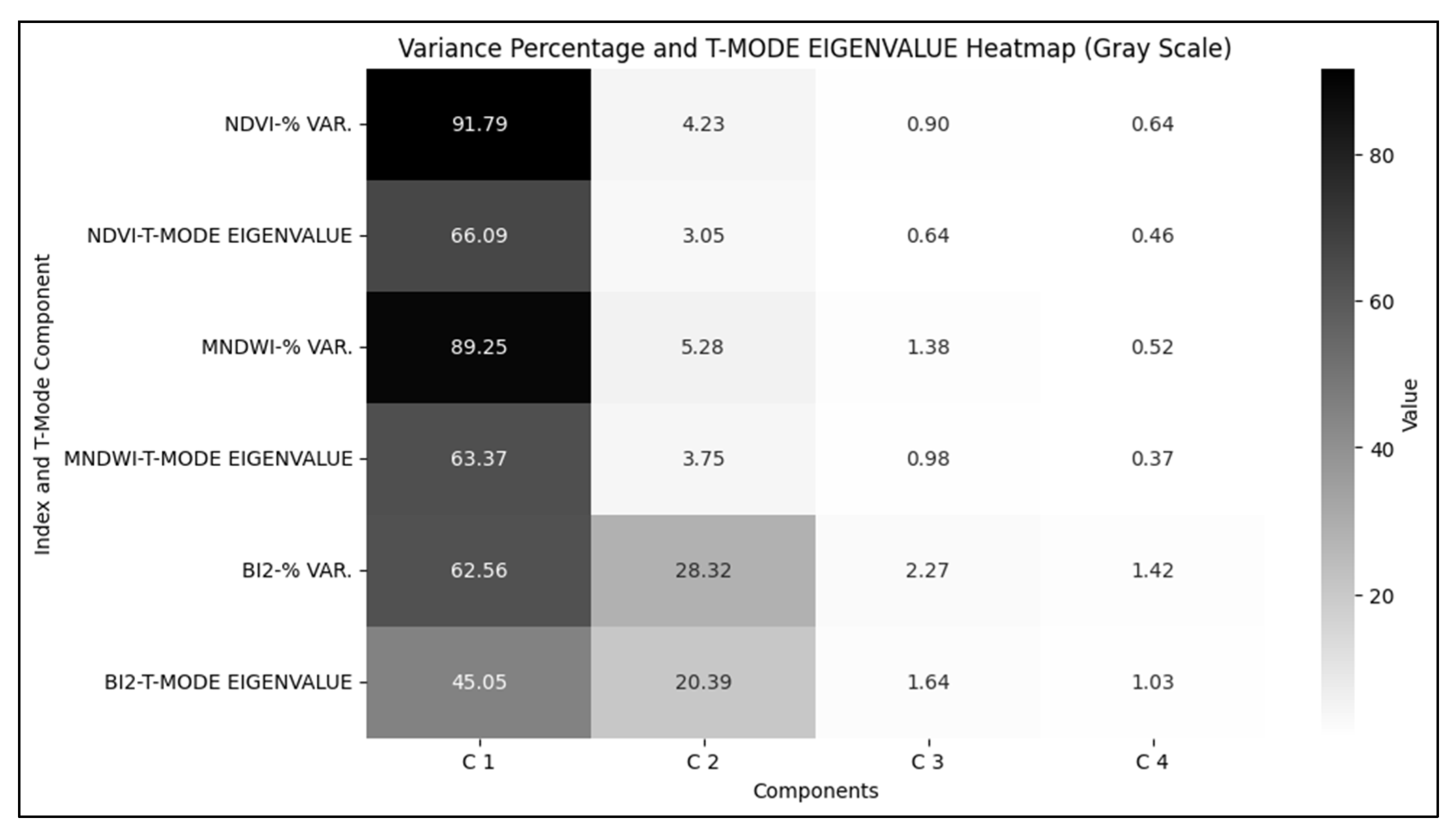
Figure 4.
Loading profiles by hydrogeomorphological attribute, T-Mode. The black, red, green, and blue lines correspond to the temporal variation (C1, C2, C3, C4), respectively; and loading profiles (a) of NDVI; (b) of MNDWI; (c) of BI2.
Figure 4.
Loading profiles by hydrogeomorphological attribute, T-Mode. The black, red, green, and blue lines correspond to the temporal variation (C1, C2, C3, C4), respectively; and loading profiles (a) of NDVI; (b) of MNDWI; (c) of BI2.
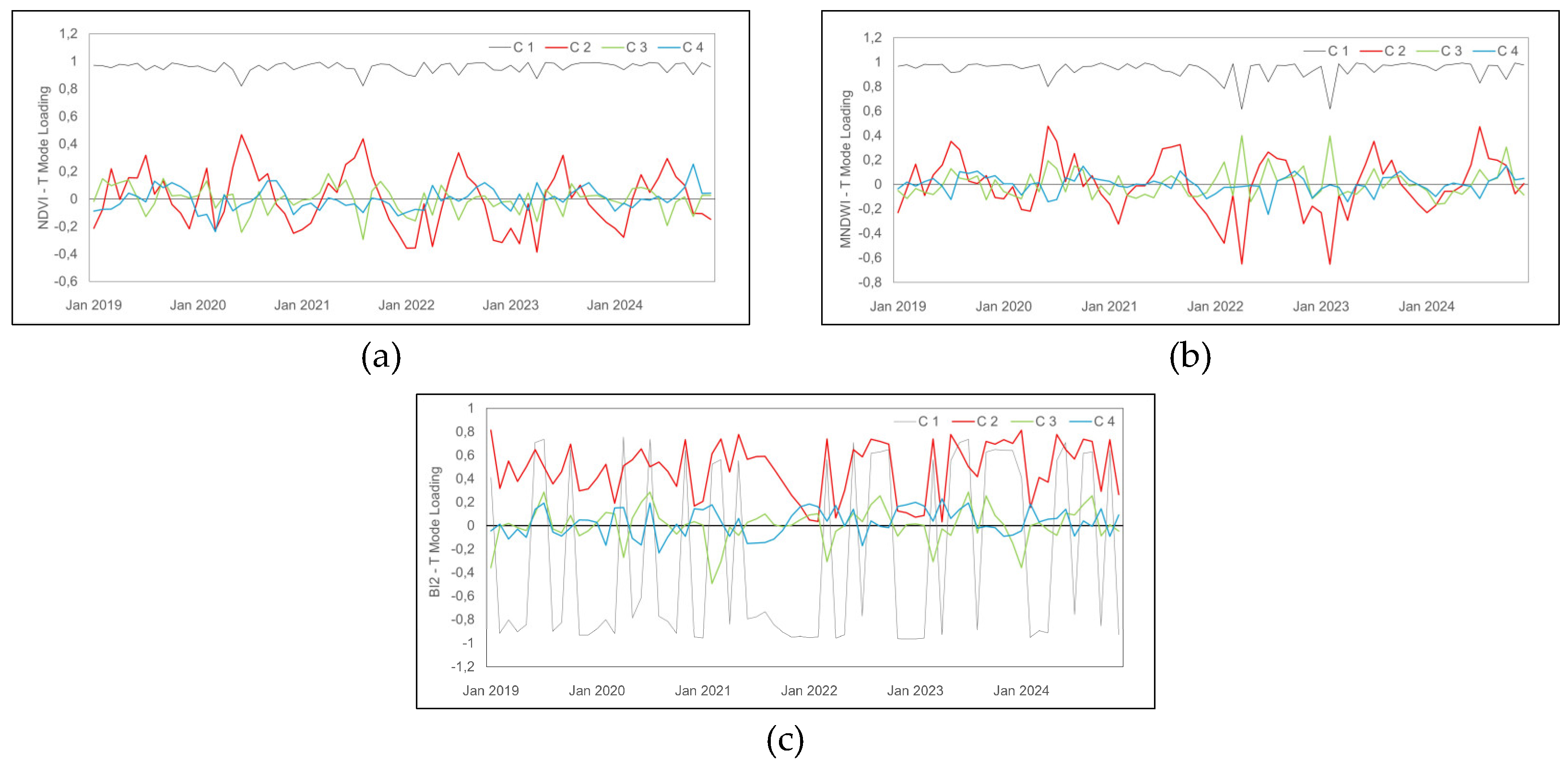
Figure 5.
Color composite of components C2, C3, and C4, T-Mode, in hydrogeomorphological compartments and wetland typologies of study area. NDVI (A, B, C, D); MNDWI (E, F, G, H); BI2 (I, J, L, M).
Figure 5.
Color composite of components C2, C3, and C4, T-Mode, in hydrogeomorphological compartments and wetland typologies of study area. NDVI (A, B, C, D); MNDWI (E, F, G, H); BI2 (I, J, L, M).
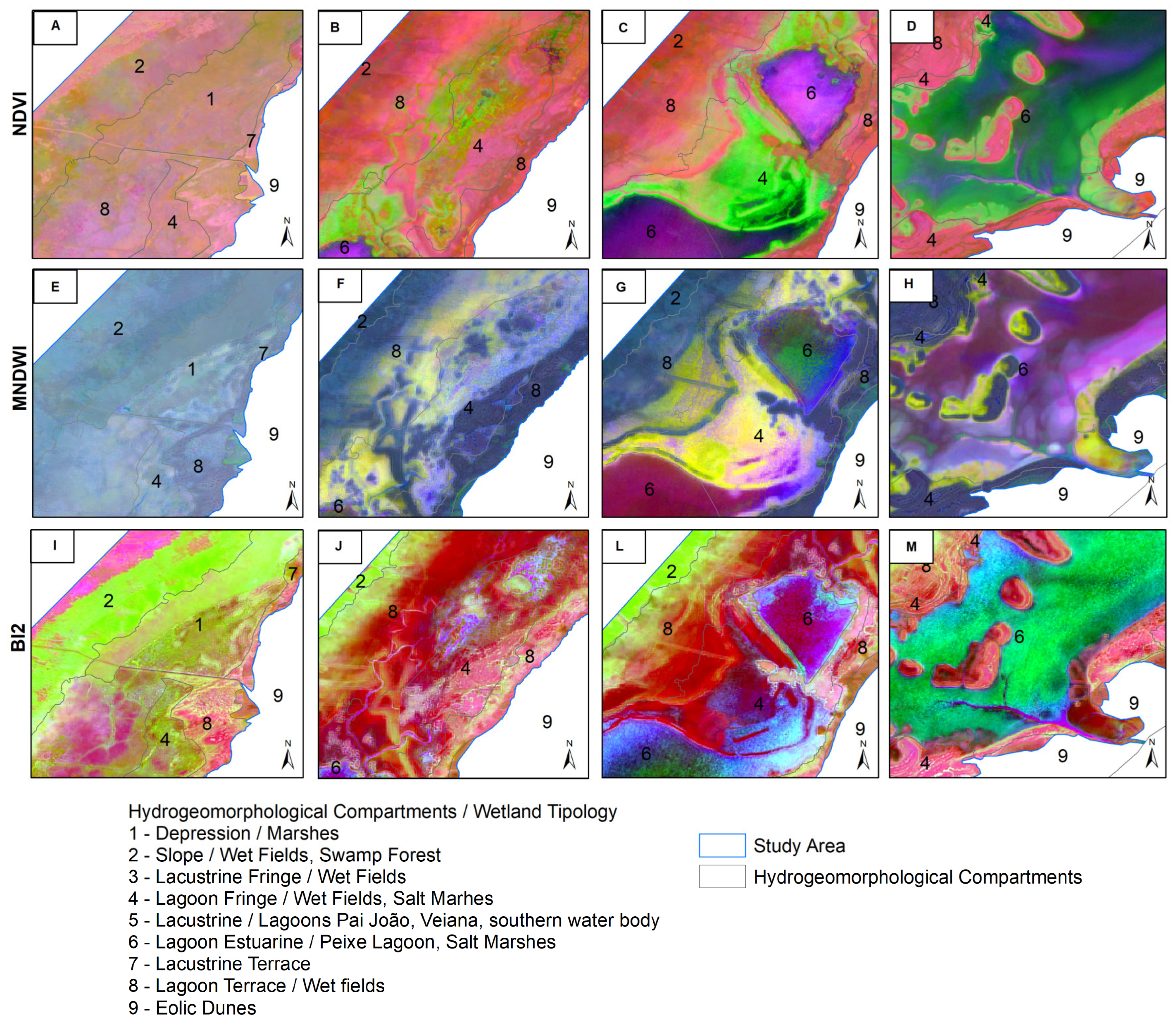
Figure 6.
Percentage of variance and eigenvalues associated with each Principal Component and Hydrogeomorphological Attribute, S-Mode.
Figure 6.
Percentage of variance and eigenvalues associated with each Principal Component and Hydrogeomorphological Attribute, S-Mode.
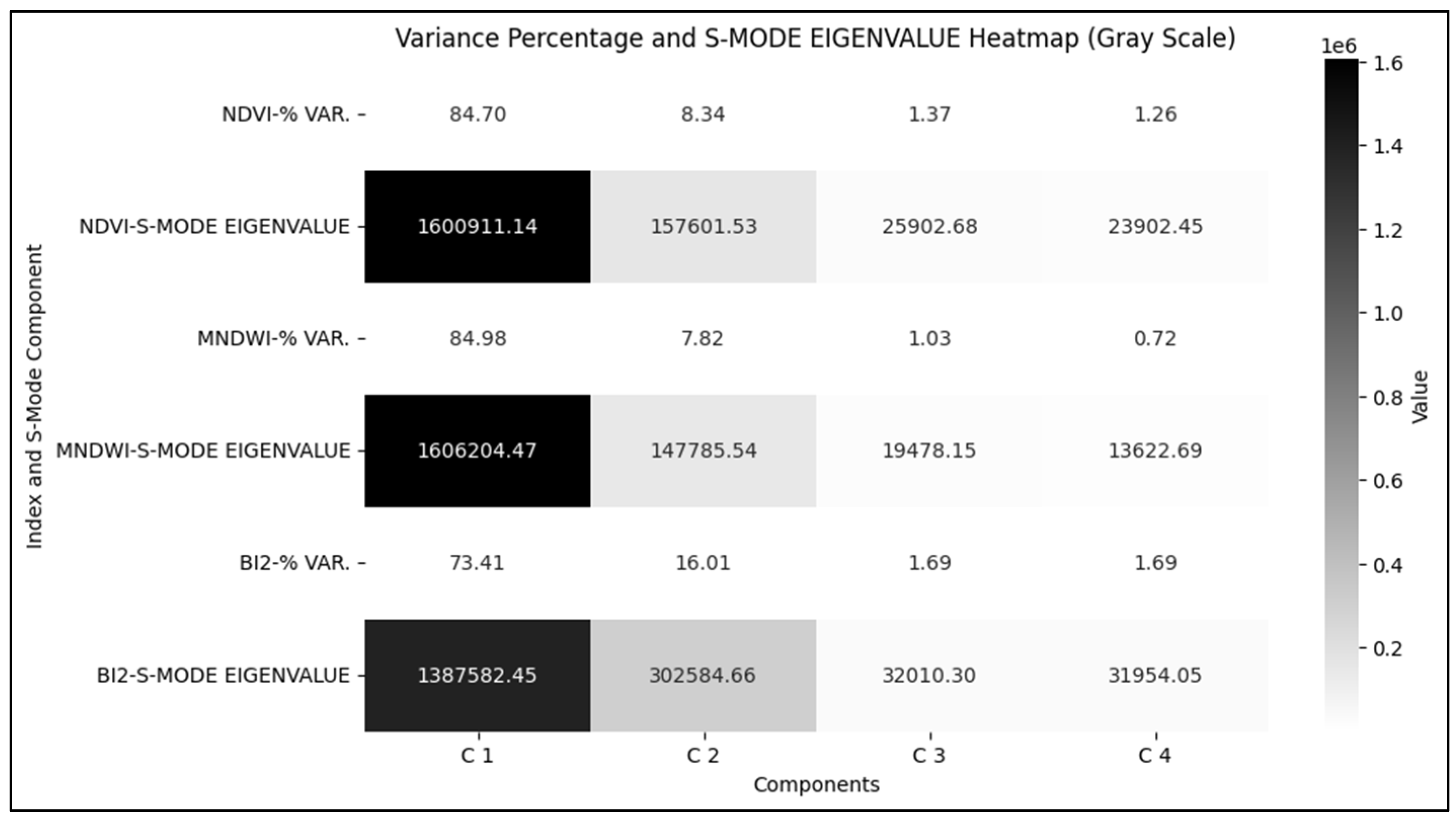
Figure 7.
Score profiles, S-Mode by hydrogeomorphological attribute in the 2019–2024 time series. The black, red, green, and blue lines represent C1, C2, C3, and C4, respectively; and loading profiles of (a) NDVI; (b) MNDWI; (c) BI2.
Figure 7.
Score profiles, S-Mode by hydrogeomorphological attribute in the 2019–2024 time series. The black, red, green, and blue lines represent C1, C2, C3, and C4, respectively; and loading profiles of (a) NDVI; (b) MNDWI; (c) BI2.
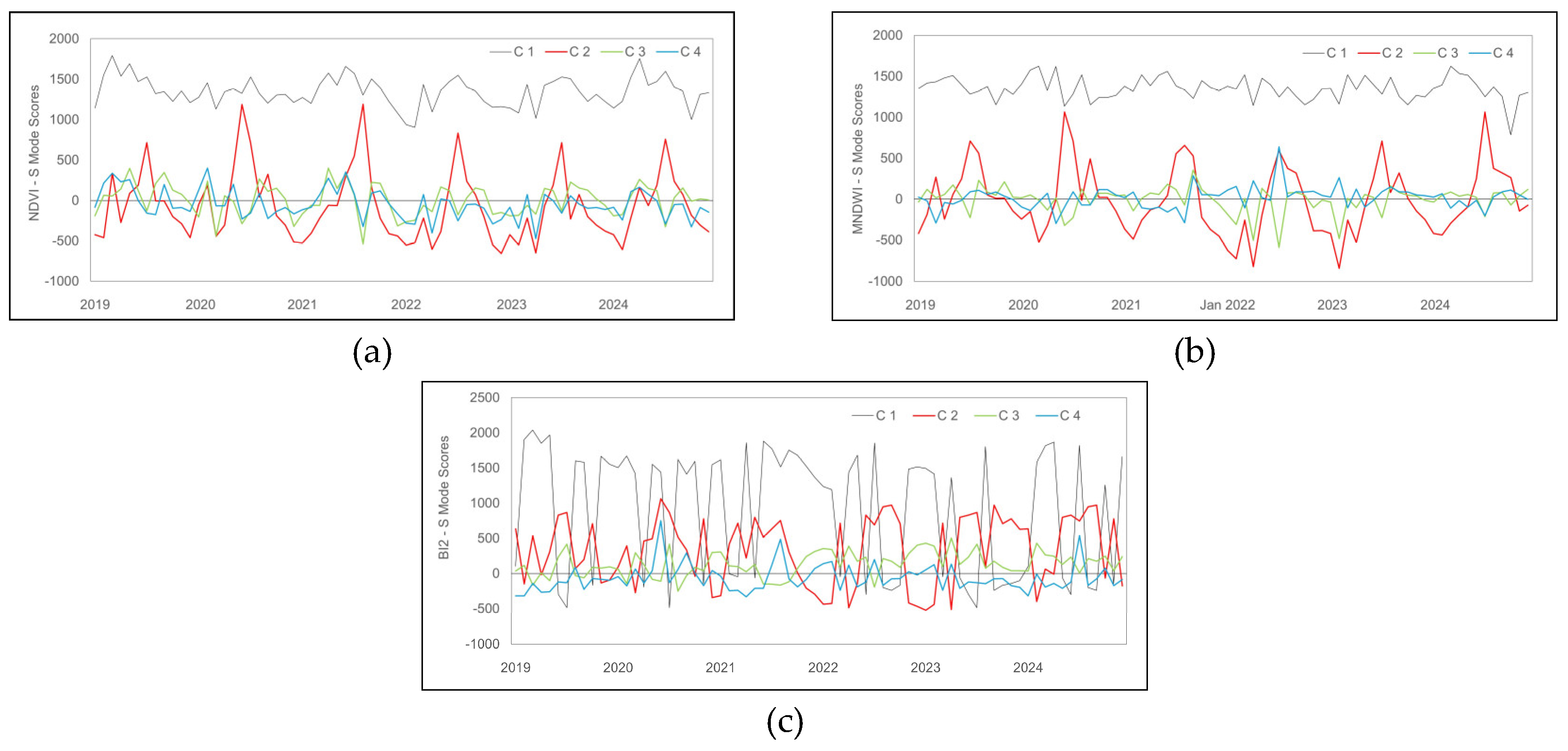
Figure 8.
False-color composite of components C2, C3, and C4, S-Mode, in hydrogeomorphological compartments and wetland typologies of study area. NDVI (N, O, P, Q); MNDWI (R, S, T, U); BI2 (V, X, Z, K).
Figure 8.
False-color composite of components C2, C3, and C4, S-Mode, in hydrogeomorphological compartments and wetland typologies of study area. NDVI (N, O, P, Q); MNDWI (R, S, T, U); BI2 (V, X, Z, K).

Figure 9.
Scatter diagrams of the hydrogeomorphological attributes, C2, T-Mode. (a): NDVI (dependent variable) and MNDWI (predictor variable); (b) BI2 (dependent variable) and MNDWI (predictor variable); (c) NDVI (dependent variable) and BI2 (predictor variable).
Figure 9.
Scatter diagrams of the hydrogeomorphological attributes, C2, T-Mode. (a): NDVI (dependent variable) and MNDWI (predictor variable); (b) BI2 (dependent variable) and MNDWI (predictor variable); (c) NDVI (dependent variable) and BI2 (predictor variable).
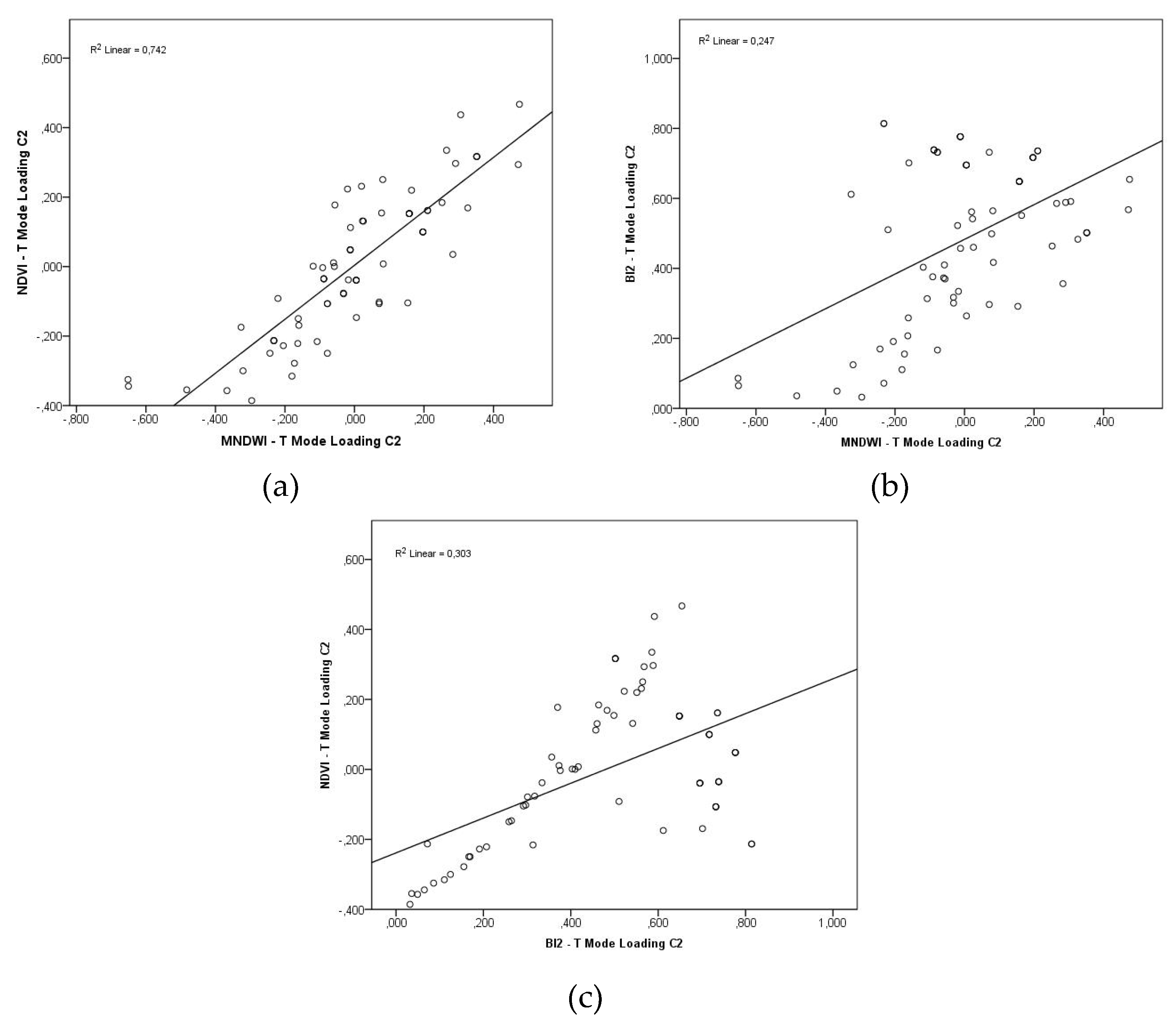
Figure 10.
Scatter diagrams of the hydrogeomorphological attributes, C2, S-Mode. (a): NDVI (dependent variable) and MNDWI (predictor variable); (b) BI2 (dependent variable) and MNDWI (predictor variable); (c) NDVI (dependent variable) and BI2 (predictor variable).
Figure 10.
Scatter diagrams of the hydrogeomorphological attributes, C2, S-Mode. (a): NDVI (dependent variable) and MNDWI (predictor variable); (b) BI2 (dependent variable) and MNDWI (predictor variable); (c) NDVI (dependent variable) and BI2 (predictor variable).
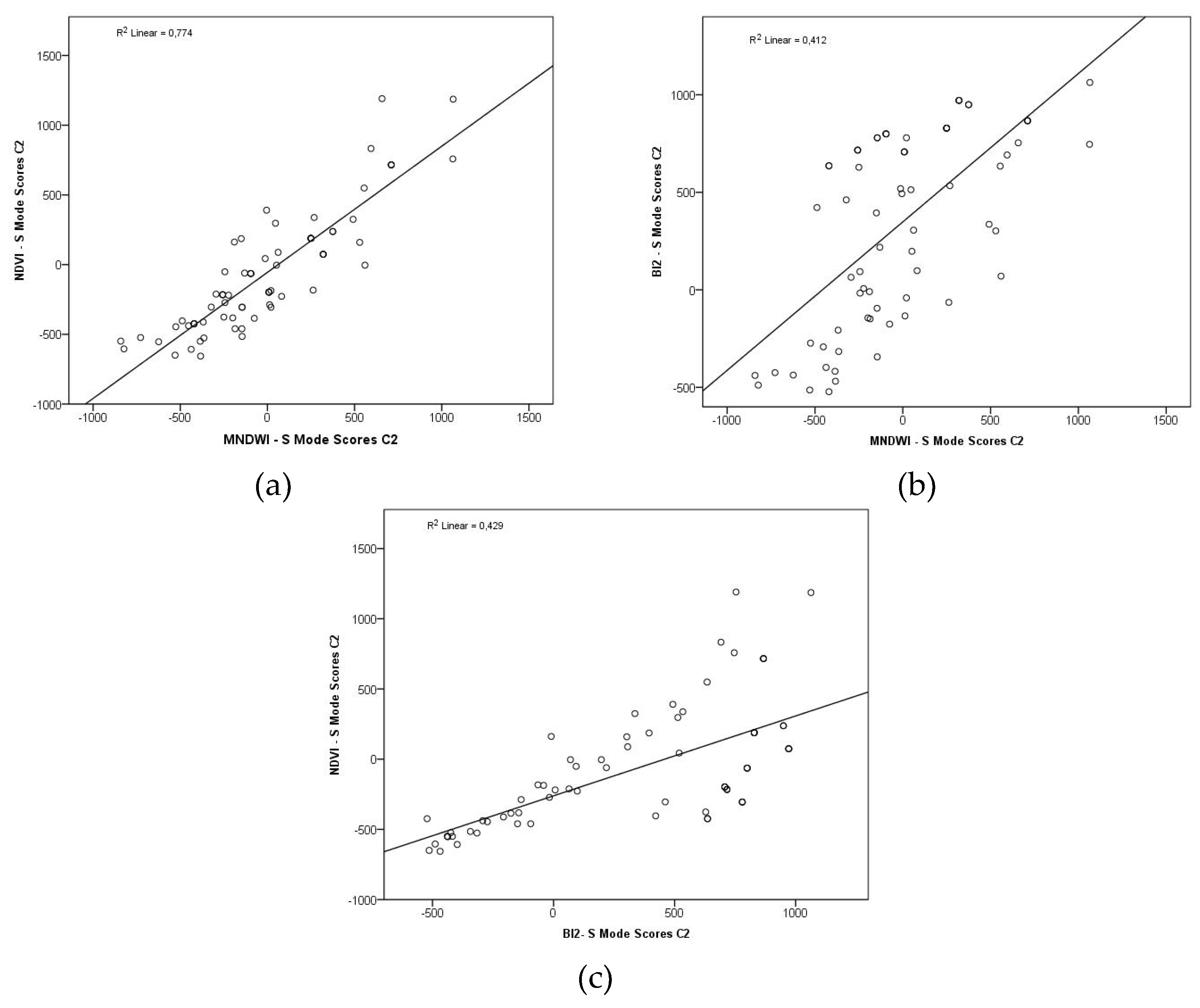
Figure 11.
Estimated values of precipitation, evapotranspiration, and water balance from 2019 to 2024. Component 2 (C2) loadings and scores by hydrogeomorphological attribute, presented for both T-Mode and S-Mode analyses. NDVI (a, d), MNDWI (b, e), BI2 (c, f).
Figure 11.
Estimated values of precipitation, evapotranspiration, and water balance from 2019 to 2024. Component 2 (C2) loadings and scores by hydrogeomorphological attribute, presented for both T-Mode and S-Mode analyses. NDVI (a, d), MNDWI (b, e), BI2 (c, f).
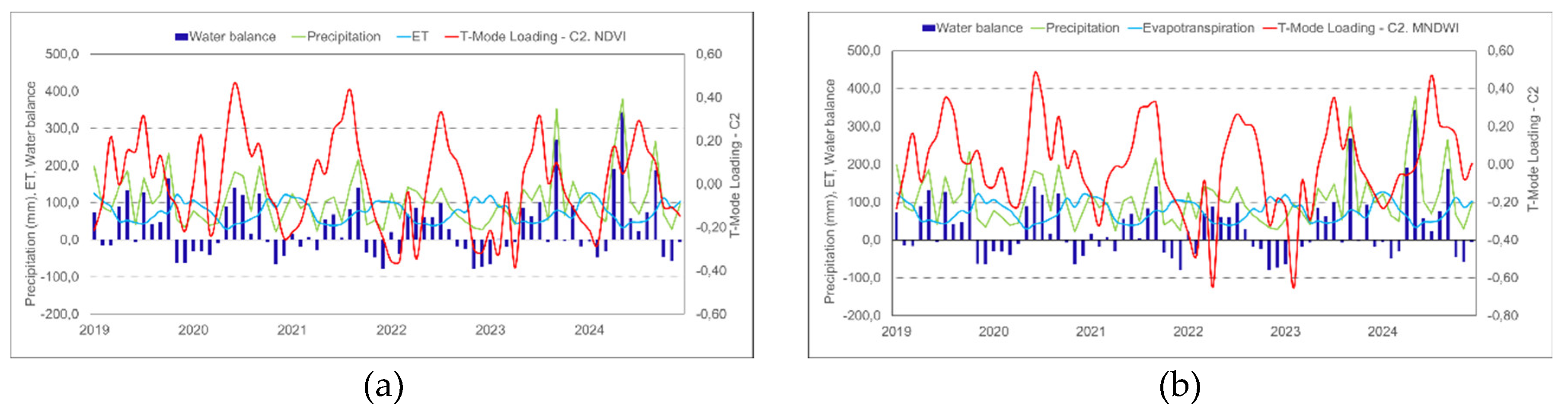
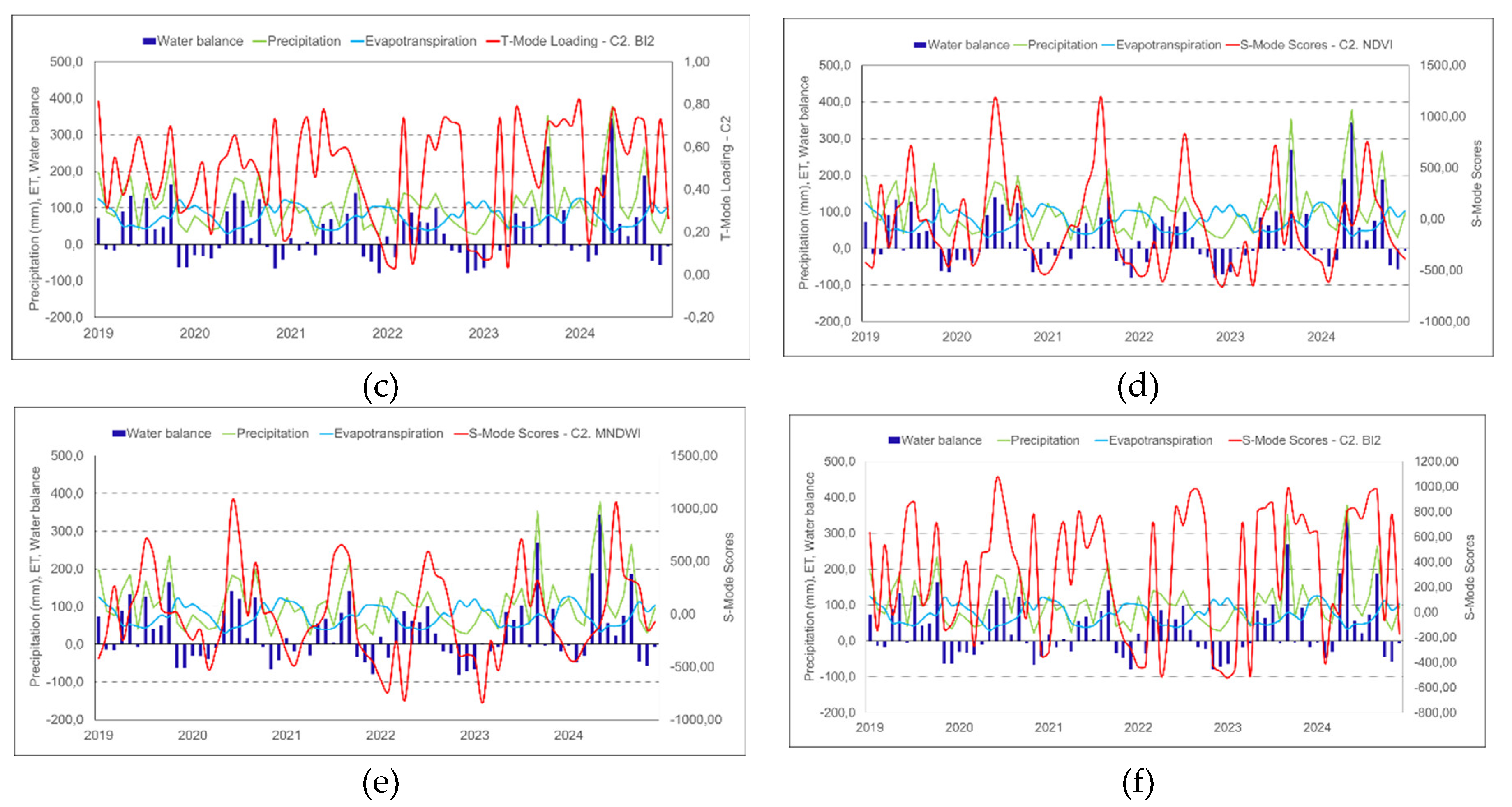

Table 1.
Spectral indices used in the study.
| Hydrogeomorphological attribute / Variable | Formula | Reference |
|---|---|---|
| Normalized Difference Vegetation Index (NDVI) | [48] | |
| Modified Normalized Difference Water Index (MNDWI) | [49] | |
| Second Brightness Index (BI2) | [50] |
Table 2.
Pearson Correlation Matrix – C2, T-Mode.
| NDVI | MNDWI | BI2 | ||
| NDVI | Pearson Correlation | - | - | - |
| p value | - | - | - | |
| MNDWI | Pearson Correlation | 0,862 | - | - |
| p value | p < 0,001 | - | - | |
| BI2 | Pearson Correlation | 0,550 | 0,497 | - |
| p value | p < 0,001 | p < 0,001 | - | |
Table 3.
Pearson Correlation Matrix – C2, S-Mode.
| NDVI | MNDWI | BI2 | ||
| NDVI | Pearson Correlation | - | - | - |
| p value | - | - | - | |
| MNDWI | Pearson Correlation | 0,880 | - | - |
| p value | p < 0,001 | - | - | |
| BI2 | Pearson Correlation | 0,655 | 0,642 | - |
| p value | p < 0,001 | p < 0,001 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



