Submitted:
29 April 2025
Posted:
29 April 2025
You are already at the latest version
Abstract
Piperine, a phytochemical alkaloid, exhibits anticancer properties on several cancer cells. The present study explored the important oxidative stress and signaling mechanisms by which piperine provokes cell death and apoptosis in colorectal DLD-1 cancer cells. We investigated the cell viability, cell cycle, and apoptosis-inducing of piperine by performing MTT assay, flow cytometry, gene overexpression, and Western blot analysis. Piperine treatment of DLD-1 cells diminished cell viability, and stimulated cell cycle G1 arrest and apoptosis. Piperine induced intracellular reactive oxygen species (ROS) generation by regulating mitochondrial complex III, nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, and xanthine oxidase. Piperine inhibited the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) pathway and activated the p-38 and p-extracellular signal-regulated kinase (ERK) pathways. Pretreatment with antimycin A (a mitochondrial complex III inhibitor), apocynin (an NADPH oxidase inhibitor), allopurinol (a xanthine oxidase inhibitor), and PD98059 (an ERK inhibitor) and the overexpression of p-Akt significantly recovered cell viability and reduced apoptosis. This study was the first to demonstrate a multiple oxidative stress mechanism and the critical regulating PI3K/Akt and ERK signaling pathways of piperine, inducing apoptosis on colorectal cancer cells.
Keywords:
piperine
; apoptosis
; cell cycle arrest
; oxidative stress
; PI3K/Akt
; MAPK
; colorectal cancer
1. Introduction
Piperine is a phytochemical alkaloid that exhibits the potential to provide anticancer activities in colorectal cancer. In human HRT-18 rectal cancer cells, piperine-induced apoptosis and cytotoxicity are partially achieved through ROS [1]. Piperine prompts autophagy-related cell death in colorectal cancer cells by increasing ROS level and obstructing Akt/mTOR signaling [2]. Piperine lessens ADP-ribosylation factors, such as GTPase 3-intermediated endoplasmic reticulum stress reaction, disrupts cell cycle development, and inhibits epithelial-to-mesenchymal transition in colorectal adenocarcinoma cells [3]. In several colorectal cell lines, piperine can downregulate the Wnt/β-catenin pathway to suppress cellular migration and proliferation [4]. Moreover, piperine can be used in adjuvant radiotherapy and chemotherapy to enhance sensitivity to colorectal cancer. Piperine combined with γ-radiation radiotherapy enhances cell cycle G2/M phase arrest, increases estrogen receptor beta expression, and induces HT-29 colorectal cancer cells to conduct apoptosis [5]. Piperine can be used as a chemosensitizer to 5-fluorouracil in chemoresistant colorectal cancer cells [6]. It can mediate Wnt/β-catenin signaling to synergistically suppress celecoxib-treated cell proliferation in colon cancer [7]. Hence, piperine exhibits a valuable potential against colorectal cancer.
ROS refer to the highly reactive substances including oxygen radicals. Major ROS specimens include superoxides, hydrogen peroxides, hydroxyl radicals, hypochlorous acid, singlet oxygen, and alpha-oxygen [8]. In mammalian cells, ROS are generated through various types of biochemical responses inside cellular organelles, such as mitochondria, peroxisome, and endoplasmic reticulum [8]. Mitochondria are known for generating about 90% of cellular ROS [9]. Many cellular enzymatic reactions can induce ROS production, such as cyclooxygenase, xanthine oxidase, and NADPH oxidase [10,11,12]. The increased amount of ROS can delay the growth of tumors by continuously augmenting during cell cycle inhibition. The augmented concentration of ROS can stimulate apoptosis through extrinsic and intrinsic pathways [8]. A high ROS level is believed to be a tumor-destroying mediator because the generation of intracellular ROS is used in most chemotherapeutic drugs to induce cancer cell death [8]. Piperine can stimulate cell cycle arrest and apoptosis in colorectal cancer by inducing ROS production. However, the detailed mechanisms by which piperine induces ROS production in colorectal cancer cells is currently unclear.
Many anticancer drugs can stimulate the apoptosis and inhibit the proliferation of colorectal cancer cells by stimulating ROS generation to regulate the PI3K/Akt and MAPK signaling pathways. Pyrvinium pamoate, an anthelmintic drug, can effectually prevent cell proliferation through the ROS-intervened Akt-reliant signaling pathway in colorectal cancer [13]. Antrodin C, a maleimide derivative compound extracted from Antrodia cinnamomea induces apoptosis via the ROS/Akt/ERK/p38 signaling pathway in colorectal cancer cells [14]. Metformin augments cisplatin to exhibit sensitivity in colorectal cancer cells via the ROS-related PI3K/Akt signaling pathway [15]. Lycorine reduces cell proliferation and cytostatic effects in colorectal cancer by triggering the ROS/p38 and Akt signaling pathways [16]. Delicaflavone prompts ROS-intervened apoptosis and impedes the PI3K/Akt/mTOR and Ras/MEK/Erk signaling pathways in colorectal cancer cells [17]. Fibulin-5 causes to apoptosis in colorectal cancer cell through the Akt and ROS/MAPK signaling routes [18]. Polyphyllin I stimulates apoptosis via the ROS-reduced Akt/mTOR signaling pathway in colon cancer cells [19]. Quinalizarin stimulates apoptosis via the ROS-intervened MAPK signaling pathway in colorectal cancer [20]. Manumycin and cocoa tea (Camellia ptilophylla) trigger mitochondrion-related apoptosis in HCT116 colorectal cells through the PI3K/Akt signaling pathway and ROS production [21,22]. The piperine-induced ROS mechanisms and the control of the MAPK and PI3K/Akt signaling pathways of colorectal cancers deserves further study.
In the current work, we intend to characterize the anticancer effects of piperine. This study aims to fill the gap among detailed oxidative stress mechanisms, PI3K/Akt, and MAPK signaling, offering new opinions into potential role of piperine in colorectal cancer progression.
2. Materials and Methods
2.1. Reagents and Chemicals
Penicillin streptomycin–glutamine and fetal bovine serum (FBS) were bought from Gibco Inc. (Billings, MT, USA). RPMI 1640 medium was obtained from Hyclone (South Logan, UT, USA). Piperine, trypan blue, crystal violet, DMSO, propidium iodide (PI), 2’,7’-dichlorodihydrofluorescein diacetate (DCFH-DA), MTT, and other chemicals were bought from Sigma-Aldrich (St. Louis, MO, USA). X-tremeGENE™ HP DNA transfection reagent was purchased from Roche (Raleigh, NC, USA). The protein assay kit was acquired from Bio-Rad Laboratories (Richmond, CA, USA). Primary antibodies against p53, Bax, p27, poly (ADP-ribose) polymerase (PARP), cyclin E, p-Akt, Akt, p-ERK, p-p38, p-JNK, tubulin, and GAPDH were bought from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). The 1036 pcDNA3 Myr HA Akt1 vector was obtained from Addgene Company (Watertown, MA, USA). The pcDNA 3.1(+) vector was bought from Thermo Fisher Scientific Company (Taiwan).
2.2. Cell Culture
Human colorectal DLD-1 cancer cells were cultured in RPMI 1640 medium. The medium was supplemented with 10% FBS, 2 mM of L-glutamine, 100 units/mL of penicillin G, and 100 μg/mL of streptomycin. All cells were retained at 37 °C in a 5% CO2 incubator. Stock solutions of piperine were suspended in DMSO, and all treated concentrations were attuned in the culture medium. The concentration of DMSO did not exceed 0.05%.
2.3. MTT Assay for Cell Viability
MTT assay was performed to determine cell viability. Approximately 4 × 104 cells/well in 0.5 mL of RPMI 1640 medium were incubated in 24-well plates. After growing overnight, the DLD-1 cells were treated with piperine (0, 62.5, 125, and 250 μM), or various compounds for various time points. The plates were then added with 0.5 mg/mL of MTT solution and maintained at 37 °C for additional 2 h. The supernatant was removed, and the formazan crystals were dispersed in 1 mL of DMSO. An aliquot of the DMSO lysed solution (200 μL) was collected from every well and transferred to 96-well reader plates. Optical density was measured with a microplate reader (Bio-Rad, Richmond, CA, USA) at 570 nm.
2.4. Colony Formation Assay
DLD-1 cells were placed in 24-well culture plates (5 × 104 cells/ well) and cultured with piperine (0, 62.5, 125, and 250 μM) for 48 h. The colonies were then washed with PBS, fixed with a fixing solution (methanol:glacial acetic acid; 3:1) for 10 min, cleaned again with PBS, and stained with 1% crystal violet solution for 30 min. Images were captured, and stained cell colony densities were determined using Image J software for analysis. Each well insert was added with 200 μL of the dissolving solution (33% acetic acid) to disperse the cells, and 100 μL of each dissolving solution was disperse and measured at 570 nm by using a microplate reader (Bio-Rad, Richmond, CA, USA).
2.5. Cell Cycle Analysis
DLD-1 cells (5×105) were incubated in six-well plates for overnight and then cultured with piperine (0, 62.5, 125 and 250 μM) for 48 h. Cells were collected and fixed in methanol solution (methanol:PBS = 2:1) at 4 °C for at least 24 h. After removing the methanol solution, cells were stained with a PI solution (40 μg/mL) that contained DNase-free RNase A (40 μg/mL) for 30 min at room temperature in the dark and later assessed via flow cytometry. In the resulting histogram, the X-axis denotes PI fluorescence intensity while the Y-axis denotes the number of cells.
2.6. Terminal Deoxynucleotidyl Transferase dUTP Nick end Labeling (TUNEL) Analysis
Apoptosis was assessed via the TUNEL assay. After piperine incubation, DLD-1 cells were washed with PBS and fixed with 4% paraformaldehyde for 30 min at room temperature. Cells were cleaned with PBS and then added with a TUNEL reaction kit for 1 h at 37°C. This procedure was followed by cleaning with PBS and PI addition for 30 min. TUNEL-positive cells were distinguished through flow cytometry.
2.7. Western Blot
Proteins were separated using SDS–polyacrylamide gel electrophoresis and transferred to immobilon polyvinyldifluoride (PVDF) membranes. The PVDF membranes were blocked and probed using primary antibodies (1:1000) at 4°C overnight. The blots were then incubated with secondary antibodies at room temperature for 1 h. The antigen–antibody complexes signals were evaluated via the enhanced chemiluminescence (Amersham Pharmacia Biotech, Piscataway, NJ, USA) by using a chemiluminescence analyzer.
2.8. Determination of Intracellular ROS
Intracellular ROS production was assessed via flow cytometry by using DCFH-DA staining after treating DLD-1 cells (5×105 cells/6 cm dish) with piperine (250 μM) for 1, 3, 6, and 24 h with 10 μM of DCFH-DA at 37°C for 30 min in the dark and then stained with PI (4 μg/mL) to eliminate dead cells. Intracellular ROS production was identified through the mean intensity of green fluorescence (2’,7’-DCF) via flow cytometry.
2.9. Overexpression of Akt in Cancer Cells
Approximately 6 μg of 1036 pcDNA3 Myr HA Akt1 vector (Addgene, Cambridge, MA, USA) or pcDNA 3.1(+) vector (an empty vector; Thermo-Fisher Scientific, Taiwan) was transfected to DLD-1 cells via an X-treme transfection reagent. Approximately 4 × 104 DLD-1 colorectal cancer cells were cultured in a six-well plate and maintained in a 37 °C, 5% CO2 incubator for 24 h. Thereafter, 2 mL of serum-free RPMI 1640 medium was substituted. Approximately 0.2 mL of the serum-free RPMI 1640 medium was added into an Eppendorf tube with 6 μL of X-tremeGENE HP DNA transfection reagent and 2 μg of vectors, gently pipetted and mixed smoothly. After reacting at room temperature for 15 min, approximately 400 μL was collected and placed in a dish. The cells were mixed smoothly, returned to the incubator at 37 °C for 24 h, and then incubated with of 1 μg/mL puromycin antibiotics for 14 days to create a steady Akt overexpression cell line. The stable DLD-1 cell line was cultured in RPMI 1640 medium with 10% FBS and then subjected to piperine for 48 h.
2.10. Statistical Analysis
Statistical analysis was performed using Student’s t-test with SigmaPlot 10.0 software. Data were presented as the mean ± standard deviation from at least three independent experiments. A P value of < 0.05 was considered statistically significant.
3. Results
3.1. Piperine Inhibits Cell Viability and Colony Formation in DLD-1 Cells
We analyzed the cell viability influence of piperine (0–250 μM) on DLD-1 cells for 48 h. Our results showed that piperine treatment significantly diminished DLD-1 cell viability and inhibition concentrations were 125 μM and 250 μM (Figure 1A). To further evaluate the cell proliferation effect of piperine, we treated DLD-1 cells with various concentrations of piperine (0, 62.5, 125, and 250 μM) for 48 h to assess colony formation. The colony formation capability of DLD-1 cells was inhibited significantly by incubation with piperine and inhibition was dose-dependent (Figure 1B).
3.2. Piperine Induces Cell Cycle Arrest in DLD-1 Cells
To investigate whether the piperine-induced colony formation inhibition was a consequence of cell cycle arrest, we operated a cell cycle assay via flow cytometry. In DLD-1 cells, piperine treatment for 48 h exhibited a time-dependent decrease in S phase populations (Figure 2A). Moreover, piperine at 250 μM evidently induced cell cycle G1 arrest (Figure 2A). The cell cycle G1 arrest was further verified via Western blot assay for cell cycle-regulated proteins, such as cyclin E, which demonstrated downregulation, and p27, which exhibited upregulation upon piperine treatment (Figure 2B). These results indicate that cell cycle arrest is responsible for the anticancer effect of piperine on human colorectal DLD-1 cancer cells.
3.3. Piperine Induces Apoptosis in DLD-1 Cells
Next, we verified the cell death mode by which piperine intermediates its cytotoxic influences on DLD-1 cells. To examine whether piperine induces apoptosis in DLD-1 cells, we treated DLD-1 cells with piperine for 48 h, followed by quantified TUNEL assay by using flow cytometry. A trend toward dose-dependent increase in TUNEL-positive cells was detected in piperine treated DLD-1 cells, representing the occurrence of apoptosis in DLD-1 cells (Figure 3A). These results were further verified through apoptosis regulated proteins, cleaved PARP, and Bax, via Western blot assay. The expression of cleaved PARP and Bax exhibited upregulation upon piperine treatment (Figure 3B).
3.4. Piperine Induces ROS in DLD-1 Cells
To measure the probable participation of the intracellular ROS level of cells in the apoptotic effect of piperine, we determined intracellular ROS generation in piperine-treated cells. Piperine treatment significantly increased intracellular ROS generation at 1 h and 3 h (Figure 4A). To investigate the possible sources of ROS generation induced by piperine, DLD-1 cells were pretreated with special inhibitors of mitochondrial complex I, II or III, NADPH oxidase, lipoxygenase, and xanthine oxidase for 1 h and then incubated with piperine for 1 h. Cells were estimated for intracellular ROS generation. Carboxin (a mitochondrial complex II inhibitor), antimycin A (a mitochondrial complex III inhibitor), Nordy (a lipoxygenase inhibitor), AEBSF and apocynin (NADPH oxidase inhibitors), and allopurinol (a xanthine oxidase) reduced the piperine-induced ROS production (Figure 4B). These results indicate that sources of ROS generation stimulated by piperine appears via mitochondrial respiratory chain complexes II and III, lipoxygenase, NADPH oxidase, and xanthine oxidase.
3.5. Piperine-Mediated ROS Induces Cell Death and Apoptosis in DLD-1 Cells
To investigate whether piperine-induced ROS sources were the critical events that caused DLD-1 cell death, we pretreated the inhibitors of mitochondrial complexes II and III, lipoxygenase, NADPH oxidase, and xanthine oxidase, treated them with piperine, and then evaluated cell viability via MTT assay. The inhibition of cell viability by piperine was significantly recovered by antimycin A (mitochondrial complex III inhibitor), apocynin (NADPH oxidase inhibitor), and allopurinol (xanthine oxidase) (Figure 5A). Moreover, the piperine-induced cleaved PARP was inhibited by pretreatment with antimycin A (mitochondrial complex III inhibitor), apocynin (NADPH oxidase inhibitor), and allopurinol (xanthine oxidase) (Figure 5B). These results indicate that ROS overgeneration from mitochondrial complex III, NADPH oxidase, and xanthine oxidase induced by piperine is a critical factor for DLD-1 cell death and apoptosis.
3.6. Piperine Inhibits p-Akt and Regulates MAPKs in DLD-1 Cells
ROS overproduction can mediate the PI3K/Akt and MAPK signaling pathways, followed by inducing apoptosis. To investigate whether piperine treatment can affect the PI3K/Akt and MAPK signaling pathways, we evaluated the expression of p-Akt, p-38, p-ERK, and p-JNK via Western blot. The expression of p-Akt was repressed by piperine treatment, and inhibition was concentration-dependent (Figure 6A). By contrast, the expression of p-p38 and p-ERK was enhanced by piperine treatment (Figure 6B). However, the expression of p-JNK was repressed by piperine treatment (Figure 6B).
3.7. Piperine Induces Apoptosis through Akt and ERK Signaling Regulation
To investigate whether the PI3K/Akt, p38 and ERK signaling pathways are included in piperine-induced cell death and apoptosis, we utilized the p-Akt overexpression plasmid to perform exogenic p-Akt expression, pretreated with SB203580 (a p38 inhibitor) and PD98059 (an ERK inhibitor), treated it with piperine for 48 h, and then assessed cell viability and cleaved PARP. Cell viability was significantly recovered in p-Akt overexpression and SB203580 and PD98059 pretreatment (Figure 7A,B). Meanwhile, cleaved PARP level was decreased by p-Akt overexpression (Figure 7C) and PD98059 pretreatment (Figure 7D), suggesting that the regulation of the PI3K/Akt and ERK signaling pathways by piperine was involved in inducing cell death and apoptosis.
4. Discussion
ROS play principal roles in cell death, growth, and differentiation by modifying several signaling molecules [23,24,25]. Compared with their normal counterparts, cancer cells exhibit integrally increased ROS intensities. Augmented ROS levels are strictly related to drug resistance, metastasis, and cancer initiation [26,27]. Although moderate increases in ROS levels can be favorable to cancer cells, extreme amounts of ROS can trigger cell death [28,29]. Hence, anticancer substances may effectually destroy cancer cells by regulating ROS levels. In fact, several small compounds, such as furanodienone, have been found to destroy cancer cells by increasing ROS levels in these cells [30]. In the current study, we determined that piperine exposure significantly increased intracellular ROS during early periods (1 h and 3 h) (Figure 4A), and then reduced it after treatment with carboxin (a mitochondrial complex II inhibitor), antimycin A (a mitochondrial complex III inhibitor), Nordy (a lipoxygenase inhibitor), AEBSF and apocynin (NADPH oxidase inhibitors), and allopurinol (a xanthine oxidase) (Figure 4B). These results indicates that piperine induces multiple sites of intracellular ROS generation. Among these ROS inhibitors, only antimycin A, apocynin, and allopurinol can recover cell viability (Figure 5A) and apoptosis (Figure 5B), indicating that ROS generated from mitochondrial complex III, NADPH oxidase, and xanthine oxidase are the major factors for piperine-induced cell death.
Under normal physiological conditions, the mitochondrial electron transfer chain provides 90% the intracellular ROS content in cells [31]. Some electrons are directly transferred to O2 to generate ROS in the electron transfer chain [32]. Many natural products and phytochemicals can kill cancer cells by inducing intracellular ROS through mitochondrial dysfunction in cancer cells. Azxoystrobin inhibits the development of oral cancer via the specific inhibition of mitochondrial complex III activity, resulting in ROS increase, and finally inducing apoptosis [33]. Ginsenoside Rh2, a monomeric compound from ginseng, is highly likely to cause mitochondrial ROS generation and stimulate apoptosis of cervical cancer cells by targeting the electron transfer chain complex, particularly electron transfer chain complex III [34]. Taxodione diminishes the activities of mitochondrial respiratory chain complexes III and V, generating ROS in leukemia cells [35]. Miltirone results in a dose-dependent decrease in complex III activity that involves mitochondrial dysfunction and increased ROS production in leukemia cells [36]. Kari et al. determined that mitochondrial electron transfer chain complex III bypasses electron transfer chain complex I to increase intracellular ROS in guanine nucleotide binding protein coupled receptor 17, signaling activation in glioblastoma multiforme and providing new opportunities to develop developing targeted therapy for glioblastoma multiforme [37]. These results agree with our finding that pretreatment with antimycin A prevent the repressive effects of piperine on DLD-1 cell death and apoptosis, confirming that the induction of oxidative stress by piperine from mitochondrial complex III is the mechanism behind its action in colorectal cancer cells.
One significant endogenic ROS source in tumorigenesis is NADPH oxidase [38]. As stated earlier, the subunit Nox1 of NADPH oxidase is expressed in human colon cancer cells [39]. Various stimuli can activate NADPH oxidase in cancer cells [40]. At present, numerous studies have indicated that many anticancer compounds can affect NADPH oxidase activity, inducing intracellular ROS overproduction. For example, lutein-stimulated ROS generation is reliant on NADPH oxidase, which mediates apoptosis and NF-κB activation in gastric cancer AGS cells [41]. Imidazo [1,2-a]pyridine-based derivatives evidently stimulate cytotoxicity by notably augmenting NADPH oxidase activity, which leading the stimulation of ROS-mediated apoptosis in A549 lung cancer cells [42]. Furanodienone-induced apoptosis in colorectal cancer cells is introduced by mitochondrial ROS derived from NADPH oxidase 4 [43]. Glycyrrhetinic acid can promote the productions of ROS by activating NADPH oxidases, subsequently triggering ferroptosis in triple-negative breast cancer cells [44]. The present study found that pretreatment with the NADPH oxidase inhibitor apocynin, combined with piperine treatment, effectively decreased intracellular ROS level (Figure 4B). Moreover, cell viability exhibited significant restoration under pretreatment with apocynin (Figure 5A), while cleaved PARP was demonstrated less in this group than in the piperine only group (Figure 5B). This finding proved that the repression of cell viability and the enhancement of apoptosis in DLD-1 cells by piperine partially resulted from the generation of intracellular ROS derived from NADPH oxidase. Thus, considering that piperine’s anticancer efficiency may be partly attributed to its capacity to elevate intracellular ROS derived from an NADPH oxidase, initiating a sequence of mechanism cascade that leads to cellular apoptosis, is reasonable.
Xanthine oxidase is a metabolic enzyme for purine catalysis and xenobiotic substrate oxidation, producing ROS species [45]. Under hypoxia or ischemia-reperfusion circumstances, xanthine oxidase activation has been associated with ROS-dependent tissue damage [46]. A previous report indicated that alternol, a fermented extract of the mutant fungus Alternaria alternata var. monosporus, works as a unique and novel xanthine dehydrogenase/xanthine oxidase activator in malignant cells to stimulate ROS accumulation and apoptosis [11]. Incubation of cells with allopurinol (an inhibitor of xanthine oxidase) prevents berberine-stimulated oxidative stress in human prostate PC-3 cancer cells [47]. In our study, we determined that pretreatment with the xanthine oxidase inhibitor allopurinol significantly increased cell viability (Figure 5A) and inhibited apoptosis (Figure 5B) compared with the piperine-treated group. These results indicate a potential mechanism of xanthine oxidase being involved in the ROS generation of piperine to induce cell death.
The PI3K/Akt pathway plays important role in drug resistance, metastasis, and cancer cell proliferation [48]. This pathway is typically regarded as a highly potential molecular target for cancer treatments. Previous investigations have shown that ROS suppress the PI3K/Akt/mTOR signaling pathways to induce apoptosis in colorectal cancer [14,17,18,49]. Delicaflavone triggers ROS-mediated cell cycle arrest and apoptosis via the mitochondrial pathway and endoplasmic reticulum stress accompanied by the suppression of the Ras/MEK/Erk and PI3K/Akt/mTOR signaling cascades [17]. Antrodin C extracted from A. Cinnamomea induces apoptosis via the ROS/Akt /ERK/p38 signaling pathway in colorectal cancer cells [14]. Fibulin-5 incites apoptosis via the Akt signal and ROS/MAPK pathways through the downregulation of the transient receptor potential cation channel subfamily V member 1 [18]. Hwang-Heuk-San-induced ROS generation is necessary for caspase-dependent apoptosis, including the suppression of the PI3K/Akt signaling pathway in HCT116 cells [49]. Our Western blot results confirmed that piperine deactivates the PI3K/Akt pathway in DLD-1 cells (Figure 6A). Piperine-induced cell death can be partly regained by the overexpression of continuously active Akt (Figure 7A). Moreover, our results revealed that the expression of cleaved PARP was reduced in Akt-overexpressed DLD-1 cells upon piperine treatment compared with the empty plasmid-transfected cells (Figure 7C). The overexpression of Akt can only partly remedy cell death and apoptosis, and thus, other pathways that are included in piperine-induced cell death probably exist.
The MAPK signaling pathway is downstream of ROS and plays a significant role in the stimulation of apoptosis [50]. Several investigations have proven that ROS increase can induce cell death via MAPK activation [51,52,53]. The p38 MAPK pathway is one of the stress response pathways triggered by increased ROS levels. The stimulation of the p38 pathway can induce apoptosis [54,55]. Moreover, the stress-triggered p38 pathway performs a crucial role in apoptosis control, cell cycle arrest, and cell growth inhibition [56]. The MAPK signaling pathway can be stimulated in response to oxidative stress in the endoplasmic reticulum [57], and the stimulated p38 MAPK induces either apoptosis or cell cycle arrest [59]. Moreover, increased ROS level in cells is related to the phosphorylation of p38 [60]. In general, the ERK pathway stimulated by K-ras and growth factors has been identified to prompt cell proliferation in cancer [61]. Nevertheless, several studies have demonstrated that ROS-dependent ERK activation induces apoptosis and cell cycle arrest in cancer cells [62]. Anticancer agents such as etoposide or cisplatin, need prolonged ERK activation to stimulate apoptosis in several transformed or immortalized cells [63]. In our study, the Western blot results revealed evident increases in the phosphorylation levels of ERK and p38, but not of JNK (Figure 6B). Pre-treatment with PD98059 (an inhibitor of ERK) and SB203580 (an inhibitor of p38) partially reversed cell death (Figure 7B) by piperine in DLD-1 colon cancer cells. However, only PD98059 pretreatment can decrease cleaved PARP compared with piperine-treated cells, suggesting that the activation of the ERK pathway is a signaling mechanism involved in piperine-induced cell death and apoptosis.
5. Conclusions
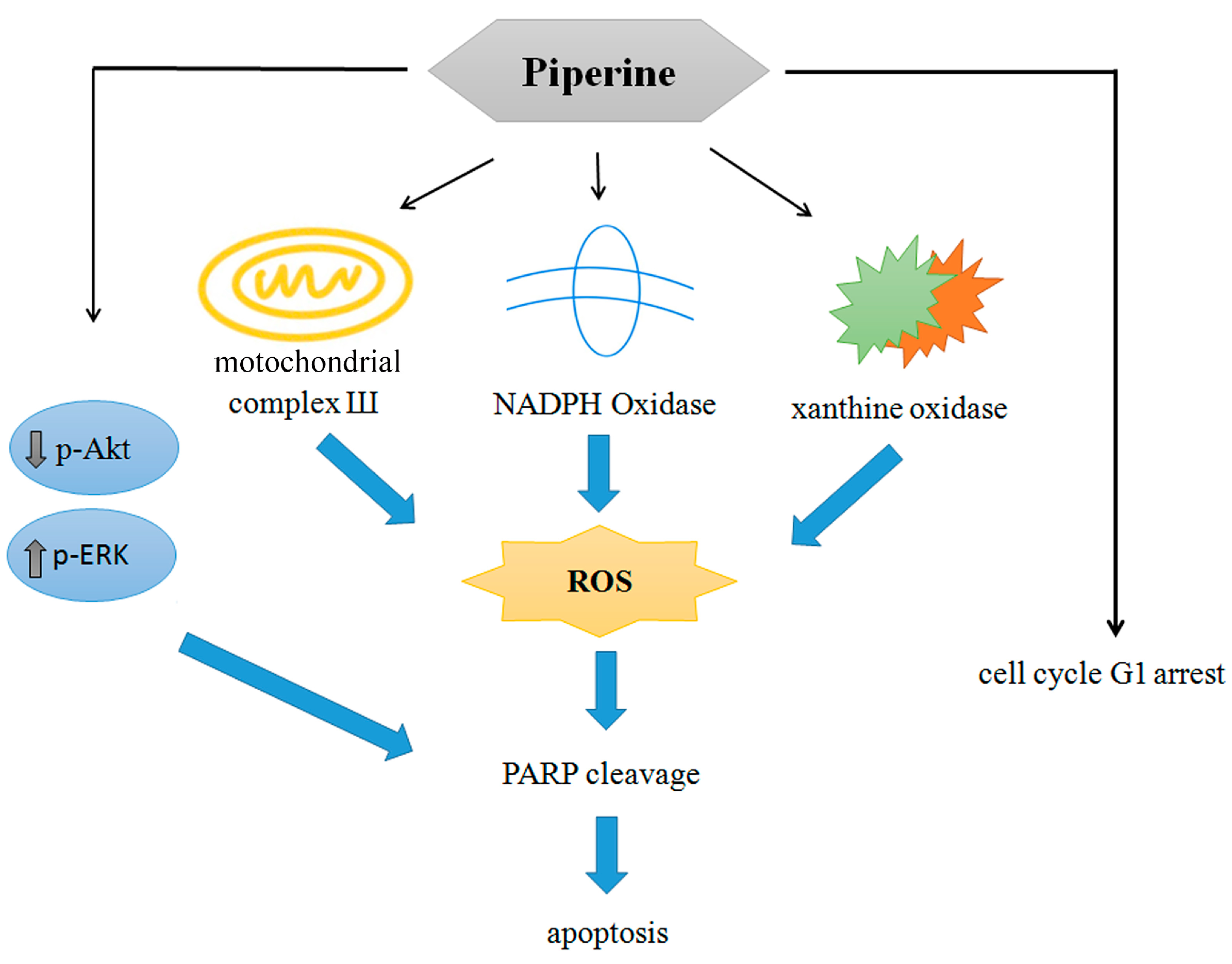
The aforementioned evidence first verifies the probability that piperine is a prospective anticancer natural compound, with the pharmaceutical capacity to inhibit colorectal cancer cell viability; induce cell cycle G1 arrest by decreasing cyclin E and increasing p27 expression; provoke apoptosis via multiple pathways, particularly by inducing multiple oxidative stress mechanisms from mitochondrial complex III, NADPH oxidase, and xanthine oxidase; and suppress the PI3K/Akt pathway and activate the ERK signaling pathway (Figure 8). However, systematic preclinical studies are still necessary to contribute scientific proof for further clinical investigations.
Author Contributions
Conceptualization, C.-H.C. and W.-L.C.; methodology, J.-Y.P., C.-L.H., P.-C.L., H.-Y.L, W.-L.C. and C.-H.C.; software, J.-Y.P., W.-L.C. and F.-J.L.; validation, C.-H.C. and W.-L.C.; resources, C.-L.H., P.-C.L. and W.-L.C.; data curation, W.-L.C., S.M.C. and C.-H.C.; writing—original draft preparation, W.-L.C., and C.-H.C.; writing—review and editing, C.-H.C.; supervision, C.-H.C. and W.-L.C.; project administration, C.-H.C. and W.-L.C.; funding acquisition, W.-L.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Chang Gung Medical Foundation, Taiwan [grant CMRPG6M0283 (W.-L.C.)].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are contained within the article.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yaffe, P.B.; Doucette, C.D.; Walsh, M.; Hoskin, D.W. Piperine impairs cell cycle progression and causes reactive oxygen species-dependent apoptosis in rectal cancer cells. Exp Mol Pathol. 2013, 94, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Guo, P.; Yang, J.; Zhang, T.; Pan, K.; Wei, H. Piperine induces autophagy of colon cancer cells: Dual modulation of AKT/mTOR signaling pathway and ROS production. Biochem Biophys Res Commun. 2024, 728, 150340. [Google Scholar] [CrossRef]
- Wu, C.; Qian, Y.; Jiang, J.; Li, D.; Feng, L. Piperine inhibits the proliferation of colorectal adenocarcinoma by regulating ARL3- mediated endoplasmic reticulum stress. Biomol Biomed. 2024, 6. [Google Scholar] [CrossRef]
- de Almeida, G.C.; Oliveira, L.F.S.; Predes, D.; Fokoue, H.H.; Kuster, R.M.; Oliveira, F.L.; Mendes, F.A.; Jose, G.; Abreu, J.G. Piperine suppresses the Wnt/β-catenin pathway and has anti-cancer effects on colorectal cancer cells. Sci Rep. 2020, 10, 11681. [Google Scholar] [CrossRef] [PubMed]
- Shaheer, K.; Somashekarappa, H.M.; Lakshmanan, M.D. Piperine sensitizes radiation-resistant cancer cells towards radiation and promotes intrinsic pathway of apoptosis. J Food Sci. 2020, 85, 4070–4079. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjya, D.; Sivalingam, N. Mechanism of 5-fluorouracil induced resistance and role of piperine and curcumin as chemo-sensitizers in colon cancer. Naunyn Schmiedebergs Arch Pharmacol. 2024, 15. [Google Scholar] [CrossRef]
- Srivastava, S.; Dewangan, J.; Mishra, S.; Divakar, A.; Chaturvedi, S.; Wahajuddin, M.; Kumar, S.; Rath, S.K. Piperine and Celecoxib synergistically inhibit colon cancer cell proliferation via modulating Wnt/β-catenin signaling pathway. Phytomedicine. 2021, 84, 153484. [Google Scholar] [CrossRef]
- Sahoo, B.M.; Banik, B.K.; Borah, P.; Jain, A. Reactive oxygen species (ROS): Key components in cancer therapies. Anticancer Agents Med Chem. 2022, 22, 215–222. [Google Scholar] [CrossRef]
- Tirichen, H.; Yaigoub, H.; Xu, W.; Wu, C.; Li, R.; Li, Y. Mitochondrial reactive oxygen species and their contribution in chronic kidney disease progression through oxidative stress. Front Physiol. 2021, 12, 627837. [Google Scholar] [CrossRef]
- Peshavariya, H. NADPH oxidase-derived ROS signaling and therapeutic opportunities. Curr Pharm Des. 2015, 21, 21,5931–2. [Google Scholar] [CrossRef]
- Xu, H.; Li, C.; Mozziconacci, O.; Zhu, R.; Xu, Y.; Tang, Y.; Chen, R.; Huang, Y.; Holzbeierlein, J.M.; Schöneich, C.; Huang, J.; Li, B. Xanthine oxidase-mediated oxidative stress promotes cancer cell-specific apoptosis. Free Radic Biol Med. 2019, 139, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Laws, K.; Eskandari, A.; Suntharalingam, K. A reactive oxygen species-generating, cyclooxygenase-2 inhibiting, cancer stem cell-potent tetranuclear copper (ii) cluster. Dalton Trans. 2017, 46, 12785–12789. [Google Scholar] [CrossRef]
- Zheng, W.; Hu, J.; Lv, Y.; Bai, B.; Shan, L.; Chen, K.; Dai, S.; Zhu, H. Pyrvinium pamoate inhibits cell proliferation through ROS-mediated AKT-dependent signaling pathway in colorectal cancer. Med Oncol. 2021, 838, 21. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.T.; Lee, K.C.; Cheng, K.C.; Lee, K.F.; Yang, Y.L.; Chu, H.T.; Lin, T.W.; Chen, C.C.; Hsieh, M.C.; Huang, C.Y.; Kuo, H.C.; Teng, C.C. Antrodin C isolated from Antrodia Cinnamomea induced apoptosis through ROS/AKT/ERK/p38 signaling pathway and epigenetic histone acetylation of TNFα in colorectal cancer cells. Antioxidants (Basel). 2023, 12, 764. [Google Scholar] [CrossRef]
- Zhang, P.; Zhao, S.; Lu, X.; Shi, Z.; Liu, H.; Zhu, B. Metformin enhances the sensitivity of colorectal cancer cells to cisplatin through ROS-mediated PI3K/Akt signaling pathway. Gene. 2020, 745, 144623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Yuan, X.; Yu, T.; Huang, H.; Yang, C.; Zhang, L.; Yang, S.; Luo, X.; Luo, J. Lycorine inhibits cell proliferation, migration and invasion, and primarily exerts in vitro cytostatic effects in human colorectal cancer via activating the ROS/p38 and AKT signaling pathways. Oncol Rep. 2021, 19. [Google Scholar] [CrossRef]
- Yao, W.; Lin, Z.; Shi, P.; Chen, B. Wang, G.; Huang, J.; Sui, Y.; Liu, Q.; Li, S.; Lin, X.; Liu, Q.; Yao, H. Delicaflavone induces ROS-mediated apoptosis and inhibits PI3K/AKT/mTOR and Ras/MEK/Erk signaling pathways in colorectal cancer cells. Biochem Pharmacol. 2020, 171, 113680. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Jin, L.; Lei, K.; Liu, H.; Yang, Y. Fibulin-5 contributes to colorectal cancer cell apoptosis via the ROS/MAPK and Akt signal pathways by downregulating transient receptor potential cation channel subfamily V member 1. J Cell Biochem. 2019, 120, 17838–17846. [Google Scholar] [CrossRef]
- Luo, Q.; Jia, L.; Huang, C.; Qi, Q.; Jahangir, A.; Xia, Y.; Liu, W.; Shi, R.; Tang, L.; Chen, Z. Polyphyllin I Promotes Autophagic Cell Death and Apoptosis of Colon Cancer Cells via the ROS-Inhibited AKT/mTOR Pathway. Int J Mol Sci. 2022, 23, 9368. [Google Scholar] [CrossRef]
- Meng, L.Q.; Wang, Y.; Luo, Y.H.; Piao, X.J.; Liu, C,; Wang, Y. ; Zhang, Y.; Wang, J.R.; Wang, H.; Xu, W.T.; Liu, Y.; Wu, Y.Q.; Sun, H.N.; Han, Y.H.; Jin, M.H.; Shen, G.N.; Fang, N.Z.; Jin, C.H. Quinalizarin induces apoptosis through reactive oxygen species (ROS)-mediated mitogen-activated protein kinase (MAPK) and signal transducer and activator of transcription 3 (STAT3) signaling pathways in colorectal cancer cells. Med Sci Monit. 2018, 24, 3710–3719. [Google Scholar] [CrossRef]
- Gao, X.; Li, X.; Ho, C.T.; Lin, X.; Zhang, Y.; Li, B.; Chen, Z. Cocoa tea (Camellia ptilophylla) induces mitochondria-dependent apoptosis in HCT116 cells via ROS generation and PI3K/Akt signaling pathway. Food Res Int. 2020, 129, 108854. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiang, H.; Xie, L.; Hu, J.; Li, L.; Yang, M.; Cheng, L.; Liu, B.; Qian, X. Antitumor effect of manumycin on colorectal cancer cells by increasing the reactive oxygen species production and blocking PI3K-AKT pathway. Onco Targets Ther. 2016, 9, 2885–2895. [Google Scholar] [CrossRef] [PubMed]
- D'Autréaux, B.; Toledano, M.B. ROS as signalling molecules: mechanisms that generate specificity in ROS homeostasis. Nat Rev Mol Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat Rev Mol Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Galano, J.M.; Durand, T.; Le Guennec, J.Y.; Lee, J.C. Physiological role of reactive oxygen species as promoters of natural defenses. FASEB J. 2017, 31, 3729–3745. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwa,l B. B. Oxidative stress, inflammation, and cancer: how are they linked? Free Radic Biol Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tafani, M.; Sansone, L.; Limana, F.; Arcangeli, T.; De Santis, E.; Polese, M.; Fini, M.; Russo, M.A. The Interplay of Reactive Oxygen Species, Hypoxia, Inflammation, and Sirtuins in Cancer Initiation and Progression. Oxid Med Cell Longev. 2016, 2016, 3907147. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free Radic Biol Med. 2017, 104, 144–164. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.S.; Chang, J.H.; Hung, W.Y.; Yang, Y.C.; Chien, M.H. The interplay of reactive oxygen species and the epidermal growth factor receptor in tumor progression and drug resistance. J Exp Clin Cancer Res. 2018, 37, 61. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jiang, Y.; Wang, X.; Hu, D. Furanodienone induces G0/G1 arrest and causes apoptosis via the ROS/MAPKs-mediated caspase-dependent pathway in human colorectal cancer cells: a study in vitro and in vivo. Cell Death Dis. 2017, 8, e2815. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gao, L.; Laude, K.; Cai, H. Mitochondrial pathophysiology, reactive oxygen species, and cardiovascular diseases. Vet Clin North Am Small Anim Pract. 2008, 38, 137–155. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int J Mol Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, H.; Li, L.; Lu, Y.; Shen, Y.; Zhang, M.; Ge, L.; Wang, M.; Yang, J.; Tian, Z.; Tang, X. Azoxystrobin Reduces Oral Carcinogenesis by Suppressing Mitochondrial Complex III Activity and Inducing Apoptosis. Cancer Manag Res. 2020, 12, 11573–11583. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liu, Y.; Yu, S.; Xing, X.; Qiao, J.; Yin, Y.; Wang, J.; Liu, M.; Zhang, W. Ginsenoside Rh2 stimulates the production of mitochondrial reactive oxygen species and induces apoptosis of cervical cancer cells by inhibiting mitochondrial electron transfer chain complex. Mol Med Rep. 2021, 24, 873. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Uchihara, Y.; Tago, K.; Taguchi, H.; Narukawa, Y.; Kiuchi, F.; Tamura, H.; Funakoshi-Tago, M. Taxodione induces apoptosis in BCR-ABL-positive cells through ROS generation. Biochem Pharmacol. 2018, 154, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Jiang, L.; Xu, M.; Liu, Q.; Gao, N.; Li, P.; Liu, E.H. Miltirone exhibits antileukemic activity by ROS-mediated endoplasmic reticulum stress and mitochondrial dysfunction pathways. Sci Rep. 2016, 6, 20585. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kari, S.; Kandhavelu, J.; Murugesan, A.; Thiyagarajan, R.; Kidambi, S.; Kandhavelu, M. Mitochondrial complex III bypass complex I to induce ROS in GPR17 signaling activation in GBM. Biomed Pharmacother. 2023, 162, 114678. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.S. The signaling mechanism of ROS in tumor progression. Cancer Metastasis Rev. 2006, 25, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, A.; Markel, S.; Gaur, S.; Liu, H.; Lu, J.; Jiang, G.; Wu, X.; Antony, S.; Wu, Y.; Melillo, G.; Meitzler, J.L.; Haines, D.C.; Butcher, D.; Roy, K.; Doroshow, J.H. NADPH oxidase 1 supports proliferation of colon cancer cells by modulating reactive oxygen species-dependent signal transduction. J Biol Chem. 2017, 292, 7866–7887. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bánfi, B.; Clark, R.A.; Steger, K.; Krause, K.H. Two novel proteins activate superoxide generation by the NADPH oxidase NOX1. J Biol Chem. 2003, 278, 3510–3513. [Google Scholar] [CrossRef] [PubMed]
- Eom, J.W.; Lim, J.W.; Kim, H. Lutein Induces Reactive Oxygen Species-Mediated Apoptosis in Gastric Cancer AGS Cells via NADPH Oxidase Activation. Molecules. 2023, 28, 1178. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bhavya, K.; Mantipally, M.; Roy, S.; Arora, L.; Badavath, V.N.; Gangireddy, M.; Dasgupta, S.; Gundla, R.; Pal, D. Novel imidazo [1,2-a]pyridine derivatives induce apoptosis and cell cycle arrest in non-small cell lung cancer by activating NADPH oxidase mediated oxidative stress. Life Sci. 2022, 294, 120334. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, Y.; Wang, Y.; Li, X. Furanodienone induces apoptosis via regulating the PRDX1/MAPKs/p53/caspases signaling axis through NOX4-derived mitochondrial ROS in colorectal cancer cells. Biochem Pharmacol. 2024, 227, 116456. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Chen, H.; Zhang, L.; Wu, M.; Zhang, F.; Yang, D.; Shen, J.; Chen, J. Glycyrrhetinic acid induces oxidative/nitrative stress and drives ferroptosis through activating NADPH oxidases and iNOS, and depriving glutathione in triple-negative breast cancer cells. Free Radic Biol Med. 2021, 173, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Winston, G.W.; Moore, M.N.; Kirchin, M.A.; Soverchia, C. Production of reactive oxygen species by hemocytes from the marine mussel, Mytilus edulis: lysosomal localization and effect of xenobiotics. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol. 1996, 113, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.S.; Lu, L.Q.; Jiang, Y.Q.; Li, N.S.; Luo, X.J.; Peng, J. W, Peng, J. Allopurinol attenuates oxidative injury in rat hearts suffered ischemia/reperfusion via suppressing the xanthine oxidase/vascular peroxidase 1 pathway. Eur J Pharmacol. 2021, 908, 174368. [Google Scholar] [CrossRef] [PubMed]
- Meeran, S.M.; Katiyar, S.; Katiyar, S.K. Berberine-induced apoptosis in human prostate cancer cells is initiated by reactive oxygen species generation. Toxicol Appl Pharmacol. 2008, 229, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Cheng, H.; Roberts, T.M.; Zhao, J.J. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev Drug Discov. 2009, 8, 627–644. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lee, M.H.; Hong, S.H.; Park, C.; Kim, G.Y.; Leem, S.H.; Choi, S.H.; Keum, Y.S.; Hyun, J.W.; Kwon, T.K.; Hong, S.H.; Choi, Y.H. Hwang-Heuk-San induces apoptosis in HCT116 human colorectal cancer cells through the ROS-mediated activation of caspases and the inactivation of the PI3K/Akt signaling pathway. Oncol Rep. 2016, 36, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fan, D. Ginsenoside Rg5 induces G2/M phase arrest, apoptosis and autophagy via regulating ROS-mediated MAPK pathways against human gastric cancer. Biochem Pharmacol. 2019, 168, 285–304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; He, J.; Ye, X.; Zhu, J.; Hu, X.; Shen, M.; Ma, Y.; Mao, Z.; Song, H.; Chen, F. β-Thujaplicin induces autophagic cell death, apoptosis, and cell cycle arrest through ROS-mediated Akt and p38/ERK MAPK signaling in human hepatocellular carcinoma. Cell Death Dis. 2019, 10, 255. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, L.J.; Lee, Y.C.; Huang, C.H.; Shi, Y.J.; Chen, Y.J.; Pei, S.N.; Chou, Y.W.; Chang, L.S. Non-mitotic effect of albendazole triggers apoptosis of human leukemia cells via SIRT3/ROS/p38 MAPK/TTP axis-mediated TNF-α upregulation. Biochem Pharmacol. 2019, 162, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Ren, D.; Wang, J.; Liu, X.; Zhang, H.; Wu, M.; Yang, G. Bruceine D induces lung cancer cell apoptosis and autophagy via the ROS/MAPK signaling pathway in vitro and in vivo. Cell Death Dis. 2020, 11, 126. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Osone, S.; Hosoi, H.; Kuwahara, Y.; Matsumoto, Y.; Iehara, T.; Sugimoto, T. Fenretinide induces sustained-activation of JNK/p38 MAPK and apoptosis in a reactive oxygen species-dependent manner in neuroblastoma cells. Int J Cancer. 2004, 112, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiang, D.; Liu, J.; Ye, S.; Xiao, S.; Wang, W.; Sun, Z.; Xie, Y.; Wang, J. Compound K induces apoptosis of bladder cancer T24 cells via reactive oxygen species-mediated p38 MAPK pathway. Cancer Biother Radiopharm. 2013, 28, 607–614. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol Mol Biol Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Darling, N.J.; Cook, S.J. The role of MAPK signalling pathways in the response to endoplasmic reticulum stress. Biochim Biophys Acta. 2014, 1843, 2150–2163. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Penninger, J.M. Mitogen-activated protein kinases in apoptosis regulation. Oncogene. 23, 2838–2849. [CrossRef] [PubMed]
- Ko, Y.H.; Kim, S.K.; Kwon, S.H.; Seo, J.Y.; Lee, B.R.; Kim, Y.J.; Hur, K.H.; Kim, S.Y.; Lee, S.Y.; Jang, C.G. 7,8,4'-Trihydroxyisoflavone, a Metabolized Product of Daidzein, Attenuates 6-Hydroxydopamine-Induced Neurotoxicity in SH-SY5Y Cells. Biomol Ther (Seoul). 2019, 27, 363–372. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kulisz, A.; Chen, N.; Chandel, N.S.; Shao, Z.; Schumacker, P.T. Mitochondrial ROS initiate phosphorylation of p38 MAP kinase during hypoxia in cardiomyocytes. Am J Physiol Lung Cell Mol Physiol. 2002, 282, L1324–9. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.J.; Der, C.J. Targeting the Raf-MEK-ERK mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene. 2007, 26, 3291–3310. [Google Scholar] [CrossRef] [PubMed]
- Bastola, T.; An, R.B.; Kim, Y.C.; Kim, J.; Seo, J. Cearoin Induces Autophagy, ERK Activation and Apoptosis via ROS Generation in SH-SY5Y Neuroblastoma Cells. Molecules. 2017, 22, 242. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cagnol, S.; Chambard, J.C. ERK and cell death: mechanisms of ERK-induced cell death--apoptosis, autophagy and senescence. FEBS J. 2010, 277, 2–21. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Piperine inhibits cell viability and colony formation in DLD-1 cells. (A) Cell viability of DLD-1 cells incubated for 48 h with different concentrations of piperine assessed via MTT assays. (B) Colony formation in DLD-1 cells incubated with piperine (0, 62.5, 125, and 250 μM) for 48 h. Significant differences in the 0 μM-treated group are presented P<0.05 (*) and P<0.001 (***).
Figure 1.
Piperine inhibits cell viability and colony formation in DLD-1 cells. (A) Cell viability of DLD-1 cells incubated for 48 h with different concentrations of piperine assessed via MTT assays. (B) Colony formation in DLD-1 cells incubated with piperine (0, 62.5, 125, and 250 μM) for 48 h. Significant differences in the 0 μM-treated group are presented P<0.05 (*) and P<0.001 (***).
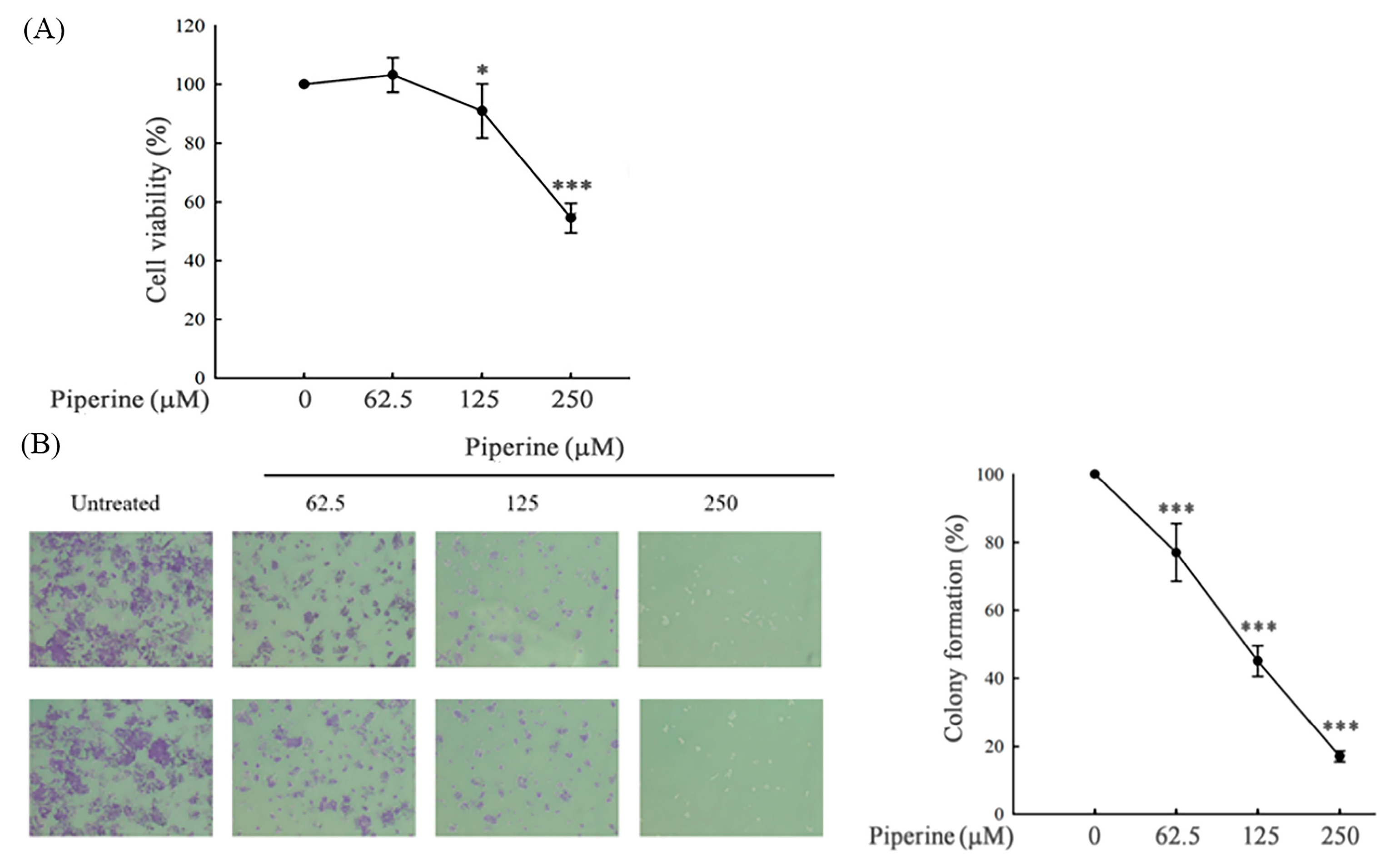
Figure 2.
Piperine induces cell cycle G1 arrest in DLD-1 cells. (A) Cell cycle of DLD-1 cells cultured for 48 h with different concentrations of piperine assessed via PI staining and flow cytometry. (B) Expression of cell cycle-regulated proteins, cyclin E, and p27 in DLD-1 cells incubated with piperine (0, 62.5, 125, and 250 μM) for 48 h.
Figure 2.
Piperine induces cell cycle G1 arrest in DLD-1 cells. (A) Cell cycle of DLD-1 cells cultured for 48 h with different concentrations of piperine assessed via PI staining and flow cytometry. (B) Expression of cell cycle-regulated proteins, cyclin E, and p27 in DLD-1 cells incubated with piperine (0, 62.5, 125, and 250 μM) for 48 h.
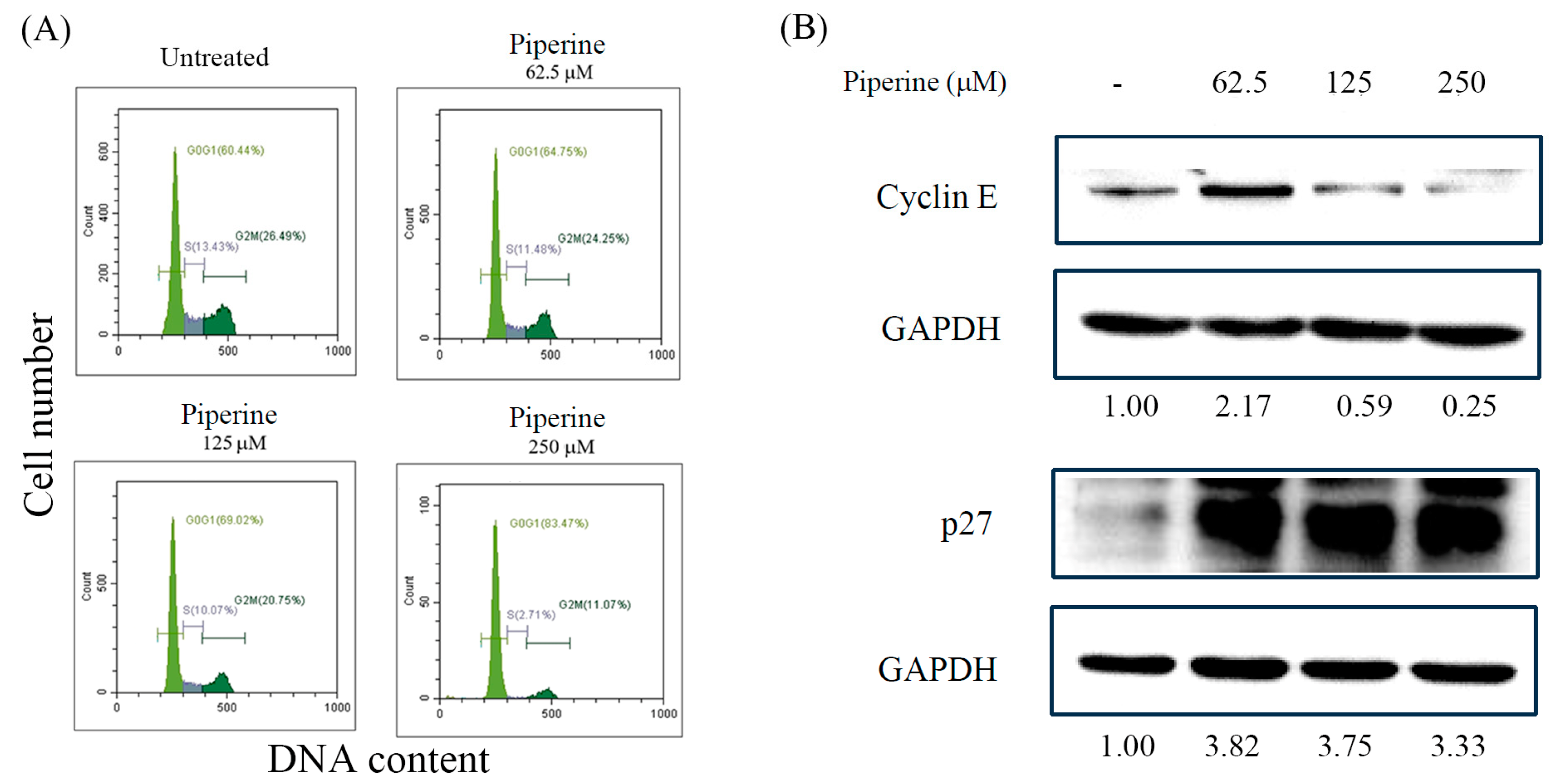
Figure 3.
Piperine induces apoptosis in DLD-1 cells. (A) Percentages of apoptosis in DLD-1 cells incubated for 48 h with piperine (0, 62.5, 125, and 250 μM) assessed via TUNEL assay and flow cytometry. (B) Expression of apoptosis marker proteins, cleaved PARP, and Bax in DLD-1 cells incubated with piperine (0, 62.5, 125, and 250 μM) for 48 h.
Figure 3.
Piperine induces apoptosis in DLD-1 cells. (A) Percentages of apoptosis in DLD-1 cells incubated for 48 h with piperine (0, 62.5, 125, and 250 μM) assessed via TUNEL assay and flow cytometry. (B) Expression of apoptosis marker proteins, cleaved PARP, and Bax in DLD-1 cells incubated with piperine (0, 62.5, 125, and 250 μM) for 48 h.
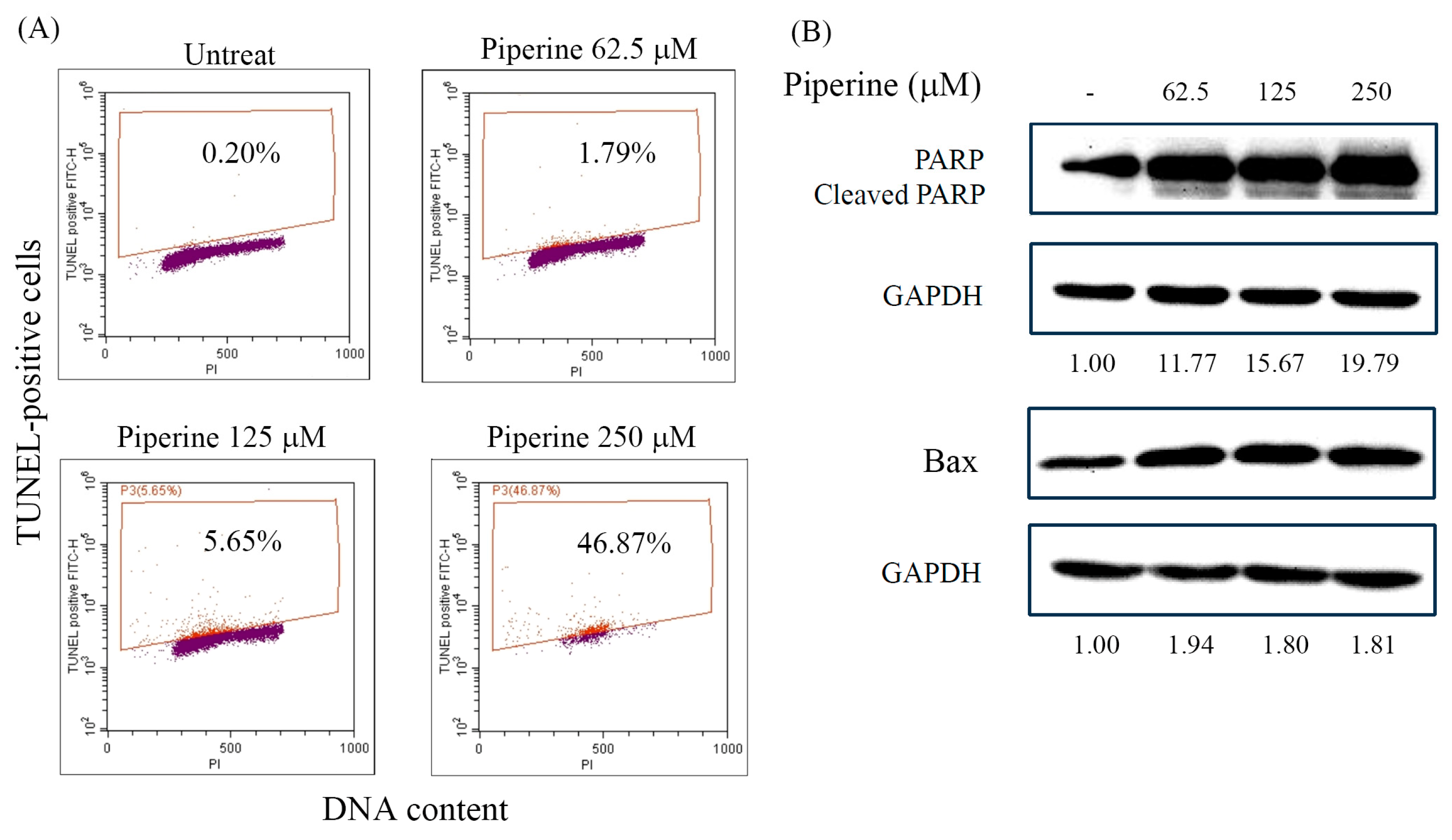
Figure 4.
Piperine induces ROS in DLD-1 cells. (A) Intracellular ROS of DLD-1 cells incubated for 1, 3, 6, and 24 h with 250 μM of piperine assessed via DCFH-DA staining and flow cytometry. (B) Intracellular ROS of DLD-1 cells pre-treated for various ROS-generated inhibitors for 1 h and then incubated with 250 μM of piperine for 1 h. After treatment, intracellular ROS was assessed via DCFH-DA staining and flow cytometry. Significant differences in the untreated group are exhibited P<0.05 (*) and P<0.001 (***). Significant differences in the piperine-incubated group are presented P<0.01 (##) and P<0.001 (###).
Figure 4.
Piperine induces ROS in DLD-1 cells. (A) Intracellular ROS of DLD-1 cells incubated for 1, 3, 6, and 24 h with 250 μM of piperine assessed via DCFH-DA staining and flow cytometry. (B) Intracellular ROS of DLD-1 cells pre-treated for various ROS-generated inhibitors for 1 h and then incubated with 250 μM of piperine for 1 h. After treatment, intracellular ROS was assessed via DCFH-DA staining and flow cytometry. Significant differences in the untreated group are exhibited P<0.05 (*) and P<0.001 (***). Significant differences in the piperine-incubated group are presented P<0.01 (##) and P<0.001 (###).
Figure 5.
Piperine-mediated ROS induces cell death and apoptosis in DLD-1 cells. DLD-1 cells were incubated with various ROS-generated inhibitors for 1 h and then with 250 μM of piperine for 48 h. After treatment, cell viability and cleaved PARP were analyzed via (A) MTT assay and (B) Western blot. Significant differences in the piperine-treated group are exhibited P<0.05 (*), P<0.01 (**), and P<0.001 (***).
Figure 5.
Piperine-mediated ROS induces cell death and apoptosis in DLD-1 cells. DLD-1 cells were incubated with various ROS-generated inhibitors for 1 h and then with 250 μM of piperine for 48 h. After treatment, cell viability and cleaved PARP were analyzed via (A) MTT assay and (B) Western blot. Significant differences in the piperine-treated group are exhibited P<0.05 (*), P<0.01 (**), and P<0.001 (***).
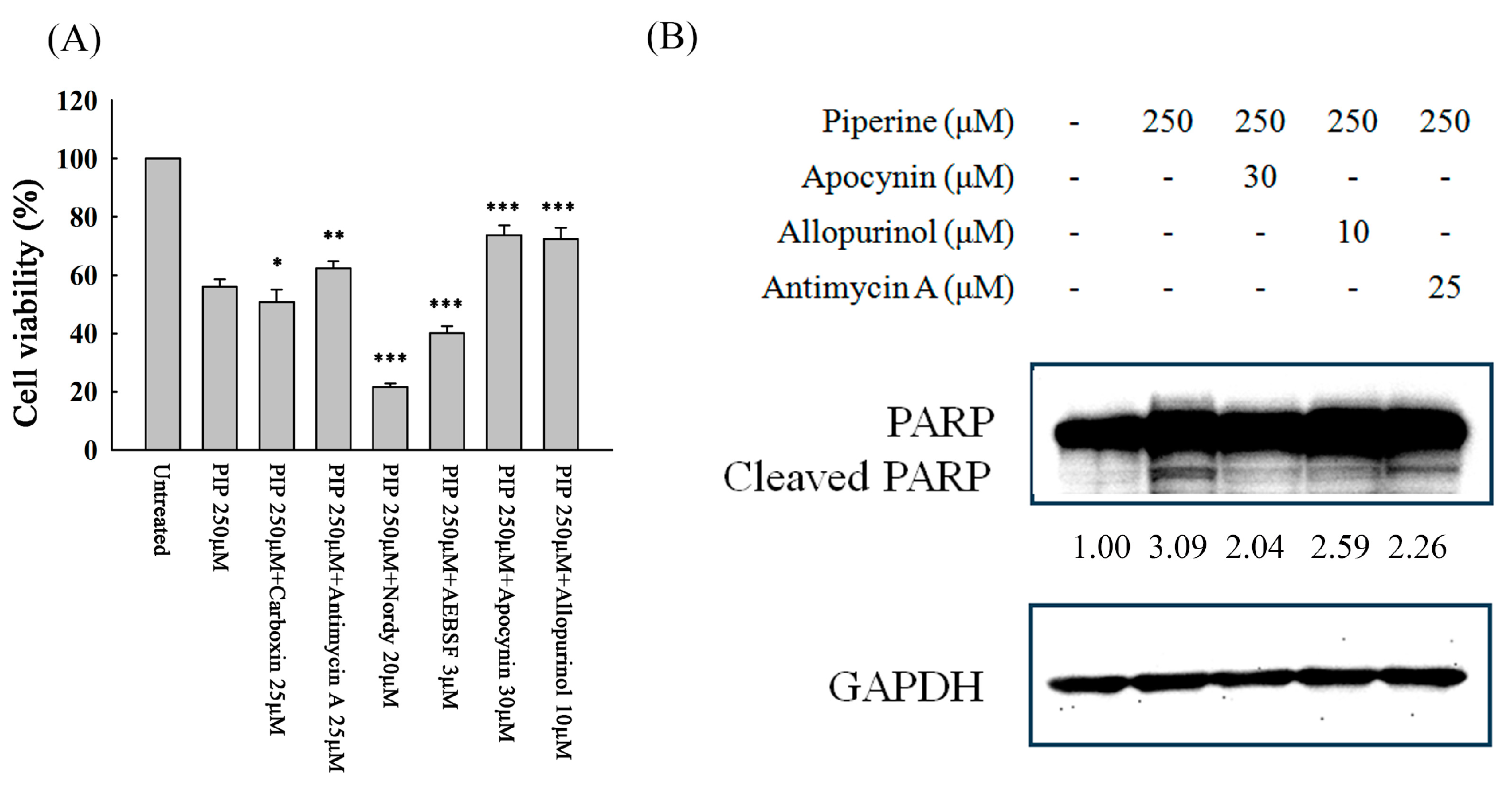
Figure 6.
Piperine inhibits p-Akt and regulates MAPKs in DLD-1 cells. Expression of (A) p-Akt and Akt and (B) MAPK proteins in DLD-1 cells incubated with piperine (0, 62.5, 125, and 250 μM) for 48 h.
Figure 6.
Piperine inhibits p-Akt and regulates MAPKs in DLD-1 cells. Expression of (A) p-Akt and Akt and (B) MAPK proteins in DLD-1 cells incubated with piperine (0, 62.5, 125, and 250 μM) for 48 h.
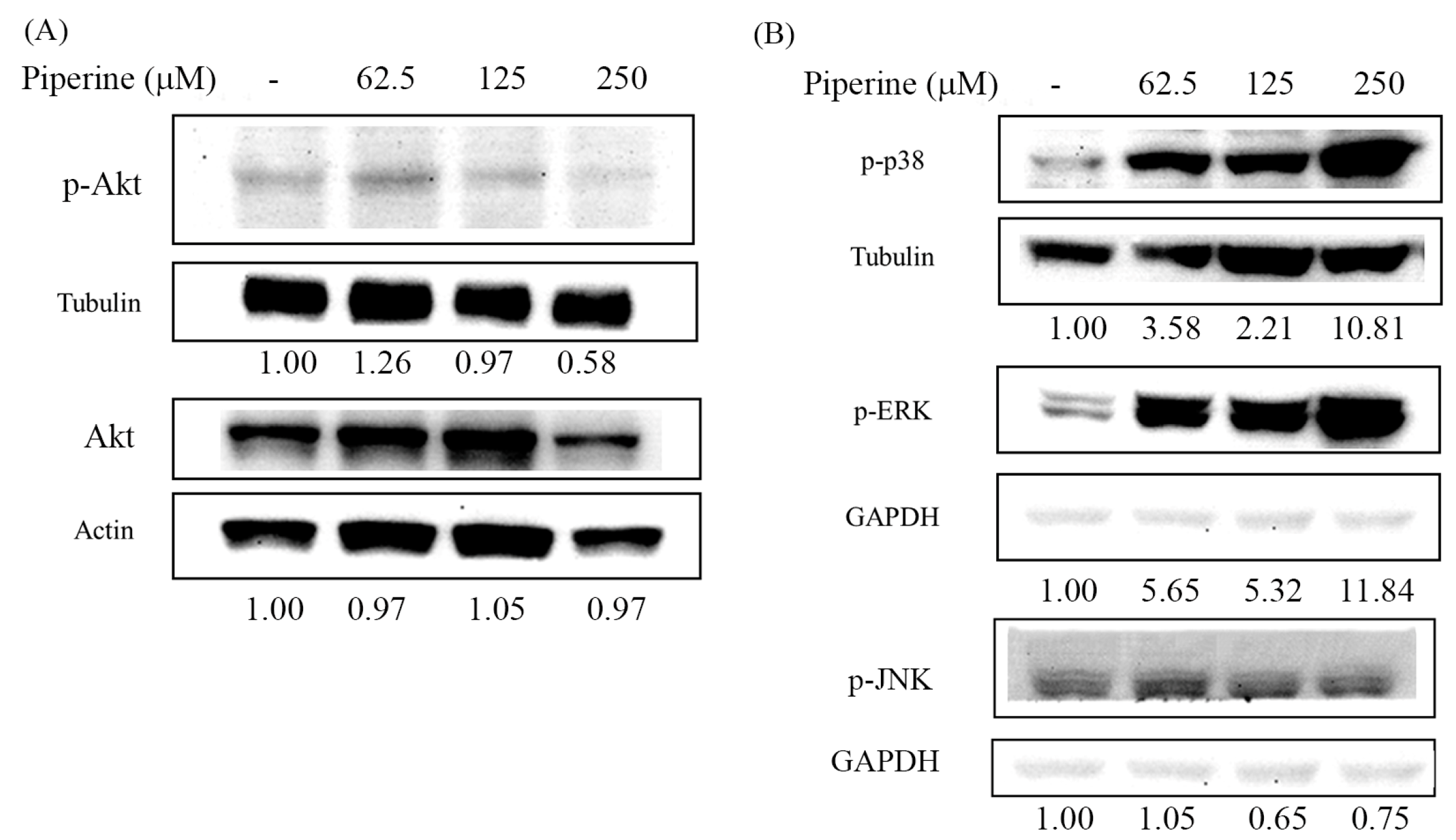
Figure 7.
Piperine induces cell death and apoptosis through the Akt, p38, and ERK signaling pathways. (A) DLD-1 cells were transfected with empty plasmid or pAkt-overexpressed plasmid and then treated with 250 μM of piperine for 48 h. After treatment, cell viability was assessed via MTT assay. Significant differences in the untreated and piperine-treated empty plasmid groups are presented P<0.001 (***) and P<0.001 (###), respectively. (B) Pretreatment of DLD-1 cells with 50 μM SP600125 or 50 μM PD98059 for 1 h, then incubation with 250 μM of piperine for 48 h. Cell viability was analyzed via MTT assay. Significant differences in the piperine-treated group are presented P<0.01 (**). (C) p-Akt expression was detected in the DLD-1, empty plasmid-, or pAkt-overexpressed plasmid transfected DLD-1 cells. Cells were incubated with 250 μM of piperine for 48 h. Cleaved PARP was measured via Western blot. (D) Pretreatment of DLD-1 cells with 50 μM SP600125 or 50 μM PD98059 for 1 h, then incubation with 250 μM of piperine for 48 h. Cleaved PARP was measured via Western blot.
Figure 7.
Piperine induces cell death and apoptosis through the Akt, p38, and ERK signaling pathways. (A) DLD-1 cells were transfected with empty plasmid or pAkt-overexpressed plasmid and then treated with 250 μM of piperine for 48 h. After treatment, cell viability was assessed via MTT assay. Significant differences in the untreated and piperine-treated empty plasmid groups are presented P<0.001 (***) and P<0.001 (###), respectively. (B) Pretreatment of DLD-1 cells with 50 μM SP600125 or 50 μM PD98059 for 1 h, then incubation with 250 μM of piperine for 48 h. Cell viability was analyzed via MTT assay. Significant differences in the piperine-treated group are presented P<0.01 (**). (C) p-Akt expression was detected in the DLD-1, empty plasmid-, or pAkt-overexpressed plasmid transfected DLD-1 cells. Cells were incubated with 250 μM of piperine for 48 h. Cleaved PARP was measured via Western blot. (D) Pretreatment of DLD-1 cells with 50 μM SP600125 or 50 μM PD98059 for 1 h, then incubation with 250 μM of piperine for 48 h. Cleaved PARP was measured via Western blot.
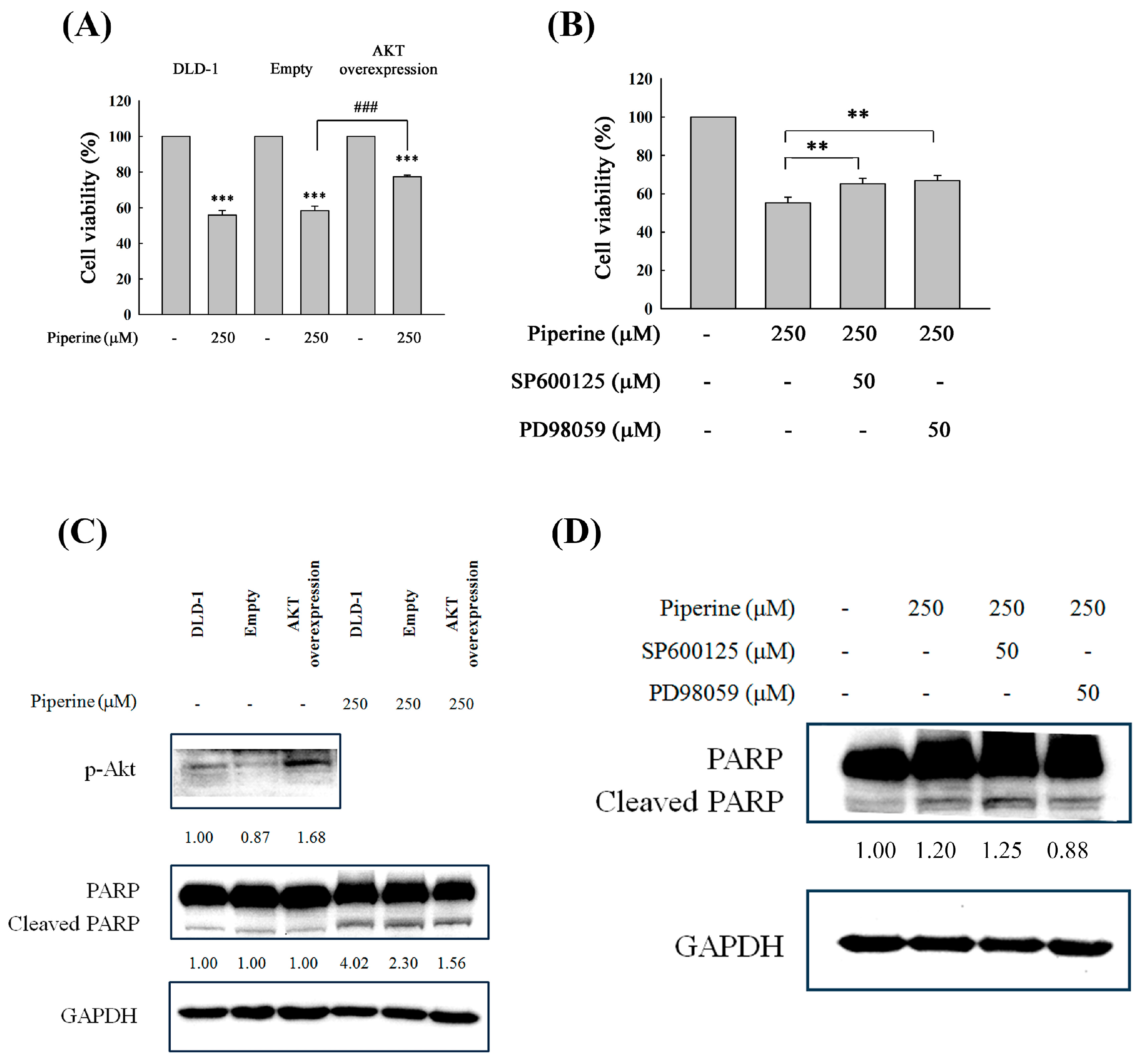
Figure 8.
Proposed model of piperine induces apoptosis in colorectal DLD-1 cancer cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



