Submitted:
01 May 2025
Posted:
05 May 2025
You are already at the latest version
Abstract
OPN is a phosphorylated glycoprotein found in all vertebrate organisms and expressed in many tissues and secretions. It’s a pleiotropic protein that has different roles in many pathological and physiological processes. OPN is involved in many tissue transformation situations such as intestinal, brain development, immune system regulation of the ac-tivation of immune cells, inflammatory response and the immune system. This protein increases the functionality of the digestive system by regulating the intestinal microbiome and may help strengthen the intestinal barrier. OPN can also affect cognitive devel-opment and behavior. In addition, recently the association of OPN with cancer has been of critical importance. It has been shown that the expression of OPN increases in many types of cancer studied, which can promote the metastasis of tumor cells. OPN is also effective against the interaction with bacteria and infections, it can prevent the adhesion of bacteria, and this has supported the creation of new therapeutic approaches for oral care. Also, the supplement of OPN to infant formula has positively affected the immune and intestinal health of infants. Many studies conducted in recent years have focused on these issues. This article contains a review and comparison of existing information on the structure and functions of OPN. It emphasizes how milk OPN affects human, infant health and illness.
Keywords:
Osteopontin
; Biomarker
; Cancer
; Infant health
; Intestinal development
; Temporal ex-pression of cytokines
; Immunomodulator
; Milk
1. Introduction
Osteopontin (OPN) is a multifunctional protein present in many human tissue and body fluids including bone, skin, urine, milk and blood [1]. OPN is essentially explained as a phosphoprotein associated with transformation, and it was defined in an extracellular matrix of bovine bone [2,3]. Oldberg et al. recommended the term OPN ("osteo" is a Greek for bone and "pontin " is an originated from Latin word " pons" is a Latin for bridge because of its capability to form a bridge between cells and mineral parts of bone [4]. OPN, also referred to as secreted phosphoprotein 1 (SPP1), bone sialoprotein I (BSP I), T-lymphocyte activation-1 (ETA-1), 2ar and lactopontin [5,6,7,8,9]. OPN is found in small integrin-binding ligand N-linked glycoprotein (SIBLING) [10]. OPN composed of 314 amino acids abundant in aspartate, glutamate, and serine residues [11] with a molecular weight range 44-75 kDa [3], encoded through the action of the human SPP1 gene. The OPN-encoding gene is found on chromosome 4 region 22 and consisting of 7 exons. OPN contains a number of preserved structural domains, the RGD do-main (arginine-glycine-aspartate), the SVVYGLR domain (serine-valine-valine-tyrosine-glutamate-leucine-arginine), the heparin-binding domain, the calcium-binding domain, the sites of cleavage for matrix metalloproteinases, and the CD44 receptor-binding domain [10]. OPN, which attaches to integrin and CD44 receptors, works by participating in cellular signaling and cell-matrix interactions [12].
When OPN, which is bound to more than one integrin receptor, is stimulated by these integrin receptors, it increases the migration and invasion of cells [13]. OPN is expressed in many species and is found in humans and rodents among these species [7,14,15,16,17]. OPN-expressing cells: Osteoclasts, osteoblast, breast, kidney and skin epithelial cells, nerve cells, vascular smooth muscle cells and endothelial cells [7,18,19,20,21]. Active cells of the immune system, such as T cells, natural killer cells (NK cells), macrophages and Kupffer cells produce OPN, and the produced OPN is dispersed to body fluids [22,23,24].
OPN is found in many physiological and pathological processes. Behavior of OPN differs in different conditions such as physiological and pathological. In physiological settings, amount of OPN expression is lower and it has crucial functions in regulating biomineralization, wound healing, and developmental processes; in pathological set-tings, OPN is frequently considerable increase and exhibits broad-spectrum actions, in a number of inflammatory, autoimmune, degenerative, fibrosis, and oncologic disorders, including cancer, diabetes, stroke, kidney damage, and cardiac, fibrosis [25]. In addition, using OPN can provide benefits in many areas such as the immune system [26,27,28,29], biomarker [30,31,32,33,34,35,36], tissue repair, bone [37] and dental health [38,39,40], neurological conditions [41,42,43]and microencapsulation [37].
2. Structure and Chemical Properties of Osteopontin
OPN is a highly phosphorylated glycoprotein and has acidic features [44]. OPN in-cludes 30 monosaccharides high in aspartic acid, glutamic acid, and serine (Figure 1). OPN is a glycoprotein comprising approximately 314 amino acids and incorporating O-linked and N-linked oligosaccharides. The molecular weight range of OPN can change between 44-75 kDa [45,46]. This change occurs due to the OPN being subjected to various post-translational modifications (PTM) (glycosylation, phosphorylation, sulfation, etc.), therefore increasing functional diversity [47].
Differences in the amount of amino acid between the mature human and bovine OPN were shown by studies. According to these studies, the amount of mature human milk OPN contains 298 amino acids, and bovine milk (bmOPN) includes 262 amino acids. The main reason for the deficiency is the absence of 22 residues in bovine OPN, which correspond to 188-209 amino acids in the mature human OPN. The OPN sequences of bovine and mature human are similar to each other in 182 locations. Moreover, each type of OPN has a lot of aspartic acid, glutamic acid, and phosphorylated residues; these residues give an acidic character to OPN. The pI of OPN was measured without the addition of phosphorylation residues, and the result was found to be 4.1, which proves that OPN has an acidic character in this case [5]. It is acknowledged that OPN contains an argi-nine-glycine-aspartic acid (RGD)-binding area, two binding areas, one thrombin-binding area, and a calcium-binding area. Furthermore, OPN is divided by matrix metalloproteinases 3 and 7 (MMP-3,7). When proteases divide OPN, N-terminal and C-terminal fragments are generally formed. While the N-terminal contains integrin receptors, the C-terminal includes two heparin-binding areas and is moreover connected with CD44 [49]. Additionally, the RGD and SVVYGLR sequences, two integrin-binding areas, exclusively interact with integrin when OPN undergoes proteolysis. Bovine OPN also carries the SVAYGLK sequence [47,50]. While approximately 22 phosphorylated amino acids are included in bovine milk and dispersed in 28 distinct areas, approximately 25 phosphorylated amino acids are composed and dispersed in 34 phosphoserines and 2 phosphothreonines. A study identifies OPN as the most phosphorylated protein in the human colostrum [51]. Protein phosphorylation which is post-translational modification is critical for regulated cellular functions, activation signal transmission pathways etc. During this process the kinases that cause the phosphorylation of certain release proteins were not identified. Subsequently, FAM20C was found in Golgi lumen and was identified. As a result of mutations in this enzyme, errors occurred in the bone structure, so this enzyme was accepted as casein kinase [52].
2.1. Biosynthesis of Osteopontin
OPN, first expressed by Senger in 1979 and then named by Franzen in 1985, is a phosphoprotein [53]. OPN in humans is a protein that is sequenced on chromosome 4 and encoded by the SPP1 gene [54]. The SPP1 gene is located in the chromosome region 4q22.1. It also occupies 7.7 kb of space containing 7 exons [55]. There are 314 amino acid residues in human OPN (hOPN) [56]. The hOPN protein, which has a molecular weight of approximately 35 kDa, is exposed to too many posttranslational modifications. For this reason, it is observed between 40-80 kDa on the SDS-PAGE [51,57]. OPN has a predominant serine phosphorylation and acidic amino acids. For this reason, OPN has a negative charge [58]. There are two accepted heparin binding sites and a calcium binding site in OPN [59]. Also, the specific RGD sequences can be identified, connected and expressed through integrins located on the cell surface. The biosynthesis process of OPN has begun with the acquisition of the primary structure of a bone-specific sialoprotein from cloned cDNA. It has 1473 base pairs long and encodes a protein that has 317 amino acid residues. The sialoprotein mRNA is a coding region containing the AUG initiation codon and the signal peptide sequence [4]. The process of biosynthesis begins with the transcription of the SPP1 gene from DNA to mRNA. Then ribosomes synthesize the production of OPN protein by using this produced mRNA. During this synthesis, alternative splicing may cause the formation of different isoforms [55]. These are alternative splicing isoforms and alternative translation isoforms. OPNa, OPNb and OPNc are examples of splicing isoforms. OPNa contains exons 2-7, but OPNb does not have exon 5, while OPNc does not have 4. All human alternative splicing isoforms have a signal sequence that protects protein sequence, calcium binding domain, thrombin and other critical OPN domains and regions. For in-stance, amino acid sequences containing SVYGLR and RGD, along with cut-off points by matrix metalloproteases (MMPs). It is an alternative translational isoform, which is another OPN isoforms. The SPP1 gene blocks the secretion of OPN when the signal sequence is removed or not used to create alternative translational isoforms and allows the production of intracellular OPN (iOPN), which creates a secreted OPN (sOPN) if the SPP1 transcript contains the signal sequence [60]. The polypeptide chain formed as a result of translation undergoes post-translational modification. Phosphorylation takes place specific to the cell depending on the characteristics and type of tissue. OPN is exposed to various post-translational modifications including serine and threonine phosphorylation. As an example, human milk OPN has undergone a high amount of phosphorylation. In addition, human milk contains 36 phosphate zones of OPN. These are 34 phosphoserine, 2 phospho-threonine, 5 O-glycosylated threonine and N-glycosylation have been indicated. 29 of the phosphorylation sites are found in the region also called MGCK (mammalian milk casein kinase) or Golgi kinase. These phosphorylations are Ser8, Ser10, Ser11, Ser46, Ser47, Thr50, Ser60, Ser62, Ser65, Ser83, Ser86, Ser89, Ser92, Ser104, Ser110, Ser113, Thr169, Ser179, Ser208, Ser218, Ser238, Ser247, Ser254, Ser275, Ser287, Ser292, and Ser294. In addition, 6 phosphorylations are present in the CKII (casein kinaseII) sequence. These phosphorylations are Ser101, Ser107, Ser175, Ser199, Ser212, and Ser251. However, Ser203 is not included in the MGCK or CKII target sequences. Also, there are 5 threonine residues that have under-gone O-glycosylation. These are Thr118, Thr122, Thr127, Thr131 and Thr136 [51]. Studies conducted on OPN (hOPN) found in human milk show that it has been found that there are some characteristics of O-glycosylation occurring in 5 threonine residues. It has been found that the resulting O-glycosylation shows resistance to pepsin digestion of integrin binding motifs and shows the property of aiding cell adhesion [61]. In another study, it was found that phosphorylation and glycosylation, which occur if modification sites are located in and near the integrin binding sites, can modulate the interaction of OPN and integrin, for example the phosphorylation of Ser146 has been found to critically reduce the RGD-mediated αvβ3 integrin interaction [61,62]. As a result, studies support that exposure of the OPN to post-translational modifications may affect the functions of OPN.
3. Effects of Environmental Conditions and Genetic Factors on OPN Levels
The World Health Organization (WHO), the United Nations Children's Fund, and the American Academy of Pediatrics support the idea that human milk is the most effective and the only food source that is worth gold during the first 6 months of life for newborn children [63,64]. Breast milk is a possible source of nutrition for babies and is the main nutrient that babies physiologically need for growth and development [65]. Breastfed babies are less likely to develop certain health problems than non-breastfed babies and have a protective effect on the intestinal health and development of breastfed babies [66,67,68]. At the same time, breast milk also has a potential effect on cognitive development [69]. It’s formed thanks to many bioactive elements such as proteins and peptides with antimicrobial and immune-stimulating properties such as lactoferrin, lactoperoxidase, lysozyme, and IgA found in human milk . It was found that OPN, which is important among the bioactive components, has many functions including immune system recovery, neurodevelopment, and protecting infants from[28] disease-causing pathogens [70]. Some studies have found that the OPN concentrations in different geographies are different from each other [65,71,72]; at the same time, the OPN concentrations in the milk of mothers in the same region were found to be different from each other in some studies [72,73]. Several studies conducted in China have examined the change of OPN concentrations in breast milk [71,74,75]. In one of the studies, the components of breast milk were tested by ELISA and the protein content in breast milk was tested by the BCA method. 318 milk samples were taken from 106 mothers. These samples were taken from mothers with different breastfeeding periods. 343.2 ± 163.5 mg/L, 228.4 ± 121.5 mg/L, 204.8 ± 100.6 mg/L were measured from mothers at 1-14 days, 2-4 months, and 5-7 months, respectively. It was shown that amount of OPN measured was very high during the first 2 weeks, and the amount of OPN began to decrease over time. In other words, the amount of OPN in colostrum and transition milk was higher than in mature milk. In addition, the study found that the amount of OPN contained in milk was associated with the mother’s muscle mass. Also, the study showed that OPN is associated with fetal factors during different breastfeeding periods [71]. A study conducted in 2014 similarly looked at the amount of OPN, but in this study, the amounts of OPN in the umbilical cord blood were examined. As a result of this examination, it was realized that the amount of OPN was related to the mother's gestational age [76]. In another study conducted in China, milk samples were taken from 105 breastfeeding women after childbirth. Samples were collected over a period of 1-5 days, 8-14 days, 1 month, 6 months, and the average amount of OPN was measured at 718 mg/L, 586 mg/L, 450 mg/L and 236 mg/L, respectively. In this study ultra-performance liquid chromatography combined with mass spectrometry was used to detect protein concentrations. As a result of the study, it was found that the amount of OPN decreases during breastfeeding, and in addition, the milk proteins present in the mother may change depending on variables related to childbirth [74]. Another study conducted in China on the same topic examined the amount of OPN found in the milk of mothers who gave birth just on time and early. In this study, milk samples were taken from 131 mothers who gave birth at different times. These times are divided into 4 groups to be term, medium-late preterm (MPT), very preterm (VPT), and extreme preterm (EPT). The milk samples used in the study were collected on the 7th, 14th, 28th, and 120th days after birth. The amount of OPN was determined by ultra-performance liquid chromatography-tandem mass spectrometry (UPLC-MS/MS) and multiple reaction monitoring (MRM) techniques. As in the other study, it was found that the amount of OPN decreased over time in this study. Pre-pregnancy body mass index (BMI) affects colostrum and transitional milk. Therefore, BMI is one of the factors that affect OPN. As a result of the study, it was found that the amount of OPN may vary depending on maternal factors. Differing amounts of OPN may influence the growth and development of infants [75]. Another study was carried out in America. Although there is an abundant amount of OPN in human milk, it is not found in infant formula. In this study, it was shown that supplementing infant formula with bmOPN will lead to better results for the immunity of infants. The amounts of OPN in human milk were measured with ELISA method. In this study milk samples were taken at times 1-7 days, 8-14 days, 1 month, 4 months, and 12 months and, the OPN were taken respectively 178.0 ± 17.9 mg/L, 134.8 ± 18.5 mg/L, 65,8 ± 13.7 mg/L, 55,9 ± 13.8 mg/L, 48.3 ± 10.2 mg/L has been found. In this study, it was shown that the amount OPN decreases over time [72]. Another study conducted in America is to investigate milk proteins and the specific glycosylation to which these proteins belong, and to analyze changes in the lactation process. A total of 231 milk samples were taken from 33 mothers with 7 different breastfeeding periods. Tandem mass spectrometry was used in these milk samples. After the samples were taken from colostrum, the amounts of OPN found were taken at 2, 5,10,13,17, and 24 weeks and average were analyzed as 180, 320, 300, 260, 190, 200, 150, respectively. In this study the amount of OPN in general decreased over time, but the amount of OPN in 2 weeks is approximately 2 times the amount of OPN in colostrum [73]. Another country where the study was conducted is Türkiye. In the study, the association between the amount of OPN in breast milk and maternal factors was investigated, as well as the importance of the amount of OPN for neonatal health. Samples were taken from a total of 85 mothers. These mothers are grouped according to different characteristic features. These groups were ranked according to age, birth method, BMI, weight gain during pregnancy, and smoking use. Firstly, 4 groups were formed according to their ages. OPN quantities of these groups are respectively 137.8 ± 65.8, 141.4 ± 53.8 mg/L, 153.6 ± 49.3 mg/L, 121.9 ± 55.9 mg/L measured. Two groups were determined depending on the method of birth. These are the cervical vaginal route and the cesarean section. The measured OPN amounts were 160.6 ± 48.8 mg/L, 99.9 ± 48.5 mg/L depending on these birth methods. For BMI, it is divided into two groups as pre-pregnancy and postpartum. It was examined under the headings of underweight, optimal, overweight and obese in both groups. Pre-pregnancy results, respectively 140.6 ± 37.9 mg/L, 143.6 ± 53.6 mg/L, 111.9 ± 56.8 mg/L, 159.1 ± 73.5 mg/L were measured. The underweight not measured in postpartum, the remaining results are respectively 156.4 ± 46.2 mg/L, 140.8 ± 61.2 mg/L, 78.9 ± 28.8 mg/L were measured. 3 groups were determined depending on weight gain during pregnancy. These groups are defined as insufficient, adequate, excessive and the amount of OPN respectively 158.2 ± 40.3 mg/L,149.0 ± 60.4 mg/L, 119.8 ± 57.4 mg/L values have been determined. Finally, OPN concentrations were examined according to smoking status. In this study, 2 groups were formed as pre-pregnancy and postpartum/lactation. The mothers in the pre-pregnancy groups were divided into groups according to their cigarette consumption before and during pregnancy. The OPN values of mothers who smoked before pregnancy were 102.0 ± 41.4 mg/L, and those of mothers who did not smoke were 160.4 ± 53.8 mg/L. And it is 103.1 ± 50.6 mg/L of mothers who consumed cigarettes during pregnancy, while it is 143.8 ± 55.9 mg/L of mothers who did not use it. The amount of OPN of mothers who consumed cigarettes in the postpartum/lactation group was 98.3 ± 49.2 mg/L, while the amount of OPN of mothers who did not use was 144.1 ± 55.5 mg /L. When these results were examined, the OPN amounts of mothers who consume cigarettes are less than those of mothers who do not use. It was found that amounts of OPN contained in breast milk may vary based on maternal factors. In addition, these changes may impact on the growth, development and immunity of infants [65]. Another study was conducted in Japan. Cytokines in human milk were examined in 1989 and 2013. Colostrum samples were taken from 48 breastfeeding Japanese mothers and mature milk samples were taken from 49 mothers. The amount of OPN of colostrum measured in 1989 was approximately 318.1 mg/L, while in 2013 it was 137 mg/L, at the same time, the amount of OPN of mature milk measured in 1989 was 300.8 mg/L, while in 2013 it was 280 mg/L. The amount of OPN present in colostrum in 1989 was found to be significantly higher but no difference was observed in mature milk [77].
As a result of these studies, it has been found that the amount of OPN varies depending on maternal factors and different geographical regions, and these changes can affect the growth, development and immune system of infants. However, the reasons why there are different amounts of OPN in different geographical regions are not stated. More detailed research is needed on this issue.
4. Biological Functions of Osteopontin
The OPN, a pleiotropic protein, plays significant roles in all vertebrates. OPN plays a role in many biological processes (Figure 2) [45]. It also has multifunctional activity in many fields such as inflammation, biomineralization, biomarkers [32,33,34,35,36], dental health [38,39,40], cancer [78,79,80,81], diabetes, and wound healing, among others [45]. Thanks to multifunctional properties of OPN, it has been shown that it can play important roles in the diagnosis and treatment of many diseases (Table 1) [32,33,34,35,36,78,79,80,81]. Also, it allows the development of various treatment strategies.
4.1. Osteopontin as a Biomarker
OPN is a glycoprotein found in all vertebrates and plays a significant role in the cellular process and is therefore involved in many tissues and physiological secretions in many cells. It was first identified in the extracellular matrix of bovine bone [5]. In fact, although it was first identified for bone metabolism, it has been demonstrated to play a significant function in many biological and pathological processes such as inflammation, immune response, and cancer development along with studies over time [88]. In addition to these properties of OPN, it has also been associated with cell motility and cell survival pathways, affects cell migration, adhesion and differentiation, and thus has a critical importance in the repair and regeneration of tissues [89]. In recent years, OPN levels in tissues and serum have been studied for diagnosis and prognosis of various diseases, and studies have been conducted on the potential biomarker properties [30,31,32,33,35,36]. OPN is a versatile protein involved in the differentiation of Th1 and Th17 in the remodeling of tissue in inflammation. It was aimed to use OPN to determine the diagnosis and prognosis value of the disease in interstitial lung diseases (ILDs). In this study, which involved 344 ILD patients, approximately 70% of the patients were men, and 66.7% had a smoking history. The level of OPN in serum was measured using the ELISA method and the expression of OPN RNA using qPCR. In addition, serum OPN amounts of 140 healthy people were analyzed in this study. It was indicated that serum OPN levels of median ILD patients were very high in the interquartile range as well. There was no notable difference between serum OPN and RNA expression in different ILD diagnoses [30]. Hepatocellular carcinoma (HCC) is one of the most prevalent malignant tumors commonly found in Egypt. This disease, which has a poor prognosis and survival rate, is one of the diseases that are difficult to treat. For this reason, early diagnosis of HCC disease was very important for patients. It was evaluated whether OPN secreted by T cells, osteoblasts and macrophages could be a potential marker in HCC patients with hepatitis C virus (HCV). 140 patients were included in this study, their ages were between 27-79. 4 groups were formed. The first of these groups was a healthy control group with 20 people, the second group has 40 patients with HCV, the third group has 40 patients with HCC, and there are 40 patients with HCV and HCC in the fourth group. Western blotting method and sandwich ELISA were used. The OPN level was measured as 0.01189 ± 0.0075 mg/L for the first group, 0.04734 ± 0.00271 mg/L for the second group, 0.201 ± 0.00585 mg/L for the third group and finally 0.25152 ± 0.01963 mg/L for the fourth group. As a result, it was shown that OPN levels were seriously high in HCC patients. In other words, they could be distinguished compared to people who did not have HCC. The use of OPN for HCC disease is very important for early diagnosis and survival monitoring. CA.CA19.9 and AFP are also biomarkers used in diagnosis of cancer, but OPN was found to have a higher sensitivity and specificity than other biomarkers. In the early diagnosis of HCC, OPN is a potential biomarker [31]. The objective of a 2020 study was to assess the bioactivity of bmOPN using knockout mouse models. In the study, the effect of bmOPN on intestinal growth, immune response, and brain development was observed. Also, wild-type pups were breastfed by WT dams, KO dams, or KO dams with bmOPN supplementation for 1–21 days. As a result, it was found that oral bmOPN was not susceptible to gastrointestinal digestion in vivo. It has similar effects to mouse milk (mmOPN); these effects were shown to help the growth of the small intestine, as shown by histological analysis of duodenal villi height and crypt depth at P10. It alters the TNF-α response to an LPS challenge at p30; and helps the myelination of the brain by improving cognitive development and production of myelin-associated glycoprotein and myelin basic protein. Although mmOPN has a different amino acid sequence, it has a similar bioactivity to bmOPN, so it is thought that adding bmOPN to the formula may also be effective in human infants [32]. It was found that OPN is very important in many biological processes, such as atheromatosis and vascular calcification (VC), after many studies were looked at. Furthermore, research reveals that OPN contributes to pro-inflammatory processes in atherosclerotic cardiovascular disease (ASCVD) and intricately interacts with VC. From a clinical point of view, most studies support the idea that OPN is a biomarker in the presence, severity, and prognosis of coronary artery disease (CAD) [33]. In a study conducted in 2024, they investigated the relationship between OPN and post-COVID-19 symptoms, lung function and imaging abnormalities. The data of 122 COVID-19 patients and 181 visitors hospitalized were recorded at 4-84 weeks. ELISA was used to determine the circulating OPN. An increased level of circulating OPN in hospitalized patients has been associated with persistent COVID-19 symptoms. It was found that patients who showed symptoms had a higher level of OPN compared to patients who did not show symptoms. It has been found that patients with dyspnea (shortness of breath) have higher OPN levels compared to patients without. It was also shown that the OPN level was higher when patients had more than one symptom. m-MRC (modified- Medical Research Council) is used to score the level of shortness of breath. It was found that patients with a score greater than one had a higher OPN level than patients with a score between 0-1. In other words, the OPN level of patients with a high score is higher. As a result, it was found that increased OPN in the circulation of patients was associated with the deterioration of severe dyspnea and worsening of quality of life [35]. The researchers observed the effects of OPN on mild cognitive impairment (MCI), Alzheimer’s disease (AD) or pre-symptomatic AD. The cerebrospinal fluid (CSF) OPN level was measured to investigate AD and synaptic biomarkers. Two groups participated in the study, 167 people with cognitively unimpaired (CU) and 399 people with mild cognitive impairment (MCI) participated. These participants were selected from the PREVENT-AD project and the Alzheimer’s Disease Neuroimaging Initiation (ADNI) cohort. The examination of amyloid beta (Aβ) and tau proteins and changes in OPN levels were examined in the participants. Survival analyses were performed in the study. These analyses examined the relationship of conversion rates to Alzheimer’s disease with OPN levels. As a result, CSF OPN levels had a direct positive relationship with synaptic biomarkers. In addition, in the PREVENT-AD and ADNI studies, it was found that a positive amyloid beta (Aβ) 42/40 ratio and positive tau protein level were associated with Alzheimer’s disease [36].
Therefore, OPN is an important glycoprotein that is associated with many diseases and carries a biomarker characteristic that attracts attention with its increased levels in various pathological conditions. There is proof that the levels of OPN found in clinical trials show how the disease will progress and what the outlook is for it. This is especially true for long-term kidney diseases, interstitial lung diseases, coronary artery diseases, symptoms that come back after COVID-19, and more. Furthermore, predictions suggest that patients with HCC may have a unique advantage in the early detection of their diseases. The role of OPN as a potential biomarker is important for diagnosing the progression of diseases and evaluating patients' responses to treatment. The study of OPN levels in the future plays a significant function in the advancement of new treatment strategies. Future research will further develop and focus on the role of OPN in biomarker property.
4.2. Osteopontin and Cancer
One of the leading causes of death in the world is cancer. According to GLOBOCAN 2020, produced by the International Agency for Research on Cancer, approximately 19.3 million new cases and 10 million deaths have been recorded in 2020. It’s expected that there will be 28.4 million cases due to a 47% increase [90]. Although research on the tumor is progressing, it is difficult to develop anticancer drugs. For this reason, a better understanding of the molecules that affect tumor progression is necessary for the development of new treatment processes. This section examined the effect of OPN on cancer [45]. Numerous cells including tumor cells, macrophages, T cells, B cells, and dendritic cells produce the OPN protein. Several studies demonstrate the association of OPN with cancer, yet its functional contribution remains unproven [91]. Different receptors interact with OPN, which starts a number of different cancers signaling pathways that help cancer cells grow, migrate, invade, and stay alive. OPN also changes how immune cells behave around tumors, which could make the immune system less able to fight off tumors, which could lead to immunosuppression [92]. In a study that was performed on 346 gastric tumor samples to examine the expression of OPN, E-cadherin, β-cadherin, and cyclooxygenase 2. Immunohistochemistry facilitates these analyses. Peritoneal relapse, dispersed histotype, and elevated histological grade were associated with overexpression of OPN. According to the result, high expression of OPN is closely correlated with metastasis, recurrence, and survival in patients who have resected gastric cancer [78]. In another study, prostate tissue samples from 40 people with prostate cancer and 30 patients with benign prostate hyperplasia were examined. The expression of OPN splicing isoforms was studied in these prostate tissue samples. In this analysis, anti-OPN antibodies were examined by immunohistochemical staining test. Because of this study, it is thought that OPN may be useful in diagnosing and tracking prostate cancer because it is highly expressed in this type of cancer [93]. The study involved 33 patients with endometrioid endometrial cancer (EEC) and 30 patients with ovarian endometrioid cancer (OEC). The OPN levels of EEC and OEC cancer patients were analyzed and compared. Quantitative reverse transcriptase-polymerase chain reaction (RT-PCR) and quantitative PCR were performed in this study. As a result of the study, it was found that the release of OPN was less in early-stage cancer patients than in advanced-stage cancer patients [79]. OPN is a protein associated with hypoxia. In this research, plasma samples were taken from 172 patients, and they examined the relationship between the amount of OPN in their blood plasma (which is associated with hypoxia) and their health status. As a result of the study, without considering any treatment method, the survival rates of patients with low OPN levels are better than those with high amounts of OPN. This suggests that patients with low OPN levels have a higher chance of survival [81]. A recent study examined some aspects of the transcriptomic expression profile of SPP1, namely OPN. These are conditions where it is activated or dysregulated, biological pathways, and different mechanisms that change how it works. Tissues from breast, prostate, kidney, and skin cancer have been studied to see how differences in splicing patterns affect protein expression and function. As a result of the analyses performed, it was found that OPN was at a higher level than normal in most of the tissues. In addition, it has also been noticed that the expression of OPN is higher than normal, especially at the advanced tumor stage of kidney and skin cancer, and the application of treatment methods targeting the genes to which OPN is associated has led to a decrease in its effect on these types of cancer. The fact that the treatment methods can improve patients' chances of survival and their response to treatment by starting up immune systems in these types of cancer is one of the most important facts [80].
Nowadays, cancer still causes morbidity and mortality. For this reason, the availability of specific biomarkers is vital for the development of early detection and treatment methods. In this context, the OPN protein involved in the pathogenesis and progression of cancer may take a very important place [45]. Increased OPN expression in various types of cancer has an important role in cancer progression. Furthermore, researchers observed a significant relationship between the level of OPN and patient survival. Considering all this information, researchers have observed that OPN has the potential to serve as a biomarker. Thanks to future studies, early diagnosis and treatment methods using OPN can be improved and will allow us to better understand the relationship of OPN with cancer [80].
4.3. Osteopontin and Gut Health
OPN has multifunctional effects on intestinal development, which is a strong cytokine. Many investigations have shown that the endogenous expression of OPN in intestinal epithelial cells impacts intestinal barriers within both healthy and sick tissues. Endogenous production of OPN in a healthy gut suggests that the intestine affects immune homeostasis [94]. The level of inflammation correlates with the low expression of OPN. The response of mucosal T cells to OPN differs in healthy or sick individuals. As in Chorn disease, OPN increases the pro-inflammatory cytokine response due to insufficient IL-10 production. However, an increase in the amount of IL-10 is effective in maintaining homeostasis in healthy people [95]. In a study on the subject, the reason for the differences in developmental processes between formula-fed and breast-fed newborns is thought to be due to the reduced milk OPN content of the formula. The impact of OPN on intestinal health has been the subject of several studies. Prematurely born piglets served as the model for the study. On the eighth day, the OPN group had higher monocyte and lymphocyte counts and higher villus-crypt ratios than the raw milk group. However, intestinal functions were similar between both groups [83]. Another study examined the impacts of milk OPN on improving intestinal role in pregnant rats fed a high-fat diet (HFD). Pregnant rats were given bmOPN supplement at a dose of 6 mg/kg body weight. Thanks to this reinforcement, bmOPN supplementation reduced colon inflammation in female rats fed with HFD. Adding bmOPN made more of the proteins ZO-1 (zona occludens-1) and claudin-4 that help keep the intestines closed. These proteins are significant molecules that provide the wholeness and role of the intestinal barrier. BmOPN obtained some changes in the intestinal microbiome of rats; especially, it increased the relative abundance of the Bacteroidetes while decreasing the abundance of bacterial species such as Proteobacteria, Helicobacteraceae, and Desulfovibrionaceae. Short-chain fatty acid levels were higher in rats that were fed a high-fat diet (HFD) that also got bmOPN supplements than in rats that did not get bmOPN supplements. These fatty acids play an essential role in intestinal health and combating inflammation. Adding bmOPN may have changed four important biochemical processes in the microbiome of pregnant rats that were fed a high-fat diet. One of these processes was the production of bile acid. Intercalary, thanks to bmOPN supplementation, increased the level of several liver-sourced bile acids. These bile acids are taurochenodeoxycholic acid, tauroursodeoxycholic acid, and taurohyodeoxycholic acid [84]. In a separate study, researchers exposed 96 piglets to four distinct dietary treatments. These treatments are standard milk (CTR) for the first group; probiotics and prebiotics (galacto-oligosaccharides 4.36 g/L and human milk oligosaccharides 0.54 g/L) together with CTR for the second group; OPN (0.43 g/L) with CTR for the third group; and finally, the group that consumes prebiotics, OPN with CTR, which is the control group. The researchers recorded the piglets' data for a period of 15 days. The results demonstrated that adding synbiotic OPN to piglet milk formulas or OPN alone may change the formation of the intestinal microbiota, the development of digestion, and most importantly, the attachment response of the newborn. Thanks to these supplements, diarrhea attacks have decreased in puppies, beneficial microbiota types have been promoted at least slightly, and dysbiotic species have decreased. Most importantly, OPN supplements increase the concentration of SCFA, decrease the level of ammonia, and decrease the intraepithelial lymphocyte count [85]. There are also many studies on OPN's effectiveness in digestive system inflammation. In this study, DSS-induced acute colitis in OPN-null mice has higher tissue damage and lower TNF-a production. Additionally, OPN-null mice exhibit abnormal cell death. These cell deaths (necroptosis) arise randomly. OPN is a cytokine that has an important function in regulating intestinal inflammation and controlling tissue damage [96].
In summary, OPN deficiency may cause microbiome damage and increased inflammation. OPN has many benefits as it affects the gut microbiome. More research is needed to fully understand the effects of OPN on gut health and gut disease treatment.
4.4. Osteopontin and Immunological Effects
OPN is referred to as an early T lymphocyte activation 1 protein (ETA-1) that occurs when T cells are active [8]. There are a lot of immune processes that OPN is linked to, including inflammatory conditions such as cancer, allergic reactions, tissue damage, and autoimmune diseases such as MS and rheumatoid arthritis [55]. Numerous body fluids, tissues, and cells, including macrophages and dendritic cells, express OPN [26,55].
Figure 3.
In different cell types, OPN expression and its effects. OPN receptors, integrins and CD44 are commonly synthesized on the surfaces of the cell types shown here analyzed in detail in the peripheral immune system, sOPN supports cell migration and effector functions such as cell proliferation, immune response and inflammatory processes are involved in influencing. Also, iOPN is a form of OPN found inside the cell and is involved in regulating immune responses and signal transmission. In macrophages, sOPN supports phogocytosis and cell migration, in dendritic cell, sOPN is involved in the migration of Th1, Th2 and Th17 cell. SOPN promotes migration degranulation in eosinophil, neutrophil and mast cell [55].
Figure 3.
In different cell types, OPN expression and its effects. OPN receptors, integrins and CD44 are commonly synthesized on the surfaces of the cell types shown here analyzed in detail in the peripheral immune system, sOPN supports cell migration and effector functions such as cell proliferation, immune response and inflammatory processes are involved in influencing. Also, iOPN is a form of OPN found inside the cell and is involved in regulating immune responses and signal transmission. In macrophages, sOPN supports phogocytosis and cell migration, in dendritic cell, sOPN is involved in the migration of Th1, Th2 and Th17 cell. SOPN promotes migration degranulation in eosinophil, neutrophil and mast cell [55].
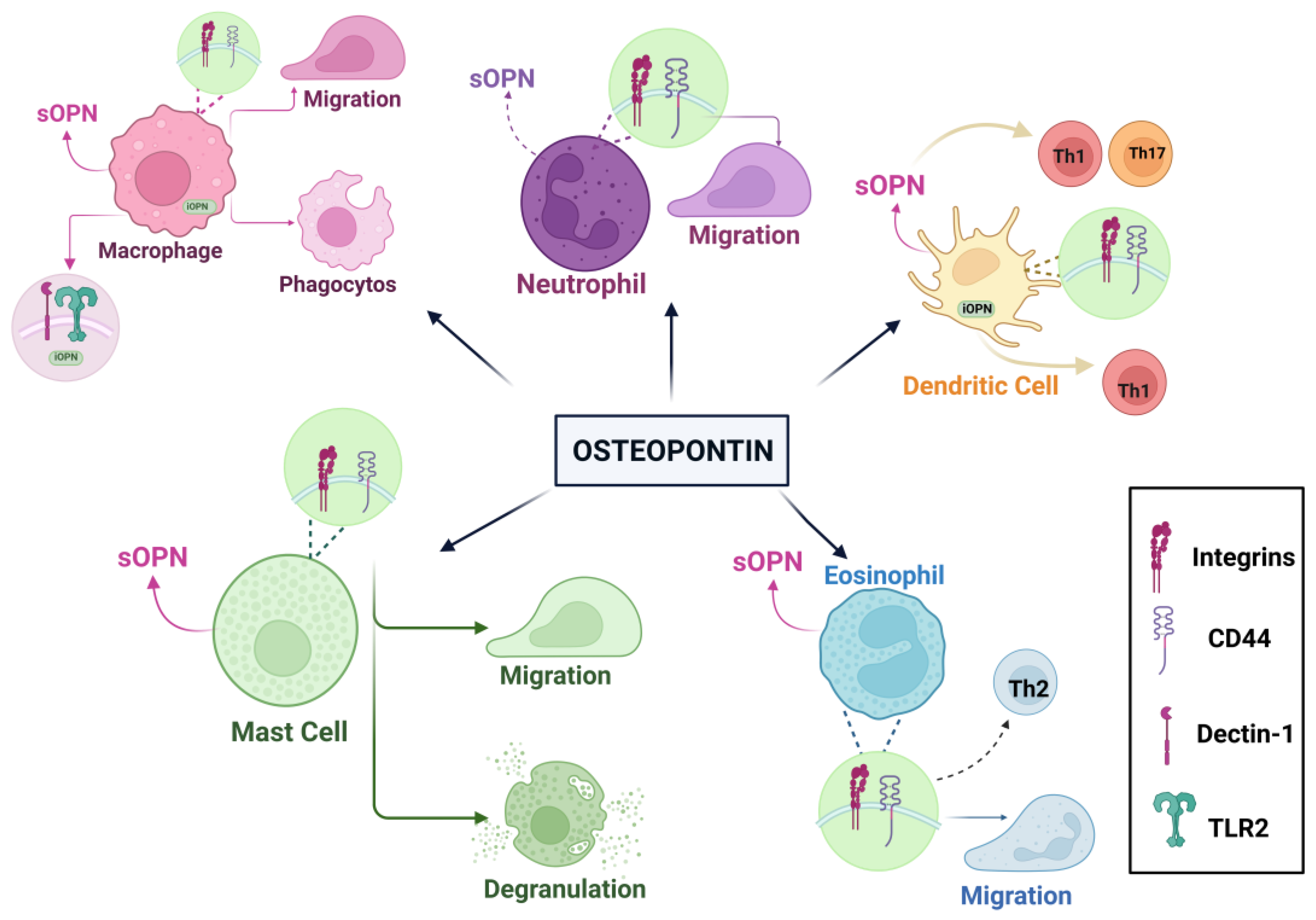
SOPN has many important functions in macrophages, such as cell migration, proliferation, phagocytosis, and MMP production. Thanks to sOPN, monocytes transform into macrophages via integrins. The iOPN strengthens and regulates immune responses in macrophages. It regulates antiviral responses in peritoneal macrophages. Dendritic cells produce sOPN, which enhances TH1 polarization by aiding in the production of TNF-a and IL-12. In addition, iOPN replicates IFN-I production while regulating Th17 responses in dendritic cells. It also facilitates interactions between MSC and dendritic cells, acting as a repressor of OPN protein in the presence of proinflammatory cytokines. Neutrophils, the most abundant leukocytes in the bloodstream, also produce OPN, albeit less than macrophages. Independent from CD44, sOPN increases the migration of neutrophils. Neutrophils have a disruptive duty against pathogenic microbes. However, in the process of fulfilling this role, neutrophils can occasionally cause harm to host cells. Eosinophils also produce OPN. This secretes OPN, which contributes to the formation of airway angiogenesis in asthma disease. One study used sputum samples from asthmatic patients. Researchers have revealed a relationship between the amounts of OPN and the eosinophils present in sputum. This study concluded that sOPN could serve as a biomarker for disease detection. Mast cells are located in mucosal areas. These cells carry large granules and contain heparin, histamine, proteases, and proinflammatory cytokines [55]. Another study analyzed the amounts of OPN in breast milk, bovine milk, and infant formula. These measurements revealed an average of 138 mg/L of OPN in breast milk. This measurement led to the conclusion that it represents 2.1% of the overall protein in breast milk (weight/weight). At the same time, the amounts of OPN in bovine milk and infant formula are approximately 18 mg/L and 9 mg/L. It has been found that OPN can increase the production of IL-2 and thus affect the immune response. Additionally, researchers analyzed the amounts of OPN in 3-month-old infants using plasma samples from both pregnant and non-pregnant women. In this study, it was found that the levels of OPN in plasma taken from 3-month-old babies and baby cords were 7–10 times higher. It’s considered that high OPN levels are very important for babies and can increase the production of cytokines in newborn babies [28]. In a study, interleukin-10-knockout (IL-10 KO) mice were used to look at the role of OPN in the development of intestinal inflammation. OPN/IL-10 and IL-10 were then compared. Researchers have observed a faster development of colitis in OPN/IL-10 mice. That is, not having enough OPN messes up the microbiota in the gut and the normal functions of macrophages, which makes inflammation in the gut worse [26]. Another study examined the relationship between B cell aggregates in the central nervous system and OPN. B cells in multiple sclerosis (MS) brain tissue express OPN. One of the most striking points of the findings is the different effects of OPN on B cells. One of them is that OPN directs down the expression of the CD80 and CD86 helper stimulating molecules and has the ability to reduce B cell activation. Another effect is that it can help B cells form clusters, which can increase B cells' activation. On the other hand, OPN has both anti-inflammatory and pro-inflammatory effects on autoimmune disease. It can stimulate the production of IL-6 and auto-antibodies by affecting B cells; this is due to the pro-inflammatory cytokine property of OPN. Among its anti-inflammatory properties. there is the potential to regulate tissue repair. In addition, OPN is associated not only with autoimmune diseases, but also with chronic neuroinflammation and has been associated with the clustering of B cells. Taking these findings into account, it is known that OPN interacts with many receptors thanks to its multifunction. These anti-inflammatory and pro-inflammatory properties that OPN has may have potential for different studies. More importantly, it may enable the identification of new treatment methods for chronic neuroinflammatory patients [27].
A clinical study investigated the immunological activity of OPN. There were three groups used in the research. These are standard formulas, 65 mg OPN/L (50% of the OPN concentration in human milk) or 130 mg OPN/L (equal to 100% of the OPN concentration in human milk), with milk containing 1, 4, and 6-month-old babies fed spontaneously, and the obtained data were compared. As a result of this comparison, it was found that formula-fed infants had higher levels of the pro-inflammatory cytokine TNF-α in their plasma than breast-fed infants. It was also found that TNF-α levels were lower in infants fed with foods with added OPN. More importantly, the way babies who are fed milk with OPN use amino acids and their cytokine responses is similar to how babies who are fed breast milk. One of the most important things to note is that babies who take OPN supplements have fewer feverish days (pyrexia) than babies who don't take any supplements. Additionally, babies fed milk containing 130 mg OPN/L have higher T cells compared to babies fed other food groups. These findings have determined that OPN also serves a significant function in immune development [29]. In another study, premature pigs were given OPN at a body weight of 46 mg/kg per day, while some pigs were given raw bovine milk, and the relationship between them was examined. Pigs exhibit similar immune statuses. However, higher levels of monocytes and lymphocytes were found in the intestines of OPN-fed pigs after the eighth day compared to pigs fed raw bovine milk [83].
As a result, looking at the studies and results, it was concluded that giving OPN to babies is good for their immune systems and is very important for finding new ways to treat some diseases.
4.5. Dental Health
During the formation process of a tooth, OPN is located in non-mineralized tissues and has a function that reduces or stops mineralization during the production of periodontal ligaments [97]. OPN, which is involved in the healthy development and repair of mineralized tissues, also helps to maintain tissue integrity by ensuring cell-matrix and matrix-matrix/mineral adhesion [98]. OPN is highly phosphorylated sialoprotein and serves a noteworthy function in the mineralized extracellular matrices of teeth. Also OPN has been shown to play an important role in the fluoride remineralization process. In a study conducted, the effects of bacterial enamel demineralization of OPN on remineralization were investigated. Mine demineralization was designed at a depth of 180-200 µm and exposed to Streptococcus mutans for 21 days. Calcium phosphate solution containing 0.499 mg/L OPN was used. As a result, remineralization with OPN occurred at the same time, fluoride acted only on the surface in the absence of OPN. In addition, this condition can stabilize the fluoride ions of OPN [39]. In another study, the aim was to focus on the relationship between hydroxyapatite surfaces and OPN and to prevent bacteria from adhering to the tooth surface. The result indicated that OPN binds stably to the hydroxyapatite surface but decreases in the event of rinsing. Fewer streptococci colonies have been detected in areas with OPN [40]. In another study, the binding properties of bacteria to saliva-coated surfaces of OPN were examined. Actinomyces naeslundii, Lactobacillus paracasei subsp. paracasei and Streptococcus mitis bacteria were used in study. It has been found that OPN is the most effective protein that inhibits adhesion [38]. In summary, OPN is a potential protein for dental health. It has been determined that the use of this protein may be effective in the future, in the treatment of periodontal diseases or in the removal of caries and prevention of dental biofilms.
4.6. Osteopontin and Brain development- Cognitive Function
OPN, a pro-inflammatory cytokine, is expressed in many cells and tissues. For example, activated macrophages and T–lymphocytes. It is known that OPN is among the components of the extracellular matrix of the central nervous system (CNS). It is also known that the level of OPN increases in case of inflammation and injury that may occur in the central nervous system [99]. It has been shown that OPN has an assignment in multiple sclerosis [100,101], Parkinson’s disease [102], Alzheimer’s disease [36,103], and many neurodegenerative diseases. In the central nervous system, it is microglia into macrophage-like cells located intermittently. These microglia show resistance to damage occurring in the CNS, but also help in restoring and maintaining the integrity of the CNS. In addition, it promotes the progression of neurodegenerative disease [104]. In one study, research was conducted on bmOPN cognitive development and its effects on the brain. During the study, 20 male pigs were given two different diets for up to 34 days after birth. These are a standard soy proteins isolated-based milk powder substitute and an OPN-supplemented dietary form. Magnetic resonance imaging (MRI) technique and a new object recognition task were used. The MRI technique was used to analyze brain development, while the new object recognition task evaluated learning and memory characteristics. Pigs fed an OPN-supplemented diet reacted to the object they saw in a shorter time. and it was found that relative brain volumes increased in the brain regions of the pigs in this group. In addition, the study found that regular consumption of OPN is safe in pig models [41]. In another study, OPN, which has the property of protecting nerve cells and brain tissue in ischemic brain injuries, is neuroprotective in hypoxia ischemic (HI) conditions and research has been conducted on whether damage to neurological functions can be corrected. The right carotid artery of the rat pups was blocked and blood flow to one side of the brain was prevented, and then brain injury was performed by placing them in a low-oxygen environment. The effect of OPN supplementation on HI was investigated in a study. As a result, it was seen that OPN decreases with increasing age, endogenous OPN occurs again. It was shown that endogenous OPN reaches its highest level at the 48th hour, and this happens to protect itself after brain damage that occurs in the rats’ body. In addition, exogenous OPN has been shown to reduce the volume of the damaged brain and eliminate neurological consequences. More importantly, OPN acted as an apoptosis inhibitor by suppressing caspase 3. It has improved neurological functions in long-term treatments [42]. In another study, the effect of OPN on sectional spinal cord injury (SCI) was investigated. It has been observed that the level of OPN increases significantly during the formation of SCI. SCI mice were used in the study. the level of OPN and the positions of the cells formed in the spinal cord were examined. In addition, the effect of OPN level on neuropathic pain and movement ability was investigated. As a result of the study, it was shown that the protein whose OPN level increased the most after SCI. As a result of suppression of the OPN level, it was found that the ability to move worsened and pain increased. In other words, it has been proven that OPN contributes to the healing process, supports the recovery of motor functions, and can suppress the pain that occurs [43]. Other study, the effects of OPN on ischemic stroke were investigated. As a result of the research, it was shown that OPN reduces cerebral damage and supports recovery [105]. In one study, the effect of consuming diets supplemented with bmOPN on brain development was investigated. As a result, there was no effect of OPN supplementation on cognitive development [83]. In a study, the effects of milk-derived OPN on brain development were examined. 2 groups were formed, and mice were used in this study. One group was fed milk containing OPN, another group was fed milk without OPN. As a result of the analyses performed, it has been shown that the OPN taken from milk is transported to the brain. It was noticed that the amount of OPN directly increased in the brains of mice fed with OPN-supplemented milk. It was found that cognitive development was worse in mice fed milk without OPN supplementation [106]. Considering all the studies, OPN to be used as a supplement has been shown to affect cognitive development and behavior and affect memory development. It can be used to support the formation of new cells and for therapeutic purposes in cases where the brain is damaged.
5. Nutritional Potential of Osteopontin
Newborn babies need breast milk to protect against infections in the first period of their lives. Milk is rich in nutrients and bioactive components [28]. It is very important for babies to be fed with breast milk in the first 6 months after birth, when the immune system has not yet developed [107]. Currently, it has been found that infants fed with formula contract more infections than infants fed breast milk [108]. This is due to the lack of certain proteins with antibacterial, immunostimulating and protective properties contained in enough milk in the food consumed by formula-fed babies. Breast milk is produced to meet all the needs of a baby due to its structure and content. However, infants who cannot be fed with breast milk are fed with formula. For this reason, many studies are trying to enrich the contents of formula, to make it similar to breast milk and to meet the necessary needs of babies in this way. One of the important proteins that may be involved in the immune development of infants is OPN. It is a multifunctional glycoprotein that plays a role in many biological and physiological processes. The concentration of OPN in milk has the highest concentration compared to other tissues and body fluids [28].
Figure 4.
Nutritional Potential of Osteopontin.
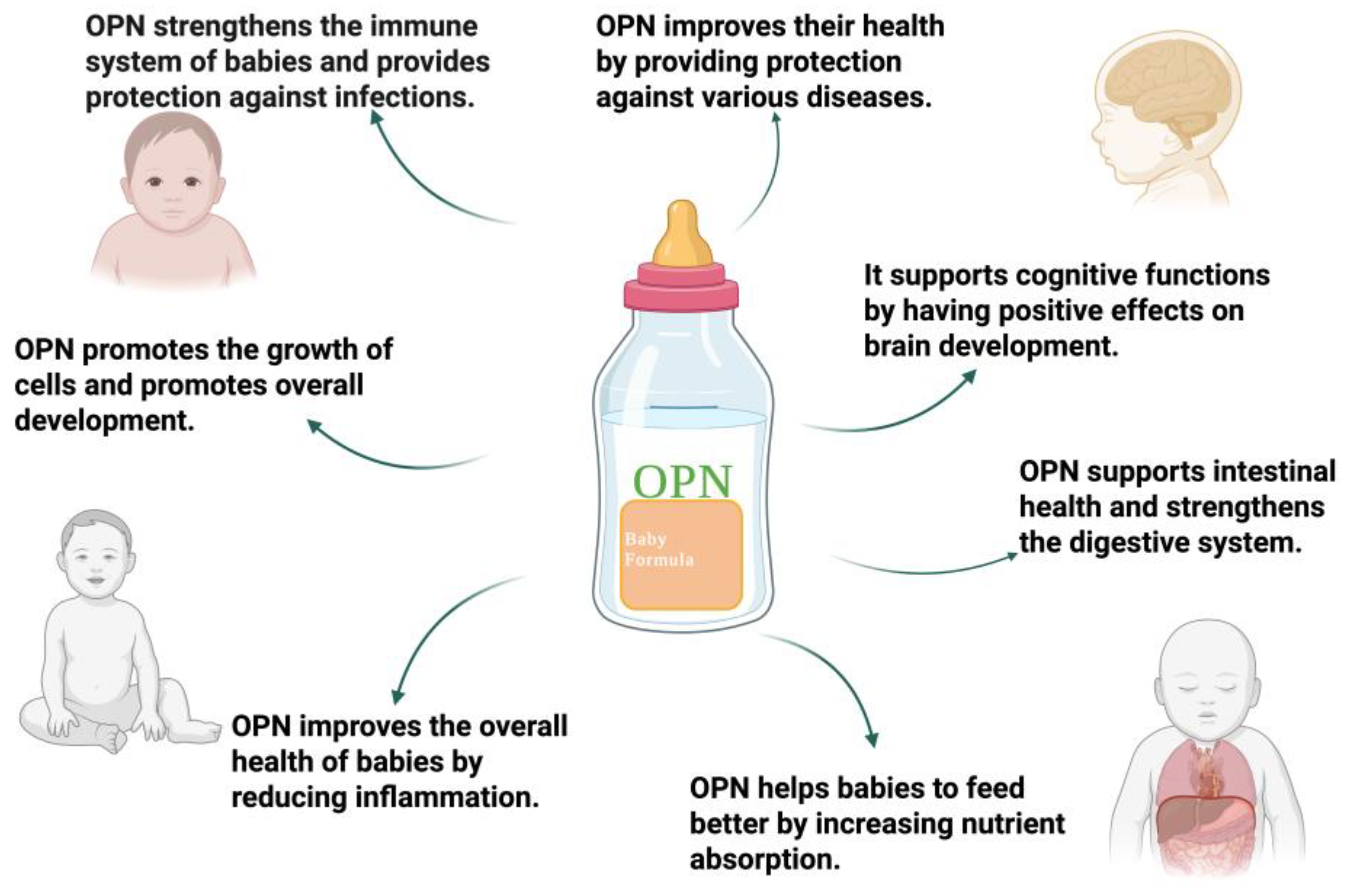
The amount of OPN in bovine milk, breast milk, and infant formula was analyzed. As a result of these analysis, OPN concentrations of approximately 138 mg/L in human milk, 18 mg/L in bovine milk, and 9 mg/L in infant formula were determined. This means that the amount of OPN consumed by formula-fed babies is quite low [28]. It has been shown that significant differences in gene expression occur in the intestines of breast-fed monkey babies. At the same time, it was noticed that half of the gene expression babies who tried formulas with bmOPN was similar to monkey babies who were fed breast milk. That is, OPN consumption has been shown to positively affect the intestines of infants. In addition, the regulation of cell migration of OPN, its interactions with integrins also support these results [86]. High plasma levels of newborn babies indicate that OPN is of critical importance for the baby. As in the case of a baby monkey, OPN consumption can support neonatal intestinal immunity and stimulate cytokine production. OPN affects the high monocyte and lymphocyte levels of pigs consuming OPN and the deeper villi-crypt level. This has a positive effect on the immune and intestinal of OPN consumption [83]. Also, the use of OPN supplements by women during pregnancy limited the occurrence of colon inflammation, encouraged the support of the intestinal barrier. In addition, it’s present in the intestinal flora, supports the microbiota population by inducing the production of SCFA, and promotes the synthesis of bile acid. In this case, OPN improves intestinal health [84]. Some of the milk proteins, for example OPN, are resistant to digestive enzymes, and mobile proteins that are biologically active have been identified [109]. OPN consumption is the protective effect of OPN on its ability to regulate and protect the structural and functional integrity of the stomach, intestines, as well as reducing liver inflammation [110]. Additionally, another study, Lacprodan® OPN-10 was found to be non-mutagenic as a result of bacterial reverse mutation testing, non-clastogenic as a result of chromosomal aberration tests, and clean genotoxicity according to in vivo micronucleus testing. In addition, according to the 13-week subchronic toxicity study, 1208 mg/kg body weight was determined in male mice and 1272 mg/kg body weight /day in females. These values are the highest safe dose amounts that can be used for mice. Also, the 2.50 g/kg Lacprodan® OPN-10 applied on pregnant females did not have a negative effect on the pregnancy rate and fetal development. As a result, the amount of Lacprodan® OPN-10 found suitable for a breastfed baby in this study is an average of 25 mg/kg body weight. More importantly, it has been stated that even 35 times more than the recommended amount is nor harmful. The result of this study has also shown that OPN consumption is safe for infants [111].
Given these results, it has been determined that OPN has important roles in supporting human health, especially in regulating the growth development and immune responses of infants. In particular, feeding babies who cannot be fed with breast milk with OPN supplemented foods is very critical for the development of babies. As a result, OPN is a potential protein that can be used in infant formula, but more studies are needed before it can be used.
6. Conclusions and Future Remarks
OPN is a versatile phosphoglycoprotein that plays a role in many biological and pathological processes. OPN is known to be involved in many biological processes such as infection-tissue regeneration, cell adhesion, immune system support, and fetal development. From another point of view, the idea that OPN concentration can be used as a potential biomarker that can be used in the management of inflammation, especially in the early diagnosis and treatment of cancer, has been supported. Also, the amount of OPN present in the milk of people living in different geographical regions varies. In addition, studies show that OPN concentration begins to decrease after childbirth, and more importantly, the amount of OPN present in mothers’ milk varies depending on maternal factors. However, more studies are needed to determine the effect of different geographical regions, maternal factors and other factors on OPN concentration. OPN is partially resistant to digestion by intestinal proteases, and thanks to this, it was found that it crosses the intestinal barrier and participates in circulation, and thus supports physiological durations. In addition, in studies conducted using OPN supplementation, the level of deep villi-crypt in the intestines of infants fed with OPN supplementation increased. In this way, it has supported the development of intestinal health, gene transcription, and the gut microbiome in infants, and OPN has been shown to affect cognitive development and behavior in infants. In the future, the use of OPN in cancer treatment may help in the development of new treatment strategies. In addition, the use of OPN as a potential biomarker for early diagnosis of cancer patients in the future is being investigated. Furthermore, OPN has important roles in the development of intestinal flora and regulation of intestinal inflammation. In other words, the use of OPN-fortified products in the future may be very important for intestinal health. Especially in infants who cannot be fed with breast milk, the consumption of OPN-fortified formula in the future is very critical for the improvement of intestinal health of infants and the development of mental functions. Since it has important tasks in diagnosing many neurological diseases, reducing neurological damage, improving neurological function and developing nerve cells, it will allow new treatment methods to emerge in the future.
As a result, OPN is a multifunctional protein whose new properties are detected every passing day, and its importance is increasing day by day. OPN can be an important goal in the development of new treatment strategies, as well as help to take an important revolutionary step in the treatment of various diseases in the future.
Author Contributions
Conceptualization, S.K., M.B.; writing—original draft preparation, B.K., H.D.; writing—review and editing, S.K., M.B., H.D., B.K, visualization, B.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kläning, E.; Christensen, B.; Sørensen, E.S.; Vorup-Jensen, T.; Jensen, J.K. Osteopontin Binds Multiple Calcium Ions with High Affinity and Independently of Phosphorylation Status. Bone 2014, 66, 90–95. [Google Scholar] [CrossRef]
- Senger, D.F.; Wirth, D.F.; Hynes, R. 0 Transformed Mammalian Cells Secrete Specific Proteins and Phosphoproteins. Cell 1979, 16, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Franzitn, A.; Heinegard, D. Isolation and Characterization of Two Sialoproteins Present Only in Bone Calcified Matrix. Biochem. J 1985, 232, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Oldberg, A.; Franzfn, A.; Heinegard, D. Cloning and Sequence Analysis of Rat Bone Sialoprotein (Osteopontin) CDNA Reveals an Arg-Gly-Asp Cell-Binding Sequence (Cell Adhesion/Bone Matrix Proteins/Glycosylation). Biochemistry 1986, 83, 8819–8823. [Google Scholar]
- Sørensen, E.S.; Christensen, B. Milk Osteopontin and Human Health. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Wrana, J.L.; Zhang, Q.; Sodek, J. Nucleic Acids Research Full Length CDNA Sequence of Porcine Secreted Phosphoprotein-I (SPP-I, Osteopontin). 17, 1989.
- Craig, A.M.; Denhardt, D.T. The Murine Gene Encoding Secreted Phosphoprotein 1 (Osteopontin): Promoter Structure, Activity, and Induction in Vivo by Estrogen and Progesterone (Promoter Analysis; Gene Structure; Bone Sialoprotein; Transformation-Associated Phosphoprotein; Early T-Lymphocyte Activation 1; Tumor Promoter Induction). Gene 1991, 100, 163–171. [Google Scholar]
- Patarca, R.; Freeman, G.J.; Singh, R.P.; Wei, F.Y.; Durfee, T.; Blattner, F.; Regnier, D.C.; Kozak, C.A.; Mock, B.A.; Morse, H.C.; et al. Structural and Functional Studies of the Early T Lymphocyte Activation 1 (Eta-1) Gene. Definition of a Novel T Cell-Dependent Response Associated with Genetic Resistance to Bacterial Infection. Journal of Experimental Medicine 1989, 170. [Google Scholar] [CrossRef]
- Craig, A.M.; Nemir, M.; Mukherjee, B.B.; Chambers, A.F.; Denhardt, D.T. Identification of the Major Phosphoprotein Secreted by Many Rodent Cell Lines as 2AR/Osteopontin: Enhanced Expression in H-RAS-Transformed 3T3 Cells. Biochem Biophys Res Commun 1988, 157. [Google Scholar] [CrossRef]
- Jia, Q.; Ouyang, Y.; Yang, Y.; Yao, S.; Chen, X.; Hu, Z. Osteopontin: A Novel Therapeutic Target for Respiratory Diseases. Lung 2024, 202, 25–39. [Google Scholar] [CrossRef]
- Clemente, N.; Raineri, D.; Cappellano, G.; Boggio, E.; Favero, F.; Soluri, M.F.; Dianzani, C.; Comi, C.; Dianzani, U.; Chiocchetti, A. Osteopontin Bridging Innate and Adaptive Immunity in Autoimmune Diseases. J Immunol Res 2016, 2016. [Google Scholar] [CrossRef]
- Wai, P.Y.; Kuo, P.C. The Role of Osteopontin in Tumor Metastasis. Journal of Surgical Research 2004, 121, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, T.; Matsuda, K.; Uchibori, T.; Sugano, M.; Uehara, T.; Honda, T. Upregulation of Osteopontin Expression via the Interaction of Macrophages and Fibroblasts under IL-1b Stimulation. Cytokine 2018, 110, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Attur, M.G.; Dave, M.N.; Stuchin, S.; Kowalski, A.J.; Steiner, G.; Abramson, S.B.; Denhardt, D.T.; Amin, A.R. Osteopontin: An Intrinsic Inhibitor of Inflammation in Cartilage. Arthritis Rheum 2001, 44, 578–584. [Google Scholar] [CrossRef]
- Hwang, S.-M.; Wilson, P.D.; Laskin, J.D.; Denmardt, D.T. Age and Development-Related Changes in Osteopontin and Nitric Oxide Synthase MRNA Levels in Human Kidney Proximal Tubule Epithelial Cells: Contrasting Responses to Hypoxia and Reoxygenation. J Cell Physiol 1994, 160, 6148. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.M.; Lopez, C.A.; Heck, D.E.; Gardner, C.R.; Laskin, D.L.; Laskin, J.D.; Denhardt, D.T. Tm JOURNAL OF B I O ~ I C A L C m m y Osteopontin Inhibits Induction of Nitric Oxide Synthase Gene Expression by Inflammatory Mediators in Mouse Kidney Epithelial Cells*. 1994, 269, 711–715.
- Scott, J.A.; Lynn Weir, M.; Wilson, S.M.; Xuan, J.W.; Chambers, A.F.; Mccormack, D.G.; Burton, A.C.; Weir, M.L. Osteopontin Inhibits Inducible Nitric Oxide Synthase Activity in Rat Vascular Tissue. 1998.
- Denhardt, D.T.; Guo, X. Osteopontin: A Protein with Diverse Functions. The FASEB Journal 1993, 7. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Noda, M. Osteopontin Expression and Function: Role in Bone Remodeling. J Cell Biochem 1998, 72, 92–102. [Google Scholar] [CrossRef]
- O’Regan, A.; Berman, J.S. Osteopontin: A Key Cytokine in Cell-Mediated and Granulomatous Inflammation. Int J Exp Pathol 2000, 81, 373–390. [Google Scholar] [CrossRef]
- Weber, G.F. The Metastasis Gene Osteopontin: A Candidate Target for Cancer Therapy.
- Tenen, D.G.; Senger, D.R.; Perruzzi, C.A.; Gracey, C.F.; Papadopoulos, A. Secreted Phosphoproteins Associated with Neoplastic Transformation: Close Homology with Plasma Proteins Cleaved during Blood Coagulation. Cancer Res 1988, 48. [Google Scholar]
- Senger, D.R.; Perruzzi, C.A.; Papadopoulos, A.; Tenen, D.G. Purification of a Human Milk Protein Closely Similar to Tumor-Secreted Phosphoproteins and Osteopontin. Biochimica et Biophysica Acta (BBA)/Protein Structure and Molecular 1989, 996. [Google Scholar] [CrossRef]
- Bautista, D.S.; Denstedt, J.; Chambers, A.F.; Harris, J.F. Low-Molecular-Weight Variants of Osteopontin Generated by Serine Proteinases in Urine of Patients with Kidney Stones. J Cell Biochem 1996, 61, 402–409. [Google Scholar] [CrossRef]
- Moorman, H.R.; Poschel, D.; Klement, J.D.; Lu, C.; Redd, P.S.; Liu, K. Osteopontin: A Key Regulator of Tumor Progression and Immunomodulation. Cancers (Basel) 2020, 12, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Toyonaga, T.; Nakase, H.; Ueno, S.; Matsuura, M.; Yoshino, T.; Honzawa, Y.; Itou, A.; Namba, K.; Minami, N.; Yamada, S.; et al. Osteopontin Deficiency Accelerates Spontaneous Colitis in Mice with Disrupted Gut Microbiota and Macrophage Phagocytic Activity. PLoS One 2015, 10. [Google Scholar] [CrossRef]
- Chunder, R.; Schropp, V.; Marzin, M.; Amor, S.; Kuerten, S. A Dual Role of Osteopontin in Modifying B Cell Responses. Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Schack, L.; Lange, A.; Kelsen, J.; Agnholt, J.; Christensen, B.; Petersen, T.E.; Sørensen, E.S. Considerable Variation in the Concentration of Osteopontin in Human Milk, Bovine Milk, and Infant Formulas. J Dairy Sci 2009, 92, 5378–5385. [Google Scholar] [CrossRef] [PubMed]
- West, C.E.; Kvistgaard, A.S.; Peerson, J.M.; Donovan, S.M.; Peng, Y.M.; Lönnerdal, B. Effects of Osteopontin-Enriched Formula on Lymphocyte Subsets in the First 6 Months of Life: A Randomized Controlled Trial. Pediatr Res 2017, 82, 63–71. [Google Scholar] [CrossRef]
- Iturbe-Fernández, D.; Pulito-Cueto, V.; Mora-Cuesta, V.M.; Remuzgo-Martínez, S.; Ferrer-Pargada, D.J.; Genre, F.; Alonso-Lecue, P.; López-Mejías, R.; Atienza-Mateo, B.; González-Gay, M.A.; et al. Osteopontin as a Biomarker in Interstitial Lung Diseases. Biomedicines 2024, 12. [Google Scholar] [CrossRef]
- Othman, O.A.; Qasem, A.; Taha, G.; Amer, H. Evaluation of Osteopontin as a Potential Biomarker in Hepatocellular Carcinoma in a Sample of Egyptian Patients. Egyptian Journal of Cancer and Biomedical Research 2024, 8, 47–53. [Google Scholar] [CrossRef]
- Jiang, R.; Lönnerdal, B. Evaluation of Bioactivities of Bovine Milk Osteopontin Using a Knockout Mouse Model. J Pediatr Gastroenterol Nutr 2020, 71, 125–131. [Google Scholar] [CrossRef]
- Kadoglou, N.P.E.; Khattab, E.; Velidakis, N.; Gkougkoudi, E. The Role of Osteopontin in Atherosclerosis and Its Clinical Manifestations (Atherosclerotic Cardiovascular Diseases)—A Narrative Review. Biomedicines 2023, 11. [Google Scholar] [CrossRef]
- Günaydın, S.; Özlü, T.; Özsu, S.S.; Örem, A. Role of Osteopontin and NGAL in Differential Diagnosis of Acute Exacerbations of COPD and Pneumonia. Medical Science and Discovery 2024, 11, 150–156. [Google Scholar] [CrossRef]
- Pappas, A.G.; Eleftheriou, K.; Vlahakos, V.; Magkouta, S.F.; Riba, T.; Dede, K.; Siampani, R.; Kompogiorgas, S.; Polydora, E.; Papalampidou, A.; et al. High Plasma Osteopontin Levels Are Associated with Serious Post-Acute-COVID-19-Related Dyspnea. J Clin Med 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Quesnel, M.J.; Labonté, A.; Picard, C.; Bowie, D.C.; Zetterberg, H.; Blennow, K.; Brinkmalm, A.; Villeneuve, S.; Poirier, J. Osteopontin: A Novel Marker of Pre-Symptomatic Sporadic Alzheimer’s Disease. Alzheimer’s and Dementia 2024. [CrossRef]
- Huang, Y.; Lu, Z.; Liu, F.; Lane, J.; Chen, J.; Fu, X.; Huang, Q.; Hu, R.; Zhang, B. Osteopontin Enhances the Probiotic Viability of Bifidobacteria in Pectin-Based Microencapsulation Subjected to in Vitro Infant Gastrointestinal Digestion. Food Hydrocoll 2024, 149. [Google Scholar] [CrossRef]
- Kristensen, M.F.; Sørensen, E.S.; Del Rey, Y.C.; Schlafer, S. Prevention of Initial Bacterial Attachment by Osteopontin and Other Bioactive Milk Proteins. Biomedicines 2022, 10. [Google Scholar] [CrossRef]
- Nurrohman, H.; Carter, L.; Barnes, N.; Zehra, S.; Singh, V.; Tao, J.; Marshall, S.J.; Marshall, G.W. The Role of Process-Directing Agents on Enamel Lesion Remineralization: Fluoride Boosters. Biomimetics 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- Burling, H.S.S.B.H.J.A.G.G. Use of Osteopontin in Dental Formulations 2005.
- Joung, S.; Fil, J.E.; Heckmann, A.B.; Kvistgaard, A.S.; Dilger, R.N. Early-Life Supplementation of Bovine Milk Osteopontin Supports Neurodevelopment and Influences Exploratory Behavior. Nutrients 2020, 12, 1–20. [Google Scholar] [CrossRef]
- Chen, W.; Ma, Q.; Suzuki, H.; Hartman, R.; Tang, J.; Zhang, J.H. Osteopontin Reduced Hypoxia-Ischemia Neonatal Brain Injury by Suppression of Apoptosis in a Rat Pup Model. Stroke 2011, 42, 764–769. [Google Scholar] [CrossRef]
- Weng, Y.; Lu, F.; Li, P.; Jian, Y.; Xu, J.; Zhong, T.; Guo, Q.; Yang, Y. Osteopontin Promotes Angiogenesis in the Spinal Cord and Exerts a Protective Role Against Motor Function Impairment and Neuropathic Pain After Spinal Cord Injury. Spine (Phila Pa 1976) 2024, 49. [Google Scholar] [CrossRef]
- Sodek, J.; Ganss, B.; McKee M., D. OSTEOPONTIN. Critical Reviews in Oral Biology & Medicine 2000, 11, 279–303. [Google Scholar]
- Icer, M.A.; Gezmen-Karadag, M. The Multiple Functions and Mechanisms of Osteopontin. Clin Biochem 2018, 59. [Google Scholar] [CrossRef]
- Butler, W.T. The Nature and Significance of Osteopontin. Connect Tissue Res 1989, 23. [Google Scholar] [CrossRef]
- Bayless, K.J.; Davis, G.E. Identification of Dual A4β1 Integrin Binding Sites within a 38 Amino Acid Domain in the N-Terminal Thrombin Fragment of Human Osteopontin. Journal of Biological Chemistry 2001, 276, 13483–13489. [Google Scholar] [CrossRef]
- Tan, Y.; Zhao, L.; Yang, Y.G.; Liu, W. The Role of Osteopontin in Tumor Progression Through Tumor-Associated Macrophages. Front Oncol 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Standal, T.; Borset, M.; Sundan, A. Role of Osteopontin in Adhesion, Migration, Cell Survival and Bone Remodeling. Exp Oncol 2004, 26. [Google Scholar]
- Yokosaki, Y.; Matsuura, N.; Sasaki, T.; Murakami, I.; Schneider, H.; Higashiyama, S.; Saitoh, Y.; Yamakido, M.; Taooka, Y.; Sheppard, D. The Integrin A9β1 Binds to a Novel Recognition Sequence (SVVYGLR) in the Thrombin-Cleaved Amino-Terminal Fragment of Osteopontin. Journal of Biological Chemistry 1999, 274, 36328–36334. [Google Scholar] [CrossRef]
- Christensen, B.; Nielsen, M.S.; Haselmann, K.F.; Petersen, T.E.; Sørensen, E.S. Post-Translationally Modified Residues of Native Human Osteopontin Are Located in Clusters: Identification of 36 Phosphorylation and Five O-Glycosylation Sites and Their Biological Implications. Biochemical Journal 2005, 390, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Tagliabracci, V.S.; Wiley, S.E.; Guo, X.; Kinch, L.N.; Durrant, E.; Wen, J.; Xiao, J.; Cui, J.; Nguyen, K.B.; Engel, J.L.; et al. A Single Kinase Generates the Majority of the Secreted Phosphoproteome. Cell 2015, 161, 1619–1632. [Google Scholar] [CrossRef]
- Kothari, A.N.; Arffa, M.L.; Chang, V.; Blackwell, R.H.; Syn, W.K.; Zhang, J.; Mi, Z.; Kuo, P.C. Osteopontin—a Master Regulator of Epithelial-Mesenchymal Transition. J Clin Med 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Shou, P.; Zhang, L.; Xu, C.; Zheng, C.; Han, Y.; Li, W.; Huang, Y.; Zhang, X.; Shao, C.; et al. An Osteopontin-Integrin Interaction Plays a Critical Role in Directing Adipogenesis and Osteogenesis by Mesenchymal Stem Cells. Stem Cells 2014, 32, 327–337. [Google Scholar] [CrossRef]
- Lin, E.Y.H.; Xi, W.; Aggarwal, N.; Shinohara, M.L. Osteopontin (OPN)/SPP1: From Its Biochemistry to Biological Functions in the Innate Immune System and the Central Nervous System (CNS). Int Immunol 2023, 35. [Google Scholar] [CrossRef]
- Weber, G.F. The Phylogeny of Osteopontin—Analysis of the Protein Sequence. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Sørensen, E.S.; Petersen, T.E.; Højrup, P. Posttranslational Modifications of Bovine Osteopontin: Identification of Twenty-eight Phosphorylation and Three O-glycosylation Sites. Protein Science 1995, 4, 2040–2049. [Google Scholar] [CrossRef] [PubMed]
- Kazanecki, C.C.; Uzwiak, D.J.; Denhardt, D.T. Control of Osteopontin Signaling and Function by Post-Translational Phosphorylation and Protein Folding. J Cell Biochem 2007, 102, 912–924. [Google Scholar] [CrossRef] [PubMed]
- Kon, S.; Ikesue, M.; Kimura, C.; Aoki, M.; Nakayama, Y.; Saito, Y.; Kurotaki, D.; Diao, H.; Matsui, Y.; Segawa, T.; et al. Syndecan-4 Protects against Osteopontin-Mediated Acute Hepatic Injury by Masking Functional Domains of Osteopontin. Journal of Experimental Medicine 2008, 205, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.L.; Kim, H.-J.; Kim, J.-H.; Garcia, V.A.; Cantor, H. Alternative Translation of Osteopontin Generates Intracellular and Secreted Isoforms That Mediate Distinct Biological Activities in Dendritic Cells. 2008.
- Christensen, B.; Karlsen, N.J.; Jørgensen, S.D.S.; Jacobsen, L.N.; Ostenfeld, M.S.; Petersen, S. V.; Müllertz, A.; Sørensen, E.S. Milk Osteopontin Retains Integrin-Binding Activity after in Vitro Gastrointestinal Transit. J Dairy Sci 2020, 103, 42–51. [Google Scholar] [CrossRef]
- Schytte, G.N.; Christensen, B.; Bregenov, I.; Kjøge, K.; Scavenius, C.; Petersen, S. V.; Enghild, J.J.; Sørensen, E.S. FAM20C Phosphorylation of the RGDSVVYGLR Motif in Osteopontin Inhibits Interaction with the Avβ3 Integrin. J Cell Biochem 2020, 121, 4809–4818. [Google Scholar] [CrossRef]
- WHO Evidence for the Ten Steps to Successful Breastfeeding. 1998.
- WHO Protecting, Promoting and Supporting Breast-Feeding: The Special Role of Maternity Services. A Joint WHO/UNICEF Statement. Geneva: World Health Organization; 1989; 2017;
- Aksan, A.; Erdal, I.; Yalcin, S.S.; Stein, J.; Samur, G. Osteopontin Levels in Human Milk Are Related to Maternal Nutrition and Infant Health and Growth. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Bezirtzoglou, E.; Tsiotsias, A.; Welling, G.W. Microbiota Profile in Feces of Breast- and Formula-Fed Newborns by Using Fluorescence in Situ Hybridization (FISH). Anaerobe 2011, 17, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Another Reason to Favor Exclusive Breastfeeding: Microbiome Resilience. J Pediatr (Rio J) 2018, 94. [Google Scholar] [CrossRef]
- Hanson, L.Å.; Korotkova, M. The Role of Breastfeeding in Prevention of Neonatal Infection. Seminars in Neonatology 2002, 7, 275–281. [Google Scholar] [CrossRef]
- Lee, H.; Park, H.; Ha, E.; Hong, Y.C.; Ha, M.; Park, H.; Kim, B.N.; Lee, B.; Lee, S.J.; Lee, K.Y.; et al. Effect of Breastfeeding Duration on Cognitive Development in Infants: 3-Year Follow-up Study. J Korean Med Sci 2016, 31, 579–584. [Google Scholar] [CrossRef]
- Demmelmair, H.; Prell, C.; Timby, N.; Lönnerdal, B. Benefits of Lactoferrin, Osteopontin and Milk Fat Globule Membranes for Infants. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Tang, Q.; Zhao, X.; Zhang, Y.; Zhao, X.; Xiang, Y.; Geng, W.; Feng, Y.; Cai, W. The Levels of Osteopontin in Human Milk of Chinese Mothers and Its Associations with Maternal Body Composition. Food Science and Human Wellness 2022, 11, 1419–1427. [Google Scholar] [CrossRef]
- Jiang, R.; Lönnerdal, B. Osteopontin in Human Milk and Infant Formula Affects Infant Plasma Osteopontin Concentrations. Pediatr Res 2019, 85, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Goonatilleke, E.; Huang, J.; Xu, G.; Wu, L.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Human Milk Proteins and Their Glycosylation Exhibit Quantitative Dynamic Variations during Lactation. Journal of Nutrition 2019, 149, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, A.; Lai, S.; Yuan, Q.; Jia, X.; Wang, P.; Zhang, Y. Longitudinal Changes in the Concentration of Major Human Milk Proteins in the First Six Months of Lactation and Their Effects on Infant Growth. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Zhu, J.; Yu, X.; Wang, Y.; Bai, S.; Lai, J.; Tong, X.; Xing, Y. Longitudinal Changes of Lactopontin (Milk Osteopontin) in Term and Preterm Human Milk. Front Nutr 2022, 9. [Google Scholar] [CrossRef]
- Joung, K.E.; Christou, H.; Park, K.H.; Mantzoros, C.S. Cord Blood Levels of Osteopontin as a Phenotype Marker of Gestational Age and Neonatal Morbidities. Obesity 2014, 22, 1317–1324. [Google Scholar] [CrossRef]
- Takahashi, T.; Ueno, H.M.; Yamaide, F.; Nakano, T.; Shiko, Y.; Kawasaki, Y.; Mitsuishi, C.; Shimojo, N. Comparison of 30 Cytokines in Human Breast Milk between 1989 and 2013 in Japan. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Di Bartolomeo, M.; Pietrantonio, F.; Pellegrinelli, A.; Martinetti, A.; Mariani, L.; Daidone, M.G.; Bajetta, E.; Pelosi, G.; de Braud, F.; Floriani, I.; et al. Osteopontin, E-Cadherin, and β-Catenin Expression as Prognostic Biomarkers in Patients with Radically Resected Gastric Cancer. Gastric Cancer 2016, 19, 412–420. [Google Scholar] [CrossRef]
- Hashiguchi, Y.; Tsuda, H.; Bandera, C.A.; Nishimura, S.; Inoue, T.; Kawamura, N.; Berkowitz, R.S.; Mok, S.C. Comparison of Osteopontin Expression in Endometrioid Endometrial Cancer and Ovarian Endometrioid Cancer. Medical Oncology 2006, 23. [Google Scholar] [CrossRef]
- Kundu, G.; Elangovan, S. Investigating the Role of Osteopontin (OPN) in the Progression of Breast, Prostate, Renal and Skin Cancers. Biomedicines 2025, 13. [Google Scholar] [CrossRef]
- Mack, P.C.; Redman, M.W.; Chansky, K.; Williamson, S.K.; Farneth, N.C.; Lara, P.N.; Franklin, W.A.; Le, Q.T.; Crowley, J.J.; Gandara, D.R. Lower Osteopontin Plasma Levels Are Associated with Superior Outcomes in Advanced Non-Small-Cell Lung Cancer Patients Receiving Platinum-Based Chemotherapy: SWOG Study S0003. Journal of Clinical Oncology 2008, 26. [Google Scholar] [CrossRef]
- Das, S.; Song, Z.; Han, H.; Ge, X.; Desert, R.; Athavale, D.; Babu Komakula, S.S.; Magdaleno, F.; Chen, W.; Lantvit, D.; et al. Intestinal Osteopontin Protects From Alcohol-Induced Liver Injury by Preserving the Gut Microbiome and the Intestinal Barrier Function. CMGH 2022, 14, 813–839. [Google Scholar] [CrossRef] [PubMed]
- Aasmul-Olsen, K.; Henriksen, N.L.; Nguyen, D.N.; Heckmann, A.B.; Thymann, T.; Sangild, P.T.; Bering, S.B. Milk Osteopontin for Gut, Immunity and Brain Development in Preterm Pigs. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Han, L.; Li, Q.; Du, M.; Mao, X. Bovine Milk Osteopontin Improved Intestinal Health of Pregnant Rats Fed a High-Fat Diet through Improving Bile Acid Metabolism. J Dairy Sci 2024, 107, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Ferreres-Serafini, L.; Martín-Orúe, S.M.; Sadurní, M.; Jiménez, J.; Moreno-Muñoz, J.A.; Castillejos, L. Supplementing Infant Milk Formula with a Multi-Strain Synbiotic and Osteopontin Enhances Colonic Microbial Colonization and Modifies Jejunal Gene Expression in Lactating Piglets. Food Funct 2024, 15, 6536–6552. [Google Scholar] [CrossRef] [PubMed]
- Donovan, S.M.; Monaco, M.H.; Drnevich, J.; Kvistgaard, A.S.; Hernell, O.; Lönnerdal, B. Bovine Osteopontin Modifies the Intestinal Transcriptome of Formula-Fed Infant Rhesus Monkeys to Be More Similar to Those That Were Breastfed. Journal of Nutrition 2014, 144, 1910–1919. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, P.; Gong, L.; Zhang, X.; Ling, Z.; Bi, K.; Shi, F.; Wang, K.; Zhang, Q.; Jiang, J.; et al. Osteopontin Exacerbates High-Fat Diet-Induced Metabolic Disorders in a Microbiome-Dependent Manner. mBio 2022, 13. [Google Scholar] [CrossRef]
- Layton, G.R.; Antoun, I.; Copperwheat, A.; Khan, Z.L.; Bhandari, S.S.; Somani, R.; Ng, A.; Zakkar, M. Osteopontin as a Biomarker for Coronary Artery Disease. Cells 2025, 14. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Noda, M.; O’Regan, A.W.; Pavlin, D.; Berman, J.S. Osteopontin as a Means to Cope with Environmental Insults: Regulation of Inflammation, Tissue Remodeling, and Cell Survival. Journal of Clinical Investigation 2001, 107, 1055–1061. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Furger, K.A.; Menon, R.K.; Tuck, A.B.; Bramwell, V.H.; Chambers, A.F. The Functional and Clinical Roles of Osteopontin in Cancer and Metastasis. Curr Mol Med 2001, 1. [Google Scholar]
- Alpana Kumari, D.K.V.K.G. Osteopontin in Cancer. In Advances in Clinical Chemistry; Elsevier, 2024; Vol. 118, pp. 87–110.
- Tilli, T.M.; Thuler, L.C.; Matos, A.R.; Coutinho-Camillo, C.M.; Soares, F.A.; da Silva, E.A.; Neves, A.F.; Goulart, L.R.; Gimba, E.R. Expression Analysis of Osteopontin MRNA Splice Variants in Prostate Cancer and Benign Prostatic Hyperplasia. Exp Mol Pathol 2012, 92, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Gassler, N.; Autschbach, F.; Gauer, S.; Bohn, J.; Sido, B.; Otto, H.F.; Geiger, H.; Obermüller, N. Expression of Osteopontin (Eta-1) in Crohn Disease of the Terminal Ileum. Scand J Gastroenterol 2002, 37, 1286–1295. [Google Scholar] [CrossRef] [PubMed]
- Agnholt, J.; Kelsen, J.; Schack, L.; Hvas, C.L.; Dahlerup, J.F.; Sørensen, E.S. Osteopontin, a Protein with Cytokine-like Properties, Is Associated with Inflammation in Crohn’s Disease. Scand J Immunol 2007, 65, 453–460. [Google Scholar] [CrossRef]
- Da Silva, A.P.B.; Pollett, A.; Rittling, S.R.; Denhardt, D.T.; Sodek, J.; Zohar, R. Exacerbated Tissue Destruction in DSS-Induced Acute Colitis of OPN-Null Mice Is Associated with Downregulation of TNF-α Expression and Non-Programmed Cell Death. J Cell Physiol 2006, 208, 629–639. [Google Scholar] [CrossRef]
- Macneil, R.L.; Berry, J.; D’errico, J.; Strayhorn, C.; Piotrowski, B.; Somerman, M.J. Role of Two Mineral-Associated Adhesion Molecules, Osteopontin and Bone Sialoprotein, during Cementogenesis. Connect Tissue Res 1995, 33, 1–7. [Google Scholar] [CrossRef]
- McKee, M.D.; Nanci, A. Osteopontin at Mineralized Tissue Interfaces in Bone, Teeth, and Osseointegrated Implants: Ultrastructural Distribution and Implications for Mineralized Tissue Formation, Turnover, and Repair. Microsc Res Tech 1996, 33, 141–164. [Google Scholar] [CrossRef]
- Yu, H.; Liu, X.; Zhong, Y. The Effect of Osteopontin on Microglia. Biomed Res Int 2017, 2017. [Google Scholar] [CrossRef]
- Stampanoni Bassi, M.; Buttari, F.; Gilio, L.; Iezzi, E.; Galifi, G.; Carbone, F.; Micillo, T.; Dolcetti, E.; Azzolini, F.; Bruno, A.; et al. Osteopontin Is Associated with Multiple Sclerosis Relapses. Biomedicines 2023, 11. [Google Scholar] [CrossRef]
- Chiocchetti, A.; Comi, C.; Indelicato, M.; Castelli, L.; Mesturini, R.; Bensi, T.; Mazzarino, M.C.; Giordano, M.; D’Alfonso, S.; Momigliano-Richiardi, P.; et al. Osteopontin Gene Haplotypes Correlate with Multiple Sclerosis Development and Progression. J Neuroimmunol 2005, 163, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhou, M.; Dai, W.; Guo, W.; Qiu, J.; Zhang, Z.; Mo, M.; Ding, L.; Ye, P.; Wu, Y.; et al. Bone-Derived Factors as Potential Biomarkers for Parkinson’s Disease. Front Aging Neurosci 2021, 13. [Google Scholar] [CrossRef]
- Wirths, O.; Breyhan, H.; Marcello, A.; Cotel, M.C.; Brück, W.; Bayer, T.A. Inflammatory Changes Are Tightly Associated with Neurodegeneration in the Brain and Spinal Cord of the APP/PS1KI Mouse Model of Alzheimer’s Disease. Neurobiol Aging 2010, 31, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Gehrmann, J.; Matsumoto, Y.; Kreutzberg, G.W. BRAIN RESEARCH REVIEWS Full-Length Review Microglia: Intrinsic Immuneffector Cell of the Brain. Brain Res Rev 1995, 20, 269–287. [Google Scholar] [CrossRef]
- Zhu, Q.; Luo, X.; Zhang, J.; Liu, Y.; Luo, H.; Huang, Q.; Cheng, Y.; Xie, Z. Osteopontin as a Potential Therapeutic Target for Ischemic Stroke. Curr Drug Deliv 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Prell, C.; Lönnerdal, B. Milk Osteopontin Promotes Brain Development by Up-Regulating Osteopontin in the Brain in Early Life. FASEB Journal 2019, 33, 1681–1694. [Google Scholar] [CrossRef]
- Mosca, F.; Giannì, M.L. Human Milk: Composition and Health Benefits. Pediatria Medica e Chirurgica 2017, 39. [Google Scholar] [CrossRef]
- Dewey, K.G.; Heinig, M.J.; Nommsen-Rivers, L.A. Differences in Morbidity between Breast-Fed and Formula-Fed Infants. 1995.
- Chatterton, D.E.W.; Rasmussen, J.T.; Heegaard, C.W.; Sørensen, E.S.; Petersen, T.E. In Vitro Digestion of Novel Milk Protein Ingredients for Use in Infant Formulas: Research on Biological Functions. In Proceedings of the Trends in Food Science and Technology; July 2004; Vol. 15, pp. 373–383.
- Ge, X.; Lu, Y.; Leung, T.-M.; Sørensen, E.S.; Nieto, N. Milk Osteopontin, a Nutritional Approach to Prevent Alcohol-Induced Liver Injury. American Journal of Physiology-Gastrointestinal and Liver Physiology 2013, 304, 929–939. [Google Scholar] [CrossRef]
- Kvistgaard, A.S.; Matulka, R.A.; Dolan, L.C.; Ramanujam, K.S. Pre-Clinical in Vitro and in Vivo Safety Evaluation of Bovine Whey Derived Osteopontin, Lacprodan® OPN-10. Food and Chemical Toxicology 2014, 73, 59–70. [Google Scholar] [CrossRef]
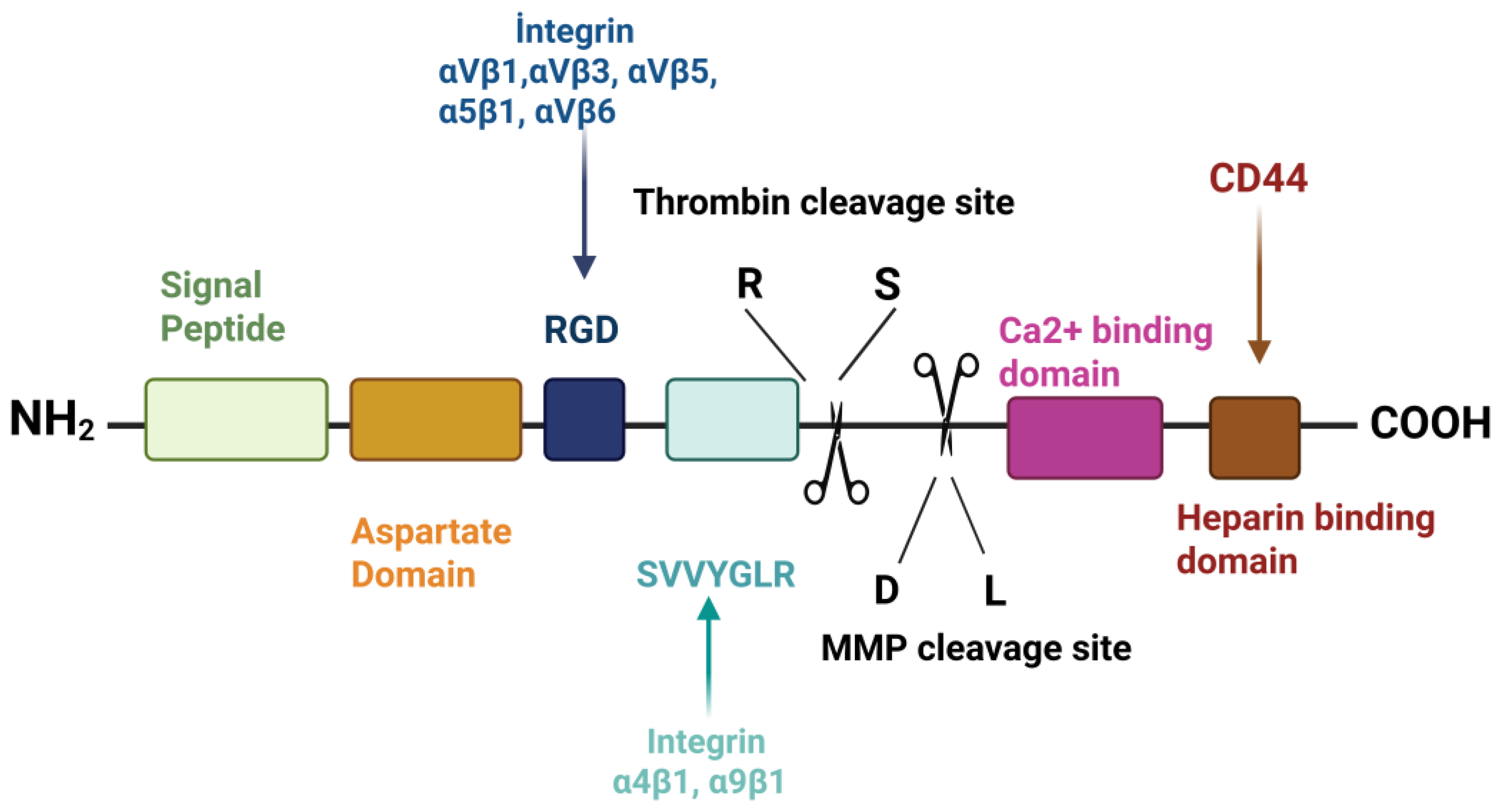
Figure 2.
Functions of Osteopontin [26,27,28,29,30,31,32,33,34,35,36,38,39,40,41,42,43,78,79,80,81,82,83,84,85,86,87].
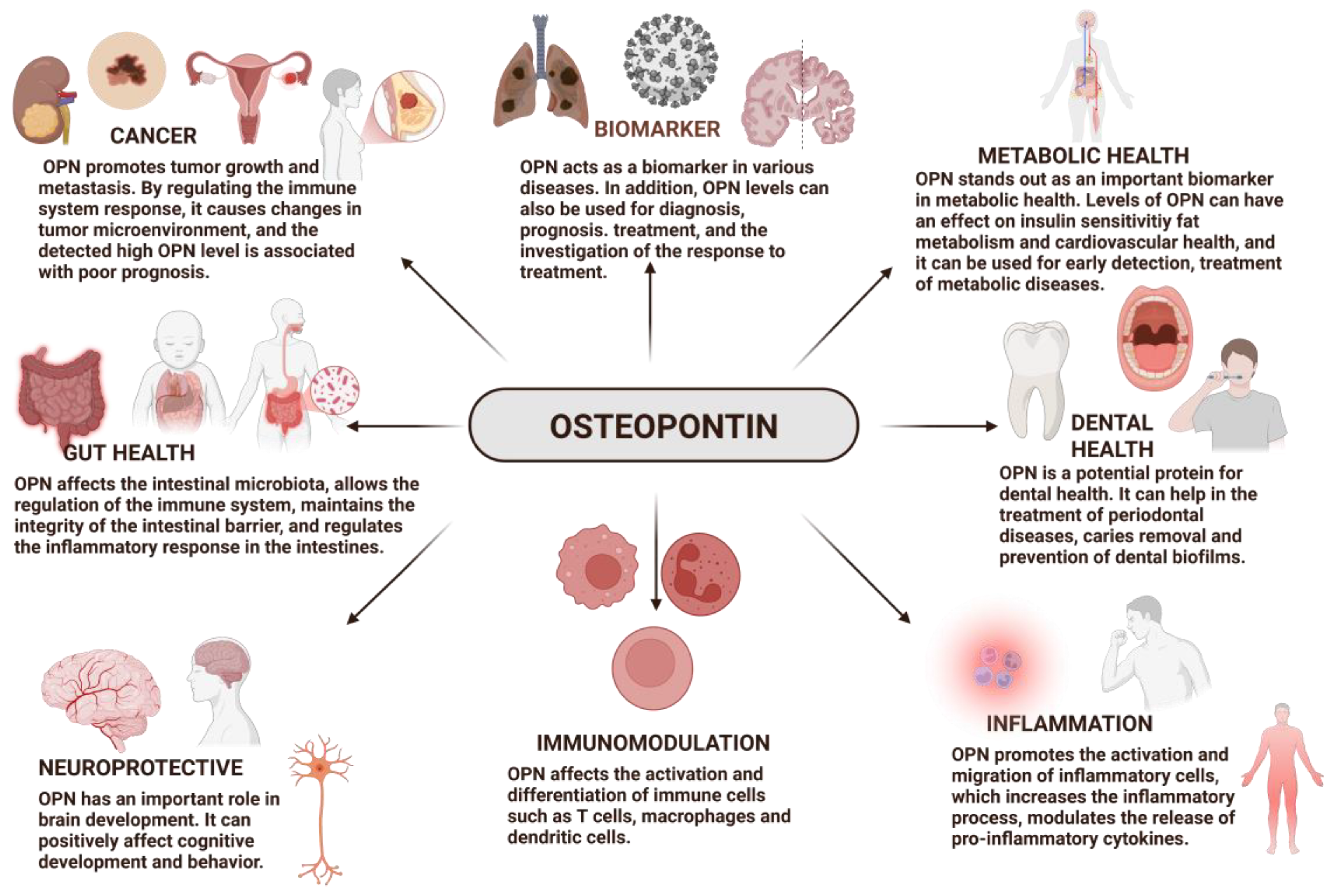

Table 1.
Reported Effects of Osteopontin in In vitro and In vivo Studies.
| Function | Dose | Effect | Study design | Ref |
|---|---|---|---|---|
| Neuroprotective | The OPN-supplemented formula was designed to use 250 mg/L OPN. | Dietary supplementation of bmOPN enhanced the relative volume of several brain regions and altered behaviors in the novel object recognition task. | In vivo | [41] |
| Neuroprotective | OPN (0.03 μg or 0.1 μg) was administered intracerebroventricularly at 1-hour post-HI. | Exogenous OPN reduced infarct volume and augmented neurological outcomes 7 weeks after HI injury. OPN-induced neuroprotection was inhibited by an integrin antagonist. | In vivo | [42] |
| Neuroprotective | - | The possible advantages of OPN increase in improving locomotor function and alleviating neuropathic pain subsequent to a spinal cord injury. | In vivo | [43] |
| Gut Health | - | It was found that OPN deficiency worsens alcohol-related disease (ALD). In addition, it has been shown that increasing the amount of OPN in intestinal epithelial cells may be beneficial for maintaining the intestinal microbiome and protecting intestinal barrier function and to destroy ALD. | In vivo | [82] |
| Gut, Immunity and Brain Development | 16 premature pigs fed diets with 46 mg/(kg.g) of OPN added. | It has been found that pigs fed with OPN supplementation have greater villus-crypt depth and a higher number of monocytes and lymphocytes. The T-maze test turned out to be similar in all groups, so their cognitive development is similar. In addition, improvements in intestinal structure and immunity have been observed thanks to OPN supplementation. | In vivo | [83] |
| Gut Health | BmOPN was enhanced at a dose of 6 mg/kg body weight. | It has been noticed that use of OPN supplements by pregnant women during pregnancy prevents colon inflammation and increases the function of the intestinal barrier. The intestinal population has been regulated by SCFA production. Bovine OPN provides support for the secretion of bile acid. | In vivo | [84] |
| Gut health and Immune function | 0.436 g/L OPN was added to the milk formulation. | It has been proven to improve digestion, intestinal microbiota, and the newborn immune system in piglets administered OPN. Synbiotic tablets enhance healthy bacteria breeds in piglets' intestines. Supports reducing hazardous genera. It also reduced diarrhea in OPN or synbiotic-fed piglets. Most notably, an increase in SCFA in offspring feeding OPN and the synbiotic formula has reduced ammonia, which prevents pathogen growth. | In vivo | [85] |
| Gut Health | Formula fed supplemented with 25 mg/L bovine OPN. | Newborn rhesus monkeys fed breast milk or OPN-supplemented bovine milk grow similarly. Different nutritional groups express genes differently. Additionally, monkeys administered bovine OPN have similar gene expression to those fed breast milk. OPN may affect newborn rhesus monkey intestines. | In vivo | [86] |
| Biomarker and Bioactivity | 12µg/g OPN was used as a supplement every morning. | BmOPN has been found to improve the growth of the small intestine, inhibit lipopolysaccharide-induce TNF-α secretion, increase the amount of brain myelination and support cognitive development. | In vivo | [32] |
| Biomarker | - | It has been found that OPN is associated with atherosclerotic cardiovascular diseases (ASCVD) and vascular calcifications (VC) and plays a pro-inflammatory role. It is known that OPN has different effects on various diseases. In addition, OPN level may be a potential biomarker in patients with coronary artery disease. | In vitro and In Vivo | [33] |
| Biomarker | - | It has been evaluated that the examination of OPN levels is important in chronic obstructive pulmonary disease (COPD) and pneumonia and that OPN may show a potential biomarker property. | In vivo | [34] |
| Biomarker | - | It was noticed that patients hospitalized due to COVID-19 had increased amounts of OPN in their circulation, and these patients showed severe symptoms. Thanks to this finding, it has been thought that OPN may be a biomarker. | In vitro | [35] |
| Biomarker | - | This study examined the diagnosis and prognosis of interstitial lung disorders (ILDs) using OPN levels. According to the study, ILD patients had 0.00105 mg/L OPN, while healthy people had 0.00081 mg/L. OPN levels are inversely linked to required vital capacity and high in ILD patients who died or had lung transplants. Based on these findings, increased OPN levels may help identify ILD patients. | In vivo | [36] |
| Biomarker | - | This study examined the diagnosis and prognosis of interstitial lung disorders (ILDs) using OPN levels. According to the study, ILD patients had 0.00105 mg/L OPN, while healthy people had 0.00081 mg/L. OPN levels are inversely linked to required vital capacity and high in ILD patients who died or had lung transplants. Based on these findings, increased OPN levels may help identify ILD patients. | In vivo | [30] |
| Biomarker | - | In Egypt, biomarker was considered to help diagnose hepatocellular carcinoma (HCC). The study examined a potential biomarker of OPN concentration in HCV patients. According to the study, OPN is more effective and sensitive than other markers. This suggests that OPN can be employed as a biomarker for early illness diagnosis. | In vivo | [31] |
| Dental Health | - | OPN is effective process-directing agents utilized in the PILP process to stabilize and convey mineral ions and the regulate remineralization had the potential to boost the performance of fluoride. | In vitro | [39] |
| Dental Health | Although 46 µM OPN was used in previous studies, the amount of OPN used in this study is 50 µM, as the desired optimal effect was not observed. | Different milk proteins have been researched to inhibit bacteria adhesion to saliva-coated surfaces. OPN prevents adhesion best among milk proteins. These proteins may improve oral health and prevent dental biofilm. | In vitro | [38] |
| Dental Health | The amount of OPN determined for use in the study is in the range of 100 mg/kg to 1000 mg/kg. But the amount of OPN that is considered suitable for use the most is approximately 350 mg/kg. | It has been found that OPN could bind to hydroxyapatite surfaces, as well as the ability to prevent bacteria from adhering to the tooth surface. | In vitro | [40] |
| Immunomodulation | - | A study on OPN's role in inflammatory intestinal inflammation indicated that colitis progressed quicker in OPN/IL-10 DKO mice than in IL-10 KO animals. Thus, OPN deficiency has advanced colitis. OPN expression is also elevated in IL-10 KO mouse epithelial cells. In OPN/IL-10 DKO mice, Clostridium subset XIVa was less abundant and cluster XVIII was more abundant. | In vivo | [26] |
| Immunomodulation | 2µg/well (1µg/mL) recombinant OPN was added to B cells. | OPN was detected in MS brain tissue B cell clusters and reduces B cells by diminishing CD80 and CD86 helper stimulus molecule expression. OPN also increases B cell clustering, which is linked to persistent neuroinflammation. OPN boosts IL-6 and autoantibodies. IL-6 was down-regulated and IL-10 up-regulated in B cells treated with human recombinant OPN (rOPN). | In vitro | [27] |
| Immunomodulation | - | OPN plasma from 3-month-old babies' umbilical cords is 7-10 times higher than normal. Infant formula's low OPN concentration is insufficient for infants' intestinal immune systems. For this reason, newborn cells are unlikely to express enough cytokines. | In vitro | [28] |
| Immunomodulation | Milk formulas with 65 mg OPN/L or 130 mg OPN/L added were used. | The amount of T cells and monocytes in the circulations of infants fed with (formula 130) increased. It is also thought to have a possible role in the development and progress of immunity in infants. | In vivo | [29] |
| Cancer | - | Cancer can result from high OPN levels. It also promotes tumor growth and metastasis, which can spread cancer. Gastric cancer patients' survival can be predicted by OPN levels. | In vitro | [78] |
| Cancer | - | In particular, the OPN levels of EEC (endometrioid endometrial cancer) and OEC (ovarian endometrioid cancer) cancer types are similar to each other. It was found that the OPN level was higher in advanced tumors. | In vitro | [79] |
| Cancer | - | It was found that OPN expression was high in breast, prostate, kidney and skin cancers. It was noticed that the amount of OPN was even higher in advanced kidney and skin cancers than in other types of cancer. When treatment targets for these types of cancer were applied, the level of OPN decreased. | In vitro | [80] |
| Cancer | - | Patients with strong OPN-expression had a shorter lifespan. OPN levels before chemotherapy are linked to survival, disease development after treatment, and treatment responses in non-small cell lung cancer (NSCLC) patients. | In vivo | [81] |
| Metabolic Health | - | Identify OPN's role in HFD-induced lipid deposition. OPN-deficient (KO) mice may tolerate HFD-induced lipid accumulation and metabolic circumstances. HFD-fed mice's intestinal lipid and fatty acid metabolism is actively regulated by OPN. By altering the gut flora, OPN deficiency may prevent HFD-induced dyslipidemia. | In vitro – in vivo | [87] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



