
Submitted:
07 May 2025
Posted:
07 May 2025
You are already at the latest version
Abstract
Background: Envenomation by poisonous creatures is a major global cause of mortality. Its concomitant impact on the postmortem corpse decomposition and associated insect succession pattern is yet poorly understood. Purpose of the study: This study comparatively investigates the impact of envenomization with the snake, Walterinnesia aegyptia L., versus the scorpion, Androctonus crassicauda L., venoms on rabbit corpse decomposition and beetle succession. Methods: Three groups of rabbits (5 animals each) were injected with the snake venom, the scorpion venom, or 0.9% saline (control) prior to euthanasia with CO2. Corpse decomposition stages and beetle succession were monitored over 11 days. Results: Four stages of decomposition, with venom-dependent duration variation, were observed. The scorpion-envenomized corpses showed a longer decay stage and a delayed dry stage. A total of 1094 beetles belonging to 27 species of 14 families were reported. Histeridae, Dermestidae, Scarabaeidae, and Tenebrionidae were the most diversified and prevalent families. Chrysomelidae, Elateridae, Hybosoridae, and Ptinidae were distinctively attracted to control corpses, while Nitidulidae and Zopheridae were only found on control and snake-envenomized ones. Four species belonging to the families Anthicidae, Histeridae, Scarabaeidae, and Tenebrionidae were predominant on all corpses. While four species belonging to the families Chrysomelidae, Curculionidae, Elateridae, and Hybosoridae were distinctively associated with the control corpses. Conclusion: These findings provided evidence that envenomation impacted the decomposition process and beetle succession in a venom-dependent manner, which could be significant for forensic investigations.
Keywords:
beetles
; corpse
; decomposition
; envenomization
; forensic
; snake
; scorpion
; venoms
1. Introduction
Forensic Entomology is a rapidly growing field of study that involves the use of insects to aid in criminal investigations. In this field, necrophagous insects and other arthropods are used as tools of forensic investigation to detect, elucidate, and establish evidence during forensic investigation [1]. This is due to the capability of these insects to reach a corpse within minutes post-death [2]. The diversity of these corpse-seeking insects, their successional behavior towards the corpse, the relationship between their arrival time and the developmental process of their immature stages, and their feeding manner on the corpse all provide valuable data to forensic entomologists that help them solve crime secrets [3,4,5] and estimate the post-mortem interval (PMI) [6].
There are four groups of corpse-seeking insects, which are categorized based on their feeding manner [7]. The first group is the sarcosaprophages, which include dipteran flies from Calliphoridae, Muscidae, and Sarcophagidae, as well as coleopteran beetles from the Dermestidae, which feed on decomposing corpses. The second group is the coprophages, which include beetles from Scarabaeidae and flies from Muscidae, which are attracted to the herbivores’ rumen contents. The third group is the dermatophages, which include beetles from Dermestidae, which feed on the cadaver remnants such as bones, hair, and dried skin. The fourth group is the predaceous insects, such as Staphylinid and Histerid beetles and Formicid ants, which feed mainly on corpse-colonizers like dipteran larvae. These insects colonize the corpse along with bacteria, where they work to decompose the corpse [8]. During decomposition, the corpse goes through five stages (fresh, bloating, active decay, advanced decay, and remain), where each stage is characterized by a specific attraction of insects [7].
Necrophagous beetles constitute a forensically determinant factor due to their ability to inhabit the majority of the corpse environment, offering valuable data for forensic investigation [9]. They belong to Coleoptera, the largest Order comprising over one-third of all known insect species [10]. These corpse-associated insects provide valuable data that help in estimating PMI of dried corpses, as well as evaluating the damage and variations that may have occurred to the corpse status [7]. The most forensically important beetle families include Cleridae, Dermestidae, Histeridae, Staphylinidae, and Scarabaeidae [7]. Nevertheless, much less research on corpse-seeking insects has given attention to these beetles compared to flies [11,12,13,14]. Therefore, the present study was implemented to participate in compensating for this deficiency.
Envenomation-related death cases constitute a vital and considerable aspect from the forensic point of view. This is based on the fact that venomous snakes and scorpions are distributed all over the world [15,16,17,18,19,20,21,22,23,24] and affect millions of people [24,25,26,27,28,29,30,31,32,33] and, consequently, envenomation is considered a significant cause of death worldwide [16,24,34,35,36,37]. In this regard, the majority of envenomation-related deaths are caused by arthropods [36,38], followed by snakes and spiders [24,26,35]. In regions like Saudi Arabia, where snakebites [33,39,40] and scorpion stings [39,41,42,43] are prevalent, understanding the impact of envenomization on decomposition is crucial for forensic science. The desert black snake, Walterinnesia aegyptia L., and the fat-tail scorpion, Androctonus crassicauda L., are significant contributors to envenomation-related deaths, yet there is a lack of research on how their venoms affect the decomposition process and beetle succession. Thus, this study aims to bridge this gap by comparing the effects of W. aegyptia and A. crassicauda venoms on rabbit corpses, focusing on the decomposition stages and beetle succession patterns. By addressing this knowledge gap, this research seeks to enhance the PMI estimation and provide valuable insights into differentiating between causes of death involving venomous animals, ultimately contributing to the advancement of forensic science in regions with high envenomation rates.
2. Materials and Methods
2.1. Meteorological Parameters
Experiments of this study ran for 11 days in the summer (from 6th to 17th of June, 2023). The atmospheric parameters were determined over the study period via the Saudi National Center for Meteorology [44]. In addition, the on-site atmospheric parameters were also reported manually at the experimental site in a daily manner (at midday) during the study period following [45,46]. The relative humidity (RH) and ambient temperature (°C) were measured using a hygrometer and digital thermometer devices (Elitch, China) following their instruction manuals. The Skywatch Wind Meter device (Skywatch®, Switzerland) was used for monitoring wind speed following its instruction manual.
2.2. Experimental Site
Experiments were carried out in the botanical garden of the Botany and Microbiology Department, College of Science, King Saud University, Riyadh, Saudi Arabia (N 24°43.174’, E 46°36.954’) (Figure 1A). This garden covers an area of about 10,000 m² with a clayey sandy soil with abundant grasses and herbs. It houses a significant number of plant species, ranging from wild, medicinal, and endangered plants to economically important plants [47].
2.3. Experimental Animals
2.3.1. Rabbits and Mice
Swiss albino mice (≈ 18-20 g each) were obtained from the Animal House, Zoology Department, College of Science, King Saud University for carrying out the lethality of snake and scorpion venoms following [48]. Male domestic rabbits, Oryctolagus cuniculus domesticus (Linnaeus, 1758) (≈ 2.8-3.0 kg each) were purchased at a specialized local farmer’s market in Riyadh City, Saudi Arabia. They were used as experimental models for conducting the forensic experiments in this study according to [49] and following other studies [46,50,51,52,53]. Prior to use for experiments, rabbits were first kept for acclimatization for 2 days in the animal house. They were housed in standard rearing conditions with suitable feed and aeration by skillful specialists in accordance with the Research Ethical Committee at King Saud University (approval code: KSU-SE-23-83).
2.3.2. Snakes and Scorpions
The Saudi desert black cobra snake, W. aegyptia Lataste, 1887 (Squamata, Elapidae) [18,54,55,56] (Figure 1B; from [57]) and the fat-tail scorpion, A. crassicauda (Olivier, 1807) (Scorpiones, Buthidae) [21,58,59,60] (Figure 1 C; from [61]) were used from the Animal House, Zoology Department, College of Science, King Saud University for carrying out this study. Snakes and scorpions are being housed in standard laboratory conditions in the animal house with feed and aeration, according to [62] and [63], respectively. Dealing with snakes and scorpions took place by a qualified specialist from the Herpetology laboratory in Zoology Department and in accordance with the Research Ethical Committee at King Saud University (approval code: KSU-SE-23-83).
2.4. Collection of Venoms
The snake crude venom was milked from five adult snakes, according to [62,64]. While scorpion crude venom was collected from 100 scorpions by inducing electric shock according to [65] and as detailed in [63]. Collected samples of each venom were pooled, lyophilized, and stored at -20°C until used. Prior to experimental use, an aliquot from each venom was freshly prepared in 0.9% saline (pH 7.2), according to [66] in a final concentration of 1.0 mg/ml (w/v). Carrying out venom collection took place by a qualified specialist from the Herpetology Laboratory in Zoology Department and in accordance with the Research Ethical Committee at King Saud University (approval code: KSU-SE-23-83).
2.5. Lethality Assay
The lethality, in terms of both LD50 and LD95, for each venom was determined following the World Health Organization guidelines [67] and as per detailed in our previously published study [48]. Briefly, a preliminary dose-finding experiment was first carried out using Swiss albino mice. The preliminary screening was carried out using a wide range of 8 descending dilutions from each venom aliquot (1,000, 500, 250, 125, 62, 31, 15, and 7 µg/ml). Then, an amount of 0.2 ml from each concentration was subcutaneously injected into each of eight mice (one mouse for each dilution). Mortalities were then recorded at 24 hours post-envenomization. The range of each venom concentration was then narrowed to the required concentration for subsequently conducting the main full lethality assays.
Based on the preliminary screening of each venom lethality, the 4 actively effective concentrations of each venom (data not shown) that caused mice mortality ranging from 0.0 to 100% were used to carry out a full lethality test to determine the LD50 and LD95. In this experiment, 5 groups (one group for each concentration) of mice (5 mice/group; n = 5) were subcutaneously injected with the venom. Mortalities were recorded at 24h post-envenomization, and both LD50 and LD95 were calculated by the Probit analysis according to [68] and following [69].
2.6. Envenomization of Rabbits
Upon determining the LD95 of each venom in mice, it was converted into the equivalent doses for rabbits according to [70] and following [48]. Fifteen rabbits were divided into three groups: A, B, and C (5 rabbits each; n = 5). Each animal in groups A and B was injected with 0.5 ml venom aliquot that contained the equivalent LD95 of the W. aegyptia snake (0.264 mg/rabbit) and A. crassicauda scorpion (10.064 mg/rabbit), respectively. Venoms were intravenously injected into rabbits via the ear vein (Figure 1D) following [71]. The use of intravenous injection was chosen for its precision in delivering a controlled dose, although it does not perfectly mimic natural bites or stings. Envenomized rabbits were deceased within 10 to 20 min post-envenomization. In parallel, the control group (group C) was injected with 0.9% saline prior to euthanasia with CO2 according to [72] and following [46]. Dealing with and killing animals took place in accordance with the Research Ethical Committee at King Saud University (approval code: KSU-SE-23-83).
2.7. Experimental Design
Within a maximum of 1 h from confirmed death, rabbit corpses were immediately translocated to the experimental site. Corpses were individually placed in a metal cage (50 × 40 × 25 cm each) (Figs. 1E) to protect them against predation, as detailed in [45,46]. Cages were placed a minimum of 10 meters away from one another to provide isolated resources for corpse-seeking insects, as recommended by [73] and to reduce the effect of odor interference, which could affect insect attraction [48,74]. Three pitfall traps (each of 10 cm in diameter, 5 cm in depth) were placed adjacent to each corpse (Figure 1E) to collect the attracted insects beyond the collection times [75]. Each trap was containing a solution composed of 250 ml of water, 5% soap powder, and 5% NaCl according to [45]. Since direct sunlight impacts insect succession compared to shadow [76], all experimental corpses were standardized by placing them in shady places underneath trees.
2.8. Decomposition Process
Each corpse was examined and investigated for 5 minutes in a daily manner, from the first day of the exposure until complete skeletonization, according to [48,77]. The four known decomposition stages: fresh, bloating, decayed, and dry (Figure 3) were observed and described following [45,46,48]. The duration of each stage (in days) was reported until complete skeletonization (dryness).
2.9. Beetles Collection and Identification
Corpse-attracted beetles were collected from corpses on an hourly basis (2 minutes for each corpse) from 6 a.m. to 4 p.m. over the first three days of the experiment. Then, insects were collected once a day at 6 a.m. until the end of the experiment according to [78] and following [45,48]. Only adult were included in the counting of collected beetles during this study. Beetles were collected from and underneath corpses using soft forceps and a spatula (3 cm in width and 10 cm in length) following [79]. Pitfall traps were also used not only to maintain monitoring beyond the time of collection but also to reduce the disturbance of carcasses-inhabitant insects for later sampling. Collected beetles were preserved in 70% ethanol and stored at 4°C until used for counting and identification as detailed in [79]. Beetles were morphologically identified to the species level at the King Saud University Museum of Arthropods (https://cfas.ksu.edu.sa/en/node/3075) by the 3rd and 4th authors, H. Al-Dhafer and M. Abdel-Dayem, respictively.
2.10. Statistical Analysis
Mice-corrected mean lethality values were used to calculate the LD50 and LD95 of each venom using Probit Analysis following [68]. The resulting mean values from Probit analysis are considered significant (P < 0.5) if their 95% confidence limits (Upper to Lower) are not overlapped, according to [80,81]. The Minitab software (MINITAB, State College, PA, version 18.1, 2018, UK) was used to statistically analyze the results of total counts of beetles. Prior to any further analysis, the normality of insects’ counts was tested using the Anderson–Darling Normality Test, according to [82]. Because the overall data of beetle counts were normally distributed, the One-way ANOVA was used for the comparisons between the treated groups, and the Multiple Tukey’s Pairwise Comparison test was used to analyze the differences between means, according to [82]. However data sets pertaining to the coleopteran families and species counts were not normally distributed and, thus, were analyzed using the non-parametric Mann–Whitney U-test. Finally, singleton or doubleton is considered when only one or two individuals of a particular beetle species are reported, respectively, according to [83] and following [45,46,48,76]. All results are presented as means of 5 replicates using five different individual rabbit corpses (n = 5) ± standard errors (SE), as determined by the Basic Statistical Analyses
3. Results
3.1. Venoms Lethality
As shown in Table 1, Probit analysis revealed LD50 of the W. aegyptia snake venom was 26.7 times higher than that of the A. crassicauda scorpion (P < 0.05), with non-overlapping confidence limits. In addition, the LD95 of W. aegyptia venom was 38.12 times higher than that of A. crassicauda (P < 0.05), with non-overlapping confidence limits. These LD95 values were converted into the equivalent (4×) for rabbits (0.264 and 10.064 mg/rabbit, respectively) and were considered the lethal doses used for rabbit envenomization.
3.2. Meteorological Parameters
3.2.1. Atmospheric Parameters
The atmospheric parameters over the course of the 11-day experiment in Riyadh City are shown graphically in Figure 2. The maximum, minimum, and mean temperatures were constant (Figure 2A), with averages of 41.0, 29.27, and 35.14°C, respectively (Figure 2B). The relative humidity (RH) was fluctuating from 5% during the first day to 13% during the last day of the experiment (Figure 2A), with an average of 7.8% (Figure 2B). Wind temperature was constant with an average of 32.03°C, while wind speed was fluctuating from 5 to 31 km/h during days 7 and 9, respectively (Figure 2A), with an average of 13.15 km/h (Figure 2B). These atmospheric parameters are within the natural range during this time of the year in Riyadh city [44].
3.2.2. On-site Recorded Weather Parameters
The on-site manually recorded weather parameters (Figure 2C and D) were fluctuating over the experimental period compared to the officially recorded ones by the Saudi National Center for Meteorology for Riyadh City (Figure 2A and B). The temperature recorded a minimum of 29°C on day 6 and a maximum of 38°C on day 4 (Figure 2C), with an average of 34°C (Figure 2D). Relative humidity recorded a minimum of 30% and a maximum of 36% (Figure 2C), with an average of 32% (Figure 2D). Wind temperature recorded a minimum of 29°C and a maximum of 34°C (Figure 2C), with an average of 31.37°C (Figure 2D). Fluctuations were noticeable in the on-site recorded temperature, RH, and wind temperature. The maximum fluctuation was recorded in wind speed, as most days recorded 0.0 km/h, while the maximum speed was 18 km/h on day 10 (Figure 2C), with an average of 3.25 km/h (Figure 2D).
3.3. Decomposition Process
The four reported decomposition stages over the study period are the fresh, bloating, decay, and dry (Figure 3). In all three treatments, rabbit corpses appeared in the fresh stage as if they were alive in terms of softness and flexibility at the beginning of death (Figure 3A). This stage lasted for 31 hours in the control group, whereas it lasted for 21 hours in both envenomized corpses (Table 2). It was noticeable that snake- and scorpion-envenomized corpses became stiffened and somewhat rigid by 30 minutes and 6 hours postmortem, respectively, and both began to emit slight unpleasant odors. This may indicate that envenomization have shortened the duration of the fresh stage as the bloating stage started earlier compared to the control (Table 2).
In the bloating stage (Figure 3B), corpses started to swell from the abdominal area, progressing to the chest and neck, releasing offensive odor, but with different durations between treatments. This stage lasted for 10 and 24 hours in snake- and scorpion-envenomized corpses, respectively, compared to 14 hours in the control ones (Table 2). It was noticeable that scorpion-envenomized corpses were releasing stronger offensive odor in this stage compared to those of snake-envenomized and control ones. This may indicate that envenomization with snake venom shortened the bloating stage as the decay stage started earlier compared to those of control and scorpion-envenomized ones.
The decay stage also varied among the three treatments (Figure 3C1). The control corpses showed distributed decomposition across body areas, partial appearance of larvae from body openings, strong offensive odors, fluid exudation, and lasted for about 48 hours (Table 2). Snake-envenomized corpses started decomposition earlier and was observed in the chest and neck areas, the abdominal contents were noticeably spreading around the corpse, significant fluid exudation, and lasted for approximately 62 hours (Table 2). While scorpion-envenomized corpses started the decomposition with the control ones that was evident in the neck, chest, and forelimbs, with a stronger offensive odor. There weren’t as many corpse-colonizing larvae as in the control and snake-envenomized ones (Figure 3C2). The smell was also worse and stronger than in the other groups, and lasted for about 72 hours (Table 2). This may indicate that envenomization prolonged the duration of the decay stage compared to that of the control.
In the dry stage, all corpse groups were characterized by complete dryness and rigidity (Figure 3D). This stage started at approximately 117 hours (4.9 days) postmortem in scorpion-envenomized corpses compared to 93 hours (3.9 days) in both control and snake-envenomized ones (Table 2). This may indicate that envenomization with scorpion venom delayed the start of the dry stage (up to 1 day) since the durations of the preceding stages have increased.
3.4. Abundance of Corpse Associated Beetles
A total of 1094 corpse-attracted beetles were collected from all the experimental corpses during the entire experimental period of this study. Out of them, 36.2% (396) and 29.1% (319) beetles were collected from snake- and scorpion-envenomized corpses, respectively, compared to 34.7% (379) collected from the control ones. As shown in Figure (4A), beetles attracted to all corpses from the first day of exposure in fewer numbers increased to the maximum during days 4 and 5, after which it reduced again to the minimum at the end of the experiment. The Interval Plot of beetles succession in each treatment versus days postmortem over the experimental period (from day 1 to day 11) was calculated and created by the Pooled Standard Deviation, which revealed four major succession peaks (waves) of attracted beetles to corpses during the period from day 2 to day 7, with the highest peak on day 4 (Figure 4B)
When comparing the envenomized corpses over the 11 day-experimental period, the one way ANOVA revealed significant more beetles attracted to the snake-envenomized corpses on days 2 and 3, about the same number on day 4, and significantly fewer on day 5 compared to the scorpion-envenomized ones (F32,132 = 12.36, P < 0.05, n = 5) (Figure 4A). However, scorpion-envenomized corpses attracted significant more beetles later on day 5 (Figure 4A). These data may indicate that snake-envenomized corpses attracted more beetles during the first 4 days of exposure compared to the scorpion-envenomized ones, which attracted more beetles later. This may suggest a distinctive difference in beetles’ succession in a venom-dependent manner.
3.5. Differential Abundance of Beetles
The impact of venom type on the differential abundance of attracted coleopteran families is represented in Figure 5A. Overall, the most abundant families were Histeridae followed by Dermestidae, Scarabaeidae, and Tenebrionidae, while the least abundant ones were Zopheridae, followed by Ptinidae, Hybosoridae, Chrysomelidae, Nitidulidae, and Elateridae (Figure 5A). As shown in Figure 5B, the Interval Plot of the pooled standard deviations revealed three highest waves of beetles’ succession distinctively to the snake-envenomized corpses represented by (Histeridae, followed by Dermestidae and Scarabaeidae), followed by another five waves to the control ones by (Dermestidae followed by Histeridae, Scarabaeidae, Curculionidae, and Tenebrionidae). While only three moderate waves, represented by (Histeridae followed by Dermestidae and Scarabaeidae), were attracted to the scorpion-envenomized corpses. These data clearly show that Dermestidae, Scarabaeidae, and Histeridae are the 3 prevalent families attracted to all corpses, while Curculionidae and Tenebrionidae were distinctively attracted to control corpses only. These data may indicate that beetles are less attracted to envenomized corpses compared to control ones, the intensity of attracted families varies based on the type of venom; in a venom-dependent succession manner, and that scorpion-envenomized corpses attracted less families compared to the snake-envenomized ones (see also Figure 4).
The overall Kruskal–Wallis test revealed significant differences in the mean number of attracted beetles within treatments (α < 0.05, H = 182.74, DF = 41, P < 0.05). As shown in Figure 5A, Mann-Whitney U-test revealed that snake-envenomized corpses attracted significant higher numbers of beetles from Anthicidae and Staphylinidae but significant lower of Cleridae and Curculionidae, compared to control ones (P < 0.05, n = 5). However, scorpion-envenomized corpses attracted significantly higher numbers of beetles of Histeridae and Staphylinidae, but lower of Curculionidae and Tenebrionidae compared to control ones. When comparing the envenomized corpses, the snake-envenomized ones attracted significantly higher numbers of Anthicidae, Curculionidae, and Tenebrionidae but significant lower of Cleridae compared to scorpion-envenomized ones (P < 0.05, n = 5). While control corpses distinctively attracted Chrysomelidae, Elateridae, and Hybosoridae. These data may indicate that the recorded families are distinctively attracted to corpses in a venom-dependent succession manner and that more beetles were attracted to snake-envenomized corpses compared to scorpion-envenomized ones.
3.6. Differential Succession of Beetles
A total of 27 species of beetles belonging to 14 families were reported and identified in this study (Table 3). Of them, Anthicidae is represented by 38 specimens of Omonadus formicarius (Goeze, 1777); Chrysomelidae is represented by 4 Caryedon acaciae (Gyllenhal, 1833); Cleridae is represented by 23 Necrobia rufipes (Fabricius, 1781) and 8 Necrobia sp.; Curculionidae is represented by 2 Coccotrypes rhizophorae (Hopkins 1915) and 57 Dinoderus sp.; Dermestidae is represented by 9 Attagenus posticalis Fairmaire, 1879, 127 Dermestes maculatus De Geer, 1774, and 114 Dermestes frischi Kugelman, 1792; Elateridae is represented by 3 Aeoloides grisescens (Germar, 1844); Histeridae is represented by 294 Saprinus chalcites (Illiger, 1807) and 4 Saprinus caerulescens (Hoffmann, 1803); Hybosoridae is represented by 3 Hybosorus illigeri Reiche, 1853; Nitidulidae is represented by 8 Carpophilus hemipterus (Linnaeus, 1792) and a singleton (1 individual beetle) of Urophorus humeralis (Fabricius, 1798); Ptinidae is represented by 3 Stegobium paniceum (Linnaeus, 1758); Scarabaeidae is represented by 181 Aphodius adustus Klug, 1855, 27 Malader insanabilis (Brenske, 1894), and 10 Rhyssemus saoudi Pittino, 1984; Staphylinidae is represented by 4 Leptacinus sp. and 62 Philonthus sp.; Tenebrionidae is represented by a singleton of Adesmia cancellate Klug,1830, 11 Alphitobius diapernius (Pancer, 1797), 22 Mesostena pincticollis Solier, 1835, 45 Opatroides punctulatus Brulle, 1832, and 31 Thriptera crinite (Klug,1830); and Zopheridae is represented by a doubleton (2 individual beetles) of Synchita sp (Table 3).
Figure 6 represents the differential succession of the 14 coleopteran families during the four decomposition stages. Out of them, 8 families were attracted to all corpses in various succession manners (Figure 6A, C, D, E, G, K, L, and M). The non-parametric Mann-Whitney U-test revealed that control corpses attracted significant more beetles from Curculionidae, Histeridae, Nitidulidae, and Staphylinidae during the decay and/or dry stages comparted to the envenomized ones (P < 0.05, n = 5) (Figure 6C, D, K, and M). For comparing the envenomized corpses, the snake-envenomized ones attracted significant more beetles from Anthicidae, Dermestidae, and Histeridae during the decay stages compared to the scorpion envenomized ones (P < 0.05, n = 5) (Figure 6A, E, and G). While scorpion-envenomized corpses attracted significant more beetles from Cleridae and Histeridae during the dry stages compared to the snake-envenomized ones (P < 0.05, n = 5) (Figure 6C and G). This may indicate a differential succession to corpses in a distinctive venom-dependent manner.
The other 6 families were attracted to corpses in a selective manner. This was evidenced by Chrysomelidae, Elateridae, and Hybosoridae that were distinctively attracted to the control corpses only during most of the decomposition stages (Figure 6B, F and H). While Nitidulidae, Ptinidae, and Zopheridae were distinctively attracted to the control and snake-envenomized corpses only during the decay and dry stages (Figure 6I, J, and N). While, none was distinctively attracted to scorpion-envenomized corpses. This may indicate that these particular families are selectively attracted to corpses in a venom-dependent manner during definitive decomposition stages.
At the species level, data in Table 3 reveal that C. acaciae (Chrysomelidae) and A. grisescens (Elateridae), followed by a doubleton of C. rhizophorae (Curculionidae) and H. illigeri (Hybosoridae) were distinctively associated with control corpses only. While O. formicarius (Anthicidae), S. chalcites (Histeridae), A. adustus (Scarabaeidae), and O. punctulatus (Tenebrionidae) were predominantly associated with all corpses during most decomposition stages (Table 3). There was a singleton of U. humeralis (Nitidulidae) and A. cancellate (Tenebrionidae) reported distinctively associated with snake-envenomized corpses. While a double ton of Synchita sp. (Zopheridae) was distinctively associated with control and snake-envenomized corpses during the decay stages (Table 3). These data may suggest a possible definitive relationship between the occurrence of a single species and a particular stage of decomposition in a venom-dependent manner.
4. Discussion
The present study proposed a forensic scenario for the envenomation-related death and its impact on the postmortem decomposition process and the succession pattern of the corpse-attracted beetles. It is important to clarify three points: a) neurotoxic venoms from two different local venomous animals, the W. aegyptia snake and the A. crassicauda scorpion, have been targeted to achieve the goal of this study, b) venoms were freshly extracted from the animals and their lethality were determined prior to carrying out this study, and c) the experimental rabbits have been envenomized by intravenous-injection with the lethal doses of venoms aliquots rather than being directly bitten or stung with the snake or scorpion, respectively.
It is well-known that the process of corpses decomposition constitutes a crucial step in the recirculation of organic materials throughout the food-chain in the nutrition cycle [77,85]. From the forensic point of view, this process provide vital evidence in the legal investigations as it has unique spatial- and temporal-dependent successive stages that aid the investigators in determining the PMI [86,87]. This process usually takes place in 4-5 stages [7] depending on many interacting environmental and non-environmental factors. The environmental factors include the atmospheric temperature and humidity [88,89], the habitat [79,90,91], and the type of soil in scene [92]. While the non-environmental factors include the corpse’s chemical composition [8], microbial composition and activity [93,94], the activity of colonizing insects [95], the barriers such as clothes and coverages [96,97], and the cause of death [48,98,99,100].
Based on the aforementioned parameters, and in order to avoid any possible discrepancies, the current study was carried out in a botanical garden with a homogeneous clayey sandy soil, and all experimental groups of corpses were placed in identical metal cages in similar shady places away from the direct sunlight. Further, this study was conducted during June, which is a hot and dry month and characterized by stable climatic conditions with the normal atmospheric averages of temperature, humidity, and wind speed of this time of the year in Riyadh city [44]. Thus, through the 11-day experimental duration, .corpses reached the dry stage within 4 to 5 days in these atmospheric conditions. Therefor, we would expect that there was no external climatic impact on the decomposition process and beetle succession in this study, and hypothesize that the reported variation of the abundance and succession of beetles between the treated corpses may be attributed to the antemortem envenomization. Consequently, the current study reported four main successive decomposition stages: fresh, bloating, decaying, and dried, as per reported in our previously published works [45,46,48].
Envenomation by venomous animals is a considerable global death-leading cause [28,29,33,101,102]. In the Middle East and Africa, thousands of snakebites as well as scorpion and spider stings were documented [16,30,33,103,104]. Yet, few studies in some countries have investigated the decomposition process and succession of forensic insects on corpses upon death by envenomation with scorpion venoms [100,105] and snakes [48,51,106]. To the best of our knowledge, no such studies have been conducted in Saudi Arabia, and hence, the present study was undertaken to address this gap. Our data showed clearly that envenomization with the snake and scorpion venoms both reduced the duration of the fresh stage and elongated that of the decay stage. The reported significant variation in lethality between the W. aegyptia snake and the A. crassicauda scorpion has been reflected as a variation in the rate of decomposition. The prolonged decomposition rate was more pronounced in the scorpion-envenomized corpses in terms of longer duration in the bloating and decay stages. Consequently, the overall duration of the decomposition process was elongated in terms of a delay (up to one day) at the start of the dry stage compared to that of the snake-envenomized ones. In contrary, our per published work [48] reported an acceleration in the decomposition rate of rabbit corpses upon envenomization with venoms from the Egyption Naja haje and Cerastes cerastes snakes. These findings may indicate that the postmortem decomposition rate is venom-dependent and, hence, could lead to bias in the estimation of PMI. There are evidences for this conclusion provided by other studies that investigated intoxication with different types of toxic substances have differently impacted the postmortem carcass decomposition and insect succession [53,107,108,109]. Comprehending these venom-dependent phenomena is essential for forensic entomologists, as it facilitates the enhancement of PMI approximations by considering the particular venom variant implicated.
The lethality has been estimated differently in different studies for the W. aegyptia snake (from 0.170 to 0.180 mg/kg) [62,110] and the A. crassicauda scorpion (from 1.1 to 1.7 mg/kg) [63,69]. These variations could be attributed to the variation of the laboratory and experimental conditions between investigators and to many other factors [111,112]. Our data revealed similar lethality (1.4 mg/kg) to that previously reported. While it was around 3.3 times greater than that determined by Al-Sadoon’s group for the snake venom. This difference in lethality could be attributed to the variation in the injection routes and in both the type and body weight of the experimental animals [63,113]. In this regard, Al-Sadoon and his colleagues used the intraperitoneal route in rats (weighing 200–250 g each), while we used the subcutaneous route in mice (weighing 18-20 g each), as well as for other factors [111]. Our data also determined the snake venom lethality of 26.7 times that of the scorpion, which could be attributed to its chemical components as it contains many different kinds of enzymes [114,115] and non-enzymatic proteins [116]. Consequently, it has multiple modes of action, mainly cardio and neurotoxicity, interruption of many vital systemic functions [62,110,117,118,119], and finally systemic dysfunction leading to death [120]. While the A. crassicauda scorpion venom contains smaller neurotoxin polypeptides of low molecular weight simple proteins with lethal neurotoxic and paralytic effects [69] resulting in cardio-respiratory failure and finally death [120,121]. This variation in the chemical composition and mode of action, may explain the concomitant variation of the decomposition process, as scorpion-envenomized corpses decomposed slower than the control and snake-envenomed ones. In contrary, other causes of death, like antemortem heroin-injection, was found to accelerate the decomposition of rabbit corpses [71]. These findings may suggest that the antemortem cause of death could potentially impact the postmortem decomposition process and that envenomation impacts differently the decomposition process based on the type of venom.
It is well known that dipteran flies are the first to attract to corpses during the very early decomposition stages, while others, like coleopteran beetles, are usually attracted to corpses during the latter stages [7]. Our data showed a maximum abundance of corpse-attracted beetles between days 4 and 5 (during the decay stage). This unique pattern of insect succession could be due to the effect of unique cadaveric volatile organic components which give cadavers their unique smell and attract a wide range of cadaver-seeking insects. In this context, up to 104 cadaveric volatile chemical compounds were identified during the decomposition process [122]. Therefore, the stronger and more unpleasant smell that we smelled coming from scorpion-envenomized corpses, compared to those of the other groups, may be attributed to the effect of certain chemicals in the scorpion venom [63,65]. Evidence for this is provided by [122], who reported cadaveric volatile components differently in different biotopes. This may explain the attraction of few beetles to the scorpion-envenomized corpses compared to those of the control and snake-envenomized ones. Moreover, the delay in the abundance of attracted beetles on the scorpion-envenomized corpses (up to day 5), compared to the snake-envenomed ones, may be attributed to the difference in the chemical components and the mode of action of venoms. A recent study conducted by [51] reported fewer number of attracted beetles to envenomed carcasses compared to controls, and fewer attracted beetles to N. haje snake envenomed carcasses compared to C. cerastes snake-envenomed ones. However, other causes of death, like heroin injection, showed no impact on the postmortem beetles succession patterns [71].
Data of differential abundance revealed variation in the succession of the coleopteran families. The most predominant corpses-attracted families were Histeridae (298 beetles) during all decomposition stages except the fresh stage. The second was Dermestidae (250 beetle) followed by Scarabaeidae (218 beetles) both during all stages except fresh and bloating ones. The third was Tenebrionidae (110 beetles) during selective decomposition stages. The least was Zopheridae (2 beetles) during the decay stage. In agreement with our results, Dermestidae, Histeridae, and Scarabaeidae were also reported as predominant families associated with rabbit corpses [79,123] and with pig corpses [124]. These data suggest two differential succession manners: the first is a differential predominant manner shown by four families (Curculionidae, Histeridae, Nitidulidae, and Staphylinidae), three families (Anthicidae, Dermestidae, and Histeridae), and two families (Cleridae and Histeridae) that were particularly predominant on control, snake-envenomized, and scorpion-envenomized corpses, respectively, during the decay and/or dry stages. Histeridae was the only predominant family on all corpses, regardless of the type of treatment. The second manner is a distinctive predominant manner showed by three families (Chrysomelidae, Elateridae, and Hybosoridae) that were distinctively attracted to control corpses only, and three families (Nitidulidae, Ptinidae, and Zopheridae) that were attracted to both control and snake-envenomized corpses only. While no definitive families were distinctively attracted to scorpion-envenomized corpses. These differentially attracted beetles could serve as potential indicators for differentiating between the types of venoms and, consequently, suggest a venom-dependent succession manner. On the other hand, the unexpected presence of Chrysomelidae, Curculionidae, Elateridae, and Zopheridae on corpses provides insight into the intricate ecological interactions surrounding the corpses. These particular families are known primarily for their plant-feeding or wood-boring behaviors rather than being necrophagous [125]. This may be attributed to either the surrounding vegetation of the experimental site or their opportunistic feeding on fungi growing on corpses. This, in fact, may suggest a broader ecological context for the decomposition process, a potential impact on the succession pattern of more conventional necrophagous insects, and ecological indication of the crime scene, which could be of forensic relevance. In forensic investigations, the accurate identification of particular species of beetles may assist in ascertaining whether a fatality resulted from envenomation by a snake or a scorpion, thereby offering critical evidence in circumstances where the etiological factors of death remains ambiguous.
At the species level, C. acaciae (Chrysomelidae) was distinctively associated with control corpses only during all decomposition stages except the bloating stage. While O. formicarius (Anthicidae), S. chalcites (Histeridae), A. adustus (Scarabaeidae), and O. punctulatus (Tenebrionidae) were predominantly associated with all corpses during most decomposition stages. These data may indicate an association of a certain beetle species with the treatment but not with the decomposition stages. Further, the reported singleton and doubleton were associated with the fresh and decay stages, which may suggest an association of the singleton or doubleton with particular decomposition stages of a particular treatment. This is in contrary to the findings of [79], who reported no association between the attraction of a single species and a particular stage of decomposition. This may be due to the difference in the experimental sites, as they were comparing between different habitats (agricultural, desert, and urban). No definitive species was distinctively reported as associated with the scorpion envenomized corpses. Reporting certain corpse-associated beetles during the early stages of decomposition, like fresh and bloating stages, might be due to their seasonal appearance rather than linking to the decomposition stage [126].
5. Conclusions
This study revealed that antemortem envenomization of rabbits with snake and scorpion venoms has significantly affected the corpses’ decomposition rate, with the impact being more pronounced in scorpion-envenomized ones. This impact was evidenced by the elongation of the bloating and decay stages, as well as a delay of up to one day in reaching the dry stage compared to the control corpses. The differential abundance and succession pattern of corpse-associated beetles varied significantly between the envenomized and control corpses, suggesting a venom-dependent succession. Notably, families such as Histeridae, Dermestidae, Scarabaeidae, and Tenebrionidae were predominant in all corpses. While others showed distinctive associations with specific treatments or decomposition stages. At the species level, there were unique associations between certain beetle species and specific treatments or decomposition stages. For instance, the singleton of U. humeralis (Nitidulidae) and A. cancellate (Tenebrionidae) were distinctively associated with snake-envenomized corpses. The doubleton of Synchita sp. (Zopheridae) was distinctively associated with the control and snake-envenomized corpses. While, C. acaciae (Chrysomelidae) and A. grisescens (Elateridae), followed by a doubleton of C. rhizophorae (Curculionidae) and H. illigeri (Hybosoridae), were distinctively associated with control corpses solely. These findings may indicate that the succession of the reported beetles is venom-dependent, which could be helpful as envenomation markers. This may aid in understanding the influence of various venom types of various lethality from various venomous animals on both corpse decomposition and insect succession, as well as identifying which venom type has the most significant postmortem impact. Finally, the herbivorous (non-necrophagous) families, Chrysomelidae, Curculionidae, Elateridae, and Zopheridae, were unexpectedly reported associated with corpses, which could be potential ecological markers of the crime scene. This, in fact, may provide additional valuable forensic evidence in forensic investigation and updating the database of the envenomation-related corpse decomposition process and the associated beetle taxa in Saudi Arabia. These findings can enhance PMI estimation and aid in differentiating between causes of death involving venomous animals, ultimately contributing to more accurate crime scene reconstructions and legal investigations.
Author Contributions
Conceptualization, A.M.A. and M.S.Al-K; methodology, A.S.Al-Q.; software, A.M.A and A.S.Al-Q.; validation, A.M.A., M.S.Al-K, H.M.Al-D., and M.S.A-D.; formal analysis, A.M.A. and M.S.A-D.; investigation, A.S.Al-Q. and M.S.A-D.; resources, A.M.A. and A.S.Al-Q.; data curation, A.M.A. and A.S.Al-Q; writing—original draft preparation, A.M.A, A.S.Al-Q., and H.E.; writing—review and editing, A.M.A, A.S.Al-Q., M.S.A-D., and H.E.; visualization, A.M.A. and H.E.; supervision, A.M.A., M.S.Al-K, and H.M.Al-D.; project administration, A.M.A.; funding acquisition, A.M.A. and A.S.Al-Q. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been funded by the Researchers Supporting Project number (RSPD2025R695), King Saud University, Riyadh, Saudi Arabia.
Data Availability Statement
All data pertinent to this work are presented in this article. Further requests should be directed to the corresponding author.
Acknowledgments
We thank Researchers Supporting Project number (RSPD2025R695), King Saud University, Riyadh, Saudi Arabia, for funding this research. We wish to acknowledge Prof. Mohamed K. Al-Sadoon and his research group (Herpetology Laboratory) for preparing and providing the snake’s and scorpion’s venoms. Additionally, we would like to thank the Botany and Microbiology Department, College of Science, King Saud University, Riyadh, Saudi Arabia, for granting us access to the botanical garden for conducting the fieldwork of this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Byrd, J.H.; Brundage, A. Forensic entomology. In: Byrd JH, Norris P, Bradley-Siemens N, editors. Veterinary forensic medicine and forensic sciences. 1st ed. Boca Raton (London, New York): CRC Press, Taylor and Francis Group. 2020, 67–111.
- Amendt, J.; Richards, C.S.; Campobasso, C.P.; Zehner, R.; Hall, M.J. Forensic entomology: applications and limitations. Forensic science, medicine, and pathology 2011, 7, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Charabidze, D.; Gosselin, M.; Hedouin, V. Use of necrophagous insects as evidence of cadaver relocation: myth or reality? Peerj 2017, 5, e3506. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kumawat, R.; Singh, G.; Jangir, S.S.; Kushwaha, P.; Rana, M. Forensic entomology: a novel approach in crime investigation. GSC Biological and Pharmaceutical Sciences 2022, 19, 165–174. [Google Scholar] [CrossRef]
- Correa, R.C.; Caneparo, M.F.C.; Vairo, K.P.; de Lara, A.G.; Moura, M.O. What have we learned from the dead? A compilation of three years of cooperation between entomologists and crime scene investigators in Southern Brazil. Revista Brasileira De Entomologia 2019, 63, 224–231. [Google Scholar] [CrossRef]
- Hu, G.L.; Wang, M.; Wang, Y.; Liao, M.Q.; Hu, J.Y.; Zhang, Y.N.; Yu, Y.M.; Wang, J.F. Estimation of post-mortem interval based on insect species present on a corpse found in a suitcase. Forensic Science International 2020, 306, 110046. [Google Scholar] [CrossRef]
- Byrd, J.H.; Castner, J.L. Forensic entomology: the utility of arthropods in legal investigations; 2019; Volume CRC press, Taylor & Francis Group, Boca Raton, FL 33487-2742. ISBN: 978-0-8153-5020-0.
- Coe, J. Postmortem chemistry: practical considerations and a review of the literature. Journal of Forensic Sciences 1974, 19, 13–32. [Google Scholar] [CrossRef]
- Kulshrestha, P.; Satpathy, D.K. Use of beetles in forensic entomology. Forensic Sci Int 2001, 120, 15–17. [Google Scholar] [CrossRef]
- McKenna, D.D.; Farrell, B.D. Beetles (Coleoptera). In: SB, Hedges and S, Kumar, editors. The timetree of life. Oxford University Press, USA 2009, 278-289.
- Alajmi, R.; Abdel-Gaber, R.; Haddadi, R. Molecular identification of forensically important beetles in Saudi Arabia based on mitochondrial16 s rRNAgene. Entomological Research 2020, 50, 343–350. [Google Scholar] [CrossRef]
- Charabidze, D.; Colard, T.; Vincent, B.; Pasquerault, T.; Hedouin, V. Involvement of larder beetles (Coleoptera: Dermestidae) on human cadavers: a review of 81 forensic cases. International Journal of Legal Medicine 2014, 128, 1021–1030. [Google Scholar] [CrossRef]
- Lyu, Z.; Wan, L.H.; Yang, Y.Q.; Tang, R.; Xu, L.Z. A checklist of beetles (Insecta, Coleoptera) on pig carcasses in the suburban area of southwestern China: A preliminary study and its forensic relevance. Journal of Forensic and Legal Medicine 2016, 41, 42–48. [Google Scholar] [CrossRef]
- Pushkin, S.V.; Tsymbal, B.M.; Nagdalian, A.A.; Nuzhnaya, K.V.; Sutaeva, A.N.; Ramazanova, S.Z.; Maschenko-Grigorova, A.N.; Mishvelov, A.E. The use of model groups of necrobiont beetles (Coleoptera) for the diagnosis of time and place of death. Entomology and Applied Science Letters 2019, 6, 46–56. [Google Scholar]
- White, J. Venomous animals: Clinical Toxinology. In: Luch, A, editor. Molecular, clinical and environmental Toxicology: Volume 2: Clinical Toxicology. Birkhäuser Basel, Germany. 2010, 233-291. [CrossRef]
- Chippaux, J.-P. Snakebite in Africa: current situation and urgent needs. In: Handbook of venoms and toxins of reptiles, 2nd ed. CRC Press, Taylor & Francis Group, London, UK. [CrossRef]
- Fathinia, B.; RASTEGAR, P.N.; Darvishnia, H.; Rajabizadeh, M. The snake fauna of Ilam Province, southwestern Iran. Iranian Journal of Animal Biosystematics 2010, 6, 9–23. [Google Scholar]
- Al-Sadoon, M.K.; Paray, B.A.; Al-Otaibi, H. Survey of the reptilian fauna of the Kingdom of Saudi Arabia. VI. The snake fauna of Turaif region. Saudi Journal of Biological Sciences 2017, 24, 925–928. [Google Scholar] [CrossRef]
- McCartney, J.A.; Seiffert, E.R. A late Eocene snake fauna from the Fayum Depression, Egypt. Journal of Vertebrate Paleontology 2016, 36, e1029580. [Google Scholar] [CrossRef]
- Ernst, C.H.; Ernst, E.M. Snakes of the United States and Canada; Smithsonian Books Washington, DC: 2003; Volume 790.
- Al-Asmari, A.K.; Al-Saif, A.A.; Abdo, N.; Al-Moutaery, K.; Al-Harbi, N. A review of the scorpion fauna of Saudi Arabia. Egyptian Journal of Natural History 2013, 6, 1–21. [Google Scholar] [CrossRef]
- Lourenço, W.R. A historical approach to scorpion studies with special reference to the 20th and 21st centuries. Journal of Venomous Animals and Toxins including Tropical Diseases 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Kachel, H.S.; Al-Khazali, A.M.; Hussen, F.S.; Yağmur, E.A. Checklist and review of the scorpion fauna of Iraq (Arachnida: Scorpiones). Arachnologische Mitteilungen 2021, 61, 1–10. [Google Scholar] [CrossRef]
- Chippaux, J.-P. Snake venoms and envenomations; Krieger Publishing Company. Malabar, Florida, USA: 2006.
- White, J. Bites and stings from venomous animals: a global overview. Therapeutic drug monitoring 2000, 22, 65–68. [Google Scholar] [CrossRef]
- Kularatne, S.; Senanayake, N. Venomous snake bites, scorpions, and spiders. Handbook of clinical neurology 2014, 120, 987–1001. [Google Scholar] [CrossRef]
- Forrester, J.A.; Holstege, C.P.; Forrester, J.D. Fatalities from venomous and nonvenomous animals in the United States (1999–2007). Wilderness & environmental medicine 2012, 23, 146–152. [Google Scholar] [CrossRef]
- Forrester, J.A.; Weiser, T.G.; Forrester, J.D. An update on fatalities due to venomous and nonvenomous animals in the United States (2008–2015). Wilderness & Environmental Medicine 2018, 29, 36–44. [Google Scholar] [CrossRef]
- Forrester, J.D.; Forrester, J.A.; Tennakoon, L.; Staudenmayer, K. Mortality, hospital admission, and healthcare cost due to injury from venomous and non-venomous animal encounters in the USA: 5-year analysis of the National Emergency Department Sample. Trauma surgery & acute care open 2018, 3, e000250. [Google Scholar] [CrossRef]
- Chippaux, J.-P.; Goyffon, M. Epidemiology of scorpionism: a global appraisal. Acta tropica 2008, 107, 71–79. [Google Scholar] [CrossRef]
- Dehghani, R.; Fathi, B. Scorpion sting in Iran: a review. Toxicon 2012, 60, 919–933. [Google Scholar] [CrossRef]
- Santos, M.S.; Silva, C.G.; Neto, B.S.; Grangeiro Júnior, C.R.; Lopes, V.H.; Teixeira Júnior, A.G.; Bezerra, D.A.; Luna, J.V.; Cordeiro, J.B.; Júnior, J.G. Clinical and epidemiological aspects of scorpionism in the world: a systematic review. Wilderness & Environmental Medicine 2016, 27, 504–518. [Google Scholar] [CrossRef]
- Amr, Z.S.; Baker, M.A.A.; Warrell, D.A. Terrestrial venomous snakes and snakebites in the Arab countries of the Middle East. Toxicon 2020, 177, 1–15. [Google Scholar] [CrossRef]
- Schneemann, M.; Cathomas, R.; Laidlaw, S.; El Nahas, A.; Theakston, R.D.G.; Warrell, D.A. Life-threatening envenoming by the Saharan horned viper (Cerastes cerastes) causing micro-angiopathic haemolysis, coagulopathy and acute renal failure: clinical cases and review. Qjm 2004, 97, 717–727. [Google Scholar] [CrossRef]
- Rahav, G.; Weiss, A.T. Scorpion sting-induced pulmonary edema: scintigraphic evidence of cardiac dysfunction. Chest 1990, 97, 1478–1480. [Google Scholar] [CrossRef]
- du Toit-Prinsloo, L.; Morris, N.K.; Meyer, P.; Saayman, G. Deaths from bee stings: a report of three cases from Pretoria, South Africa. Forensic Science, Medicine, and Pathology 2016, 12, 81–85. [Google Scholar] [CrossRef]
- Hughes, R.L. A fatal case of acute renal failure from envenoming syndrome after massive bee attack: A case report and literature review. American Journal of Forensic Medicine and Pathology 2019, 40, 52–57. [Google Scholar] [CrossRef]
- Haddad Junior, V.; Amorim, P.C.H.d.; Haddad Junior, W.T.; Cardoso, J.L.C. Venomous and poisonous arthropods: identification, clinical manifestations of envenomation, and treatments used in human injuries. Revista da sociedade brasileira de medicina tropical 2015, 48, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Al-Kathiri, W.H.; Balkhi, B.; Samrkandi, O.; Al-Khalifa, M.S.; Asiri, Y. The burden of bites and stings management: Experience of an academic hospital in the Kingdom of Saudi Arabia. Saudi pharmaceutical journal 2020, 28, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadoon, M.K.; Albeshr, M.F.; Paray, B.A.; Al-Mfarij, A.R. Envenomation and the bite rate by venomous snakes in the kingdom of Saudi Arabia over the period (2015–2018). Saudi Journal of Biological Sciences 2021, 28, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Alhamoud, M.A.; Al Fehaid, M.S.; Alhamoud, M.A.; Alzoayed, M.H.; Alkhalifah, A.A.; Menezes, R.G. Scorpion stings in Saudi Arabia: an overview. Acta Bio Medica: Atenei Parmensis 2021, 92. [Google Scholar] [CrossRef]
- Al-Sadoon, M.; Jarrar, B. Epidemiological study of scorpion stings in Saudi Arabia between 1993 and 1997. Journal of venomous animals and toxins including tropical diseases 2003, 9, 54–64. [Google Scholar] [CrossRef]
- Jarrar, B.M.; Al-Rowaily, M.A. Epidemiological aspects of scorpion stings in Al-Jouf province, Saudi Arabia. Annals of Saudi medicine 2008, 28, 183–187. [Google Scholar] [CrossRef]
- National Center for Meteorology. Meteorological services. https://ncm.gov.sa/Ar/EService/met/Pages/default.aspx (accessed on Joune, 2023). Available online: https://ncm.gov.sa/Ar/EService/met/Pages/default. (accessed on Joune, 2023).
- Al-Qurashi, A.S.; Mashaly, A.M.; Alagmi, R.; Al-Khalifa, M.S.; Mansour, L.; Al-Omar, S.Y.; Sharaf, M.R.; Aldawood, A.S.; Al-Dhafer, H.M.; Hunter, T. A preliminary investigation of rabbit carcass decomposition and attracted ants (Hymenoptera: Formicidae) on the seaward coastal beach of Al-Jubail City, Saudi Arabia. Journal of Medical Entomology 2023, 61, 318–330. [Google Scholar] [CrossRef]
- Almutawa, M.e.Y.; Al-Khalifa, M.S.; Al-Dhafer, H.M.; Abdel-Dayem, M.S.; Ebaid, H.; Ahmed, A.M. Forensic investigation of carcass decomposition and dipteran fly composition over the summer and winter: a comparative analysis of indoor versus outdoor at a multi-story building. Journal of Medical Entomology 2024, 61, 877–890. [Google Scholar] [CrossRef]
- Al Farhan, A.H.; Aldjain, I.M.; Thomas, J.; Miller, A.G.; Knees, S.G.; Llewellyn, O.; Akram, A. Botanic gardens in the Arabian Peninsula. Sibbaldia: the International Journal of Botanic Garden Horticulture. [CrossRef]
- Khalil, A.; Zidan, M.M.; Alajmi, R.; Ahmed, A.M. Impact of envenomation with snake venoms on rabbit carcass decomposition and differential adult dipteran succession patterns. Journal of Medical Entomology 2023, 60, 40–50. [Google Scholar] [CrossRef]
- Mapara, M.; Thomas, B.S.; Bhat, K. Rabbit as an animal model for experimental research. Dental research journal 2012, 9, 111. [Google Scholar] [CrossRef]
- Dautartas, A.; Kenyhercz, M.W.; Vidoli, G.M.; Jantz, L.M.; Mundorff, A.; Steadman, D.W. Differential decomposition among pig, rabbit, and human remains. Journal of Forensic Sciences 2018, 63, 1673–1683. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.; Salem, A.M.; Shaurub, E.-S.H.; Ahmed, A.M.; Al-Khalaf, A.A.; Zidan, M.M. Envenomation with snake venoms as a cause of death: A forensic investigation of the decomposition stages and the impact on differential succession pattern of carcass-attracted coleopteran beetles. Insects 2024, 15, 902. [Google Scholar] [CrossRef]
- Park, J.K.; Kim, C.J.; Do, Y. Post mortem insect colonization and body weight loss in rabbit carcasses. Entomological Research 2020, 50, 594–600. [Google Scholar] [CrossRef]
- Zeariya, M.; Hammad, K.M.; Fouda, M.A.; Al-Dali, A.G.; Kabadaia, M.M. Forensic-insect succession and decomposition patterns of dog and rabbit carcasses in different habitats. Journal of Entomology and Zoology Studies 2015, 3, 473–482. [Google Scholar]
- Alqahtan, A.R. Ecological notes on snake diversity in Tathleeth District, Aseer Region, Southwest of Saudi Arabia. Egyptian Academic Journal of Biological Sciences, B. Zoology 2017, 9, 61–70. [Google Scholar] [CrossRef]
- Masood, M.F. Ecological distribution of snakes’ fauna of Jazan region of Saudi Arabia. Egyptian Academic Journal of Biological Sciences, B. Zoology 2012, 4, 183–197. [Google Scholar] [CrossRef]
- Alshammari, A.M. Four new records of snake species in Ar’ar region, Northern border of Saudi Arabi. Jordan Journal of Natural History 2022, 9, 41–48. [Google Scholar]
- Wikipedia. Sinai-Desert-Cobra. By Ltshears. https://en.wikipedia.org/wiki/File:Sinai-Desert-Cobra.jpg. (Visited on 24). Available online: https://commons.wikimedia.org/wiki/File:Sinai-Desert-Cobra. 20 December.
- Alqahtani, A.R.; Badry, A.; Abd Al Galil, F.M.; Amr, Z.S. Morphometric and meristic diversity of the species Androctonus crassicauda (Olivier, 1807)(Scorpiones: Buthidae) in Saudi Arabia. PeerJ 2022, 10, e14198. [Google Scholar] [CrossRef]
- Al-Asmari, A.; Al-Saief, A.; Abdo, N.; Al-Moutaery, K. New additions to the scorpion fauna of Riyadh region, Saudi Arabia. Journal of Venomous Animals and Toxins including Tropical Diseases 2009, 15, 612–632. [Google Scholar] [CrossRef]
- Alqahtani, A.R.; Elgammal, B.; Ghaleb, K.I.; Badry, A. The scorpion fauna of the southwestern part of Saudi Arabia. Egyptian Academic Journal of Biological Sciences, B. Zoology 2019, 11, 19–29. [Google Scholar] [CrossRef]
- Timokhanov, V. Scorpion Androctonus crassicauda. https://freelance.ru/science_art/work-298408.html (visited on December, 2024). 2009.
- Al-Sadoon, M.K.; Orabi, G.M.; Badr, G. Toxic effects of crude venom of a desert cobra, Walterinnesia aegyptia, on liver, abdominal muscles and brain of male albino rats. Pakistan Journal of Zoology 2013, 45. [Google Scholar]
- Boghozian, A.; Nazem, H.; Fazilati, M.; Hejazi, S.H.; Sheikh Sajjadieh, M. Toxicity and protein composition of venoms of Hottentotta saulcyi, Hottentotta schach and Androctonus crassicauda, three scorpion species collected in Iran. Veterinary Medicine and Science 2021, 7, 2418–2426. [Google Scholar] [CrossRef]
- Calvete, J.J.; Pla, D.; Els, J.; Carranza, S.; Damm, M.; Hempel, B.-F.; John, E.B.; Petras, D.; Heiss, P.; Nalbantsoy, A. Combined molecular and elemental mass spectrometry approaches for absolute quantification of proteomes: application to the venomics characterization of the two species of desert black cobras, Walterinnesia aegyptia and Walterinnesia morgani. Journal of proteome research 2021, 20, 5064–5078. [Google Scholar] [CrossRef]
- Al-Asmari, A.K.; Kunnathodi, F.; Al Saadon, K.; Idris, M.M. Elemental analysis of scorpion venoms. Journal of venom research 2016, 7, 16. [Google Scholar]
- Broad, A.; Sutherland, S.; Coulter, A.R. The lethality in mice of dangerous Australian and other snake venom. Toxicon 1979, 17, 661–664. [Google Scholar] [CrossRef]
- WHO. World Health Organization Expert Committee on Biological Standardization, sixty-seventh report; Technical Report Series- World Health Organization. https: //iris.who.int/bitstream/handle/10665/255657/9789241210133-eng.pdf. (Visited on December, 2024): 2017; Volume 1004. [Google Scholar]
- Finney, D.J. Probit analysis 3rd edition. Cambridge University, London, UK. 1971, Volume 333. [CrossRef]
- Ozkan, O.; Kar, S.; Güven, E.; Ergun, G. Comparison of proteins, lethality and immunogenic compounds of Androctonus crassicauda (Olivier, 1807) (Scorpiones: Buthidae) venom obtained by different methods. Journal of Venomous Animals and Toxins including Tropical Diseases 2007, 13, 844–856. [Google Scholar] [CrossRef]
- Jacob, S.; Nair, A.B.; Morsy, M.A. Dose conversion between animals and humans: A practical solution. Indian J. Pharm. Educ. Res 2022, 56, 600–607. [Google Scholar] [CrossRef]
- Al-Qahtni, A.; Mashaly, A.; Haddadi, R.; Al-Khalifa, M. Seasonal impact of heroin on rabbit carcass decomposition and insect succession. Journal of Medical Entomology 2021, 58, 567–575. [Google Scholar] [CrossRef]
- Conlee, K.; Stephens, M.; Rowan, A.N.; King, L.A. Carbon dioxide for euthanasia: concerns regarding pain and distress, with special reference to mice and rats. Laboratory animals 2005, 39, 137–161. [Google Scholar] [CrossRef]
- Velásquez, Y. A checklist of arthropods associated with rat carrion in a montane locality of northern Venezuela. Forensic Science International 2008, 174, 68–70. [Google Scholar] [CrossRef]
- Lewis, A.; Benbow, M. When entomological evidence crawls away: Phormia regina en masse larval dispersal. Journal of Medical Entomology 2011, 48, 1112–1119. [Google Scholar] [CrossRef]
- Majer, J.D. The use of pitfall traps for sampling ants–a critique. Memoirs of the Museum of Victoria 1997, 56, 323–329. [Google Scholar] [CrossRef]
- Mashaly, A.; Mahmoud, A.; Ebaid, H. Relative insect frequency and species richness on sun-exposed and shaded rabbit carrions. Journal of Medical Entomology 2020, 57, 1006–1011. [Google Scholar] [CrossRef]
- Parmenter, R.R.; MacMahon, J.A. Carrion decomposition and nutrient cycling in a semiarid shrub–steppe ecosystem. Ecological Monographs 2009, 79, 637–661. [Google Scholar] [CrossRef]
- Goff, M.L. Early postmortem changes and stages of decomposition. In: Amendt, J., Goff, M. L., Campobasso, C. P., & Grassberger, M. (Eds). Current concepts in forensic entomology. Dordrecht: Springer Dordrecht Heidelberg London New York. 2010, 1-24. [CrossRef]
- Mashaly, A.M.A. Carrion beetles succession in three different habitats in Riyadh, Saudi Arabia. Saudi journal of biological sciences 2017, 24, 430–435. [Google Scholar] [CrossRef]
- Litchfield, J.j.; Wilcoxon, F. A simplified method of evaluating dose-effect experiments. Journal of pharmacology and experimental therapeutics 1949, 96, 99–113. [Google Scholar] [CrossRef]
- Matsuda, K. Changes in the insecticide susceptibility of the American serpentine leafminer, Liriomyza trifolii (Diptera: Agromyzidae), in indoor successively reared and crop field populations over 25 years. Applied Entomology and Zoology 2021, 1–10. [Google Scholar] [CrossRef]
- Morrison, D.A. How to improve statistical analysis in parasitology research publications. International Journal for Parasitology 2002, 32, 1065–1070. [Google Scholar] [CrossRef]
- Novotný, V.; Basset, Y. Rare species in communities of tropical insect herbivores: pondering the mystery of singletons. Oikos 2000, 89, 564–572. [Google Scholar] [CrossRef]
- Sakuma, M. Probit analysis of preference data. Applied Entomology and Zoology 1998, 33, 339–347. [Google Scholar] [CrossRef]
- Weathers, K.C.; Strayer, D.L.; Likens, G.E. Fundamentals of ecosystem science; Elsevier, Academic Press. SanDiego, USA: ISBN: 978-0-12-088774-3: 2012.
- Haglund, W.D.; Sorg, M.H. Advances in forensic taphonomy: Method, theory, and archaeological perspectives (1st ed.). CRC Press. Boca Raton London New York Washington, DC. 2002.
- Vanin, S.; Zanotti, E.; Gibelli, D.; Taborelli, A.; Andreola, S.; Cattaneo, C. Decomposition and entomological colonization of charred bodies–a pilot study. Croatian medical journal 2013, 54, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.C.; Giardina, C.P.; Litton, C.M.; Francisco, K.S.; Pacheco, C.; Thomas, N.; Uehara, T.; Metcalfe, D.B. Impacts of insect frass and cadavers on soil surface litter decomposition along a tropical forest temperature gradient. Ecology and Evolution 2022, 12, e9322. [Google Scholar] [CrossRef]
- Iancu, L.; Dean, D.E.; Purcarea, C. Temperature influence on prevailing necrophagous Diptera and bacterial taxa with forensic implications for postmortem interval estimation: A review. Journal of Medical Entomology 2018, 55, 1369–1379. [Google Scholar] [CrossRef]
- Mashaly, A.M.A.; Al-Mekhlafi, F.A. Differential Diptera succession patterns on decomposed rabbit carcasses in three different habitats. Journal of Medical Entomology 2016, 53, 1192–1197. [Google Scholar] [CrossRef]
- Dalal, J.; Sharma, S.; Bhardwaj, T.; Dhattarwal, S.K.; Verma, K. Seasonal study of the decomposition pattern and insects on a submerged pig cadaver. Journal of Forensic and Legal Medicine 2020, 74. [Google Scholar] [CrossRef]
- Fancher, J.; Aitkenhead-Peterson, J.; Farris, T.; Mix, K.; Schwab, A.; Wescott, D.; Hamilton, M. An evaluation of soil chemistry in human cadaver decomposition islands: Potential for estimating postmortem interval (PMI). Forensic science international 2017, 279, 130–139. [Google Scholar] [CrossRef]
- Dangerfield, C.R.; Frehner, E.H.; Buechley, E.R.; Sekercioglu, C.H.; Brazelton, W.J. Succession of bacterial communities on carrion is independent of vertebrate scavengers. Peerj 2020, 8. [Google Scholar] [CrossRef]
- Cobaugh, K.L.; Schaeffer, S.M.; DeBruyn, J.M. Functional and structural succession of soil microbial communities below decomposing human cadavers. PloS one 2015, 10, e0130201. [Google Scholar] [CrossRef]
- Soon, L.P.; See, K.L.; Ahmad, N.W.; Abdullah, K.; Hasmi, A.H. A scoping review on factors affecting cadaveric decomposition rates. Journal of Forensic Sciences and Criminal Investigation 2017, 2, 555584. [Google Scholar] [CrossRef]
- Card, A.; Cross, P.; Moffatt, C.; Simmons, T. The effect of clothing on the rate of decomposition and Diptera colonization on Sus scrofa carcasses. Journal of Forensic Sciences 2015, 60, 979–982. [Google Scholar] [CrossRef]
- Mashaly, A.M.; Mahmoud, A.; Ebaid, H. Influence of clothing on decomposition and presence of insects on rabbit carcasses. Journal of Medical Entomology 2019, 56, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Al-Khalifa, M.; Mashaly, A.; Al-Qahtni, A. Impacts of antemortem ingestion of alcoholic beverages on insect successional patterns. Saudi Journal of Biological Sciences 2020, 28, 685–692. [Google Scholar] [CrossRef]
- Al-Qahtni, A.; Mashaly, A.; Haddadi, R.; Al-Khalifa, M. Seasonal impact of heroin on rabbit carcass decomposition and insect succession. Journal of Medical Entomology 2020, 58, 567–575. [Google Scholar] [CrossRef]
- Abd El-Aziz, F.E.; Eldeeb, S.M.; Abdellah, N.Z.; Shaltout, E.S.; Ebrahem, N.E. Influence of scorpion venom on decomposition and arthropod succession. Egypt Acad J Biol Sci B Zool 2022, 14, 209–219. [Google Scholar] [CrossRef]
- Edstrom, A. Venomous and poisonous animals; Krieger Publishing Company, Malabar, Florida, USA: 1992.
- Biery, T.L. Venomous Arthropod Handbook: Envenomization symptoms/treatment, identification, biology and control. Disease Surveillance Branch, Epidemiology Division, USAF School of Aerospace. U.S. Government Printing Office: 1977 O—251-048, Washington, DC. 20402, USA; 1977.
- Cichutek, K.; Epstein, J.; Griffiths, E.; Hindawi, S.; Jivapaisarnpong, T.; Klein, H.; Minor, P.; Moftah, F.; Reddy, V.; Slamet, L. WHO Expert Committee on Biological Standardization Sixty-seventh report. Technical Report Series-World Health Organization 2017, 1004, 1–591. [Google Scholar]
- Coelho, P.; Sousa, P.; Harris, D.; van der Meijden, A. Deep intraspecific divergences in the medically relevant fat-tailed scorpions (Androctonus, Scorpiones). Acta tropica 2014, 134, 43–51. [Google Scholar] [CrossRef]
- El-Aziz, A.; Kasem, S.M.; Ebrahem, N.E. Evaluation of the toxicity of scorpion venom and digoxin on human cardiovascular system and in decomposition arthropods succession using rat carrions. Egyptian Academic Journal of Biological Sciences, B. Zoology 2022, 14, 1–16. [Google Scholar] [CrossRef]
- Abdou, R.H.; Ibrahim, A.E. Effects of Egyptian cobra (Naja haje) venom on postmortem changes and some biochemical parameters in rats. Forensic Science 2015, 4. [Google Scholar]
- Abd El-Gawad, A.; Badawy, R.M.; Abd El-Bar, M.M.; Kenawy, M.A. Successive waves of dipteran flies attracted to warfarin-intoxicated rabbit carcasses in Cairo, Egypt. The Journal of Basic and Applied Zoology 2019, 80, 1–10. [Google Scholar] [CrossRef]
- Al-Qahtni, A.; Mashaly, A.; Haddadi, R.; Al-Khalifa, M. Seasonal impact of heroin on rabbit carcass decomposition and insect succession. J Med Entomol 2020, 58, 567–575. [Google Scholar] [CrossRef]
- Al-Khalifa, M.; Mashaly, A.; Al-Qahtni, A. Impacts of antemortem ingestion of alcoholic beverages on insect successional patterns. Saudi journal of biological sciences 2021, 28, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadoon, M.K.; Fahim, A.; Salama, S.F.; Badr, G. The effects of LD50 of Walterinnesia aegyptia crude venom on blood parameters of male rats. African journal of microbiology research 2012, 6, 653–659. [Google Scholar] [CrossRef]
- Oukkache, N.; Jaoudi, R.E.; Ghalim, N.; Chgoury, F.; Bouhaouala, B.; Mdaghri, N.E.; Sabatier, J.-M. Evaluation of the lethal potency of scorpion and snake venoms and comparison between intraperitoneal and intravenous injection routes. Toxins 2014, 6, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Biologyinsights. LD50 values: Mechanisms, influences, and pharmacological applications. In: Pathology and Diseases. 2024. https://biologyinsights.com/ld50-values-mechanisms-influences-and-pharmacological-applications/?form=MG50AV53 (Visiten on December, 2024).
- WHO. World Health Organization: Progress in the characterization of venoms and standardization of antivenoms; WHO: Geneva, Switzerland. https://iris.who.int/bitstream/handle/10665/37282/WHO_OFFSET_58.pdf?sequence=1 (Visited on December, 2024); 1981. [Google Scholar]
- Al-Saleh, S.S.; Khan, S. Purification and characterization of phosphodiesterase i from Walterinnesia aegyptia venom. Preparative Biochemistry & Biotechnology 2011, 41, 262–277. [Google Scholar] [CrossRef]
- Abid, I.; Jemel, I.; Alonazi, M.; Ben Bacha, A. A new group II phospholipase A2 from Walterinnesia aegyptia venom with antimicrobial, antifungal, and cytotoxic potential. Processes 2020, 8, 1560. [Google Scholar] [CrossRef]
- Aziz, T.M.A.E.; Bourgoin-Voillard, S.; Combemale, S.; Beroud, R.; Fadl, M.; Seve, M.; De Waard, M. Fractionation and proteomic analysis of the Walterinnesia aegyptia snake venom using OFFGEL and MALDI-TOF-MS techniques. Electrophoresis 2015, 36, 2594–2605. [Google Scholar] [CrossRef]
- Al-Jammaz, I. Physiological effect of LD50 of Walterinnesia aegyptia crude venom on rat metabolism over various periods of time. Pakistan Journal of Biological Sciences 2001, 4, 1429–1431. [Google Scholar]
- Alhazza, I. Effect of Walterinnesia aegyptia, Cerastes cerastes and Bitis arietans venoms on liver functions of male rats. Pakistan Journal of Zoology 2001, 33, 157–165. [Google Scholar]
- Khalil, F.; Zaki, K.; Naguib, M. Effect of black snake (Walterinnesia aegyptia) venom on the respiratory activity of some tissues of the rabbit. Zeitschrift für vergleichende Physiologie 1964, 48, 270–276. [Google Scholar] [CrossRef]
- Postma, T.L. Neurotoxic animal poisons and venoms. Clinical Neurotoxicology 2009, 55, 463–489. [Google Scholar]
- Safdarian, M.; Vazirianzadeh, B.; Ghorbani, A.; Pashmforoosh, N.; Baradaran, M. Intraspecific differences in Androctunus crassicauda venom and envenomation symptoms. EXCLI journal 2022, 21, 1222. [Google Scholar] [CrossRef] [PubMed]
- Dekeirsschieter, J.; Verheggen, F.; Gohy, M.; Hubrecht, F.; Bourguignon, L.; Lognay, G.; Haubruge, E. Cadaveric volatile organic compounds released by decaying pig carcasses (Sus domesticus L.) in different biotopes. Forensic Science International 2009, 189, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Al-Dakhil, A.A.; Alharbi, S.A. A preliminary investigation of the entomofauna composition of forensically important necrophagous insects in Al-Madinah Al-Munawwarah region, Kingdom of Saudi Arabia. Journal of Taibah University for Science 2020, 14, 1127–1133. [Google Scholar] [CrossRef]
- Sonker, R.; RAWAT, S.; SINGH, K. Succession and life cycle of beetles on the exposed carcass. Int J Sci Innov Res 2015, 1, 46–50. [Google Scholar]
- Evans, A.V.; Bellamy, C.L. An Inordinate Fondness for Beetles; University of California Press. Nevraumont Publishing Company, New York: 2000.
- VanLaerhoven, S.L.; Anderson, G.S. Insect succession on buried carrion in two biogeoclimatic zones of British Columbia. J Forensic Sci 1999, 44, 32–43. [Google Scholar] [CrossRef]
Figure 1.
A displaying plate shows A: part of the botanical garden targeted by the study, B: the desert black snake, W. aegyptia (from [57]), C: the fat-tail scorpion, A. crassicauda (from [61]), D: the ear intravenous injection of rabbits, and E: an example of rabbit corpses in its protective metal cage along with the pitfall traps.
Figure 1.
A displaying plate shows A: part of the botanical garden targeted by the study, B: the desert black snake, W. aegyptia (from [57]), C: the fat-tail scorpion, A. crassicauda (from [61]), D: the ear intravenous injection of rabbits, and E: an example of rabbit corpses in its protective metal cage along with the pitfall traps.

Figure 2.
The meteorological parameters over the experimental period show the atmospheric parameters (A) and their overall averages (B) in Riyadh city, and the on-site manually recorded daily weather parameters (C) and their overall averages (D) at the experimental sites. Max.T: maximum temperature, Min.T: minimum temperature, M.T: the mean of maximum and minimum temperatures (n = 11), RH: relative humidity, WT: wind temperature, and WS: wind speed.
Figure 2.
The meteorological parameters over the experimental period show the atmospheric parameters (A) and their overall averages (B) in Riyadh city, and the on-site manually recorded daily weather parameters (C) and their overall averages (D) at the experimental sites. Max.T: maximum temperature, Min.T: minimum temperature, M.T: the mean of maximum and minimum temperatures (n = 11), RH: relative humidity, WT: wind temperature, and WS: wind speed.
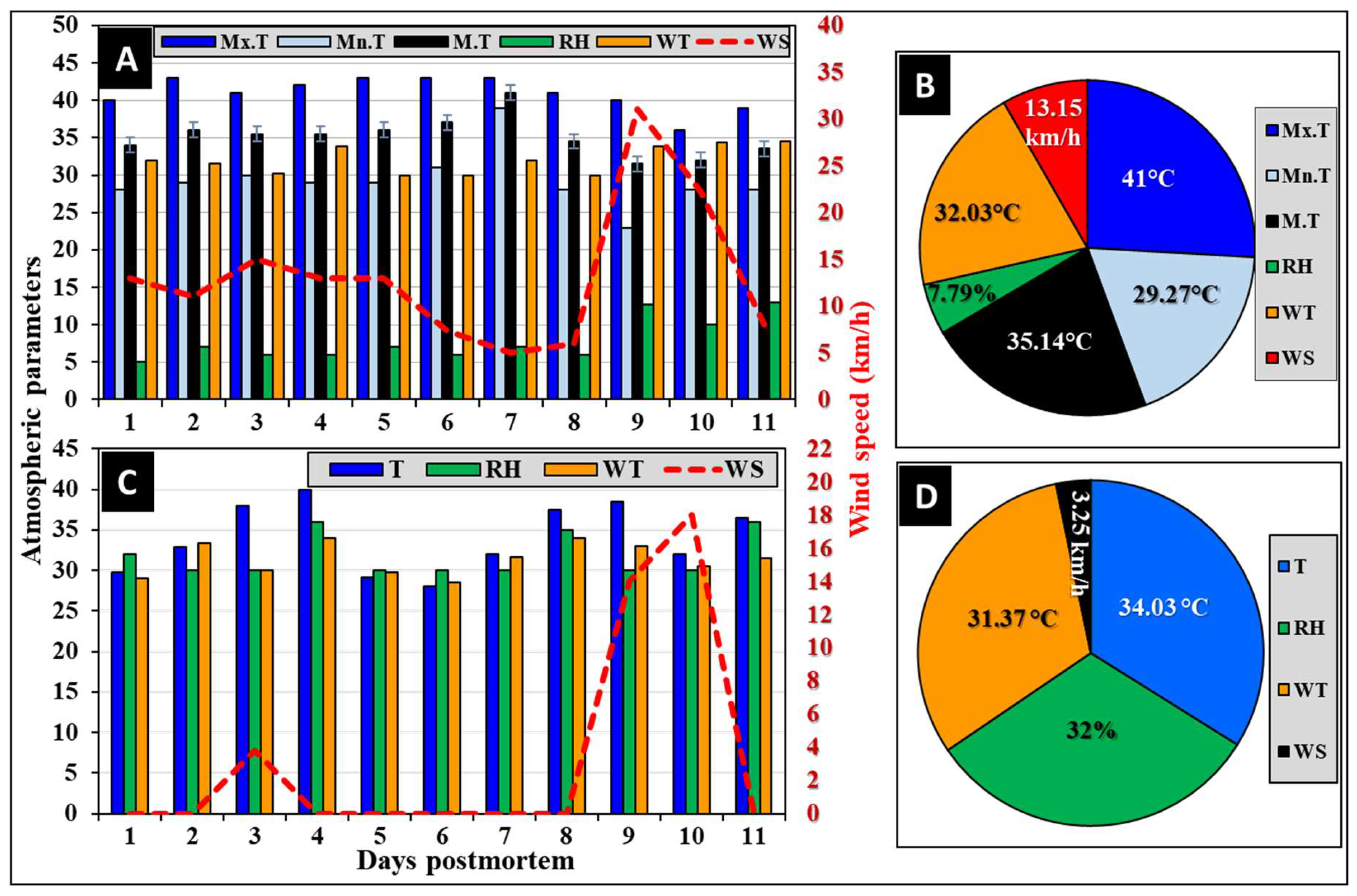
Figure 3.
The postmortem characteristic features of the reported four decomposition stages of the rabbit corpses inside their metal cages. A: Fresh stage, B: bloating stage, C1 & C2: decayed stage & the associated colonizing larvae, respectively, and D: dry stage.
Figure 3.
The postmortem characteristic features of the reported four decomposition stages of the rabbit corpses inside their metal cages. A: Fresh stage, B: bloating stage, C1 & C2: decayed stage & the associated colonizing larvae, respectively, and D: dry stage.

Figure 4.
The abundance of corpse-attracted beetles post-envenomization (A), and the Interval Plot of mean numbers of beetles (B) in each treatment versus days over the experimental period (from D1 to D11). C: control, Sn: snake-envenomized, and Sc: scorpion-envenomized corpses. Error bars represent standard errors of means of 5 replicates (n = 5). The sign (*) indicates significant differences compared to the control, while sign (§) indicates significant differences between envenomized corpses (P < 0.05, n = 5).
Figure 4.
The abundance of corpse-attracted beetles post-envenomization (A), and the Interval Plot of mean numbers of beetles (B) in each treatment versus days over the experimental period (from D1 to D11). C: control, Sn: snake-envenomized, and Sc: scorpion-envenomized corpses. Error bars represent standard errors of means of 5 replicates (n = 5). The sign (*) indicates significant differences compared to the control, while sign (§) indicates significant differences between envenomized corpses (P < 0.05, n = 5).
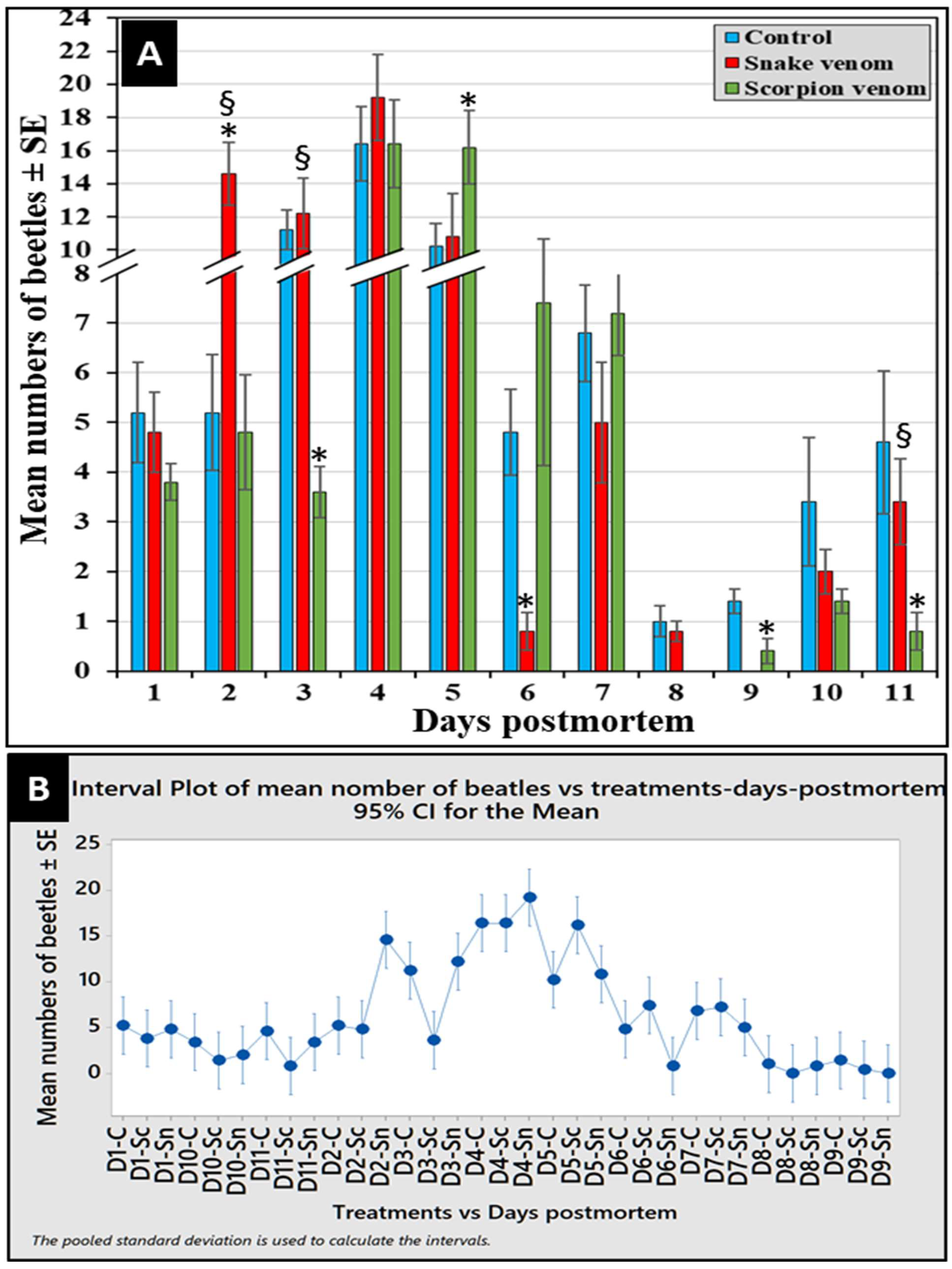
Figure 5.
The abundance of reported corpse-attracted coleopteran families post-envenomization over the experimental period (A). Error bars represent the standard errors (SE) of means of 5 replicates (n = 5). The Interval Plot of the pooled standard deviations (B) shows various succession waves of coleopteran families versus envenomization with snake- (Sn), scorpion-envenomized (Sc), and control (C). An: Anthicidae, Ch: Chrysomelidae, Cl: Cleridae, Cu: Curculionidae, De: Dermestidae, El: Elateridae, Hi: Histeridae, Hy: Hybosoridae, Ni: Nitidulidae, Pt: Ptinidae, Sc: Scarabaeidae St: Staphylinidae, Te: Tenebrionidae, and Zo: Zopheridae. The sign (*) indicates significant differences compared to the control, while sign (§) indicates significant differences between envenomized corpses (P < 0.05).
Figure 5.
The abundance of reported corpse-attracted coleopteran families post-envenomization over the experimental period (A). Error bars represent the standard errors (SE) of means of 5 replicates (n = 5). The Interval Plot of the pooled standard deviations (B) shows various succession waves of coleopteran families versus envenomization with snake- (Sn), scorpion-envenomized (Sc), and control (C). An: Anthicidae, Ch: Chrysomelidae, Cl: Cleridae, Cu: Curculionidae, De: Dermestidae, El: Elateridae, Hi: Histeridae, Hy: Hybosoridae, Ni: Nitidulidae, Pt: Ptinidae, Sc: Scarabaeidae St: Staphylinidae, Te: Tenebrionidae, and Zo: Zopheridae. The sign (*) indicates significant differences compared to the control, while sign (§) indicates significant differences between envenomized corpses (P < 0.05).

Figure 6.
Differential abundance of corpse-attracted coleopteran families during the decomposition stages: fresh (Fr), bloating (Bl), decayed (De), and dry (Dr). Individual figures represent families Anthicidae (A), Chrysomelidae (B), Cleridae (C), Curculionidae (D), Dermestidae (E), Elateridae (F), Histeridae (G), Hybosoridae (H), Nitidulidae (I), Ptinidae (J), Scarabaeidae (K), taphylinidae (L), Tenebrionidae (M), and Zopheridae (N). The sign (*) indicates significant difference compared to the control, while (§) indicates significant difference between envenomized corpses (P < 0.05, n = 5).
Figure 6.
Differential abundance of corpse-attracted coleopteran families during the decomposition stages: fresh (Fr), bloating (Bl), decayed (De), and dry (Dr). Individual figures represent families Anthicidae (A), Chrysomelidae (B), Cleridae (C), Curculionidae (D), Dermestidae (E), Elateridae (F), Histeridae (G), Hybosoridae (H), Nitidulidae (I), Ptinidae (J), Scarabaeidae (K), taphylinidae (L), Tenebrionidae (M), and Zopheridae (N). The sign (*) indicates significant difference compared to the control, while (§) indicates significant difference between envenomized corpses (P < 0.05, n = 5).
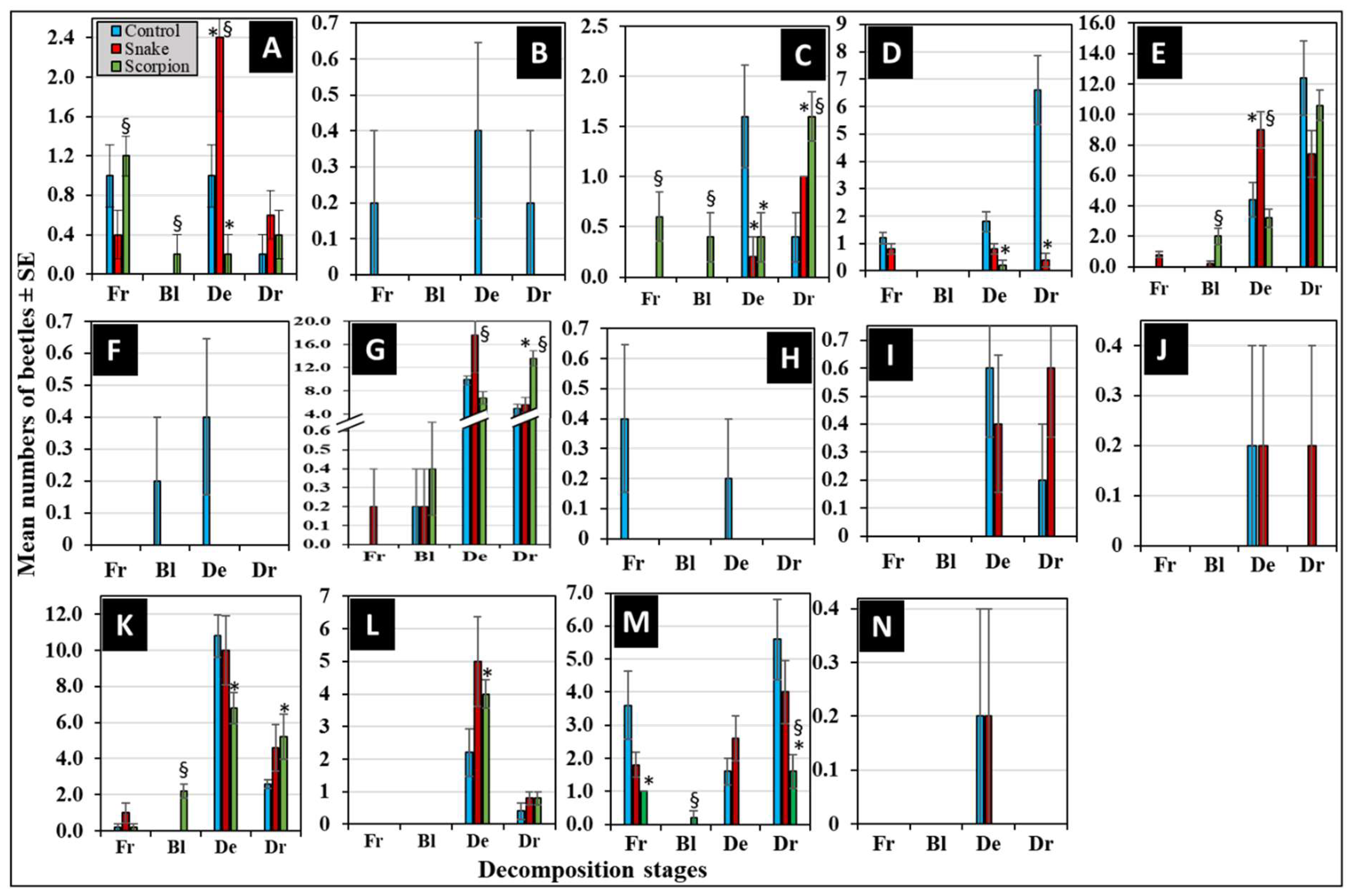

Table 1.
Results of Probit analysis showing LD50 and LD95 of snake and scorpion venoms in mice at 24h post-envenomization.
Table 1.
Results of Probit analysis showing LD50 and LD95 of snake and scorpion venoms in mice at 24h post-envenomization.
| Venom types | LD50 (mg/kg) (lower-upper) |
LD95 (mg/kg) (lower-upper) |
Slope ± SE |
|---|---|---|---|
| W. aegyptia | 0.053* (0.052-0.054) |
0.066** (0.063-0.069) |
18.29 ± 3.22 |
| A. crassicauda | 1.416* (1.288-1.558) |
2.516** (2.251-2.813) |
6.59 ± 0.91 |
Table 2.
Duration of the decomposition stages of envenomized corpses post-envenomization with the snake and scorpion venoms.
Table 2.
Duration of the decomposition stages of envenomized corpses post-envenomization with the snake and scorpion venoms.
| Types of venoms | Days postmortem | |||||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||||||||
| Control | ||||||||||||||||||
| W. eagiptia | ||||||||||||||||||
| A. crassicauda | ||||||||||||||||||
| Keys | Fresh stage | Bloated stage | Decay stage | Dry stage | ||||||||||||||
Table 3.
Recorded coleopteran families and species at different stages of decomposition of the experimental copses.
Table 3.
Recorded coleopteran families and species at different stages of decomposition of the experimental copses.
| Coleopteran Families (Total number) |
Beetle Species (Total number) |
Control | W. aegyptia | A. crassicauda | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fr | Bl | De | Dr | Fr | Bl | De | Dr | Fr | Bl | De | Dr | ||
| Anthicidae (38) | O. formicarius (38) | + | – | + | + | + | – | + | + | + | + | + | + |
| Chrysomelidae (4) | C. acaciae (4) | + | – | + | + | – | – | – | – | – | – | – | – |
| Cleridae (31) | N. rufipes (23) | – | – | + | + | – | – | + | + | – | – | + | + |
| Necrobia sp. (8) | – | – | + | – | – | – | – | + | + | + | – | – | |
| Curculionidae (59) | Dinoderus sp. (57) | + | – | + | + | + | – | + | + | – | – | + | – |
| C. rhizophorae (2) | + | – | – | – | – | – | – | – | – | – | – | – | |
| Dermestidae (250) | D. maculatus (127) | – | – | + | + | – | + | + | + | – | + | + | + |
| D. frischi (114) | – | – | + | + | – | – | + | + | – | + | + | + | |
| A. posticalis (9) | – | – | + | + | + | – | + | – | – | – | + | – | |
| Elateridae (3) | A. grisescens (3) | – | + | + | – | – | – | – | – | – | – | – | – |
| Histeridae (298) | S. chalcites (294) | – | + | + | + | + | + | + | + | – | + | + | + |
| S. caerulescens (4) | – | – | – | – | – | – | + | – | – | – | + | + | |
| Hybosoridae (3) | H. illigeri (3) | + | – | + | – | – | – | – | – | – | – | – | – |
| Nitidulidae (9) | C. hemipterus (8) | – | – | + | + | – | – | + | + | – | – | – | – |
| U. humeralis (1) | – | – | – | – | – | – | + | – | – | – | – | – | |
| Ptinidae (3) | S. paniceum (3) | + | – | + | – | – | – | – | – | – | – | – | – |
| Scarabaeidae (218) | A. adustus (181) | + | – | + | + | + | – | + | + | – | + | + | + |
| R. saoudi (10) | – | – | + | + | + | – | + | + | – | + | + | + | |
| M. insanabilis (27) | – | – | + | + | + | – | + | + | + | + | + | + | |
| Staphylinidae (66) | Philonthus sp. (62) | – | – | + | + | – | – | + | + | – | – | + | + |
| Leptacinus sp. (4) | – | – | – | – | – | – | + | – | – | – | – | – | |
| Tenebrionidae (110) | M. pincticollis (22) | + | – | + | + | + | – | – | + | + | – | – | – |
| T. crinite (31) | + | – | + | + | + | – | – | + | + | – | – | + | |
| A. diapernius (11) | – | – | – | + | – | – | – | – | – | – | – | – | |
| O. punctulatus (45) | + | – | + | + | + | – | + | + | + | + | – | + | |
| A. cancellate (1) | – | – | – | – | + | – | – | – | – | – | – | – | |
| Zopheridae (2) | Synchita sp.(2) | – | – | + | – | – | – | + | – | – | – | – | – |
Fr: fresh stage, Bl: bloating stage, De: decayed stage, Dr: dry stage. The positive sign (+) indicates recorded beetle while negative sign (-) indicates no record.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



