
Submitted:
03 May 2025
Posted:
06 May 2025
You are already at the latest version
Abstract
The presence of various inhibitory compounds in lignocellulosic hydrolysates presents a significant challenge for bioethanol production, requiring yeasts with exceptional multistress tolerance. This study investigated the nonconventional yeast Saccharomycodes ludwigii APRE2 as a potential solution for this industrial challenge. Initial experiments revealed a promising capability: S. ludwigii APRE2 effectively produced ethanol from undetoxified sugarcane bagasse hydrolysate at 37 °C, eliminating the need for costly detoxification pretreatments typically required in industrial processes. To optimize fermentation efficiency, a central composite design (CCD) approach was implemented. This statistical method identified precise optimal parameters: sugar concentration (143.95 g/L), diammonium phosphate (4.99 g/L), pH (4.98), yeast extract (8.94 g/L), and magnesium sulfate (2.22 g/L). The optimized conditions yielded impressive results: a maximum ethanol concentration of 38.11 g/L, productivity of 1.59 g/L·h, and yield of 0.45 g/L. These metrics position S. ludwigii APRE2 competitively among other high-performing nonconventional ethanologenic yeasts. This study establishes S. ludwigii as a viable candidate for sustainable, industrial-scale bioethanol production from lignocellulosic biomass, potentially reducing production costs through elimination of detoxification steps.
Keywords:
biofuel
; lignocellulosic materials
; multistress-tolerant yeast
; nonconventional yeast
; Sacharomycodes ludwigii
; sugarcane bagasse
1. Introduction
Sugarcane (Saccharum officinarum) stands as one of the most economically important crops grown worldwide. According to the Food and Agriculture Organization of the United Nations [1], global sugarcane production reached approximately 2 billion tons in 2023, with Brazil leading production at 782.59 million tons (mt), followed by India (490.53 mt), China (104.57 mt), and Thailand (93.98 mt).
The sugar production process generates sugarcane bagasse as its major by-product, accounting for approximately 37% to 40% of the raw material [2]. While some bagasse serves as fuel for heat generation in sugar factories, a considerable portion remains unutilized. This by-product is particularly valuable due to its high content of insoluble carbohydrates, primarily cellulose and hemicellulose. Multiple studies have documented the composition of sugarcane bagasse, with Pereira et al. [3] reporting 36-40% cellulose and 28-32% hemicellulose on a dry weight basis. Subsequent research by de Souza et al. [4] found similar concentrations of 39.2% cellulose and 37.9% hemicellulose, while de Araujo Guilherme et al. [5] documented slightly different proportions at 38.6% cellulose and 27.9% hemicellulose. More recent studies have indicated even higher cellulose content, with Chamnipa et al. [6] reporting 47.9% cellulose and 31.4% hemicellulose, and Dolpatcha et al. [7] finding 49.09% cellulose and 29.30% hemicellulose. These carbohydrates can be converted into fermentable sugars to produce high-value products. The substantial cellulose and hemicellulose content makes sugarcane bagasse a promising feedstock for various bioproducts, particularly second-generation bioethanol, which has gained recognition as an important alternative biofuel used worldwide.
The pretreatment and hydrolysis of sugarcane bagasse into fermentable sugars represent critical processes for utilizing this agricultural waste as feedstock for bioethanol production. Several pretreatment platforms have been developed and employed, including chemical, physical, physico-chemical, and biological approaches [8]. Among these techniques, chemical processes—particularly dilute acid pretreatment—are commonly utilized due to their operational advantages. These advantages include ease of handling, low-cost operation resulting from lower temperature requirements, high efficiency in separating plant cell wall components, substantial yield of fermentable sugars, and broad applicability across diverse biomass sources [6,9−13].
Despite these benefits, a significant drawback of dilute acid pretreatment is the generation of inhibitory compounds during the process. These inhibitors include organic acids (such as acetic acid and formic acid), furan derivatives (furfural and 5-hydroxymethyl furfural or 5-HMF), and various phenolic compounds [11,14]. The concentrations of these inhibitors vary depending on the raw materials and pretreatment conditions; however, their accumulation can adversely affect microbial growth and metabolic activity during ethanol fermentation, thereby reducing overall conversion efficiency and ethanol yield. To overcome this challenge, researchers have developed several mitigation strategies. These include implementing detoxification processes to remove or reduce inhibitors in the hydrolysate and developing multistress-tolerant ethanol-producing microorganisms. Among these approaches, the application of multistress-tolerant ethanologenic microbes has emerged as particularly promising. These specialized microorganisms can withstand the presence of inhibitory compounds while maintaining efficient fermentation performance, thereby maximizing the potential of sugarcane bagasse as a sustainable feedstock for second-generation bioethanol production. This biological approach not only addresses the inhibitor challenge but also potentially reduces processing costs by eliminating the need for separate detoxification steps, ultimately enhancing the economic viability of the overall bioethanol production process.
Multistress-tolerant yeasts have gained significant attention in bioethanol production due to their ability to withstand various stress conditions during fermentation processes. Numerous studies have successfully isolated and identified multistress-tolerant yeasts capable of efficient bioethanol production [12,15,16]. However, research has predominantly focused on specific genera such as Pichia kudriavzevii [12,15−19], Kluyveromyces marxianus [16,20−22], and Candida tropicalis [12,16]. In contrast, relatively limited information exists regarding other potentially valuable multistress-tolerant yeasts, particularly Saccharomycodes ludwigii [23]. This nonconventional yeast species has been isolated from diverse ecological niches, including sweet wine [24], soil [25], and coconut inflorescence sap [26]. S. ludwigii demonstrates considerable potential for various fermentation applications, including biofuel production, winemaking, and low-alcohol beer brewing [27]. Most notably, recent studies have established that this yeast exhibits remarkable multi-stress tolerance toward critical fermentation inhibitors, including heat, ethanol, acetic acid, furfural, and 5-(hydroxymethyl) furfural [23].
Despite these promising characteristics, a significant research gap exists in understanding the bioethanol production capabilities of S. ludwigii when utilizing lignocellulosic biomass, particularly sugarcane bagasse. To the best of our knowledge, no studies have documented the ethanol production efficiency of this nonconventional yeast using sugarcane bagasse hydrolysate as feedstock. Therefore, this study aims to comprehensively evaluate the ethanol fermentation efficiency of S. ludwigii using sugarcane bagasse hydrolysate as the primary carbon source. Additionally, the research will determine the optimal fermentation conditions for maximum ethanol production by this multistress-tolerant yeast through statistical experimental design approaches. This investigation will contribute valuable insights into expanding the range of effective yeast strains for lignocellulosic bioethanol production and potentially establish S. ludwigii as a promising candidate for industrial applications.
2. Materials and Methods
2.1. Chemicals
Chemicals for culture medium preparation, including yeast extract, malt extract, and peptone (bacteriological grade), were purchased from TM media (Titan Biotech Ltd., Delhi, India). Other chemicals, such as isopropanol and ethanol (HPLC grade), glucose, and magnesium sulfate (MgSO4), were obtained from KemAusTM (New South Wales, Australia). Sulfuric acid was purchased from Merck (Merck Ltd, Darmstadt, Germany), while phenol was obtained from Sigma-Aldrich (St. Louis, MI, USA).
2.2. Microbial Strain and Culture Conditions
In this study, the multistress-tolerant yeast Saccharomycodes ludwigii APRE2, previously isolated from tropical acidic fruits [23], was utilized. The strain was maintained on yeast malt (YM) agar slants (3 g/L yeast extract, 3 g/L malt extract, 5 g/L peptone, 10 g/L glucose, and 20 g/L agar) at 4 °C. For inoculum preparation, a loopful of yeast cells was transferred to YM broth and incubated in a controlled incubator shaker (JSR, Gongju, Republic of Korea) at 35 °C and 150 rpm for 16 h. These cells were then transferred to fresh YM broth at an initial concentration of approximately 10⁵ cells/mL and incubated under identical conditions for 9 h. The resulting yeast culture served as the starter culture for all subsequent experiments.
2.3. Sugarcane Bagasse Hydrolysate (SCH) Preparation
Sugarcane bagasse was obtained from Mitr Phu Wiang Sugar Factory in Khon Kaen, Thailand. The bagasse was dried in a hot air oven at 60 °C until reaching constant weight, then pulverized into small pieces of approximately 0.5 × 0.5 cm. The prepared bagasse was subjected to acid pretreatment following the method described by Sritrakul et al. [11]. This pretreatment involved transferring 10% (w/v) of dried sugarcane bagasse into 3% (v/v) sulfuric acid solution and keeping it at room temperature for 16 h. The mixture was subsequently heated at 121 °C and 15 psi for 1 h. After pretreatment, the liquid fraction (acid hydrolysate) was collected by filtration through a muslin cloth and used directly as feedstock for ethanol production without detoxification.
The remaining solid fraction of the acid-pretreated sugarcane bagasse was washed with running tap water and dried at 60 °C until constant weight before enzymatic hydrolysis. For this process, 5% (w/v) of the acid-pretreated solid fraction was transferred into 50 mM sodium citrate buffer (pH 4.8) with cellulase (Cellic® CTec2) added at a concentration of 15 filter paper units (FPU)/g dry solid. After incubation at 50 °C and 200 rpm for 24 h, the enzymatic reaction was terminated by heating the reaction mixture in boiling water for 5 min. The liquid fraction (enzymatic hydrolysate) was collected by centrifugation at 8000 rpm for 10 min and used as feedstock for ethanol production in subsequent experiments.
The concentrations of sugars and lignocellulosic inhibitors in both the acid and enzymatic hydrolysates were determined using high-performance liquid chromatography (HPLC).
2.4. Ethanol Production from Acid and Enzymatic Hydrolysate of Sugarcane Bagasse
A comparative analysis of ethanol production by S. ludwigii was conducted using acid and enzymatic hydrolysates as feedstocks in a batch fermentation process. The fermentations were performed in 250-mL Erlenmeyer flasks, each containing 100 mL of the respective hydrolysate supplemented with 1.5 g/L of yeast extract and 3.0 g/L of peptone [7]. Prior to fermentation, each medium was inoculated with a starter culture of S. ludwigii at an initial concentration of approximately 1 × 107 cells/mL. The process was carried out in a controlled incubator shaker at 37 °C and 150 rpm. Samples were collected at predetermined time intervals throughout the fermentation period to monitor ethanol concentration using gas chromatography (GC) and sugar concentration using the phenol-sulfuric acid method.
2.5. Optimization Conditions for Ethanol Production from SBH
For this experiment, the enzymatic hydrolysate of sugarcane bagasse was used as feedstock for ethanol production. Response surface methodology (RSM) based on central composite design (CCD) was employed to determine the optimal conditions for ethanol production from SBH by S. ludwigii. Based on literature review, various independent variables that may affect ethanol production were tested, including sugar concentration [28−30], diammonium phosphate (DAP) [12,31], pH [29,31,32], yeast extract [28,30,31], and magnesium sulfate (MgSO4) [31−33]. The experimental codes and actual values (low and high) of each variable are summarized in Table 1.
Regression analysis was performed on data generated from the experimental design using Design Expert version 13 (Demo version) (Stat-Ease Inc., Minneapolis, MN, USA), with ethanol concentration as the response variable. The predictive model of the design was presented as a second-order polynomial function in Equation (1):
Where Y is the predicted response, xᵢ and xⱼ are the independent variables, β₀ is a constant term, βᵢ represents the linear coefficients, βᵢᵢ represents the squared coefficients, and βᵢⱼ is the interaction coefficient.
Analysis of variance (ANOVA) was performed using Design-Expert version 13 (Demo version) to evaluate the statistical significance of independent variables. The coefficient of determination (R²) was determined to monitor the reliability of the experimental design. The optimized fermentation conditions obtained from the response surface plots and the prediction equation established from the statistical experimental model were verified through batch mode fermentation in a flask scale, as previously described.
2.6. Analytical Methods and Statistical Analysis
Total sugar was determined by the phenol-sulfuric acid method using glucose as a standard [34]. Ethanol concentration (P, g/L) was measured by GC (GC-14B, Shimadzu, Kyoto, Japan) following the procedure described by Laopaiboon et al. [35]. Volumetric ethanol productivity (Qp, g/L·h) and ethanol yield (Yp/s, g ethanol produced/g glucose consumed) were calculated according to the method of Nuanpeng et al. [28]. Sugar compositions (glucose, xylose, and arabinose) and inhibitors (acetic acid, formic acid, furfural, 5-hydroxymethyl furfural (5-HMF), and vanillin) in the SBH were analyzed using HPLC (Shimadzu, Kyoto, Japan) equipped with an Aminex HPX-87H column. Sugars were detected using a refractive index detector, while inhibitors were detected using a UV-VIS detector at 210 nm wavelength. A 5 mM sulfuric acid (H₂SO₄) mobile phase was used at a flow rate of 0.6 mL/min at 40 °C. All experiments were performed twice, each with two replications, with results expressed as means ± standard deviations (SDs).
3. Results and Discussion
3.1. Ethanol Production Potential of S. ludwigii Using Acid and Enzymatic Sugarcane Bagasse Hydrolysate as Feedstock
The chemical compositions of sugarcane bagasse hydrolysates used in the current study were comprehensively analyzed by Dolpatcha et al. [7], who reported detailed findings on sugar content and inhibitory compounds. The acid hydrolysate contained 25.81 g/L of total sugar, with xylose as the predominant component (20.32 g/L), followed by arabinose (2.90 g/L) and glucose (2.59 g/L). In contrast, the enzymatic hydrolysate yielded significantly higher total sugar content at 63.88 g/L, with glucose being the major constituent (43.54 g/L), followed by xylose (19.29 g/L) and arabinose (1.05 g/L).
Regarding lignocellulosic inhibitors, the profiles differed markedly between the two hydrolysis methods. Acetic acid was the primary inhibitor in the acid hydrolysate, measuring 3.77 g/L, compared to only 0.89 g/L in the enzymatic hydrolysate. Conversely, formic acid and furfural were the predominant inhibitors in the enzymatic hydrolysate (2.80 g/L and 1.57 g/L, respectively), whereas these compounds appeared in much lower concentrations in the acid hydrolysate (0.26 g/L formic acid and 0.08 g/L furfural). 5-HMF was detected in minimal amounts in both hydrolysates, while vanillin was present exclusively in the acid hydrolysate and absent in the enzymatic preparation. As previously reported, these compositional differences highlight the distinct chemical profiles resulting from acid versus enzymatic hydrolysis techniques [7].
A primary objective of this research was to evaluate the direct utilization potential of both hydrolysates for ethanol production without implementing detoxification procedures. Fermentation experiments using acid hydrolysate as substrate yielded no detectable ethanol production, as depicted in Figure 1. This outcome can be attributed to the insufficient glucose concentration (2.59 g/L) in the acid hydrolysate, as glucose constitutes the essential carbon source for both cellular growth and ethanol biosynthesis in the investigated yeast strain. Throughout the fermentation period, xylose and arabinose concentrations remained essentially constant, confirming previous findings by Vejarano [27] and Pilap et al. [23] that this nonconventional yeast lacks the metabolic pathways necessary for pentose sugar utilization.
When the enzymatic hydrolysate was employed as the fermentation substrate, the higher glucose concentration (43.54 g/L) facilitated substantial ethanol production, reaching 20.54 g/L after 24 hours with a conversion efficiency of 0.48 g ethanol per gram of glucose consumed. This yield approaches the theoretical maximum (0.51 g/g), demonstrating that the inhibitory compounds present in the enzymatic hydrolysate did not significantly impair fermentation performance. As observed in the acid hydrolysate fermentation, the pentose sugars (xylose and arabinose) remained unconsumed. The exceptional fermentation efficiency despite the presence of inhibitors can be attributed to the remarkable tolerance of this nonconventional yeast strain toward lignocellulosic inhibitors, with previous research by Pilap et al. [23] establishing its capacity to withstand acetic acid (up to 10 g/L), furfural (up to 1.5 g/L), and 5-HMF (up to 2.5 g/L) without significant metabolic impairment.
Based on these findings, the enzymatic hydrolysate was selected as feedstock for ethanol production in subsequent experiments. Before use, the hydrolysate underwent evaporation to concentrate its total sugar content to approximately 300 g/L.
3.2. Optimization Conditions for Ethanol Production from SBH
Based on a comprehensive literature review, several key factors have been identified that significantly influence ethanol fermentation efficiency and yield, particularly under high-temperature conditions. These factors include sugar concentration [28−30], DAP [12,31], pH of the fermentation medium [29,31,32], yeast extract [28,30,31], and MgSO₄ [31−33]. To investigate these parameters, their impact on ethanol fermentation from the enzymatic hydrolysate of sugarcane bagasse at 37 °C using S. ludwigii was systematically optimized. The optimization process employed CCD through the Design-Expert program, which generated 47 experimental runs with varying combinations of the selected factors. The actual values for each variable in these experimental runs are summarized in Table 2.
The fermentation experiments were conducted on a flask scale using 100 mL of fermentation medium. The actual ethanol concentrations obtained from the experimental runs ranged from 7.87 g/L to 37.03 g/L, which aligned closely with the predicted values (6.87 - 36.26 g/L). The highest ethanol concentration of 37.03 g/L was achieved in run 35, which utilized specific parameter values: 150 g/L sugar concentration, 5.0 g/L DAP, pH 5.0, 8.5 g/L yeast extract, and 2.5 g/L MgSO₄. In contrast, the lowest ethanol yield of 7.87 g/L was observed in run 29, which employed a significantly higher sugar concentration of 268.92 g/L while maintaining the same levels of other parameters (5.0 g/L DAP, pH 5.0, 8.5 g/L yeast extract, and 2.5 g/L MgSO₄) (Table 2). This substantial reduction in ethanol production can be attributed to the excessive sugar concentration creating high osmotic pressure, which previous studies have shown to adversely affect microbial growth and metabolic activity [36,37]. This finding underscores the critical importance of optimizing sugar concentration in ethanol fermentation processes, as concentrations that are too high can paradoxically reduce overall yield despite providing more fermentable substrate.
A quadratic polynomial equation was developed to predict ethanol concentration based on experimental data from the CCD experiment. The equation is:
P (g/L) = 36.26 − 1.08A − 0.10B − 0.15C + 0.42D − 0.89E − 0.2AB + 0.24AC + 0.11AD + 0.24AE − 0.48BC + 0.21BD − 0.19BE − 0.15CD − 0.10CE − 0.003DE − 4,74A2 − 0.97B2 − 3.93C2 − 1.62D2 − 1.61E2
Where P is the ethanol concentration (g/L), A, B, C, D, and E are the code factors for sugar concentration, DAP, pH, yeast extract, and MgSO4, respectively.
The statistical analysis of variance (ANOVA) of the experimental data obtained from the CCD is summarized in Table 3. Results indicate that the established model was statistically significant, while the lack of fit was not, implying that the model is reliable for predicting response values. The relatively high determination coefficient (R2 = 0.98) demonstrates close alignment between predicted and actual values of the response variable, suggesting that the established model could explain 98% of the ethanol concentration variance.
Based on the linear and quadratic terms shown in Table 3, three variables, specifically sugar concentration (A), yeast extract (D), and MgSO4 (E), significantly affected ethanol production from SBH by S. ludwigii (p < 0.05). These findings align with previous studies by Nuanpeng et al. [28], Techaparin et al. [29], Nuanpeng et al. [30], Phong et al. [31], Pereira et al. [33], and Nguyen et al. [32]. Although no interaction effects were found among these significant variables, an interaction effect between DAP and pH (BC) was detected, suggesting these two factors are also important parameters for ethanol fermentation by this nonconventional yeast, particularly when their combined effect is considered, rather than as individual factors.
To examine the interactive effects of the five selected variables on ethanol production by S. ludwigii, three-dimensional (3-D) response surfaces and contour plots were generated using the established model and experimental data from the CCD experiment. As illustrated in Figure 2, each variable pair exhibited distinct interactive effects on ethanol production. When analyzing the interactive effect of sugar concentration with other variables, ethanol content increased as sugar concentration rose to approximately 150 g/L, beyond which ethanol concentration decreased. This pattern suggests the existence of substrate inhibition at higher sugar concentrations, a phenomenon commonly observed in yeast fermentation [31,38,39]. Varying DAP concentration showed no significant impact on ethanol production by S. ludwigii (Figure 2A), indicating that this strain may efficiently utilize nitrogen sources already present in the medium.
With respect to pH, a slight increase in ethanol content was observed when the initial pH of the fermentation medium was raised from 4.0 to 5.0, followed by a minor decrease at pH values exceeding 5.0 (Figure 2B). This optimal pH range aligns with previous studies [28,29]. Similar to DAP, minimal changes in ethanol concentration were detected when varying either yeast extract concentration (Figure 2C) or MgSO4 content (Figure 2D), while maintaining different sugar concentrations. These results collectively suggest that among the parameters studied, sugar concentration exerts the most significant influence on ethanol production by S. ludwigii, a finding consistent with fundamental principles of fermentation stoichiometry, where sugar serves as the primary carbon source for conversion to ethanol.
With sugar concentration held constant at the middle point (150 g/L), most variable pairs showed no interactive effects on ethanol production, with one notable exception. No significant interaction was observed between DAP and yeast extract (Figure 2F) or between DAP and MgSO4 (Figure 2G). However, a clear interactive effect was detected between DAP and pH (Figure 2E). Ethanol production increased with pH, reaching its maximum at approximately pH 5.0, after which ethanol concentration declined as pH continued to increase. Similarly, when both sugar and DAP concentrations were fixed at their middle points, varying the concentrations of either yeast extract (Figure 2H) or MgSO4 (Figure 2I) resulted in minimal changes to ethanol production. This indicates that when other parameters are optimized, the individual effects of these nutrients become less pronounced. The interaction between yeast extract and MgSO4 (Figure 2J) also showed no significant impact on ethanol production when the other variables were held constant. Despite the minimal interaction effects, the highest ethanol content was achieved at specific concentrations of yeast extract and MgSO4—approximately 9.0 g/L and 2.5 g/L, respectively. These findings suggest that while these nutrients are necessary for optimal fermentation, their concentrations become less critical once they reach certain threshold levels, provided that other key parameters (sugar concentration, pH, and DAP) are appropriately regulated.
Based on the second-order polynomial model and response surface analysis, the predicted optimum conditions for ethanol production from SBH by S. ludwigii at 37°C were determined to be: sugar concentration of 143.95 g/L, DAP concentration of 4.99 g/L, pH of 4.98, yeast extract concentration of 8.94 g/L, and MgSO4 concentration of 2.22 g/L. To validate these theoretical predictions, a verification experiment was conducted in a 250-mL flask-scale fermentation using these optimized parameters. As shown in Figure 3, ethanol concentration increased significantly during fermentation, reaching its maximum value at 24 hours. This peak in ethanol production corresponded directly with the reduction in total sugar and glucose concentrations in the medium. Notably, xylose concentration remained constant throughout the fermentation period, confirming that S. ludwigii lacks the metabolic capability to utilize this pentose sugar.
Under these optimized conditions, the maximum ethanol concentration achieved was 38.11 g/L, with a volumetric ethanol productivity of 1.59 g/L·h and an ethanol yield of 0.45 g ethanol produced per g of glucose consumed. The experimental ethanol concentration (38.11 g/L) closely matched the model's predicted value (36.48 g/L), with a difference of only 4.5%. This strong agreement between predicted and experimental results validates the accuracy and reliability of the established model for predicting ethanol production from SBH using S. ludwigii. These findings demonstrate the effectiveness of response surface methodology for optimizing bioethanol production parameters in this nonconventional yeast system.
A comparative analysis was conducted examining ethanol production from SBH by S. ludwigii relative to other ethanologenic yeasts using various lignocellulosic materials as feedstock. As summarized in Table 4, S. ludwigii APRE2 demonstrated superior ethanol production capacity compared to several other yeast species and strains. Specifically, the ethanol content produced by S. ludwigii APRE2 exceeded that reported for P. kudriavzevii RGB3.2 and K. marxianus RGB4.5 using rice straw [16], K. marxianus CECT10875 using woody and herbaceous biomass [40], S. cerevisiae TC-5 using corncob residue [41], and S. cerevisiae PTCC5052 using wheat straw [42] as feedstock. In comparison with P. kudriavzevii RZ8-1 using sugarcane bagasse [17] and S. cerevisiae using pomelo peel waste [43], the ethanol content produced by S. ludwigii was comparable. Notably, the ethanol concentration, productivity, and yield obtained from S. ludwigii APRE2 using sugarcane bagasse were relatively similar to those values reported when using pineapple waste as feedstock [23].
Regarding inhibitory compounds, although the concentration of lignocellulosic inhibitors in the concentrated enzymatic hydrolysate was not directly measured, we estimate their concentration likely increased approximately 2.2-fold compared to the original hydrolysate. This estimation is based on the proportional increase observed in sugar content during the concentration process. Despite this potential increase in inhibitory compounds, the glucose utilization profile and ethanol production efficiency of S. ludwigii remained robust. The fermentation performance was not significantly affected by these inhibitors, suggesting that S. ludwigii APRE2 possesses notable tolerance to lignocellulosic inhibitors commonly present in hydrolysates. This inhibitor tolerance, combined with the competitive ethanol yields demonstrated in our comparative analysis, positions S. ludwigii APRE2 as a promising candidate for industrial-scale lignocellulosic ethanol production processes.
Various cellular mechanisms are involved in stress response in yeast cells, including the regulation of cell wall integrity, remodeling of cellular structure and membrane transport, activation of antioxidative systems, and changes in signaling pathway and gene transcription under stress conditions [44−48]. Different species of yeasts respond differently to stress conditions. Thus, further study to elucidate the cellular mechanisms in response to stress conditions in this multistress-tolerant S. ludwigii is needed.
Future research should focus on applying advanced transcriptomic and proteomic approaches to comprehensively map the stress response pathways in S. ludwigii under lignocellulosic hydrolysate conditions [49,50], while comparative genomic analyses with conventional bioethanol yeasts could reveal unique genetic elements responsible for superior stress tolerance [46,49,51]. Investigating epigenetic regulation mechanisms [52], membrane lipid composition changes [53,54], and key transcription factors governing multistress responses [55,56] would provide crucial insights for developing robust industrial strains. The acquired information provides not only basic knowledge on how the yeast responds to stress but also gives useful information to develop a robust yeast with desirable traits, as well as modification of the fermentation conditions for a highly efficient lignocellulosic ethanol fermentation process.
4. Conclusions
This study demonstrates that the multistress-tolerant yeast S. ludwigii APRE2 exhibits exceptional potential for ethanol production using undetoxified sugarcane bagasse hydrolysate as feedstock. Initial testing under unoptimized conditions revealed that this nonconventional yeast produced 20.54 g/L of ethanol with a yield of 0.48 g/g from the enzymatic hydrolysate. Statistical analysis through linear and quadratic regression identified sugar concentration, yeast extract, and MgSO₄ as the three critical variables significantly influencing ethanol production. Under optimized conditions through CCD (143.95 g/L sugar concentration, 4.99 g/L DAP, pH 4.98, 8.94 g/L yeast extract, and 2.22 g/L MgSO₄), S. ludwigii APRE2 achieved the highest ethanol concentration of 38.11 g/L, along with a productivity of 1.59 g/L·h and yield of 0.45 g/L. These performance metrics are comparable to other leading ethanologenic yeasts. The results establish S. ludwigii as a promising multistress-tolerant yeast for sustainable bioethanol production from lignocellulosic biomass, offering significant advantages for industrial applications.
Author Contributions
Conceptualization, P.K., S.T., J.A., K.F., R.L., M.Y. and P.T.; methodology, P.K., S.T., W.P. and P.T.; software, P.T.; validation, P.K., S.T., W.P. and P.T.; formal analysis, P.K., S.T., W.P. and P.T.; investigation, P.K., S.T., W.P. and P.T.; resources, P.T.; data curation, P.K., S.T. and P.T.; writing—original draft preparation, P.K., S.T., W.P. and P.T.; writing—review and editing, P.K., S.T., M.Y. and P.T.; visualization, P.T.; supervision, J.A., M.Y. and P.T.; project administration, P.T.; funding acquisition, J.A. and P.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Fundamental Fund (FF) of Khon Kaen University and the National Science, Research, and Innovation Fund (NSRF), year 2025. A part of this work was also supported by the Ministry of Higher Education, Science, Research and Innovation through Reinventing University (2025), Khon Kaen University, Thailand.
Data Availability Statement
The experimental data are contained within the article.
Acknowledgments
The authors thank the Mitr Phu Wiang Sugar Factory, Khon Kaen, Thailand, for providing sugarcane bagasse. The authors also thank Wutthichai Janmahong and Kannika Boonkert for technical assistance in chemical analysis and the Department of Biotechnology, Faculty of Technology, and the Research Center for Value Added Agricultural Products (FerVAAPs), Khon Kaen University, for all research facilities.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Food and Agriculture Organization of the United Nations. Available online: https://ourworldindata.org/grapher/sugar-cane-production?country=USA~BRA~IND~CHN~European+Union~THA (accessed on 19 April, 2025).
- Junpen, A.; Pansuk, J.; Garavait, S. Estimation of reduced air emissions as a result of the implementation of the measure to reduce burned sugarcane in Thailand. Atmosphere. 2020, 11, 366. [Google Scholar] [CrossRef]
- Pereira, S.C.; Maehara, L.; Machado, C.M.M.; Farinas, C.S. 2G ethanol from the whole sugarcane lignocellulosic biomass. Biotechnol. Biofuels. 2015, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.F.R.; Dutra, E. D.; Leite, F.C.B.; Cadete, R.M.; Rosa, C.A.; Stambuk, B.U.; Stamford, T.L.M.; de Morais, M.A.Jr. Production of ethanol fuel from enzyme-treated sugarcane bagasse hydrolysate using d-xylose-fermenting wild yeast isolated from Brazilian biomes. 3 Biotech. 2018, 8, 312. [Google Scholar]
- de Araujo Guilherme, A.; Dantas, P.V.F.; Padilha, C.E.A.; Dos Santos, E.S.; de Macedo, G.R. Ethanol production from sugarcane bagasse: Use of different fermentation strategies to enhance an environmental-friendly process. J Environ Manage. 2019, 234, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Chamnipa, N.; Klanrit, P.; Thanonkeo, S.; Thanonkeo, P. Sorbitol production from a mixture of sugarcane bagasse and cassava pulp hydrolysates using thermotolerant Zymomonas mobilis TISTR548. Ind. Crop. Prod. 2022, 188, 115741. [Google Scholar] [CrossRef]
- Dolpatcha, S.; Phong, H.X.; Thanonkeo, S.; Klanrit, P.; Yamada, M.; Thanonkeo, P. Adaptive laboratory evolution under acetic acid stress enhances the multistress tolerance and ethanol production efficiency of Pichia kudriavzevii from lignocellulosic biomass. Sci. Rep.-UK. 2023, 13, 21000. [Google Scholar] [CrossRef]
- Yusuf, A.A.; Inambao, F.L. Bioethanol production techniques from lignocellulose biomass as alternative fuel: a review. Int. J. Mech. Eng. Technol. 2019, 10, 34–71. [Google Scholar]
- Dussan, K.J.; Silva, D.D.V.; Moraes, E.J.C.; Arruda, P.V.; Felipe, M.G.A. Dilute-acid hydrolysis of cellulose to glucose from sugarcane bagasse. Chem. Engineer. Trans. 2014, 38, 433–438. [Google Scholar]
- Azhar, S.H.M.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Faik, A.A.M.; Rodrigues, K.F. Yeasts in sustainable bioethanol production: A review. Biochem. Biophys. Rep. 2017, 10, 52–61. [Google Scholar]
- Sritrakul, N.; Nitisinprasert, S.; Keawsompong, S. Evaluation of dilute acid pretreatment for bioethanol fermentation from sugarcane bagasse pith. Agri. Natural Resour. 2017, 51, 512–519. [Google Scholar] [CrossRef]
- Phong, H.X.; Klanrit, P.; Dung, N.T.P.; Yamada, M.; Thanonkeo, P. Isolation and characterization of thermotolerant yeasts for the production of second-generation bioethanol. Ann. Microbiol. 2019, 69, 765–776. [Google Scholar] [CrossRef]
- Sunkar, B.; Bhukya, B. Bi-phasic hydrolysis of corncobs for the extraction of total sugars and ethanol production using inhibitor resistant and thermotolerant yeast, Pichia kudriavzevii. Biomass Bioenerg. 2021, 153, 106230. [Google Scholar] [CrossRef]
- Chandel, A.K.; Kapoor, R.K.; Singh, A.; Kuhad, R.C. Detoxification of sugarcane bagasse hydrolysate improves ethanol production by Candida shehatae NCIM 3501. Bioresource Technol. 2007, 98, 1947–1950. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Park, E.H.; Kim, M.D. Isolation of thermotolerant yeast Pichia kudriavzevii from nuruk. Food Sci. Biotechnol. 2017, 26, 1357–1362. [Google Scholar] [CrossRef]
- Avchar, R.; Lanjekar, V.; Kshirsagar, P.; Dhakephalkar, P.K.; Dagar, S.S.; Baghela, A. Buffalo rumen harbours diverse thermotolerant yeasts capable of producing second-generation bioethanol from lignocellulosic biomass. Renew. Energ. 2021, 173, 795–807. [Google Scholar] [CrossRef]
- Chamnipa, N.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. The potential of the newly isolated thermotolerant yeast Pichia kudriavzevii RZ8-1 for high-temperature ethanol production. Braz. J. Microbiol. 2018, 49, 378–391. [Google Scholar] [CrossRef]
- Aouine, M.; Elalami, D.; Koraichi, S.I.; Haggoud, A.; Barakat, A. Exploring natural fermented foods as a source for new efficient thermotolerant yeasts for the production of second-generation bioethanol. Energies. 2022, 15, 4954. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, P.; Zhou, X.; Zheng, J.; Ma, Y.; Liu, C.; Wu, T.; Li, H.; Wang, X.; Wang, H.; Zhao, X.; Mehmood, M.A.; Zhu, H. Isolation, identification, and characterization of an acid-tolerant Pichia kudriavzevii and exploration of its acetic acid tolerance mechanism. Fermentation. 2023, 9, 540. [Google Scholar] [CrossRef]
- Charoensopharat, K.; Thanonkeo, P.; Thanonkeo, S.; Yamada, M. Ethanol production from Jerusalem artichoke tubers at high temperature by newly isolated thermotolerant inulin-utilizing yeast Kluyveromyces marxianus using consolidated bioprocessing. A. Van. Leeuw. J. Microb. 2015, 108, 173–190. [Google Scholar] [CrossRef]
- Hemansi.; Himanshu.; Patel, A.K.; Saini, J.K.; Singhania, R.R. Development of multiple inhibitor tolerant yeast via adaptive laboratory evolution for sustainable bioethanol production. Bioresource Technol. 2022, 344, 126247.
- Pattanakittivorakul, S.; Tsuzuno, T.; Kosaka, T.; Murata, M.; Kanesaki, Y.; Yoshikawa, H.; Limtong, S.; Yamada, M. Evolutionary adaptation by repetitive long-term cultivation with gradual increase in temperature for acquiring multi-stress tolerance and high ethanol productivity in Kluyveromyces marxianus DMKU 3-1042. Microorganisms. 2022, 10, 798. [Google Scholar] [CrossRef]
- Pilap, W.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. The potential of multistress tolerant yeast, Saccharomycodes ludwigii, for second-generation bioethanol production. Sci. Rep-UK. 2022, 12, 22062. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage yeasts in the wine industry. Int. J. Food Microbiol. 2003, 86, 23–50. [Google Scholar] [CrossRef]
- Thammasittirong, S.N.; Chamduang, T.; Phonrod, U.; Sriroth, K. Ethanol production potential of ethanol-tolerant Saccharomyces and non-Saccharomyces yeasts. Pol. J. Microbiol. 2012, 61, 219–221. [Google Scholar] [CrossRef]
- Udomsaksakul, N.; Kodama, K.; Tanasupawat, S.; Savarajara, A. Diversity of ethanol fermenting yeasts in coconut inflorescence sap and their application potential. ScienceAsia. 2018, 44, 371–381. [Google Scholar] [CrossRef]
- Vejarano, R. Saccharomycodes ludwigii, control and potential uses in winemaking processes. Fermentation. 2018, 4, 71. [Google Scholar] [CrossRef]
- Nuanpeng, S.; Thanonkeo, S.; Yamada, M.; Thanonkeo, P. Ethanol production from sweet sorghum juice at high temperatures using a newly isolated thermotolerant yeast Saccharomyces cerevisiae DBKKU Y-53. Energies. 2016, 9, 253. [Google Scholar] [CrossRef]
- Techaparin, A.; Thanonkeo, P.; Klanrit, P. High-temperature ethanol production using thermotolerant yeast newly isolated from Greater Mekong Subregion. Braz. J. Microbiol. 2017, 48, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Nuanpeng, S.; Thanonkeo, S.; Klanrit, P.; Yamada, M.; Thanonkeo, P. Optimization conditions for ethanol production from sweet sorghum juice by thermotolerant yeast Saccharomyces cerevisiae: using a statistical experimental design. Fermentation. 2023, 9, 450. [Google Scholar] [CrossRef]
- Phong, H.X.; Klanrit, P.; Dung, N.T.P.; Thanonkeo, S.; Yamada, M.; Thanonkeo, P. High-temperature ethanol fermentation from pineapple waste hydrolysate and gene expression analysis of thermotolerant yeast Saccharomyces cerevisiae. Sci. Rep-UK. 2022, 12, 13965. [Google Scholar] [CrossRef]
- Nguyen, P.V.; Nguyen, K.H.V.; Nguyen, N.L.; Ho, X.T.T.; Truong, P.H.; Nguyen, K.C.T. Lychee-derived, thermotolerant yeasts for second-generation bioethanol production. Fermentation. 2022, 8, 515. [Google Scholar] [CrossRef]
- Pereira, F.; Guimarães, P.M.R.; Teixeira, J.A.; Domingues, L. Optimization of low-cost medium for very high gravity ethanol fermentations by Saccharomyces cerevisiae using statistical experimental designs. Bioresour. Technol. 2010, 101, 7856–7863. [Google Scholar] [CrossRef]
- Mecozzi, M. Estimation of total carbohydrate amount in environmental samples by the phenol−sulphuric acid method assisted by multivariate calibration. Chemometr. Intell. Lab. 2005, 79, 84–90. [Google Scholar] [CrossRef]
- Laopaiboon, L.; Nuanpeng, S.; Srinophakun, P.; Klanrit, P.; Laopaiboon, P. Ethanol production from sweet sorghum juice using very high gravity technology: Effects of carbon and nitrogen supplementations. Bioresour. Technol. 2009, 100, 4176–4182. [Google Scholar] [CrossRef] [PubMed]
- Limtong, S.; Sringiew, C.; Yongmanitchai, W. Production of fuel ethanol at high temperature from sugar cane juice by a newly isolated Kluyveromyces marxianus. Bioresour. Technol. 2007, 98, 3367–3374. [Google Scholar] [CrossRef]
- Pilap, W.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. The potential of the newly isolated thermotolerant Kluyveromyces marxianus for high-temperature ethanol production using sweet sorghum juice. 3 Biotech. 2018, 8, 126. [Google Scholar] [CrossRef]
- Bai, F.W.; Chen, L.J.; Zhang, Z.; Anderson, W.A.; Moo-Young, M. Continuous ethanol production and evaluation of yeast cell lysis and viability loss under very high gravity medium conditions. J. Biotechnol. 2004, 110, 287–293. [Google Scholar] [CrossRef]
- Ozmihci, S.; Kargi, F. Comparison of yeast strains for batch ethanol fermentation of cheese-whey powder (CWP) solution. Lett. Appl. Microbiol. 2007, 44, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, I.; Oliva, J.M., Negro, M. J., Manzanares, P. & Ballesteros, M. Enzymic hydrolysis of steam exploded herbaceous agricultural waste (Brassica carinata) at different particule sizes. Process Biochem. 38, 187−192 (2002).
- Boonchuay, P.; Techapun, C.; Leksawasdi, N.; Seesuriyachan, P.; Hanmoungjai, P.; Watanabe, M.; Chaiyaso, T. Bioethanol production from cellulose-rich corncob residue by the thermotolerant Saccharomyces cerevisiae TC-5. J. Fungi. 2021, 7, 547. [Google Scholar] [CrossRef]
- Valamonfared, J.; Javanmard, A.S.; Ghaedi, M.; Bagherinasab, M. Bioethanol production using lignocellulosic materials and thermophilic microbial hydrolysis. Biomass Conv. Bior. 2024, 14, 16589–16601. [Google Scholar] [CrossRef]
- Huang, R.; Cao, M.; Guo, H.; Qi, W.; Su, R.; He, Z. Enhanced ethanol production from pomelo peel waste by integrated hydrothermal treatment, multienzyme formulation, and fed-batch operation. J. Agric. Food Chem. 2014, 62, 4643–4651. [Google Scholar] [CrossRef]
- Saini, P.; Beniwal, A.; Kokkiligadda, A.; Vij, S. Response and tolerance of yeast to changing environmental stress during ethanol fermentation. Process Biochem. 2018, 72, 1–12. [Google Scholar] [CrossRef]
- Eardley, J.; Timson, D.J. Yeast cellular stress: Impacts on bioethanol production. Fermentation. 2020, 6, 109. [Google Scholar] [CrossRef]
- Takagi, H. Molecular mechanisms and highly functional development for stress tolerance of the yeast Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 2021, 85, 1017–1037. [Google Scholar] [CrossRef]
- Postaru, M.; Tucaliuc, A.; Cascaval, D.; Galaction, A.I. Cellular stress impact on yeast activity in biotechnological processes-A short overview. Microorganisms. 2023, 11, 2522. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Wang, W.B.; Wang, Y.T.; Zhao, X.Q. Regulatory mechanisms underlying yeast chemical stress response and development of robust strains for bioproduction. Curr. Opinion. Biotechnol. 2024, 86, 103072. [Google Scholar] [CrossRef]
- Xu, J.R.; Mehmood, M.A.; Wang, L.; Ahmad, N.; Ma, H.J. Omics-based strategies to explore stress tolerance mechanisms of Saccharomyces cerevisiae for efficient fuel ethanol production. Front. Energy Res. 2022, 10, 884582. [Google Scholar] [CrossRef]
- Topaloğlu, A.; Esen, Ö.; Turanli-Yildiz, B.; Arslan, M.; Çakar, Z.P. From Saccharomyces cerevisiae to ethanol: Unlocking the power of evolutionary engineering in metabolic engineering applications. J. Fungi. 2023, 9, 984. [Google Scholar] [CrossRef]
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic expression programs in the response of yeast cells to environmental changes. Mol. Biol. Cell. 2000, 11, 4241–4257. [Google Scholar] [CrossRef]
- Kubiak-Szymendera, M.; Pryszcz, L.P.; Bialas, W.; Celinska, E. Epigenetic response of Yarrowia lipolytica to stress: Tracking methylation level and search for methylation patterns via whole-genome sequencing. Microorganisms. 2021, 9, 1798. [Google Scholar] [CrossRef]
- Vázquez, J.; Grillitsch, K.; Daum, G.; Mas, A.; Beltran, G.; Torija, M.J. The role of the membrane lipid composition in the oxidative stress tolerance of different wine yeasts. Food Microbiol. 2019, 78, 143–154. [Google Scholar] [CrossRef]
- Ribeiro, R.A.; Bourbon-Melo, N.; Sá-Correia, I. The cell wall and the response and tolerance to stresses of biotechnological relevance in yeasts. Front. Microbiol. 2022, 13, 953479. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Qi, X.; Lin, Y.; Guo, Y.; Zhang, Y.; Wang, Q. A hierarchical transcriptional regulatory network required for long-term thermal stress tolerance in an industrial Saccharomyces cerevisiae strain. Front. Bioeng. Biotechnol. 2022, 9, 826238. [Google Scholar] [CrossRef] [PubMed]
- Özel, A.; Topaloğlu, A.; Esen, Ö.; Holyavkin, C.; Baysan, M.; Çakar, Z.P. Transcriptomic and physiological meta-analysis of multiple stress-resistant Saccharomyces cerevisiae strains. Stresses. 2024, 4, 714–733. [Google Scholar] [CrossRef]
Figure 1.
The ethanol production from acid (A) and enzymatic hydrolysate (B) of sugarcane bagasse without detoxification at 37 °C using S. ludwigii.
Figure 1.
The ethanol production from acid (A) and enzymatic hydrolysate (B) of sugarcane bagasse without detoxification at 37 °C using S. ludwigii.
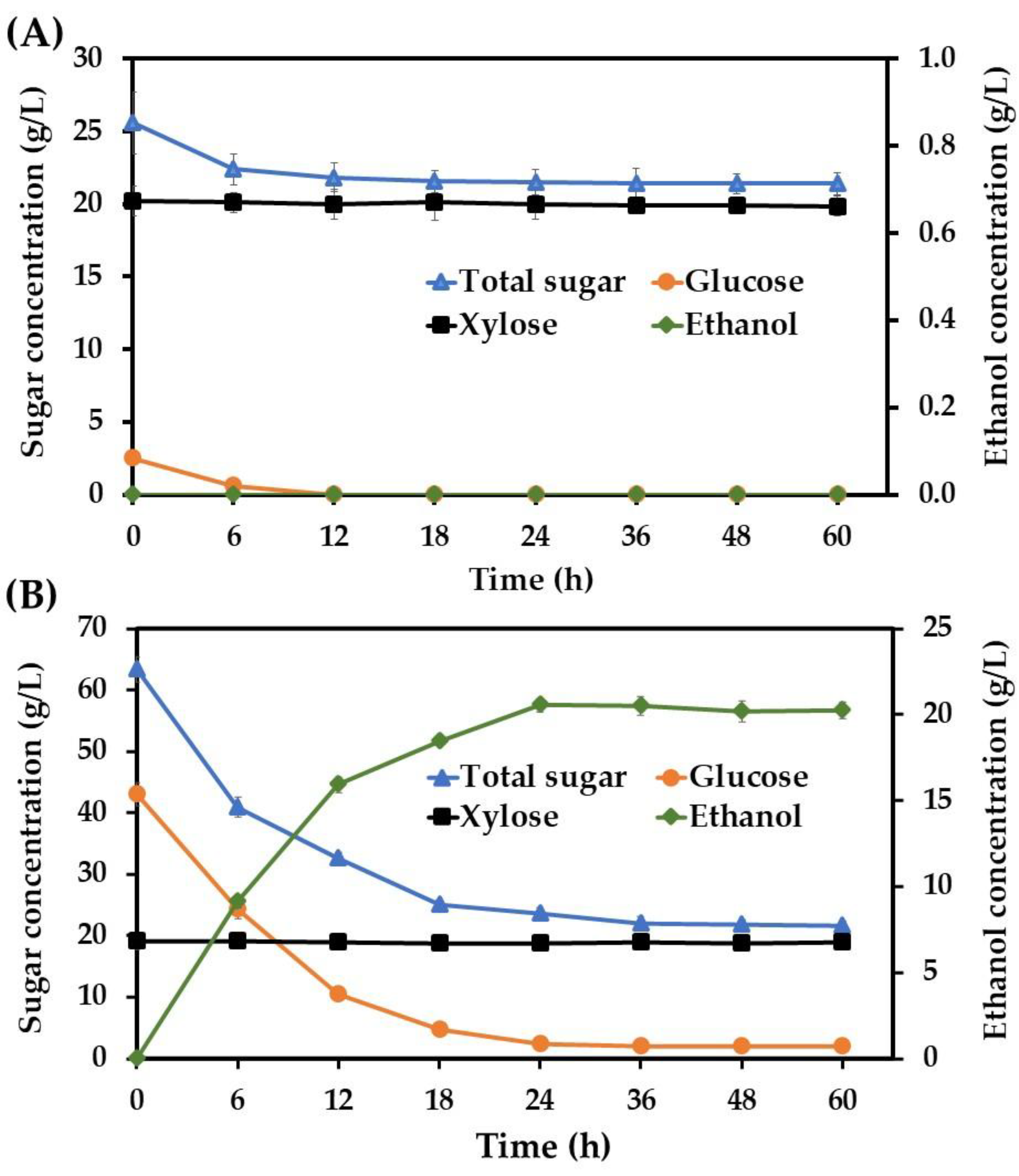
Figure 2.
Response surfaces and contour plots of interaction between sugar and DAP (A), sugar and pH (B), sugar and yeast extract (C), sugar and MgSO4 (D), DAP and pH (E), DAP and yeast extract (F), DAP and MgSO4 (G), pH and yeast extract (H), pH and MgSO4 (I), and yeast extract and MgSO4 (J) on ethanol production from SBH at 37 °C by S. ludwigii.
Figure 2.
Response surfaces and contour plots of interaction between sugar and DAP (A), sugar and pH (B), sugar and yeast extract (C), sugar and MgSO4 (D), DAP and pH (E), DAP and yeast extract (F), DAP and MgSO4 (G), pH and yeast extract (H), pH and MgSO4 (I), and yeast extract and MgSO4 (J) on ethanol production from SBH at 37 °C by S. ludwigii.
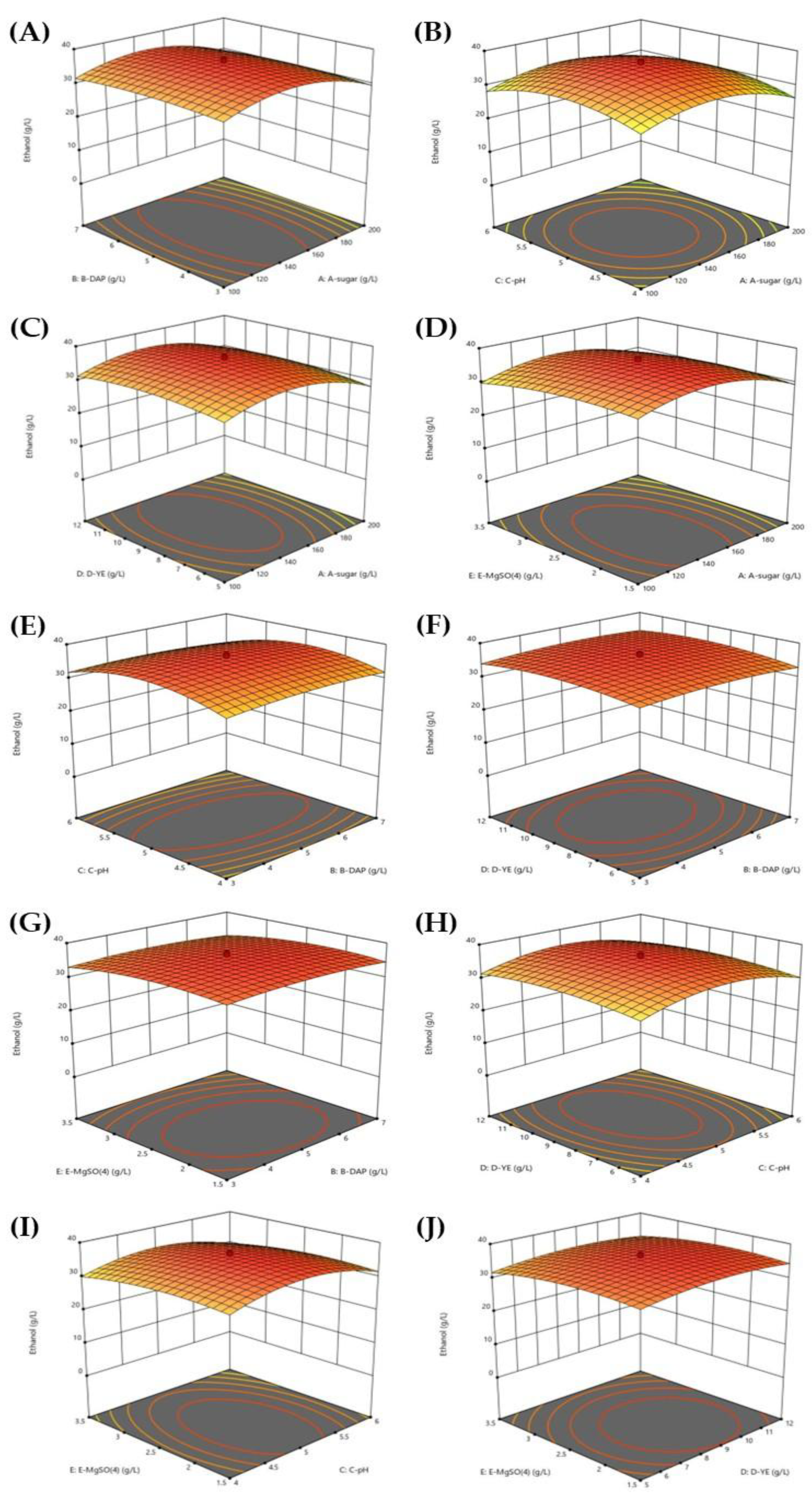
Figure 3.
The sugar utilization and ethanol production profiles of S. ludwigii at 37 °C using SBH as feedstock.
Figure 3.
The sugar utilization and ethanol production profiles of S. ludwigii at 37 °C using SBH as feedstock.
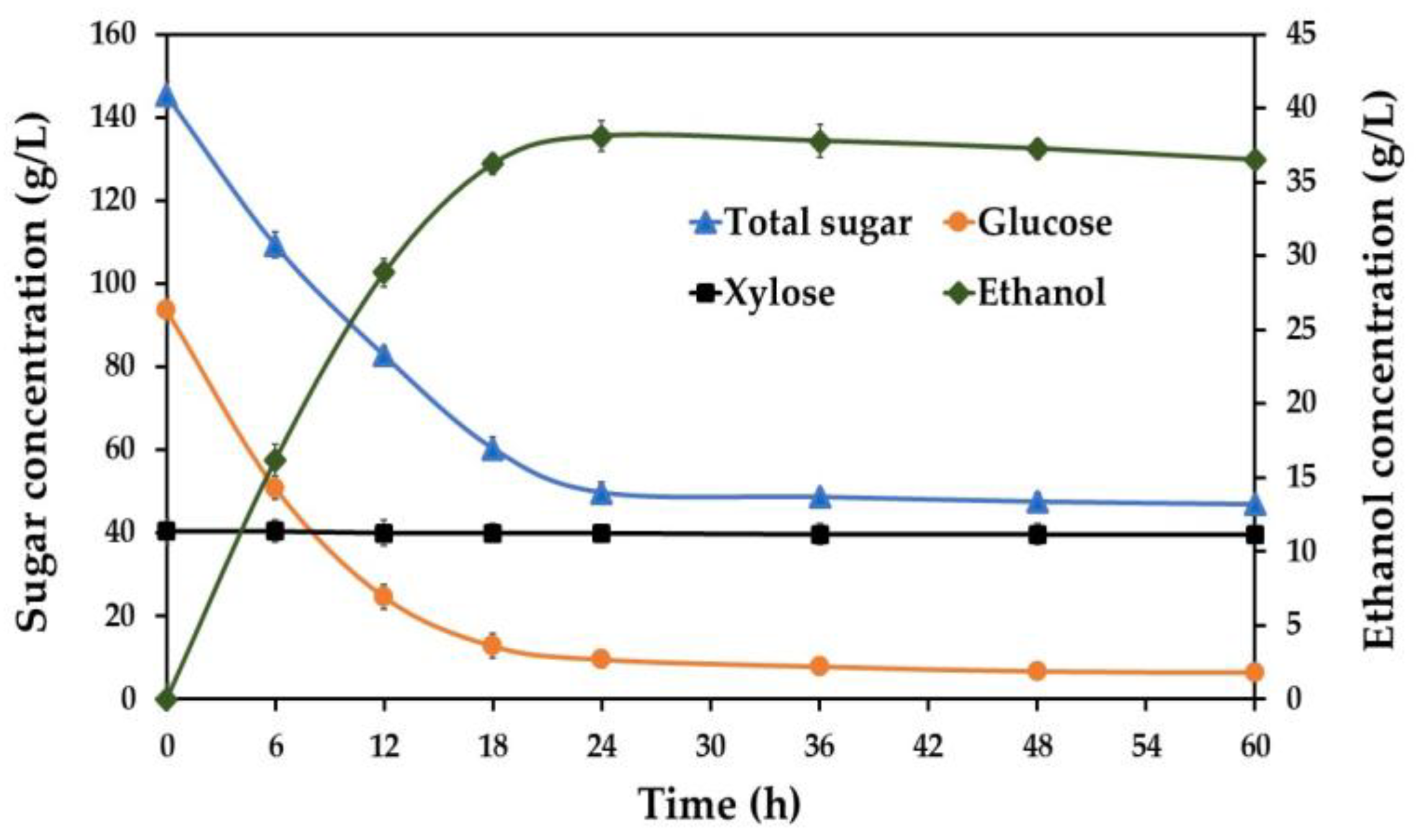

Table 1.
Codes and actual values of the independent variables for the optimization experiment using central composite design (CCD).
Table 1.
Codes and actual values of the independent variables for the optimization experiment using central composite design (CCD).
| Code | Variable | Unit | Level | ||||
| −2.37 | −1 | 0 | +1 | +2.37 | |||
| A | Sugar concentration | g/L | 31.08 | 100.00 | 150.00 | 200.00 | 268.92 |
| B | DAP | g/L | 0.24 | 3.00 | 5.00 | 7.00 | 9.76 |
| C | pH | - | 2.62 | 4.00 | 5.00 | 6.00 | 7.38 |
| D | Yeast extract | g/L | 0.18 | 5.00 | 8.50 | 12.00 | 16.82 |
| E | MgSO4 | g/L | 0.12 | 1.50 | 2.50 | 3.50 | 4.88 |
Table 2.
Codes and actual values of each independent variable, and the predicted and actual values of ethanol concentrations from the central composite design (CCD) experiment.
Table 2.
Codes and actual values of each independent variable, and the predicted and actual values of ethanol concentrations from the central composite design (CCD) experiment.
| Run | A-sugar (g/L) |
B-DAP (g/L) |
C-pH | D-YE (g/L) |
E-MgSO4 (g/L) |
Ethanol concentration (P, g/L) | |
| Predict value | Actual value | ||||||
| 1 | 200.00 | 3.00 | 4.0 | 12.00 | 1.50 | 22.70 | 20.34 |
| 2 | 150.00 | 5.00 | 5.0 | 16.82 | 2.50 | 28.08 | 27.40 |
| 3 | 150.00 | 5.00 | 5.0 | 8.50 | 4.88 | 25.04 | 23.14 |
| 4 | 100.00 | 3.00 | 6.0 | 12.00 | 3.50 | 23.54 | 24.06 |
| 5 | 200.00 | 7.00 | 6.0 | 5.00 | 3.50 | 20.27 | 20.81 |
| 6 | 200.00 | 7.00 | 6.0 | 5.00 | 1.50 | 22.14 | 21.59 |
| 7 | 200.00 | 7.00 | 4.0 | 5.00 | 3.50 | 20.93 | 19.89 |
| 8 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 36.62 |
| 9 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 35.80 |
| 10 | 100.00 | 7.00 | 4.0 | 12.00 | 3.50 | 24.69 | 25.10 |
| 11 | 100.00 | 3.00 | 4.0 | 12.00 | 3.50 | 23.85 | 24.34 |
| 12 | 100.00 | 3.00 | 4.0 | 12.00 | 1.50 | 25.55 | 26.20 |
| 13 | 100.00 | 7.00 | 6.0 | 12.00 | 1.50 | 25.30 | 25.04 |
| 14 | 150.00 | 5.00 | 2.6 | 8.50 | 2.50 | 14.37 | 13.35 |
| 15 | 200.00 | 3.00 | 4.0 | 5.00 | 3.50 | 21.03 | 22.76 |
| 16 | 200.00 | 3.00 | 6.0 | 5.00 | 3.50 | 22.28 | 20.40 |
| 17 | 200.00 | 3.00 | 6.0 | 12.00 | 1.50 | 23.75 | 24.60 |
| 18 | 150.00 | 5.00 | 5.0 | 8.50 | 0.12 | 29.28 | 30.20 |
| 19 | 100.00 | 7.00 | 6.0 | 5.00 | 3.50 | 21.73 | 23.56 |
| 20 | 100.00 | 3.00 | 4.0 | 5.00 | 1.50 | 25.08 | 25.82 |
| 21 | 200.00 | 7.00 | 4.0 | 12.00 | 3.50 | 22.71 | 23.57 |
| 22 | 150.00 | 5.00 | 5.0 | 0.18 | 2.50 | 26.10 | 25.80 |
| 23 | 31.08 | 5.00 | 5.0 | 8.50 | 2.50 | 12.00 | 10.02 |
| 24 | 100.00 | 3.00 | 4.0 | 5.00 | 3.50 | 23.38 | 23.15 |
| 25 | 100.00 | 3.00 | 6.0 | 12.00 | 1.50 | 25.63 | 26.04 |
| 26 | 100.00 | 7.00 | 4.0 | 12.00 | 1.50 | 27.14 | 28.40 |
| 27 | 200.00 | 3.00 | 4.0 | 5.00 | 1.50 | 21.76 | 21.50 |
| 28 | 150.00 | 9.76 | 5.0 | 8.50 | 2.50 | 30.54 | 29.80 |
| 29 | 268.92 | 5.00 | 5.0 | 8.50 | 2.50 | 6.87 | 7.87 |
| 30 | 200.00 | 7.00 | 4.0 | 5.00 | 1.50 | 22.42 | 22.80 |
| 31 | 100.00 | 7.00 | 4.0 | 5.00 | 3.50 | 23.37 | 23.90 |
| 32 | 150.00 | 5.00 | 7.4 | 8.50 | 2.50 | 13.67 | 13.71 |
| 33 | 200.00 | 7.00 | 6.0 | 12.00 | 3.50 | 21.46 | 21.80 |
| 34 | 200.00 | 3.00 | 4.0 | 12.00 | 3.50 | 21.95 | 22.90 |
| 35 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 37.03 |
| 36 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 35.90 |
| 37 | 200.00 | 7.00 | 4.0 | 12.00 | 1.50 | 24.20 | 24.19 |
| 38 | 200.00 | 7.00 | 6.0 | 12.00 | 1.50 | 23.34 | 22.75 |
| 39 | 150.00 | 0.24 | 5.0 | 8.50 | 2.50 | 31.04 | 30.80 |
| 40 | 100.00 | 7.00 | 4.0 | 5.00 | 1.50 | 25.81 | 25.76 |
| 41 | 200.00 | 3.00 | 6.0 | 5.00 | 1.50 | 23.40 | 23.88 |
| 42 | 200.00 | 3.00 | 6.0 | 12.00 | 3.50 | 22.62 | 22.42 |
| 43 | 100.00 | 3.00 | 6.0 | 5.00 | 1.50 | 25.74 | 24.65 |
| 44 | 100.00 | 7.00 | 6.0 | 12.00 | 3.50 | 22.46 | 22.38 |
| 45 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 35.30 |
| 46 | 100.00 | 3.00 | 6.0 | 5.00 | 3.50 | 23.65 | 25.02 |
| 47 | 100.00 | 7.00 | 6.0 | 5.00 | 1.50 | 24.56 | 24.40 |
* DAP, diammonium phosphate; YE, yeast extract.
Table 3.
An analysis of variance (ANOVA) of central composite design (CCD) for ethanol production from sugarcane bagasse hydrolysate (SBH) at 37 °C by S. ludwigii.
Table 3.
An analysis of variance (ANOVA) of central composite design (CCD) for ethanol production from sugarcane bagasse hydrolysate (SBH) at 37 °C by S. ludwigii.
| Source | Sum Square | df | Mean Square | F-value | p-value* | Remark |
| Model | 1619.550 | 20 | 80.980 | 51.540 | < 0.0001 | Significant |
| A | 50.420 | 1 | 50.420 | 32.090 | < 0.0001 | |
| B | 0.471 | 1 | 0.471 | 0.300 | 0.5885 | |
| C | 0.935 | 1 | 0.935 | 0.595 | 0.4474 | |
| D | 7.520 | 1 | 7.520 | 4.790 | 0.0379 | |
| E | 34.560 | 1 | 34.560 | 22.000 | < 0.0001 | |
| AB | 0.014 | 1 | 0.014 | 0.009 | 0.9266 | |
| AC | 1.910 | 1 | 1.910 | 1.220 | 0.2802 | |
| AD | 0.414 | 1 | 0.414 | 0.264 | 0.6120 | |
| AE | 1.850 | 1 | 1.850 | 1.180 | 0.2875 | |
| BC | 7.350 | 1 | 7.350 | 4.680 | 0.0399 | |
| BD | 1.440 | 1 | 1.440 | 0.920 | 0.3464 | |
| BE | 1.100 | 1 | 1.100 | 0.702 | 0.4098 | |
| CD | 0.684 | 1 | 0.684 | 0.436 | 0.5150 | |
| CE | 0.300 | 1 | 0.300 | 0.191 | 0.6656 | |
| DE | 0.0002 | 1 | 0.0002 | 0.000 | 0.9911 | |
| A2 | 1086.690 | 1 | 1086.690 | 691.660 | < 0.0001 | |
| B2 | 45.170 | 1 | 45.170 | 28.750 | < 0.0001 | |
| C2 | 746.940 | 1 | 746.940 | 475.410 | < 0.0001 | |
| D2 | 126.960 | 1 | 126.960 | 80.810 | < 0.0001 | |
| E2 | 125.030 | 1 | 125.030 | 79.580 | < 0.0001 | |
| Residual | 40.850 | 26 | 1.570 | |||
| Lack of fit | 38.950 | 22 | 1.770 | 3.730 | 0.105 | Not significant |
| Pure error | 1.900 | 4 | 0.475 | |||
| Cor Total | 1660.400 | 46 | ||||
| R2 | 0.975 | |||||
| Adj. R2 | 0.957 | |||||
| CV (%) | 5.140 |
* p = 0.05. .
Table 4.
The ethanol production from SBH by S. ludwigii and other ethanologenic yeasts using different lignocellulosic materials as feedstock.
Table 4.
The ethanol production from SBH by S. ludwigii and other ethanologenic yeasts using different lignocellulosic materials as feedstock.
| Microbe | Feedstock | Fermentation parameter 1 | References | ||||
| S (g/L) | T (°C) | P (g/L) | Qp (g/L.h) | TY (%) | |||
| P. kudriavzevii RZ8-1 | Sugarcane bagasse | 85 | 37 | 35.51 | 1.48 | 81.75 | [17] |
| 85 | 40 | 33.84 | 1.41 | 77.91 | |||
| P. kudriavzevii RGB3.2 | Rice straw | 19.10 | 40 | 9.32 | 0.39 | 95.49 | [16] |
| K. marxianus RGB4.5 | Rice straw | 19.10 | 40 | 8.03 | 0.33 | 82.27 | |
| K. marxianus CECT10875 | Woody and herbaceous biomass | NR | 42 | 19.0 | NR | 71.2 | [40] |
| S. cerevisiae | Pomelo peel waste | NR | 30 | 36.00 | 0.75 | 73.50 | [43] |
| S. cerevisiae TC-5 | Corncob residue | NR | 40 | 31.96 | 0.22 | NR | [41] |
| S. cerevisiae PTCC5052 | Wheat straw | NR | 25 | 24.02 | 0.25 | NR | [42] |
| S. ludwigii APRE2 | Pineapple waste | 105.65 | 37 | 38.02 | 1.58 | 82.35 | [23] |
| S. ludwigii APRE2 | Sugarcane bagasse | 143.95 | 37 | 38.11 | 1.59 | 88.24 | This study |
1 S, initial sugar concentration; T, fermentation temperature; P, ethanol concentration; Qp, volumetric ethanol productivity; TY, theoretical yield of ethanol; and NR, not reported.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



