
Submitted:
07 May 2025
Posted:
08 May 2025
You are already at the latest version
Abstract
Global warming is driven by the increasing atmospheric emissions of greenhouse gases. Soils are highly sensitive to climate change and can shift from carbon reservoirs to carbon sources under warmer and wetter conditions. This study is the first to simultaneously measure trace gas fluxes in Euterpe oleracea (açaí) plantations in upland areas, contrasting them with floodplain areas managed for açaí production in the eastern Amazon. Flux measurements were conducted during both the rainy and dry seasons using the closed dynamic chamber technique. In upland areas, CO2 fluxes exhibited spatial (plateau vs. lowland) and temporal (hourly, daily, and seasonal) variations. During both the rainy and dry months, CH4 uptake in upland soils was higher in lowland areas compared to the plateau. When comparing the two ecosystems, upland area emitted more CO2 during the rainy season, while floodplain areas released more CH4 into the atmosphere. Unexpectedly, during the dry season, floodplain soils produced more CO2 and captured more CH4 from the atmosphere compared to upland soils. In upland areas, CO2-equivalent production reached 59.1 Mg CO2-eq ha-1 yr-1, while in floodplain areas, it reached 49.3 Mg CO2-eq ha-1 yr-1. Soil organic matter plays a vital role in preserving water and microorganisms, enhancing ecosystem productivity in uniform açaí plantations and intensifying the transfer of CH4 from the atmosphere to the soil. However, excessive soil moisture can create anoxic conditions, block gas diffusion, reduce soil respiration, and potentially turn the soil from a sink into a source of CH4.
Keywords:
soil carbon flux
; forest management
; soil microorganism
; Amazon
1. Introduction
Global warming is caused by the increase in atmospheric emissions of greenhouse gas (GHG), resulting from the burning of fossil fuels and land-use changes [1]. This rise in atmospheric temperature is already significantly impacting the behavior of terrestrial ecosystems, such as the reduction in soil moisture (IPCC, 2018), which is an important regulator of GHG [2], especially in estuarine regions. In natural systems, soil respiration significantly contributes to the flux of CO2 into the atmosphere [3]. It is estimated that soil can store 23.8 Gt of CO2-eq annually on a global scale [4]. Recent studies indicate that tropical soils are susceptible to climate change and may shift from being a carbon reservoir to a carbon producer under warmer and wetter conditions [5]. This is due to the influence of soil temperature and moisture [6] on substrate quality, managed area, and the diversity of soil organisms [2], factors that affect GHG fluxes.
The Brazilian Amazon is primarily divided into terra firme (upland) and várzea (floodplain), representing 87% and 13%, respectively, of its total area of 5.5 million km² [7]. Soil saturation, whether in upland or floodplain, alters organic matter decomposition processes due to changes in redox conditions [8,9]. Soil GHG emissions are closely linked to biological activities, which interact with flooding patterns and land management practices [10]. In the Amazon estuarine region, floodplain areas experience daily cycles of flooding and dry, driven by oceanic tides [11]. These tidal patterns are indirectly influenced by lunar phases and seasonal rainfall variations [12].
The upland region also experiences zones of anoxia during rainy periods, leading to the CH4 production [9]. This gas can also be released due to increased termite activity resulting from higher litter accumulation [13]. However, there is limited understanding of CO2 and CH4 fluxes following the transformation of degraded areas into productive lands or secondary forests. Agricultural soils are a primary source of GHG emissions, with approximately 70% of CH4 production originating from anthropogenic sources and 30% from natural processes [14]. The pressure to convert tropical forests into productive regions is alarming [15]; yet, vast areas of already degraded land hold significant potential for restoration and productivity.
Açaí (Euterpe oleracea) is a hyperdominant species native to the Amazon, thriving naturally in floodplain regions with hydromorphic soils that drain water twice daily along river margins [11]. The increasing demand for açaí fruit is transforming floodplain forests into monocultures, significantly altering ecosystem structures and services [16,17]. The economic demand for açaí has driven its cultivation to upland irrigated areas [18], transitioning from an extractive production of 4.2 Mg fruits ha-1 to irrigated upland plantations producing 15.0 Mg fruits ha-1 [19]. Typically, açaí cultivation in uplands (ATF) is carried out in abandoned or already degraded areas [20]. This expansion in açaí production, both in floodplains and uplands, may obscure environmental risks [21]. This research aims to evaluate the contribution of CO2 and CH4 fluxes in intensively managed areas for açaí production in floodplain and irrigated upland regions of the eastern Amazon, to enhance global GHG models.
2. Material and Methods
The floodplain area of the estuary (AV) studied (Figure 1C) is located in the municipality of Belém (1°30’02.0” S and 48°27’31.6” W). The climate type is classified as Af according to the Köppen classification [22], with an annual average temperature of 27 °C and annual precipitation of 3,286 mm [23]. The soils are classified as Haplic Gleysol, characterized by a high proportion of silt and clay, and a low proportion of sand [24]. The floristic composition was previously more diverse [11] but is now predominantly composed of Euterpe oleracea Mart. [17] with 960 clumps per hectare, averaging three stems per clump.
The upland area (ATF) studied (Figure 1D) is located in the municipality of Santa Maria do Pará (1°20’10.0” S and 47°30’04.0” W). The climate is classified as Af according to the Köppen classification [22], with an average annual air temperature of 27 °C and annual precipitation of 2,250 mm [23]. The soils are classified as Yellow Latosol with a medium sandy texture [24]. In this already degraded area, açaí was planted in 2011 with a spacing of 5 m x 5 m, resulting in a density of 400 clumps per hectare. Each clump can contain up to three stems and is organically fertilized, without the application of soluble chemical fertilizers.
2.1. Experimental Design
2.1.1. Experiment on Upland
The measurements to detail spatial variation in ATF were conducted in September 2020 (dry season), simultaneously in high topography (plateau, Top1) and low topography (lowland, Top2). In each topography four rings were fixed in the planting line (L) and four in street (R) where the machines operate, in a homogeneous açaí plantation area (Figure 2A) located in the municipality of Santa Maria do Pará (Figure 1B and D). Simultaneous measurements were taken hourly, from 09:00 to 17:00 (local time), from September 21 to 25, 2020. For the same time interval, measurements in AV was conducted one week earlier, over two consecutive days, in high and low floodplain areas, respectively, using eight flux chambers at a single time point each day (Figure 2b).
2.1.2. Comparison Between Dry Land and Floodplain
During the rainy season (April 3 to 7, 2021), only the high topography was evaluated in ATF and AV. The measurements were conducted simultaneously from 08:00 to 17:00. In all GHG flux analyses in ATF, due to the uniformity of the planting, four sequential measurements were taken every hour in L and four in R (Figure 2A) over five consecutive days. In AV, the measurement points were randomly allocated in a 700 cm diameter circle (Figure 2B). The measurements took place on the same days and times as the measurements conducted in ATF.
2.2. Trace Gas Flux Measurements
The closed dynamic chamber methodology [9] was utilized to measure soil CO2 (FCO2) and CH4 (FCH4) fluxes. The flux chambers were constructed with polyvinyl chloride (PVC) rings measuring 12.0 cm in height and 20.0 cm in diameter, inserted approximately 4.0 cm deep into the soil (Figure 2). During each measurement period, the rings were sequentially closed with a lid for three minutes, forming the flux chamber. This chamber was connected to a Los Gatos portable gas analyzer (Ultra-portable Greenhouse Gas Analyzer, U.S.A.), which recorded the gas concentrations (ppm) inside the chamber at two second intervals [25]. After the measurements, the height of each ring was measured at four equidistant points using a ruler. The FCO2 and FCH4 values were calculated based on the increase/decrease in GHG concentrations within the chamber [6,26]. The flux was considered zero when the linear regression achieved an R2 < 0.30 [27].
2.3. Soil Sampling and Analysis and Environmental Characterization
After each measurement period (ATF and AV), six soil samples were collected using an auger at a depth of 0-10 cm. In the ATF, three samples were collected in the L and three in the R at each flux measurement point (Figure 2). The samples were appropriately conditioned and sent to the Chemical Analysis Laboratory of the Emílio Goeldi Museum, located in Belém (PA).
The concentration of C and N in microbial biomass was investigated using the soil microwave irradiation method [28], conducted only during the dry period due to a lack of chemicals in the laboratory. The determination of microbial biomass carbon (Cm) was performed through Dichromate oxidation [29,30]. The quantification of nitrogen in microbial biomass (Nm) followed the method by Brookes et al., (1985), substituting fumigation with irradiation. For this, we used the equation proposed by Jenkinson (1988), which calculates the difference between the amount of N in irradiated and non-irradiated soil divided by the constant k (k = 0.45). Samples of fine (diameter ≤ 0.2 mm; RF) and coarse (diameter > 0.2 mm; RG) root biomass were collected during both dry and rainy seasons, at a depth of 10 cm, in the previously described ATF and AV locations. After separation from the soil, the roots were oven-dried at 65 °C for 72 hours and weighed using an analytical balance [26]. Soil pH was measured using a potentiometer in deionized water, calibrated with standard solutions of pH 4.0 and pH 7.0 [33].
2.4. Environmental Characterization
Precipitation data for the AV were provided by the National Institute of Meteorology [23], with the automatic data collection station located in Belém (1°26’09.00” S and 48°26’14.00” W). For the ATF, precipitation data were supplied by the National Water and Basic Sanitation Agency [34], with the meteorological station located in Santa Maria do Pará (1°21’24.5” S; 47°34’27.5” W). Soil moisture determination (Us; %) was conducted using the Gravimetric Method [33]. During the trace gas flux measurements, soil temperature (Ts, °C) was quantified using a portable digital thermometer (TP101). Additionally, air temperature (Ta, °C) and relative humidity (HR, %) were recorded every 5 minutes using a Hobo pro V2 data logger.
2.5. Statistical Analysis
The topographical variation in ATF was compared to that of the estuary's AV. In the ATF, hourly and daily analyses were conducted using subdivided plots, with two treatments (high and low topography) divided into line (L) and street (R). The Shapiro-Wilks method was used to assess the normality of the FCH4 and FCO2 data and the soil physicochemical parameters. When normality was not achieved, logarithmic transformations were applied. Student's t-test was used to determine if there were differences in the means between the times and days of data collection at the same site and between the studied sites. Significant differences (p < 0.05) in flux between the different locations were evaluated using ANOVA and Tukey's LSD test. Pearson correlation coefficients were calculated to establish the connections between environmental variables and gas fluxes in the months (dry and rainy seasons) when soil chemical characteristics were analyzed simultaneously with gas flux measurements. The free statistical software Infostat 2015® was used to perform the statistical analyses.
3. Results
3.1. Carbon Dioxide and Methane Flux
3.1.1. Spatial Analysis of Homogeneous Açaí Planting in the Dry Season
The analysis of gas flux across different topographies (Top1 and Top2) was conducted simultaneously along the line (L) and street (R) in a homogeneous açaí plantation on upland soil during the dry season. The results are presented as mean ± standard error. The FCO2 was significantly higher (p < 0.01) in Top2 (19.561 ± 0.522 g CO2 m-2 d-1) compared to Top1 (7.416 ± 0.292 g CO2 m-2 d-1). In Top1, there was significant variation (p < 0.01) between the analyzed days. The highest fluxes were observed on the fifth day of sampling (15.997 ± 0.229 g CO2 m-2 d-1), compared to the second day (9.827 ± 0.688 g CO2 m-2 d-1), with both being significantly higher (p < 0.01) than the first (6.900 ± 0.229 g CO2 m-2 d-1) and third days (5.057 ± 0.187 g CO2 m-2 d-1). The third day did not differ statistically (p > 0.05) from the fourth day of analysis (4.772 ± 0.215 g CO2 m-2 d-1). In Top2, there was no significant variation (p > 0.05) in FCO2 over the five days of analysis.
Only in the plateau topography (Top1) did FCO2 vary significantly between the analyzed time periods, being higher (p < 0.05) between 09:00 and 12:00 compared to other times (Figure 3A). In Top1, FCO2 levels were higher in the early morning (09:00 to 12:00) compared to other times (Figure 3C), with significantly higher values (p < 0.01) in L (8.187 ± 0.449 g CO2 m-2 d-1) compared to R (6.582 ± 0.362 g CO2 m-2 d-1). However, in Top2, the fluxes did not vary between time periods, but were significantly higher (p < 0.001) in L (25.834 ± 0.734 g CO2 m-2 d-1) compared to R (13.017 ± 0.253 g CO2 m-2 d-1) across all time periods (Figure 3E).
On all analyzed days, there was a consumption of CH4 from the atmosphere by soil bacteria (Figure 3). On average, the CH4 influx was significantly higher (p < 0.001) in Top2 (-0.540 ± 0.037 mg CH4 m-2 d-1) compared to Top1 (-0.291 ± 0.038 mg CH4 m-2 d-1). In Top1, the CH4 influx was significantly higher on the fifth day of sampling (-0.835 ± 0.155 mg CH4 m-2 d-1), while no significant differences were observed between the other days (-0.236 ± 0.038 mg CH4 m-2 d-1). In Top2, no significant variation (p > 0.05) in CH4 influx was observed across the analyzed days. The CH4 influx was higher during the morning (until 12:00) compared to the afternoon in Top1 (Figure 3D), with no significant difference (p = 0.221) in fluxes between L (-0.259 ± 0.054 mg CH4 m-2 d-1) and R (-0.383 ± 0.057 mg CH4 m-2 d-1). In Top2, the influx was also higher in the morning compared to the afternoon (Figure 3F), with a significant variation (p < 0.01) in FCH4 between L (-0.683 ± 0.054 mg CH4 m-2 d-1) and R (-0.440 ± 0.052 mg CH4 m-2 d-1).
3.1.2. Simultaneous Flow Measurements in Upland and Floodplains
Simultaneous flux measurements were conducted during the rainy season in March 2021, exclusively in Top1 (higher topography), covering both upland (ATF) and floodplain (AV) areas. When comparing simultaneous measurements in ATF and AV, the distribution of GHG fluxes did not follow a normal distribution (p < 0.001) in either location, even after logarithmic transformation. Consequently, a non-parametric test was selected for comparing the means. The average FCO2 in ATF (20.691 ± 0.563 g CO2 m-2 d-1) was significantly higher (H = 101.532; p < 0.001) than that observed in AV (12.869 ± 0.475 g CO2 m-2 d-1). During the rainy season, FCO2 in ATF did not show significant variation (H = 6.906, p = 0.647) across the sampled times. However, in AV, FCO2 levels were significantly lower (H = 45.171, p < 0.001) in the late afternoon, starting from 15:00 (Figure 4A).
During the rainy season, there was an influx of CH4 in ATF (-0.464 ± 0.038 mg CH4 m-2 d-1), which was significantly lower (H = 137.451, p < 0.001) than the efflux observed in AV (1.278 ± 0.255 mg CH4 m-2 d-1). In ATF, the CH4 influx was significantly higher (H = 62.835, p < 0.001) between 10:00 and 13:00 compared to the other analyzed time periods (Figure 4B). However, during the same rainy season, in AV, the fluxes did not vary significantly (H = 4.755, p = 0.804) across the analyzed time periods (Figure 4B).
3.2. Seasonal Flux of Greenhouse Gases
When comparing GHG fluxes along the L and R in ATF during the two months of the rainy season on the plateau (Top1-Rainy) and the dry season on both the plateau (Top1-Dry) and lowland (Top2-Dry), a significant difference was observed between the sampled locations (Figure 5A). During both seasons, in Top1 and Top2 (only during the dry season), FCO2 was significantly higher (p < 0.001) in L compared to R (Figure 5). However, regarding FCH4, only in Top2 during the dry season was the CH4 influx significantly higher in L compared to R (Figure 5B), while no significant differences (p > 0.05) were observed in the other measurements.
In March (rainy season), when fluxes were measured simultaneously, FCO2 was significantly higher (H = 101.532, p < 0.001) in ATF (20.691 ± 0.563 g CO2 m-2 d-1) compared to AV (12.869 ± 0.475 g CO2 m-2 d-1) (Figure 6A). However, in September (dry season), FCO2 was significantly lower (H = 290.921, p < 0.001) in ATF at Top1 (7.416 ± 0.292 g CO2 m-2 d-1) than at Top2 (19.561 ± 0.522 g CO2 m-2 d-1), where the efflux did not differ significantly from FCO2 in AV (20.647 ± 1.741 g CO2 m-2 d-1) (Figure 6C). When considering both topographies in ATF as a single high topography (Top1), the fluxes in AV were significantly higher (H = 15.664, p < 0.001) than in ATF (13.590 ± 0.400 g CO2 m-2 d-1). In ATF, the FCO2 measured during the rainy season was significantly higher (H = 102.696, p < 0.001) than the efflux during the dry season. Conversely, in AV, the CO2 efflux during the dry season was significantly higher (H = 23.636, p < 0.001) than during the rainy season.
In March (rainy season), when the flows were measured simultaneously in the two places, the FCH4 was higher (H = 137.451, p < 0.001) in AV (1.278 ± 0.255 mg CH4 m-2 d-1) compared to the influx in ATF (-0.464 ± 0.0375 mg CH4 m-2 d-1) (Figure 6B). However, in September (dry season), the CH4 influx in the high topography (Top1) of AV (-0.798 ± 0.179 mg CH4 m-2 d-1) did not differ from the CH4 influx in the lowland (Top2) of ATF (-0.539 ± 0.037 mg CH4 m-2 d-1), both of which were higher (H = 52.422, p < 0.001) than the CH4 influx on the Top1 (-0.292 ± 0.039 mg CH4 m-2 d-1) (Figure 6D). Considering the average flux rates of the two topographies in the ATF, the CH4 influx in AV was higher (H = 10.302, p < 0.001) than the CH4 influx in ATF (-0.418 ± 0.0273 mg CH4 m-2 d-1). In ATF, the CH4 influx measured during the rainy season did not differ (p > 0.05) from that during the dry season. However, in AV, the CH4 efflux during the rainy season was higher (H = 33.407, p < 0.001) than the influx during the dry season.
3.3. Environmental Variables
The total annual precipitation (from August 2020 to July 2021) was 2,634.0 mm in the Santa Maria do Pará region (ATF) and 3,915.3 mm in the Belém region (AV) (Figure 7A). For both ATF and AV, the wettest months span from December to May, with climatological totals of 1,588.0 mm and 1,760.0 mm, accounting for 81.5% and 83.6% of the total annual precipitation, respectively (Figure 7B). Precipitation was consistently lower in ATF compared to AV, except during the months of May, July, and August (Figure 7A). During the study period, rainfall exceeded the climatological averages by 538 mm and 1,596 mm in ATF and AV, respectively, coinciding with a La Niña event (Figure 7B). In both locations, precipitation was above the climatological average (values above zero) for most of the months studied, particularly during the dry season (Figure 7B).
Air temperature and relative humidity (UR) did not follow a normal distribution, necessitating the use of non-parametric tests to compare the means. Temperatures were highest between 12:00 and 14:00 (Figure 8) in both ATF (H = 264.327, p < 0.001) and AV (H = 128.898, p < 0.001). Additionally, temperatures were higher during the dry season compared to the rainy season in both ATF (H = 215.845, p < 0.001) and AV (H = 97.727, p < 0.001). The highest temperatures (H = 405.651, p < 0.001) were recorded in AV during the dry season (30.98 ± 0.10 °C), followed by ATF during the dry season (29.05 ± 0.12 °C). These values were significantly higher than those recorded in ATF (27.92 ± 0.14 °C) and AV (27.38 ± 0.10 °C) during the rainy season, respectively.
Relative humidity (UR) was higher at 09:00 and 17:00 (Figure 8) in both ATF (H = 175.494, p < 0.001) and AV (H = 157.412, p < 0.001). As expected, UR was higher during the rainy season compared to the dry season in both ATF (H = 368.694, p < 0.001) and AV (H = 124.181, p < 0.001). There was a significant variation when comparing the locations and seasons. During the rainy season, UR was higher in AV (89.59 ± 0.37%) compared to ATF (83.13 ± 0.47%), which did not differ significantly from AV during the dry season (82.65 ± 0.45%). Both of these values were higher than those recorded in ATF during the dry season (67.27 ± 0.38%).
In ATF, soil temperature (Ts) was higher than in AV during both the rainy season (H = 30.10, p < 0.001) and the dry season (H = 16.78, p < 0.001). At both study sites, there was significant variation in Ts across the different sampling times during both the rainy and dry seasons. In ATF, the warmest Ts values were recorded between 11:00 and 15:00 (Figure 8A and 8C), whereas AV did not show significant variation in Ts throughout the day (Figure 8B and 8D).
Soil moisture (Us) was higher during the rainy season (Table 1) in both ATF (H = 14.29, p < 0.001) and AV (H = 7.50, p < 0.01). Additionally, Us was consistently higher in the floodplain area (AV) compared to ATF during both the rainy (H = 9.38, p < 0.001) and dry seasons (H = 10.59, p < 0.001). Soil pH did not show statistically significant differences (p > 0.05) between the seasons in either of the sampled areas (Table 1). However, pH was higher in ATF during both the rainy (H = 7.38, p < 0.01) and dry seasons (H = 4.27, p < 0.05) when compared to AV. Fine root biomass did not differ significantly (p > 0.05) either seasonally within each site or between the sites within the same season (Table 1).
In ATF, root biomass (RG) was higher during the rainy season (H = 8.20, p < 0.01), whereas in AV, it was higher during the dry season (H = 4.34, p < 0.05). However, when comparing the two sites, RG differed only during the rainy season (Table 1), being higher in ATF (H = 13.95, p < 0.001) compared to AV (Table 1). Microbial carbon (Cmic) and microbial nitrogen (Nmic) were analyzed only during the dry season, with both Cmic and Nmic being higher in AV (HCmic = 8.00, p < 0.01; HNmic = 8.00, p < 0.01) compared to ATF (Table 1).
3.4. Correlations Between Flow and Environmental Variable
In ATF during the rainy season, FCO2 showed positive correlations with Us, RG, TR, and Ta, while exhibiting negative correlations with RF, UR, Pa, and pH (Table 2). Conversely, FCH4 was positively correlated with Us, UR, and Pa, and negatively correlated with Ts and Ta (Table 2). During the dry season, FCO2 was positively correlated with Us, RF, Cm, and pH, but negatively correlated with FCH4, Ts, and RG (Table 2). Meanwhile, FCH4 was positively correlated with Ts, RG, and Nm, and negatively correlated with Us, RF, Pa, Cm, and pH (Table 2).
In AV during the rainy season, FCO2 was positively correlated with FCH4, Ts, RF, RG, TR, and Ta, while showing negative correlations with Us, UR, Pa, and pH (Table 2). In the same season, FCH4 did not show significant correlations with any of the analyzed variables, except for a positive correlation with FCO2. During the dry season, only FCO2 exhibited positive correlations with Ta and UR (Table 2).
4. Discussion
4.1. Soil Carbon Flux in Açaí Plantation on Dry Land
During the dry season, in a La Niña year when rainfall significantly exceeded the climatological average (Figure 7B), the soil FCO2 on the plateau (Top1) in ATF was considerably lower than the efflux in the lowland (Top2) area (Figure 6B). These results differ markedly from those observed in a mangrove forest in the Amazon estuary, where simultaneous CO2 effluxes were higher in elevated topography compared to low-lying areas [26]. On the other hand, in a pristine tropical forest in the Guianas, no variation in CO2 fluxes was found across different topographies, including plateaus and lowlands [35]. An important consideration is that, in our study, as well as in [26], the fluxes were measured simultaneously in both topographies using laboratory-calibrated equipment, with no statistical variation between the instruments.
To explain the difference in CO2 efflux, fine root (RF) biomass was significantly higher (LSD = 0.814; p < 0.01) in Top2 (3.762 ± 0.222 Mg ha-1) compared to Top1 (2.212 ± 0.309 Mg ha-1). Similarly, soil moisture (Us) was significantly higher (LSD = 2.425; p < 0.05) in Top2 (11.085 ± 1.036%) than in Top1 (8.641 ± 0.508%). Additionally, microbial biomass was significantly greater (LSD = 0.106; p < 0.001) in Top2 (0.537 ± 0.047 kg Cmic kg-1 soil) compared to Top1 (0.293 ± 0.021 kg Cmic kg-1 soil). In this context, the higher moisture levels in the lowland area directly influence nutrient and organic matter dynamics, leading to increased fine root biomass [35] and microbial biomass [36], which in turn result in significantly higher CO2 efflux. This may explain the greater soil respiration observed in the lowland area of an organic açaí plantation on upland terrain. Thus, soil organic matter plays a crucial role in maintaining water and microbial activity, enhancing ecosystem productivity in homogeneous açaí plantations.
When comparing the lines (L) and streets (R), the FCO2 during the dry season in Top1 was 8.154 ± 0.450 g CO2 m-2 d-1 in L, which was significantly lower (H = 172.510; p < 0.001) than the 25.825 ± 0.667 g CO2 m-2 d-1 in L in Top2. In R, the FCO2 in ATV in Top1 was 6.672 ± 0.361 g CO2 m-2 d-1, which was also significantly lower (H = 117.794; p < 0.001) than the 13.297 ± 0.263 g CO2 m-2 d-1 in Top2. When comparing the rainy season to the dry season, only in Top1 were both FCO2 in L and R significantly higher (p < 0.001) during the rainy season compared to the dry season (Figure 3A). The results reveal that, in both L and R, the lowland topography (Top2) emits more CO2 into the atmosphere compared to the plateau (Top1), a finding that contrasts with observations in an Acacia mangium plantation in Indonesia [37] and in a primary upland forest in the Amazon [38]. On the other hand, studies conducted in a closed-canopy upland forest in the central Amazon [39] and in a floodplain forest in the estuary (Aroni et al., n.d.) identified significant topographic variation, although, unlike the results presented here, the fluxes were higher in elevated topography compared to lowland areas. In an analysis of the annual average FCO2, the efflux in L (21.025 ± 0.543 g CO2 m-2 d-1) was significantly higher (H = 166.519; p < 0.001) than in R (11.251 ± 0.275 g CO2 m-2 d-1), which could not be explained by root biomass (fine, coarse, or total), as no statistical variation (p > 0.05) was observed between L and R. However, the data show that soil moisture (Us) was significantly higher (H = 5.859; p < 0.05) in L (28.458 ± 3.434%) compared to R (18.423 ± 3.262%). During the dry season, irrigation is applied only to the açaí planting rows (L), which may enhance soil biological activity in upland areas of the Amazon region.
In the upland açaí plantation, the FCO2 in the plateau (Top1) topography was significantly higher (H = 253.615, p < 0.001) during the rainy season (20.691 ± 0.563 g CO2 m-2 d-1) compared to the dry season (7.388 ± 0.292 g CO2 m-2 d-1). Despite daily irrigation during the dry season and the occurrence of a La Niña year, the fluxes were considerably higher during the rainy season, both when comparing the L and the R. Root biomass did not differ (p > 0.05) between the seasons; however, soil moisture (Us) mirrored the differences in FCO2, being significantly higher (H = 19.355, p < 0.001) during the rainy season (23.118 ± 0.765%) compared to the dry season (8.641 ± 0.508%). On the other hand, soil temperature (Ts), although showing only a slight difference, was significantly higher (H = 30.171, p < 0.001) during the dry season (27.989 ± 0.071 °C) compared to the rainy season (27.371 ± 0.075 °C). Several studies in tropical forests have revealed pronounced seasonal fluctuations in CO2 gas fluxes, with higher emissions during the rainy season compared to the dry season [37,40,41,42], a pattern also observed in an agroforestry system in Panama [43]. The data suggest that, despite irrigation during the dry season, it may not be sufficient to maintain soil productivity in upland açaí plantations.
During the dry season, the influx of CH4 from the atmosphere into the soil of the açaí plantation was significantly higher (H = 42.392, p < 0.001) in Top2 (-0.539 ± 0.034 mg CH4 m-2 d-1) compared to Top1 (-0.332 ± 0.024 mg CH4 m-2 d-1). Only in Top2 was the CH4 influx significantly higher (H = 8.134, p < 0.01) in the lines (L) (-0.670 ± 0.051 mg CH4 m-2 d-1) compared to the streets (R) (-0.409 ± 0.052 mg CH4 m-2 d-1) during the dry season. These results differ from those observed in an A. mangium plantation in Indonesia [37], where the CH4 influx from the atmosphere was higher in the plateau compared to the lowland. As previously mentioned, both soil moisture (Us) and microbial biomass were higher in Top2, explaining the greater microbial activity and, consequently, the higher CH4 influx from the planetary atmosphere. We understand that the higher Us in the lowland (11.085 ± 1.036%) was not sufficient to negatively influence the permeability of CH4 and atmospheric O2 into the soil [9,44], allowing the influx of CH4 from the atmosphere to be consumed in the soil.
Only in Top2 was the influx of atmospheric CH4 significantly higher (H = 8.134, p < 0.01) in the L (-0.670 ± 0.051 mg CH4 m-2 d-1) compared to the R (-0.409 ± 0.052 mg CH4 m-2 d-1). As previously shown, in this same topography, soil moisture (Us) was higher in L compared to R, meaning that the activity of methanotrophs was not hindered at Us levels close to 29%. In fact, the higher Us in the planting lines enhanced the activity of these organisms compared to the streets, and the diffusion of CH4 and atmospheric O2 into the soil was not impaired. When comparing only Top1, seasonality did not influence the influx of atmospheric CH4 (p > 0.05), with values of -0.360 ± 0.031 mg CH4 m-2 d-1 during the rainy season and -0.291 ± 0.038 mg CH4 m-2 d-1 during the dry season.
Methane (CH4) has a global warming potential 32 times greater than that of CO2 over a 100-year period [45]. In this context, the removal of 1 Mg (Mega gram) of CH4 has a significant impact on global warming [46]. Additionally, with an average tropospheric lifetime of about 10 years [47], land-use practices that reduce CH4 emissions or contribute to the capture of atmospheric CH4 can play a major role in global climate change mitigation. In this regard, organic açaí plantations in upland areas are estimated to absorb an average of 1.6 ± 0.080 kg CH4 ha-1 yr-1, slightly lower than the values predicted in previous studies for different ecosystems, particularly when compared to the 2.7 kg CH4 ha-1 yr-1 for tropical soils [48] and significantly lower than the 2.5 - 4.0 kg CH4 ha-1 yr-1 removed by a secondary forest enriched with leguminous trees [41]. However, the atmospheric methane uptake is quite close to the 1.0 kg CH4 ha-1 yr-1 reported for a secondary forest in the eastern Amazon [9] and similar to the 1.6 ± 0.9 kg CH4 ha-1 yr-1 removed by the soil in the Tapajós National Forest [49]. Most importantly, land use with organic and homogeneous açaí plantations in upland areas does not produce methane, even with the use of irrigation.
4.2. Soil Carbon Flux During the Rainy Season in Upland Planting Compared to Estuary Floodplain
It is important to note that during the rainy season, measurements in ATF and AV were conducted simultaneously using two laboratory-calibrated devices with no statistical variation between them. The FCO2 in ATF (16.176 ± 0.347 g CO2 m-2 d-1) was significantly higher (H = 19.286, p < 0.001) than the flux in AV (12.813 ± 0.477 g CO2 m-2 d-1). On the sampling days and at all sampled times, the FCO2 was higher in ATF compared to the efflux in AV (Figure 4A). Studies comparing CO2 efflux between upland and floodplain ecosystems are scarce in the literature, especially those that measure simultaneously in both locations. Therefore, our comparison can only be made across distinct sites, though efforts were made to maintain similar ecosystem variations. Two-year measurements conducted in the Caxiuanã National Forest showed that the flux on the plateau (sandy soil) was 21.0% higher than in the lowland (clay soil) [38]. A study conducted in a region with significant topographic variation (25 m between plateau and lowland) in China, found that the plateau area produced 11.9% more CO2 [50]. Similarly, in a topographic variation study in Guyana, the FCO2 was 11.7% higher on the plateau compared to the lowland [35]. Consistent with these studies, our results during the rainy season show a 20.8% higher CO2 production in the upland area (sandy soil - plateau) compared to the floodplain area of the Amazon estuary (clay soil - lowland).
Some studies suggest that the difference in soil FCO2 between plateau and lowland is related to higher root biomass in sandy soils compared to clay soils [51]. In the present study, the higher CO2 efflux in ATF during the rainy season can be explained by root biomass, as the total live root biomass in ATF (11.839 ± 1.041 Mg ha-1) was twice as high (p < 0.001) compared to AV (5.643 ± 0.891 Mg ha-1). However, FCO2 during the rainy season was negatively correlated with fine root (RF) and positively correlated with coarse root (RG) and total root (TR) (Table 2). Unfortunately, we were unable to measure microbial carbon and nitrogen during this period, which would have allowed for a more detailed discussion of these results. Nevertheless, the entry of water into the soil, whether through rainfall, irrigation, or tidal movement, is associated with the immediate replacement of air-filled pores with water. This influx of water can initially expel air from the soil pores and subsequently form an insulating layer, hindering the diffusion of CO2 from the soil to the atmosphere [52], thereby reducing soil respiration [53]. Supporting this, during the rainy season in ATF, FCO2 was positively correlated with soil moisture (Us) and soil temperature (Ts), while negatively correlated with relative humidity (UR) and atmospheric pressure (Pa) (Table 2). Other studies have also indicated correlations between FCO2 and temperature [54,55] as well as soil moisture [56].
During the same rainy season, the CH4 efflux in AV (1.033 mg CH4 m-2 d-1) was significantly higher (H = 122.733; p < 0.001) than the CH4 influx in ATF (-0.360 mg CH4 m-2 d-1). On all analyzed days, a high production of CH4 was observed in AV, while an influx of CH4 was recorded in ATF (Figure 4B). The production/influx of CH4 are microbiological processes predominantly controlled by the absence/presence of oxygen, respectively, and the availability of easily degradable biomass [57]. Consequently, the higher soil moisture (Us) in AV (Table 1) facilitates the proliferation of methanogenic processes (microbial production of CH4). In contrast, aerobic soils in ATF promote methanotrophy (microbial consumption of CH4). This explains the negative correlation with soil temperature (Ts) and the positive correlation with soil moisture (Us) (Table 2), as Ts negatively affects the physiology and proliferation of microorganisms, while Us positively influences root growth, respiration, and microbial activity. In upland areas (ATF) with homogeneous organic açaí plantations, 34.9% of the CH4 produced in managed floodplain areas for açaí production in the estuary is removed daily.
Soils in tropical upland areas are generally considered sinks for atmospheric CH4, and changes in land use, such as the homogeneous planting of açaí in this case, can reduce the intensity of this sink [58]. The influx of atmospheric CH4 in the organic açaí plantation in upland areas during the rainy season was 1.7 ± 0.1 kg CH4 ha-1 yr-1, while the production of CH4 in the estuarine floodplain during the same period was 4.6 ± 0.9 kg CH4 ha-1 yr-1. Concurrent measurements during the rainy season revealed that CH4 production in AV was 2.7 times higher compared to the uptake in ATF (Figure 5B). Studies in the state of Rondônia (Brazil) showed that during the rainy season, pasture soils produced 6.1 kg CH4-C m-2 yr-1, whereas soils in primary forest areas consumed 1.1 kg CH4-C m-2 yr-1 [59]. It is understood that in the Amazon, soils with less than 35–40% of pore space filled with water act as CH4 sinks, while those above these values act as sources [60]. In ATF, the soil had an average water content of 23.1%, while in AV it was 53.3%, which appears to align with this premise. The results presented here fall within the range of consumption reported for fine-textured soils in the Amazon rainforest, which was 1.5 - 2.0 kg CH4 ha-1 yr-1 [9]. Considering that CH4 has a global warming potential 32 times greater than CO2 over a 100-year period [45], the production of CO2-equivalents (CO2-eq) in ATF was 59.1 Mg CO2-eq ha-1 yr-1, while in AV it was 49.3 Mg CO2-eq ha-1 yr-1.
4.3. Annual Soil Carbon Flux in Upland Planting Compared to Estuary Floodplain
FCO2 was positively correlated with FCH4 in AV during the rainy season and negatively correlated in ATF during the dry season (Table 2). In ATF, the sandy soil facilitates air exchange with the atmosphere, enhancing soil respiration and methane oxidation by methanotrophic organisms [61]. This process is hindered in AV due to the highly clayey soil and elevated moisture levels [9,60], as well as the anoxia caused by tidal water movement, which creates conditions favorable for the proliferation of methanogenic bacteria [62].
The increase in CO2 efflux between the rainy and dry seasons (Figure 5A) is likely due to an increase in labile carbon available to microorganisms, greater soil oxygenation, resulting in intense microbial activity, and subsequent degradation of soil organic matter [63,64], which intensifies soil respiration. The onset of the rainy season leads to lower CO2 efflux, which may be necessary to increase soil carbon and recover the carbon lost through decomposition during the dry season [65], when the soil is more aerated.
The efflux/influx of CH4 in ATF during the rainy season (Figure 5B) was positively correlated with soil moisture (Us), relative humidity (UR), and atmospheric pressure (Pa), while negatively correlated with soil temperature (Ts) (Table 2). In contrast, during the dry season in ATF, FCH4 was positively correlated with Ts, coarse roots (RG), total roots (TR), and microbial nitrogen (Nm), and negatively correlated with Us, fine roots (RF), Pa, microbial carbon (Cm), and pH (Table 2). In AV, no significant correlations were found between FCH4 and the studied variables during either season (Table 2). Our study revealed that an increase in soil moisture driven by precipitation during the wet season leads to higher CH4 production. In contrast, during the dry season, elevated soil moisture resulting from irrigation reduces CH4 production. Other studies have similarly demonstrated a correlation between CH4 flux and soil moisture [66,67], highlighting that soil moisture regulates CH4 uptake by controlling the diffusion of methane and oxygen within the soil atmosphere. Thus, we can confirm that waterlogged conditions promote the activity of methanogenic bacteria due to anoxia. However, as soil moisture decreases, reduced moisture levels favor the proliferation of methanotrophic bacteria, enhancing the diffusion and consumption of atmospheric CH4 [68]. This shift in soil moisture may explain the high rates of CH4 consumption observed in AV (Figure 6D). These results demonstrate that high floodplain areas are not always sources of CH4 emissions to the atmosphere; under certain conditions, they can also function as significant sinks for this gas.
A significant positive correlation between variations in soil respiration and soil moisture indicates that, within a certain range, soil moisture is directly proportional to soil respiration. However, excessive increases in soil moisture can create anoxic conditions, hinder gas exchange, reduce respiration, and shift soil from CH4 inflow to efflux. In this sense, the main environmental factors controlling CH4 exchange between soil and the atmosphere are the proportion of pore space occupied by water, the availability and quality of carbon, and soil temperature [44,69,70]. Additionally, root exudates can increase CH4 production in the soil due to the heightened demand for O2 by decomposer organisms, thereby reducing methanotrophy from soil-derived carbon and stimulating intense, short-term changes in soil organic matter recycling (priming effect) [71]. This may explain the strong positive correlation between coarse roots (RG) and total roots (TR) with CH4 flux during the dry season.
We agree that a limited number of observations of CH4 fluxes, and the numerous environmental variables associated with its exchange between soil and atmosphere, constrain the parameterization of process based biogeochemical models [72]. This limitation is particularly significant given the complex interplay of factors influencing methane production, oxidation, and transport, such as soil moisture, temperature, organic matter availability, and microbial community dynamics. As a result, models often rely on simplified representations of these processes, which can lead to uncertainties in predicting CH4 emissions under varying environmental conditions.
To improve model accuracy, there is a pressing need for more comprehensive and high-resolution datasets that capture the spatial and temporal variability of CH4 fluxes across different ecosystems. Additionally, integrating multi-disciplinary approaches, such as combining remote sensing, eddy covariance measurements, and laboratory experiments, could enhance our understanding of the underlying mechanisms driving methane dynamics. Advances in machine learning and data assimilation techniques also offer promising avenues for refining model parameterizations and reducing uncertainties. Ultimately, a more robust representation of CH4 fluxes in biogeochemical models is crucial for accurately projecting future methane emissions and informing climate change mitigation strategies.
5. Conclusion
The upland area produced a total of 59.1 Mg CO2-eq ha-1 year-1, while the floodplain area produced 49.3 Mg CO2-eq ha-1 year-1. During the dry season, spatial heterogeneity in the greenhouse gases flux was identified, with soil CO2 flux and CH4 consumption being higher in the lowland compared to the plateau, across all hours and days analyzed. In the rainy season, simultaneous measurements revealed that the CO2 flux is higher in the upland, while CH4 flux was greater in the floodplain, with atmospheric CH4 consumption on upland. In the floodplain, CH4 production was observed in the rainy month and consumption in the dry month. In the upland açaí plantation area, CO2 fluxes were higher in the planting row compared to the streets, in both seasons of the year, with no variation in the atmospheric CH4 consumption. Soil moisture and temperature appear to control gas fluxes only in the upland, but in contrasting ways during the two seasons analyzed. Soil organic matter plays a vital role in preserving water and microorganisms, enhancing ecosystem productivity in uniform açaí plantations and intensifying the transfer of CH4 from the atmosphere to the soil. However, excessive soil moisture can create anoxic conditions, block gas diffusion, reduce soil respiration, and potentially turn the soil from a sink into a source of CH4. More detailed studies are needed to understand how greenhouse gas fluxes vary across different topographies in upland areas of the Amazon under agricultural production systems.
References
- IPCC Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri R. K, Meyer, L.A., Eds.; Geneva, 2014;
- Verchot, L. V.; Dannenmann, M.; Kengdo, S.K.; Njine-Bememba, C.B.; Rufino, M.C.; Sonwa, D.J.; Tejedor, J. Land-Use Change and Biogeochemical Controls of Soil CO2, N2O and CH4 Fluxes in Cameroonian Forest Landscapes. J. Integr. Environ. Sci. 2020, 17, 45–67. [Google Scholar] [CrossRef]
- Adachi, M.; Ito, A.; Yonemura, S.; Takeuchi, W. Estimation of Global Soil Respiration by Accounting for Land-Use Changes Derived from Remote Sensing Data. J. Environ. Manage. 2017, 200, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Bossio, D.A.; Cook-Patton, S.C.; Ellis, P.W.; Fargione, J.; Sanderman, J.; Smith, P.; Wood, S.; Zomer, R.J.; von Unger, M.; Emmer, I.M.; et al. The Role of Soil Carbon in Natural Climate Solutions. Nat. Sustain. 2020, 3, 391–398. [Google Scholar] [CrossRef]
- Naidu, D.G.T.; Bagchi, S. Greening of the Earth Does Not Compensate for Rising Soil Heterotrophic Respiration under Climate Change. Glob. Chang. Biol. 2021, 27, 2029–2038. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.L. Educational Programs for Team Delivery. Interdisciplinary Education of Health Associates. Acad. Med. 1975, 50, 111–117. [Google Scholar] [CrossRef]
- Alves, N. de O.; Brito, J.; Caumo, S.; Arana, A.; Hacon, S. de S.; Artaxo, P.; Hillamo, R.; Teinilä, K.; Medeiros, S.R.B. de; Vasconcellos, P. de C. Biomass Burning in the Amazon Region: Aerosol Source Apportionment and Associated Health Risk Assessment. Atmos. Environ. 2015, 120, 277–285. [Google Scholar] [CrossRef]
- Denardin, L.G. de O.; Alves, L.A.; Ortigara, C.; Winck, B.; Coblinski, J.A.; Schmidt, M.R.; Carlos, F.S.; Toni, C.A.G. de; Camargo, F.A. de O.; Anghinoni, I.; et al. How Different Soil Moisture Levels Affect the Microbial Activity. Ciência Rural 2020, 50, 1–10. [Google Scholar] [CrossRef]
- Verchot, L. V.; Davidson, E.A.; Cattânio, J.H.; Ackerman, I.L. Land-Use Change and Biogeochemical Controls of Methane Fluxes in Soils of Eastern Amazonia. Ecosystems 2000, 3, 41–56. [Google Scholar] [CrossRef]
- Lira-Guedes, A.C.; Leal, G.; Fischer, G.; Aguiar, L.J.; Júnior, N.J.; Baia, A.L.; Guedes, M. Carbon Emissions in Hydromorphic Soils from an Estuarine Floodplain Forest in the Amazon River. Brazilian J. Environ. Sci. 2021, 56, 413–423. [Google Scholar] [CrossRef]
- Cattanio, J.H.; Anderson, A.B.; Carvalho, M.S. Floristic Composition and Topographic Variation in a Tidal Floodplain Forest in the Amazon Estuary. Rev. Bras. Botânica 2002, 25, 419–430. [Google Scholar] [CrossRef]
- Salo, M.; Sirén, A.; Kalliola, R.; Wild, D.; Harvest, S. Açaí: The Forest Farms of the Amazon Estuary. In Diagnosing Wild Species Harvest; Salo, M., Sirén, A., Kalliola, R., Eds.; Elsevier, 2013; pp. 191–202 ISBN 9780123972040.
- Cattânio, J.H.; Davidson, E.A.; Nepstad, D.C.; Verchot, L.V.; Ackerman, I.L. Unexpected Results of a Pilot Throughfall Exclusion Experiment on Soil Emissions of CO2, CH4, N2O, and NO in Eastern Amazonia. Biol. Fertil. Soils 2002, 36. [Google Scholar] [CrossRef]
- Mosier, A.; Wassmann, R.; Verchot, L.; King, J.; Palm, C. Methane and Nitrogen Oxide Fluxes in Tropical Agricultural Soils: Sources, Sinks and Mechanisms. Environ. Dev. Sustain. 2004, 6, 11–49. [Google Scholar] [CrossRef]
- Carvalho, W.D.; Mustin, K.; Hilário, R.R.; Vasconcelos, I.M.; Eilers, V.; Fearnside, P.M. Deforestation Control in the Brazilian Amazon: A Conservation Struggle Being Lost as Agreements and Regulations Are Subverted and Bypassed. Perspect. Ecol. Conserv. 2019, 17, 122–130. [Google Scholar] [CrossRef]
- Campbell, A.J.; Carvalheiro, L.G.; Maués, M.M.; Jaffé, R.; Giannini, T.C.; Freitas, M.A.B.; Coelho, B.W.T.; Menezes, C. Anthropogenic Disturbance of Tropical Forests Threatens Pollination Services to Açaí Palm in the Amazon River Delta. J. Appl. Ecol. 2018, 55, 1725–1736. [Google Scholar] [CrossRef]
- Freitas, M.A.B.; Vieira, I.C.G.; Albernaz, A.L.K.M.; Magalhães, J.L.L.; Lees, A.C. Floristic Impoverishment of Amazonian Floodplain Forests Managed for Açaí Fruit Production. For. Ecol. Manage. 2015, 351, 20–27. [Google Scholar] [CrossRef]
- Brondizio, E.S.; Moran, E.F. Human Dimensions of Climate Change: The Vulnerability of Small Farmers in the Amazon. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1803–1809. [Google Scholar] [CrossRef]
- Santos, J.C. dos; Sena, A.L. dos S.; Homma, A.K.O. Viabilidade Econômica Do Manejo de Açaizais No Estuário Amazônico: Estudo de Caso Na Região Do Rio Tauerá-Açu, Abaetetuba – Estado Do Pará. In Proceedings of the Sociedade Brasileira de Economia, Administração e Sociologia Rural; Vitória, 2012; pp. 22–25. [Google Scholar]
- Farias Neto, J.T.; Resende, M.D.V.; Oliveira, M. do S.P. Seleção Simultânea Em Progênies de Açaizeiro Irrigado Para Produção e Peso Do Fruto. Rev. Bras. Frutic. 2011, 33, 532–539. [Google Scholar] [CrossRef]
- Tavares, G. dos S.; Homma, A.K.O.; Menezes, A.J.E.A. de; Palheta, M.P. Análise Da Produção e Comercialização de Açaí No Estado Do Pará, Brasil. Int. J. Dev. Res. 2020, 10, 35215–35221. [Google Scholar]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated World Map of the Köppen-Geiger Climate Classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- INMET Instituto Nacional de Meteorologia. Available online: http://www.inmet.gov.br/portal/index.php?r=bdmep/bdmep (accessed on 19 November 2019).
- EMBRAPA Mapas de Solo - Brasil.
- Wilkinson, J.; Bors, C.; Burgis, F.; Lorke, A.; Bodmer, P. Measuring CO2 and CH4 with a Portable Gas Analyzer: Closed-Loop Operation, Optimization and Assessment. PLoS One 2018, 13, e0193973. [Google Scholar] [CrossRef]
- Castellón, S.E.M.; Cattanio, J.H.; Berrêdo, J.F.; Rollnic, M.; Ruivo, M. de L.; Noriega, C. Greenhouse Gas Fluxes in Mangrove Forest Soil in an Amazon Estuary. Biogeosciences 2022, 19, 5483–5497. [Google Scholar] [CrossRef]
- Sundqvist, E.; Vestin, P.; Crill, P.; Persson, T.; Lindroth, A. Short-Term Effects of Thinning, Clear-Cutting and Stump Harvesting on Methane Exchange in a Boreal Forest. Biogeosciences 2014, 11, 6095–6105. [Google Scholar] [CrossRef]
- Islam, K.R.; Weil, R.R. Microwave Irradiation of Soil for Routine Measurement of Microbial Biomass Carbon. Biol. Fertil. Soils 1998, 27, 408–416. [Google Scholar] [CrossRef]
- Kalembasa, S.J.; Jenkinson, D.S. A Comparative Study of Titrimetric and Gavimetric Methods for Determination of Organic Carbon in Soil. J. Sci. Food Agric. 1973, 24, 1085–1090. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An Extraction Method for Measuring Soil Microbial Biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform Fumigation and the Release of Soil Nitrogen: A Rapid Direct Extraction Method to Measure Microbial Biomass Nitrogen in Soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Jenkinson, D.S. Determination of Microbial Biomass C and N in Soil. In Advances in Nitrogen Cycling in Agricultural Ecosystems; Wilson, J.R., Ed.; CAB International: Wallingford, 1988; pp. 368–386. [Google Scholar]
- Embrapa Manual de Métodos de Análises de Solo; Teixeira, P. C., Donagemma, G.K., Fontana, A., Teixeira, W.G., Eds.; 3rd ed.; Brasília, DF: Embrapa: Brasília, DF, 2017; ISBN 9788570357717. [Google Scholar]
- ANA Agência Nacional de Águas e Saneamento Básico: Rede Hidrometeorológica Nacional. Available online: https://www.snirh.gov.br/hidroweb/mapa (accessed on 15 March 2022).
- Courtois, E.A.; Stahl, C.; Van den Berge, J.; Bréchet, L.; Van Langenhove, L.; Richter, A.; Urbina, I.; Soong, J.L.; Peñuelas, J.; Janssens, I.A. Spatial Variation of Soil CO2, CH4 and N2O Fluxes Across Topographical Positions in Tropical Forests of the Guiana Shield. Ecosystems 2018, 21, 1445–1458. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Separating Root and Soil Microbial Contributions to Soil Respiration: A Review Ofmethods and Observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Konda, R.; Ohta, S.; Ishizuka, S.; Heriyanto, J.; Wicaksono, A. Seasonal Changes in the Spatial Structures of N2O, CO2, and CH4 Fluxes from Acacia Mangium Plantation Soils in Indonesia. Soil Biol. Biochem. 2010, 42, 1512–1522. [Google Scholar] [CrossRef]
- Sotta, E.D.; Veldkamp, E.; Guimarães, B.R.; Paixão, R.K.; Ruivo, M.L.P.; Almeida, S.S. Landscape and Climatic Controls on Spatial and Temporal Variation in Soil CO2 Efflux in an Eastern Amazonian Rainforest, Caxiuanã, Brazil. For. Ecol. Manage. 2006, 237, 57–64. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Higuchi, N.; Teixeira, L.M.; Dos Santos, J.; Laurance, S.G.; Trumbore, S.E. Response of Tree Biomass and Wood Litter to Disturbance in a Central Amazon Forest. Oecologia 2004, 141, 596–611. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, S.; Tsuruta, H.; Murdiyarso, D. An Intensive Field Study on CO2, CH4, and N2O Emissions from Soils at Four Land-Use Types in Sumatra, Indonesia. Global Biogeochem. Cycles 2002, 16, 22–1. [Google Scholar] [CrossRef]
- Verchot, L.V.; Brienza, S.; de Oliveira, V.C.; Mutegi, J.K.; Cattânio, J.H.; Davidson, E.A. Fluxes of CH4, CO2, NO, and N2O in an Improved Fallow Agroforestry System in Eastern Amazonia. Agric. Ecosyst. Environ. 2008, 126. [Google Scholar] [CrossRef]
- Werner, C.; Kiese, R.; Butterbach-Bahl, K. Soil-Atmosphere Exchange of N2O, CH4, and CO2 and Controlling Environmental Factors for Tropical Rain Forest Sites in Western Kenya. J. Geophys. Res. Atmos. 2007, 112, 3308. [Google Scholar] [CrossRef]
- Murphy, M.; Balser, T.; Buchmann, N.; Hahn, V.; Potvin, C. Linking Tree Biodiversity to Belowground Process in a Young Tropical Plantation: Impacts on Soil CO2 Flux. For. Ecol. Manage. 2008, 255, 2577–2588. [Google Scholar] [CrossRef]
- Le Mer, J.; Roger, P. Production, Oxidation, Emission and Consumption of Methane by Soils: A Review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Etminan, M.; Myhre, G.; Highwood, E.J.; Shine, K.P. Radiative Forcing of Carbon Dioxide, Methane, and Nitrous Oxide: A Significant Revision of the Methane Radiative Forcing. Geophys. Res. Lett. 2016, 43, 12–614. [Google Scholar] [CrossRef]
- Collins, W.J.; Webber, C.P.; Cox, P.M.; Huntingford, C.; Lowe, J.; Sitch, S.; Chadburn, S.E.; Comyn-Platt, E.; Harper, A.B.; Hayman, G.; et al. Increased Importance of Methane Reduction for a 1.5 Degree Target. Environ. Res. Lett. 2018, 13, 054003. [Google Scholar] [CrossRef]
- Dlugokencky, E.J.; Nisbet, E.G.; Fisher, R.; Lowry, D. Global Atmospheric Methane: Budget, Changes and Dangers. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 2058–2072. [Google Scholar] [CrossRef]
- Dutaur, L.; Verchot, L. V. A Global Inventory of the Soil CH4 Sink. Global Biogeochem. Cycles 2007, 21, GB4013. [Google Scholar] [CrossRef]
- Davidson, E.A.; Nepstad, D.C.; Ishida, F.Y.; Brando, P.M. Effects of an Experimental Drought and Recovery on Soil Emissions of Carbon Dioxide, Methane, Nitrous Oxide, and Nitric Oxide in a Moist Tropical Forest. Glob. Chang. Biol. 2008, 14, 2582–2590. [Google Scholar] [CrossRef]
- Fang, Y.; Gundersen, P.; Zhang, W.; Zhou, G.; Christiansen, J.R.; Mo, J.; Dong, S.; Zhang, T. Soil-Atmosphere Exchange of N2O, CO2 and CH4 along a Slope of an Evergreen Broad-Leaved Forest in Southern China. Plant Soil 2009, 319, 37–48. [Google Scholar] [CrossRef]
- Silver, W.L.; Neff, J.; Mcgroddy, M.; Veldkamp, E.; Keller, M.; Cosme, R. Effects of Soil Texture on Belowground Carbon and Nutrient Storage in a Lowland Amazonian Forest Ecosystem. 2000, 193–209. [CrossRef]
- Bunnell, F.L.; Tait, D.E.N.; Flanagan, P.W.; Van Clever, K. Microbial Respiration and Substrate Weight Loss-I. A General Model of the Influences of Abiotic Variables. Soil Biol. Biochem. 1977, 9, 33–40. [Google Scholar] [CrossRef]
- Ishikura, K.; Hirano, T.; Okimoto, Y.; Hirata, R.; Kiew, F.; Melling, L.; Aeries, E.B.; Lo, K.S.; Musin, K.K.; Waili, J.W.; et al. Soil Carbon Dioxide Emissions Due to Oxidative Peat Decomposition in an Oil Palm Plantation on Tropical Peat. Agric. Ecosyst. Environ. 2018, 254, 202–212. [Google Scholar] [CrossRef]
- Chanda, A.; Akhand, A.; Manna, S.; Dutta, S.; Das, I.; Hazra, S.; Rao, K.H.; Dadhwal, V.K. Measuring Daytime CO2 Fluxes from the Inter-Tidal Mangrove Soils of Indian Sundarbans. Environ. Earth Sci. 2014, 72, 417–427. [Google Scholar] [CrossRef]
- Lessard, R.; Rochette, P.; Topp, E.; Pattey, E.; Desjardins, R.L.; Beaumont, G. Methane and Carbon Dioxide Fluxes from Poorly Drained Adjacent Cultivated and Forest Sites. Can. J. Soil Sci. 1994, 74, 139–146. [Google Scholar] [CrossRef]
- Davidson, E.A.; Verchot, L. V; Cattânio, J.H.; Ackerman, I.L.; Carvalho, J.E.M. Effects of Soil Water Content on Soil Respiration in Forests and Cattle Pastures of Eastern Amazonia; 2000; Vol. 48;
- Segers, R. Methane Production and Methane Consumption--a Review of Processes Underlying Wetland Methane Fluxes [Review]. Biogeochemistry 1998, 41, 23–51. [Google Scholar] [CrossRef]
- Keller, M.; Mitre, M.E.; Stallard, R.F. Consumption of Atmospheric Methane in Soils of Central Panama: Effects of Agricultural Development. Global Biogeochem. Cycles 1990, 4, 21–27. [Google Scholar] [CrossRef]
- Fernandes, S.A.P.; Bernoux, M.; Cerri, C.C.; Feigl, B.J.; Piccolo, M.C. Seasonal Variation of Soil Chemical Properties and CO2 and CH4 Fluxes in Unfertilized and P-Fertilized Pastures in an Ultisol of the Brazilian Amazon. Geoderma 2002, 107, 227–241. [Google Scholar] [CrossRef]
- Steudler, P.A.; Melillo, J.M.; Feigl, B.J.; Neill, C.; Piccolo, M.C.; Cerri, C.C. Consequence of Forest-to-Pasture Conversion on CH4 Fluxes in the Brazilian Amazon Basin. J. Geophys. Res. Atmos. 1996, 101, 18547–18554. [Google Scholar] [CrossRef]
- Sihi, D.; Davidson, E.A.; Savage, K.E.; Liang, D. Simultaneous Numerical Representation of Soil Microsite Production and Consumption of Carbon Dioxide, Methane, and Nitrous Oxide Using Probability Distribution Functions. Glob. Chang. Biol. 2019, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Roslev, P.; King, G.M. Regulation of Methane Oxidation in a Freshwater Wetland by Water Table Changes and Anoxia. FEMS Microbiol. Ecol. 1996, 19, 105–115. [Google Scholar] [CrossRef]
- Fang, X.; Zhao, L.; Zhou, G.; Huang, W.; Liu, J. Increased Litter Input Increases Litter Decomposition and Soil Respiration but Has Minor Effects on Soil Organic Carbon in Subtropical Forests. Plant Soil 2015, 392, 139–153. [Google Scholar] [CrossRef]
- Fanin, N.; Hättenschwiler, S.; Barantal, S.; Schimann, H.; Fromin, N. Does Variability in Litter Quality Determine Soil Microbial Respiration in an Amazonian Rainforest? Soil Biol. Biochem. 2011, 43, 1014–1022. [Google Scholar] [CrossRef]
- RoyChowdhury, T.; Bramer, L.; Hoyt, D.W.; Kim, Y.-M.; Metz, T.O.; McCue, L.A.; Diefenderfer, H.L.; Jansson, J.K.; Bailey, V. Temporal Dynamics of CO2 and CH4 Loss Potentials in Response to Rapid Hydrological Shifts in Tidal Freshwater Wetland Soils. Ecol. Eng. 2018, 114, 104–114. [Google Scholar] [CrossRef]
- Kiese, R.; Hewett, B.; Graham, A.; Butterbach-Bahl, K.; Kiese, C. :; Hewett, B.; Graham, A.; Butterbach-Bahl, K. Seasonal Variability of N2O Emissions and CH4 Uptake by Tropical Rainforest Soils of Queensland, Australia. Global Biogeochem. Cycles 2003, 17, 1043. [Google Scholar] [CrossRef]
- Werner, C.; Zheng, X.; Tang, J.; Xie, B.; Liu, C.; Kiese, R.; Butterbach-Bahl, K. N2O, CH4 and CO2 Emissions from Seasonal Tropical Rainforests and a Rubber Plantation in Southwest China. Plant Soil 2006, 289, 335–353. [Google Scholar] [CrossRef]
- Conrad, R. The Global Methane Cycle: Recent Advances in Understanding the Microbial Processes Involved. Environ. Microbiol. Rep. 2009, 1, 285–292. [Google Scholar] [CrossRef]
- Whalen, S.C. Biogeochemistry of Methane Exchange between Natural Wetlands and the Atmosphere. Environ. Eng. Sci. 2005, 22, 73–94. [Google Scholar] [CrossRef]
- Turetsky, M.R.; Kotowska, A.; Bubier, J.; Dise, N.B.; Crill, P.; Hornibrook, E.R.C.; Minkkinen, K.; Moore, T.R.; Myers-Smith, I.H.; Nykänen, H.; et al. A Synthesis of Methane Emissions from 71 Northern, Temperate, and Subtropical Wetlands. Glob. Chang. Biol. 2014, 20, 2183–2197. [Google Scholar] [CrossRef]
- Waldo, N.B.; Hunt, B.K.; Fadely, E.C.; Moran, J.J.; Neumann, R.B. Plant Root Exudates Increase Methane Emissions through Direct and Indirect Pathways. Biogeochem. 2019 1451 2019, 145, 213–234. [Google Scholar] [CrossRef]
- Bridgham, S.D.; Cadillo-Quiroz, H.; Keller, J.K.; Zhuang, Q. Methane Emissions from Wetlands: Biogeochemical, Microbial, and Modeling Perspectives from Local to Global Scales. Glob. Chang. Biol. 2013, 19, 1325–1346. [Google Scholar] [CrossRef]
Figure 1.
Location of the study areas: estuarine floodplain area (AV), in the municipality of Belém (B and C), and upland area (ATF), in the municipality of Santa Maria do Pará (B and D.
Figure 1.
Location of the study areas: estuarine floodplain area (AV), in the municipality of Belém (B and C), and upland area (ATF), in the municipality of Santa Maria do Pará (B and D.
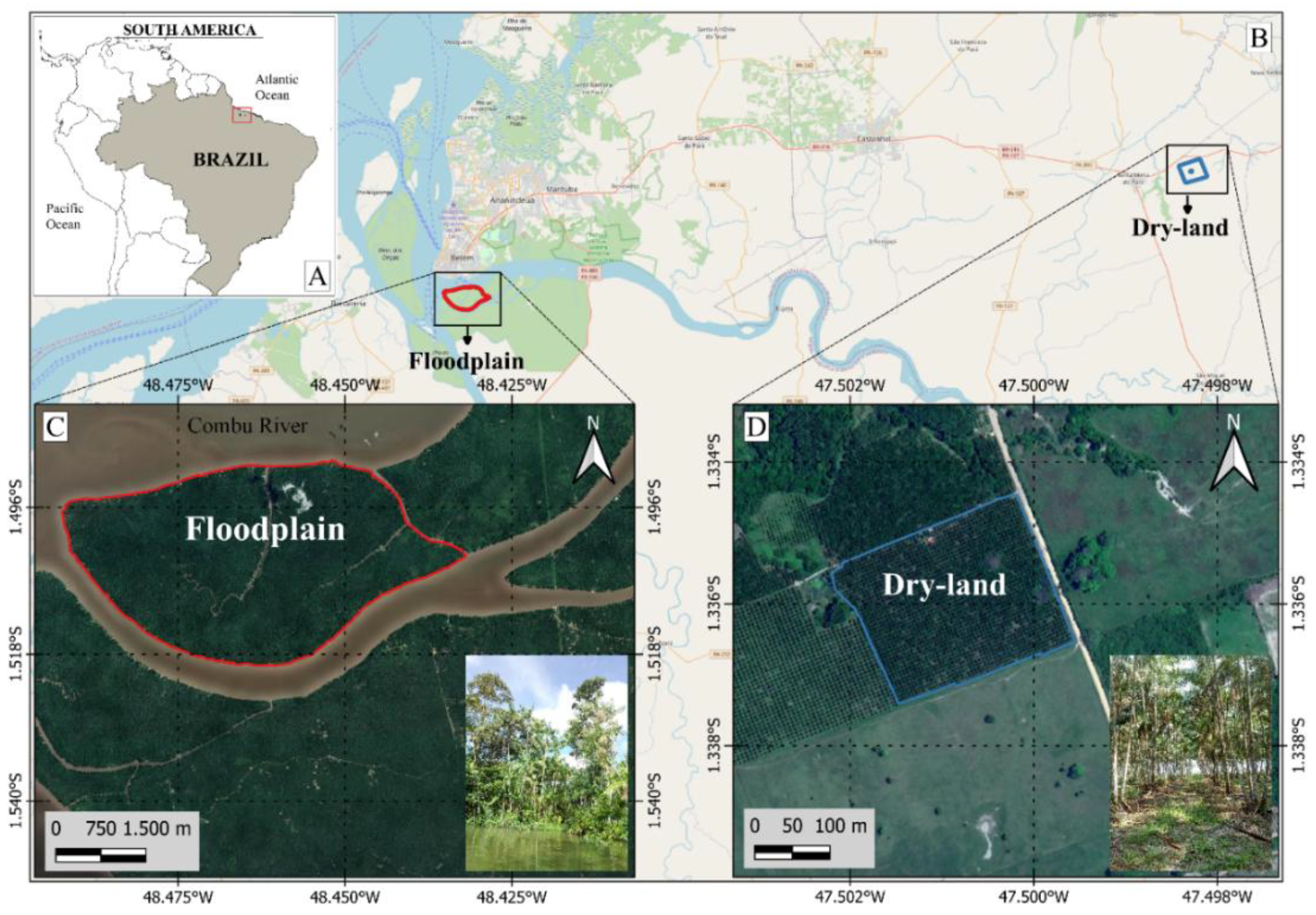
Figure 2.
Experimental design for flow analysis in the area: (a) açaí plantation on upland (ATF), with flow chambers (blue dots) in the planting lines (L) around the clumps, and on the street (R) where the machinery passes, in Santa Maria do Pará, and (b) in the floodplain area (AV), in the estuary in Belém, with the flow chambers randomly distributed in a circle.
Figure 2.
Experimental design for flow analysis in the area: (a) açaí plantation on upland (ATF), with flow chambers (blue dots) in the planting lines (L) around the clumps, and on the street (R) where the machinery passes, in Santa Maria do Pará, and (b) in the floodplain area (AV), in the estuary in Belém, with the flow chambers randomly distributed in a circle.
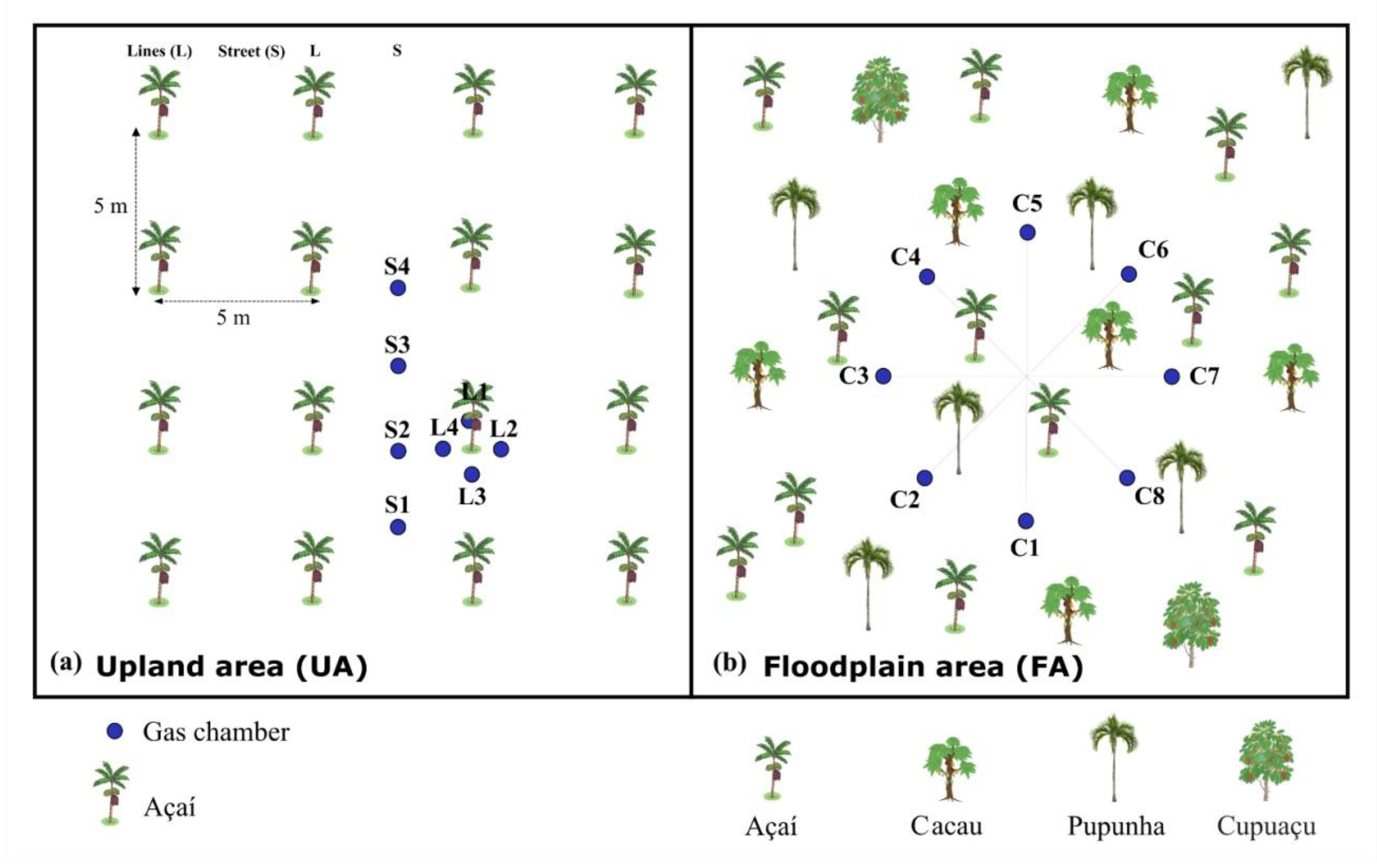
Figure 3.
Figure 3. A) CO2 flux (FCO2; g CO2 m-2 d-1) in high topography (Top1) and low topography (Top2) in the dry period; B) CH4 flux (FCH4; mg CH4 m-2 d-1) in Top1 and Top2 in the dry period; C) FCO2 in Top1 comparing lines (L) with streets (R) in the dry period; D) FCH4 in Top1 comparing L with R in the dry period; E) FCO2 in Top2 comparing L with R; F) FCH4 in Top2 comparing L with R in the dry period, in an area of homogeneous açaí plantation on dry land, in the municipality of Santa Maria do Pará (Brazil).
Figure 3.
Figure 3. A) CO2 flux (FCO2; g CO2 m-2 d-1) in high topography (Top1) and low topography (Top2) in the dry period; B) CH4 flux (FCH4; mg CH4 m-2 d-1) in Top1 and Top2 in the dry period; C) FCO2 in Top1 comparing lines (L) with streets (R) in the dry period; D) FCH4 in Top1 comparing L with R in the dry period; E) FCO2 in Top2 comparing L with R; F) FCH4 in Top2 comparing L with R in the dry period, in an area of homogeneous açaí plantation on dry land, in the municipality of Santa Maria do Pará (Brazil).
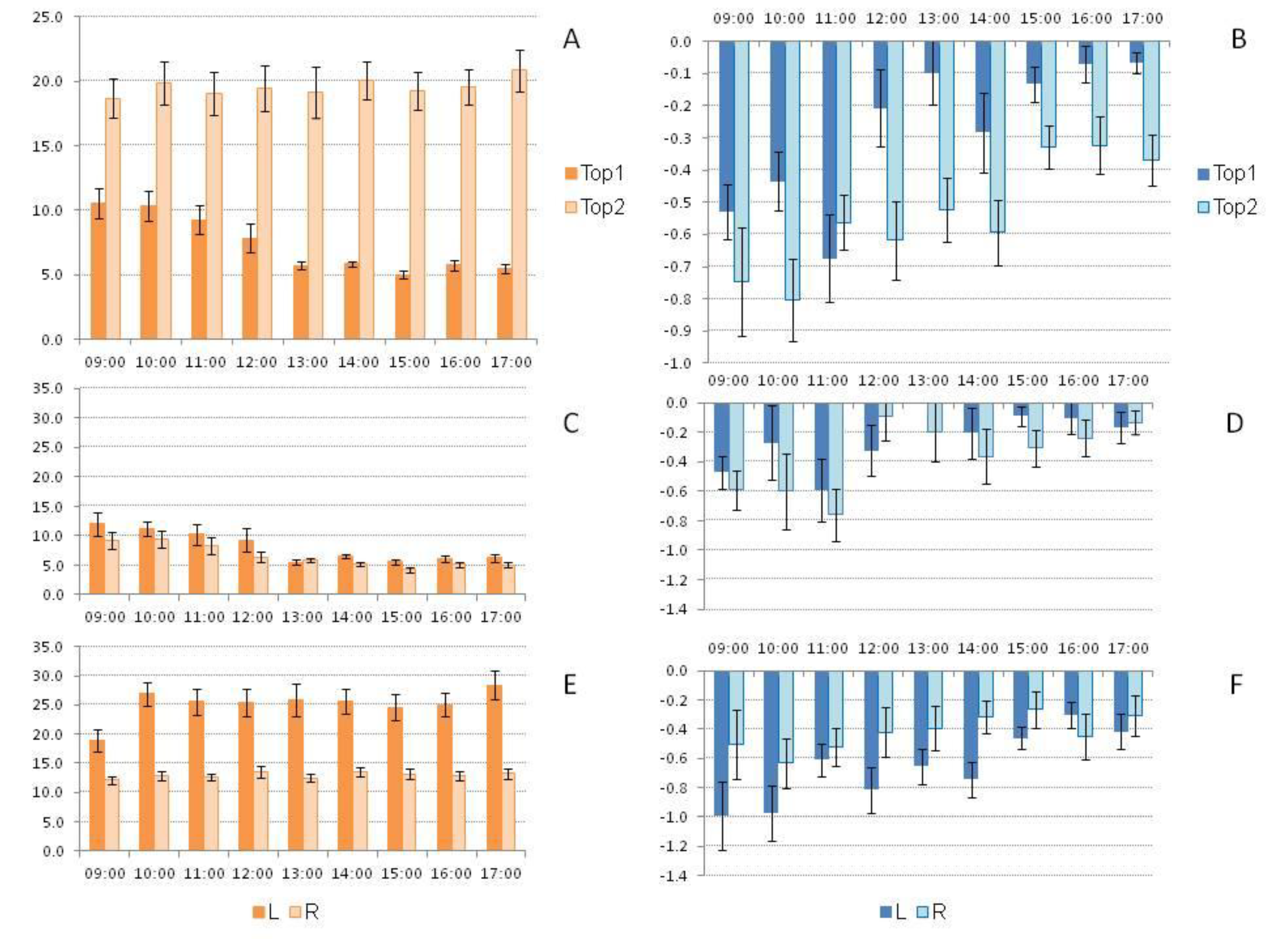
Figure 4.
A) CO2 flux (FCO2; g CO2 m-2 d-1) in homogeneous açaí plantation in upland (ATF) and in managed floodplain forest (VA) in the rainy season; B) CH4 flux (FCH4; mg CH4 m-2 d-1) in homogeneous açaí plantation in upland (ATF) and in managed floodplain (VA) area, in the rainy season.
Figure 4.
A) CO2 flux (FCO2; g CO2 m-2 d-1) in homogeneous açaí plantation in upland (ATF) and in managed floodplain forest (VA) in the rainy season; B) CH4 flux (FCH4; mg CH4 m-2 d-1) in homogeneous açaí plantation in upland (ATF) and in managed floodplain (VA) area, in the rainy season.
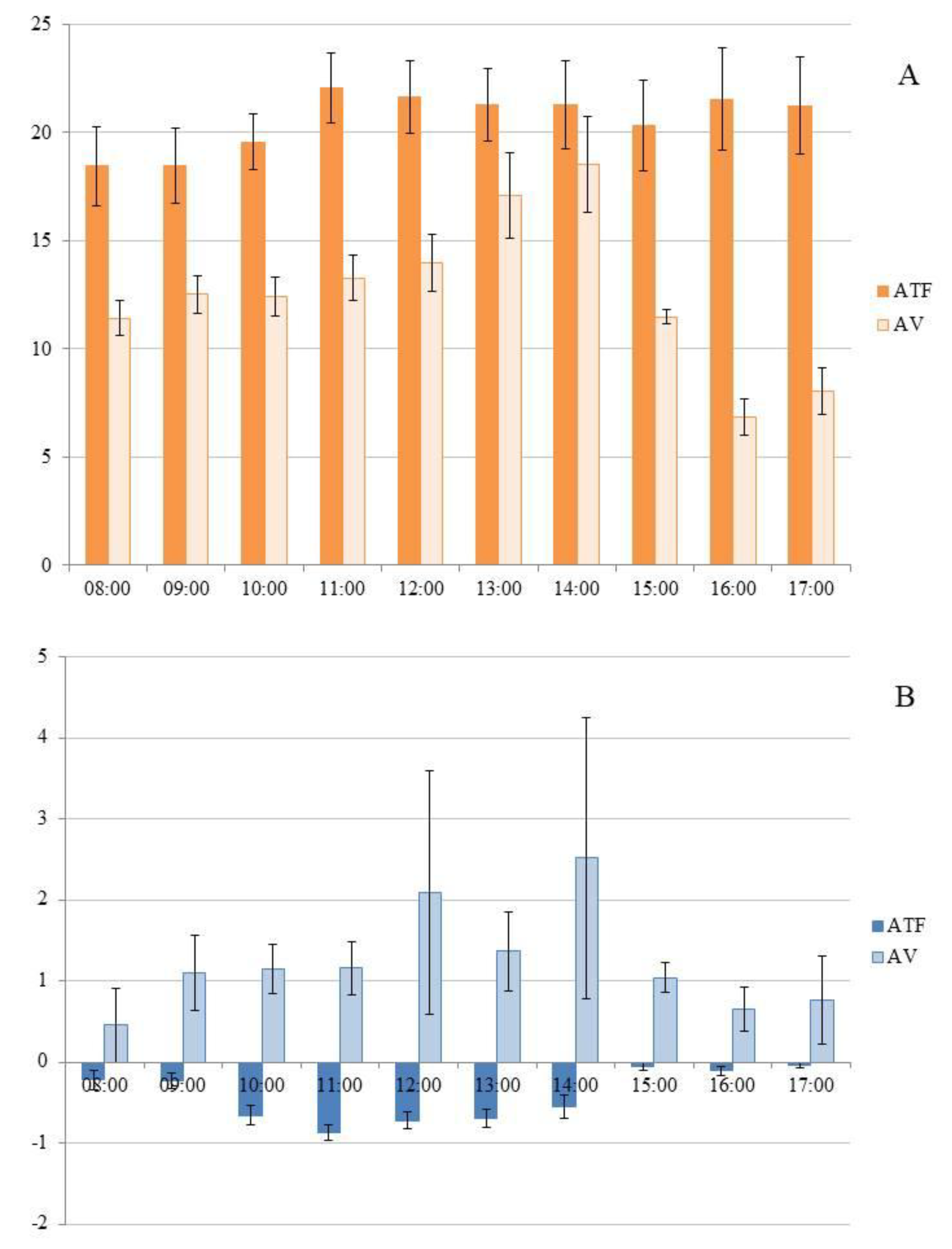
Figure 5.
A) CO2 efflux (FCO2) in the upland area (ATF) in the plateau (Top1), shoal (Top2), and average (TopA) topography during the months sampled in the wet and dry season. B) CH4 influx (FCH4) in the upland area (ATF) in the plateau (Top1), shoal (Top2), and average (TopA) topography, during the months sampled in the wet and dry season. The bars represent the standard error of the mean.
Figure 5.
A) CO2 efflux (FCO2) in the upland area (ATF) in the plateau (Top1), shoal (Top2), and average (TopA) topography during the months sampled in the wet and dry season. B) CH4 influx (FCH4) in the upland area (ATF) in the plateau (Top1), shoal (Top2), and average (TopA) topography, during the months sampled in the wet and dry season. The bars represent the standard error of the mean.
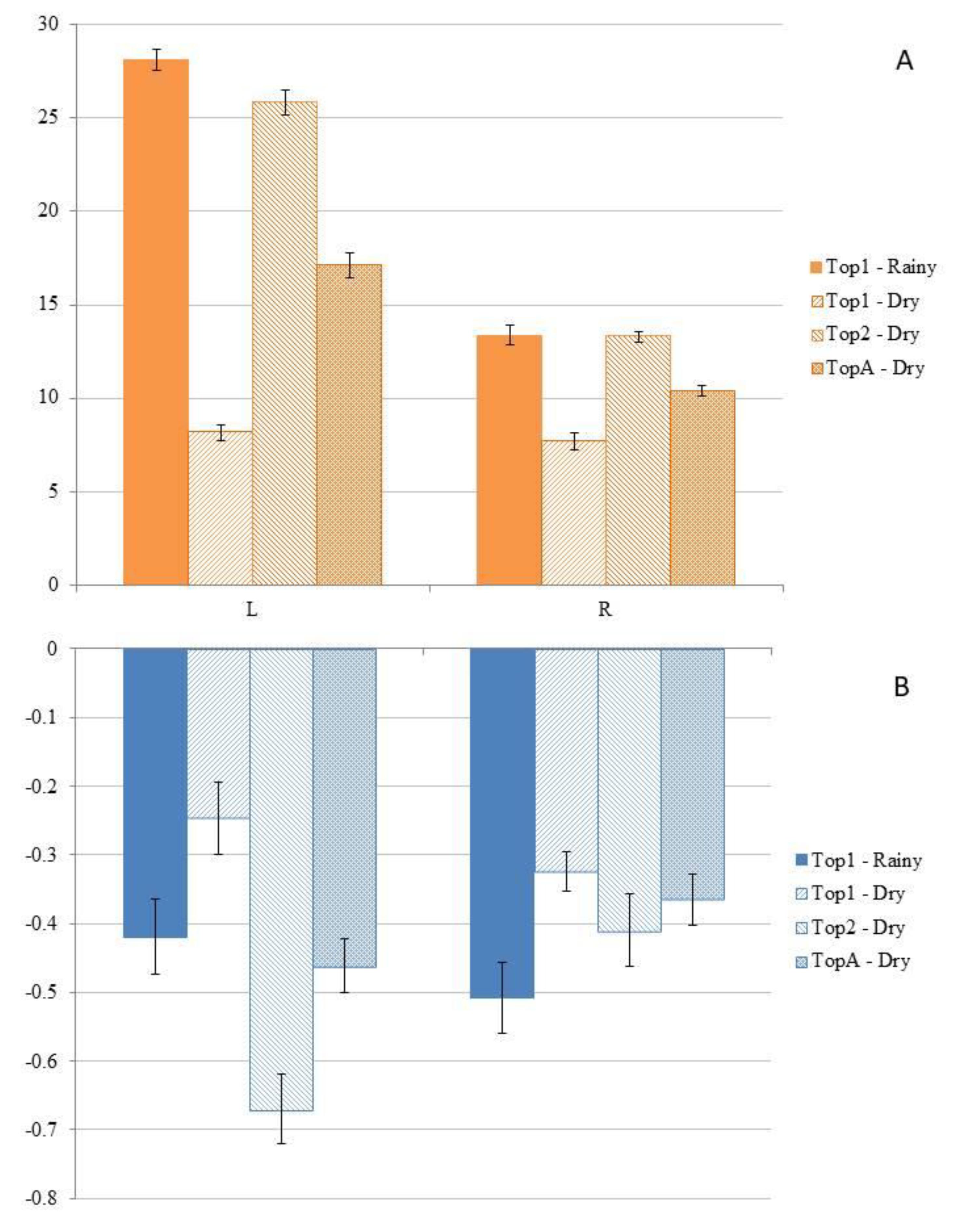
Figure 6.
A) CO2 flux (FCO2; g CO2 m-2 d-1) in the rainy season (March) in homogeneous açaí plantation in upland (ATF) and in managed floodplain (AV), in high topography (Top1); B) CH4 flux (FCH4; mg CH4 m-2 d-1) in the rainy season (March) in homogeneous açaí plantation in Terra Firme (ATF) and in managed Várzea forest (AV, in high topography (Top1); C) CO2 flux (FCO2; g CO2 m-2 d-1) in the dry season (September) in homogeneous açaí plantation in upland (ATF) in high topography (Plateau; Top1) and low topography (Lowlands; Top2), and in managed floodplain (VA), in high topography (Top1); D) CH4 flux (FCH4; mg CH4 m-2 d-1) in the dry season (September) in homogeneous açaí plantation in upland (ATF) in high topography (Top1) and low topography (Top2), and in managed floodplain (AV) in high topography (Top1).
Figure 6.
A) CO2 flux (FCO2; g CO2 m-2 d-1) in the rainy season (March) in homogeneous açaí plantation in upland (ATF) and in managed floodplain (AV), in high topography (Top1); B) CH4 flux (FCH4; mg CH4 m-2 d-1) in the rainy season (March) in homogeneous açaí plantation in Terra Firme (ATF) and in managed Várzea forest (AV, in high topography (Top1); C) CO2 flux (FCO2; g CO2 m-2 d-1) in the dry season (September) in homogeneous açaí plantation in upland (ATF) in high topography (Plateau; Top1) and low topography (Lowlands; Top2), and in managed floodplain (VA), in high topography (Top1); D) CH4 flux (FCH4; mg CH4 m-2 d-1) in the dry season (September) in homogeneous açaí plantation in upland (ATF) in high topography (Top1) and low topography (Top2), and in managed floodplain (AV) in high topography (Top1).
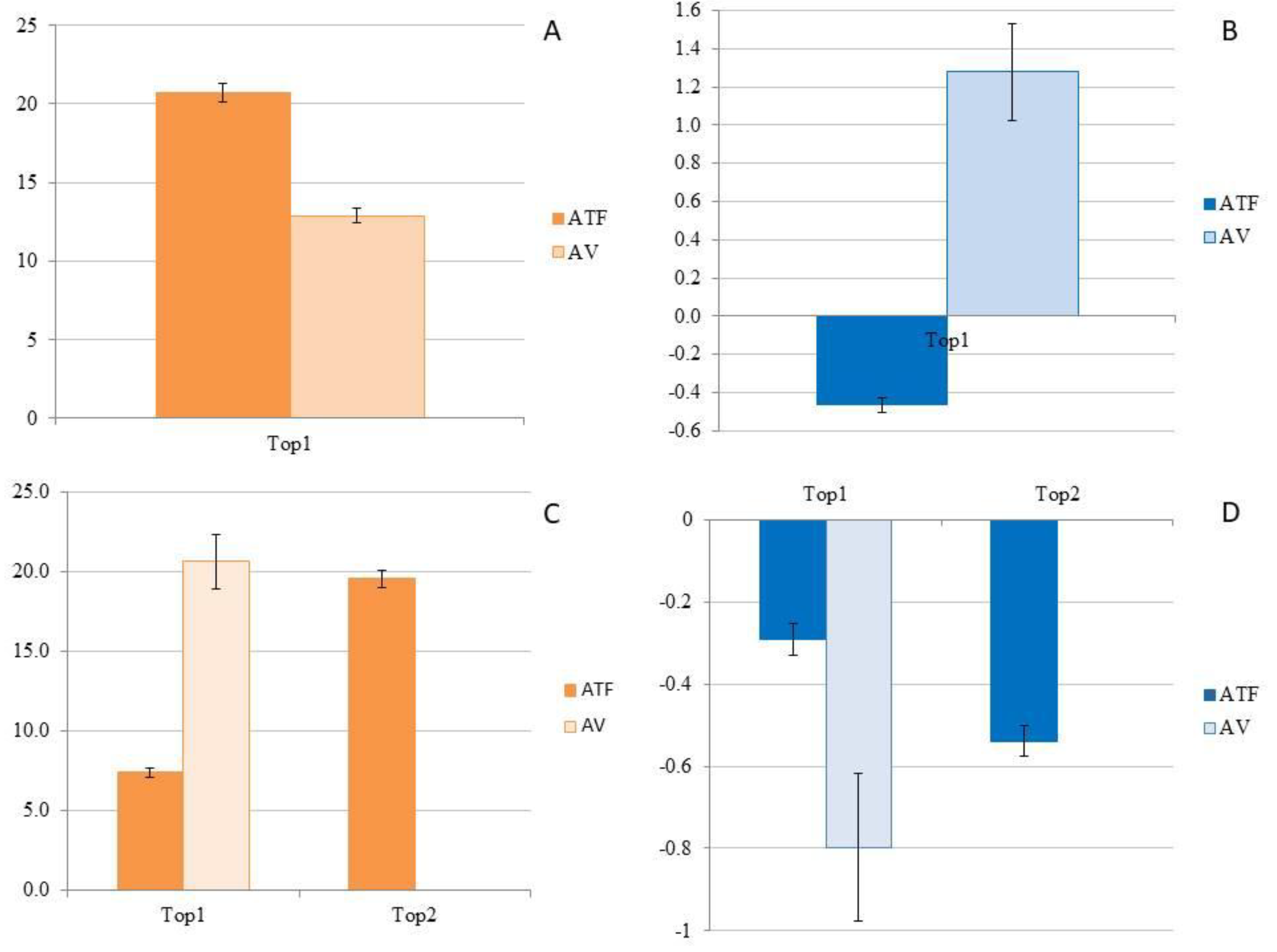
Figure 7.
A) Precipitation (mm) in the dry land area (ATF) in Santa Maria do Pará, compared to the floodplain area (AV) in Belém, and the difference in precipitation between the two locations (ATF-AV), from 2020 to 2021; B) Climatology (mm) in the dry land area (Climatology ATF) in Santa Maria do Pará in the floodplain area (Climatology AV) in Belém, and the difference between precipitation (mm) during the study period and the Climatology in ATF (ATF - Clim_ATF) and the Climatology in AV (AV - Clim_AV).
Figure 7.
A) Precipitation (mm) in the dry land area (ATF) in Santa Maria do Pará, compared to the floodplain area (AV) in Belém, and the difference in precipitation between the two locations (ATF-AV), from 2020 to 2021; B) Climatology (mm) in the dry land area (Climatology ATF) in Santa Maria do Pará in the floodplain area (Climatology AV) in Belém, and the difference between precipitation (mm) during the study period and the Climatology in ATF (ATF - Clim_ATF) and the Climatology in AV (AV - Clim_AV).
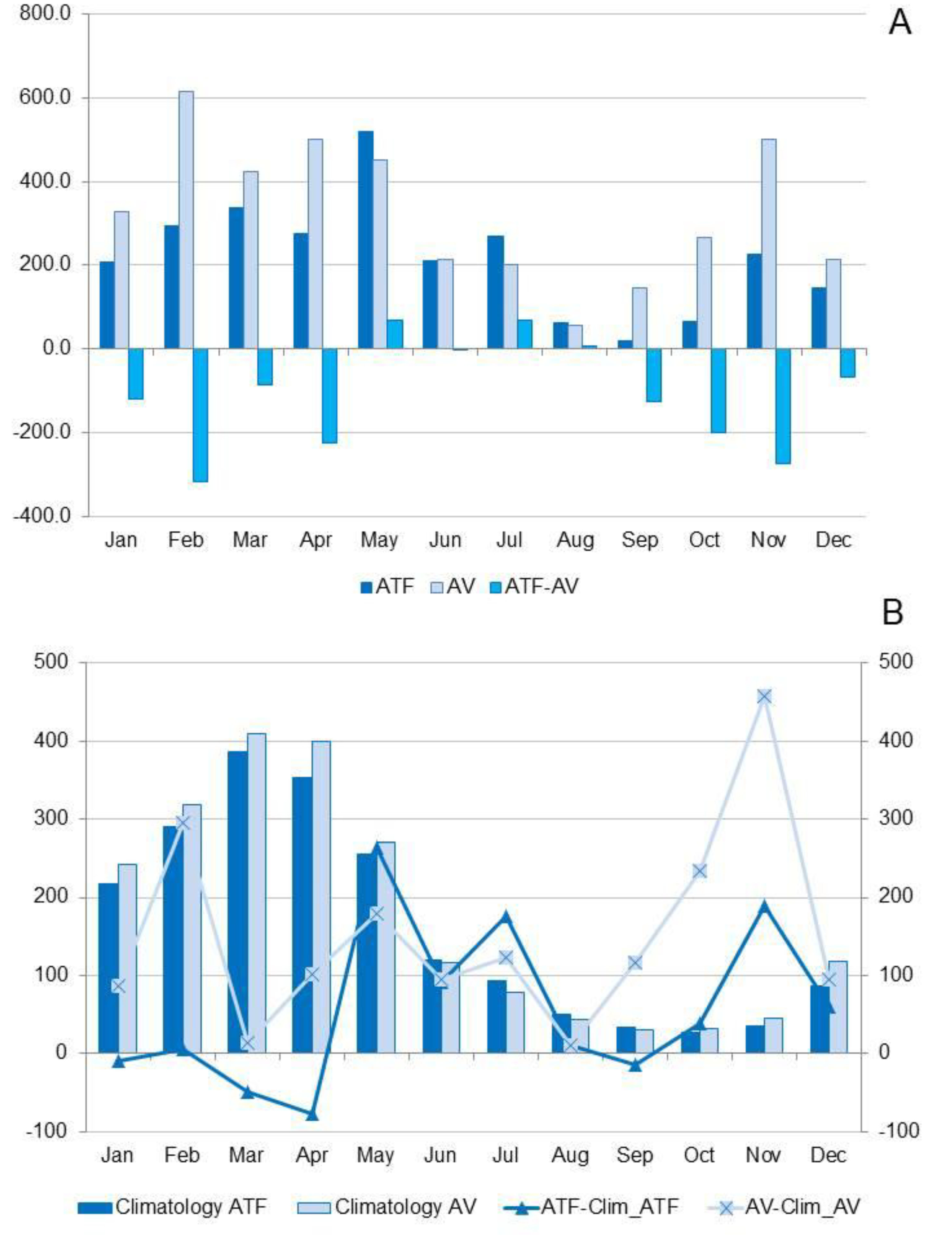
Figure 8.
Behavior of Temperature (Ta, °C) and Relative Humidity (RH, %) of the air, and Soil Temperature (Ts, °C) in: A) dry land area (ATF) in the rainy season; B) floodplain area (AV) in the rainy season; C) ATF in the dry season; D) AV in the dry season. Times that have no value were due to the impossibility of collecting the data.
Figure 8.
Behavior of Temperature (Ta, °C) and Relative Humidity (RH, %) of the air, and Soil Temperature (Ts, °C) in: A) dry land area (ATF) in the rainy season; B) floodplain area (AV) in the rainy season; C) ATF in the dry season; D) AV in the dry season. Times that have no value were due to the impossibility of collecting the data.
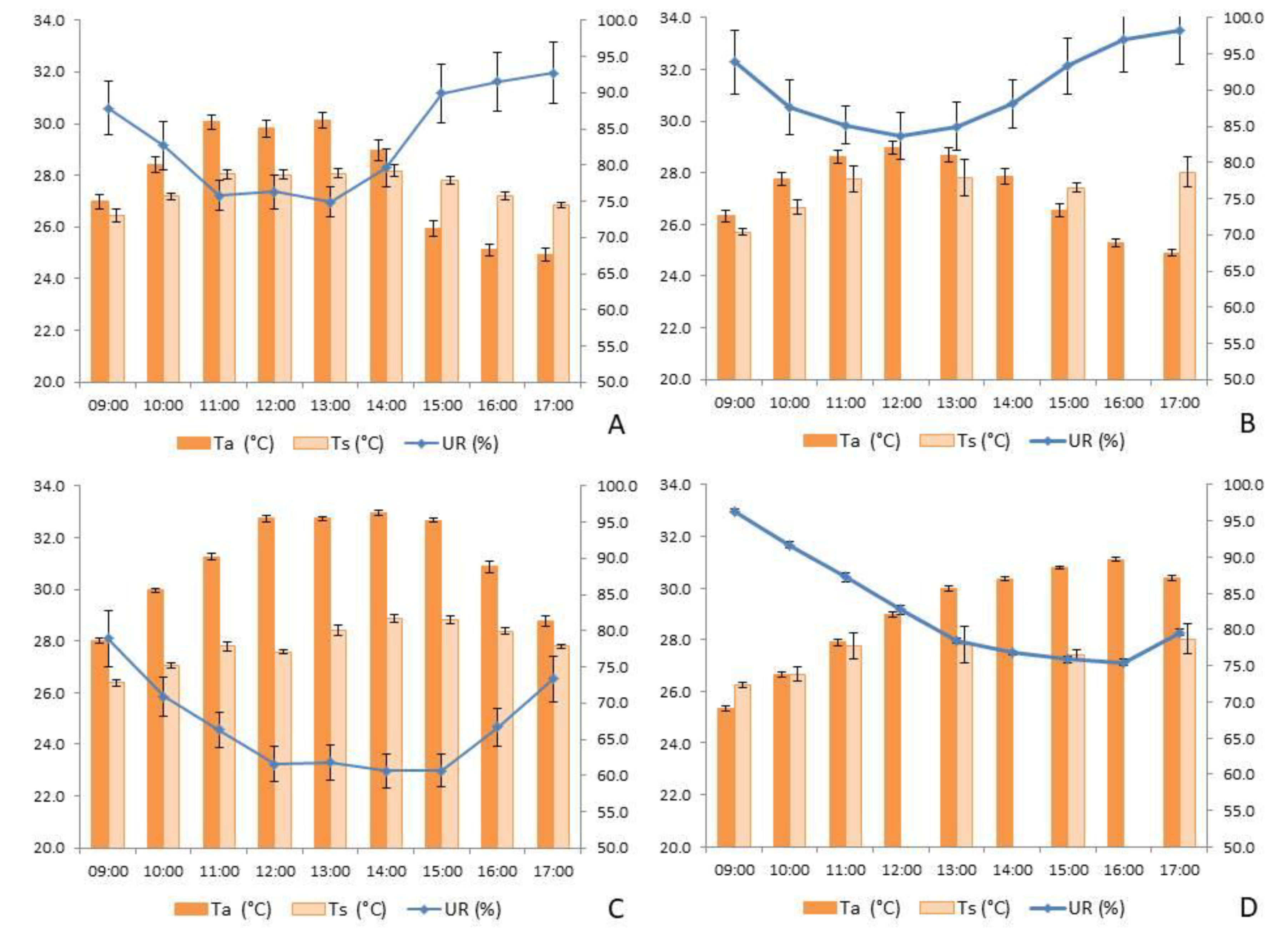

Table 1.
Seasonality of some environmental data in upland (ATV) and floodplain (AV) treatments in the eastern Amazon region. Numbers report the mean ± standard error, with lowercase letters comparing the seasonality in each treatment, and uppercase letters comparing treatments in the same seasonality. ND means that there was no analysis.
Table 1.
Seasonality of some environmental data in upland (ATV) and floodplain (AV) treatments in the eastern Amazon region. Numbers report the mean ± standard error, with lowercase letters comparing the seasonality in each treatment, and uppercase letters comparing treatments in the same seasonality. ND means that there was no analysis.
| Upland | Floodplain | |||
| Dry | Rainy | Dry | Rainy | |
| Soil moisture (Us, %) | 8.64 ± 0.51bB | 23.12 ± 0.77aB | 26.44 ± 0.70bA | 53.27 ± 1.12aA |
| pH | 4.83 ± 0.21aA | 5.16 ± 0.05aA | 4.15 ± 0.01aB | 4.15 ± 0.04aB |
| Fine roots (Mg ha-1) | 2.61 ± 0.28aA | 2.96 ± 0.36aA | 4.64 ± 0.88aA | 2.25 ± 0.42aA |
| Thick roots (Mg ha-1) | 11.38 ± 1.17aA | 8.87 ± 0.77bA | 5.91 ± 0.91aA | 3.39 ± 0.60bB |
| Total Roots (Mg ha-1) | 13.59 ± 1.38aA | 11.84 ± 1.04aA | 6.57 ± 0.40aB | 5.64 ± 0.89aB |
| Microbial carbon (g kg-1) | 0.42 ± 0.03B | ND | 1.56 ± 0.10A | ND |
| Microbial nitrogen (mg kg-1) | 6.12 ± 0.74B | ND | 46.28 ± 2.12A | ND |
Table 2.
Correlation coefficient between CO2 (FCO2) and CH4 (FCH4) fluxes with soil temperature (Ts), soil moisture (Us), fine root biomass (RF), coarse root biomass (RG), total root biomass (TR), air temperature (Ta), relative humidity (UR), atmospheric pressure (Pa), microbial carbon (Cm), microbial nitrogen (Nm), pH in açaí monoculture grown on dry land (ATF) and açaí agroforestry in the estuary floodplain area (AV), in the rainy and dry seasons.
Table 2.
Correlation coefficient between CO2 (FCO2) and CH4 (FCH4) fluxes with soil temperature (Ts), soil moisture (Us), fine root biomass (RF), coarse root biomass (RG), total root biomass (TR), air temperature (Ta), relative humidity (UR), atmospheric pressure (Pa), microbial carbon (Cm), microbial nitrogen (Nm), pH in açaí monoculture grown on dry land (ATF) and açaí agroforestry in the estuary floodplain area (AV), in the rainy and dry seasons.
| Rainy Season | |||||||||||||
| ATF | FCH4 | Ts | Us | RF | RG | TR | Ta | UR | Pa | Cm | Nm | pH | |
| FCO2 | -0,111 | 0,250** | 0,287** | -0,174** | 0,293** | 0.156** | 0,259** | -0,245** | -0,156** | ND | ND | -0,743** | |
| FCH4 | 1,000 | -0,453** | 0,137* | -0,088 | 0,028 | -0.009 | -0,493** | 0,500** | 0,142* | ND | ND | -0,067 | |
| AV | |||||||||||||
| FCO2 | 0,163** | 0,448** | -0,135* | 0,209** | 0,295** | 0,297** | 0,553** | -0,562** | -0,153* | ND | ND | -0,173** | |
| FCH4 | 1,000 | -0,043 | -0,081 | -0,078 | 0,025 | -0,019 | 0,079 | -0,099 | 0,017 | ND | ND | -0,007 | |
| Dry Season | |||||||||||||
| ATF | FCH4 | Ts | Us | RF | RG | TR | Ta | UR | Pa | Cm | Nm | pH | |
| FCO2 | -0,316** | -0,151** | 0,626** | 0,570** | -0,207** | -0,009 | -0,053 | 0,062 | 0,056 | 0.441** | 0,001 | 0,419** | |
| FCH4 | 1,000 | 0,173** | -0,173** | -0,100** | 0,204** | 0,158** | 0,057 | -0,078 | -0,215** | -0,116** | 0,116* | -0,166** | |
| AV | |||||||||||||
| FCO2 | -0,053 | 0,197 | 0,209 | 0,207 | 0,257 | 0,245 | 0,406* | -0,461* | -0,370 | 0,112 | -0,073 | 0,248 | |
| FCH4 | 1,000 | -0,307 | -0,209 | -0,119 | -0,200 | -0,169 | 0,121 | -0,241 | -0,148 | -0,019 | -0,019 | -0,175 | |
* Significant p < 0.05; ** significant p < 0.01; ND not determined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



